
Submitted:
04 March 2023
Posted:
06 March 2023
You are already at the latest version
Abstract
Research and development on innovative packaging materials have advanced significantly to safeguard packaged food against microbial contamination and oxidation. To combat demanding issues, active packaging has evolved as a viable method for minimizing oxidation/microbial growth in packaged goods, extending their shelf life, and ensuring the consumer's safety. Active food packaging includes O2, CO2 scavengers, moisture absorbers, U.V. barriers, and carriers of antioxidant and antimicrobial agents. Various antimicrobial agents are carried and/or incorporated into food packaging formulations. Consumers demand natural antimicrobials over chemical/synthetic ones, such as bacteriocins, bacteriophages, and essential oils. Bacteriophages (viruses) have emerged as a feasible option for decontaminating and eliminating infections from food sources. These viruses can target specific food-borne pathogens without impairing beneficial bacteria and, most critically, without causing disease in humans or animals. Fortifying bacteriophages into food packaging films will not only kill specific food microorganisms but has evolved as a new weapon to combat antimicrobial-resistant (AMR) issues. The present review summarises recent developments in active antimicrobial packaging focused particularly on bacteriophage-food packaging applications and advantages, drawbacks, and future trends for active food packaging.
Keywords:
Virus-fortification
; antimicrobial
; active packaging films
; bacteriophages
1. Introduction
The innovation in the food packaging sector over the past few decades, driven by the more demanding and changing desire of consumers and food packaging industries, led to the evolution of advanced packaging technologies with augmented protection attributes more than conventional packages [1,2,3]. Conventional packing materials are considered passive, with their primary role being protection from extraneous environmental surroundings and ease of handling [4].
Packaging materials can be segmented into traditional or passive, active, intelligent, and smart [4,5]. Out of the listed, active packaging materials show more attention as they can actively react to the packaged foods’ internal and external environmental changes [6]. Currently, most active packaging materials are broadly classified as antioxidants-based, antimicrobials-based, gas scavengers/emitters based, etc. [7]. Most active-antimicrobial packaging constituents/materials are broad-spectrum antimicrobial agents that do not target specific bacterial pathogens; hence, there is an urgent need to fabricate antimicrobial materials with high (host) specificity to target only pathogenic organisms without hampering beneficial bacterial population [8]. Specificity in antimicrobial activity is crucial since pathogens may only make up a tiny percentage of the total microbial load in food systems. [9]. In addition, many recent advances in dairy and nutraceutical-based foods rely on the presence of non-pathogenic microorganisms. Therefore, it may be possible to increase antibacterial potential by limiting interactions with non-targeted bacteria if pathogen-specific antibacterial active packing materials can be developed [4,6,10].
According to the centers for disease control and prevention (CDC), following just four easy steps-namely, “clean,” “separate,” “cook,” and “chill,” can considerably control foodborne illness at household-level [11]. However, with a growing new generation of microbial threats, including antimicrobial resistance, consumer demands are pushing the scientist and food industry toward strategies to increase sensory attributes, shelf life, real-time monitoring, packaging, and improving overall quality characteristics of the foods [12,13].
In view of the above discussions, one potential approach recently attracted much attention is bacteriophages, a green and sustainable nano-tool targeting specific pathogenic bacteria without impacting beneficial microbiota [9,14]. In 1896, a British bacteriologist named Ernest Hanbury Hankin discovered that water purified from India’s Ganges and Jamuna rivers had the bactericidal activity of bacteriophages against Vibrio cholerae, publishing this work in the Annals of the Pasteur Institute [15,16,17]. In 1915, Frederick Twort described the antimicrobial efficacies of bacteriophages while studying the growth studies of the vaccinia virus on culture media (cell-free). After 2 years, in 1917, the scientist Flix d’Herelle used bacteriophages for therapeutic purposes to treat dysentery [17]. However, mainstream research nearly neglected phages due to antibiotic discoveries. Later in the 1980s, the inactivation of E. coli using phage in mice confirmed bacteriophages’ better efficacy than antibiotics [18].
Among the different forms of active packaging materials, employing bacteriophages as antimicrobial agents have attracted much interest [19]. Numerous brief reviews on active packaging features have been documented recently. The present review focuses on recent advances in applying bacteriophages as active agents in biopolymer-based packaging.
An attempts to highlight the recent research findings on food packaging films/coatings by fortifying them with bacteriophages to fulfill the need of the hour, considering the rising AMR issues worldwide has been done.
2. Understanding bacteriophages
Bacteriophages or phages are viruses that infect bacteria, varying in size (10-250 nm) and shape, as depicted in Figure 1 [20]. Bacteriophages are considered the most common creatures on the earth because they are present in any habitat containing the bacteria that serve as their hosts, and are ubiquitous [18,21]. The vast majority of bacteriophages, approximately 96% of those discovered to date, are placed in the order of Caudovirales [22]. This order comprises bacteriophages with tails and double-stranded DNA, and bacteriophages usually infect their bacterial hosts in a species- or even strain-specific manner [23,24]. Based on the length of their life cycles, they may be classified as either virulent or temperate phages [22]. Virulent phages result in the lytic cycle where the phage binds/attaches itself to its bacterial host by injecting its genome, starts further multiplications by utilizing the host’s cellular machinery, and lyses the host cell, concurrently releasing its posterity [25]. Lysins and holins are two types of proteins commonly used by lytic phages to destroy their host cell [23,24]. The holins puncture the bacterial cytoplasmic layer and work as a synergy tool for the endolysins, destroying the bacteria’s cell wall. On the other end, temperate phages infect the host by opening a lysogenic cycle, in which the phage genome remains latent as a prophage, replicates with their host, and may sporadically burst into a lytic phage under specific activations [26,27].
The Bacterial and Archaeal Subcommittee (BAVS), which is part of the International Committee on Taxonomy of Viruses (ICTV), is responsible for categorizing and classifying bacteriophages [28,29,30,31,32,33]. The classification is based on the different characteristic properties that a bacteriophage possesses, such as the type of nucleic acid that it uses as its genetic material (DNA or RNA), the structure of its capsid (tailed, polyhedral, filamentous, or pleomorphic), its activity spectrum against various hosts, their sequence similarity, and their pathogenicity [22]. About 95% of all known bacteriophages belong to the Caudovirales, sometimes known as the “Order of Tail Phage.” This order comprises the three major families, Myoviridae, Podoviridae, and Siphoviridae, all of which include phages of double-stranded DNA (dsDNA) as their genetic material [34]. Polyhedral viruses comprise five families Microviridae, Corticoviridae, Tectiviridae, Fiersviridae, and Cystoviridae [35]. Filamentous viruses type contains Inoviridae, Lipothrixviridae, and Rudiviridae [36]. Other important pleomorphic viruses comprise Plasmaviridae, Fuselloviridae, Guttaviridae, Ampullaviridae, Bicaudaviridae, and Globuloviridae [30]. Figure 2 details the classification of bacteriophages based on their morphology and nucleic acid content.
3. Bacteriophages applications in various sectors:
Antibiotics have been utilized in animal husbandry and plant pathogen management since World War II. Antibiotic misuse in agriculture has led to an increase in AMR bacteria’s prevalence globally [37,38]. For instance, streptomycin-resistant Erwinia amylovora is becoming a significant issue since it is prevalent in many places where antibiotics are misused, such as pears and apples. This strain of the pathogen is getting more difficult to treat. Bacteriophages are excellent for preventing or reducing animal illnesses (phage treatment), disinfecting raw materials and carcasses, such as fresh vegetables and fruit, cleaning equipment, and rigid contact interfaces [39]. Using phages in place of antibiotics in agricultural practices is a viable alternative to preserve animal and plant health and reduce the spread of AMR and zoonotic diseases that can be dangerous to consumers [39,40].
