Submitted:
14 March 2023
Posted:
15 March 2023
You are already at the latest version
Abstract
The emergence of drug resistant pathogenic bacteria is increasingly challenging conventional antibiotics. Plant derived flavonoids are considered as potential alternatives to antibiotics due to their antimicrobial properties. However, the mechanisms by which flavonoids modulate pathogenic microorganisms’ growth are not fully understood. In our previous studies we found that Rutin, a kind of flavonoids showed inhibition against Klebsiella pneumoniae strains. In order to better understand its inhibitory mechanism on strain growth, we used both Rutin and Luteolin as treatments to incubate K. pneumoniae ATCC700603 strain. After incubation for 4 hours, the homogenous and differential effects of two different flavonoids on the growth of K. pneumoniae were evaluated. The integrated metabolomic and transcriptomic analysis was performed. The expression levels of 5,483 genes and the contents of 882 metabolites were measured. The differentially expressed genes (DEGs) and metabolites (DEMs) were screened and analyzed. The correlation between DEGs and DEMs were also studied. Our results showed that Rutin could inhibit the strain growth by changing metabolic pathways and ABC transporters pathways. Our study also revealed FU841_RS17580 and FU841_RS19145 as functional genes that played vital roles in the strain growth.
Keywords:
drug resistance
; flavonoids
; RNA-seq
; Klebsiella pneumoniae
; Rutin
; antibacterial agents
1. Introduction
The infections caused by pathogenic bacteria are threatening human beings’ health worldwide [1]. These bacteria are very likely to develop drug resistance during the treatment process with the use of antibiotics [2]. Once become drug resistant ones, the microorganisms are often difficult to eradicate by conventional antibiotics [3]. In addition, the drug resistant bacteria have evolved to spread drug resistance within populations via horizontal gene transfer, as well as gene mutations, which lead to persistent infections [4,5]. As a result, new strategies and approaches in finding effective antibacterial agents that are not easily to generate drug resistance are becoming more and more urgent, especially when high frequency of antibiotics resistance occurs [6].
Plant derived flavonoids are bioactive compounds in traditional herbal medicines and have been considered as potential antibacterial agents due to their antimicrobial properties [7]. With a C6-C3-C6 skelecton, flavonoids represent a large group of plant secondary metabolites and are widely distributed in plant kingdom [8,9]. Flavonoids are consist of more than 8,000 compounds and provide a huge resource pool for novel antibacterial drug discovery [10]. Most importantly, with low toxicity and no side effects, flavonoids, especially dietary flavonoids are much safer and not easily to develop drug resistance [11].
The antibacterial effects of flavonoids have been extensively investigated and studied for many years [12,13]. Flavonoids extracts derived from Quercus brantii L. fruit exhibited significant inhibitory effect on both Staphylococcus aureus (MIC 500µg/mL) and Enterococcus fecalis (MIC 600 µg/mL) [14]. The extract from a shrub (Sambucus australis) also showed antibacterial activity against Salmonella typhimurium (MIC 250µg/mL) and Klebsiella pneumoniae (MIC 250µg/mL) [15]. Regarding to a single compound, naringenin showed inhibitory effect on clinical strains of clarithromycin-resistant Helicobacter pylori, vancomycin-resistant E. faecalis, methicillin-resistant S. aureus, and betalactam-resistant Acinetobacter baumannii and K. pneumoniae [16]. In addition, some other reports also demonstrated that flavonoids, such as naringenin [17] and apigenin [18] showed enhanced antimicrobial effect when used in combination with various conventional antibiotics. As a result, the strategy of using flavonoids with antibiotics in clinical, would enhance antimicrobial therapy, reduce medicine dosage and slow down the development of drug resistance.
The antibacterial mechanism of flavonoids on pathogenic microorganisms varied from bacteria to bacteria, even though they were different isolates affiliated to a same genus [19]. Briefly, the flavonoids can inhibit the cell envelop synthesis [20], bacterial motility [21], nucleic acid synthesis [22], biofilm formation [23] and quorum sensing [24] in various pathogenic bacteria. In addition, some flavonoid showed up to sixfold stronger antibacterial activities than conventional antibiotics [13]. Nevertheless, these plant derived components remain a poorly characterized reservoir of anti-infective agents [25]. Although the antioxidant and antibacterial properties of flavonoids, either natural or synthetic, have been studied extensively, the antibacterial mechanism has not been fully understood.
To understand the susceptibility of pathogenic bacteria in response to flavonoids, it is crucial to reveal the interaction between gene expression and metabolites, especially the changes before and after flavonoids treatments. However, most previous antibacterial studies performed through the paper disk method [26] and/or minimum inhibition concentration (MIC) evaluation [27]. The mechanism of flavonoids showed inhibitory effects was not fully understood. Recently, in our previous studies, we also evaluated the antioxidant and antibacterial effects of 10 flavonoids. Nevertheless, consistent with some previous studies [28,29], we found most flavonoids were ineffective in inhibiting the growth of Klebsiella pneumoniae except Rutin [30]. Therefore, in present work, a flask of a liquid culture of Rutin and K. pneumoniae bacterium were pre-incubated, effectively avoiding influence of environmental factors. Another type of flavonoids, Luteolin was also used as control. The RNA-seq and metabolome sequencing were conducted, respectively. The relationship between differentially expressed metabolites (DEMs) and differentially expressed genes (DEGs) in K. pneumoniae after pre-incubation with flavonoids treatments was assessed, revealing the molecular mechanism determining the inhibitory effects of Rutin against K. pneumoniae strain.
2. Materials and Methods
2.1. Bacteria Growth Conditions and Treatments
The standard bacterial strain K. pneumoniae ATCC700603 was purchased from China National Health Inspection Center and used as a model to evaluate the inhibition of Rutin. This strain was cultured in Luria-Bertani broth (LB, Haibo, Qingdao, China) at 37℃ with shaking or as static cultures, as required.
Flavonoids including Rutin and Luteolin were purchased from Aladdin Chemistry Co., Ltd. (Shanghai, China). The flavonoids stocks were prepared in mixture of methanol and ethanol (V:V=1:1) and the concentration was made to 1024 µg/mL. All chemicals used in this study were of chromatography-grade. Ultra pure water was used throughout all experiments.
Briefly, 10mL bacterial suspension (10^6 cfu/mL) was transferred to a flask of 89 mL LB liquid culture. Then, 1mL of each dissolved flavonoid stock was added to each flask, respectively, making the final volume to 100 mL in each flask. All experiments were performed in triplicate. The empty control was conducted in 3 flasks filled with 90 mL LB and 10 mL bacterial cells (10^6 cfu/mL). The flasks were covered and incubated for 4h at 37℃with shaking (150 rpm). At last, two specimens randomly selected from the flavonoids treatment groups and two specimens from the control group were used for transcriptomic analysis.
2.2. RNA Extraction, Library Preparation and Illumina Hiseq Sequencing
The bacterium cells were collected from 10 mL culture medium after centrifugation at 1000 g for 10 min at 4℃. Total RNA was isolated from bacterial cells using TRIzol® Reagent (Invitrogen, Carlsbad, CA, USA), according to the method[31]. Genomic DNA was removed using DNase I (TaKaRa, Japan). Then RNA quality was determined using 2100 Bioanalyser (Agilent, CA, USA) and quantified using the NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies, Delaware, USA). High-quality RNA sample (OD260/280=1.8~2.2, OD260/230≥2.0, RIN≥6.5, 23S/16S≥1.0) was used to construct sequencing library.
A total amount of 5 μg RNA per sample was extracted from the strain cells and used as input material for library construction. First, the mRNA was interrupted by ion using TruseqTM Stranded RNA sample prep Kit (Illumina, San Diego, USA). Then the dUTP was used instead of dTTP in the second chain synthesis for double stranded cDNA synthesis, and the index adaptor was added after double strained cDNA synthesis using TruseqTM Stranded RNA sample prep Kit. After that, the second chain of cDNA was degraded using UNG (Uracil-N-Glycosylase) enzyme (Illumina, San Diego, USA). The library was enriched by PCR (polymerase chain reaction) using cBot Truseq PE Cluster Kit v3-cBot-HS (Illumina, San Diego, USA). Second, the library was separated and purified using 2% certified low range ultra agarose gel electrophoresis (Bio-Rad, Herales, USA). The target gene fragments were isolated by gel extraction kit. Then, the library was amplified using a cBot Truseq PE Cluster Kit v3-cBot-HS (Illumina, San Diego, USA). At last, the library preparations were sequenced on Illumina platform (Beijing iGeneTech Biotechnology Co., Ltd., Beijing, China) using the Truseq SBS Kit (300cycles) (Illumina, San Diego, USA), following their manufacturer’s recommendations, and index codes were added to attribute sequences in each sample.
The raw paired end reads were trimmed and quality controlled by Trimmomatic with parameters (SLIDINGWINDOW:4, z15 MINLEN:75) (version 0.36) (http://www.usadellab.org/cms/uploads/supplementary/Trimmomatic). Then clean reads were separately aligned to reference genome with orientation mode using Rockhopper (http://cs.wellesley.edu/btjaden/Rockhopper/) software. Rockhopper was a comprehensive and user-friendly system for computational analysis of bacterial RNA-seq data. As input, Rockhopper takes RNA sequencing reads generated by high-throughput sequencing technology. This software was also used to calculate gene expression levels with default parameters.
RNA-seq and bioinformatics analysis were executed by Beijing iGeneTech Biotechnology Cp. Ltd. (Beijing, China). The sequencing libraries of samples were constructed by a TruSeq RNA Sample Preparation Kit (Illumina, San Diego, CA, USA), and the libraries were sequenced on the Illumina Hiseq X platform. The reference genome was extracted from Genome Database for Klebsiella pneumoniae (BioProject: PRJNA559783), and gene annotations were also acquired from the genome. The index of the reference genome was built with Bowtie 2 (2.2.6), and the clean reads were mapped to the reference genome using Tophat (2.0.14). HTSeq (0.9.1) was applied to the expression and quantification levels of genes to calculate fragments per kilobase per million fragments (FPKM). Then, the DEGs were identified by DESeq (1.30.0) with screening conditions as follows: expression difference multiple |log2 Fold Change|>1 and significant P-value < 0.05. Subsequently, clustering analysis of all differential genes was performed using the R language Pheatmap (1.0.8) software package. Each point had three biological replicates for RNA-seq analysis.
To identify DEGs (differentially expressed genes) between two compared groups, the expression level for each transcript was calculated using the RPKM method. The edgeR (https://bioconductor.org/packages/release/bioc/html/edgeR.html) was used for differential expression analysis. The DEGs between two samples were selected using the following criteria: the logarithmic of fold change was greater than 2 and the false discovery rate (FDR) should be less than 0.05. To understand the functions of the differentially expressed genes, GO functional enrichment and KEGG pathway analysis were carried out by Goatools (https://github.com/tanghaibao/Goatools) and KOBAS (http://kobas.cbi.pku.edu.cn/home.do) respectively. DEGs were significantly enriched in GO terms and metabolic pathways when their Bonferroni-corrected P-value was less than 0.05.
