
Submitted:
28 February 2023
Posted:
01 March 2023
You are already at the latest version
Abstract
Selectins are responsible for the early stages of cell migration as they control cell adhesion. They make the microenvironment permissive for metastatic events by promoting the activation of other cell adhesion molecules (CAMs). We employed several robust bioinformatics tools to evaluate gene sequence; single nucleotide polymorphisms (SNPs) in intronic and UTR regions; and missense SNPs with amino acid change in L-selectin (SELL), E-selectin (SELE), P-selectin (SELP) and PSGL-1 (SELPLG). We demonstrated that gene polymorphisms rs2229569, rs1131498, rs4987360, rs4987301 and rs2205849; polymorphisms rs3917777, rs2205894 and rs2205893 of SELP gene; rs7138370, rs7300972 and rs2228315 variants of SELPLG gene; and rs1534904 and rs5368 polymorphisms of SELE gene may produce important alterations in the DNA structure and consequent alterations in the morphology and function of the corresponding proteins. These Selectin polymorphic variants deserve further investigation in cancer patients, as they may become useful clinical markers for risk determination, diagnostic and prognostic biomarkers or even targets for targeted therapies.
Keywords:
bioinformatics analysis
; selectins
; cancer
1. Introduction
Single nucleotide polymorphisms (SNP) are variations in DNA that cause a single nitrogenous base substitution in the gene sequence. Most SNPs are neutral, but some may contribute to the predisposition to disease, consequently acting as genetic markers [1]; or influence their evolution and serve as useful outcome markers; or even determine the responses to the treatment of diseases such as cancer [2]. The main advantages of using SNPs as biomarkers are related to their stability (because they have a low mutation rate), high frequency (they are present in more than 1% of the population), and facilitate the optimization of analysis techniques through automation [3,4].
Depending on its location, the SNP can promote nucleotide substitutions that lead to different alterations in the DNA; alterations in the formation of proteins (to their structure, function and stability); alterations in protein formation (regarding its structure, function and stability) [5] and in the regulation of protein-protein interactions [6]; changes in the mechanisms of splicing, transcription, localization and degradation of mRNAs [7]; functional changes in transcription factor binding sites, intron/ exon splicing sites, exonic splicing promoter sites, and miRNA binding sites [7].
The impact of the effects of amino acid substitution on the structure and function of a given protein is essential for a better understanding of the complex mechanisms involved in diseases caused or related to this protein. Likewise, it is important to understand the effect of SNPs on the regulation of gene expression [3,4].
Cell adhesion molecules (CAMs) are transmembrane proteins that play important roles in cell-cell communication and interaction [8,9,10]. Selectins are a family of CAMs that are involved in the initial steps of cell adhesion and are particularly important in the immune response. Selectins are able to bind to specific carbohydrates on the surface of different cells, helping to bring these cells closer together. This is important for the immune response, as it allows leukocytes to move from the blood vessels to sites of infection or injury in tissues [11,12]. The interaction between selectins and carbohydrates is weak and reversible allowing cells to adhere and detach from each other quickly and efficiently [11,12]. In addition to their role in the immune response, selectins have also been implicated in other processes, such as tumor metastasis [13,14]. This effect has enabled the development of target-directed drugs that may become important in the treatment of these conditions.
There are three types of selectins: L-selectin, P-selectin, and E-selectin [12,15,16]. L-selectin is encoded by SELL gene, located on the long arm of chromosome 1 (1q24.2) [17]. It is believed that the glycosylation patterns of this protein may dictate its functions in the cell, but the mechanisms involved in these functions are still unclear [17]. Initially its expression was considered to be exclusive to the leukocyte surface, including myeloid cells, naïve T cells and some activated T cells [17,18], but this molecule expression has more recently been observed in several types of cancer and other cell types [13,14,19,20,21,22]. Overall, L-selectin favors interactions that allow both leukocytes [18] and metastatic tumor cells [19,22] leave the bloodstream, come into contact with activated endothelial cells and start the rolling process [15,18,23]. In addition to its main role in the process of initial capture of immune cells and cell adhesion, it is also relevant in acute and chronic inflammatory processes [10].
P-selectin has also been suggested to play a role in cancer [24,25] promoting the spread of cancer cells by facilitating their migration using CAM’s migrations system. The protein is encoded by SELP gene, located on chromosome 1 (1q24.2). It plays a role in the process of blood clotting and inflammation and is found on the surface of platelets and endothelial cells [26,27]. In addition, P-selectin has been shown to be involved in the recruitment of immune cells to tumor site [14,28]. One of the most important ligands of P-selectin is PSGL-1, a protein encoded by SELPLG gene, also located on chromosome 12 (12q24.11). It is expressed on the surface of various types of immune cells including T cells, B cells, and neutrophils. Recent studies [29] have shown that PSGL-1 may promote cancer progression by promoting the adhesion and migration of immune cells to the tumor microenvironment. This can lead to the recruitment of pro-tumor immune cells and the suppression of anti-tumor immune responses, ultimately promoting tumor growth and metastasis [30]. PSGL-1 has been found to be upregulated in various types of cancer, including breast, lung, and colorectal cancer. This upregulation is associated with more aggressive tumors and poorer prognosis [30]. In addition, targeting PSGL-1 has been considered a potential approach for cancer immunotherapy [31,32,33,34,35,36,37,38].
E-selectin is also expressed on endothelial cells and is involved in the recruitment of immune cells to sites of inflammation [12]. It is encoded by SELE gene, located on the long arm of chromosome 1 (1q24.2). E-selectin plays an important role in the process of inflammation by facilitating the adhesion and migration of immune cells to sites of inflammation [39]. Recent studies have shown that E-selectin may also be involved in cancer, since it was found to be upregulated in various types of cancer including breast [24,40], lung [41], and pancreatic cancer [42], and its expression levels are associated with more aggressive tumors and poorer prognosis in cancer patients [43].
Because of their relevant role in the migration of immune system and metastatic tumor cells, the study of SNPs in SELL, SELP, SELE and SELPLG is fundamental for the identification of potential biomarkers of susceptibility, diagnosis, prognosis and even possible therapeutic targets for cancer. The objective of this study was to understand the alterations caused by the presence of polymorphisms in the DNA structure of these genes and the consequent alterations in the morphology and function of the corresponding proteins through bioinformatics tools.
2. Materials and Methods
The nsSNPs were retrieved from dbSNP of NCBI (https://www.ncbi.nlm.nih.gov/snp/). The selection was performed considering a minor allele frequency (MAF) between 0.1 and 1.0. The gene sequences and FASTA sequences of the proteins were obtained from Genome Browser (https://genome.ucsc.edu/cgi-bin/hgGateway) and Universal Protein Resource – UniProt [44] (https://www.uniprot.org/; SELL ID: P14151, SELE ID: P16581, SELP ID: P16109 and SELPLG ID: Q14242), respectively.
Analysis of gene sequence
All nsSNPs with MAF>0.1 were analyzed by PredictSNP2.0 [45] (https://loschmidt.chemi.muni.cz/predictsnp2/). This software consists of 5 tools that analyze the impacts of nucleotide substitutions on DNA: CADD [46] associates the presence of SNPs with deleterious functions (insertion or deletion) in the human genome; DANN [47] uses Neutral Network for deleterious gene sequence annotations; FATHMM [48] predicts possible functional consequences caused by the presence of SNPs in coding and non-coding regions; FunSeq2 [49] prioritizes the analysis of somatic alterations related to the appearance of neoplasms and GWAVA [50] evaluates the functional impact of changes in non-coding regions. In addition, we used PredictSNP2.0 [45] to complement the computational analyses with experimental annotations from eight databases (ClinVar, dbSNP, Ensembl Genome Browser, GenBank, HaploReg, OMIM, RegulomeDB, and UCSC Genome Browser), enabling correlation of the data obtained in silica with the existing literature available in these databases.