Necrotizing enterocolitis, which can manifest clinically or asymptomatically in broilers when caused by Clostridium perfringens, is one of the most critical challenges facing the poultry industry [41]. This disease can be effectively controlled by a cocktail of phages (five types) that can also lead to an increased feed conversion rate and overall weight of the chicken [42]. Phages have been utilized as growth promoters in chicken and have also been researched as potential replacements for antibiotics, typically employed for this reason. The method of administration of bacteriophages is important in determining their efficacy against various bacterial strains. Aquaculture has shown phages to be a profitable and environmentally acceptable alternative to antibiotics. Vibrio anguillarum is the most widespread disease of estuarine fish and marine biota caused by Vibriosis [43]. Vibriosis infections cause higher mortality rates in fish, particularly in larvae [44]. The studies reported that V. anguillarum-related infection could be successfully treated using a single phage in Atlantic salmon [43].
Bacteriophages can also be administered against plant pathogenic bacteria to prevent crop diseases and increase yield [45]. The first practical indication that phages may be linked to plant pathogenic bacteria was presented when it was discovered that a filtrate collected from decaying cabbage could suppress cabbage rot caused by Xanthomonas campestris. Xylella fastidiosa is a pathogen of numerous plant species, but its economic impact is highest on grapes [46]. Since the pathogen is confined to the xylem of grapes, disease control strategies are restricted and difficult. In greenhouse studies, phage cocktails were able to significantly reduce the development of pathogens and symptoms in grapes using therapeutic and preventative therapies [46]. Highly variable seasonal fluctuations in biological controls are common and represent one of the biggest obstacles to commercializing phages in agriculture. For this reason, few studies on applying phage as a promising alternative to antibiotics in crop protection exist. Nonetheless, as discussed above, the promising results provide confidence and a need for further research on phage therapy for crop protection in agriculture [45,46].
One of the oldest facilities applying phage therapy to common bacterial diseases associated with urology, gynecology, internal medicine, and pediatrics is the Eliaba Institute of Bacteriophage, Microbiology, and Virology, Georgia [47]. More than ninety-five percent of patients undergoing phage treatment showed significant improvement and recovery without side effects [48]. With the progression of multiple-drug-resistant (MDR) or AMR bacterial strains, phage therapy is gaining popularity again as infected patients are left without effective treatment options. In 2016, Tom Patterson at the University of California contracted a multidrug-resistant Acinetobacter baumannii infection that he could treat using antibiotics [49] and treated for the disease with a successful and effective intravenous bacteriophage. He fully recovered from AMR bacterial infection after phage treatment, becoming the first successful treatment case in the U.S. During the lytic cycle, the adsorption of phage particles on surfaces of the bacterial cell is the 1st step in which the tail fibers attach to specific receptors located on a bacterial cell wall. Viral DNA enters the host through a hollow tube in the tail in the second step, injection. Protein synthesis and host hijacking are the third steps. Viral genes regulate the synthesis of viral proteins by using the host’s machinery. Viral genome synthesis and assembly are steps 4 and 5. The release is performed in step 6 by a viral peptidoglycan hydrolase (endolysin) that triggers the lysis of the host cell and releases up to 200 infectious phages. Despite the exciting therapeutic potential of phages, numerous challenges must be overcome before phage therapy can be used in the clinical setting. These challenges include a narrow/limited host range, poor phage stability in the blood circulation system, safety issues, and commercial viability issues. However, with modern synthetic biology approaches, phage properties can be modified to solve many of the abovementioned problems [48,49,50].
According to target specificity, phages are classified into broad-spectrum and narrow-spectrum bacteriophages [48]. Bacteriophages are highly target-specific and are not known to be detrimental to the human microbiome. Broad-spectrum bacteriophages are multivalent bacteriophages capable of binding to more than one receptor site on the target cell surface, whereas narrow-spectrum bacteriophages are monovalent [50]. They are limited and attach to specific receptor sites. Nonetheless, these two classes of bacteriophages can be engineered/developed and interconverted using point mutations in the phage genome to produce desired changes at the receptor-binding site. This method efficiently solved the bacteriophage’s narrow host range problem.
In addition, various methods have been devised to solve the stability problem related to phage in the body of human beings. Encapsulation of phages in stable materials such as polyethylene glycol (PEG) or liposomes effectively improved the stability of phage particles in the blood circulation system. Encapsulated phages can remain in circulation without being eliminated by the reticuloendothelial system [51]. Furthermore, liquid phages are converted into powder form using spray drying technology to facilitate inhalation in treating respiratory infections. The powder can be formulated into tablets, bandages, and wound dressings. Recent developments in bacteriophage therapy include using phages and phage-acquired products, including endolysins, as antimicrobial agents using complete bacteriophages as a substitute for conventional antibiotics [50,51]. Endolysins are phage proteins that perform specific functions when phages invade bacterial cells. For example, endolysins, such as virion-associated peptidoglycan hydrolases, help break down bacterial cell wall peptidoglycan during the bacteriophage lytic cycle. Endolysins have an advantage over full bacteriophages because their genome is not directly involved in the treatment, eliminating the possibility of mutagenesis. In addition, endolysins do not develop significant resistance, supporting excellent conservation and high host specificity [48,49,50,51].
Food manufacturers employ many multi-methods worldwide to ensure their products’ safety, including heat pasteurization, high-pressure processing (HPP), microwave irradiation, chemical sanitizers, and natural additives/antioxidants, each with their drawbacks [52,53,54]. Thermal pasteurization results in the food being cooked and, therefore, unsuitable for fresh food items. At the same time, high-pressure treatment has deteriorating effects on the nutritional quality and appearance of foods such as fresh produce and meat [53,55,56]. Although irradiation is more effective and superior to the methods discussed above, high-level applications hurt the organoleptic properties of foods [53,57]. Meanwhile, chemical disinfectants and additives erode food processing equipment and decrease consumer acceptance; the demand for pesticide-free organic food grows rapidly [58,59,60,61]. In addition to all these disadvantages, the preservation methods mentioned indiscriminately kill microorganisms, including beneficial ones.
Bacteriophages have also been studied as antimicrobial agents to achieve food safety from microorganisms [62]. Bacteriophage-mediated food safety practices, commonly called “bacteriophage biocontrol,” are gradually emerging and gaining popularity among food technologists, addressing the shortcomings of conventional food preservation methods [63]. Bacteriophage biological control does not affect not only the beneficial microflora of food but also its quality characteristics. Lytic bacteriophages (Wild-type) can be employed as pre-harvest, like in live animals, or may be supplied via animal feed and/or post-harvest, and may be useful to food surfaces in the packaging materials to limit pathogen contamination [64]. Biocontrol employed using bacteriophage has also been shown with disinfecting activities on food processing surfaces. Several surveys on pre-harvest (on farm animals) and post-harvest (on meat, fresh produce, and packaged goods) were conducted. These studies control various endemic and emerging foodborne pathogens, including Salmonella, Listeria, Campylobacter, and Escherichia [62,63]. Various investigators have evaluated the intervention studies addressing phage biological control of food-eating pathogens before and after harvest [18,20,21,24,65].
4. Recent updates on bacteriophage-based food packaging:
“Active packaging” refers to packaging in which supplemental/active ingredients have been purposely added/infused in or on either the packaging matrix/components or in the packages’ headspace to produce the packaging system’s more efficient performance [66,67]. As discussed earlier, various active agents (antimicrobial-based) have been explored to combat foodborne illnesses. The use of phages as antimicrobial agents has increased due to their omnipresence and host specificity [22]. Many commercial producers are participating in developing phage and phage-based derivatives like PhageGuard Listex®, PhageGuard S.®, PhageGuard E.® etc.
