
Submitted:
28 February 2023
Posted:
03 March 2023
You are already at the latest version
Abstract
This study is the first report of the successful cultivation of Dinophysis norvegica isolated from Japanese coastal waters (7 of 48 isolated cells were established as the clonal cultures, 14.5%) and their toxin content and production. The strains were maintained at relatively high abundance (>2,000 cells mL-1) for more than one year when fed on Mesodinium rubrum with the addition of Teleaulax amphioxeia. At the end of the one-month incubation, the total amounts of pectenotoxin-2 (PTX2), dinophysistoxin-1 (DTX1) and okadaic acid (OA) were 132.0-375.0 ng mL-1 (n = 7), 0.7-3.6 ng mL-1 (n = 3) and trace level OA (n = 1), respectively. Similarly, the cell quota of PTX2 and DTX1 were 60.6-152.4 pg cell-1 (n = 7) and 0.5-1.2 pg cell-1 (n = 3), respectively. These data indicated that toxin production varies depending on the strains in this species. In the growth experiment, D. norvegica grew, but it was very slow first 12 days. However, after that they grew exponentially with the maximum growth rate of 0.56 divisions day-1 (during Day 24-27), reaching a maximum concentration of 3,000 cells mL-1 at the end of incubation (Day 36), suggesting that they have a long lag phase. In the toxin production, OA was not at a detectable level (≤ 0.010 ng mL-1) during the 36 days of incubation except for Day 6. The concentration of DTX1 and PTX2 showed similar patterns as described for vegetative growth, but the toxin production still increased on Day 36 (1.3 ng mL-1 and 154.7 ng mL-1 in DTX1 and PTX2, respectively). The findings of this study provide novel information on the toxin content and production in D. norvegica as well as details on the culturing and maintenance of the species.
Keywords:
Diarrhetic shellfish poisoning (DSP)
; Dinophysis norvegica
; Mesodinium rubrum
; Teleaulax amphioxeia
; Pectenotoxin-2 (PTX2)
; Dinophysistoxin-1 (DTX1)
; Okadaic acid (OA)
1. Introduction
Harmful Algal Blooms (HABs) are caused by several species of microalgae in freshwater, marine, and brackish environments, which may lead toimportant ecosystematic and socioeconomic impacts as well as human illnesses [1,2,3]. There has been a marked increase in the occurrence of HABs worldwide over the past decades [1,2,3,4,5]. The rise has been associated with climate change and intensified anthropogenic activities, notably eutrophication, transport of species with maritime activities, alteration of natural habitats, and growth of the aquaculture industry [4,6,7,8,9,10,11,12,13,14].
In marine waters, dinoflagellates form the majority of toxin-producing HAB species and are responsible for several human poisoning syndromes, including Ciguatera Fish Poisoning (CFP), the Neurotoxic Shellfish Poisoning (NSP), Paralytic Shellfish Poisoning (PSP), and the Diarrhetic Shellfish Poisoning (DSP) [15]. For example, gastrointestinal poisoning in humans is caused by the consumption of shellfish contaminated with DSP toxins [16,17]. These toxins are produced by dinoflagellates from the genera Dinophysis, Phalacroma, and Prorocentrum. Ten species of Dinophysis and two species of Phalacroma are known to produce lipophilic Diarrheic Shellfish Toxins, or DSTs, i.e. Okadaic Acid (OA) and its analogues the dinophysistoxins (DTX) principally DTX1, DTX2 and DTX3; in addition to the bioactive pectenotoxins, the PTX [17,18,19,20,21,22,23,24,25,26,27,28].
Typically, Dinophysis spp. do not attain high cell densities but form dense patches of populations, which sets them apart from other HAB species and makes their monitoring and prediction of shellfish contaminations with DSTs more difficult, especially since molecular tools have been hard to develop due to insufficient resolution to differentiate between the species [21,29,30,31,32,33,34,35,36].
Despite the availability of extensive studies, little is known about the ecophysiology, bloom mechanisms, and toxin production of Dinophysis spp. due to difficulties in establishing and maintaining cultures [21,37,38,39]. The discovery of mixotrophy in Dinophysis spp. [37,38,39,40] and plastids of cryptophyte origin [40,41,42,43,44,45,46,47] led to the first success in establishing cultures of Dinophysis acuminata [48]. Seven species were subsequently cultured based on feeding Dinophysis spp. with the ciliate Mesodinium rubrum grown with the cryptophyte Teleaulax sp., namely D. fortii [49], D. acuta [50], D. sacculus [51], D. tripos [52], D. cf. ovum [53], D. caudata [54], and D. infundibulum [55]. Mainly growth and, in some cases, toxin production in the established cultures have been reported. A few studies have investigated the effects of temperature, prey, and irradiance on the growth and toxin production of Dinophysis spp. in these cultures [33,48,56,57,58,59,60,61,62,63,64,65].
There is evidence of the global expansion of Dinophysis species related to both climate change and aquaculture activities [14,66], resulting in hardship to fisheries and aquaculture industries through extended closure of shellfish production [67]. Among the toxigenic species, six of the toxic Dinophysis species have a wide, global distribution, including D. norvegica [36,68,69]. A boreal to cold-temperate species, D. norvegica is commonly reported from the Northern Hemisphere, for example from the coastal waters around Scotland and Norway, the Baltic Sea, and the Arctic Sea [29,30,71,72,73,74,75,76]. Recently, it was reported for the first time at very low occurrence in oceanic samples in the Southern Hemisphere, in southern Argentine Sea [77]. It forms dense blooms in the Baltic Sea and Eastern Canada with mild DSP outbreaks [78,79,80]. The earliest information based on cells picked from environmental samples showed the production of OA and DTX1 in Norway [17], and high content of OA in Eastern Canada [81]. More recently, LC-MS have shown the production of PTX2, PTX12, and traces of OA by strains from Norway [22]. In the Baltic Sea, D. norvegica produces OA, PTX2, and PTX2SA [82] leading to the contamination of blue mussels and flounders with OA [83,84]. One recent study reported the production of Dihydrodinophysistoxin-1 in picked cells from environmental samples and cultures of D. norvegica from the Gulf of Maine, USA [85], with a complete absence of OA, DTX1, and DTX2 following analyses with LC-MS/MS. In Japan, high levels of PTX2 have been reported for the first time in cells of D. norvegica picked from environmental samples [86]. In a later study, PTX2 was confirmed as the dominant toxin in D. norvegica, although some of the picked cells had trace levels of OA and DTX1 [87]. In the present study, we report the successful cultivation of D. norvegica isolated from Japanese waters for the first time. The toxin productions in seven strains of D. norvegica are provided as well as the information on the growth and toxin production of one strain during a 36-day culture experiment.
2. Results
2.1. Species identification
Cells of Dinophysis norvegica are generally large, ovoid, and robust. The posterior end tapers to a triangular shape (Figure 1). Dinophysis norvegica is very similar to D. acuta in morphology, therefore they may be misidentified. These species can be differentiated by their size (although it overlaps) and the location of thewidest position: D. acuta is larger and widest below the mid-section, whereas D. norvegica is smaller and widest in the middle region of the cell [88,89]. A phylogenetic analysis based on the D1/D2 region (735 bp) supported that the strains isolated from Funka Bay, Japan belong to the D. norvegica clade (Figure 2), and are closely related to the strains from Canada, Norway, and the USA from the Atlantic Ocean..
2.2. Feeding behavior and growth of Dinophysis norvegica in culture experiments
Only seven cultures of 48 single-cell isolates grew with the addition of the ciliate Mesodinium rubrum as the prey species. However, they were successfully established as clonal strains and the success rate for isolation was 14.6% (7/48). The cultures reached maximum cell densities of 1,057-3,050 cells ml-1 (2,020 ± 702) (mean ± SD, n = 7) at end of the one-month incubation. Similar to the case of other Dinophysis species, Dinophysis norvegica was able to feed on M. rubrum and grew. The microphotographs showed the large nucleus which occupied the upper half of the cell and food vacuoles at the lower part (Figure 1A) and numerous chloroplasts (Figure 1B).