2.3. Metabolites Measurement and Analysis
Another 10 mL pre-incubated culture medium was centrifuged for collection of bacteria cells. The metabolites identification and quantification were also conducted by Beijing iGeneTech Biotechnology Company. Briefly, the freeze-dried samples were mashed with liquid nitrogen, and 50 mg of powder for each sample was transferred into a 1.5 mL EP tube containing 5 mL precooled methanol/water (3:1, v/v) and subjected to vigorous vortexing. Each sample point had 3 independent biological replicates. All samples were extracted at 4˚C overnight. Then, the samples were centrifuged at 13,000 rpm for 15 min at 4˚C and the filtered supernatant (0.22 μm membrane filter) was blow-dried with nitrogen. The samples were redissolved with 50μL isopropanol/acetonitrile/water (2:7:1, v/v/v) and extracted by centrifugation before LC-MS/MS analysis.
The extracted samples of strains were separated on the UPLC system (Dionex UltiMate Ultimate 3000) equipped with ACQUITY UPLC HSS T3 column (100 mm*2.1 mm, 1.7 μm, Waters, USA). The analysis conditions were as follows: column temperature, 40 ℃; injection volume, 1.5 μL; flow rate, 0.35 mL/min. The mobile phases were water (0.1% formic acid) (phase A) and acetonitrile (phase B). The metabolites were eluted with the following gradient: 0~1.0 min, 5% B; 1.0~9.0 min, 5~100% B; 9.0~12.0 min, 1000% B; 12.0~15.0 min 5% B. The loading volume of each sample was 5 μL. Samples were inserted into quality control (QC) samples in queue mode to monitor and evaluate the stability of the system and the reliability of the experimental data. Each sample was operated in both positive and negative ion modes by the electrospray ionization (ESI) source parameters. Mass spectrometry was carried out by the Q-Exactive spectrometer (Thermo Scientific, San Jose, CA, USA) after the sample was separated by UPLC. The MS conditions were as follows: mass range (m/z) 80~1200; aux gas flow rate, 15 arb; sheath gas flow rate, 45 arb; source temperature, 320˚C; spray voltage, 3.5 kV in positive mode and 3.2 kV in negative mode. After the original data were adjusted for peak alignment, peak area extraction, retention time correction and feature extraction using the Compound Discoverer 3.0 program, the metabolite structure was identified by accurate mass MS matching (<25 ppm), and the MS1 and MS2 matching search MZcloud and ChemSpider database were used to determine the metabolite structure. For MZcloud database, the metabolites were identified by exact mass (m/z), molecular formula, and fragmentation spectrum (MS2). The relative content of metabolites was expressed by peak area. After preprocessed by pattern recognition and Pareto-scaling, the peak area data for all metabolites were further analyzed by SIMCA-P 14.1 software (Umetrics, Umea, Sweden), including unsupervised principal component analysis (PCA).
2.4. Gene Expression Verification
To validate the RNA-seq results, 10 genes (5 from the metabolic pathways, 5 from the ABC transporters, both pathways were shown to be significantly impacted by Rutin treatment) were randomly selected for quantitative real-time polymerase chain reaction (qRT-PCR). Primers were designed based on cDNA sequences and 16S [32] was used as reference gene. Relative gene expression of each gene was calculated by using 2-∆∆CT method [33]. The fold change in gene expression was obtained by the normalization of each gene to the 16S rRNA internal control.
The qRT-PCR was performed by the 7500 RT-PCR system (Applied Biosystems, Foster City, USA) using a SYBR Green RT-PCR Kit (Qiagen, Hilder, Germany). According to the manufacturer’s instructions, 20 µL PCR reaction mixture were composed of 2 µL of cDNA, 1.5 µl of each pair of target primers (200 nM), 5 µL of ddH2O and 10 µL of 2 × SYBR® Green Supermix (Bio-Rad Laboratories, Shanghai, China). The PCR conditions were as follows: 95˚C for 5 min (initial denaturation), followed by 40 cycles of denaturation at 95 ˚C for 15 s, annealing at 60 ˚C for 25 s, and extension at 72 ˚C for 30 s. The specificity of each PCR reaction was confirmed by melting curve analysis.
2.5. Statistical Analysis
To conduct the statistical analyses and generate data graphs, SPSS (V19.0) software and GraphPad Prism (V9.1.0) software were used. Data with a normally distributed distribution were expressed as mean ± SD. The nonnormal distribution data was shown as median (interquartile distribution). The t-test or one-way ANOVA test was used to quantify the differences between different groups. All measurements and calculations are presented as mean of 3 standard error of the mean (s.e.m.). Student’s t-test was used to compare data between each two groups and P < 0.05 was considered statistically significant. Pearson correlation analysis was performed. A P value < 0.05 was considered statistically significant correlation for all tests. Cytoscape (V3.8.2) software was used to construct the correlation network.
3. Results
3.1. Inhibition of Rutin and Luteolin on the Growth of K. pneumoniae Strain
In a previous study, we evaluated the antioxidant capacity of 10 flavonoids and found that flavonol class posessed higher free radical scavenging capacity compared to those of flavone class [30]. Also, we have uncovered that the minimum inhibitory concentrations (MICs) of Rutin against K. pneumoniae were lower than that of Luteolin as no MIC was found in Luteolin. In present study, both Rutin (from flavonol class) and Luetolin (from flavone class) were used as treatments in omics sequencing.
The inhibitory effects of Rutin against the growth of K. pneumoniae ATCC700603 were identified by both the Confocal Laser Scanning Microscopy (CLSM) and spectrophotometry methods. The inhibition was observed at 4h and 8h, respectively (Figure 1A). In the first 4h, Rutin was found to have inhibition against strain growth at the concentrations of either 256 μg/mL or 512 μg/mL. However, when the concentration was 256μg/mL, the inhibition of Rutin against the strains was not significant at 8h. Gentamicin (50 μg/mL) was used as reference antibiotic and showed obvious inhibitory effects through detection. The solvent (methanol : ethanol= 1:1) only treatment showed no obvious influence on the bacteria at two time points. The bacterial density was also evaluated by spectrophotometry. Consistent with the results of CLSM, the bacterial growth was inhibited by Rutin at 4h. When Rutin’s concentration was lowered to 256 g/mL, no obvious inhibition was found at 8h (Figure 1B).
The function of Luteolin against K. pneumoniae ATCC700603 was also evaluated. However, the inhibitory effects was not obvious, even the concentration was increased to 1024 μg/mL.
3.2 Differentially Expressed Genes and Metabolites in Response to Rutin and Luteolin Treatments
Both Rutin and Luteolin treatments were used for integrative analysis of transcriptomic and metabolomic sequencing. Well-grown liquid bacteria without any treatments was used as wild control.An average of 17,066,492 clean reads were obtained per sample from 37,497,710 raw reads for all samples (Supplementary material, Data sheet 1, Table S1). The Q20 value and Q30 value were 98.96% and 96.23%, respectively. The average mapping rate for all clean reads to the reference transcripts achieved to 65.59%. To search for key genes that are in response to flavonoids treatments, the expression levels of the transcripts were calculated utilizing the RPKM approach. We categorized these genes into high, medium and low groups according to their RPKM. In all the 6 libraries, more genes had low expression (RPKM ≤1, avg. 2,843), fewer genes had medium (RPKM 1~10, avg. 1,274) and high expression (RPKM ≥10, avg. 1,366) (Figure 2A). According to their RPKM values, genes in each library were divided into three groups, including high, medium and low. In present study, genes in a same RPKM group were also compared. These results were shown in venn diagrams (Figure 2B~E). Overall, there was no big difference in the total amount of genes induced by treatments or without treatments within a same type RPKM group. However, in addition to the genes that they shared, the numbers of those unique to treatments or without treatments varied. The most obvious difference was that, within a same type RPKM group, the amount of unique gene induced by Rutin was less than that of Luteolin treatment. It indicates that Rutin inhibited the expression of these unique genes. However, the number of unique genes measured in wild strains was more than that of any treatments, except within the high RPKM group. These results indicate that, compared to wild control, no matter Luteolin or Rutin treatments could inhibit the medium and low RPKM genes. Compared to the number (40) of highly expressed genes unique to wild control, Luteolin treatment induced more highly expressed genes (80) while Rutin treatment decreased the number of highly expressed genes (14). As a result, the inhibitory effects of Rutin against K. pneumoniae ATCC700603 could be due to the decreased number of expressed genes, especially the highly expressed ones.
To gain a holistic understanding of the results, pairwise comparisons between wild control group (KP_C) and Rutin treatment group (KP_R) and, KP_R and Luteolin treatment group (KP_L) were compared and analyzed, respectively. More DEGs were identified after Luteolin treatment while less DEGs yielded after Rutin treatment. The numbers of DEGs were found to be 374 and 159 in KP_R vs KP_C and KP_R vs KP_L, respectively (Supplementary Material, Data sheet 1, Tables S2 and S3).
In total, 543 metabolic molecules under positive ion mode and 339 under negative ion mode were detected using an UPLC-MS detection platform (Supplementary material, Data sheet 2, Tables S1 and S2). To better understand the overall metabolic differences among the samples and the degree of variations between the samples within the groups, the metabolites were analyzed by principal component analysis (PCA). The results showed that, under positive ion mode, the principal component 1 (PC1) and principal component 2 (PC2) were accounted for 55.7% and 24.4% of the total variations, respectively (Figure 2F). Under the negative ion mode, the PC1 and PC2 were 45.4% and 31.3%, respectively (Figure 2G). Furthermore, metabolites measured in present study of these three groups (Rutin treatment group, Luteolin treatment group and control group) were clearly distinguished from each other, and the repeated samples were compactly gathered together, indicating that our experiments were reproducible and reliable.
In this project, the differentially expressed metabolites (DEMs) were screened according to the VIP (Variable Importance in the Projection) value (threshold ≥1) derived from the PLS-DA model analysis and the P-value (≤ 0.05) of the independent sample t-test. Interestingly, compared to wild control, Rutin and Luteolin treatments induced 38 and 38 DEMs, respectively (Figure 2H). There were 30 DEMs were identified in KP_R vs KP_L group. Although the total amount of DEMs was almost equal, the components of DEMs varied among different groups.
3.3. Annotation and Enrichment Analysis of DEGs
The total amount of the genes induced by Rutin treatment or Luteolin treatment were basically equal. After Rutin and Luteolin treatments, 3,682 and 3,703 genes were detected, respectively. Compared to wild control, Rutin induced 374 DEGs, among which 170 genes were down regulated while 204 genes were up regulated (Figure 3A). Analysis of their expression levels showed that, 214 DEGs were moderately expressed. The other 97 and 63 out of these DEGs were that with low and high expression levels, respectively.
