
Submitted:
13 March 2023
Posted:
15 March 2023
You are already at the latest version
Abstract
The winged bean is an underutilized legume that has the potential to contribute to nutrition and food security globally, especially in sub-Saharan Africa (SSA). However, very little research attention is paid to exploiting its full potential due to a lack of adequate knowledge of the existing genetic diversity in the available winged bean germplasm. To bridge this gap, seed morphometric and selected agro-morphological traits which are crucial for yield and seed quality determination in crop plants were used to assess the genetic diversity of thirty accessions of winged bean sourced from the Gene bank of the International Institute of Tropical Agriculture (IITA), Ibadan. The thirty winged accessions were evaluated under field trials at three agro-ecological zones in Nigeria. The agro-morphological data as well as the seed morphometric dataset obtained from a Multi-Spectral Imaging (MSI) system were subjected to Analysis of Variance (ANOVA), Principal Component (PC) analysis, cluster analysis, and correlation analysis. Significant differences (p < 0.05) were observed among the accessions for all measured traits. The first three PCs accounted for 93% of the variation observed among the accessions for all measured traits. Accessions were grouped into three clusters based on the agro- morphological traits and three clusters under the MSI system. Seed yield per plant had the highest estimates of significant positive correlation with hundred seed weight, pod weight per plant, etc. at both phenotypic and genotypic levels. TPt-31 had the highest seed yield per plant, pod weight per plant, and early maturity while TPt-7 had an extended flowering and maturity period, the highest number of pods per peduncle and pods per plant as well as the lowest seed yield. These accessions could serve as a good resource for future winged bean improvement programs. The result also confirmed that the MSI system is an invaluable tool for discriminating among accessions of the same crop species.
Keywords:
Genetic diversity
; Underutilized legume
; Multispectral imaging system
; Agro-morphological traits
; Winged bean
; Morphometric traits
1. Introduction
Winged bean (Psophocarpus tetragonolobus (L.) DC.) is a neglected and underutilized legume that thrives well in hot, humid equatorial countries [1]. It belongs to the family Fabaceae. It is predominantly grown in Papua New Guinea, Malaysia, Thailand, Bangladesh, Indonesia, Ghana and to a lesser extent in Nigeria [2]. It is nutritious with an exceptionally high amount of proteins, vitamins, and fatty oils that could play a major role in the diet improvement of people in SSA countries [3]. It is a multipurpose legume grown for its edible seeds, vegetable and root tuber. The dried tubers and mature seeds contain about 17–19% and 28-45% proteins, respectively, as well as a high oil contents of about 14-20.4% similar to that of soybean [4,5,6]. A study by Amoo et al. [7] revealed that winged bean flour has the potential to reduce dietary deficiency in children and adults of SSA when blended and prepared into different meals. Due to the increasing global food demand, the winged bean is a prospective crop that when cultivated and utilized, could enhance nutrition and food security especially in the developing countries of SSA [2].
Despite the nutritional potentials of winged bean, very little attention is paid to exploiting it due to its long maturation period, indeterminate growth habit, low seed yield and the need of trellis for support, as well as the presence of anti- nutritive factors including chymotrypsin inhibitors, hemagglutinins and trypsin inhibitors [4]. In addition, there is limited information about the existing genetic diversity among the available winged bean germplasm [8]. Understanding the genetic diversity of existing landraces from various geographical regions is critical for their conservation and subsequently, their utilization in breeding efforts [2]. Information on genetic diversity is needed by plant breeders in developing effective approaches as well as provides the information on heterotic combinations in crop improvement programs [9]. Several efforts have been made by researchers to assess the genetic diversity of winged bean using agro-morphological traits [10,11,12,13]. For example, Yulianah et al. [10] assessed genetic diversity of 21 Indonesian winged bean accessions based on morphological characters and they reported that phenotypic traits including pod length, number of pods per plant and total pod weight per plant are useful in discriminating among the winged bean accessions. Similarly, Wong et al. [13] evaluated 24 winged bean accessions sourced from Nigeria, Malaysia, Bangladesh, Papua New Guinea, Indonesia, Sri Lanka and Liberia based on their yield-related traits. Apart from agro-morphological traits, molecular tools have also been employed to assess the genetic diversity of winged bean accessions using molecular markers. Mohanty et al. [6] used Inter Simple Sequence Repeat (ISSR) and Random Amplified Polymorphic DNA (RAPD) markers to assess the genetic diversity of 24 winged bean accessions. In another study, Laosatit et al. [8] employed 14 simple sequence repeats (SSR) markers to assess the genetic diversity and the population structure in 457 accessions of winged bean collected from six geographical regions in Thailand. However, it is important to assess the genetic diversity of newly collected winged accessions together with already existing accessions in the IITA-Gene bank.
