
Submitted:
01 March 2023
Posted:
06 March 2023
You are already at the latest version
Abstract
Corals are an important habitat for a diverse fauna which is especially rich in branching species. Each colony harbors several species and dozens of individuals forming an integrated symbiotic community. Factors affecting the diversity and abundance of coral symbionts are poorly understood. The present study tested experimentally the relationship between the population density of coral Pocillopora verrucosa and the diversity and abundance of symbionts in planted coral fragments exposed during three months. Colony size and bleaching were also considered as two additional factors. We demonstrated that even at the early stage of the colonies’ formation, the diversity of obligate symbiontson on these corals was higher than in any other studied local faunas of the World Ocean.Species richness and abundance of symbionts were positively related to colony size and density, while no impact of bleaching was found. Species richness and abundance of obligate symbionts increased along with the host population density, while in facultative symbionts, these indexes decreased. We hypothesized that the negative effect of the host density on species richness and abundance of facultative symbionts is caused by the competitive pressure of obligates, which increases with their abundance. Reasons for the different responses of obligate and facultative symbionts to host density were considered.

Keywords:
Symbionts Pocillopora verrucose
; Diversity
; Species richness
; Abundance
; Obligate sym-biont
; Facultative symbiont
1. Introduction
Almost all scleractinian corals provide shelter to various invertebrate and fish species [1,2]. Some of them cannot survive without the host corals (obligate symbionts), while others can be free-living (facultative symbionts) [1]. Due to their extremely high abundance and biomass, symbionts are a key component of the coral reef food webs and, by extension, the entire reef ecosystem [3,4]. Another important role of symbionts in reef ecosystems is maintaining coral fitness [5,6,7,8] and protecting them from predator attacks [9,10,11,12,13]. The great diversity of these organisms is partly attributed to the habitat diversity provided by scleractinian corals [14], which in turn depends on the growth form of corals and is higher in branching than in massive, laminate, or encrusting coral colonies [1]. Accordingly, the highest diversity of symbionts has been recorded in association with branching acroporid and pocilloporid corals [1,15]. In some cases, the number of species and individuals inhabiting one coral head reaches 10–20 species and 70–80 individuals, respectively, thus forming complex symbiotic communities [16,17,18].
The species composition and structure of coral symbiotic communities may depend on various environmental factors, such as the morphology and size of the host colony [19,20,21], host density [20,22,23,24], surface chlorophyll a [24], wave energy [25,26], flow regime [21,24] and others. The relationship of some of them, i.e., the size and morphology of the host colony, with the characteristics of the community is well documented [18,19,20,21,22,27,28], and it should be considered as a methodological basis for the analysis of coral symbiotic communities. At the same time the impact of other specific environmental factors on the structure of symbiotic communities is poorly studied or controversial [20,22,24]. Specifically, a comparison of the diversity and abundance of large crustaceans inhabiting coral Madracis auretenra (previously named M. mirabilis) in two sites with different colonies densities showed that these measures were higher in communities associated with individual isolated colonies than with continuous aggregations. However, this effect may also be associated with the shape of the colonies, which is different in isolated heads and continuous aggregations [20]. According to Counsell et al. [24] the density of Pocillopora meandrina colonies did not have a significant effect on cryptofaunal species richness and showed minimal variation in species-specific responses. On the contrary, a comparison of Pocillopora damicornis symbiont communities showed that their abundance was higher on the reef flat, where the density of corals was higher than on the reef slope [22]. These differences were explained by the high pressure of predators on the symbionts during their host-to-host migrations on the reef slope, where the distances between coral colonies are higher, which leads to a decrease in their abundance. However, the authors [22] believe that the density of hosts is not the only factor affecting the abundance of symbionts since the compared colonies were located in different biotopes subject to the influence of various environmental factors [22]. A trend towards increased diversity and abundance in coral aggregations, which was more distinct for obligate symbionts, was also shown for another coral species, Pocillopora verrucosa [23]. Thus, the available data on the relationship between the spatial arrangement of host corals and the structure of their symbiotic communities are contradictory and require further investigation.
Another factor that may have a significant impact on the symbiotic community is the bleaching phenomenon. Coral bleaching associated with anthropogenic climate change is one of the most important factors responsible for the degradation of coral reefs [29,30]. Bleaching can affect the physiological state of symbiotic fish and invertebrates, increase intra- and interspecific competition, and decrease population density of symbionts [30,31,32,33,34,35], which, in turn, implies a change in the structure of the symbiotic community as a whole. However, the response of the entire symbiotic community to bleaching events remains unclear.
This work aimed to experimentally test the hypothesis of a positive relationship between the population density of hosts and the diversity and abundance of their symbionts. Taking into account that obligate and facultative symbionts can respond differently to changes in the density of the host population, we consider the effect of density both on the symbiotic community as a whole and on these two components of the community separately. Since short-term bleaching happened in the area of research during the experiment, an assessment of its impact on the symbiotic community was performed as well.
2. Materials and Methods
2.1. Host coral and sampling location
The most diverse symbiont communities are associated with branching corals of the Acroporidae and Pocilloporidae families, which form complex three-dimensional structures with numerous well-protected microbiotopes. Of the nearly 900 species known as coral symbionts, about 250 species are associated with corals of the genus Pocillopora [1]. In addition, species of Pocillopora are fast-growing, commonly used in aquaculture [36], and are widely distributed in Nha Trang Bay, Vietnam, where our study was conducted. Therefore, we chose one of the species of the genus, Pocillopora cf. verrucosa (Ellis & Solander, 1786) (hereafter P. verrucosa), as the object of study (Figure 1A). The symbiont fauna of Pocillopora ssp. has been well documented during the last decades for several regions of the Indo-West Pacific [15,28,37,38,39], including Nha Trang Bay, where 16 species of obligate and 28 species of facultative symbionts were noted in association with P. verrucosa [23,40,41].