In the growth experiment, M. rubrum grew exponentially during the first 9 days, reaching until 5,600± 346 cells mL-1 (mean ± SD, n = 3) (Figure 3). After that, cell abundances of M. rubrum decreased sharply and disappeared by Day 30, probably due to the active consumption by D. norvegica and natural death. Dinophysis norvegica grew, but it showed very slow growth for the first 12 days, suggesting that they have a long lag phase. After that they grew exponentially with a maximum growth rate of 0.56 divisions day-1 (during Day 24-27), reaching a maximum concentration of 2,883 ± 104 cell mL-1 at the end of incubation (Day 36), and the growth rates in every three days from Day 0 to Day 30 were -0.06 to 0.56 divisions day-1 (0.18 ± 0.18) (Figure 3).
2.3. Toxin production
Low levels of DTX1 were found but only in cells of three strains of D. norvegica at 0.5 pg cells-1 for DN05, 0.7 pg cell-1 for DN08, and 1.2 pg cell-1 for DN06. All seven strains, however, produced PTX2 with cell quotas varying from 60.6 pg cell-1 to 152.4 pg cell-1 (Table 1). All strains of Dinophysis norvegica did not produce OA, except for Stain DN08 which detected a trace level of OA. In cultures, the toxin concentration ranged from 0.705 ng mL-1 to 3.55 ng mL-1 in the case of DTX1 and from 126 ng mL-1 to 375 ng mL-1 in the case of PTX2 (Table 1).
Strain DN16062021FUN-06 of D. norvegica produced DTX1 and PTX2 throughout the 36-day culture experiment (Figure 4). Both productions of DTX1 and PTX2 started to increase from 0.092 ± 0.009 pg cell-1 on Day 12 to 1.280 ± 0.185 pg cell-1 on Day 36, and from 9.3 ± 0.5 pg cell-1 on Day 12 to 154.7 ± 24.2 pg cell-1 on Day 36, respectively. The specific toxin production rates for DTX1 and PTX2 during the exponential growth phase were 0.291 ± 0.020 pg mL-1 day-1 and 0.291 ± 0.023 pg mL-1 day-1, respectively. The net toxin production rate (Rtox) during the exponential growth phase was 0.001±0.0003 ng mL-1 day-1 for DTX1 and 0.13±0.03 ng mL-1 day-1 for PTX2 in the exponential phase. DTX2 concentrations that were above the detection limit were available from Day 18 to Day 36 and ranged from 0.132 to 1.28 ng mL-1. For PTX2, the concentrations above the detection limit were measured from Day 3 until the end of the experiment Day 36, and were in the range of 3.4 to 154.7 ng mL-1.
3. Discussion
Similar to the case of other Dinophysis species [49,50,51,52,53,54,55] , D. norvegica was able to feed on M. rubrum. Dinophysis norvegica swam around the prey and inserted the peduncle into the cell of M. rubrum, the ciliate became immobile and their cilia were shed from the cell within 1-5 minutes. The cytoplasm of the prey was actively ingested through the peduncle. It took 45-100 min until whole cell content of M. rubra was consumed in D. fortii and D. tripos [49,52]. Dinophysis acuminata, D. caudata, D. fortii and isolated from Japanese coastal waters displayed growth rates of 0.50-1.03 divisions day-1, reaching maximum concentrations of 2,200-11,000 cells mL-1 at temperature ranging from 18 to 25 °C [36] . In this study, D. norvegica showed a similar growth rate at a lower temperature of 12.5 °C than other species (Figure 2). The culture strains were maintained successfully at high densities (>2,000 cells mL-1) for more than one year. This is an advancement compared to previous findings, where clonal cultures of D. norvegica isolated from Funka Bay and lake Notoroko in Hokkaido, Japan, grew well (>1,000 cell mL-1) in the first incubation (24/96, 25%), however, when they were reinoculated into fresh Mesodinium cultures, no further growth was confirmed resulting in the failure to establish cultures (0/96, 0%) [52]. This led to the conclusion that the predator and the prey from different regions may be incompatible and cause of the failure in culturing [52]. In the growth experiment using the established cultures, D. norvegica had much longer log phase (12 days) (Figure 3) than those in D. acuminata, D. caudata and D. fortii (3-4 days) [49,54,55]. Perhaps, D. norvegica may become unstable when they are reinoculated into fresh Mesodinium cultures because of sudden environmental changes such as in pH levels. It is suggested that the reinoculation of D. norvegica cells with relatively high concentration (>250 cells mL-1) would achieve the successful maintenance of the cultures for long periods. The ranges of water temperatures and salinity in which this species appears in southern Hokkaido including Funka Bay were 2-16 °C and 24.3-33.9 PSU during 2016-2020 (https://www.hro.or.jp/list/fisheries/research/central/section/kankyou/kaidoku/j12s220000000dgw.html). Influence of the temperature (12.5 °C) on the success of establishing cultures is still unclear in this study.
Establishment of culture strains in the toxic Dinopshysis species is important for investigating toxin production and how it is influenced by changes in the physical-chemical conditions. Until now, information on the concentration of toxins produced by D. norvegica has been available based on picked and pooled cells from field samples from two locations in Norway and Japan [19,22,87]. Similarly to the previous records based on isolated cells, OA concentrations in cultured strains remained mainly below the detection limit [19,22,87]. The maximum DTX1 concentrations in cultured strains were lower than reported based on cells from field samples (1.16 pg cell-1 vs 14 pg cell-1) [19,22,87]. Interestingly the PTX2 maximum concentrations were much higher in the cultured strains (152 pg cell-1), than in the cells collected from the field in Japan (89 pg cell-1) and 1.7 pg cell-1 in Norway [19,22,87]. From 1987 until 2022, 43 HABs associated with D. norvegica have been recorded globally with the majority of those associated with DSP [90]. A study based on Scottish shellfish farms estimated that a 1% change in the production of toxins produced by Dinophysis spp. led to an annual loss of 1.37 million £ (GBP) [91]. Thus, further experiments with cultured strains can lead to an improved understanding of the variability in toxin production between the strains from the same and different geographic locations as well as the influence of changes in physical-chemical variables [69,92].
4. Materials & Methods
4.1. Isolation of clonal strains and establishment of clonal cultures
Mesodinium rubrum and Teleaulax amphioxeia were isolated from Inokushi Bay (32.7998 N, 131.8923 E) in Oita Prefecture, Japan, at the end of February 2007 [49]. The M. rubrum culture was maintained by mixing 50 ml of the culture (7.0–9.0 × 103 cells mL–1) with 100 ml of a modified f/2 medium [93,94], with the addition of 25-100 µL of T. amphioxeia culture (containing 0.5–2.0 × 104 cells) as a food source. The culture medium was prepared with 1/3 nitrate, phosphate, metals, and 1/10 vitamins, plus any enrichment from the autoclaved natural seawater collected from Tokyo Bay (35.3460 N, 139.6570 E). Seawaters were filtrated through three cartridge filters with 5, 1, and 0.5 µm (STG-10-5, STG-10-1, STG-10-0.5, Kankyotechnos, Japan) at the sampling site and brought back to the lab. Salinity was adjusted to 30 practical salinity units (PSU) before autoclaving. A part of the ciliate culture was transferred into a fresh culture medium containing the food source once a week, and they were maintained at a temperature of 18 ˚C under a photon irradiance of 100 μmol m–2 s–1, provided by cool-white fluorescent lamps, with a 12:12 h light: dark cycle.
Cells of Dinophysis norvegica (48 cells in total) were isolated by micropipetting from a seawater sample collected from Funka Bay, Japan (42.28N, 140.35E) in June 2021 and incubated in individual wells of a 48-well microplate (Iwaki, Japan). Each cell was grown in 1.0 mL of the culture medium prepared for the maintenance of the M. rubrum strain, containing ca. 1.0 × 103 cells of the marine ciliate M. rubrum as the prey species. Dinophysis cells were incubated under the same light conditions as those for the M. rubrum culture, but at the temperature of 12.5 °C. After one month of incubation, several strains grew well and were established as clonal strains (7 strains in total). Small aliquots (0.1 mL) of the established cultures in each strain were inoculated into 2.9 mL of fresh M. rubrum culture (ca. 2 × 103 cells mL–1, just after reinoculation for the maintenance without adding Teleaulax culture) in 12-well microplates to maintain these strains, and they were incubated for one month under the same conditions as mentioned above. After one month of incubation, 1 mL of each culture was sampled for toxin analysis, and cell abundances in each stain were counted using a Nikon TE-300 (Tokyo, Japan) inverted microscope. Also, the orange auto-florescence derived from the chloroplasts of M. rubrum in D. norvegica cells was observed using an epifluorescence microscope under blue light excitation (Zeiss Axioskop 2 (Carl Zeiss, Göttingen, Germany) with digital camera Axiocam 305 color (Carl Zeiss, Göttingen, Germany).