Multi Spectral Imaging (MSI) system is an integrated computer-aided image analysis system for assessing morphometry and distinguishing variations in seeds [14]. It measures a variety of seed morphometric features and provides a more accurate assessment of the seed features such as size, color, shape, and texture to quantitatively discriminate among different accessions [15]. Additionally, it replaces subjective and perception- based evaluation with a more accurate objective quantification in the analysis of seed morphometric variation [14,15]. Venora et al. [16] used an image analysis system for varietal identification of 15 Italian common bean landraces based on the seed size and color descriptors. Similarly, Kilic et al. [17] developed a computer vision system and artificial neural network based on seed size and color for classification of 69 common bean samples. In addition, Geetha et al. [18] used seed image analyzer to characterize the shape descriptors of 9 mustard genotypes. They found most of the seed geometric features including area, perimeter, circularity and length useful in discriminating the genotypes. However, the application and effectiveness of MSI system in assessing the genetic diversity among winged bean accessions is yet to be reported. The MSI system offers to be an invaluable tool in the phenotypic characterization of winged bean accessions in the IITA-Gene bank. Hence, the objective of this study was to assess the genetic diversity of thirty winged bean accessions using agro-morphological traits and seed morphometric traits derived from a MSI system.
2. Materials and Methods
2.1. Genetic Materials, Experimental Locations and Experimental Design
Thirty accessions of winged bean sourced from IITA-Gene bank (Table 1) were evaluated under rainfed conditions during the growing seasons of year 2020 at three (3) IITA research stations (Ubiaja - Humid rainforest; 6°65ʹN 6°38ʹE; Ikenne - Lowland humid forest; 6°92ʹN 3°73ʹE and Ibadan - Forest-savanna transition zone; 7°48ʹN 3°89ʹE). The trials were laid out in a Randomized Complete Block Design (RCBD) with three replications. Seeds of each accession were sown on a 14 m row plot with a spacing of 1 m inter and intra row spacing. Hand weeding was carried out to get a clean field throughout the experiment. Data on yield and other seed traits were collected using the Bioversity revised descriptor list for winged bean as a guide https://www.bioversityinternational.org/e-library/publications/detail/winged-bean-descriptors-revised/.
2.1.1. Data Analyses
The agro-morphological data were subjected to Analysis of Variance (ANOVA) using the ‘lme4’ package in R (R Core Team, 2022) [19]. Phenotypic and genotypic correlation analyses of traits were carried out using ‘variability’ package in R and was used to determine the level of association among the traits measured [20]. Principal Component Analysis (PCA) was used to obtain information on the traits that mostly contributed to the overall variability observed among the accessions. Cluster Analysis was used to group accessions based on similarity among agro-morphological traits and this was done using the Ward minimum variance method [21].
2.2. Multispectral imaging analysis
VideometerLab4TM (Videometer A/S, Herlev, Denmark) was used to capture images of ten seeds from each of the thirty winged bean accessions with three replications. Feature information was extracted from the scanned images with the aid of the Videometer lab software version 3.14.9 as described by Galetti et al. [22]. Data on seed morphometric traits including size and shape features such as seed length, width, seed area, seed perimeter, seed form factor, seed eccentricity, seed rectangularity, seed compactness and seed hue were captured. Other seed shape parameters such as seed circularity index and seed eccentricity index were estimated from the measured features using methods from Cervantes et al. [23]; Daniel et al. [24]; and Adewale et al. [25] (Table 2). The means from the ten individual samples in the three replicates for each accession were computed for the eleven morphometric traits. Pearson’s correlation analysis was performed to determine the level of association among seed morphometric characters. Single Linkage Cluster analysis (SLCA) was used to group accessions. Morphometric data were subjected to descriptive statistics.