Compared to Luteolin treatment, Rutin induced more down regulated genes and less up regulated genes (Figure 3C). These genes included 2552 down regulated ones and 1111 up regulated ones. Among these 3,663 genes, 104 were identified as down regulated DEGs while the other 55 were identified as up regulated DEGs. In addition, among these DEGs, 80 were moderately expressed genes, the other 64 and 15 were that with low and high expression levels, respectively.
The annotation and KEGG enrichment analysis was performed using the DEGs. The KEGG enrichment analysis of the DEGs induced by Rutin yielded 82 pathways, including metabolic pathways, ABC transporters, microbial metabolism in diverse environments etc (Supplementary material, Data sheet 1, Table S4). It is worth mentioning that, 62 DEGs were enriched in metabolic pathways, 30 DEGs were enriched in ABC transporters, and 26 DEGs were enriched in microbial metabolism in diverse environments. These 82 pathways are mainly classified into 4 categories, which were metabolism, environmental information processing, cellular processes and genetic information processing. According to the P value and FDR, the top scored 30 pathways were shown in Figure 3B. The results showed that ABC transporters was a dominant pathway that was responsible for Rutin treatment.
To find out the difference in efficacy of Rutin and Luteolin against strain growth, the DEGs induced by Rutin and Luteolin treatments were also compared and analyzed. Notably, more DEGs (104) were found to be down regulated and less DEGs (55) were found to be up regulated. These DEGs were enriched into 57 pathways (Supplementary material, Data sheet 1, Table S5). Among these pathways, 50 were metabolism related pathways, 3 were environmental information processing related pathways, 2 were genetic information processing related pathways and 2 were cellular processes related pathways. Further analysis showed that 38 DEGs were enriched in metabolic pathways, 14 DEGs were enriched in ABC transporters, 12 DEGs were enriched in biosynthesis of secondary metabolites and 11 DEGs were enriched in microbial metabolism in diverse environments. In addition, the top 30 enriched pathways identified according to p and FDR values were represented in Figure 3D. These results showed that both Rutin and Luteolin could cause changes in various pathways. Among those pathways, the metabolism related pathways were the most abundant. Although many pathways were enriched, only several of them contained a high number of DEGs.
3.4. Annotation and Enrichment Analysis of DEMs
Based on the OPLS-DA results, 38 DEMs were identified in KP_R vs KP_C group, and 30 DEMs were identified in KP_R vs KP_L group, respectively (Supplementary material, Data sheet 2, Tables S3 and S4). Among the 38 DEMs, 20 were successfully annotated to KEGG. However, among the 30 DEMs, 15 were successfully annotated to KEGG. Further analysis uncovered that 17 DEMs were found in both KP_R vs KP_C group and KP_R vs KP_L group. Compared to wild control, Rutin treatment induced 4 down regulated DEMs and 34 up regulated DEMs. However, when compared to Luteolin treatment, Rutin treatment induced 18 and 12 DEMs to be up and down regulated, respectively.
The annotation of the DEMs showed that, compared to wild control, the DEMs induced by Rutin were annotated to 25 pathways (Supplementary material, Data sheet 2, Table S5). These pathways were mainly classified into metabolism, genetic information processing, environmental information processing and organismal systems related pathways. Interestingly, the metabolism related pathways were also the most abundant as 19 out of the 25 annotated pathways were involved in metabolism, like microbial metabolism in diverse environments and ABC transporters. This result was consistent with the enrichment result in DEGs. Particularly worth mentioning is that all the annotated DEMs in KP_R vs KP_C group were up regulated. It indicated that Rutin induced the production of most of the DEMs and all the annotated ones. These DEMs were enriched in 25 pathways and shown in Figure 4A.
Similarly, the DEMs derived from the KP_R vs KP_L group were also annotated to KEGG (Supplementary material, Data sheet 2, Table S6). Together with 7 metabolism related pathways and 2 environmental information processing related pathways, the DEMs in this section were annotated to 9 pathways (Figure 4B). Unlike that in KP_R vs KP_C group, there were some up regulated DEMs and some down regulated DEMs in this group. These results indicated that the metabolism of Rutin in strains was different to that of Luteolin. Interestingly, except for the purine metabolism pathway (ko00230), all the other pathways enriched in KP_R vs KP_L group were contained in the pathways which were enriched in KP_R vs KP_C group. Further analysis showed that, the top 4 enriched pathways (ko00944, ko00943, ko00941 and ko01061) were found both in KP_R vs KP_C and KP_R vs KP_L group. These pathways are responsible for flavonoids biosynthesis or decomposition. Therefore, we concluded that, Rutin aroused the flavonoids metabolism and further damaged the balance of the growth in strains, further exhibited inhibition.
3.5. Integrated Analysis of Transcriptomics and Metabolomics
The integrated analysis of transcriptomics and metabolomics were performed. In this section, a pathway was annotated by both the transcriptome and metabolome that could be identified as a shared pathway. In KP_R vs KP_C group, 12 pathways were identified as shared pathways (Table 1). Of these pathways, 11 were metabolism related pathways and 1 was environmental information processing related. These results indicated that, Rutin caused changes in a series of metabolic pathways such as amino acid metabolism, biosynthesis of other secondary metabolites, xenobiotics biodegradation and metabolism, metabolism of cofactors and vitamins, metabolism of terpenoids and polyketides and, global and overview maps. In addition, within the metabolism related pathways, the metabolic pathways (ko01100) was the most enriched, since 62 DEGs and 8 DEMs were enriched in it. Another metabolism related pathway that was enriched was biosynthesis of secondary metabolites (ko01110). In this pathway, 24 DEGs and 9 DEMs were contained. Besides, a third enriched pathway was microbial metabolism in diverse environments (ko01120). These results proved that Rutin could change various metabolism related pathways by modulating both genes’ expression and changes in metabolites. But beyond that, Rutin also influenced environmental information processing, which was responsible for ABC transporters in membrane transport. Further analysis showed that 30 DEGs and 2 DEMs were enriched in ABC transporters (ko02010).
Similarly, the shared pathways derived from integrated metabolomic and transcriptomic analysis of KP_R vs KP_L uncovered the functional distinctions between Rutin and Luteolin. However, when compared these 2 treatments, only 4 pathways were screened as shared pathways (Table 2). Of these 4 pathways, 3 were responsible for metabolism and 1 was environmental information processing related. Interestingly, these pathways also consisted of two major blocks, metabolism and environmental information processing. The metabolic pathways were the most enriched since 38 DEGs and 7 DEMs were found in it. And then, 12 DEGs and 7 DEMs were enriched in biosynthesis of secondary metabolites. Another 14 DEGs and 1 DEMs were enriched in ABC transporters, which was responsible for membrane transporters. Lastly, 1 DEG and 1 DEM were enriched in purine metabolism.
The results derived from KP_R vs KP_C group revealed that the inhibitory function of Rutin could be due to the changes in the 12 pathways. However, the results derived from KP_R vs KP_L group could tell the reason that Luteolin exhibited no inhibitory effects. All the changes of genes and metabolites induced by Rutin and Luteolin were contained in such 4 shared pathways.
3.6. Correlation Network Between the Metabolites and Genes
The correlation between the DEMs and DEGs were analyzed and represented in heat maps (Supplementary material, Data sheet 3, Figure S1 and S2). In KP_R vs KP_C group, more DEGs showed positive correlation with DEMs and less DEGs showed negative correlation with the DEMs. This indicated that Rutin treatment promoted more gene’s expression and inhibited less genes’ expression. Herein, this phenomena may be responsible for the inhibitory effect of Rutin on strain growth by modulating expression levels of DEGs. However, in KP_R vs KP_C group, when compared to results derived from the KP_R vs KP_C group, two major differences were obtained. The first difference was that much less pathways were found in KP_R vs KP_L group. This indicated, as a flavonoid, Luteolin would participate in the same pathways and played similar roles in metabolism and growth in K. pneumoniae strains like Rutin. However, the inhibitory effect of Luteolin was not obvious. Therefore, we hypothesized the pathways which were specifically enriched in KP_R vs KP_L group, played vital roles. The second difference, one DEG from KP_R vs KP_C showed only one type of correlation with all the DEMs, positive or negative. But the DEGs derived from KP_R vs KP_L group showed both positive and negative correlation to the DEMs, even to one single DEM. These results reconfirmed the differences in efficacy and properties between Rutin and Luteolin, and also pushed us to found out more evidence.
To narrow down the DEGs and DEMs, only those with significant correlation coefficient were screed and used to build correlation network. When compared Rutin treatment to wild control, 4 pathways consisted of 23 genes and 9 metabolites were identified to be functional in strain growth (Figure 5A). Among these metabolites, C00389 (Quercetin, C15H10O7) and C00188 (L-Threonine, C4H9NO3) were the most and least abundant substances, respectively. What we can infer is that L-Threonine could be a vital substance as it was correlated with 16 genes’ expression, including 2 negatively related genes and 14 positively related ones. Further analysis showed that these genes were involved in 4 pathways, including metabolic pathways (ko01100), microbial metabolism in diverse environments (ko01120), biosynthesis of amino acids (ko01230) and ABC transporters (ko02010), further indicating that L-Threonine was functional in multiple pathways and regulated by multiple genes. Therefore, we hypothesized that Rutin induced the accumulation of L-Threonine, further inhibited strain growth through regulating genes’ expression. Another vital substance was C00255 (Vitamin B2, C17H20N4O6), which was found to be correlated with 14 genes and simultaneously associated with 3 pathways. Of these 14 genes, 9 were involved in ABC transporters. Among these ABC transporters related genes, FU841_RS01625 and FU841_RS19145 were negatively correlated with Vitamin B2, the other 7 genes were positively correlated with it. Herein, we concluded that Rutin induced the production of Vitamin B2, further changed the ABC transporters pathway by modulating genes. In addition, the results also showed that C00509 (Naringenin, C15H12O5), C05903 (Kaempferol, C15H10O6) and C00601 (Phenylacetaldehyde, C8H8O), each had 11 genes that related to them, respectively.
Similarly, the results derived from Rutin treatment and Luteolin treatment were also compared and analyzed. More DEGs were found to be clustered in the metabolic pathways while less DEGs were found in the ABC transporters pathway (Figure 5C). The metabolic pathways contained 20 genes and 1 metabolite while the ABC transporters pathway covered 3 genes and 1 metabolite. In this section, the most abundant metabolite was also Quercetin, followed by C05625 (Rutin, C27H30O16), C00858 (Formononetin, C16H12O4), Kaempferol, C00814 (Biochanin A, C16H12O5), C00387 (Guanosine, C10H13N5O5), C01514 (Luteolin, C15H10O6) and C01477 (Apigenin, C15H10O5). Here, we can see the big differences between Rutin and Luteolin treatments. Among these 8 metabolites, Apigenin, Luteolin and Guanosine were accumulated much less in Rutin treatment than that in Luteolin treatment. On the contrary, the other 5 metabolites were accumulated more in Rutin treatment than that in Luteolin treatment.