4.2. Growth experiments
The Mesodinium rubrum culture grown until the late exponential growth phase (ca. 5.0 × 103 cells mL–1) was diluted with the fresh culture media to give initial concentrations of ca. 2.0 × 103 cells mL–1, and 7.5 mL aliquots of the mixed culture were inoculated into the wells of 6-well microplates (Iwaki, Japan). Next, 125 µL of a D. norvegica culture (strain, DN16062021FUN-06) containing 375 cells was added into the M. rubrum culture to give an initial concentration of 50 cells mL-1. The growth experiment was conducted for 36 days under the same light and temperature conditions used for maintaining the culture of D. norvegica. In the growth experiment, 1 mL of the cultures (triplicate) was sampled every three days except for Day 33. The cell densities of D. norvegica and M. rubrum were counted using an inverted microscope. The specific growth rate (µ, divisions day-1) of D. norvegica was determined during the exponential growth phase according to [95].
4.3. Sequences of 28S rDNA (D1-D2 region)
Genomic DNAs were extracted from several cells in each strain with 5% Chelex buffer in four of the established strains [96] . PCR amplification was carried out on a thermal cycler (PC-808, ASTEC, Fukuoka, Japan) with a reaction mixture consisting of 1 μL template DNA, 1 μM each of D1/D2 primer sets [97] , 0.2 mM of each dNTP, 1× PCR buffer, 1.5 mM Mg2+, 1U KOD-Plus-Ver.2 (TOYOBO, Osaka, Japan), and RNA-free dH2O to bring up to 25 μL volume. The PCR cycling conditions were as follows: 2 min at 94 °C, 30 cycles at 94 °C for 15 sec, 55 °C for 30 sec, and 68 °C for 40 sec. The PCR products were transformed into DH5α cells (Promega, Madison, WI, USA) after ligation into the pGEM T-Easy Vector (Promega). The plasmid DNAs were purified after color selection. DNA sequences were determined using a Dynamic ET terminator cycle sequencing kit (GE Healthcare, Little Chalfont, UK) in combination with M13 Reverse and U19 primers and analyzed on a DNA sequencer (ABI3730, Applied Biosystems, Foster City, CA, USA). BLAST search was performed to determine closely related species and their GenBank sequences were obtained for phylogenetic analyses. The sequences were aligned using AliView [98] ( and identical sequences were compiled into a single sequence. All newly obtained sequences were deposited into the DDBJ databank (accession numbers: LC634028- LC634030).
A phylogenetic tree was constructed based on maximum likelihood (ML) using MEGA version 10 [99] with the best substitution model selected: Kimura 2-parameter model plus gamma distribution (G = 0.86). Bootstrap support (BS) values of ML and neighbor-joining (NJ) analyses for the trees were estimated using 500 replicates each. For the posterior probabilities (PP) of Bayesian inference, the best model substitution calculated by Akaike information criterion in jModelTest version 2.1.10 [100] was TIM plus gamma (G = 0.9710), and the effective sample size was calculated using Bayesian Evolutionary Analysis Sampling Trees (BEAST) and Tracer. Bayesian inference was conducted using MrBayes version 3.2.5 [101] based on the Bayesian information criteria calculated by jModelTest. 3,077,000 Markov chain Monte Carlo generations was with four chains and trees sampled every 1,000 generations and PP were estimated with 25% generations burn-in. Convergence of the chains was reconfirmed when the average standard deviations of the split frequencies were below 0.01 after calculations. Sequences from Prorocentrum micans and Prorocentrum minimum were used as outgroup.
4.4. DSP toxin analysis
The samples were frozen at –30°C until the toxins were extracted by solid-phase extraction (SPE). The SPE of toxins was modified compared to a previous method [86,102,103]. The 1-mL frozen and thawed samples were applied to the MonoSpin C18 centrifuge cartridge column (GL Science Inc., Tokyo, Japan) equilibrate with 1.0-mL each methanol and distilled water. The SPE column was washed with 0.5-mL distilled water, and the toxins were eluted with 0.1-mL methanol. The methanol elutes were directly analyzed by LC-MS/MS. LC-MS/MS analysis of the toxins was carried out according to a previous method [104]. A Nexera-20XR series liquid chromatograph (Shimadzu, Kyoto, Japan) was coupled to a QTRAP 4500 mass spectrometer (SCIEX, MA, USA) of hybrid triple quadrupole/linear ion trap. Separations were performed on LC columns (internal diameter [i.d.], 100 mm × 2.1 mm) packed with 1.9-μm Hypersil GOLD C8 (Thermo Fisher Scientific Inc., Waltham, MA, USA) and maintained at 30°C. Eluent A was water, and eluent B was acetonitrile water (95:5), containing two mM ammonium formate and 50 mM formic acid. Toxins were eluted from the column with 50% B at a flow rate of 0.3 mL·min–1. Multiple reaction monitoring (MRM) LC-MS/MS analysis with negative-mode ionization was carried out using the target parent ions and the fragment ions in Q1 and Q3 for each toxin as follows: OA, m/z 803.5 > 255.1; DTX1, m/z 817.5 > 255.1; PTX2, m/z 857.5 > 137.0; PTX1 and PTX11, m/z 873.5 > 137.0; PTX2 Seco acid (PTX2 SA), m/z 875.5 > 137.0. The lowest detection limits of OA/DTX1 and PTX2 were 0.1 and 1.2 ng·mL–1. These levels are equivalent to 0.1 pg·cell–1 of OA (and DTX1) and 1.2 pg·cell–1 of PTX2, when 100 cells of the toxic plankton were analyzed using our LC-MS/MS method. During the growth experiments, the specific toxin production rate (µtox, pg cell-1 day-1) and net toxin production rate (Rtox) were calculated for the exponential growth phase, using the previously published equations [105].
Author Contributions
Conceptualization, SN; Investigation, TK & MN; Methodology, HU, SN; Data analyses, HU, LB, NN, RM, SN, WML; writing –original draft, LB & SN; writing-review and editing: SS, LB & SN. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by JST/JICA, Science and Technology Research Partnership for Sustainable Development (JPMJSA1705) [SN], a Grant-in-Aid for Scientific Research (Kiban-B) by the Japan Society for the Promotion of Science (21H02274) [SN, NN, RM, TK], (18KK0182) [SN], and Estonian Research Council (PSG735) [SS].
Acknowledgement
We thank Drs. Toshiyuki Suzuki and Hiroshi Oikawa, Fisheries Technology Institute, Japan Fisheries Research and Education Agency, for their encouragement during this study.
Conflict of Interest
The authors declare no conflict of interest.
References
- Smayda, T.J. Novel and nuisance phytoplankton blooms in the sea: evidence for a global epidemic. In Toxic Marine Plankton, Proceedings of the 4th International Conference on Toxic Marine Phytoplankton, Elsevier, Lund, Sweden, 1990, pp. 29–40.
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine algal toxins: origins, health effects, and their increased occurrence. Environ. Health Persp. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Zingone, A.; Oksfeldt Enevoldsen, H. The diversity of harmful algal blooms: a challenge for science and management. Ocean Coast. Manage. 2000, 43, 725–748. [Google Scholar] [CrossRef]
- Allen, J.I.; Anderson, D.; Burford, M.; Dyhrman, S.; Flynn, K.; Glibert, P.M.; Granéli, E.; Heil, C.; Sellner, K.; Smayda, T.; Zhou, M. Global Ecology and Oceanography of Harmful Algal Blooms, Harmful Algal Blooms in Eutrophic Systems. GEOHAB Report 4IOC and SCOR. GEOHAB, Paris, France/Baltimore, MD, USA, 2000, pp. 1–74.