Analysis of missense SNPs with amino acid change
We employed PredictSNP1.0 [51] (https://loschmidt.chemi.muni.cz/predictsnp1/) to perform morphofunctional analysis of proteins with amino acids altered by the presence of missense SNPs. PredictSNP1.0 [51] comprises eight tools: SIFT [52] which performs an estimation of the effects of amino acid substitution on protein function based on homology and the chemical characteristics of the amino acids; PolyPhen-1 [53] and PolyPhen-2 [54] which assess the impacts on protein structure and function by empirical methods of analysis and also by comparison of physical properties of the molecules; MAP [55] which evaluates the physicochemical variations of the protein; PhD-SNP [56] uses Support Vector Machine (SVM) methodology for protein structure and sequence analysis; SNAP [57] evaluates changes in the secondary structure of the protein, as well as compares solvent accessibility in case of amino acid changes by Neural Network methodology; PANTHER [58] evaluates protein function; and nsSNPAnalyzer [59] which uses Random Forest methodology with sequence alignment and 3D structure to assess phenotypic impacts. Complementarily, MuPRO [60] (http://mupro.proteomics.ics.uci.edu/) was used to evaluate the stability of proteins.
3. Results
3.1. L-selectin (SELL gene)
We ere able to retrieve 216 polymorphisms with MAF>0.1 from dbSNP, homever, after excluding the repeats and the merged records, there were 55 unique polymorphisms and a total of 80 nucleotide alterations. The SNPs were classified as: upstream variant (4); intronic (46); 3’UTR (2); and exonic (3, including 2 nonsynonymous and 1 synonymous).
All 80 alterations were evaluated by PredictSNP2.0 tool. A total 38 variants (47.5%) were considered neutral in all tools in the consensus. Five (6.3%) SNPs were considered deleterious in at least 4 of the 6 tools as shown in Table 1.
rs4987360 is a intronic SNP wich alters an adenine to a guanine (A/G). This alteration was considered deleterious in 4 (66.7%) tools suggesting possible structural and functional DNA changes. However, FATHMM wasn’t able to evaluated this alteration. Another polimorphic variant, rs2229569, is an exonic SNP which promotes the exange of a guanine for an adenine (G/A) or for a timine (G/T). Both alterations can promote amino acid changes in the corresponding protein. The G/A alteration was considered deleterious in 4 (66.7%) PredictSNP2.0 tools suggesting that it is capable of altering the structure and folding of DNA. In addition, it promotes the aminoacid switch of a proline to a serine in position 213 (P213S). P213S alteration was considered deleterious by MAPP tool, sugesting physicalchemical alterations in protein structure. Decreased strability was observed (ΔΔG=-0.6147; MuPRO), but there was no significant alterations in the pattern of ligation with adjacents amino acids (Dynamut2.0). The G/T alteration was considered deleterious in 3 (50.0%) PredictSNP2.0 tools. This variant promotes amino acid switch of a proline to a threonine in position 213 (P213T). P213T alteration was considered deleterious by MAPP, PhdSNP and SIFT tools (Table 2) suggesting physicalchemical, structural and functional alterations in the corresponding protein. Decreased stability (ΔΔG=-0.6483; MuPRO) and rigidification of the structure (ΔΔSvibENCoM: -0.048 kcal.mol-1.K-1; Dynamut2.0) were observed, but this change did not produce significant alterations in ligation patterns with adjacent amino acids.
rs4987301 is a intronic SNP with two possible nucleotide alterations (G/A and G/T). Both alterations were considered deleterious for all PredictSNP2.0 tools (Table 1) except GWAVA, suggesting an importante role in DNA modification considering structure, pattern of interation with adjacents nucleotides and function. In a similar way, rs2205849 is a upstream SNP that promotes alteration from a timine to a citosine in DNA structure and was considered deleterious in all PredictSNP2.0 tools (Table 1).
rs1131498 is a exonic that promotes the exchange of an adenine for a guanine in the DNA structure (A/G). This alteration was considered deleterious in CADD, suggesting that it has harmful effects on the human genome (PredictSNP2.0 tools). The consequence of A/G alteration in protein is a change of a phenylalanine to a leucine in 193 position (F193L). F193L alteration was found to be neutral in all PredictSNP1.0 tools (Table 2), however, decreased stability (ΔΔG= : -0.3179; MuPRO) and increase in molecule flexibility (ΔΔSvibENCoM: 0.005 kcal.mol-1.K-1; Dynamut2.0) were observed. Furthermore, a different pattern of interaction with adjacent amino acids was observed with loss of a hydrophobic contact, probably explaining the changes in flexibility as shown in Figure 1A (Dynamut2.0).
3.2. P-selectin (SELP gene)
We retrieved 245 polymorphisms that presented MAF>0.1 from dbSNP. After removing the repeats, 88 unique polymorphisms and a total of 144 nucleotide changes were left. The SNPs were classified as: upstream variant (4); 3’UTR (1); intronic (76) and exonic (6, including 3 synonymous and 3 nonsynonymous).
All 144 alterations were evaluated by PredictSNP2.0 tool. A total 108 (75.0%) were considered neutral in all tools in the consensus, but 6 (4.2%) were identified as deleterious in at least 4 of the 5 tools as presented in Table 1.
rs3917777 (T/A, T/C and T/G), rs2205894 (T/A and T/G) and rs2205893 (T/A and T/G) are intronic SNPs and were considered deleterious in 4 out 5 PredictSNP2.0 tools (Table 1) suggesting a possible role in DNA structural and functional alterations. rs6127 (C/T; D541N) and rs6131 (C/T; S331N) were identified as neutral in all PredictSNP2.0 tools (Table 1). However, these are exonic SNPs with amino acid changes in protein structure which can cause morphofunctional alterations. Therefore we proceeded with PredictSNP1.0, MuPRO and Dynamut evaluation. In fact rs6127 (C/T; D541N) and rs6131 (C/T; S331N) were considered neutral in all PredictSNP1.0 tools (Table 2) suggesting no alteration in structure or function of E-selectin protein, however these SNPs cause decrease in protein stability (ΔΔG= -0.7148 and ΔΔG= -0.1887, respectively; MuPRO)
Also, rs6127 (C/T; D541N) promotes a change in the binding pattern with adjacent amino acids due to the disappearance of a hydrogen bond (Figure 1B) correlating with decreased protein stability and flexibility of the protein structure (ΔΔSVibENCoM: 0.643 kcal.mol-1.K-1; Dynamut).
3.3. PSGL-1 (SELPLG gene)
Ninety-five SNPs with MAF>0.1 were available in the dbSNP database. However, after removing the duplicate records, only 27 SNPs and 48 nucleotide alterations were observed. The SNPs were classified as follows: downstream variant (2), upstream variant (4), 3’UTR (1), intronic (35) and exonic (3).