In general, from the correlation network in KP_R vs KP_C, 23 DEGs and 9 DEMs involved in 4 pathways (ko01100, ko01120, ko01230 and ko02010) were found. Similarly, 29 DEGs and 8 DEMs were identified from the correlation network of KP_R vs KP_L group. These DEGs and DEMs were involved in 3 pathways which were ko01100, ko01110 and ko02010. Because Rutin showed inhibitory effect on strains growth while Luteolin did not, we hypothesized the inhibition of Rutin was due to the 4 pathways and the inefficacy of Luteolin was caused by the 3 pathways, respectively. The metabolic pathways (ko01100) and ABC transporters (ko02010) were found in both comparative groups.
Metabolic pathways- The inhibitory function of Rutin against K. pneumoniae strains could be related to the up regulating genes in metabolic pathways since all the 9 DEGs were induced to increase their expression levels after Rutin treatment (Figure 5B). The gene FU841_RS07110 (eutC) with highest expression level, which is responsible for ethanolamine ammonia-lyase subunit, was up regulated to more than 6 fold. In addition, one of the up regulated genes with the lowest expression levels, FU841_RS07100 (eutA), which is responsible for ethanolamine ammonia-lyase reactivating factor also increased its expression level to 2.5 fold. Besides, all the DEMs in this network were also up regulated. These results indicated that, Rutin induced both the accumulation of metabolites and the up regulation of genes, further disturbed the metabolic pathways and inhibited the growth of strains.
However, when we studied the DEGs involved in metabolic pathways in KP_R vs KP_L group, we found differences. Firstly, more DEGs were found and some of them were down regulated(Figure 5D). Only four genes, FU841_RS07110 (ydfG), which is responsible for bifunctional NADP-dependent 3-hydroxy acid dehydrogenase/3-hydroxypropionate dehydrogenase; FU841_RS12940, which is responsible for NAD(P)-dependent oxidoreductase; FU841_RS00370, which is responsible for GNAT family N-acetyltransferase; and FU841_RS17580 (bioC), which is responsible for malonyl-ACP O-methyltransferase showed higher expression after Rutin treatment than that of Luteolin treatment. All the other 16 genes were down regulated. Secondly, unlike KP_R vs KP_C group, some of the DEMs in KP_R vs KP_L group were down regulated. The results showed that 2 flavonoids, Apigenin and Luteolin, and 1 riboside, Guanosine were heavily accumulated after Luteolin treatment.
Rutin, Kaempferol, Quercetin and Biochanin-A were found in both comparative groups (Supplementary material, Data sheet 2, Table S7). In this pathway, among the genes that showed correlation with Rutin, only FU841_RS17580 was found in both comparative groups. However, it showed positive correlation with Rutin in KP_R vs KP_C group while showed negative correlation with Rutin in KP_R vs KP_L group. Among the genes that correlated with Kaempferol, FU841_RS07100, FU841_RS07110 and FU841_RS17580 were found in both comparative groups. However, only FU841_RS17580 gene showed positive correlation with Kaempferol of consistency in different groups. In addition, FU841_RS07100 and FU841_RS17580 were found to be correlated with Quercetin in both comparative groups. But only FU841_RS17580 showed positive correlation with consistency in two comparative groups. Besides, no genes from two comparative groups were found to be simultaneously correlated with Biochanin A. Herein, we can conclude that although within a same pathway, Rutin and Luteolin performed properties through different manners. The gene, FU841_RS17580 which was responsible for malonyl-ACP O-methyltransferase, could play vital roles in the inhibition of Rutin against K. pneumoniae strains.
ABC transporters- Rutin exhibited inhibitory function against K. pneumoniae strains by inhibiting genes’s expression in ABC transporters. The expression of the DEGs varied in KP_R vs KP_C group. Expression of two genes, FU841_RS01625 (livF), which is responsible for high-affinity branched-chain amino acid ABC transporter ATP-binding protein, and FU841_RS19145, which is responsible for sugar ABC transporter ATP-binding protein were inhibited by Rutin. However, expression of the other genes involved in the network were promoted by Rutin. Interestingly, the expression of all three genes in KP_R vs KP_L group were inhibited by Rutin. These genes were FU841_RS29030, FU841_RS19145 and FU841_RS09200 (araG), and responsible for ABC transporter permease, sugar ABC transporter ATP-binding protein and L-arabinose ABC transporter ATP-binding protein, respectively. Once again, these results demonstrated that Rutin showed different functions to that of Luteolin. Compared to wild control, Rutin could simultaneously promote and inhibit genes’ expression. However, when compared to Luteolin trement, all selected DEGs showed down-regulation. This results derived from two comparative groups consisted with each other, further indicated that, the inhibition of Rutin against K. pneumoniae strains was more likely due to the down regulated genes’ expression in ABC transporters. Among 4 shared metabolites, no genes were found to be correlated with them simultaneously in KP_R vs KP_C and KP_R vs KP_L. Further analysis revealed that FU841_RS19145, which was responsible for sugar ABC transporter ATP-binding protein, could play vital roles in the inhibition of Rutin against K. pneumoniae strains.
3.8. Gene Validation
As we have described above, two genes FU841_RS17580 and FU841_RS19145 were screened as key functional genes. Results derived from the RNA-seq data showed that FU841_RS17580 was specifically induced by Rutin as no transcript was obtained in wild control. Another gene FU841_RS19145 was not specifically induced by Rutin since its transcripts could be detected among all samples. Further analysis showed that the lowest expression level of FU841_RS19145 was found in strains which were treated by Rutin. As a result, we extrapolated that FU841_RS19145 could be suppressed by Rutin and further limited the strain growth. Consequently, we conducted qPCR to validate their expression profiles.
The results proved that low expression of FU841_RS17580 was measured in wild control group (Figure 6A). But when the concentration of Rutin was 256 μg/mL, the expression of FU841_RS17580 was up regulated. Furthermore, when Rutin’s concentration increased to 512 μg/mL, the relative expression level of FU841_RS17580 was higher. However, when the concentration of Rutin increased, the relative expression level of FU841_RS19145 decreased (Figure 6B). The highest expression level was detected in wild control group while the lowest expression was found in the group which were treated by 512 μg/mL Rutin.
To validate the results of DEGs identified by RNA-seq, 10 candidate modulated genes were selected for quantitative real-time PCR (qRT-PCR). Results showed that the expression levels of DEGs detected by RNA-seq were consistent with that of qPCR method (Supplementary Material, Data_sheet_3, Figure S3).
4. Discussion
The emergence of antimicrobial resistance brings great challenges to clinical anti-infective therapy. This underlines the urgent need for new discovery and development of effective drugs as current treatment options for drug-resistant pathogen infections are severely limited [34]. Plant derived flavonoids are increasingly attracting attention among researchers not only because they have been indicated to possess effective properties against pathogenic microorganisms [35], but also because these phytomolecules are free of toxicity and environmental hazards [36]. However, a series of studies have focused on the inhibitory effects of flavonoids against bacteria but gained no great success [37]. Therefore, there are no classic (bacteriostatic or bactericidal) antibiotics derived from plant flavonoids on the market.
Flavonoids have been reported to possess a wide range of pharmacological properties, like antimicrobial and antiviral activities [38], but the antibacterial activities of different flavonoids components varied [39]. Rutin, a kind of flavonols, showed efficient biofilm inhibition of Streptococcus suis without impairing its growth [40]. Quercetin was reported not only to inhibit the carbapenemase and efflux pump, but also exerted bactericidal activity through disrupting cell-wall/membrane integrity and altering cellular morphology when used in combination with meropenem in carbapenem-resistant K. pneumoniae strain [41]. Another study demonstrated that Rutin showed higher antibacterial activity against K. pneumoniae RSKK 574 standard strain than its ESBL+ isolates [42]. According to our previous studies, Rutin was found to exhibit growth inhibition against K. pneumoniae strains, but other flavonoids showed weak or even no inhibitory effects [30]. In this regard, we hypothesized the inhibitory mechanism of Rutin is unique and different to other flavonoids.
To understand the inhibitory mechanism of Rutin on strain growth, both the transcriptomic and metabolomic sequencing were studied. We measured 5,483 genes and 882 metabolites and identified DEGs and DEMs. In KP_R vs KP_C group, 12 pathways were filtered out. In KP_R vs KP_L group, 4 pathways were identified. The results in present study uncovered that the inhibition of Rutin against K. pneumoniae strains mainly due to the its modulation of the metabolic pathways and ABC transporters pathways. Further analysis demonstrated that, Rutin could specifically induce the expression of FU841_RS17580 and up regulate malonyl-ACP O-methyltransferase, further inhibit the strain growth. Besides, Rutin inhibited the expression of FU841_RS19145 and further inhibited the function of sugar ABC transporter ATP-binding protein, then inhibited strain growth.
We demonstrated the mechanism through which Rutin could inhibit the strain growth by narrowing down the DEGs and DEMs. There were 13 DEMs that were identified in this study. In addition to 4 DEMs that they shared, two comparative groups also have their own unique metabolites. The unique substances identified in KP_R vs KP_C were L-Threonine, Naringenin, Vitamin_B2, C00196 (2,3-Dihydroxybenzoic_acid, C7H6O4) and Phenylacetaldehyde. Each of these metabolites was correlated with several genes, indicated that they played vital roles in the pathways. In addition, these metabolites included amino acids, flavonoids and Vitamin, indicating the inhibitory effects on K. pneumoniae strains were involved in multiple pathways and complicated. Regarding to the metabolites that unique to KP_R vs KP_L, big difference was found. Firstly, among the 4 kinds of metabolites, Apigenin, Luteolin and Formononetin were flavonoids, and one Guanosine. Secondly, the ko01110 pathway was unique in this group. Therefore, we supposed different flavonoids induced different changes in K. pneumoniae strains. However, some of the changes could influence the growth while some others could not.
Klebsiella pneumoniae is a common, Gram-negative organism. It is a significant pathogenic factor of severe infections in humans. Due to its natural resistance to antibiotics, infection with this pathogen can cause severe therapeutic problems. In present study we have provided valuable information about metabolic pathways and ABC transporters pathways leading to an better understanding of the inhibitions of K. pneumoniae strains, which can further lead to new drug development of new diagnostics and treatment strategies.
5. Conclusions
The present study suggested that Rutin has the ability to inhibit K. pneumoniae growth by modulating the metabolic pathways and ABC transporters pathways. Also, the inhibitory mechanism of Rutin was different to that of Luteolin. Although these two flavonoids shared two same pathways in K. pneumoniae strains, the DEGs and DEMs were different. Two genes, FU841_RS17580 and FU841_RS19145 were identified as key genes playing vital roles in the process of inhibition. In conclusion, these results derived from both the associative omics study and intracellular experiments are encouraging for further biological or phytochemical studies aimed at new development of antimicrobial agents of plant origin.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1, title, Table S1, title, Video S1, title.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This research was funded by Sichuan Science and Technology Program (22022, 2022YFS0633), Scientific Startup Funds for Doctors (19076) and Postdoctoral fellows(20003) of the Affiliated Hospital of Southwest Medical University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Please refer to suggested Data Availability Statements in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics. You might choose to exclude this statement if the study did not report any data.