Experimental studies were carried out from April 26 to July 23, 2020, in Dam Bay Research Station coastal area located in Nha Trang Bay of the South China Sea, Vietnam (Figure 2). Fixed rectangular nurseries for corals’ planting were installed on the sandy bottom among bioherms at a depth of 3 m (Figure 1B and Figure 2C). Planted colonies were raised above the ground to a height of 40 cm.
A temperature logger (HOBO U22-001) was deployed at a depth of 3 m to monitor the temperature regime in the area of the experiment. It recorded the temperature two times per day. During the observation period the daily temperatures varied from 25.3 to 31.2°C. Seasonal upwelling mitigates thermal anomalies [42], thus the average daily temperatures exceeded 30°C only six days in June and three days in the first decade of July 2020 (Figure S1).
2.2. Experimental design
For the experiment, fragments of coral branches 4-6 cm high were collected from the large colonies grown in the same area at the coral nursery. Fragments were measured before planting and sampled coral colonies. The volume between coral branches is the major factor influencing the number of animals inhabiting a coral, as this is the space where the animals live. As demonstrated earlier, this volume is highly correlated with the total coral colony volume calculated by multiplying the height and the large and small diameters [43]. Therefore, in this study, we used these three parameters to determine the volume of colonies, as it speeds up measurements when processing a large number of colonies. The fragments were then washed in an emulsion of clove oil (15 drops per liter) to anesthetize and remove symbionts. Then, fragments were examined with the naked eye, and all found symbionts were removed. After processing, the fragments were planted on five rectangular frames 1.5 x 1.0 m in size to form settlements of different densities (20, 30, 40, 50, and 60 fragments per frame, 200 fragments in total). The distance between individual fragments was 20, 13.3, 10, 8, and 6.7 cm, respectively.
During the experiment, individual photo registration of coral colonies and their cleaning from fouling and debris were carried out twice a month. Bleaching was assessed based on the images of colonies according to the 6-point Siebeck's color reference card [44]. Often the bleaching of Pocillopora verrucosa is characterized by spotting: colonies are darkest in the central part and lightest at the ends of the branches. Therefore, the arithmetic mean of the scores of the lightest and darkest colony parts was used to estimate the level of bleaching of each individual colony. Since the impact of bleaching on symbionts develops gradually [31], the accumulated bleaching score for the last 2 censuses of colonies before sampling (07/03/2020 and 07/21/2020) was used for statistical analysis. After exposure for three months in the sea (Figure 1C), the colonies were removed from the frames, measured, photographed, and washed in clove oil emulsion (Figure 1D). Then they were carefully examined, and the remaining symbionts were collected with tweezers. Washouts from the colonies were sieved through a 1 mm diameter mesh. All symbionts found in the washouts and on the colony were fixed in 70% ethanol.
2.3. Identification and categorization of the species
Each animal was identified to the lowest possible taxonomic level in the laboratory based on specialized literature [40,41,45,46,47]. All specimens associated with the colonies of P. verrucosa were assigned to one of two main groups, “obligatory symbionts” and “facultative symbionts,” according to the existing literature [1,23,27,37]. The latter group includes both species known as facultative symbionts and poorly studied species with an uncertain status. Juveniles of Trapezia spp. crabs, which cannot be identified to the species level, were used only for comparison of the abundance of main groups of symbionts. The identified material is deposited in the A.N. Severtsov Institute of Ecology and Evolution Russian Academy of Sciences (IPEE RAS), Moscow, Russia.
The biodiversity (the total number of symbionts and the number obligate and facultative species apart), species richness (median number of species per one colony of infested host), abundance (median number of individuals per one colony of infested hosts) were used in the analysis of the environmental impacts on the structure of symbiotic community. The terms were used with minor changes following Bush et al. [48].
2.4. Statistical analysis.
Generalized linear models, GLZ, were used to assess the impact of host colony volume, colony density, and bleaching on species richness and abundance of symbionts [49]. The value of the fit criterion χ2 was used to test the distributions of dependent variables, the results of the Kolmogorov-Smirnov test were applied to supplement the estimation. The test showed that the values of all dependent variables were distributed following the Poisson distribution pattern. The Akaike information criterion (AIC) was applied to determine the best link function for the model. The models based on the identity link function were characterized by the lowest values of AIC. Therefore, they were selected to test the impacts of given factors on the symbiotic community.
The impact of each factor was assessed using the Wald test (W). The factor’s influence was considered significant if the test results were statistically significant (p<0.5).
In order to describe the effect of host colony density on community characteristics, linear trends were plotted based on average values for each density. All tests were performed in Statistica v. 12.0.
3. Results
3.1. Taxonomic composition
A total of 69 animal species were found on 200 coral colonies, of which 26 were identified to the species level, 10 at the genus, and 33 at the family or higher taxonomic level. The richest taxonomic group was crustaceans (subphylum Crustacea) which included 41 species, of which the majority, 33 species, belong to the order Decapoda. They are followed in descending order by polychaetes – 13 species, mollusks – 9 species, and brittle stars – 2 species. Fish, sipunculans, nemerteans, and turbellarians were represented by one species each (Table S1).
Of the species found, 21 were obligate symbionts, and 48 were facultative. Among obligates, the most diverse taxa were crabs of the genera Trapezia (9 species) and Cymo (6 species). In addition, three species of shrimp and one species of molluscs and fish were recorded (Table S1).
The total number of symbiont species and the number of facultative symbionts increase with the sample size and approach the asymptote when the sample size is about 160 colonies. For obligate symbionts, saturation occurs much earlier, with a sample size of about 40 colonies (Figure 3).