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences. Estuaries 2002, 25, 562–584. [Google Scholar] [CrossRef]
- Edwards, M.; Johns, D.G.; Leterme, S.C.; Svendsen, E.; Richardson, A.J. Regional climate change and harmful algal blooms in the northeast Atlantic. Limnol. Oceanogr. 2006, 51, 820–829. [Google Scholar] [CrossRef]
- Hégaret, H.; Shumway, S.E.; Wikfors, G.H.; Pate, S.; Burkholder, J.M. Potential transfer of harmful algae through relocation of bivalve molluscs. Mar. Ecol. Progr. Ser. 2008, 361, 169–181. [Google Scholar] [CrossRef]
- Heisler, J.; Gilbert, P.M.; Burkholder, J.M.; Anderson, D.M; Cochlan, W.; Dennison, W. C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; Lewitus, A.; Magnien, R.; Marshall, H.G.; Sellner, K.; Stockwell, D.A.; Stoecker, D.K.; Suddleson, M. Eutrophication and harmful algal blooms: a scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Backer, L.C.; Fleming, L.E. Impacts of climate variability and future climate change on harmful algal blooms and human health. Environ. Health 2008, 7(2), S4. [Google Scholar] [CrossRef] [PubMed]
- Roger, A.D.; Laffoley, D.d’A. International Earth System Expert Workshop on Ocean Stresses and Impacts. Summary Report. IPSO, Oxford, 2011, 18 pp.
- Fu, F.X.; Tatters, A.O.; Hutchins, D.A. Global change and the future of harmful algal blooms in the ocean. Mar. Ecol. Progr. Ser. 2012, 470, 207–233. [Google Scholar] [CrossRef]
- Glibert, P.M.; Allen, J.I.; Artioli, Y.; Beusen, A.; Bouwman, L.; Harle, J.; Holmes, R.; Holt, J. Vulnerability of coastal ecosystems to changes in harmful algal bloom distribution to climate change: projections based on model analysis. Glob. Change Biol. 2014, 20, 3845–3858. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Dechroui Bottein, M-Y.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H.; Sunesen, I.; Pitcher, G.C.; Richardson, A.; Schweibold, L.; Tester, P.A.; Trainer, V.L.; Yñiguez, A.T.; Zingone, A. Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts. Comm. Earth Environ. 2021, 2, 117.
- Burkholder, J.M. Implication of harmful microalgae and heterotrophicdinoflagellates in management of sustainable marine fisheries. Biol. App. 1998, 8, S37–S62. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Lee, J.S.; Igarashi, T.; Fraga, S.; Dahl, E.; Hovgaard, P.; Yasumoto, T. Determination of diarrhetic shellfish toxins in various dinoflagellate species. J. Appl. Phycol. 1989, 1, 147–152. [Google Scholar] [CrossRef]
- Jun, J.H.; Sim, C.S.; Lee, C.O. Cytotoxic compounds from a two-sponge association. J. Nat. Prod. 1995, 58, 1722–1726. [Google Scholar]
- Terao, K.; Ito, E.; Yanagi, T.; Yasumoto, T. Histopatholgical studies on marine toxin poisoning. 1. Ultrastructural changes in the small intestine and liver of suckling mice induced by dinophysistoxin-1 and pectenotoxin-1. Toxicon 1986, 24, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.Y.; Kim, Y.Z. Acute toxicity of pectenotoxin 2 and its effects on hepatic metabolizing enzyme in mice. Korean J. Toxicol. 1997, 13, 183–186. [Google Scholar]
- Maestrini, S.Y. Bloom dynamics and ecophysiology of Dinophysis spp. In Physiological Ecology of Harmful Algal Blooms; Anderson, D. M.; Cembella, A. D.; Hallegraeff, G. M., Eds.; NATO ASI Series, Vol. G 41, Springer-Verlag, Berlin, 1998, pp. 243–265.
- Miles, C.O.; Wilkins, A.L.; Samdal, I.A.; Sandvik, M.; Petersen, D.; Quilliam, M.A.; Naustvoll, L.J.; Jensen, D.J.; Cooney, J.M. A novel pectenotoxin, PTX-12, in Dinophysis spp. and shellfish from Norway. Chem. Res. Toxicol. 2004, 17, 1423–1433. [Google Scholar] [CrossRef]
- Ito, E., Suzuki, T., Oshima, Y., Yasumoto, T., 2008. Studies on diarrheic activity on pectenotoxin-6 in the mouse and rat. Toxicon, 15, 707–716.
- Reguera, B.; Pizarro, G. Planktonic Dinoflagellates Which Produce Polyether Toxins of the Old—DSP Complex. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Botana, L.M., Ed.; 2nd ed., Taylor & Francis, London, UK, 2008, pp. 257–284.
- Tubaro, A.; Sosa, S.; Bornancin, A.; Hungerford, J. Pharmacology and toxicology of diarrheic shellfish toxins. In: Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; Botana, L.M., Ed.; 2nd ed., CRC Press, Boca Raton, FL, USA, 2008, pp. 229–253.
- Vilariño, N.; Espiña, B. Pharmacology of pectenotoxins. In: Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; Botana, L.M., Ed.; 2nd ed., CRC Press, Boca Raton, FL, USA, 2008, pp. 361–369.
- >Lawrence, J.; Loreal, H.; Toyofuku, H.; Hess, P.; Iddya, K.; Ababouch, L. Assessment and Management of Biotoxin Risks in Bivalve Molluscs; FAO Fisheries and Aquaculture Technical Paper 551. FAO: Rome, Italy, 2011, p. 337.
- Moestrup, Ø.; Akselmann-Cardella, R.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Komárek, J.; Larsen, J.; Lundholm, N.; Zingone, A.; (Eds.) 2009 onwards. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. http://www.marinespecies.org/hab (accessed 2022-04-25). [CrossRef]
- Edvardsen, B.; Shalchian-abrizi, K.; Jakobsen, K.S.; Medlin, L.K.; Dahl, E.; Brubak, S.; Paasche, E. Genetic variability and molecular phylogeny of Dinophysis species (Dinophyceae) from Norwegian waters inferred from single cell analyses of rDNA. J. Phycol. 2003, 39, 395–408. [Google Scholar] [CrossRef]
- Hart, M.C.; Green, D.H.; Bresnan, E.; Bolch, C.J. Large ribosomal RNA gene variation and sequence heterogeneity of Dinophysis (Dinophyceae) species from Scottish coastal waters. Harmful Algae 2007, 6, 271–287. [Google Scholar] [CrossRef]
- Raho, N.; Pizarro, G.; Escalera, L.; Reguera, B.; Marín, I. Morphology, toxin composition and molecular analysis of Dinophysis ovum Schütt, a dinoflagelate of the “Dinophysis acuminata complex”. Harmful Algae 2008, 7, 839–848. [Google Scholar] [CrossRef]
- Raho, N.; Rodríguez, F.; Reguera, B.; Marín, I. Are the mitochondrial cox1 and cobgenes suitable markers for species of Dinophysis Ehrenberg? Harmful Algae 2013, 28, 64–70. [Google Scholar] [CrossRef]
- Rodríguez, F.; Escalera, L.; Reguera, B.; Rial, P.; Riobó, P.; de Jesús da Silva, T. Morphological variability, toxinology and genetics of the dinoflagellate Dinophysis tripos (Dinophysiaceae, Dinophysiales). Harmful Algae 2012, 13, 26–33. [Google Scholar] [CrossRef]
- Taylor, M.; McIntyre, L.; Ritson, M.; Stone, J.; Bronson, R.; Bitzikos, O.; Rourke, W.; Galanis, E.; Team, O. Outbreak of diarrhetic shellfish poisoning associated with mussels, British Columbia, Canada. Mar. Drugs 2013, 11, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.F.; Amorim, A.L.; Bresnan, E. Diversity and plastid types in Dinophysis acuminata complex (Dinophyceae) in Scottish waters. Harmful Algae 2014, 39, 223–231. [Google Scholar] [CrossRef]
- Nagai, S.; Sildever, S.; Suzuki, T.; Nishitani, G.; Basti, L.; Kamiyama, T. Successful cultivation and growth characteristics of the dinoflagellate Dinophysis. In Dinoflagellates: Morphology, Life History and Ecological Significance; Suba Rao, D. V.; Ed., Nova Science Publishers, New Jersey, 2020, pp. 129–166.