Acknowledgments
We are grateful to Professor Xiaochun Zhao and his team members from Citrus Research Institute, Southwest University, China, for their good suggestions in flavonoids selection. Also, we appreciate the funding support from Luzhou Government and Southwest Medical University Cooperation Program (2020LZXNYDJ46).
Conflicts of Interest
The authors declare they have no conflict of interest. In addition, the funders had no roles in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Kupnik, K.; Primožič, M.; Vasić, K.; Knez, Ž.; Leitgeb, M. A Comprehensive Study of the Antibacterial Activity of Bioactive Juice and Extracts from Pomegranate ( Punica granatum L.) Peels and Seeds. Plants (Basel, Switzerland) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Yang. Z; Cai. Y.; Liu, H.; Li, S.; Kou. N.; Wu. J.; Zhang. Q. Effect of Bloodletting at Shaoshang and Shangyang Acupuncture Points on Outcome and Prognosis of Severe Community-Acquired Pneumonia in the Elderly. Disease markers 2021, 2021, 3295021. [Google Scholar] [CrossRef] [PubMed]
- Wang. P.; Jiang. S.; Li. Y.; Luo. Q.; Lin. J.; Hu. L.; Liu. X.; Xue. F.Virus-like mesoporous silica-coated plasmonic Ag nanocube with strong bacteria adhesion for diabetic wound ulcer healing. Nanomedicine : nanotechnology, biology, and medicine 2021, 34, 102381. [Google Scholar]
- Singh. V.; Chibale. K. Strategies to Combat Multi-Drug Resistance in Tuberculosis. Accounts of chemical research 2021, 54, 2361–2376. [Google Scholar] [CrossRef] [PubMed]
- 5. Montenegro. C.; Gonçalves. G.; Oliveira Filho. A.; Lira. A.; Cassiano. T.; Lima. N.; Barbosa-Filho. J.; Diniz. M.; Pessôa. H. In Silico Study and Bioprospection of the Antibacterial and Antioxidant Effects of Flavone and Its Hydroxylated Derivatives. Molecules (Basel, Switzerland), 2017; 22.
- Farhadi. F.; Khameneh. B.; Iranshahi. M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytotherapy research : PTR 2019, 33, 13–40. [Google Scholar] [CrossRef]
- Lin. S.; Wade. J.; Liu. S. De Novo Design of Flavonoid-Based Mimetics of Cationic Antimicrobial Peptides: Discovery, Development, and Applications. Accounts of chemical research 2021, 54, 104–119. [Google Scholar] [CrossRef] [PubMed]
- 8. Sheng. Z.; Jiang. Y.; Liu. J.; Yang. B. UHPLC-MS/MS Analysis on Flavonoids Composition in and Their Antioxidant Activity. Antioxidants (Basel, Switzerland), 2021; 10.
- Wang. Z.; Yu. Q.; Shen. W.; El Mohtar. C.; Zhao. X.; Gmitter. F. Functional study of CHS gene family members in citrus revealed a novel CHS gene affecting the production of flavonoids. BMC plant biology 2018, 18, 189. [Google Scholar]
- Li. G.; Ding. K.; Qiao. Y.; Zhang. L.; Zheng. L.; Pan. T.; Zhang. L. Flavonoids Regulate Inflammation and Oxidative Stress in Cancer. Molecules 2020, 25, 5628. [Google Scholar] [CrossRef] [PubMed]
- Harborne. JB.; Williams. CA. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Cushnie. T.; Lamb. A. Antimicrobial activity of flavonoids. International journal of antimicrobial agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Farhadi. F.; Khameneh. B.; Iranshahi. M.; Iranshahy. M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytotherapy research : PTR 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- 14. Aleebrahim-Dehkordy. E.; Rafieian-Kopaei. M. In Vitro Evaluation of Antioxidant Activity and Antibacterial Effects and Measurement of Total Phenolic and Flavonoid Contents of Quercus brantii L. Fruit Extract. 2019; 16, 408–416.
- Benevides Bahiense. J.; Marques. F.; Figueira. M.; Vargas. T.; Kondratyuk. T.; Endringer. D.; Scherer. R.; Fronza. M. Potential anti-inflammatory, antioxidant and antimicrobial activities of Sambucus australis. Pharmaceutical biology 2017, 55, 991–997. [Google Scholar] [CrossRef] [PubMed]
- 16. Duda-Madej. A.; Kozłowska. J.; Krzyżek. P.; Anioł. M.; Seniuk. A.; Jermakow. K.; Dworniczek. E. Antimicrobial O-Alkyl Derivatives of Naringenin and Their Oximes Against Multidrug-Resistant Bacteria. Molecules (Basel, Switzerland), 2020; 25.
- Xiao. Z.; Wang. X.; Wang. P.; Zhou. Y.; Zhang. J.; Zhang. L.; Zhou. J.; Zhou. S.; Ouyang. H.; Lin. X. Design, synthesis and evaluation of novel fluoroquinolone-flavonoid hybrids as potent antibiotics against drug-resistant microorganisms. European journal of medicinal chemistry 2014, 80, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Eumkeb. G.; Chukrathok. S. Synergistic activity and mechanism of action of ceftazidime and apigenin combination against ceftazidime-resistant Enterobacter cloacae. Phytomedicine 2013, 20, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice. A.; Bruni. V.; Michaud. L. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. Journal of basic microbiology 2007, 47, 496–505. [Google Scholar] [CrossRef]
- Wallace. J.; Bowlin. N.; Mills. D.; Saenkham. P.; Kwasny. S.; Opperman. T.; Williams. J.; Rock. C.; Bowlin. T.; Moir. D. Discovery of bacterial fatty acid synthase type II inhibitors using a novel cellular bioluminescent reporter assay. Antimicrobial agents and chemotherapy 2015, 59, 5775–5787. [Google Scholar] [CrossRef] [PubMed]
- Rütschlin. S.; Böttcher. T. Inhibitors of Bacterial Swarming Behavior. Chemistry (Weinheim an der Bergstrasse, Germany) 2020, 26, 964–979. [Google Scholar]
- Górniak. I.; Bartoszewski. R.; Króliczewski. J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochemistry Reviews 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Lee. J.; Regmi. S.; Kim. J.; Cho. M.; Yun. H.; Lee. C.; Lee. J. Apple flavonoid phloretin inhibits Escherichia coli O157, H7 biofilm formation and ameliorates colon inflammation in rats. Infection and immunity 2011, 79, 4819–4827. [Google Scholar] [CrossRef]
- Silva. L.; Zimmer. K.; Macedo. A.; Trentin. D. Plant Natural Products Targeting Bacterial Virulence Factors. Chemical reviews 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Tsou. L.; Lara-Tejero. M.; RoseFigura. J.; Zhang. Z.; Wang. Y.; Yount. J.; Lefebre. M.; Dossa. P.; Kato. J.; Guan. F. Antibacterial Flavonoids from Medicinal Plants Covalently Inactivate Type III Protein Secretion Substrates. Journal of the American Chemical Society 2016, 138, 2209–2218. [Google Scholar] [CrossRef] [PubMed]
- Mehta. J.; Utkarsh. K.; Fuloria. S.; Singh. T.; Sekar. M.; Salaria. D.; Rolta. R.; Begum. MY.; Gan. SH.; Rani. NNIM. Antibacterial Potential of Bacopa monnieri (L.) Wettst. and Its Bioactive Molecules against Uropathogens—An In Silico Study to Identify Potential Lead Molecule(s) for the Development of New Drugs to Treat Urinary Tract Infections. Molecules 2022, 27, 4971. [Google Scholar]
- Ble-González. EA.; Gómez-Rivera. A.; Zamilpa. A.; López-Rodríguez. R.; Lobato-García. CE.; Álvarez-Fitz. P.; Gutierrez-Roman. AS.; Perez-García. MD.; Bugarin. A.; González-Cortazar. M. Ellagitannin, Phenols, and Flavonoids as Antibacterials from Acalypha arvensis (Euphorbiaceae). Plants 2022, 11, 300. [Google Scholar]
- Cushnie. T.; Lamb. A. Recent advances in understanding the antibacterial properties of flavonoids. International journal of antimicrobial agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Lewis. K.; Ausubel. F. Prospects for plant-derived antibacterials. Nature biotechnology 2006, 24, 1504–1507. [Google Scholar] [CrossRef] [PubMed]
- Wang. Z.; Ding. Z.; Li. Z.; Ding. Y.; Jiang. F.; Liu. J. Antioxidant and antibacterial study of 10 flavonoids revealed rutin as a potential antibiofilm agent in Klebsiella pneumoniae strains isolated from hospitalized patients. Microbial pathogenesis 2021, 105121. [Google Scholar]
- Chia. L.; Hornung. B.; Aalvink. S.; Schaap. P.; de Vos. W.; Knol. J.; Belzer. C. Deciphering the trophic interaction between Akkermansia muciniphila and the butyrogenic gut commensal Anaerostipes caccae using a metatranscriptomic approach. Antonie van Leeuwenhoek 2018, 111, 859–873. [Google Scholar] [CrossRef] [PubMed]
- De Araujo. C.; Balestrino. D.; Roth. L.; Charbonnel. N.; Forestier, C. Quorum sensing affects biofilm formation through lipopolysaccharide synthesis in Klebsiella pneumoniae. Research in microbiology 2010, 161, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Livak. KJ.; Schmittgen. TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Inagaki. K.; Lucar. J.; Blackshear. C.; Hobbs. CV. Methicillin-susceptible and Methicillin-resistant Staphylococcus aureus Bacteremia: Nationwide Estimates of 30-Day Readmission, In-hospital Mortality, Length of Stay, and Cost in the United States. Clinical Infectious Diseases 2019, 69, 2112–2118. [Google Scholar]
- Acquaviva. R.; D’Angeli. F.; Malfa. GA.; Ronsisvalle. S.; Garozzo. A.; Stivala. A.; Ragusa. S.; Nicolosi. D.; Salmeri. M.; Genovese. C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Natural product research 2021, 35, 2076–2081. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafi. S.; Al-Mohammadi. A-R.; Sitohy. M.; Mosa. B.; Ismaiel. A.; Enan. G.; Osman. A. Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris. Molecules 2019, 24, 4280. [Google Scholar] [CrossRef] [PubMed]
- Khan. M.; Tang. H.; Lyles. J.; Pineau. R.; Mashwani. Z.; Quave. C. Antibacterial Properties of Medicinal Plants From Pakistan Against Multidrug-Resistant ESKAPE Pathogens. Frontiers in pharmacology 2018, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Wang. T-y.; Li. Q.; Bi. K-s. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian Journal of Pharmaceutical Sciences 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Mandalari. G.; Bennett. R.; Bisignano. G.; Trombetta. D.; Saija. A.; Faulds. C.; Gasson. M.; Narbad. A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. Journal of applied microbiology 2007, 103, 2056–2064. [Google Scholar] [CrossRef]
- Wang. S.; Wang. C.; Gao. L.; Cai. H.; Zhou. Y.; Yang. Y.; Xu. C.; Ding. W.; Chen. J.; Muhammad. I. Rutin Inhibits Streptococcus suis Biofilm Formation by Affecting CPS Biosynthesis. Front Pharmacol 2017, 8, 379. [Google Scholar] [CrossRef] [PubMed]
- Pal. A.; Tripathi. A. Quercetin inhibits carbapenemase and efflux pump activities among carbapenem-resistant Gram-negative bacteria. APMIS : acta pathologica, microbiologica, et immunologica Scandinavica 2020, 128, 251–259. [Google Scholar] [CrossRef]
- Orhan. D.; Ozçelik. B.; Ozgen. S.; Ergun. F. Antibacterial, antifungal, and antiviral activities of some flavonoids. Microbiological research 2010, 165, 496–504. [Google Scholar] [CrossRef]
Figure 1.