Food application-based phage treatment includes dipping, mixing phage solutions, and/or spraying directly into food surfaces. However, these applications (direct) methods often require a high number/quantity of phages to be effective [24,39]. An efficient way to challenge these issues might be fabricating a support basements system on which phages are immobilized/fixed before their controlled release and interaction with the food. This could be a significant advantage for food packaging, preservation, and storage. Figure 3 illustrates methods/techniques for producing phage-based bio-polymers, including casting (a), dipping/spraying (b), extrusion (c), and layer-to-layer (d) as shown in Figure 3.
Bacteriophage release and stability present a significant obstacle for scientists. In food systems, the release of active phages from the polymeric films/coatings/hydrogels occurs much slower than in watery systems [68]. Variability in phage stability has been seen in edible polymeric films and coatings and complete inhibition/suppression of the bacteriophages has been documented [40,68,69]. The exact phenomenon behind the inhibition or inactivation of phage is unclear. Encapsulation of the phages, which would increase their stability and make them more resistant to damage, has been recommended by several researchers to solve these issues.
As reported, incorporating T4 (bacteriophage) using electrospinning (suspension) led to a significantly decreased phage activity [70]. For better bacteriophage viability, they pre-encapsulated T4 in an alginate reservoir into an electrospun fiber, reporting the coaxial electrospinning process, and the activity of bacteriophage could be improved. A core/shell fiber structure was formed in this process, with the T4 bacteriophage directly fused into the fiber core. The core of fiber-encapsulated T4 showed higher bacteriophage viability for several weeks at a temperature of +4°C [70].
Another study on developing a formulation for encapsulating phage K with an improved acid shield for oral delivery [71]. They encapsulated the calcium carbonate (microparticles) with phage K into alginate microspheres for better phage survivability within in vitro acidic environments. Free phages (without encapsulations) were killed by exposure to a gastric fluid of pH 2.5. They reported that the viability of encapsulated phage K in SGF was enhanced by adding calcium carbonate to the alginate microspheres, with only a 0.17 log decrease after two hours of exposure to SGF at pH 2.5. In contrast, alginate-encapsulated phage K decreased to only 2.4 log in survivability when incubated for one hour in SGF under pH 2.5 [71]. E. coli was efficiently suppressed by an antibacterial film created by immobilizing phage T4 on a poly-caprolactone (PCL) film [72]. After being used as a packaging film for beef (raw) infected with E. coli O157:H7, fabricated PCL film showed 30-fold microbial inhibitory properties than the film containing physically adsorbed phage T4. These findings suggested that the developed PCL film incorporated with phage T4 has a good potential application as an active food packaging against E. coli.
Table 1 illustrates the recent studies on bacteriophage-based bio-polymeric films/coating for active packaging applications.
The team of researchers [80] studied the manufacturing of bacteriophage IBB-PF7A fabricated using sodium alginate to prevent microbiological meat spoiling caused by Pseudomonas fluorescens. They claimed that the bacteriophages had been loaded efficiently in films with significant vitality. They found that the number of P. fluorescens organisms dropped by 2 Logs during the first two days of storage in the refrigerator and then only dropped by 1 Log over the subsequent five days [80]. The film’s effectiveness as an antibacterial agent was established by artificially inoculating chicken breast fillets with P. fluorescens. In another study [73], the competence of acetate cellulose film added with bacteriophage against Salmonella typhimurium and observed the increased lag phase, thereby, slower bacterial growth in the environment containing bacteriophages with the films as compared to control (without phage). No significant changes were observed in the films’ mechanical and physical properties, like thickness, elongation, and puncture resistance after adding bacteriophages. However, bacteriophages remained viable in films only for 14 days after that, not detected in the acetate cellulose film [73].
Scientists developed prototypes of bioactive packaging materials based on immobilized bacteriophages to control bacterial pathogens’ growth in foods [96]. Phage-based compounds had substantial antibacterial effects when applied to artificially contaminated foods. The developed bioactive films could inhibit L. monocytogenes in ready-to-eat meat under different storage conditions by using specific lytic bacteriophage cocktails, either free or immobilized [96]. A team of scientists [90] studied the development of chitosan film embedded with developed phage to control E. coli O157:H7 in beef. Developed chitosan film containing liposome-encapsulated phage exhibited high antimicrobial efficacy against E. coli O157:H7. They observed that phage encapsulation efficacy improved by 57.66%.
5. Summary and future research
Bacteriophage and bacteriophage-derived bio-polymeric/edible coatings and films have emerged as a substitute for traditional food packaging to address various emerging issues like bacterial host specificity, AMR, etc. Many reports have claimed that adding novel phages into bio-polymeric films/coatings does not change food’s physicochemical properties and sensory qualities. However, it has also been claimed that adding these antibacterial agents leads to changes in films’ mechanical properties. The antibacterial efficacies of bacteriophage-added films and coatings have been successfully tested and proven in food systems like vegetables, meat, fruit, poultry, fish, etc. It is postulated that there are still some challenges to getting full-scale harvesting from this novel strategy to develop bacteriophage-based food packaging films for active packaging applications. Some leading challenges include phage viability/stability, phage mobility into the coatings/film, bacteriophage release from coating/film to a food matrix, and active bacterial population/availability to promote the host-action of phages. However, research has already proved that incorporating phages into films/coating is advantageous for maintaining antibacterial activity. In demand to increase the stability of phages, there is a continuing need for more research into the mechanics of phage release and the strategies of film fabrication.
There is a significant need for more research to develop encapsulation strategies/formulations for various uses along the food supply chain. These products can treat contamination caused by particular bacterial pathogens at various stages throughout the food-producing process, including spraying them, exposing them to livestock before processing, flushing food contact surfaces in production plants, and treating them post-harvest foodstuff. Bacteriophage biocontrol can be a promising tool in a multi-barrier/hurdle strategy to prevent foodborne pathogens from reaching customers. This technique is particularly promising when producers aim to preserve foods’ natural and often beneficial microbial population while targeting/removing only the pathogenic bacteria. Phage technology might enhance food safety by lowering infections in farm animals and limiting microbial burdens in the food supply chain, bioremediation of foodborne microorganisms in food items, or serving as a cleaning agent.
Author Contributions
Conceptualization, R.V.W.; writing—original draft preparation, R.V.W. and R.P.; writing—review and editing, J.W.R.; visualization, R.V.W.; project administration, J.W.R.; funding acquisition, J.W.R. All authors have read and agreed to the published version of the manuscript.