3.2. The impact of colony size, bleaching and host coral density on symbiotic communities.
The generalized linear models used to assess the influence of environmental factors on the composition and structure of the symbiotic community demonstrated that the volume of the host colony significantly affects both the species richness of all symbionts (W (1) = 9.64, p < 0.002) and the species richness of obligate (W (1) = 7.88, p < 0.005) and facultative (W (1) = 3.89, p < 0.048) symbionts separately. The volume of the host colony also has a significant effect on the abundance of all symbionts (W (1) = 30.77 and p < 0.0001), on the abundance of only obligate (W (1) = 21.02, p < 0.0001), and facultative (W (1) = 13.34, p < 0.0001) symbionts. In general, an increase in the volume of the host colony led to an increase in both the number of symbiont species and the number of individuals.
During the observation period, we recorded partial bleaching, in which the state of the colonies varied from 2 to 4 points on Siebeck’s scale from colony to colony and during the period of observation. According to the test results, bleaching did not significantly affect neither the richness (W (2) = 0.17, p = 0.920) nor the abundance of the symbiotic community as a whole (W (2) = 0.37, p = 0.830). Similar results were obtained in analysis of the effect of bleaching on the species richness and abundance of obligate (W (2) = 0.91, p = 0.635 and W (2) = 0.69, p = 0.710, respectively) and of facultative symbionts (W (2) = 0.04, p = 0.982 and (W (2) = 1.62, p = 0.444, respectively) alone.
Population density of host coral had a significant effect both on the species richness of all symbionts (W (4) = 11.65, p < 0.020) and on obligate (W (4) = 11.52, p = 0.021) and facultative ones (W (4) = 19.20, p < 0.0001) (Figure 4). Similarly, the density affected the abundance of all (W (4) = 18.40, p < 0.001), obligate (W (4) = 22.23, p < 0.0001), and facultative symbionts (W (4) = 45.35, p < 0.0001) (Figure 5).
However, the host density had different impacts on obligate and facultative symbionts (Figure 4 and Figure 5). Impact was positive and pronounced on species richness and abundance of obligate symbionts (SR = 0.89 + 0.24D and A = 0.92 + 0.38D, respectively, where SR is species richness, A – the abundance of symbionts, and CD – the density of colonies), whereas impact on these indexes of facultative symbionts was slightly negative (SR = 2.97 – 0.02CD and A = 4.40 – 0.15CD). Due to the opposite effect of the colony density on these two ecological groups, the impact of this factor on the community as a whole was weakened (SR = 3.99 + 0.18CD and A = 5.54 + 0.18CD).
4. Discussion
For the first time, we have obtained representative data on the composition of symbiotic communities of the coral Pocillopora verrucosa at the early stage of colony formation. Due to the large samples, we can conclude that the fauna of symbionts is represented in our samples to a very high extent. Facultative symbionts are often opportunistic species that do not have close relationships with their host coral and their diversity is driven mainly by external environmental factors (e.g., diversity of surrounding habitats) [18]. It is clear that the fauna of these facultative symbionts continues to increase slowly with an increase in the number of samples. This applies to a lesser extent to obligate symbionts and a given coral size group. According to our result, a sample of 40 colonies can be considered sufficient to assess their species diversity. However, the larger size of the sample we used in the present research allowed assessing the local diversity of obligate species more accurately. As a result, the local species diversity of obligate symbionts recorded in the framework of the present study (21 species) is noticeably higher than for larger colonies in other regions of the Indo-Pacific [18], including the richest local symbiotic communities of the Great Barrier Reef (15 species) [16,37,38,43] even though the present study was performed on small colonies inhabited by communities at an early stage of formation. These data are consistent with the exceptionally high diversity of crinoid symbionts reported earlier in Nha Trang Bay [50], which is probably related to the proximity of this region to the center of marine biodiversity, the Coral Triangle [51]. The present result on coral symbiont biodiversity is, in fact, one of the first reports on symbiont biodiversity in coral farms. Therefore, we can assume that another important function of coral farms (in addition to coral propagation) is the maintaining and conservation coral-reliant invertebrates.
Our study revealed that the structure of symbiotic communities of the coral P. verrucosa is significantly affected by two of the three studied parameters: the size of colonies and their density. At the same time, no significant effect of bleaching on species richness and abundance of communities was found.
Previously, it was shown that the deterioration of colonies’ fitness during bleaching leads to a change in intra- and interspecies relationships in the community of obligate symbionts [34]. The most pronounced consequence of bleaching is the displacement of female Trapezia cymodoce crab and shrimp Alpheus lottini from corals by male crab and a sharp decrease in clutch size in the remaining female crabs [31,32,34]. Obligate symbionts were absent on completely bleached colonies [52]. These data imply that bleaching should lead to a decrease in the species richness and abundance of obligate symbionts. However, our findings do not support this hypothesis. To what can it be related?
The observed discrepancy in the assessment of bleaching consequences for symbiotic communities is probably related to the different levels of bleaching recorded in our work and the studies of other authors. Specifically, several authors [31,32,52] considered colonies as in the “bleaching” state when they were completely bleached (white). This state probably corresponds to 1 point of Siebeck’s color reference card [44] or to “complete bleaching” in the work of Stella et al. [34]. At the same time, we did not observe colonies with a bleaching level below 2 points, which is in good agreement with the temperature regime in the area during the experiment (Figure S1). In contrast to 2019, when the temperature anomaly reached Bleaching Alert Level 2 in Nha Trang Bay [42], the temperature in the hottest months (May–July) of 2020 did not exceed the coral bleaching threshold. The partial bleaching observed by us (2 - 2.5 points) was probably caused by a slight excess of the threshold of 30 ° C in June and the beginning of July. Therefore, we can conclude that the level and duration of bleaching observed in a framework of this study are insufficient to induce changes in the structure of symbiotic communities.