Inhibitory effects of Rutin against the growth of K. pneumoniae ATCC700603. (A) Growth states at different time points, observation by CLSM. Up, effect of Rutin in the growth of the planktonic cells after treatments (4h). Down, effect of Rutin treatment (8h) on the planktonic cells growth of this strain. (B) Biomass accumulation with or without Rutin treatments. The wild control group was compared to all the other groups, respectively. *** indicates P<0.001 and ns indicates not significant. The working concentration of Gentamicin used in this study was 50 μg/mL.
Figure 1.
Inhibitory effects of Rutin against the growth of K. pneumoniae ATCC700603. (A) Growth states at different time points, observation by CLSM. Up, effect of Rutin in the growth of the planktonic cells after treatments (4h). Down, effect of Rutin treatment (8h) on the planktonic cells growth of this strain. (B) Biomass accumulation with or without Rutin treatments. The wild control group was compared to all the other groups, respectively. *** indicates P<0.001 and ns indicates not significant. The working concentration of Gentamicin used in this study was 50 μg/mL.
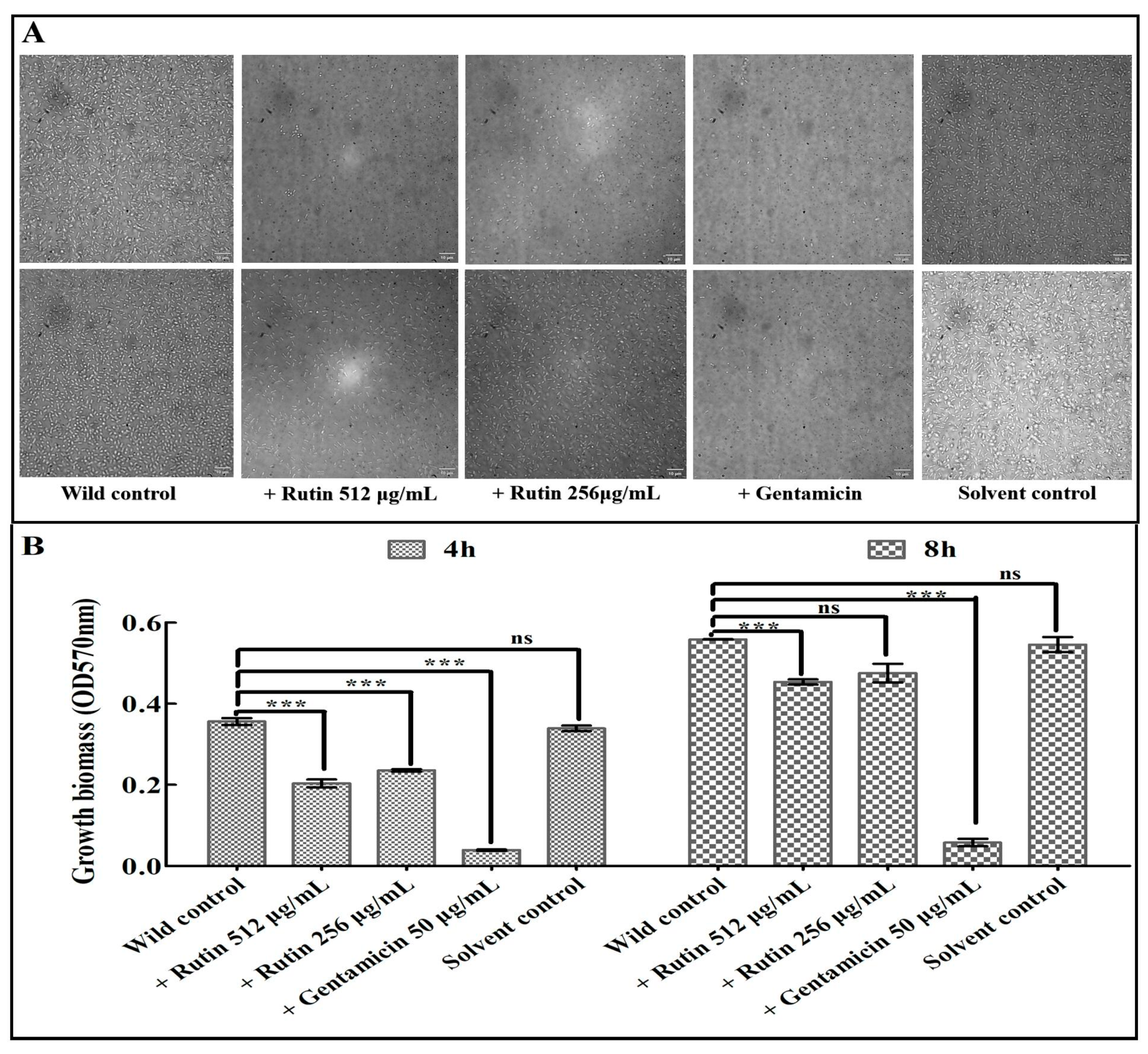
Figure 2.
Quantification of transcript expression levels and metabolites. (A) Number of genes and their expression levels measured in 6 libraries. (B) Venn diagram showing numbers of all genes measured in three groups. (C) Venn diagram showing numbers of genes with low expression levels. (D) Venn diagram showing numbers of genes with medium expression levels. (E) Venn diagram showing numbers of genes with high expression levels. (F) Scattered point plots are obtained for PCA analysis of positive ion mode. (G) Scattered point plots are obtained for PCA analysis of negative ion mode. (H) Venn diagram showing numbers of DEMs. C: wild control; R: 512 μg/mL Rutin treatment; L: 512 μg/mL Luteolin treatment. The genes shown in venn diagrams were compared in pairs among KP_R group, KP_L group and KP_C group, respectively.
Figure 2.
Quantification of transcript expression levels and metabolites. (A) Number of genes and their expression levels measured in 6 libraries. (B) Venn diagram showing numbers of all genes measured in three groups. (C) Venn diagram showing numbers of genes with low expression levels. (D) Venn diagram showing numbers of genes with medium expression levels. (E) Venn diagram showing numbers of genes with high expression levels. (F) Scattered point plots are obtained for PCA analysis of positive ion mode. (G) Scattered point plots are obtained for PCA analysis of negative ion mode. (H) Venn diagram showing numbers of DEMs. C: wild control; R: 512 μg/mL Rutin treatment; L: 512 μg/mL Luteolin treatment. The genes shown in venn diagrams were compared in pairs among KP_R group, KP_L group and KP_C group, respectively.
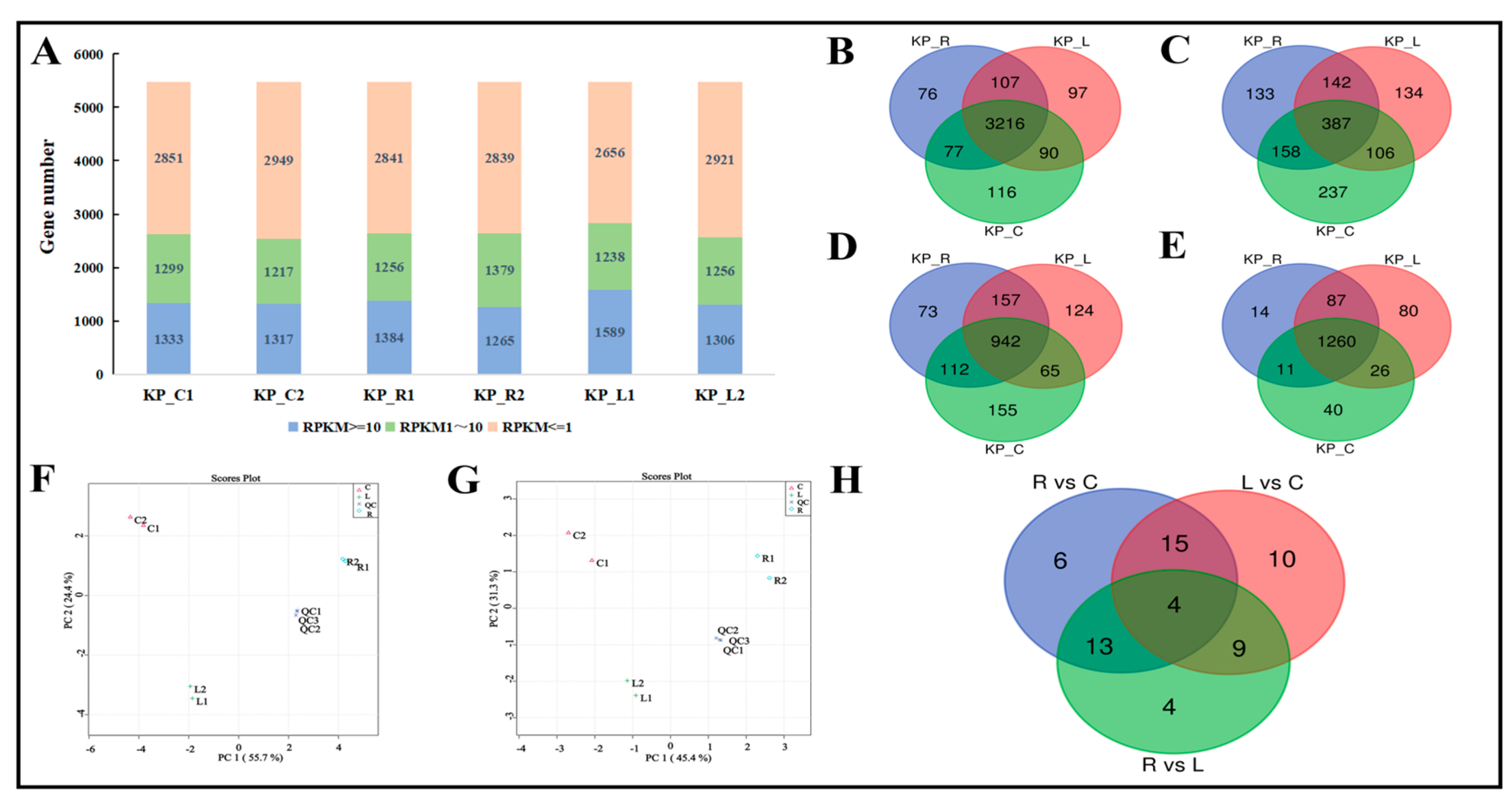
Figure 3.