Funding
The work was executed under the financial assistance for the International training provided by the Indian Council of Agricultural Research (ICAR), New Delhi, India through the National Agricultural Higher Education Project (IDP-NAHEP) running at Guru Angad Dev Veterinary and Animal Sciences University, Ludhiana, Punjab (INDIA). This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (2022R1A2B5B02001422) and Brain Pool program funded by the Ministry of Science, ICT and Future Planning through the National Foundation of Korea (2019H1D3A1A01070715).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The author R.V.W. acknowledge the support provided by GADVASU authorities for the international training provided by the Indian Council of Agricultural Research through the National Agricultural Higher Education Project (IDP) running at Guru Angad Dev Veterinary and Animal Sciences University, Ludhiana, Punjab (INDIA).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Priyadarshi, R.; Kim, S.-M.; Rhim, J.-W. Pectin/Pullulan Blend Films for Food Packaging: Effect of Blending Ratio. Food Chem 2021, 347, 129022. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Priyadarshi, R.; Ezati, P.; Rhim, J.W. Curcumin and Its Uses in Active and Smart Food Packaging Applications - a Comprehensive Review. Food Chem 2022, 375, 131885. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshi, R.; Riahi, Z.; Rhim, J.-W.; Han, S.; Lee, S.-G. Sulfur Quantum Dots as Fillers in Gelatin/Agar-Based Functional Food Packaging Films. ACS Appl Nano Mater 2021, 4, 14292–14302. [Google Scholar] [CrossRef]
- Amin, U.; Khan, M.K.I.; Maan, A.A.; Nazir, A.; Riaz, S.; Khan, M.U.; Sultan, M.; Munekata, P.E.S.; Lorenzo, J.M. Biodegradable Active, Intelligent, and Smart Packaging Materials for Food Applications. Food Packag Shelf Life 2022, 33, 100903. [Google Scholar] [CrossRef]
- Cheng, H.; Xu, H.; Julian McClements, D.; Chen, L.; Jiao, A.; Tian, Y.; Miao, M.; Jin, Z. Recent Advances in Intelligent Food Packaging Materials: Principles, Preparation and Applications. Food Chem 2022, 375, 131738. [Google Scholar] [CrossRef] [PubMed]
- Kuai, L.; Liu, F.; Chiou, B. sen; Avena-Bustillos, R.J.; McHugh, T.H.; Zhong, F. Controlled Release of Antioxidants from Active Food Packaging: A Review. Food Hydrocoll 2021, 120, 106992. [Google Scholar] [CrossRef]
- Perera, K.Y.; Jaiswal, S.; Jaiswal, A.K. A Review on Nanomaterials and Nanohybrids Based Bio-Nanocomposites for Food Packaging. Food Chem 2022, 376, 131912. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, S.; Wang, Q.; Lin, G.; Niu, B.; Guo, R.; Yan, H.; Wang, H. Sodium Alginate/Chitosan-Based Intelligent Bilayer Film with Antimicrobial Activity for Pork Preservation and Freshness Monitoring. Food Control 2023, 148, 109615. [Google Scholar] [CrossRef]
- Huang, K.; Nitin, N. Edible Bacteriophage Based Antimicrobial Coating on Fish Feed for Enhanced Treatment of Bacterial Infections in Aquaculture Industry. Aquaculture 2019, 502, 18–25. [Google Scholar] [CrossRef]
- Azman, N.H.; Khairul, W.M.; Sarbon, N.M. A Comprehensive Review on Biocompatible Film Sensor Containing Natural Extract: Active/Intelligent Food Packaging. Food Control 2022, 141, 109189. [Google Scholar] [CrossRef]
- CDC Four Steps to Food Safety-CDC. Available online: https://www.cdc.gov/foodsafety/keep-food-safe.html (accessed on 29 January 2023).
- Hofer, U. The Cost of Antimicrobial Resistance. Nature Reviews Microbiology 2018, 17, 3–3. [Google Scholar] [CrossRef] [PubMed]
- C Reygaert, W. An Overview of the Antimicrobial Resistance Mechanisms of Bacteria. AIMS Microbiol 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter Baumannii Infection. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Orlova, E.V. Bacteriophages and Their Structural Organisation; IntechOpen, 2012; ISBN 978-953-51-0272-4. [Google Scholar]
- Batinovic; Wassef; Knowler; Rice; Stanton; Rose; Tucci; Nittami; Vinh; Drummond; et al. Bacteriophages in Natural and Artificial Environments. Pathogens 2019, 8, 100. [Google Scholar] [CrossRef]
- Abedon, S.T.; Thomas-Abedon, C.; Thomas, A.; Mazure, H. Bacteriophage Prehistory. Bacteriophage 2011, 1, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A. The Future of Bacteriophage Biology. Nat Rev Genet 2003, 4, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.; López, A.; Sáez-Orviz, S.; Marcet, I.; García, P.; Rendueles, M.; Díaz, M. Effectiveness of Bacteriophages Incorporated in Gelatine Films against Staphylococcus Aureus. Food Control 2021, 121, 107666. [Google Scholar] [CrossRef]
- Hendrix, R.W. Bacteriophage Genomics. Curr Opin Microbiol 2003, 6, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.; Streisinger, G.; Horne, R.W.; Champe, S.P.; Barnett, L.; Benzer, S.; Rees, M.W. Structural Components of Bacteriophage. J Mol Biol 1959, 1, 281–292. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Phage Classification and Characterization. 2009; pp. 127–140.
- Ackermann, H.W. 5500 Phages Examined in the Electron Microscope. Arch Virol 2007, 152, 227–243. [Google Scholar] [CrossRef]
- Ackermann, H.W. Frequency of Morphological Phage Descriptions in 1995. Arch Virol 1996, 141, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.W.; Prangishvili, D. Prokaryote Viruses Studied by Electron Microscopy. Arch Virol 2012, 157, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Hay, I.D.; Lithgow, T. Filamentous Phages: Masters of a Microbial Sharing Economy. EMBO Rep 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, H.M. ICTV Virus Taxonomy Profile: Corticoviridae. Journal of General Virology 2017, 98, 888–889. [Google Scholar] [CrossRef] [PubMed]
- Prangishvili, D.; Mochizuki, T.; Krupovic, M. ICTV Virus Taxonomy Profile: Guttaviridae. Journal of General Virology 2018, 99, 290–291. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M. ICTV Virus Taxonomy Profile: Plasmaviridae. Journal of General Virology 2018, 99, 617–618. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, H.M. ICTV Virus Taxonomy Profile: Corticoviridae. Journal of General Virology 2017, 98, 888–889. [Google Scholar] [CrossRef] [PubMed]
- Poranen, M.M.; Mäntynen, S. ICTV Virus Taxonomy Profile: Cystoviridae. Journal of General Virology 2017, 98, 2423–2424. [Google Scholar] [CrossRef]
- Prangishvili, D.; Krupovic, M. ICTV Virus Taxonomy Profile: Bicaudaviridae. Journal of General Virology 2018, 99, 864–865. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Tailed Bacteriophages: The Order Caudovirales. 1998; pp. 135–201.