Generally, there is a consensus that coral colony size is positively associated with symbiont abundance and diversity [18,19,37,43]. However, this parameter is not always a good predictor of symbiont abundance. Other factors, such as exposure at low tides, differences in host colony structure, intraspecific interactions, and spatial distribution, also can affect the symbiotic community [18,21,23,24,28,53]. In particular, our study revealed a significant effect of the population density of P. verrucosa colonies on symbionts. This result is consistent with the often observed positive correlation between the host population density and the abundance in communities of parasites [54]. It may be partly explained by the general epidemiological density-dependence theory of infection transmission [55].
We suggest that the positive effect of host density on symbiotic communities is related to the recruitment patterns of symbiont populations. It is known that the recruitment of obligate symbionts occurs in two ways: by the settlement of larvae from plankton and by the immigration of symbionts from other colonies. Coral colony aggregations may be more attractive for settling larvae than single colonies due to the effect of high concentrations of metabolites [56]. Migrations of juvenile and adult invertebrate symbionts from host to host are a common phenomenon in marine communities, which has been experimentally confirmed [57,58,59], including for coral symbionts [53,60]. The reasons for migration may be the search for a partner, lack of space and food in the colony, and aggressive intra- and interspecies interactions [53]. However, obligate symbionts associated with the host throughout their life cycle are poorly adapted to the environment and are vulnerable to predators during migration. Therefore, the mortality of symbionts migrating from host to host in dense aggregations of hosts should be lower [53], which may lead to increased diversity and abundance of symbionts in host aggregations.
Previously, we suggested that the effect of host density would be more pronounced on obligate than on facultative symbionts [23]. The data obtained confirm this suggestion (Figure 4 and Figure 5). This difference between the two ecological groups of symbionts is probably the result of a lack of functional relationships between opportunistic facultative symbionts and their hosts. Therefore, the attraction of settling larvae by host metabolites and host-to-host migrations is unlikely or impossible for them. Moreover, although the differences were not statistically significant, we found a negative relationship between the density of the host population and species richness and the abundance of facultative symbionts (Figure 4 and Figure 5). We believe that this could be the result of the competitive pressure of obligate symbionts on facultative ones, which increased with an increase in the species richness and abundance of obligates. Earlier, a similar conclusion was drawn by Patton [16], observing an increase in the abundance of facultative symbionts on Acropora colonies from which the obligate symbionts were removed. However, this hypothesis requires further experimental verification.
5. Conclusions
Our studies have shown that even at the early stage of the formation of Pocillopora verrucosa colonies, they are already inhabited by obligate and facultative symbionts. The diversity of obligate symbionts appeared to be the highest for all the studied local symbiont faunas of the World Ocean. We have demonstrated that contrary to expectations, the effect of bleaching on symbiotic communities was not recognizable. This is probably due to the absence of colonies in the state of “complete bleaching” (1 point) in our samples, since the observed bleaching did not exceed 2 – 2.5 points on the bleaching color scale [44]. However, it is possible that the observed partial bleaching may affect symbionts at the population level.
It was shown that, in general, the coral population density positively affects species richness and the abundance of symbiotic communities. However, this had the opposite effect on obligate and facultative symbionts. With an increase in density, in obligate symbionts these indexes increased, while in facultative symbionts, they slightly decreased. We believe that the positive effect of the host density on symbiotic communities is related to the peculiar properties of obligate symbionts recruitment: (1) attracting settling larvae due to the effect of high concentration of metabolites in dense host aggregations and (2) reducing mortality of migrating symbionts in host aggregations. We hypothesize that a slight negative effect of the host density on species richness and the abundance of facultative symbionts is caused by the competitive pressure of obligates increasing with increase of their abundance. However, further experimental studies are necessary to verify these hypotheses.
Due to the vital role of symbionts in functioning coral colonies [4,6,7], young colonies transplanted from coral nurseries to the natural environment should contain a set of symbionts to maintain their fitness and protection from predators. Therefore, our data on the effect of density on the symbiotic community should be considered when planning the optimal fragment density in coral farms and when planting seedlings in coral reef restoration.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of symbiont taxa found on experimental colonies in association with the coral Pocillopora verrucosa; Figure S1: Temperature dynamics for 3 months from April to July 2020 at a depth of 3 m, obtained using HOBO® u22-001 thermologgers at the experimental site in Dam Bay.
Author Contributions
Conceptualization, T.A.B. and F.V.L..; methodology, T.A.B., F.V.L., Y.A.B.; software, F.V.L.; formal analysis, T.A.B., F.V.L., Y.A.B.; investigation, T.A.B., F.V.L., Y.A.B., H.T.N. and P.R.A.; resources, F.V.L., T.A.B. and H.T.N.; data curation, T.A.B.; writing—original draft, T.A.B.; writing—review and editing, T.A.B., F.V.L., Y.A.B., H.T.N. and P.R.A.; visualization, T.A.B., F.V.L., Y.A.B., and P.R.A.; supervision, T.A.B.; funding acquisition, T.A.B. and H.T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation; grant number RSF 22-24-00836. .
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data used in this study have been made available on-line, and the corresponding links are indicated in the text.
Acknowledgments
Our work in Vietnam was made possible through the logistics of the Coastal Branch of the Russian-Vietnamese Tropical Centre, and we would like to thank its directors, Mr. N.L. Filichev and Mr. N.N. Hu’ng. We are also grateful to Mr. S. Gorin for his help in planting and sampling corals, as well as for preparing an orthophotoplan of the area where the experiment was deployed and to Dr. A. Zalota for the insightful comments that helped improve this manuscript. .