KEGG enrichment of DEGs induced by Rutin and Luteolin treatments. (A) Genes induced by Rutin treatment, compared to wild control. (B) KEGG enrichment of DEGs in KP_C vs KP_R group. (C) Genes induced by Rutin treatment, compared to Luteolin treatment. (D) KEGG enrichment of DEGs derived from the KP_R vs KP_L group. Red indicates up regulated DEGs; blue indicates down regulated DEGs.
Figure 3.
KEGG enrichment of DEGs induced by Rutin and Luteolin treatments. (A) Genes induced by Rutin treatment, compared to wild control. (B) KEGG enrichment of DEGs in KP_C vs KP_R group. (C) Genes induced by Rutin treatment, compared to Luteolin treatment. (D) KEGG enrichment of DEGs derived from the KP_R vs KP_L group. Red indicates up regulated DEGs; blue indicates down regulated DEGs.
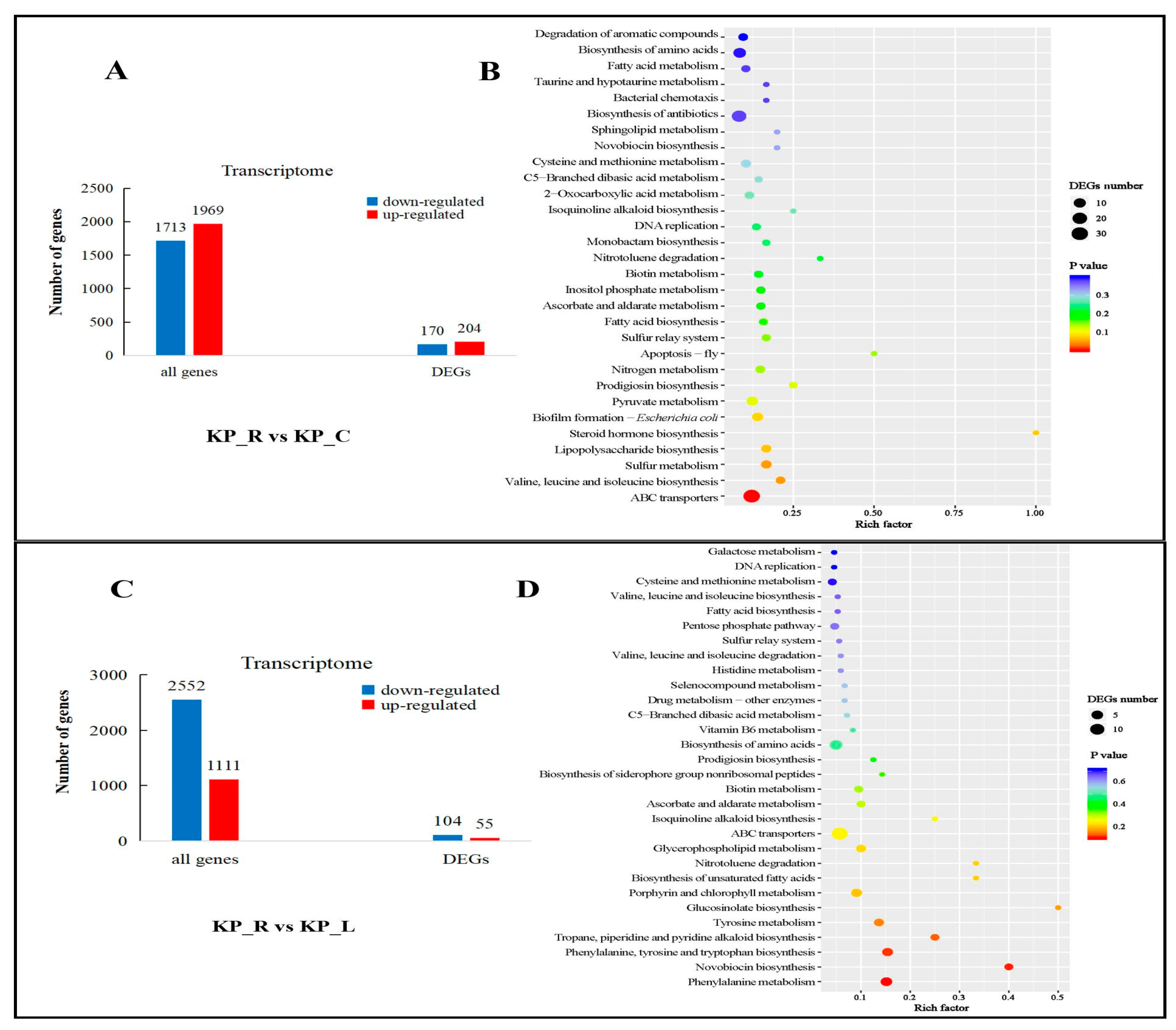
Figure 4.
KEGG enrichment of DEMs. (A) KEGG enrichment of the DEMs induced by Rutin treatment, compared to wild control. (B) KEGG enrichment of DEMs derived from KP_R vs KP_C group.
Figure 4.
KEGG enrichment of DEMs. (A) KEGG enrichment of the DEMs induced by Rutin treatment, compared to wild control. (B) KEGG enrichment of DEMs derived from KP_R vs KP_C group.
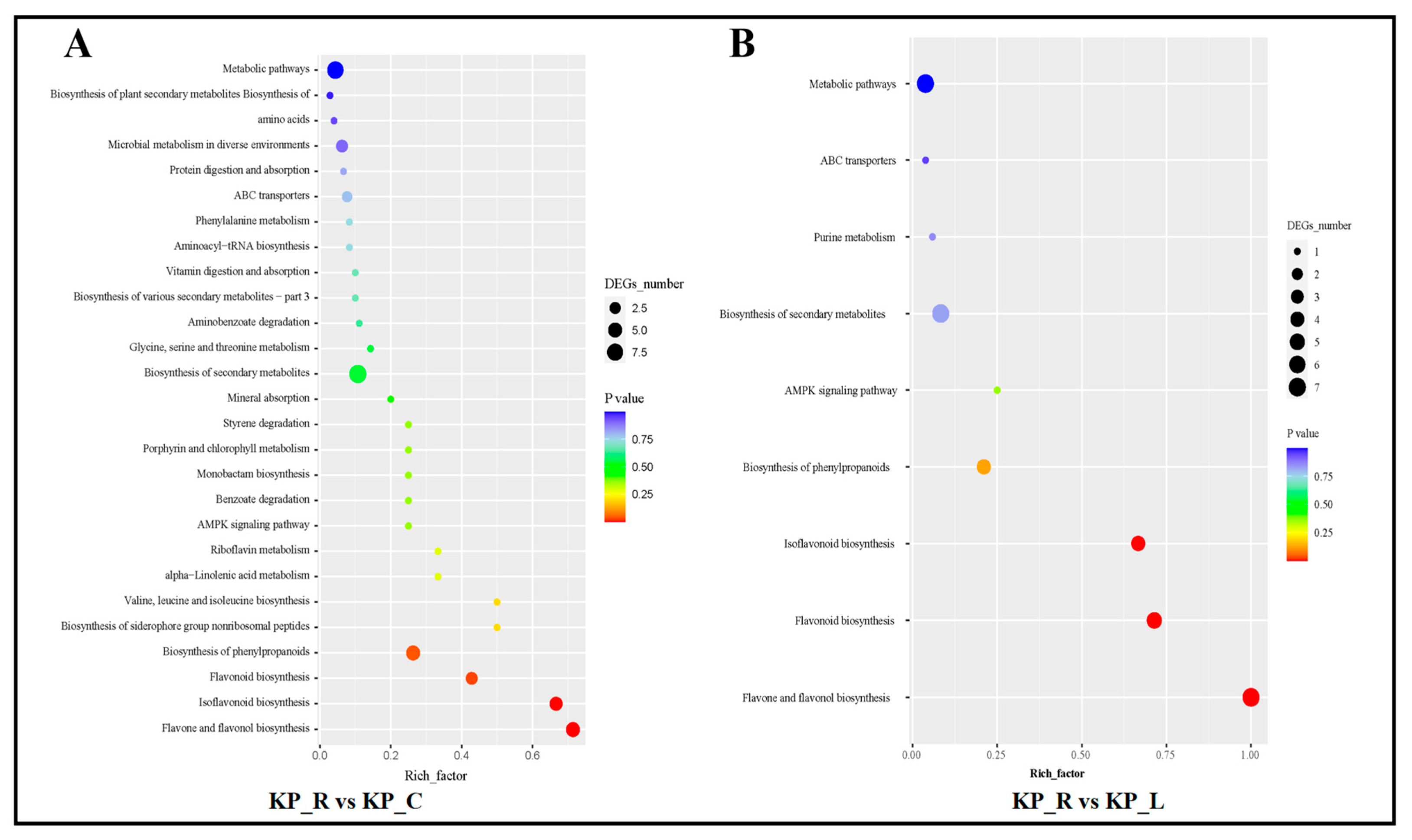
Figure 5.
Correlation network of DEGs and DEMs. (A) Correlation network of KP_R vs KP_C group. (B) The heat map of DEGs and DEMs, derived from KP_R vs KP_C group. (C) Correlation network of KP_R vs KP_L group. (D) The heat map of DEGs and DEMs, derived from KP_R vs KP_L group. In A and C, blue colour represents metabolic pathways (ko01100), purple colour represents microbial metabolism in diverse environments (ko01120), yellow colour represents biosynthesis of amino acids (ko01230), green colour represents ABC transporters (ko02010), and gray colour represents biosynthesis of secondary metabolites (ko01110). DEMs or DEGs with a same colour were involved in a same pathway. Rectangle represents gene and ellipse represents metabolite. Dash line represents negative correlation and solid line represents positive correlation. In B and D, red represents high expression, and blue represents low expression.
Figure 5.
Correlation network of DEGs and DEMs. (A) Correlation network of KP_R vs KP_C group. (B) The heat map of DEGs and DEMs, derived from KP_R vs KP_C group. (C) Correlation network of KP_R vs KP_L group. (D) The heat map of DEGs and DEMs, derived from KP_R vs KP_L group. In A and C, blue colour represents metabolic pathways (ko01100), purple colour represents microbial metabolism in diverse environments (ko01120), yellow colour represents biosynthesis of amino acids (ko01230), green colour represents ABC transporters (ko02010), and gray colour represents biosynthesis of secondary metabolites (ko01110). DEMs or DEGs with a same colour were involved in a same pathway. Rectangle represents gene and ellipse represents metabolite. Dash line represents negative correlation and solid line represents positive correlation. In B and D, red represents high expression, and blue represents low expression.