- Chipman, P.R.; Agbandje-McKenna, M.; Renaudin, J.; Baker, T.S.; McKenna, R. Structural Analysis of the Spiroplasma Virus, SpV4: Implications for Evolutionary Variation to Obtain Host Diversity among the Microviridae. Structure 1998, 6, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Prangishvili, D.; Arnold, H.P.; Götz, D.; Ziese, U.; Holz, I.; Kristjansson, J.K.; Zillig, W. A Novel Virus Family, the Rudiviridae: Structure, Virus-Host Interactions and Genome Variability of the Sulfolobus Viruses SIRV1 and SIRV2. Genetics 1999, 152, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Kochhar, R. The Virus in the Rivers: Histories and Antibiotic Afterlives of the Bacteriophage at the Sangam in Allahabad. Notes and Records: the Royal Society Journal of the History of Science 2020, 74, 625–651. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, V.; Nair, A.; Mallya, R.; Khan, T.; Omri, A. Antimicrobial Nanomaterials for Food Packaging. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin Microbiol Rev 2019, 32. [Google Scholar] [CrossRef]
- López de Dicastillo, C.; Settier-Ramírez, L.; Gavara, R.; Hernández-Muñoz, P.; López Carballo, G. Development of Biodegradable Films Loaded with Phages with Antilisterial Properties. Polymers (Basel) 2021, 13, 327. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.W.; Skinner, J.; Sulakvelidze, A.; Mathis, G.F.; Hofacre, C.L. Bacteriophage Therapy for Control of Necrotic Enteritis of Broiler Chickens Experimentally Infected with Clostridium Perfringens. Avian Dis 2010, 54, 33–40. [Google Scholar] [CrossRef]
- Keerqin, C.; McGlashan, K.; Van, T.T.H.; Chinivasagam, H.N.; Moore, R.J.; Choct, M.; Wu, S.-B. A Lytic Bacteriophage Isolate Reduced Clostridium Perfringens Induced Lesions in Necrotic Enteritis Challenged Broilers. Front Vet Sci 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Plaza, N.; Castillo, D.; Pérez-Reytor, D.; Higuera, G.; García, K.; Bastías, R. Bacteriophages in the Control of Pathogenic Vibrios. Electronic Journal of Biotechnology 2018, 31, 24–33. [Google Scholar] [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio Anguillarum Infections in Fish Larvae Production. PLoS One 2014, 9, e114197. [Google Scholar] [CrossRef]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The Future of Phage Biocontrol in Integrated Plant Protection for Sustainable Crop Production. Curr Opin Biotechnol 2021, 68, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Kutateladze, M.; Adamia, R. Bacteriophages as Potential New Therapeutics to Replace or Supplement Antibiotics. Trends Biotechnol 2010, 28, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Kutateladze, M. Experience of the Eliava Institute in Bacteriophage Therapy. Virol Sin 2015, 30, 80–81. [Google Scholar] [CrossRef] [PubMed]
- Zolnikov, T.R. Global Health in Action Against a Superbug. Am J Public Health 2019, 109, 523–524. [Google Scholar] [CrossRef]
- Azam, A.H.; Tan, X.-E.; Veeranarayanan, S.; Kiga, K.; Cui, L. Bacteriophage Technology and Modern Medicine. Antibiotics 2021, 10, 999. [Google Scholar] [CrossRef]
- Raza, A.; jamil, M.; Tahir Aleem, M.; Aamir Aslam, M.; Muhammad Ali, H.; khan, S.; Kareem, N.; Asghar, T.; Gul, K.; Nadeem, H.; et al. Bacteriophage Therapy: Recent Development and Applications. Scholars Bulletin 2021, 7, 27–37. [Google Scholar] [CrossRef]
- Singh, S.; Shalini, R. Effect of Hurdle Technology in Food Preservation: A Review. Crit Rev Food Sci Nutr 2016, 56, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Chiozzi, V.; Agriopoulou, S.; Varzakas, T. Advances, Applications, and Comparison of Thermal (Pasteurization, Sterilization, and Aseptic Packaging) against Non-Thermal (Ultrasounds, UV Radiation, Ozonation, High Hydrostatic Pressure) Technologies in Food Processing. Applied Sciences 2022, 12, 2202. [Google Scholar] [CrossRef]
- Jagtap, N.S.; Wagh, R. v.; Chatli, M.K.; Malav, O.P.; Kumar, P.; Mehta, N. Functional Goat Meat Nuggets Fortified with Novel Bioactive Carica Papaya L. and Origanum Vulgare Extracts and Storage Stability Thereof. Nutr Food Sci 2019, 50, 402–414. [Google Scholar] [CrossRef]
- Abera, G. Review on High-Pressure Processing of Foods. Cogent Food Agric 2019, 5, 1568725. [Google Scholar] [CrossRef]
- Wagh, R. v.; Chatli, M.K. Response Surface Optimization of Extraction Protocols to Obtain Phenolic Rich Antioxidant from Sea Buckthorn and Their Potential Application into Model Meat System. J Food Sci Technol 2017, 54, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J. Irradiation as a Method for Decontaminating Food. Int J Food Microbiol 1998, 44, 189–204. [Google Scholar] [CrossRef]
- Kumar, V.; Chatli, M.K.; Wagh, R. v.; Mehta, N.; Kumar, P. Effect of the Combination of Natural Antioxidants and Packaging Methods on Quality of Pork Patties during Storage. J Food Sci Technol 2015, 52, 6230–6241. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, N.S.; Wagh, R. v.; Chatli, M.K.; Malav, O.P.; Kumar, P.; Mehta, N. Chevon Meat Storage Stability Infused with Response Surface Methodology Optimized Origanum Vulgare Leaf Extracts. Agricultural Research 2020, 9, 663–674. [Google Scholar] [CrossRef]
- Jagtap, N.S.; Wagh, R. v.; Chatli, M.K.; Kumar, P.; Malav, O.P.; Mehta, N. Optimisation of Extraction Protocol for Carica Papaya L. to Obtain Phenolic Rich Phyto-Extract with Prospective Application in Chevon Emulsion System. J Food Sci Technol 2019, 56, 71–82. [Google Scholar] [CrossRef]
- Wagh, R.V.; Chatli, M.K.; Ruusunen, M.; Puolanne, E.; Ertbjerg, P. Effect of Various Phyto-Extracts on Physico-Chemical, Colour, and Oxidative Stability of Pork Frankfurters. Asian-Australas J Anim Sci 2015, 28, 1178–1186. [Google Scholar] [CrossRef]
- Tauxe, R. v. Emerging Foodborne Pathogens. Int J Food Microbiol 2002, 78, 31–41. [Google Scholar] [CrossRef]
- Hameed, F.; Bandral, J.D.; Gupta, N.; Nayik, G.A.; Sood, M.; Rahman, R. Use of Bacteriophages as a Target Specific Therapy against Food-Borne Pathogens in Food Industry-a Review: Bacteriophage. Journal of microbiology, biotechnology and food sciences 2022, 11, e2949–e2949. [Google Scholar] [CrossRef]
- Sulakvelidze, A. Using Lytic Bacteriophages to Eliminate or Significantly Reduce Contamination of Food by Foodborne Bacterial Pathogens. J Sci Food Agric 2013, 93, 3137–3146. [Google Scholar] [CrossRef]
- Ackermann, H.W. Frequency of Morphological Phage Descriptions in the Year 2000. Arch Virol 2001, 146, 843–857. [Google Scholar] [CrossRef]
- Malhotra, B.; Keshwani, A.; Kharkwal, H. Antimicrobial Food Packaging: Potential and Pitfalls. Front Microbiol 2015, 6. [Google Scholar] [CrossRef]
- Motelica, L.; Ficai, D.; Ficai, A.; Cristian Oprea, O.; Durmu¸s, D.; Kaya, A.; Andronescu, E. Biodegradable Antimicrobial Food Packaging: Trends and Perspectives. Foods. [CrossRef] [PubMed]
- López de Dicastillo, C.; Settier-Ramírez, L.; Gavara, R.; Hernández-Muñoz, P.; López Carballo, G. Development of Biodegradable Films Loaded with Phages with Antilisterial Properties. Polymers (Basel) 2021, 13, 327. [Google Scholar] [CrossRef]