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. An Annu. Rev. 2011, 49, 43–104. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Van Der Meij, S.E.T.; Fransen, C.H.J.M. The mushroom coral as a habitat. J. Mar. Biol. Assoc. United Kingdom 2012, 92, 647–663. [Google Scholar] [CrossRef]
- Enochs, I.C. Motile cryptofauna associated with live and dead coral substrates: implications for coral mortality and framework erosion. Mar. Biol. 2012, 159, 709–722. [Google Scholar] [CrossRef]
- Montano, S. The extraordinary importance of coral associated fauna. Diversity 2020, 12, 357. [Google Scholar] [CrossRef]
- Stimson, J. Stimulation of fat-body production in the polyps of the coral Pocillopora damicornis by the presence of mutualistic crabsof the genus Trapezia. Mar. Biol. 1990, 106, 211–218. [Google Scholar] [CrossRef]
- Stewart, H.L.; Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Symbiotic crabs maintain coral health by clearing sediments. Coral Reefs 2006, 25, 609–615. [Google Scholar] [CrossRef]
- Stier, A.C.; Gil, M.A.; McKeon, C.S.; Lemer, S.; Leray, M.; Mills, S.C.; Osenberg, C.W. Housekeeping mutualisms: Do more symbionts facilitate host performance? PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Pollock, F.J.; Katz, S.M.; Bourne, D.G.; Willis, B.L. Cymo melanodactylus crabs slow progression of white syndrome lesions on corals. Coral Reefs 2013, 32, 43–48. [Google Scholar] [CrossRef]
- Glynn, P.W. Some physical and biological determinants of coral community structure in the Eastern Pacific. Ecol. Monogr. 1976, 46, 431–456. [Google Scholar] [CrossRef]
- Pratchett, M.S. Influence of coral symbionts on feeding preferences of crown-of-thorns starfish Acanthaster planci in the western Pacific. Mar. Ecol. Prog. Ser. 2001, 214, 111–119. [Google Scholar] [CrossRef]
- McKeon, C.S.; Stier, A.C.; McIlroy, S.E.; Bolker, B.M. Multiple defender effects: synergistic coral defense by mutualist crustaceans. 2012, 169, 1095−1103. [CrossRef]
- Rouzé, H.; Lecellier, G.; Mills, S.C.; Planes, S.; Berteaux-Lecellier, V.; Stewart, H. Juvenile Trapezia spp. crabs can increase juvenile host coral survival by protection from predation. Mar. Ecol. Prog. Ser. 2014, 515, 151–159. [Google Scholar] [CrossRef]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. R. Soc. B. 2017, 284, 20172405. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.K.; Graham, N.A.J.; Polunin, N.V.C. Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Mar. Biol. 2007, 151, 1069–1076. [Google Scholar] [CrossRef]
- Patton, W.K. Decapod Crustacea commensal with Queensland branching corals. Crustaceana 1966, 10, 271–295. [Google Scholar] [CrossRef]
- Patton, W.K. Distribution and ecology of animals associated with branching corals (Acropora spp.) from the Great Barrier Reef, Australia. Bull. Mar. Sci. 1994, 55, 193–211. [Google Scholar]
- Stella, J.S.; Jones, G.P.; Pratchett, M.S. Variation in the structure of epifaunal invertebrate assemblages among coral hosts. Coral Reefs 2010, 29, 957–973. [Google Scholar] [CrossRef]
- Britayev, T.A.; Spiridonov, V.A.; Deart, Y.V.; El-Sherbiny, M. Biodiversity of the community associated with Pocillopora verrucosa (Scleractinia: Pocilloporidae) in the Red Sea. Mar. Biodivers. 2017, 47, 1093–1109. [Google Scholar] [CrossRef]
- Abele, L.G.; Patton, W.K. The size of coral heads and the community biology of associate decapod crustaceans. Journal of Biogeography 1976, 3, 35–47. [Google Scholar] [CrossRef]
- Lewis, J.B.; Snelgrove, P.V.R. Corallum morphology and composition of crustacean cryptofauna of the hermatypic coral Madracis mirabilis. Mar. Biol. 1990, 106, 267–272. [Google Scholar] [CrossRef]
- Pisapia, C.; Stella, J.; Silbiger, N.J.; Carpenter, R. Epifaunal invertebrate assemblages associated with branching pocilloporids in Moorea, French Polynesia. PeerJ 2020, 8, e9364. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Abele, L.G. Community patterns of coral-associated decapods. Mar. Ecol. Prog. Ser. 1983, 13, 131–139. [Google Scholar] [CrossRef]
- Britayev, T.A.; Mikheev, V.N. Clumped spatial distribution of scleractinian corals influences the structure of their symbiotic associations. Dokl. Biol. Sci. 2013, 448, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Counsell, C.W.; Donahue, M.J.; Edwards, K.F.; Franklin., E.C.; Hixon. Variation in coral-associated cryptofaunal communities across spatial scales and environmental gradients. Coral Reefs 2018, 37, 827–840. [Google Scholar] [CrossRef]
- Munday, P.L. Interactions between habitat use and patterns of abundance in coral-dwelling fishes of the genus Gobiodon. Environ. Biol. Fishes 2000 58, 355–369. [CrossRef]
- Depczynski, M.; Bellwood, D.R. Wave energy and spatial variability in community structure of small cryptic coral reef fishes. Mar. Ecol. Prog. Ser. 2005, 303, 283–293. [Google Scholar] [CrossRef]
- Black, R.; Prince, J. Fauna associated with the coral Pocillopora damicornis at the Southern Limit of its distribution in Western Australia. Journal of Biogeography 1983, 10, 135–152. [Google Scholar] [CrossRef]
- Edwards, A. ; Emberton. H. Crustacea associated with the scleractinian coral, Stylophora pistillata (Esper.) in the Sudanese Red Sea. J. Exp. Mar. Biol. Ecol. 1980, 42, 225–240. [Google Scholar] [CrossRef]