3. Results
3.1. Analysis of variance
The combined analysis of variance revealed significant differences (p<0.05 and <0.01) among the accessions and environments for all the measured traits (Table 3). Coefficients of variation were low (<21%) for all measured traits except seed yield per plant (SYD) with 43.8%, pod weight per plant (PWT) with 42.2% and number of pods per peduncle (NPP) with 30.8%.
3.2. Mean Performance of the Accessions
The mean performance among accessions (Table 4) for seed yield per plant was highest in TPt-31 (99.8 g) and lowest in TPt-7 (16.7 g) with an average seed yield of 61.6 g. The weight of hundred seeds ranged from 14.1 g in TPt-7 to 37.6 g in TPt-12 with an overall mean of 31.0 g. Days to 50% flowering ranged from 70 days for TPt- 126 to 102 days for TPt-7 with an average of 74.6 days. TPt-22 recorded maturity earliest at 83.7 days while TPt-7 recorded an extended maturity period of 140.8 Days after Sowing (DAS) with an average maturity period of 93.7 days. TPt-7 (5.33 cm) and TPt-153 (22.55 cm) had the shortest and longest pods, respectively with an average pod length of 19.53 cm. The number of pods per plant ranged from 13.6 in TPt-33 to 48.0 in TPt-7 with an average of 25.6. Comparatively, number of seeds per pod varied between 5.33 (TPt-7) and 14.56 (TPt-53). Weight of pods per plant varied from 47.09 in TPt-7 to 197.5 g in TPt-31.
3.3. Correlation Analysis
The genotypic and phenotypic correlation between the agro-morphological traits studied is presented in Table 5. SYD recorded positive significant phenotypic and genotypic correlation between with HSW (rp = 0.45**, rg = 0.52**); PWT (rp = 0.95**, rg = 0.99**); NLP (rp = 0.68**, rg = 0.76**); NSP (rp = 0.69**, rg = 0.76**), PL (rp = 0.52**, rg = 0.57**); PW (rp = 0.41**, rg = 0.45**); SL (rp = 0.48**, rg = 0.55**); SW (rp = 0.45**, rg = 0.49**) and ST (rp = 0.47**, rg = 0.52**), however, its correlation with D50P, DFF, D50F, DPM, D50M and NPd was negative but significant. HSW had a significant positive phenotypic and genotypic relationship with the three seed metric traits; SL (rp = 0.91**, rg = 0.99**); SW (rp = 0.88**, rg = 0.95**) and ST (rp = 0.81**, rg = 0.78**). There was also a positive significant association observed between PWT and NLP (rp = 0.70**, rg = 0.79**) as well as NSP ((rp = 0.71**, rg = 0.81**) at both phenotypic and genotypic levels. NPP had a significant positive phenotypic association with PWT (rp = 0.14**), however it recorded a negative and non-significant genotypic relationship (rg = -0.03). At both phenotypic and genotypic levels, the correlation between SYD and NPP was positive but non-significant (rp = 0.17, rg = 0.11). D50F was significant and positively correlated with D50M at (rp = 0.95**, rg = 1.00**). Significant but negative correlations were observed for D50F with SYD (rp = -0.58**, rg = -0.62**) at phenotypic and genotypic levels.
3.4. Principal Component Analysis
The Principal Component Analysis (PCA) based on agro-morphological traits revealed that the first three PCs accounted for 93% of the total observed variation among accessions studied (Table 6). The first PC contributed the highest (76%) followed by the second PC with 17%, while the third PC accounted for 5% of the total variations observed. The PC1 exhibited factor loadings with positive signs for days to 50% peduncle initiation, days to first flowering, days to 50% flowering, days to first pod maturity, days to 50% maturity, number of pods per peduncle, number of pods per plant while all the factors except for pod length, pod width, seed length, seed width and seed thickness registered positive factor loadings towards PC2. For PC3, seed yield per plant, 100 seed weight, pod weight per plant, number of pods per peduncle, number of pods per plant, seed length, seed width and seed thickness contributed factor loadings in positive signs.