- Sampayo, M.A.de.M. Trying to cultivate Dinophysis spp. In Toxic Phytoplankton Blooms in the Sea; Smayda, T. J.; Shimizu, Y.; Eds., Elsevier, Amsterdam, 1993, pp. 807–810.
- Jacobson, D.M.; Andersen, R.A. The discovery of mixotrophy in photosynthetic species of Dinophysis (Dinophyceae): light and electron microscopical observations of food vacuoles in Dinophysis acuminata, D. norvegica and two heterotrophic dinophysoid dinoflagellates. Phycologia 1994, 33, 97–110. [Google Scholar] [CrossRef]
- Nishitani, G.; Miyamura, K.; Imai, I. Trying to cultivation of Dinophysis caudata (Dinophyceae) and the appearance of small cells. Plankton Biol. Ecol. 2003, 50, 31–36. [Google Scholar]
- Nishitani, G.; Sugioka, H.; Imai, I. Seasonal distribution of species of the toxic dinoflagellate genus Dinophysis in Maizuru Bay (Japan), with comments on their autofluorescence and attachment of picophytoplankton. Harmful Algae 2002, 1, 253–64. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Cryptophycean-like double membrane-bound chloroplast in the dinoflagellate, Dinophysis Ehrenb.: evolutionary, phylogenetic and toxicological implications. Bot. Acta 1988, 101, 196–203. [Google Scholar] [CrossRef]
- Lucas, I.A.N.; Vesk, M. The fine structure of two photosynthetic species of Dinophysis (Dinophysiales, Dinophyceae). J. Phycol. 1990, 26, 345–357. [Google Scholar] [CrossRef]
- Hewes, C.D.; Mitchell, B.G.; Moisan, T.A.; Vernet, M.; Reid, F.M.H. The phycobilin signature of chloroplasts from three dinoflagellate species: A microanalytic study of Dinophysis caudata, D. fortii, and D. acuminata (Dinophysiales, Dinophyceae). J. Phycol. 2002, 34, 945–954. [Google Scholar]
- Takishita, K.; Koike, K.; Maruyama, T.; Ogata, T. Molecular evidence for plastid robbery (kleptoplastidy) in Dinophysis, a dinoflagellate causing diarrhetic shellfish poisoning. Protist 2002, 153, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.D.; Maranda, L.; Yoon, H.S; Bhattacharya, D. Phylogenetic evidence for the cryptophyte origin of the plastid of Dinophysis (Dinophysiales, Dinophyceae). J. Phycol. 2003, 39, 440–448. [Google Scholar] [CrossRef]
- Janson, S.; Granéli, E. Genetic analysis of the psbA gene from single cells indicates a cryptomonad origin of the plastid in Dinophysis (Dinophyceae). Phycologia 2003, 42, 473–477. [Google Scholar] [CrossRef]
- Janson, S. Molecular evidence that plastids in the toxin-producing dinoflagellate genus Dinophysis originate from the free-living cryptophyte Teleaulax amphioxeia. Environ. Microbiol. 2004, 6, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Park, M.G.; Kim, S.; Kim, H.S.; Myung, G.; Kang, Y.G.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- Nagai, S.; Nitshitani, G.; Tomaru, Y.; Sakiyama, S.; Kamiyama, T. Predation by the toxic dinoflagellate Dinophysis fortii on the ciliate Myrionecta rubra and observation of sequestration of ciliate chloroplasts. J. Phycol. 2008, 44, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Jaén, D.; Mamán, L.; Domínguez, R.; Martín, E. First report of Dinophysis acuta in culture. Harmful Algal News 2009, 39, 1–2. [Google Scholar]
- Riobó, P.; Reguera, B.; Franco, J.M.; Rodríguez, F. First report of the toxin profile of Dinophysis sacculus Stein from LC-MS analysis of laboratory cultures. Toxicon 2013, 76, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S.; Suzuki, T.; Kamiyama, T. Successful cultivation of the toxic dinoflagellate Dinophysis tripos (Dinophyceae). Plankton Benthos Res. 2013, 8, 171–177. [Google Scholar] [CrossRef]
- Mafra, Jr.L.L.; Nagai, S.; Uchida, H.; Tavares, C.P.S.; Escobar, B.P.; Suzuki, T. Harmful effects of Dinophysis to the ciliate Mesodinium rubrum: implications for prey capture. Harmful Algae 2016, 59, 82–90. [Google Scholar] [CrossRef]
- Nishitani, G.; Nagai, S.; Sakiyama, S.; Kamiyama, T. Successful cultivation of the toxic dinoflagellate Dinophysis caudata (Dinophyceae). Plankton Benthos Res. 2008, 3, 78–85. [Google Scholar] [CrossRef]
- Nishitani, G.; Nagai, S.; Takano, Y.; Sakiyama, S.; Baba, K.; Kamiyama, T. Growth characteristics and phylogenetic analysis of the marine dinoflagellate Dinophysis infundibulus (Dinophyceae). Aquat. Microb. Ecol. 2008, 52, 209–221. [Google Scholar] [CrossRef]
- Kim, S.; Kang, Y.G.; Kim, H.S.; Yih, W.; Coats, D.W.; Park, M.G. Growth and grazing responses of the mixotrophic dinoflagellate Dinophysis acuminata as functions of light intensity and prey concentration. Aquat. Microb. Ecol. 2008, 51, 301–310. [Google Scholar] [CrossRef]
- Kim, M.; Nam, S.W.; Shin, W.; Coats, D.W.; Park, M.G. Dinophysis caudata (Dinophyceae) sequesters and retains plastids from the mixotrophic ciliate prey Mesodinium rubrum. J. Phycol. 2012, 48, 569–279. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, T.; Suzuki, T. Production of dinophysis-1 and pectenotoxin-2 by a culture of Dinophysis acuminata (Dinophyceae). Harmful Algae 2009, 8, 312–317. [Google Scholar] [CrossRef]
- Hackett, J.D.; Tong, M.M.; Kulis, D.M.; Fux, E.; Hess, P.; Bire, R.; Anderson, D.M. DSP toxin production de novo in cultures of Dinophysis acuminata (Dinophyceae) from North America. Harmful Algae 2009, 8, 873–879. [Google Scholar] [CrossRef]
- Kamiyama, T.; Nagai, S.; Suzuki, T.; Miyamura, K. Effect of temperature on production of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 by Dinophysis acuminata in culture experiments. Aquat. Microb. Ecol. 2010, 60, 193–202. [Google Scholar] [CrossRef]
- Tong, M.; Kulis, D.M.; Fux, E.; Smith, J.L.; Hess, P.; Zhou, Q.; Anderson, D.M. The effects of growth phase and light intensity on toxin production by Dinophysis acuminata from northeastern United States. Harmful Algae 2011, 10, 254–264. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Nishikawa, T.; Kamiyama, T. Differences in the production and excretion kinetics of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan. J. Phycol. 2011, 47, 1326–13337. [Google Scholar] [CrossRef]
- Basti, L., Uchida, H., Kanamori, M., Matsushima, R., Suzuki, T., Nagai, S. Mortality and pathology of Japanese scallop, Patinopecten (Mizuhopecten) yessoensis, and noble scallop, Mimachlamys nobilis, fed monoclonal culture of PTX-producer, Dinophysis caudata. In Marine and Freshwater Harmful Algae; In MacKenzie, A.L.; Ed., Proceedings of the 16th International Conference on Harmful Algae, Wellington, New Zealand. Cawthron Institute, Nelson, New Zealand and International Society for the Study of Harmful Algae, Wellington, New Zealand, 2015, pp. 105–108.