All 48 alterations were evaluated in PredictSNP2.0 tool. Seventeen (35.4%) alterations were considered neutral in all tolls and only rs7138370 was considered deleterious in 4 out of 6 tools, suggesting its possible role in DNA structural and functional alterations (Table 1).
In addition, we studied the exonic SNPs (Table 2): rs2228315 (C/T; M62I), rs7300972 (T/C; M274V) and rs201851784 (A/G; V137A). rs2228315 (C/T; M62I) was considered neutral by PredictSNP2.0, but it was found deleterious by PolyPhen-1 tool (PredictSNP1.0) and MuPRO analysis indicated that this SNP can decrease protein stability (ΔΔG= -0.5872). Also, it may cause rigidification of protein structure (ΔΔSVibENCoM: -0.133 kcal.mol-1.K-1; Dynamut) and alterations in amino acid binding patterns showing a weaker water mediated hydrogen bonding (Figure 1C). rs7300972 was considered deleterious by FUNSEQ and GWAVA tools (PredictSNP2.0 – Table 1), suggesting a possible role in somatic alterations and modifications in DNA structure. This SNP was considered deleterious by MAPP and SIFT tools (Table 2), that disclosed putative structural and physicalchemistry alterations. rs201851784 (A/G; V137A) was considered neutral by all tools employed (PredictSNP1.0 – Table 2 and PredictSNP2.0 – Table 1).
3.4. E-selectin (SELE gene)
Data obtained in dbSNP indicate 97 polymorphisms with MAF between 0.1 and 1.0. However, some records were merged and, after deleting the repeats, we observed 32 unique polymorphisms and 58 nucleotide alterations. The SNPs were classified according to position in DNA: upstream variant (5); downstream variant (3); 3’UTR (2); intronic (20) and exonic (2, including 1 synonymous and 1 nonsynonymous). All 58 nucleotide alterations were evaluated by PredictSNP2.0 tool and 82.7% (n=48) were considered neutral in all tools, but one (rs1534904) was considered deleterious in 4 out of 5 tools as presented in table 01.
rs1534904 is an intronic SNP that has two possible nucleotide alterations: T/A and T/G. The T/A exchange was considered deleterious in 4 (66,7%) of the tools evaluated (Table 1) suggesting possible structural, functional and damage control pathway alterations in DNA. However, FATHMM tool was unable to evaluate this alteration. The T/G alteration was considered deleterious in 2 (33,4%) of the tools employed (Table 1) suggesting a deleterious effect to human genome.
rs5368 is the only exonic SNP with MAF>0.1 recorded for the SELE gene in dbSNP database. It is an exchange of a guanine for an adenine in the DNA structure which was considered deleterious in 2 out of 6 PredictSNP2.0 tools suggesting a possible role in DNA damage control pathway and a functional impact. This polymorphism promotes the exchange of a histidine by a tyrosine at position 468 (H468Y). It was evaluated by PredictSNP1.0 (Table 2) and was considered deleterious only by SNAP tool, indicating a possible alteration in the secondary structure of the E-selectin protein (Table 01). This exchange is not located in the region of post-translational modifications according to the MutPred2 tool. Increased protein stability (ΔΔG=-0.1524, MuPRO) and increased molecule flexibility (ΔΔSVibENCoM: 0.024 kcal.mol-1.K-1; Dynamut) were observed. Alteration in hydrogen binds with adjacent amino acids were observed (Figure 1D).
4. Discussion
For a long time, selectins were thought to be unique to the immune system or correlated cells. However, more recent solid evidence has demonstrated their overexpression in tumor cells, suggesting that these CAMs play an important role in metastatic pathways [61]. Using a wide repertoire of bioinformatics tools, we identified a series of SNPs that, due to their MAF>0.1, may be significantly present in the population and correlate with several diseases, including cancer. Our bioinformatic analysis demonstrated that rs2229569, rs1131498, rs4987360, rs4987301 and rs2205849 of SELL gene; rs3917777, rs2205894 and rs2205893 of SELP gene; rs7138370, rs7300972 and rs2228315 of SELPLG gene and; rs1534904 and rs5368 of SELE gene may be promising biomarkers of diseases, especially in cancer patients, considering the role of selectins (L-selectin, P-selectin and E-selectin) and PSGL-1 in these conditions.
L-selectin (SELL) expression has been related to several types of cancers such as endometrial [62,63], breast [64] and thyroid [20]. An in-silico study [64] using the TCGA and On-comine databases found higher expression of SELL in tumor tissues, suggesting that L-selectin could be a biomarker of inflammatory microenvironment. In addition, the authors observed higher SELL expression in breast cancer patients with better outcome. Kobawala et al. [20] analyzed 150 patients with thyroid nodules (83 papillary thyroid carcinomas - PTC and 67 benign nodules) by ELISA and immunohistochemistry techniques and observed a higher protein expression of L-selectin in cells and higher serum levels in PTC patients compared to benign thyroid diseases, suggesting a possible role of this adhesion molecule in the development of thyroid cancer.
Despite several studies reporting gene and protein expression of L-selectin, the literature is still scarce with regard to the study of its polymorphisms in cancer. We demonstrated that rs2229569 (exonic; G/A - P213S and G/T - P213T) may alter the structure and folding of DNA and promote physical-chemical alterations in protein structure with decreased stability and possible protein functional alterations. Similarly, rs1131498 was shown to be able to modify binding patterns with adjacent amino acids, thus altering protein structure at the molecular and atomic level. We also did not find reports in the literature of other possible cancer biomarkers, such as the intronic polymorphisms rs4987360 (A/G), rs4987301 (G/A and G/T) and rs2205849 (T/C), which were considered deleterious by all the tools that we used.
P-selectin (SELP) is mainly expressed on the surface of activated endothelial cells and platelets [11,26]. Its expression upregulation is correlated with the pathogenesis of various diseases, including atherosclerosis [65], thrombosis [66], diabetes [67] and cancer [14,24,25,28,68]. Because P-selectin has been shown to promote the formation of cancer metastases by facilitating the adhesion and migration of tumor cells [24,25], this molecule has emerged as a potential therapeutic strategy for the treatment of various conditions. Some studies suggested that SNPs of SELP gene could be diagnostic biomarkers of head and neck [28] and pancreatic cancer [68]. Our analysis also identified rs3917777 (T/A, T/C and T/G), rs2205894 (T/A and T/G) and rs2205893 (T/A and T/G) as promising biomarkers that have not yet been investigated in cancer patients.
PSGL-1 (P-selectin glycoprotein ligand-1) plays a key role in mediating leukocyte adhesion to activated endothelial cells and platelets, as well as facilitating leukocyte rolling and migration into sites of inflamation [33] and has been widely studied because of its role as an immune checkpoint and its promising role in immune checkpoint landscaping [32,33]. Monoclonal antibodies against PSGL-1 have been shown to inhibit tumor growth and metastasis in preclinical models of cancer [29,30,37]. However, studies on its polymorphisms and its possible clinical use are still scarce. Our data indicate that both rs7138370 and rs7300972 could promote DNA structural and functional alteration; rs2228315 (C/T; M62I) also may promote modifications in DNA and, additionally, modify structure, function, stability, rigidification and interaction with adjacent amino acids, making these SNPs interesting candidates for biomarkers. PSGL1 polymorphisms can help identify response patterns to immunological therapies to which this molecule is targeted and improve the quality of treatment offered to cancer patients.