3.5. Hierarchical clustering of winged bean accessions
Clustering based on the agro-morphological traits showed that the 30 accessions of winged bean were grouped into three major clusters (Figure 1). The first cluster consists of a singleton, TPt-7 that is characterized by the longest duration to flowering and extended days to maturity. In addition, TPt-7 is had the highest number of pods per peduncle and the number of pods per plant. Its pods and seeds were smaller than the other twenty-nine accessions, therefore, had the lowest seed yield. It is of an unknown origin. Group II consists of eight accessions (TPt-10, TPt-12, TPt-31, TPt-22, TPt-26, TPt-2, TPt-18 and TPt-48). These accessions were characterized by early flowering and consequently, early maturity, a high number of pods per plant, high pod weight per plant and seed yield per plant. Cluster III consisted of twenty- one accessions with a mid-number of days to flowering and long pods.
The clustering pattern from the mean of seed morphometric traits revealed three major clusters (Figure 2). The first cluster had a singleton (TPt-7), and it is the accession with the least seed length, seed width, seed perimeter and seed area thereby making it have the smallest seed size. Seven accessions (TPt-12, TPt-22, TPt-6, TPt-10, TPt-153, TPt-2 and TPt-32) were grouped in the second cluster. The accessions in this group had bigger seed sizes. The third cluster comprised of twenty-two accessions (TPt-5, TPt-11 and TPt-43, TPt-48, TPt-30, TPt-26, TPt-21, TPt-14, TPt-51, TPt- 4, TPt-18, TPt-17, TPt-154, TPt-16, TPt-33, TPt-19, TPt-53, TPt-126, TPt-9, TPt-31, TPt-3 and TPt-15) with moderate seed size.
3.6. Correlation coefficient of seed morphometric traits
The higher the correlation coefficient, the stronger the relationship between the seed morphometric traits. The correlation values ranged from -1 to 1. Correlation values towards the right indicates a positive association while towards the left direction indicates a negative association. A strong positive correlation was seen between seed length and seed area (0.99); seed width and seed area (0.98); seed perimeter and seed area (0.99). The positive correlation observed between seed circularity index and seed area (0.50) was moderate, however the relationship between seed formfactor and seed area (0.38); compactness and area (0.39) was positive but weak. A negative but strong correlation was observed between seed hue and seed area (-0.65); seed hue and seed width (-0.72); seed hue and seed length (-0.64); seed perimeter and seed hue (-0.66). Seed rectangularity and seed area (-0.54); seed eccentricity index and seed area (-0.42) were moderately correlated (Figure 3).
3.7. Descriptive statistics of seed morphometric traits from the multispectral imaging system
The phenotypic values for seed morphometric traits indicated substantial phenotypic variation in the winged bean germplasm (Table 7). Averagely, TPt-10 had the highest seed length and seed width of 9.63 mm and 8.53 mm respectively. TPt-7 recorded the least seed length and seed width of 6.18 mm and 5.18 mm respectively. Seed size based on seed length, seed width, seed area and seed perimeter were highest in TPt-10, TPt-12, TPt-22 and TPt-32 and least in TPt-7. In the contrary, the range of other seed shape characteristics among the accessions did not show a regular trend with other seed metric values, like seed eccentricity index, seed eccentricity and seed rectangularity were highest in TPt-7 but least in TPt-126. TPt-7 had the highest value in the seed shape characteristics; however, had the least value in seed compactness while TPt-126 had the lowest value. The seed circularity index also showed a regular trend with the seed area, seed length, seed width and seed perimeter as TPt-7 had the lowest value while TPt-154, TPt- 126, TPt-31, TPt-14 and TPt-4 had the highest value.