- Basti, L.; Uchida, H.; Matsushima, R.; Watanabe, R.; Suzuki, T.; Yamatogi, T.; Nagai, S. Influence of temperature on growth and production of Pectenotoxin-2 by a monoclonal culture of Dinophysis caudata. Mar. Drugs 2015, 13, 7124–7137. [Google Scholar] [CrossRef]
- Basti, L.; Suzuki, T.; Uchida, H.; Kamiyama, T.; Nagai, S. Thermal acclimation affects growth and lipophilic toxin production in a strain of cosmopolitan harmful alga Dinophysis acuminata. Harmful Algae 2018, 73, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Gobler, J.C.; Dohrty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. PNAS 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed]
- Guillotreau, P. ; Le Bihan, V. ; Morineau, B., Pardo, S. The vulnerability of shellfish farmers to HAB events: An optimal matching analysis of closure decrees. Harmful Algae 2021, 101, 101968. [CrossRef] [PubMed]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.J.; Janson, S.; Boje, R.; Pollenhne, F.; Chang, J. The dinoflagellate Dinophysis norvegica: biological and ecological observation in the Baltic Sea. Eur. J. Phycol. 1995, 30, 1–9. [Google Scholar] [CrossRef]
- Okolodkov, Y.B.; Dodge, J.D. Biodiversity and biogeography of planktonic dinoflagellates in the Arctic Ocean. J. Exp. Mar. Biol. Ecol. 1996, 202, 19–27. [Google Scholar] [CrossRef]
- Meyer-Harms, B.; Pollenhe, F. Alloxanthin in Dinophysis norvegica (Dinophysiales, Dinophyceae) from the Baltic Sea. J. Phycol. 1998, 34, 280–285. [Google Scholar] [CrossRef]
- Bresnan, E.; Fryer, R.; Hart, M.; Percy, L. Correlation Between Algal Presence in Water and Toxin Presence in Shellfish. Fisheries Research Services Contract Report 04/05, 2005, 27 pp.
- Jansen, S.; Riser, C.W.; Wassmann, P.; Bathmann, U. Copepod feeding behavior and egg production during a dinoflagellate bloom in the North Sea. Harmful Algae 2006, 5, 102–112. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful Algal Bloom Communities in Scottish Coastal Waters: Relationship to Fish Farming and Regional Comparisons – A Review. Natural Scotland Scottish Executive, 2006.
- Whyte, C.; Swan, S.; Davidson, K. Changing wind patterns linked to unusually high Dinophysis blooms around the Shetland Islands, Scotland. Harmful Algae 2014, 39, 365–373. [Google Scholar] [CrossRef]
- Fabro, E.; Almandoz, G.O.; Ferrario, M.; Tillmann, U.; Cembella, A.; Krock, B. Distribution of Dinophysis species and their association with lipophilic phycotoxins in plankton from the Argentine Sea. Harmful Algae 2016, 59, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Shahi, N.; Nayak, B.B.; Mallik, S.K. Dinophysis norvegica: First report of the toxic temperate water Dinophysis in Manori creek of Mumbai water. Harmful Algae News 2010, 42, 14–15. [Google Scholar]
- Hernández-Becerril, D.U. La Diversidad del Fitoplancton Marino de México. Un Acercamiento Actual. In Planctología Mexicana; Barreiro, M.T.; Meave, M.E.; Signoret, M.; Figueroa, M.G.; Eds., Sociedad Mexicana de Planctología (SOMPAC), Universidad Autónoma Metropolitana, México, 2003, pp. 1–17.
- Suba Rao, D.V.; Pan, Y.; Zitko, V.; Bugden, G.; Mackelgan, K. Diarrhetic shellfish poisoning (DSP) associated with a subsurface bloom of Dinophysis norvergica in Bedford Basin, eastern Canada. Mar. Ecol. Prog. Ser. 1993, 97, 117–126. [Google Scholar] [CrossRef]
- Cembella, A.D. Occurrence of okadaic acid, a major diarrhetic shellfish toxin, in natural populations of Dinophysis spp. from the eastern of North America. J. Appl. Phycol. 1989, 1, 307–310. [Google Scholar] [CrossRef]
- Goto, H.; Igarashi, T.; Watai, M.; Yasumoto, T.; Gomez, O.V.; Valdivia, G.L.; Noren, F.; Gisselson, L.A.; Granéli, E., Worldwide occurrence of pectenotoxins and yessotoxins in shellfish and phytoplankton. In Ninth International Conference on Harmful Algal Blooms, Tasmania, Australia, 2000, p. 20 (Abstract).
- Pimiä, V.; Kankaanpaä, H.; Kononen, K. The first observation of okadaic acid in Mytilus edulis from the Gulf of Finland. Boreal Environ. Res. 1998, 2, 381–385. [Google Scholar]
- Sipiä, V.; Kankaanpaä, H.; Meriluoto, J. The first observation of okadaic acid in flounder in the Baltic Sea. Sarsia 2000, 85, 471–475. [Google Scholar] [CrossRef]
- Deeds, J.R.; Stutts, W.L.; Celiz, M.D.; mac Leod, J.; Hamilton, A.E.; Lewis, B.J.; Miller, D.W.; Kanwit, K.; Smith, J.L.; Kulis, D.M.; McCarron, P.; Rauschenberg, C.D.; Burnell, C.A.; Archer, S.D.; Brochert, J.; Lnakford, S.K. Dihydrodinophysistoxin-1 produced by Dinophysis norvegica in the Gulf of Main, USA and its accumulation in shellfish. Toxins 2020, 12, 533. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T; Miyazono, A.; Baba, K.; Sugawara, R.; Kamiyama, T. LC-MS/MS analysis of okadaic acid analogues and other lipophilic toxins in single-cell isolates of several Dinophysis species collected in Hokkaido, Japan. Harmful Algae 2009, 8, 233–238. [CrossRef]
- Uchida, H.; Watanabe, R.; Matsushima, R.; Oikawa, H.; Nagai, S.; Kamiyma, T.; Baba, K.; Miyazono, A.; Kosaka, K.; Kaga, S.; Matsuyama, Y.; Suzuki, T. Toxin profiles of okadaic acid analogues and other lipophilic toxins in Dinophysis from Japanese coastal waters. Toxins 2018, 10, 457. [Google Scholar] [CrossRef]
- Balech, E. 1976. Some Norwegian Dinophysis species (Dinoflagellata). Sarsia 61: 75-94.
- Taylor, F.J.R., Y. Fukuyo and J. Larsen 1995. Taxonomy of harmful dinoflagellates. In: G.M. Hallegraeff, D.M. Anderson and A.D. Cembella (eds), Manual on Harmful Marine Microalgae, IOC Manuals and Guides No. 33. UNESCO, France: 283-317.
- IOC-UNESCO. The Harmful Algal Event Database (HAEDAT). http://haedat.iode.org/index.php [accessed: 18.02.2023].
- Martino, S.; Gianella, F.; Davidson, K. An approach for evaluating the economic impacts of harmful algal blooms: The effects of blooms of toxic Dinophysis spp. on the productivity of Scottish shellfish farms. Harmful Algae 2020, 99, 101912. [Google Scholar] [CrossRef]
- Séchet, V.; Sibat, M.; Billien, G.; Carpentier, L.; Rovillon, G. A.; Raimbault, V.; Malo, F.; Gaillard, S.; Perrière-Rumebe, M.; Hess, P.; Chomérat, N. Characterization of toxin-producing strains of Dinophysis spp. (Dinophyceae) isolated from French coastal waters, with a particular focus on the D. acuminata-complex. Harmful Algae 2021, 107, 101974. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.P.L., 1975. Culture of phytoplankton for feeding marine invertebrates. In: Smith, W.L., Chanley, M.H. (Eds.), Culture of Marine Inveretbrate Animals. Springer, Boston.
- Nagai, S.; Matsuyama, Y.; Oh, S-J.; Itakura, S. Effect of nutrients and temperature on encystment of the toxic dinoflagellate Alexandrium tamarense (Dinophyceae) isolated from Hiroshima Bay, Japan. Plankton Biology and Ecology 2004, 51(2), 103-109.
- Guillard, R.R.L.; Kilham, P.; Jackson, T.A. 1973. Kinetics of silicon-limited growth in the marine diatom Thalassiosira pesudonana Hasle and Heimdal (= Cyclotella nana Husted). Journal of Phycology 1973, 9, 233–237. [Google Scholar]
- Nagai, S.; Yamamoto, K.; Hata, N.; Itakura, S. Study of DNA extraction methods for use in loop-mediated isothermal amplification detection of single resting cysts in the toxic dinoflagellates Alexandrium tamarense and A. catenella. Marine Genomics 2012, 7, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C. A; Herzog, M; Sogin, M; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. Journal of Phycology 1994, 30(6), 999-1011.