- Radford, D.; Guild, B.; Strange, P.; Ahmed, R.; Lim, L.-T.; Balamurugan, S. Characterization of Antimicrobial Properties of Salmonella Phage Felix O1 and Listeria Phage A511 Embedded in Xanthan Coatings on Poly(Lactic Acid) Films. Food Microbiol 2017, 66, 117–128. [Google Scholar] [CrossRef]
- Korehei, R.; Kadla, J. Incorporation of T4 Bacteriophage in Electrospun Fibres. J Appl Microbiol 2013, 114, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Pacan, J.C.; Wang, Q.; Sabour, P.M.; Huang, X.; Xu, Y. Enhanced Alginate Microspheres as Means of Oral Delivery of Bacteriophage for Reducing Staphylococcus Aureus Intestinal Carriage. Food Hydrocoll 2012, 26, 434–440. [Google Scholar] [CrossRef]
- Choi, I.; Yoo, D.S.; Chang, Y.; Kim, S.Y.; Han, J. Polycaprolactone Film Functionalized with Bacteriophage T4 Promotes Antibacterial Activity of Food Packaging toward Escherichia Coli. Food Chem 2021, 346, 128883. [Google Scholar] [CrossRef] [PubMed]
- Gouvêa, D.M.; Mendonça, R.C.S.; Soto, M.L.; Cruz, R.S. Acetate Cellulose Film with Bacteriophages for Potential Antimicrobial Use in Food Packaging. LWT - Food Science and Technology 2015, 63, 85–91. [Google Scholar] [CrossRef]
- Vonasek, E.; Le, P.; Nitin, N. Encapsulation of Bacteriophages in Whey Protein Films for Extended Storage and Release. Food Hydrocoll 2014, 37, 7–13. [Google Scholar] [CrossRef]
- Salalha, W.; Kuhn, J.; Dror, Y.; Zussman, E. Encapsulation of Bacteria and Viruses in Electrospun Nanofibres. Nanotechnology 2006, 17, 4675–4681. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Han, P.; Dong, S.; Li, H. Preparation and Application of Bacteriophage-Loaded Chitosan Microspheres for Controlling Lactobacillus Plantarum Contamination in Bioethanol Fermentation. RSC Adv 2015, 5, 69886–69893. [Google Scholar] [CrossRef]
- Boggione, D.M.G.; Batalha, L.S.; Gontijo, M.T.P.; Lopez, M.E.S.; Teixeira, A.V.N.C.; Santos, I.J.B.; Mendonça, R.C.S. Evaluation of Microencapsulation of the UFV-AREG1 Bacteriophage in Alginate-Ca Microcapsules Using Microfluidic Devices. Colloids Surf B Biointerfaces 2017, 158, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Radford, D.; Guild, B.; Strange, P.; Ahmed, R.; Lim, L.-T.; Balamurugan, S. Characterization of Antimicrobial Properties of Salmonella Phage Felix O1 and Listeria Phage A511 Embedded in Xanthan Coatings on Poly(Lactic Acid) Films. Food Microbiol 2017, 66, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Amarillas, L.; Lightbourn-Rojas, L.; Angulo-Gaxiola, A.K.; Basilio Heredia, J.; González-Robles, A.; León-Félix, J. The Antibacterial Effect of Chitosan-based Edible Coating Incorporated with a Lytic Bacteriophage against Escherichia Coli O157:H7 on the Surface of Tomatoes. J Food Saf 2018, 38. [Google Scholar] [CrossRef]
- Alves, D.; Marques, A.; Milho, C.; Costa, M.J.; Pastrana, L.M.; Cerqueira, M.A.; Sillankorva, S.M. Bacteriophage ΦIBB-PF7A Loaded on Sodium Alginate-Based Films to Prevent Microbial Meat Spoilage. Int J Food Microbiol 2019, 291, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Kimmelshue, C.; Goggi, A.S.; Cademartiri, R. The Use of Biological Seed Coatings Based on Bacteriophages and Polymers against Clavibacter Michiganensis Subsp Nebraskensis in Maize Seeds. Sci Rep 2019, 9, 17950. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Nitin, N. Edible Bacteriophage Based Antimicrobial Coating on Fish Feed for Enhanced Treatment of Bacterial Infections in Aquaculture Industry. Aquaculture 2019, 502, 18–25. [Google Scholar] [CrossRef]
- Lin, L.; Zhu, Y.; Cui, H. Inactivation of Escherichia Coli O157:H7 Treated by Poly-L-lysine-coated Bacteriophages Liposomes in Pork. J Food Saf 2018, 38. [Google Scholar] [CrossRef]
- Novello, J.; Sillankorva, S.; Pires, P.; Azeredo, J.; Wanke, C.H.; Tondo, E.C.; Bianchi, O. Inactivation of Pseudomonas Aeruginosa in Mineral Water by DP1 Bacteriophage Immobilized on Ethylene-vinyl Acetate Copolymer Used as Seal Caps of Plastic Bottles. J Appl Polym Sci 2020, 137, 49009. [Google Scholar] [CrossRef]
- Choi, I.; Yoo, D.S.; Chang, Y.; Kim, S.Y.; Han, J. Polycaprolactone Film Functionalized with Bacteriophage T4 Promotes Antibacterial Activity of Food Packaging toward Escherichia Coli. Food Chem 2021, 346, 128883. [Google Scholar] [CrossRef]
- Kim, S.; Chang, Y. Anti-Salmonella Polyvinyl Alcohol Coating Containing a Virulent Phage PBSE191 and Its Application on Chicken Eggshell. Food Research International 2022, 162, 111971. [Google Scholar] [CrossRef]
- Weng, S.; López, A.; Sáez-Orviz, S.; Marcet, I.; García, P.; Rendueles, M.; Díaz, M. Effectiveness of Bacteriophages Incorporated in Gelatine Films against Staphylococcus Aureus. Food Control 2021, 121, 107666. [Google Scholar] [CrossRef]
- Stipniece, L.; Rezevska, D.; Kroica, J.; Racenis, K. Effect of the Biopolymer Carrier on Staphylococcus Aureus Bacteriophage Lytic Activity. Biomolecules 2022, 12, 1875. [Google Scholar] [CrossRef]
- Cui, H.; Yang, X.; Li, C.; Ye, Y.; Chen, X.; Lin, L. Enhancing Anti-E. Coli O157:H7 Activity of Composite Phage Nanofiber Film by D-Phenylalanine for Food Packaging. Int J Food Microbiol 2022, 376, 109762. [Google Scholar] [CrossRef] [PubMed]
- Kamali, S.; Yavarmanesh, M.; Habibi Najafi, M.B.; Koocheki, A. Development of Whey Protein Concentrate/Pullulan Composite Films Containing Bacteriophage A511: Functional Properties and Anti-Listerial Effects during Storage. Food Packag Shelf Life 2022, 33, 100902. [Google Scholar] [CrossRef]
- Vonasek, E.L.; Choi, A.H.; Sanchez, J.; Nitin, N. Incorporating Phage Therapy into WPI Dip Coatings for Applications on Fresh Whole and Cut Fruit and Vegetable Surfaces. J Food Sci 2018, 83, 1871–1879. [Google Scholar] [CrossRef]
- Kalkan, S. Vibrio Parahaemolyticus ATCC 17802 Inactivation by Using Methylcellulose Films Containing Encapsulated Bacteriophages. Turk J Vet Anim Sci 2018, 42, 480–485. [Google Scholar] [CrossRef]
- Tomat, D.; Soazo, M.; Verdini, R.; Casabonne, C.; Aquili, V.; Balagué, C.; Quiberoni, A. Evaluation of an WPC Edible Film Added with a Cocktail of Six Lytic Phages against Foodborne Pathogens Such as Enteropathogenic and Shigatoxigenic Escherichia Coli. LWT 2019, 113, 108316. [Google Scholar] [CrossRef]
- Choi, I.; Lee, J.-S.; Han, J. Maltodextrin-Trehalose Miscible System-Based Bacteriophage Encapsulation: Studies of Plasticizing Effect on Encapsulated Phage Activity and Food Application as an Antimicrobial Agent. Food Control 2023, 146, 109550. [Google Scholar] [CrossRef]
- Kim, S.; Chang, Y. Anti-Salmonella Polyvinyl Alcohol Coating Containing a Virulent Phage PBSE191 and Its Application on Chicken Eggshell. Food Research International 2022, 162, 111971. [Google Scholar] [CrossRef] [PubMed]
- Lone, A.; Anany, H.; Hakeem, M.; Aguis, L.; Avdjian, A.-C.; Bouget, M.; Atashi, A.; Brovko, L.; Rochefort, D.; Griffiths, M.W. Development of Prototypes of Bioactive Packaging Materials Based on Immobilized Bacteriophages for Control of Growth of Bacterial Pathogens in Foods. Int J Food Microbiol 2016, 217, 49–58. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Typical structure of bacteriophages under the microscope and schematically.