4. Discussion
The significant variations observed among the accessions for seed yield per plant and all other agro-morphological traits revealed adequate variation existed among the accessions. This implied that the accessions expressed variation for each trait, which could enable effective selection among the accessions. Additionally, the observed variation among the accessions could be used in possible combination in crop improvement programme. Yulianah et al.; Wong et al. [10,13] reported similar results in their study on winged bean accessions characterized based on their phenotypic traits. The observed significant mean squares for the environment for all traits suggest the distinctiveness of the environments in discriminating among the accessions. It however, still suggests the need for more evaluation in different environments to ascertain the genotypic integrity of the accessions. The non-significant differences observed for the interaction between the accessions and the environments implied that the performance of the winged bean accessions was consistent across the study environments.
In this study, higher genotypic correlation coefficients were observed than phenotypic correlation. This denotes that trait expressions are largely influenced by genes. The significant and positive phenotypic and genotypic correlation observed between seed yield per plant and hundred seed weight, pod weight per plant, number of locules per pod and number of seed per pod implied that these traits could be substituted for seed yield during selection in crop improvement. In addition, the correlation recorded between these desirable traits implied that the improvement of one trait simultaneously improves the other trait [26,27]. These results agrees with the findings of previous authors on the association between seed yield per plant and hundred seed weight in winged bean [26] and AYB [27]. In addition, seed yield per plant and pod weight per plant in AYB [28]; seed yield per plant and number of seed per pod in common bean [29]; days to flowering and days to maturity in winged bean [26]. The positive phenotypic and genotypic significant correlations among the yield-related traits suggested that these traits could be considered for selection to improve seed yield in winged bean. The non-significant correlation observed between number of pods per plant and seed yield per plant, indicates that the seed yield is independent of the number of pods produced by the plant.
All the 17 agro-morphological traits were found to be relevant in discriminating the winged bean accessions based on the contribution of each of the measured traits to the most informative principal components. This implied that the traits with high coefficients in the three components could be used for selection or characterization of other winged bean collections.
To assess the existing diversity among the winged bean accessions, the dendrogram generated from cluster analysis classified the 30 accessions into three distinct groups. The classification was based on their similarity for one or more agro-morphological traits studied irrespective of their geographical origin. TPt-7 was placed in cluster I and characterized by longest duration to flowering, longest days to maturity, highest number of pods per peduncle, and number of pods per plant. This accession also had the lowest seed yield. This implied that TPt-7 possesses unique characters that could be harnessed in crop improvement programme. Accessions in cluster II possess earliness to flowering and maturity, high number of pods per plant, high pod and seed yield while accessions in cluster III have early to mid-number of days to flowering with high number of pods. This result suggests that accessions were grouped based on their distinct features and could be a reliable source for hybridization when improving for reduced days to flowering and high seed yield in winged bean. This further suggests that heterosis could be exploited through selection and hybridization of distinct accessions belonging to different groups.
A positive correlation was observed between seed length and seed area; seed width and seed area; seed perimeter and seed area which implies that an increase in one trait simultaneously improves the other. A negative association was recorded between seed hue and seed area; seed eccentricity and seed area, seed rectangularity and seed area, seed eccentricity index and seed area which denotes that if one trait increases, the other trait decreases. This result agrees with Daniel et al., and Adewale et al. [24,25] who reported similar findings for AYB and maize seeds respectively. Adewale et al. [25] recommended the use of six seed traits including seed length and seed width as unique parameters of AYB based on the wide variability between them and the use of the four seed shape indices which includes eccentricity index as complement for seed shape description in AYB.
The dendrogram generated from the seed morphometry showed that the accessions clustered based on similarities between one or more seed morphometric traits. This implied that the classification formed a basis for grouping of parental lines based on seed morphometric measurements. This constitutes background data for future evaluation of crosses involving these parents. The grouping pattern in the agro-morphological and seed morphometric clusters showed the solitary clustering of TPt-7, which is due to its distinct features. This implied that TPt-7 might belong to the species scandens as reported by Yang et al. [30]. They reported a successful hybridization between Psophocarpus tetragonolobus and Psophocarpus scandens. Accessions (TPt-10, TPt-12, TPt-22, TPt-2); (TPt-33, TPt-126, TPt-21, TPt-17, TPt-9, TPt-11, TPt- 154, TPt-19, TPt-4, TPt-30, TPt-51, TPt-53, TPt-14, TPt-15, TPt-3, TPt-43, TPt-16 and TPt-5) were grouped in the same clusters in the two dendrograms generated from this study. This implied that the accessions were closely related, and the differences observed could be because of environmental influence.