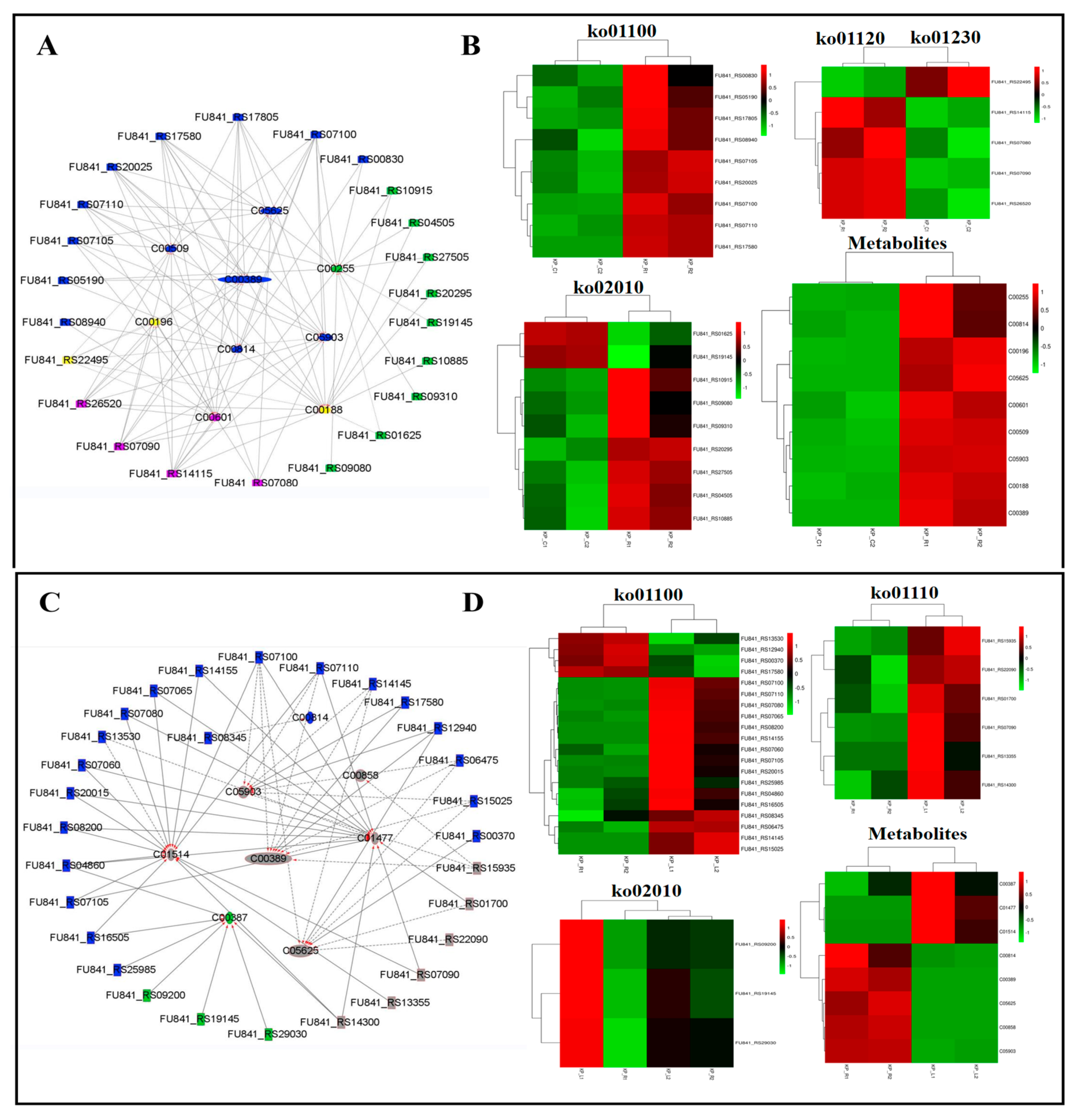
Figure 6.
Relative expression profiles of two genes. (A) The expression of FU841_RS17580 after Rutin treatment. (B) The expression of FU841_RS19145 after Rutin treatment. * indicates P<0.05, ** indicates P<0.01 and ns indicates not significant.
Figure 6.
Relative expression profiles of two genes. (A) The expression of FU841_RS17580 after Rutin treatment. (B) The expression of FU841_RS19145 after Rutin treatment. * indicates P<0.05, ** indicates P<0.01 and ns indicates not significant.
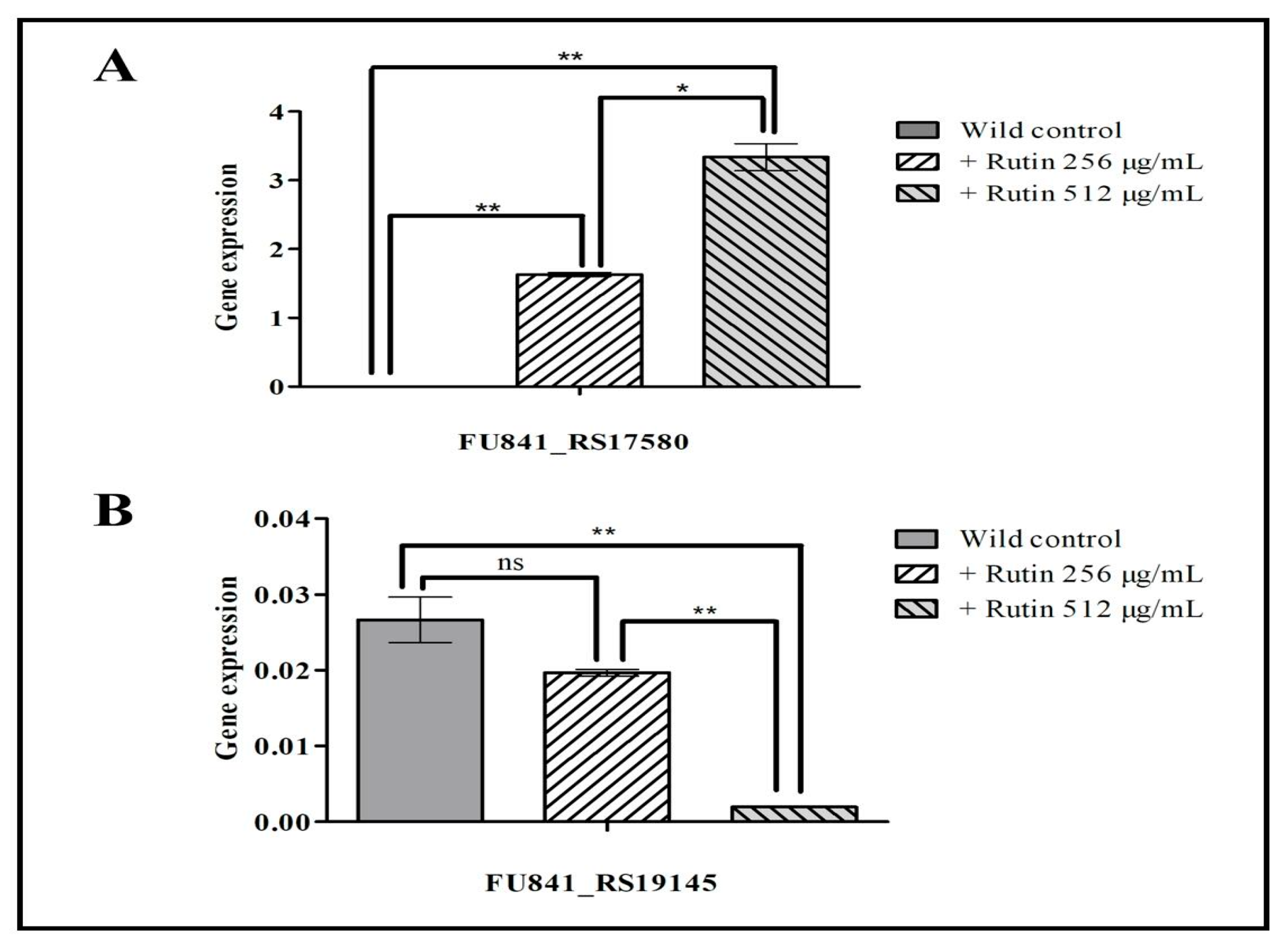

Table 1.
The pathways derived from the annotations of both the transcriptomic and metabolomic analysis in KP_R vs KP_C group.
Table 1.
The pathways derived from the annotations of both the transcriptomic and metabolomic analysis in KP_R vs KP_C group.
| z | Layer 1 | Layer 2 | Lyaer 3 | DEGs | DEMs |
|---|---|---|---|---|---|
| ko00260 | Metabolism | 1.5 Amino acid metabolism | Glycine, serine and threonine metabolism | 4 | 1 |
| ko00261 | Metabolism | 1.10 Biosynthesis of other secondary metabolites | Monobactam biosynthesis | 2 | 1 |
| ko00290 | Metabolism | 1.5 Amino acid metabolism | Valine, leucine and isoleucine biosynthesis | 4 | 1 |
| ko00360 | Metabolism | 1.5 Amino acid metabolism | Phenylalanine metabolism | 2 | 1 |
| ko00362 | Metabolism | 1.11 Xenobiotics biodegradation and metabolism | Benzoate degradation | 3 | 1 |
| ko00740 | Metabolism | 1.8 Metabolism of cofactors and vitamins | Riboflavin metabolism | 1 | 1 |
| ko01053 | Metabolism | 1.9 Metabolism of terpenoids and polyketides | Biosynthesis of siderophore group nonribosomal peptides | 1 | 1 |
| ko01100 | Metabolism | 1.0 Global and overview maps | Metabolic pathways | 62 | 8 |
| ko01110 | Metabolism | 1.0 Global and overview maps | Biosynthesis of secondary metabolites | 24 | 9 |
| ko01120 | Metabolism | 1.0 Global and overview maps | Microbial metabolism in diverse environments | 26 | 3 |
| ko01230 | Metabolism | 1.0 Global and overview maps | Biosynthesis of amino acids | 12 | 1 |
| ko02010 | Environmental Information Processing | 3.1 Membrane transport | ABC transporters | 30 | 2 |
Table 2.
The pathways derived from the annotations of both the transcriptomic and metabolomic analysis of KP_R vs KP_L group.
Table 2.
The pathways derived from the annotations of both the transcriptomic and metabolomic analysis of KP_R vs KP_L group.
| Pathway | Layer 1 | Layer 2 | Layer 3 | DEGs | DEMs |
|---|---|---|---|---|---|
| ko00230 | Metabolism | 1.4 Nucleotide metabolism | Purine metabolism | 1 | 1 |
| ko01100 | Metabolism | 1.0 Global and overview maps | Metabolic pathways | 38 | 7 |
| ko01110 | Metabolism | 1.0 Global and overview maps | Biosynthesis of secondary metabolites | 12 | 7 |
| ko02010 | Environmental Information Processing | 3.1 Membrane transport | ABC transporters | 14 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