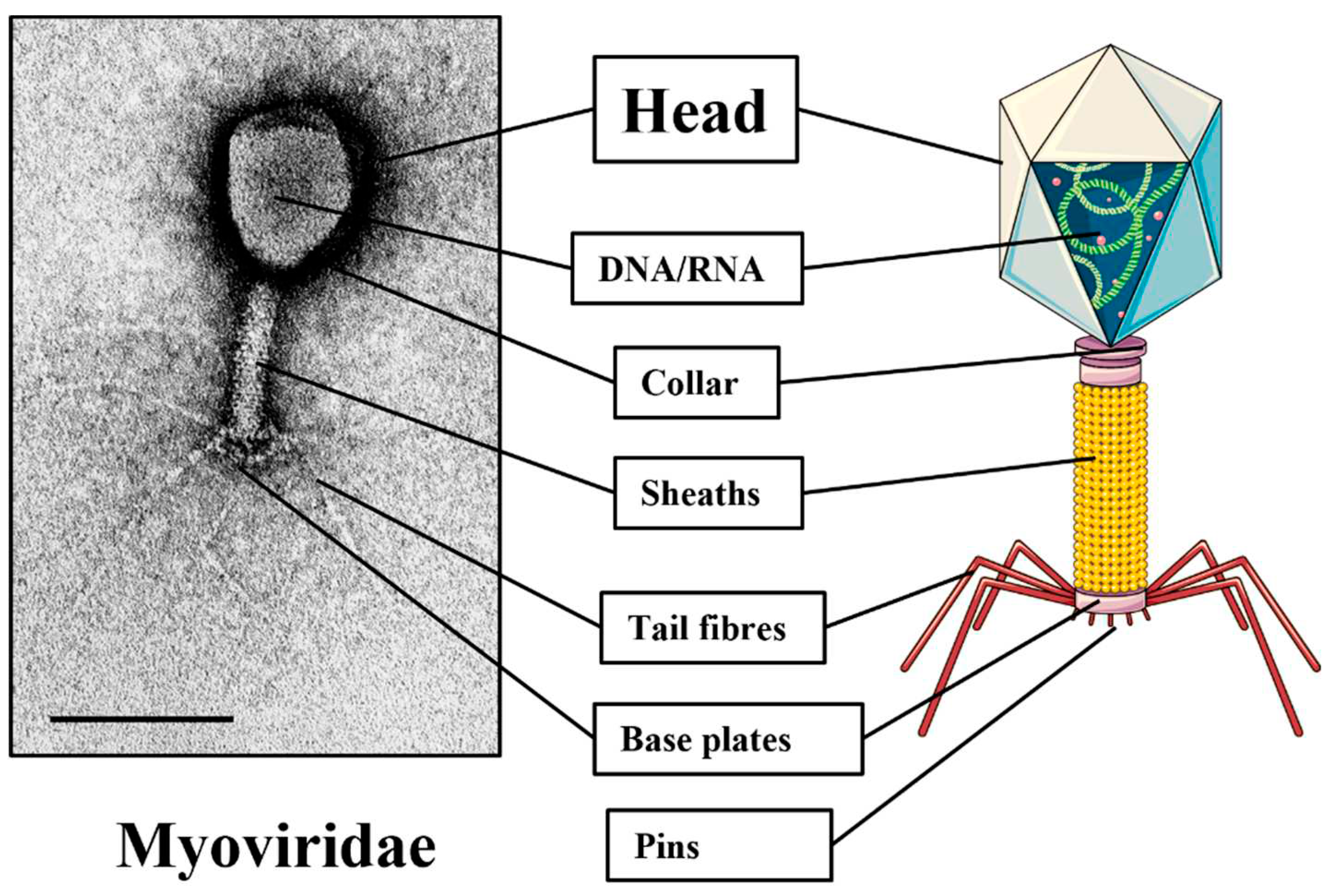
Figure 2.
Classification of bacteriophages based on their morphology and nucleic acid content.
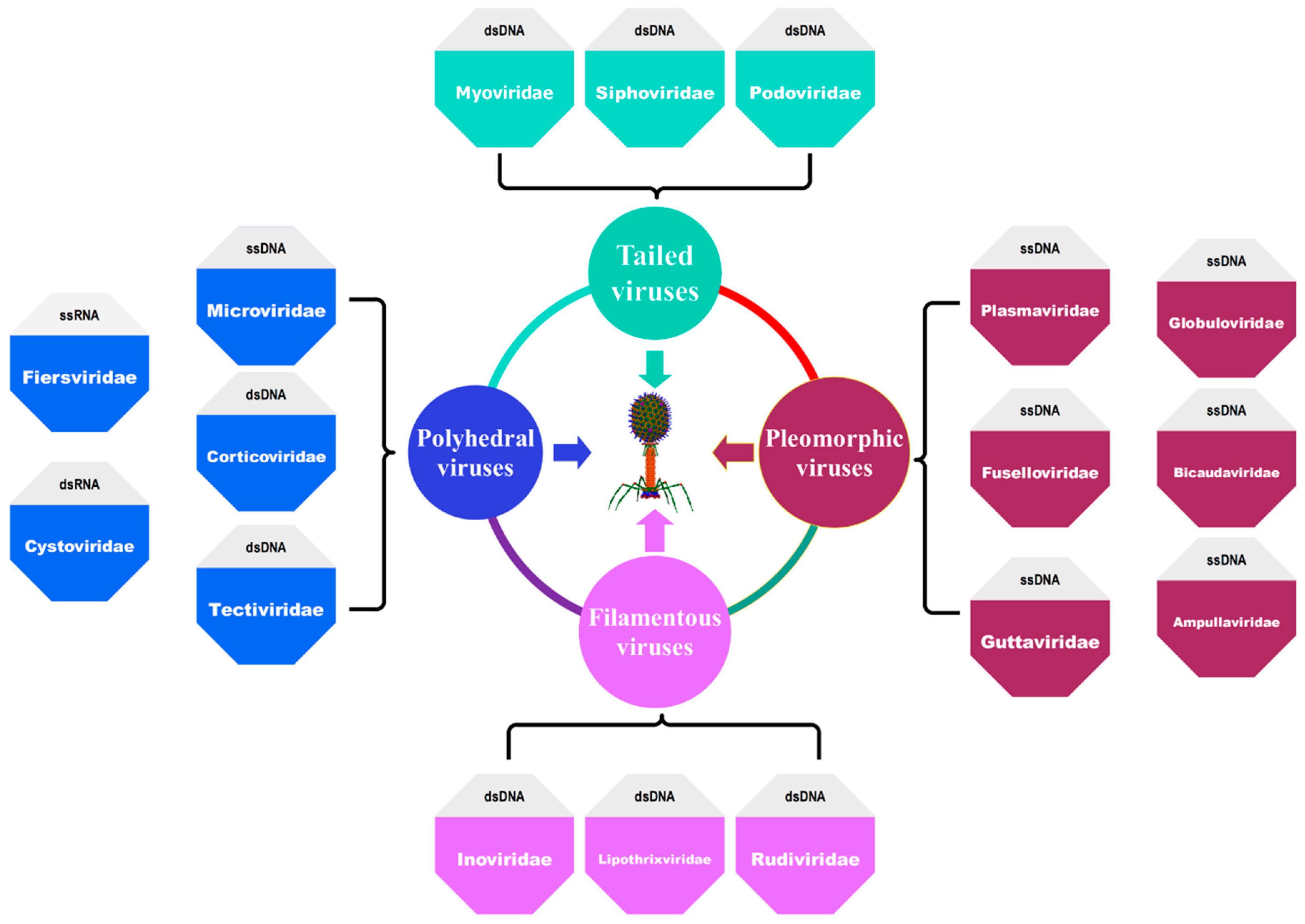
Figure 3.