Expression of E-selectin at significantly higher levels has been related to several types of cancer such as colorectal [14], gastric [69] and breast [24,40] was correlated with decrease risk of hospitalization or need for respiratory support/death in COVID-19 cases [70]. Na Li et al [71], in an in-silico study employing TCGA and the GEPIA server, found higher gene expression in tumor samples when compared to healthy samples, and in of lymph node metastasis in colorectal cancer [71]. Furthermore, when antitumor drugs were applied, gene expression levels were reduced, suggesting a possible role of E-selectin as an oncogene. Targeting E-selectin has emerged as a potential therapeutic strategy for the treatment of cancer [72,73,74]. Several approaches have been developed to target E-selectin, including monoclonal antibodies and small molecule inhibitors and have shown promise in preclinical studies by reducing tumor growth and metastasis [75,76,77,78,79]. SELE polymorphisms have been reported as possible risk indicators for a number of medical conditions such as hypertension in cases of occupational stress [80], risk of coronary artery disease [81], enlargement of renal cyst in patients with polycystic kidney disease [82], type 2 diabetes [83] and subclinical atherosclerosis and increased platelet activity in systemic lupus erythematosus [84]. We demonstrated that rs1534904 (T/A and T/G) could provoke important alterations in DNA. This polymorphism has not yet been reported in studies evaluating disease conditions. We also showed that rs5368 (H468Y) may alter the secondary structure of the E-selectin protein and is capable of increase protein stability and flexibility. Zakariya BF et al [40] found CT heterozygous genotype frequency significantly higher in breast cancer patients, confirming the importance of SELE polymorphisms in cancer risk prediction.
5. Conclusions
In conclusion, using a series of robust bioinformatics tools, we investigated the three selectins and an important selectin ligand. We demonstrated that a series of relevant polymorphisms that present relevant minor allele frequency in the population might become important biomarkers and deserve further investigation in cancer patients.
Author Contributions
Conceptualization, LTR, KCP, EST, NEB and LSW; methodology, LTR, DZV and EST; validation, LTR and DZV; formal analysis, LTR and KCP; investigation, LTR and DZV; data curation, LTR; writing—original draft preparation, LTR, KCP, DZV and LSW; writing—review and editing, LTR, NEB and LSW; visualization, LTR, KCP, NEB and LSW; supervision, NEB and LSW; project administration, LTR and DZV; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
The authors thank the group of the Laboratory of Cancer Molecular Genetics (GEMOCA) of the School of Medical Sciences (FCM-UNICAMP) for technical support. LSW is a Category 1 Research Fellow at the National Council for Scientific and Technological Development (CNPq).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, X.; Tomso, D.J.; Liu, X.; Bell, D.A. Single nucleotide polymorphism in transcriptional regulatory regions and expression of environmentally responsive genes. Toxicol Appl Pharmacol 2005, 207, 84–90. [Google Scholar] [CrossRef]
- Giacomini, K.M.; Brett, C.M.; Altman, R.B.; Benowitz, N.L.; Dolan, M.E.; Flockhart, D.A.; Johnson, J.A.; Hayes, D.F.; Klein, T.; Krauss, R.M.; et al. The pharmacogenetics research network: from SNP discovery to clinical drug response. Clin Pharmacol Ther 2007, 81, 328–345. [Google Scholar] [CrossRef]
- Cargill, M.; Altshuler, D.; Ireland, J.; Sklar, P.; Ardlie, K.; Patil, N.; Shaw, N.; Lane, C.R.; Lim, E.P.; Kalyanaraman, N.; et al. Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat Genet 1999, 22, 231–238. [Google Scholar] [CrossRef]
- Teng, S.; Srivastava, A.K.; Schwartz, C.E.; Alexov, E.; Wang, L. Structural assessment of the effects of amino acid substitutions on protein stability and protein protein interaction. Int J Comput Biol Drug Des 2010, 3, 334–349. [Google Scholar] [CrossRef]
- Kucukkal, T.G.; Petukh, M.; Li, L.; Alexov, E. Structural and physico-chemical effects of disease and non-disease nsSNPs on proteins. Curr Opin Struct Biol 2015, 32, 18–24. [Google Scholar] [CrossRef]
- Buroker, N.E. Regulatory SNPs and transcriptional factor binding sites in ADRBK1, AKT3, ATF3, DIO2, TBXA2R and VEGFA. Transcription 2014, 5, e964559. [Google Scholar] [CrossRef]
- Birney, E.; Stamatoyannopoulos, J.A.; Dutta, A.; Guigó, R.; Gingeras, T.R.; Margulies, E.H.; Weng, Z.; Snyder, M.; Dermitzakis, E.T.; Stamatoyannopoulos, J.A.; et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 2007, 447, 799–816. [Google Scholar] [CrossRef]
- Elangbam, C.S.; Qualls, C.W., Jr.; Dahlgren, R.R. Cell adhesion molecules--update. Veterinary pathology 1997, 34, 61–73. [Google Scholar] [CrossRef]
- Gupta, G.S. L-Selectin (CD62L) and Its Ligands. In Animal Lectins: Form, Function and Clinical Applications; Springer Vienna: Vienna, 2012; pp. 553–574. [Google Scholar]
- Raffler, N.A.; Rivera-Nieves, J.; Ley, K. L-selectin in inflammation, infection and immunity. Drug Discovery Today: Therapeutic Strategies 2005, 2, 213–220. [Google Scholar] [CrossRef]
- da Costa Martins, P.; van den Berk, N.; Ulfman, L.H.; Koenderman, L.; Hordijk, P.L.; Zwaginga, J.J. Platelet-monocyte complexes support monocyte adhesion to endothelium by enhancing secondary tethering and cluster formation. Arteriosclerosis, thrombosis, and vascular biology 2004, 24, 193–199. [Google Scholar] [CrossRef]
- Rosen, S.D.; Bertozzi, C.R. The selectins and their ligands. Current opinion in cell biology 1994, 6, 663–673. [Google Scholar] [CrossRef]
- Laubli, H.; Borsig, L. Selectins promote tumor metastasis. Semin Cancer Biol 2010, 20, 169–177. [Google Scholar] [CrossRef]
- Korniluk, A.; Kaminska, J.; Kiszlo, P.; Kemona, H.; Dymicka-Piekarska, V. Lectin adhesion proteins (P-, L- and E-selectins) as biomarkers in colorectal cancer. Biomarkers 2017, 22, 629–634. [Google Scholar] [CrossRef]
- Rosen, S.D. Ligands for L-selectin: homing, inflammation, and beyond. Annual review of immunology 2004, 22, 129–156. [Google Scholar] [CrossRef]