- Larsson, A. AliView: a fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C. K. T. Mega X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D. L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M. A.; Huelsenbeck, J. P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic biology 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Suzuki, T.; Mitsuya, T.; Imai, M.; Yamasaki, M. DSP toxin contents in Dinophysis fortii and scallops collected at Mutsu Bay Japan. Journal of Applied Phycolology 1997, 8, 509–515. [Google Scholar] [CrossRef]
- Suzuki, T.; Mitsuya, T.; Matsubara, H.; Yamasaki, M. Determination of pectenotoxin-2 after solid-phase extraction from seawater and from the dinoflagellate Dinophysis fortii by liquid chromatography with electrospray mass spectrometry and ultraviolet detection: evidence of oxidation of pectenotoxin-2 to pectenotoxin-6 in scallops. Journal of Chromatography A, 1998, 815, pp. 155–160. [Google Scholar]
- Suzuki, T.; Quilliam, M. LC-MS/MS analysis of Diarrhetic Shellfish Poisoning (DSP) toxins, okadaic acid and dinophysistoxin analogues, and other lipophilic toxins. Analytical Sciences 2011, 27, 571–584. [Google Scholar] [CrossRef]
- Albinsson M.E.; Negri A.P.; Blackburn S.I.; Bolch C.J.S. Bacterial Community Affects Toxin Production by Gymnodinium catenatum. PLoS ONE 2014, 9(8): e104623.
Figure 1.
Micrographs of a vegetative cell of Dinophysis norvegica in culture in a bright field (left) and with fluorescence under blue light excitation (right). Plastids of D. norvegica emit red autofluoresecence. Scale bar, 10 µm.
Figure 1.
Micrographs of a vegetative cell of Dinophysis norvegica in culture in a bright field (left) and with fluorescence under blue light excitation (right). Plastids of D. norvegica emit red autofluoresecence. Scale bar, 10 µm.
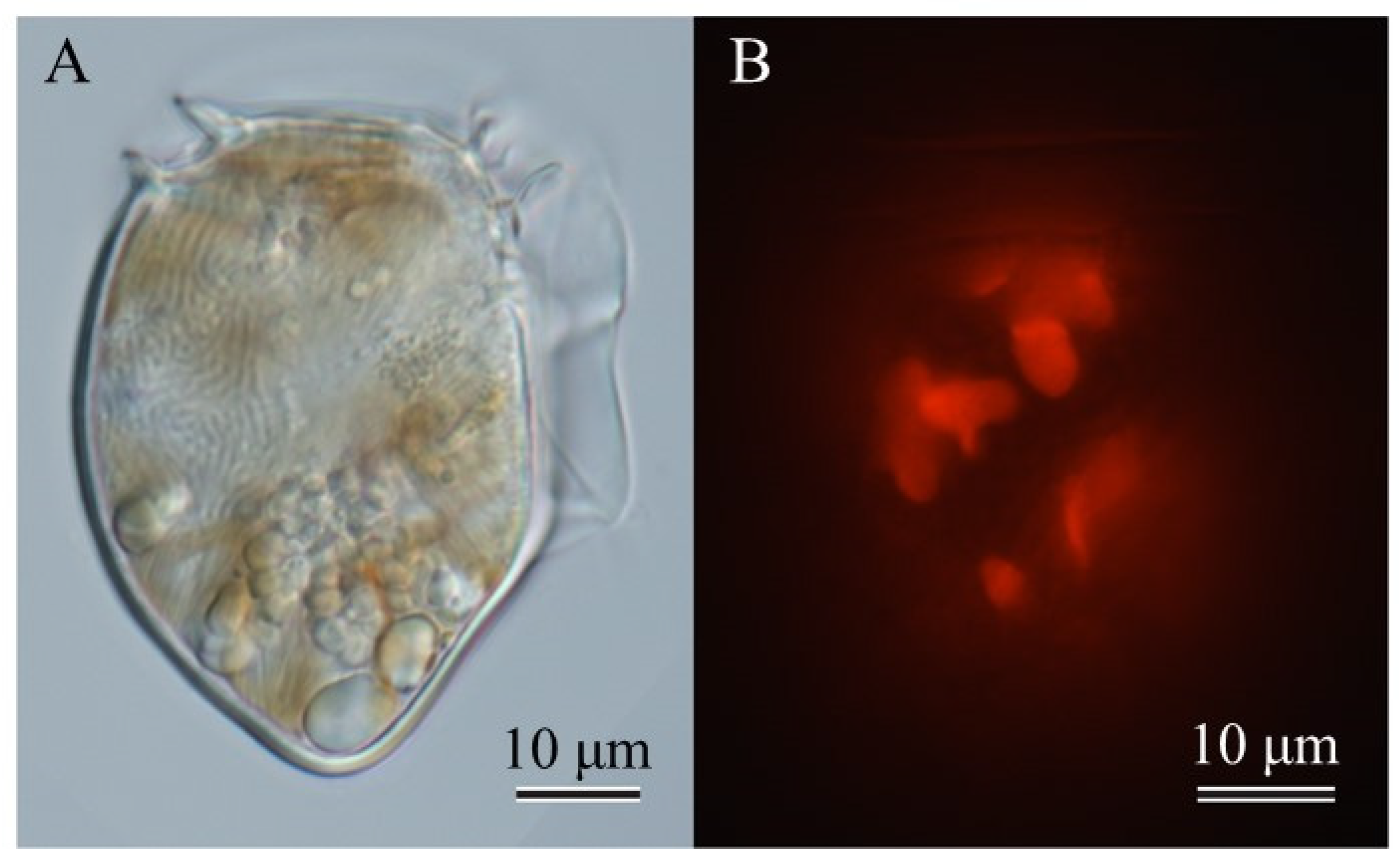
Figure 2.
Maximum likelihood (ML) tree of Dinophysis norvegica inferred from D1/D2 region (735 b.p.). Bootstrap supports of ML and neighbor-joining (NJ) and posterior probabilities (PP) of Bayesian inference are indicated at node (ML/NJ/PP). Culture strains obtained in this study are highlighted in bold.
Figure 2.
Maximum likelihood (ML) tree of Dinophysis norvegica inferred from D1/D2 region (735 b.p.). Bootstrap supports of ML and neighbor-joining (NJ) and posterior probabilities (PP) of Bayesian inference are indicated at node (ML/NJ/PP). Culture strains obtained in this study are highlighted in bold.

Figure 3.
Growth of strain DN16062021FUN-06 of Dinophyis norvegica in culture with Mesodinium rubrum.
Figure 3.
Growth of strain DN16062021FUN-06 of Dinophyis norvegica in culture with Mesodinium rubrum.