- Brown, B.E. Coral bleaching: causes and consequences. Coral Reefs 1997, 16 (Suppl. S1), S129–S138. [Google Scholar] [CrossRef]
- Douglas, A.E. Coral bleaching—how and why? Mar. Pollut. Bull. 2003, 46, 385–392. [Google Scholar] [CrossRef]
- Stella, J.S.; Munday, P.L.; Jones, G.P. Effects of coral bleaching on the obligate coral-dwelling crab Trapezia cymodoce. Coral Reefs 2011, 30, 719–727. [Google Scholar] [CrossRef]
- Tsuchiya, M. Effect of mass coral bleaching on the community structure of small animals associated with the hermatypic coral Pocillopora damicornis. Galaxea, JCRS 1999, 1, 65–72. [Google Scholar] [CrossRef]
- Munday, P.L. Habitat loss, resource specialization, and extinction on coral reefs. Global Change Biol. 2004, 10, 1642–1647. [Google Scholar] [CrossRef]
- Stella, J.S.; Munday, P.L.; Walker, S.P.W.; Pratchett, M.S.; Jones, G.P. From cooperation to combat: adverse effect of thermal stress in a symbiotic coral-crustacean community. Oecologia 2014, 174, 1187–1195. [Google Scholar] [CrossRef]
- Froehlich, C.Y. M.; Klanten, O.S.; Hing, M.L.; Dowton, M.; Wong, M.Y.L. Uneven declines between corals and cryptobenthic fish symbionts from multiple disturbances. Sci. Rep. 2021, 11, 16420. [Google Scholar] [CrossRef] [PubMed]
- Combillet, L.; Fabregat-Malé, S.; Mena, S.; Marín-Moraga, J.A.; Gutierrez, M.; Alvarado, J.J. Pocillopora spp. growth analysis on restoration structures in an Eastern Tropical Pacific upwelling area. PeerJ 2022, 10, e13248. [Google Scholar] [CrossRef] [PubMed]
- Coles, S. Species diversity of decapods associated with living and dead reef coral Pocillopora meandrina. Mar. Ecol. Prog. Ser. 1980, 2, 281–291. [Google Scholar] [CrossRef]
- Patton, W.K. Community structure among the animals inhabiting the coral Pocillopora damiconis at Heron Island, Australia. In Symbiosis in the sea; Vernberg, W.B., Ed.; University of South Carolina Press: Columbia, USA, 1974; pp. 219–243. [Google Scholar]
- Tkachenko, K.S.; Soong, K.; Deart, Y.V.; Britayev, T.A. Coral symbiotic communities from different environments of an isolated atoll: reef lagoon versus forereef. Invert. Zool. 2022, 19, 78–90. [Google Scholar] [CrossRef]
- Marin, I.N.; Britayev, T.A.; Anker, A. Pontoniine shrimps associated with cnidarians: new records and list of species from coastal waters of Viet Nam. Arthropoda Selecta 2004, 13, 199–218. [Google Scholar]
- Marin, I.N.; Spiridonov, V.A. Coral-associated crabs (Decapoda: Domecidae, Trapeziidae, Tetraliidae, Xanthidae: Cymoinae) from Nha Trang Bay. In Benthic Fauna of the Bay of Nha Trang; Britayev, T.A., Pavlov, D.S., Eds.; KMK Scientific Press: Moscow, Russia, 2007; pp. 209–234. [Google Scholar]
- Tkachenko, K.S.; Britayev, T.A.; Huan, N.; Pereladov, M.V.; Latypov, Y. Influence of anthropogenic pressure and seasonal upwelling on coral reefs in Nha Trang Bay (Central Vietnam). Mar. Ecol 2016, 37, 1131–1146. [Google Scholar] [CrossRef]
- Austin, A.D.; Austin, S.A.; Sale, P.E. Community structure of the fauna associated with the coral Pocillopora damicornis (L.) on the Great Barrier Reef. Mar. Freshwater Res. 1980, 31, 163–174. [CrossRef]
- Siebeck, U.E.; Marshall, N.J.; Klüter, A.; Hoegh-Guldberg, O. Monitoring coral bleaching using a colour reference card. Coral Reefs 2006, 25, 453–460. [Google Scholar] [CrossRef]
- Bruce, A.J. New keys for the identification of Indo-West Pacific coral associated pontoniine shrimps, with observations on their ecology: (Crustacea: Decapoda: Palaemonidae). Ophelia, 1998, 49, 29–46. [Google Scholar] [CrossRef]
- Castro, P.; Ng, P.K.L.; Ahyong, S.T. Phylogeny and systematics of the Trapeziidae Miers, 1886 (Crustacea: Brachyura), with the description of a new family. Zootaxa 2004, 643, 1–70. [Google Scholar] [CrossRef]
- Banner, D.M.; Banner, A.B. The alpheid shrimp of Australia Part III: The remaining alpheids, principally the genus Alpheus, and the family Ogyrididae. Rec. Aust. Mus. 1982, 34, 1–357. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- O'Hara, R.; Kotze, J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Britayev, T.A.; Mekhova, E.S. Assessment of hidden diversity of crinoids and their symbionts in the bay of Nhatrang, Vietnam. Org. Divers. Evol. 2011, 11, 275–285. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Delineation of the Indo-Malayan Centre of maximum marine biodiversity: the Coral Triangle. In Biogeography, time and place: distributions, barriers and islands; Renema, W., Ed.; Springer: Dordrecht, 2007; pp. 117–178. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Yamauchi, Y.; Moretzsohn, F.; Tsukiji, M. Species composition and some population traits of obligate symbiotic xanthid crabs, Trapezia and Tetralia, associated with bleached corals. In Proc 7th Intl Coral Reef Symp. 1992, 1, 56–63. [Google Scholar]
- Castro, P. Movements between corals colonies in Trapezia ferruginea (Crustacea: Brachyura), an obligate symbiont of scleractinian corals. Mar. Biol. 1978, 46, 237–245. [Google Scholar] [CrossRef]
- Arneberg, P.; Skorping, A.; Grenfell, B.; Read, A.F. Host densities as determinants of abundance in parasite communities. Proc. R. Soc. Lond B 1998, 265, 1283–1289. [Google Scholar] [CrossRef]
- Anderson, M.R.; May, R.M. Infectious diseases of humans: dynamics and control. Oxford University Press, New York, 1991.