- McEver, R.P. Selectins: initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovascular research 2015, 107, 331–339. [Google Scholar] [CrossRef]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling. Frontiers in immunology 2019, 10, 1068. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Choudhary, D.; Hegde, P.; Voznesensky, O.; Choudhary, S.; Kopsiaftis, S.; Claffey, K.P.; Pilbeam, C.C.; Taylor, J.A. , 3rd. Increased expression of L-selectin (CD62L) in high-grade urothelial carcinoma: A potential marker for metastatic disease. Urologic oncology 2015, 33, 387–e317. [Google Scholar] [CrossRef]
- Kobawala, T.P.; Trivedi, T.I.; Gajjar, K.K.; Patel, D.H.; Patel, G.H.; Ghosh, N.R. Significance of TNF-alpha and the Adhesion Molecules: L-Selectin and VCAM-1 in Papillary Thyroid Carcinoma. J Thyroid Res 2016, 2016, 8143695. [Google Scholar] [CrossRef]
- Muzza, M.; Degl'Innocenti, D.; Colombo, C.; Perrino, M.; Ravasi, E.; Rossi, S.; Cirello, V.; Beck-Peccoz, P.; Borrello, M.G.; Fugazzola, L. The tight relationship between papillary thyroid cancer, autoimmunity and inflammation: clinical and molecular studies. Clinical endocrinology 2010, 72, 702–708. [Google Scholar] [CrossRef]
- Reymond, N.; d'Agua, B.B.; Ridley, A.J. Crossing the endothelial barrier during metastasis. Nature reviews. Cancer 2013, 13, 858–870. [Google Scholar] [CrossRef]
- Kobayashi, H.; Boelte, K.C.; Lin, P.C. Endothelial cell adhesion molecules and cancer progression. Current medicinal chemistry 2007, 14, 377–386. [Google Scholar] [CrossRef]
- Nasti, T.H.; Bullard, D.C.; Yusuf, N. P-selectin enhances growth and metastasis of mouse mammary tumors by promoting regulatory T cell infiltration into the tumors. Life Sci 2015, 131, 11–18. [Google Scholar] [CrossRef]
- Richter, U.; Schröder, C.; Wicklein, D.; Lange, T.; Geleff, S.; Dippel, V.; Schumacher, U.; Klutmann, S. Adhesion of small cell lung cancer cells to E- and P-selectin under physiological flow conditions: implications for metastasis formation. Histochem Cell Biol 2011, 135, 499–512. [Google Scholar] [CrossRef]
- Celi, A.; Pellegrini, G.; Lorenzet, R.; De Blasi, A.; Ready, N.; Furie, B.C.; Furie, B. P-selectin induces the expression of tissue factor on monocytes. Proc Natl Acad Sci U S A 1994, 91, 8767–8771. [Google Scholar] [CrossRef]
- Schmalbach, B.; Stepanow, O.; Jochens, A.; Riedel, C.; Deuschl, G.; Kuhlenbäumer, G. Determinants of platelet-leukocyte aggregation and platelet activation in stroke. Cerebrovasc Dis 2015, 39, 176–180. [Google Scholar] [CrossRef]
- Powrózek, T.; Mlak, R.; Brzozowska, A.; Mazurek, M.; Gołębiowski, P.; Małecka-Massalska, T. Relationship Between -2028 C/T SELP Gene Polymorphism, Concentration of Plasma P-Selectin and Risk of Malnutrition in Head and Neck Cancer Patients. Pathol Oncol Res 2019, 25, 741–749. [Google Scholar] [CrossRef]
- DeRogatis, J.M.; Viramontes, K.M.; Neubert, E.N.; Tinoco, R. PSGL-1 Immune Checkpoint Inhibition for CD4(+) T Cell Cancer Immunotherapy. Front Immunol 2021, 12, 636238. [Google Scholar] [CrossRef]
- Hoos, A.; Protsyuk, D.; Borsig, L. Metastatic growth progression caused by PSGL-1-mediated recruitment of monocytes to metastatic sites. Cancer Res 2014, 74, 695–704. [Google Scholar] [CrossRef]
- Agrati, C.; Sacchi, A.; Tartaglia, E.; Vergori, A.; Gagliardini, R.; Scarabello, A.; Bibas, M. The Role of P-Selectin in COVID-19 Coagulopathy: An Updated Review. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- DeRogatis, J.M.; Viramontes, K.M.; Neubert, E.N.; Henriquez, M.L.; Guerrero-Juarez, C.F.; Tinoco, R. Targeting the PSGL-1 Immune Checkpoint Promotes Immunity to PD-1-Resistant Melanoma. Cancer Immunol Res 2022, 10, 612–625. [Google Scholar] [CrossRef]
- Tinoco, R.; Otero, D.C.; Takahashi, A.A.; Bradley, L.M. PSGL-1: A New Player in the Immune Checkpoint Landscape. Trends Immunol 2017, 38, 323–335. [Google Scholar] [CrossRef]
- Schmitt, C.; Abt, M.; Ciorciaro, C.; Kling, D.; Jamois, C.; Schick, E.; Solier, C.; Benghozi, R.; Gaudreault, J. First-in-Man Study With Inclacumab, a Human Monoclonal Antibody Against P-selectin. J Cardiovasc Pharmacol 2015, 65, 611–619. [Google Scholar] [CrossRef]
- Gutsaeva, D.R.; Parkerson, J.B.; Yerigenahally, S.D.; Kurz, J.C.; Schaub, R.G.; Ikuta, T.; Head, C.A. Inhibition of cell adhesion by anti-P-selectin aptamer: a new potential therapeutic agent for sickle cell disease. Blood 2011, 117, 727–735. [Google Scholar] [CrossRef]
- Mertens, P.; Maes, A.; Nuyts, J.; Belmans, A.; Desmet, W.; Esplugas, E.; Charlier, F.; Figueras, J.; Sambuceti, G.; Schwaiger, M.; et al. Recombinant P-selectin glycoprotein ligand-immunoglobulin, a P-selectin antagonist, as an adjunct to thrombolysis in acute myocardial infarction. The P-Selectin Antagonist Limiting Myonecrosis (PSALM) trial. Am Heart J 2006, 152, 125–e121. [Google Scholar] [CrossRef]
- Patel, M.S.; Miranda-Nieves, D.; Chen, J.; Haller, C.A.; Chaikof, E.L. Targeting P-selectin glycoprotein ligand-1/P-selectin interactions as a novel therapy for metabolic syndrome. Transl Res 2017, 183, 1–13. [Google Scholar] [CrossRef]
- Muz, B.; Azab, F.; de la Puente, P.; Rollins, S.; Alvarez, R.; Kawar, Z.; Azab, A.K. Inhibition of P-Selectin and PSGL-1 Using Humanized Monoclonal Antibodies Increases the Sensitivity of Multiple Myeloma Cells to Bortezomib. Biomed Res Int 2015, 2015, 417586. [Google Scholar] [CrossRef]
- Aleisa, F.A.; Sakashita, K.; Lee, J.M.; AbuSamra, D.B.; Al Alwan, B.; Nozue, S.; Tehseen, M.; Hamdan, S.M.; Habuchi, S.; Kusakabe, T.; et al. Functional binding of E-selectin to its ligands is enhanced by structural features beyond its lectin domain. J Biol Chem 2020, 295, 3719–3733. [Google Scholar] [CrossRef]
- Zakariya, B.F.; Almohaidi, A.M.S.; Şimşek, S.A.; Al-Waysi, S.A.; Al-Dabbagh, W.H.; Kamal, A.M. The relationship of E-selectin singlenucleotide polymorphisms with breast cancer in Iraqi Arab women. Genomics Inform 2022, 20, e42. [Google Scholar] [CrossRef]
- Kiziltunc Ozmen, H.; Simsek, M. Serum IL-23, E-selectin and sICAM levels in non-small cell lung cancer patients before and after radiotherapy. J Int Med Res 2020, 48, 300060520923493. [Google Scholar] [CrossRef]