Techniques for fabrications of bacteriophage-based coatings/films using bio-polymers.
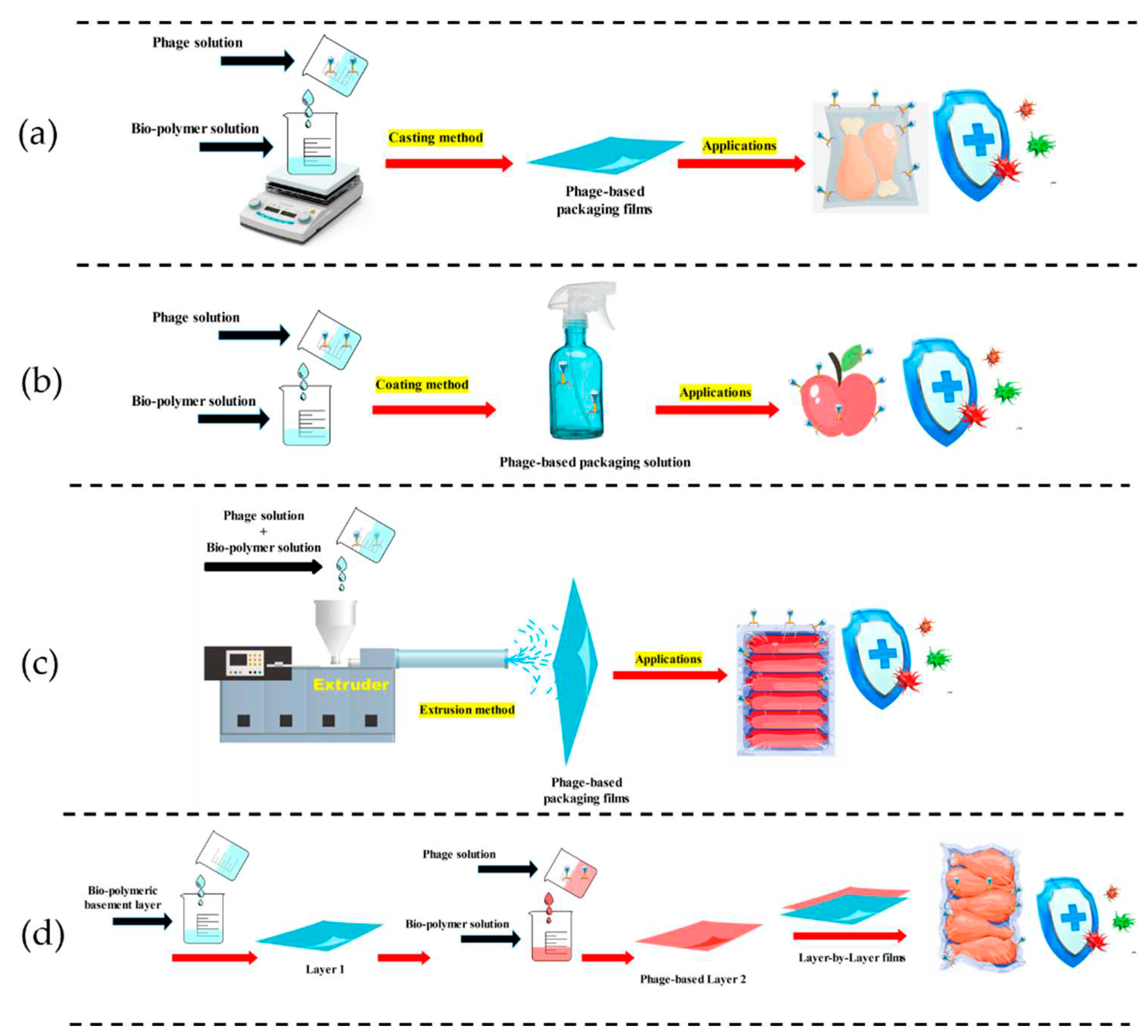

Table 1.
Various reports on bacteriophage-based bio-polymeric active packaging applications.
| Bacteriophage/ Cocktails |
Targeted pathogens |
Bio/ Polymer matrix |
Application | References |
|---|---|---|---|---|
| BFSE16, BFSE18, PaDTA1, PaDTA9, PaDTA10 and PaDTA11 | Salmonella enterica subsp. enterica serovar Typhimurium ATCC 14028 | Acetate cellulose film | Active | [73] |
| T4 bacteriophage | E. coli BL21 | Whey protein films | Active | [74] |
| T7, T4, λ | Escherichia coli, Staphylococcus albus | Poly (vinyl alcohol) | Active | [75] |
| Lactobacillus plantarum bacteriophage | L. plantarum | Chitosan microspheres | Active | [76] |
| LinM-AG8, LmoM-AG13, and LmoM-AG20 | L. monocytogenes and E. coli O104:H4 | Cellulose membranes/ alginate beads | RTE food | [76] |
| UFV-AREG1 | Escherichia coli O157:H7 | Calcium alginate matrix | - | [77] |
| Salmonella phage Felix/Listeria phage A511 | S. Typhimurium and L. monocytogenes cultures. | Poly(lactic acid) | Precooked sliced turkey breast | [78] |
| vB_EcoM34X, vB_EcoSH2Q and vB_EcoMH2W | E. coli O157:H7 CECT 4076 | Chitosan | Tomato | [79] |
| ϕIBB-PF7A | Pseudomonas fluorescens | Sodium alginate | Skinless chicken breast fillets | [80] |
| CN8 bacteriophages | Clavibacter michiganensis subsp. nebraskensis | Polyvinyl polymers with alcohol | Zea mays L. seeds. | [81] |
| T7 phages (#BAA-1025-B2) | E. coli BL21 | Whey protein isolate | Fish feed | [82] |
| E. coli O157:H7 bacteriophages | Escherichia coli O157:H7 | Poly-L-lysine | Pork suspension | [83] |
| vB_PaeM_CEB_DP1 | Pseudomonas aeruginosa | Ethylene-vinyl acetate | Mineral water bottles | [84] |
| Phage T4 | E. coli O157:H7 | Polycaprolactone film | Beef | [85] |
| FO1 | S. Enteritidis | Electrospun PHBV/nanofiber/coating films | ||
| PBSE191 | S. Enteritidis | Polyvinyl alcohol | Active | [86] |
| PhiIPLA-RODI | Staphylococcus aureus | Gelatine | Cheese | [87] |
| Pyo bacteriophages/ Staph bacteriophages | S. aureus | Chitosan and alginate | - | [88] |
| E. coli O157 | Escherichia coli O157:H7 | Sodium alginate /polyethylene oxide (PEO) nanofibers | Beef, cucumber, and cherry tomato | [89] |
| Listeria phage A511 | Listeria monocytogenes 19113 | Whey protein concentrate/pullulan | - | [90] |
| T7 bacteriophages | Escherichia coli BL21 | Whey protein isolate (WPI) | Coating | [91] |
| V. parahaemolyticus-derived phages | Vibrio parahaemolyticus ATCC 17802 | Methylcellulose | films | [92] |
| T-even type, DT1 to DT6 | E. coli DH5α | Whey protein concentrate | Fish fillets | [93] |
| Phage T4 | Escherichia coli K12 | Maltodextrin and trehalose as encapsulating agents | Nutrient broth, skimmed milk, and beef juices | [94] |
| PBSE191 | Salmonella Enteritidis | Polyvinyl alcohol (PVA) film | Chicken eggshell | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