- Marsden, J.R. Coral preference behaviour by planktotrophic larvae of Spirobranchus giganteus corniculatus (Serpulidae: Polychaeta). Coral Reefs 1987, 6, 71–74. [Google Scholar] [CrossRef]
- Thiel, M.; Zander, A.; Baeza, J.A. Movements of the symbiotic crab Liopetrolisthes mitra between its host sea urchin Tetrapygus niger. Bull. Mar. Sci. 2003, 72, 89–101. [Google Scholar]
- Dgebuadze, P. Yu.; Mehova, E.S.; Britayev, T.A. Recolonization of the Himerometra robustipinna (Himerometridae, Crinoidea) by macrosymbionts: an in situ experiment. Symbiosis 2012, 58, 253–258. [Google Scholar] [CrossRef]
- Britayev, T.A.; Mekhova, E.S. Do symbiotic polychaetes migrate from host to host? Memoirs of Museum Victoria 2014, 71, 21–25. [Google Scholar] [CrossRef]
- Preston, E.M. A computer simulation of competition among five sympatric congeneric species of xanthid crabs. Ecology 1973, 54, 469–483. [Google Scholar] [CrossRef]
Figure 1.
A. Pocillopora verrucosa colony in its natural habitat. B. Rectangular experimental frame with planted coral colonies installed on a sandy bottom. C. One of the experimental colonies, close view. D. Washout of one colony after three months of exposure: crab Trapezia tigrina, shrimps Alpheus lottini and Palaemonella rotumana are recognizable.
Figure 1.
A. Pocillopora verrucosa colony in its natural habitat. B. Rectangular experimental frame with planted coral colonies installed on a sandy bottom. C. One of the experimental colonies, close view. D. Washout of one colony after three months of exposure: crab Trapezia tigrina, shrimps Alpheus lottini and Palaemonella rotumana are recognizable.

Figure 2.
Location of the study area at the coast of Vietnam (A and B) and sampling site in Dam Bay (C, orthophotoplan taken from a height of 20 m). The area where experimental constructions with planted colonies of Pocillopora verrucosa are located is circled in red.
Figure 2.
Location of the study area at the coast of Vietnam (A and B) and sampling site in Dam Bay (C, orthophotoplan taken from a height of 20 m). The area where experimental constructions with planted colonies of Pocillopora verrucosa are located is circled in red.

Figure 3.
Accumulation of symbiont species as a function of sample size, both for the total species pool and for obligatory and facultative species associated with Pocillopora verrucosa (n=200 colonies.
Figure 3.
Accumulation of symbiont species as a function of sample size, both for the total species pool and for obligatory and facultative species associated with Pocillopora verrucosa (n=200 colonies.
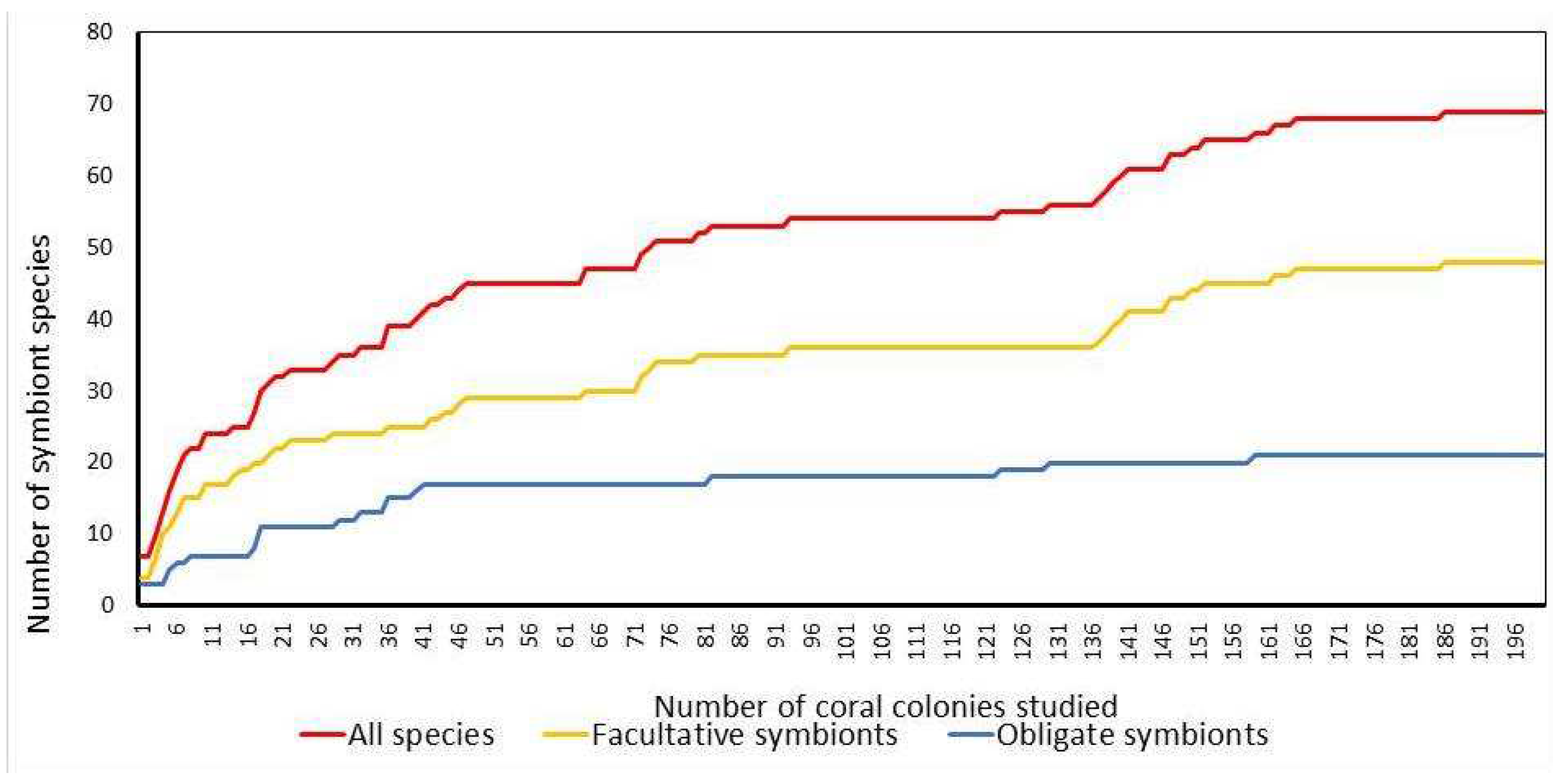
Figure 4.