- Guerrero, P.E.; Miró, L.; Wong, B.S.; Massaguer, A.; Martínez-Bosch, N.; Llorens, R.; Navarro, P.; Konstantopoulos, K.; Llop, E.; Peracaula, R. Knockdown of α2,3-Sialyltransferases Impairs Pancreatic Cancer Cell Migration, Invasion and E-selectin-Dependent Adhesion. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Tanio, M.; Muramoto, A.; Hoshino, H.; Murahashi, M.; Imamura, Y.; Yokoyama, O.; Kobayashi, M. Expression of functional E-selectin ligands on the plasma membrane of carcinoma cells correlates with poor prognosis in clear cell renal cell carcinoma. Urol Oncol 2021, 39, 302–e309. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.U. UniProt: the Universal Protein Knowledgebase in 2023. Nucleic Acids Research 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Bendl, J.; Musil, M.; Štourač, J.; Zendulka, J.; Damborský, J.; Brezovský, J. PredictSNP2: A Unified Platform for Accurately Evaluating SNP Effects by Exploiting the Different Characteristics of Variants in Distinct Genomic Regions. PLoS Comput Biol 2016, 12, e1004962. [Google Scholar] [CrossRef]
- Kircher, M.; Witten, D.M.; Jain, P.; O'Roak, B.J.; Cooper, G.M.; Shendure, J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet 2014, 46, 310–315. [Google Scholar] [CrossRef]
- Quang, D.; Chen, Y.; Xie, X. DANN: a deep learning approach for annotating the pathogenicity of genetic variants. Bioinformatics 2015, 31, 761–763. [Google Scholar] [CrossRef]
- Shihab, H.A.; Rogers, M.F.; Gough, J.; Mort, M.; Cooper, D.N.; Day, I.N.; Gaunt, T.R.; Campbell, C. An integrative approach to predicting the functional effects of non-coding and coding sequence variation. Bioinformatics 2015, 31, 1536–1543. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, Z.; Lou, S.; Bedford, J.; Mu, X.J.; Yip, K.Y.; Khurana, E.; Gerstein, M. FunSeq2: a framework for prioritizing noncoding regulatory variants in cancer. Genome Biol 2014, 15, 480. [Google Scholar] [CrossRef]
- Ritchie, G.R.; Dunham, I.; Zeggini, E.; Flicek, P. Functional annotation of noncoding sequence variants. Nat Methods 2014, 11, 294–296. [Google Scholar] [CrossRef]
- Bendl, J.; Stourac, J.; Salanda, O.; Pavelka, A.; Wieben, E.D.; Zendulka, J.; Brezovsky, J.; Damborsky, J. PredictSNP: robust and accurate consensus classifier for prediction of disease-related mutations. PLoS Comput Biol 2014, 10, e1003440. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.C.; Henikoff, S. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [PubMed]
- Ramensky, V.; Bork, P.; Sunyaev, S. Human non-synonymous SNPs: server and survey. Nucleic Acids Res 2002, 30, 3894–3900. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.A.; Sidow, A. Physicochemical constraint violation by missense substitutions mediates impairment of protein function and disease severity. Genome Res 2005, 15, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, E.; Calabrese, R.; Casadio, R. Predicting the insurgence of human genetic diseases associated to single point protein mutations with support vector machines and evolutionary information. Bioinformatics 2006, 22, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Bromberg, Y.; Rost, B. SNAP: predict effect of non-synonymous polymorphisms on function. Nucleic Acids Res 2007, 35, 3823–3835. [Google Scholar] [CrossRef] [PubMed]
- Brunham, L.R.; Singaraja, R.R.; Pape, T.D.; Kejariwal, A.; Thomas, P.D.; Hayden, M.R. Accurate prediction of the functional significance of single nucleotide polymorphisms and mutations in the ABCA1 gene. PLoS Genet 2005, 1, e83. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Zhou, M.; Cui, Y. nsSNPAnalyzer: identifying disease-associated nonsynonymous single nucleotide polymorphisms. Nucleic Acids Res 2005, 33, W480–482. [Google Scholar] [CrossRef]
- Cheng, J.; Randall, A.; Baldi, P. Prediction of protein stability changes for single-site mutations using support vector machines. Proteins 2006, 62, 1125–1132. [Google Scholar] [CrossRef]
- Kawaguchi, T. Cancer metastasis: characterization and identification of the behavior of metastatic tumor cells and the cell adhesion molecules, including carbohydrates. Curr Drug Targets Cardiovasc Haematol Disord 2005, 5, 39–64. [Google Scholar] [CrossRef]
- Karpathiou, G.; Sramek, V.; Dagher, S.; Mobarki, M.; Dridi, M.; Picot, T.; Chauleur, C.; Peoc'h, M. Peripheral node addressin, a ligand for L-selectin is found in tumor cells and in high endothelial venules in endometrial cancer. Pathol Res Pract 2022, 233, 153888. [Google Scholar] [CrossRef] [PubMed]
- Jóźwik, M.; Okungbowa, O.E.; Lipska, A.; Jóźwik, M.; Smoktunowicz, M.; Semczuk, A.; Jóźwik, M.; Radziwon, P. Surface antigen expression on peripheral blood monocytes in women with gynecologic malignancies. BMC Cancer 2015, 15, 129. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Arora, M.; Singh, J.; Chauhan, S.S.; Kumar, S.; Chopra, A. L-Selectin expression is associated with inflammatory microenvironment and favourable prognosis in breast cancer. 3 Biotech 2021, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Changfu, W.; Linyun, L.; Bing, M.; Xiaohui, H. Correlations of platelet-leukocyte aggregates with P-selectin S290N and P-selectin glycoprotein ligand-1 M62I genetic polymorphisms in patients with acute ischemic stroke. J Neurol Sci 2016, 367, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Cosan, F.; Oku, B.; Gedar Totuk, O.M.; Abaci, N.; Ustek, D.; Diz Kucukkaya, R.; Gul, A. The association between P selectin glycoprotein ligand 1 gene variable number of tandem repeats polymorphism and risk of thrombosis in Behçet's disease. Int J Rheum Dis 2018, 21, 2175–2179. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Singh, J.; Kapoor, R.; Kaur, M. Association of SELP Polymorphisms with Soluble P-Selectin Levels and Vascular Risk in Patients with Type 2 Diabetes Mellitus: A Case-Control Study. Biochem Genet 2019, 57, 73–97. [Google Scholar] [CrossRef] [PubMed]