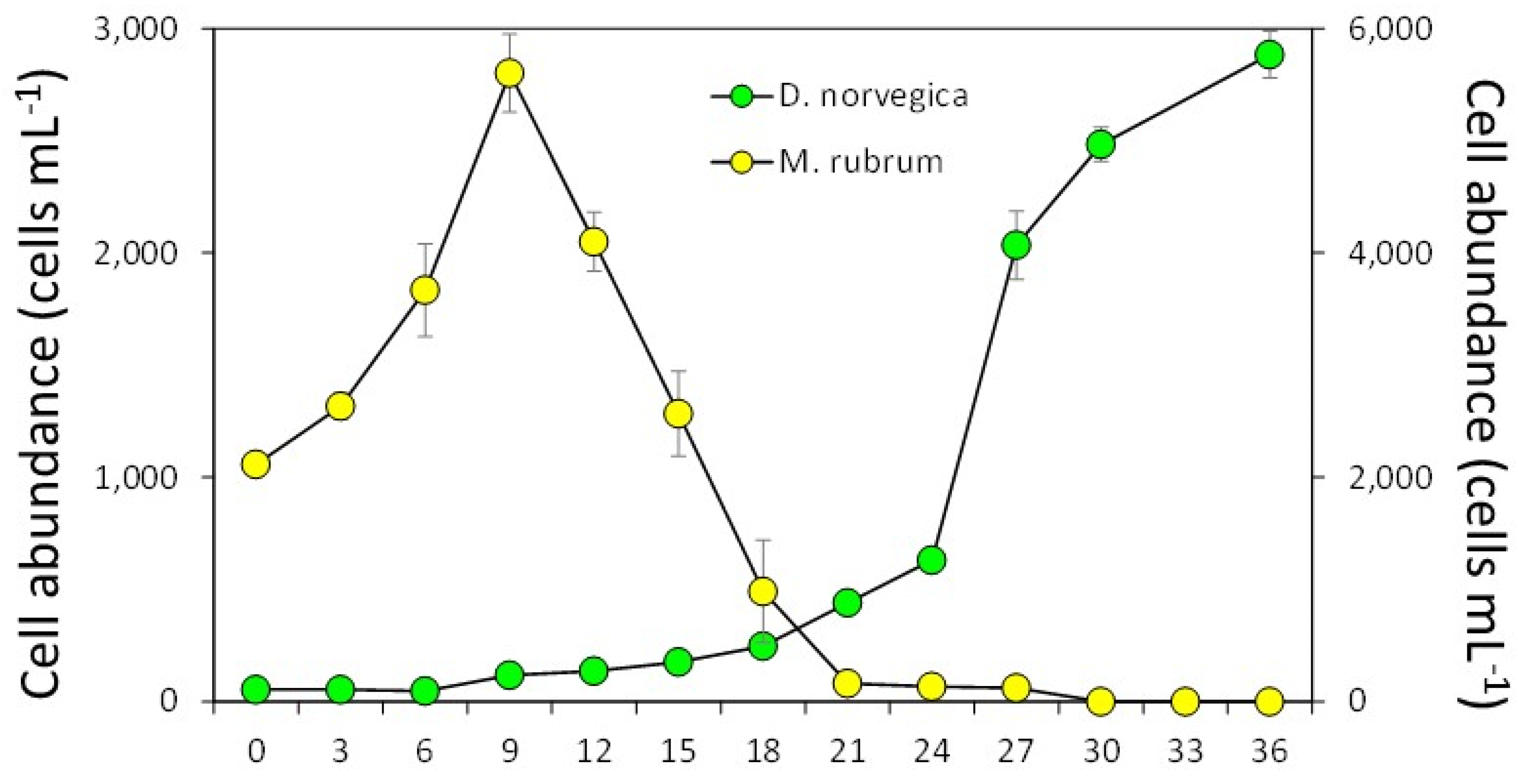
Figure 4.
Toxin production of strain DN16062021FUN-06 of Dinophysis norvegica in culture with Mesodinium rubrum. (A) Dinophysistoxin-1; (B) Pectenotoxin-2.
Figure 4.
Toxin production of strain DN16062021FUN-06 of Dinophysis norvegica in culture with Mesodinium rubrum. (A) Dinophysistoxin-1; (B) Pectenotoxin-2.
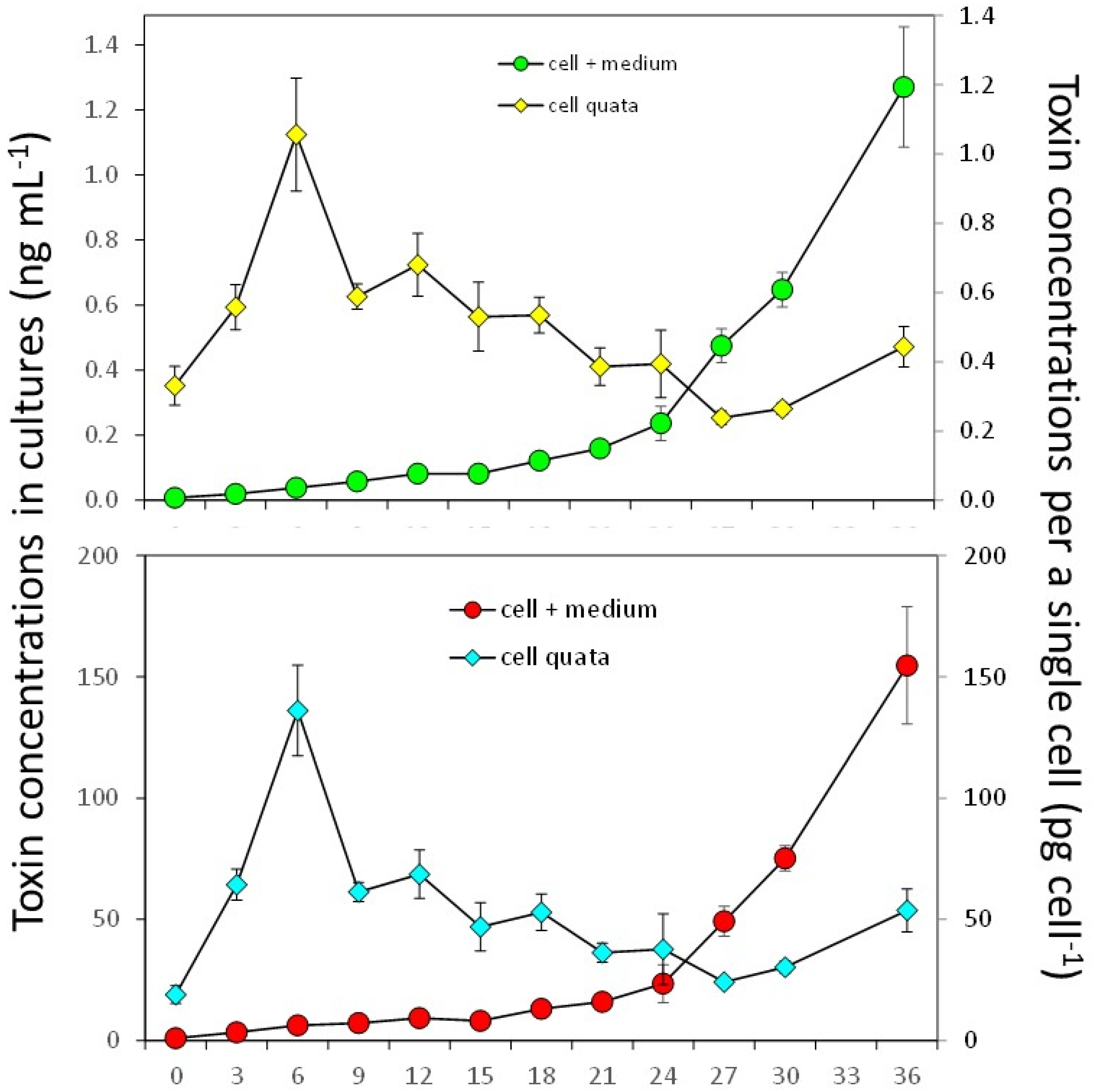

Table 1.
Toxin production in seven cultured strains of Dinophysis norvegica.
| Strains | Concentration in culture (ng ml-1) |
Number of cells |
Cell quota (pg ml-1) |
||||
|---|---|---|---|---|---|---|---|
| OA | DTX1 | PTX2 | OA | DTX1 | PTX2 | ||
| DN01 | ND (<0.1) | ND (<0.1) | 137 | 1807 | ND (<0.2) | ND (<0.2) | 75.8 |
| DN02 | ND (<0.1) | ND (<0.1) | 126 | 2080 | ND (<0.2) | ND (<0.2) | 60.6 |
| DN03 | ND (<0.1) | ND (<0.1) | 145 | 2333 | ND (<0.2) | ND (<0.2) | 62.1 |
| DN05 | ND (<0.1) | 1.44 | 375 | 2850 | ND (<0.2) | 0.5 | 131.6 |
| DN06 | ND (<0.1) | 3.55 | 316 | 3050 | ND (<0.2) | 1.2 | 103.6 |
| DN07 | ND (<0.1) | ND (<0.1) | 161 | 1057 | ND (<0.2) | ND (<0.2) | 152.4 |
| DN08 | Trace | 0.705 | 132 | 936 | Trace | 0.7 | 137.0 |
OA: Okadaic acid; DTX1: Dinophysistoxin-1; PTX2: Pectenotoxin-2; ND: not detected (limit of detection), Trace: a signal to noise ratio was about 3 by the LC/MS/MS, but lower than the limit of detection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