Effect of colony density on species richness of symbionts (mean number of species per colony).
Figure 4.
Effect of colony density on species richness of symbionts (mean number of species per colony).
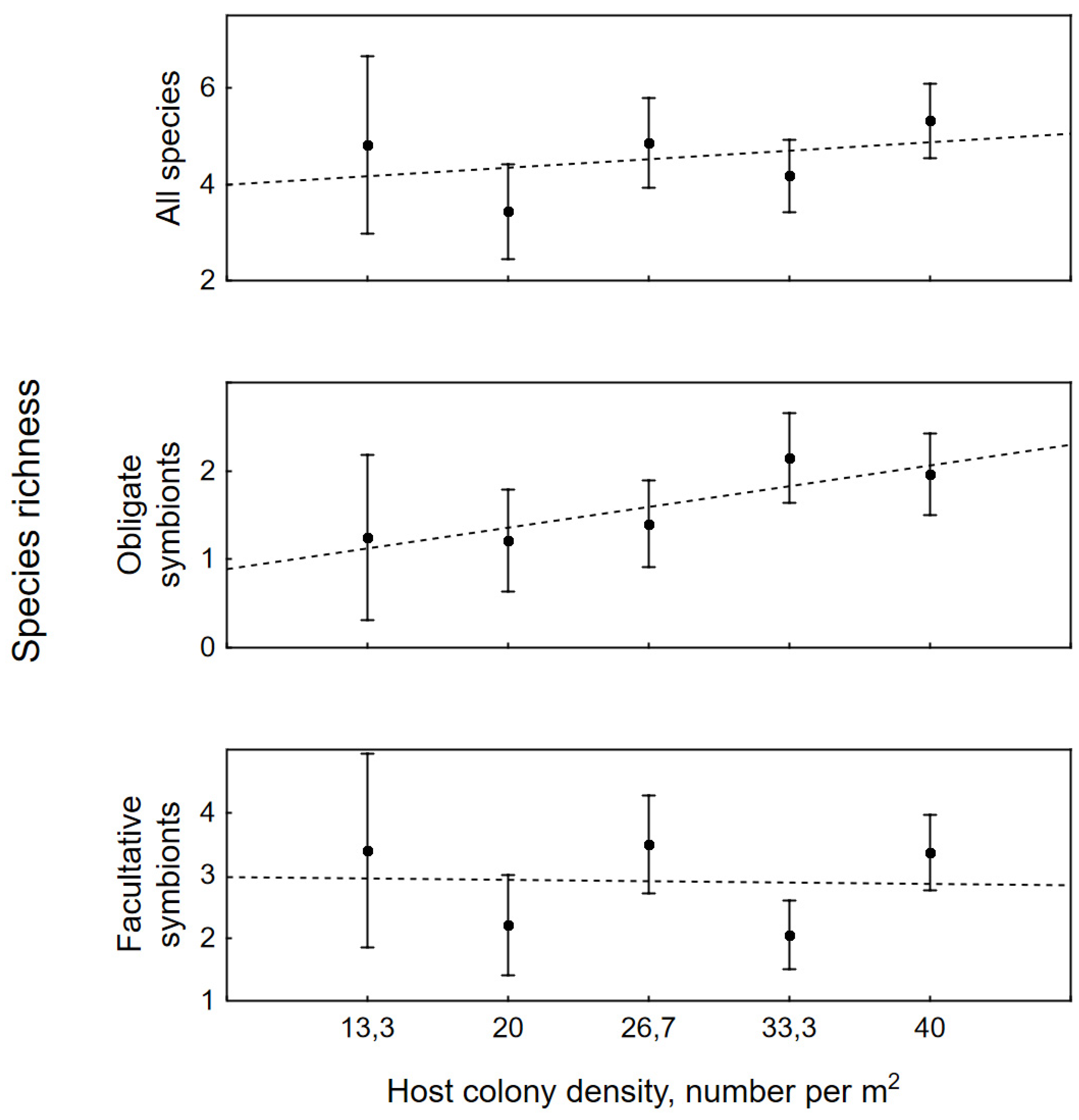
Figure 5.
Effect of colony density on abundance of symbionts (mean number of individuals per colony).
Figure 5.
Effect of colony density on abundance of symbionts (mean number of individuals per colony).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