- Avan, A.; Avan, A.; Le Large, T.Y.; Mambrini, A.; Funel, N.; Maftouh, M.; Ghayour-Mobarhan, M.; Cantore, M.; Boggi, U.; Peters, G.J.; et al. AKT1 and SELP polymorphisms predict the risk of developing cachexia in pancreatic cancer patients. PLoS One 2014, 9, e108057. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.; Freitas, R.; Ferreira, D.; Soares, J.; Azevedo, R.; Gaiteiro, C.; Peixoto, A.; Oliveira, S.; Cotton, S.; Relvas-Santos, M.; et al. Nucleolin-Sle A Glycoforms as E-Selectin Ligands and Potentially Targetable Biomarkers at the Cell Surface of Gastric Cancer Cells. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Palmos, A.B.; Millischer, V.; Menon, D.K.; Nicholson, T.R.; Taams, L.S.; Michael, B.; Sunderland, G.; Griffiths, M.J.; Hübel, C.; Breen, G. Proteome-wide Mendelian randomization identifies causal links between blood proteins and severe COVID-19. PLoS Genet 2022, 18, e1010042. [Google Scholar] [CrossRef]
- Li, N.; Xiao, H.; Shen, J.; Qiao, X.; Zhang, F.; Zhang, W.; Gao, Y.; Liu, Y.D. SELE gene as a characteristic prognostic biomarker of colorectal cancer. J Int Med Res 2021, 49, 3000605211004386. [Google Scholar] [CrossRef]
- Ribieras, A.J.; Ortiz, Y.Y.; Li, Y.; Huerta, C.T.; Le, N.; Shao, H.; Vazquez-Padron, R.I.; Liu, Z.J.; Velazquez, O.C. E-Selectin/AAV2/2 Gene Therapy Alters Angiogenesis and Inflammatory Gene Profiles in Mouse Gangrene Model. Front Cardiovasc Med 2022, 9, 929466. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, H.J.; Parikh, P.P.; Lassance-Soares, R.M.; Regueiro, M.M.; Li, Y.; Shao, H.; Vazquez-Padron, R.; Percival, J.; Liu, Z.J.; Velazquez, O.C. Gangrene, revascularization, and limb function improved with E-selectin/adeno-associated virus gene therapy. JVS Vasc Sci 2021, 2, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yu, C.; Wang, P.; Shi, Y.; Cao, W.; Cheng, B.; Chapla, D.G.; Ma, Y.; Li, J.; Rodrigues, E.; et al. Glycoengineering of NK Cells with Glycan Ligands of CD22 and Selectins for B-Cell Lymphoma Therapy. Angew Chem Int Ed Engl 2021, 60, 3603–3610. [Google Scholar] [CrossRef] [PubMed]
- Turpin, A.; Labreuche, J.; Fléjou, J.F.; Andre, T.; de Gramont, A.; Hebbar, M. Prognostic factors in patients with stage II colon cancer: Role of E-selectin gene polymorphisms. Dig Liver Dis 2019, 51, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- DeAngelo, D.J.; Jonas, B.A.; Liesveld, J.L.; Bixby, D.L.; Advani, A.S.; Marlton, P.; Magnani, J.L.; Thackray, H.M.; Feldman, E.J.; O'Dwyer, M.E.; et al. Phase 1/2 study of uproleselan added to chemotherapy in patients with relapsed or refractory acute myeloid leukemia. Blood 2022, 139, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Wamil, B.D.; Thurman, G.B.; Sundell, H.W.; DeVore, R.F.; Wakefield, G.; Johnson, D.H.; Wang, Y.F.; Hellerqvist, C.G. Soluble E-selectin in cancer patients as a marker of the therapeutic efficacy of CM101, a tumor-inhibiting anti-neovascularization agent, evaluated in phase I clinical trail. J Cancer Res Clin Oncol 1997, 123, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Paluri, R.; Madan, A.; Li, P.; Jones, B.; Saleh, M.; Jerome, M.; Miley, D.; Keef, J.; Robert, F. Phase 1b trial of nintedanib in combination with bevacizumab in patients with advanced solid tumors. Cancer Chemother Pharmacol 2019, 83, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Dampier, C.D.; Telen, M.J.; Wun, T.; Brown, R.C.; Desai, P.; El Rassi, F.; Fuh, B.; Kanter, J.; Pastore, Y.; Rothman, J.; et al. A randomized clinical trial of the efficacy and safety of rivipansel for sickle cell vaso-occlusive crisis. Blood 2023, 141, 168–179. [Google Scholar] [CrossRef]
- Yang, F.; Qiu, R.; Abudoubari, S.; Tao, N.; An, H. Effect of interaction between occupational stress and polymorphisms of MTHFR gene and SELE gene on hypertension. PeerJ 2022, 10, e12914. [Google Scholar] [CrossRef]
- Rayat, S.; Ramezanidoraki, N.; Kazemi, N.; Modarressi, M.H.; Falah, M.; Zardadi, S.; Morovvati, S. Association study between polymorphisms in MIA3, SELE, SMAD3 and CETP genes and coronary artery disease in an Iranian population. BMC Cardiovasc Disord 2022, 22, 298. [Google Scholar] [CrossRef]
- Esen, B.; Sayin, T.; Akbas, H.; Gokmen, E.S.; Atay, A.E.; Guzel, S.P.; Gorgulu, N.; Yavuz, D.G. The role of serum E-selectin level and E-selectin gene S128R polymorphism on the enlargement of renal cyst in patients with polycystic kidney disease: Genetic background of renal cyst growth Clin Nephrol 2020, 93, 34-49. [CrossRef]
- Zhang, W.J.; Cheng, J.H.; Nie, Z.C.; Ding, B.S.; Han, Z.; Zheng, W.W. Association of E-Selectin Gene +A561C Polymorphism with Type 2 Diabetes in Chinese Population. Clin Lab 2022, 68. [Google Scholar] [CrossRef]
- Clancy, R.; El Bannoudi, H.; Rasmussen, S.E.; Bornkamp, N.; Allen, N.; Dann, R.; Reynolds, H.; Buyon, J.P.; Berger, J.S. Human low-affinity IgG receptor FcγRIIA polymorphism H131R associates with subclinical atherosclerosis and increased platelet activity in systemic lupus erythematosus. J Thromb Haemost 2019, 17, 532–537. [Google Scholar] [CrossRef]
Figure 1.
Interatomic alterations observed between adjacent amino acids by Dynamut2.0. A: rs1131498 (F193L) in SELL gene promotes a different pattern of interaction with adjacent amino acids by loss of a hydrophobic contact; B: rs6127 (D541N) of SELP gene promotes a change in the binding pattern with adjacent amino acids due to the disappearance of a hydrogen bond; C: rs2228315 (M62I) of SELPLG gene prometes alterations in amino acid binding patterns showing a weaker water mediated hydrogen bonding and D: rs5368 (H468Y) of SELE gene promotes alteration in hydrogen bonds with adjacent amino acids.
Figure 1.
Interatomic alterations observed between adjacent amino acids by Dynamut2.0. A: rs1131498 (F193L) in SELL gene promotes a different pattern of interaction with adjacent amino acids by loss of a hydrophobic contact; B: rs6127 (D541N) of SELP gene promotes a change in the binding pattern with adjacent amino acids due to the disappearance of a hydrogen bond; C: rs2228315 (M62I) of SELPLG gene prometes alterations in amino acid binding patterns showing a weaker water mediated hydrogen bonding and D: rs5368 (H468Y) of SELE gene promotes alteration in hydrogen bonds with adjacent amino acids.
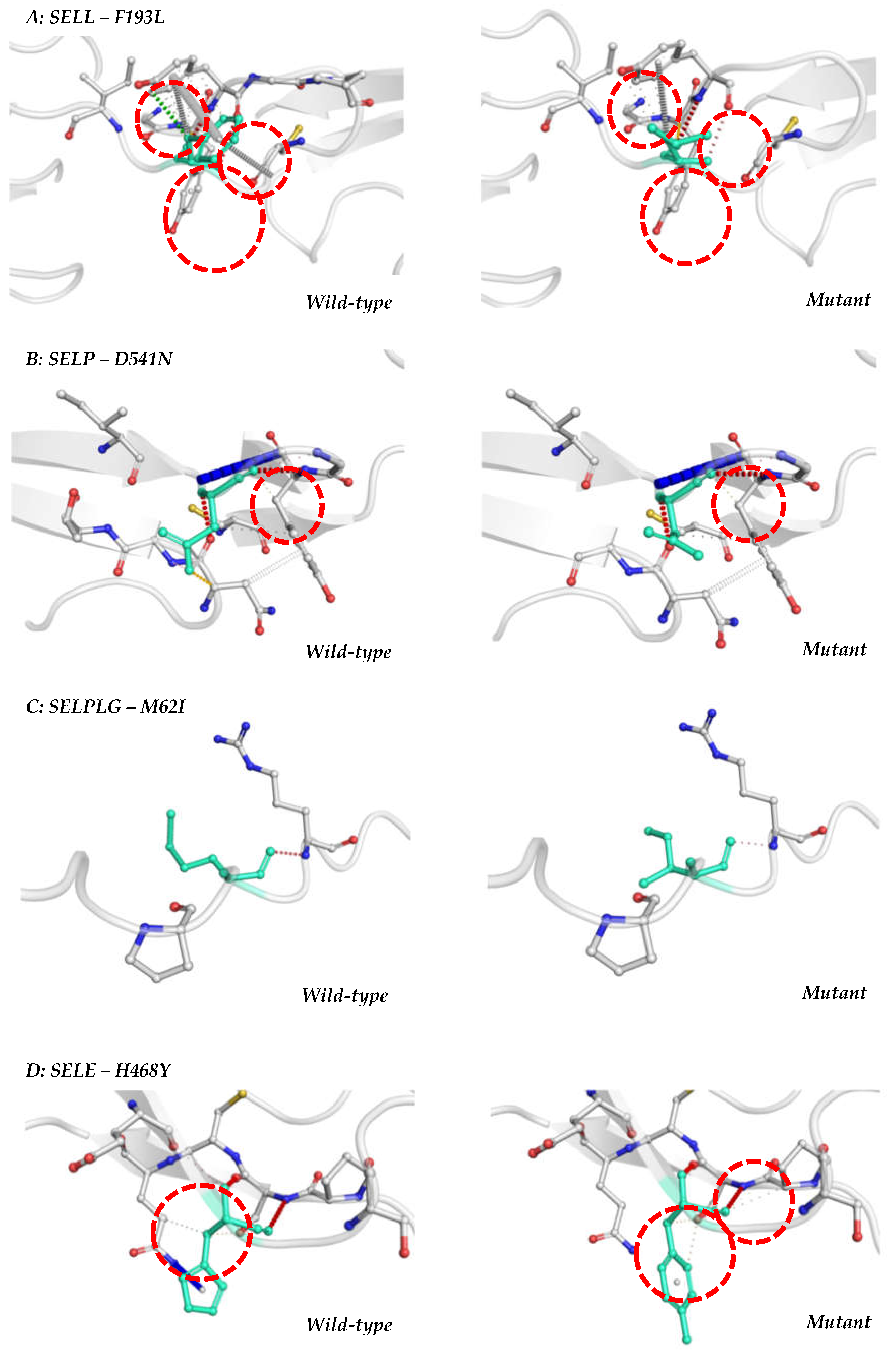

Table 1.
In silico analysis of SNPs in SELL, SELP, SELPLG and SELE genes considering DNA alterations analyzed by PredictSNP2.0.
Table 1.
In silico analysis of SNPs in SELL, SELP, SELPLG and SELE genes considering DNA alterations analyzed by PredictSNP2.0.
| Position | ID | Ref | Alt | Classification | PredictSNP2.0 | CADD | DANN | FATHMM | FUNSEQ2 | GWAVA | |
|---|---|---|---|---|---|---|---|---|---|---|---|
|
SELL Chr #1 |
169695726 | rs4987360 | A | G | intronic | D | D | D | ? | D | N |
| 169704697 | rs2229569 | G | A | exonic | D | D | D | D | N | ? | |
| 169706069 | rs4987301 | G | A | intronic | D | D | D | D | D | N | |
| 169706069 | rs4987301 | G | T | intronic | D | D | D | D | D | N | |
| 169707345 | rs1131498 | A | G | exonic | N | D | N | N | N | N | |
| 169712216 | rs2205849 | T | C | upstream | D | D | D | D | D | D | |
|
SELP Chr #1 |
169596108 | rs6133 | C | A | exonic | N | N | N | D | N | D |
| 169596108 | rs6133 | C | G | exonic | N | N | N | D | N | D | |
| 169597075 | rs6127 | C | G | exonic | N | N | N | N | N | ? | |
| 169597075 | rs6127 | C | T | exonic | N | N | N | N | N | ? | |
| 169601781 | rs3917777 | T | A | intronic | D | D | D | ? | D | N | |
| 169601781 | rs3917777 | T | C | intronic | D | D | ? | ? | D | N | |
| 169601781 | rs3917777 | T | G | intronic | D | D | D | ? | D | N | |
| 169605484 | rs2205894 | T | A | intronic | D | D | D | D | ? | N | |
| 169605484 | rs2205894 | T | G | intronic | D | D | D | D | ? | N | |
| 169605486 | rs2205893 | T | A | intronic | D | D | D | D | ? | N | |
| 169605486 | rs2205893 | T | G | intronic | D | D | D | D | ? | N | |
| 169611647 | rs6131 | C | T | exonic | N | N | N | N | N | N | |
|
SELPLG Chr #12 |
108623488 | rs7300972 | T | A | exonic | N | N | N | N | D | D |
| 108623488 | rs7300972 | T | C | exonic | N | N | N | N | D | D | |
| 108623898 | rs201851784 | A | G | exonic | N | N | N | N | N | D | |
| 108623898 | rs201851784 | A | T | exonic | N | N | N | N | N | D | |
| 108624122 | rs2228315 | C | T | exonic | N | N | N | N | N | N | |
| 108628692 | rs7138370 | G | A | intronic | N | D | D | N | D | D | |
|
SELE Chr #1 |
169727805 | rs5368 | G | A | exonic | N | N | N | N | D | D |
| 169729684 | rs1534904 | T | A | intronic | D | D | D | ? | D | N | |
| 169729684 | rs1534904 | T | G | intronic | N | D | N | N | D | N |
Abbreviations: Chr-Chromossome; ID–Identification of the SNP; Ref–Reference allele; Alt–Altered allele; N–Neutral; D-Deleterious and ?– Unknown.
Table 2.
Bioinformatics analysis of SNPs that cause amino acid alterations in L-selectin, P-selectin, PSGL-1 and E-selectin proteins according to PredictSNP1.0.
Table 2.
Bioinformatics analysis of SNPs that cause amino acid alterations in L-selectin, P-selectin, PSGL-1 and E-selectin proteins according to PredictSNP1.0.
| Gene | ID | AA Change | PredictSNP1.0 | MAPP | PhD-SNP | PolyPhen-1 | PolyPhen-2 | SIFT | SNAP |
|---|---|---|---|---|---|---|---|---|---|
| SELL | rs1131498 | F193L | N | N | N | N | N | N | N |
| rs2229569 | P213S | N | D | N | N | N | N | N | |
| rs2229569 | P213T | N | D | D | N | N | D | N | |
| SELP | rs6131 | S331N | N | N | N | N | N | N | N |
| rs6127 | D541N | N | N | N | N | N | N | N | |
| SELPLG | rs2228315 | M62I | N | N | N | D | N | N | N |
| rs201851784 | V137A | N | N | N | N | N | N | N | |
| rs7300972 | M274V | N | D | N | N | N | D | N | |
| SELE | rs5368 | H468Y | N | N | N | N | N | N | D |
Abbreviations: ID-Identification of the SNP; AA-Amino Acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