5. Conclusions
This study suggests the availability of ample genetic variation in the winged bean germplasm collection at GRC- IITA, Ibadan, Nigeria. The agro-morphological analysis clearly reflected the existence of genetic variation. The positive and significant associations recorded among agro-morphological traits indicated the possibility of simultaneous traits improvement. Accessions characterized by high seed yield per plant, high pod weight per plant, early flowering and maturity days are potential sources of gene diversity for developing desirable varieties. TPt-31 was identified as the accession with the highest seed yield, early flowering time and maturity period. In contrast, TPt-7 had the highest number of pods per peduncle and pods per plant. Its pods and seed sizes were small so had the lowest seed yield per plant and lowest pod weight per plant with a long flowering duration and an extended maturity period. The MSI system is a fast and reliable method for the characterization of winged bean accessions. The genetic diversity revealed in this study within the winged bean germplasm will serve as invaluable resources to facilitate winged bean breeding in the nearest future.
Author Contributions
“Conceptualization, A.B., E.I. and O.O.; Data curation, A.B. and U.E.; Formal analysis, A.B. and U.E.; Funding acquisition, M.A.; Investigation, A.B.; Methodology, A.B.; Project administration, M.A.; Supervision, M.A., E.I., R.P. and O.O.; Writing – original draft, A.B.; Writing – review & editing, A.B., M.A., E.I., U.E., R.P. and O.O. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by the Global Crop Diversity Trust and Consortium of International Agricultural Research Centers (CGIAR) through GRC-IITA.
Acknowledgments
The authors express their gratitude to seed bank staff at the GRC-IITA, Ibadan and the cassava breeding unit, IITA, Ubiaja for their technical assistance.
Conflicts of interest
The authors disclose that there are no conflicts of interests. “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
Consent for publication
All authors have read and agreed to the publication of this manuscript.
References
- Ojuederie, O.B.; Nkang, N.A.; Odesola, K.A.; Igwe, D.O. Genetic diversity assessment of winged bean (Psophorcarpus tetragonolobus) accessions revealed by Inter-Simple Sequence Repeat (ISSR) markers. J. Plant Biol. Crop Res. 2020, 3, 1014.
- Sriwichai S, Monkham T, Sanitchon J, Jogloy S, Chankaew S (2021) Dual-Purpose of the Winged Bean (Psophocarpus tetragonolobus (L.) DC.), the Neglected Tropical Legume Based on Pod and Tuber Yields. Plants. 2021, 10, 1746. [CrossRef]
- Tanzi, A.S.; Ho, W.K.; Massawe, F.; Mayes, S. Development and interaction between plant architecture and yield-related traits in winged bean (Psophocarpus tetragonolobus (L.) DC.). Euphytica 2019, 215, 1-14. https://rdcu.be/c4ca4. [CrossRef]
- Lepcha, P.; Egan, A.N.; Doyle, J.J.; Sathyanarayana, N. A review on current status and future prospects of winged bean (Psophocarpus tetragonolobus) in tropical agriculture. Plant Foods Hum. Nutr. 2017, 72, 225–235. [CrossRef]
- Wan, M.W.A.; Mohtar, Hamid, A.A.; Abd-Aziz, S.; Muhamad, S.K.; Saari, N. Preparation of bioactive peptides with high angiotensin converting enzyme inhibitory activity from winged bean [Psophocarpus tetragonolobus (L.) DC.] seed. JFST. 2014, 51, 3658-3668. [CrossRef]
- Mohanty, C.S.; Verma, S.; Singh, V.; Khan, S.; Gaur, P.; Gupta, P.; Niranjan, A.; Rana, T.S. Characterization of winged bean (Psophocarpus tetragonolobus (L.) DC.) based on molecular, chemical and physiological parameters. Am. J. Mol. Biol. 2013, 3, 187. [CrossRef]
- Amoo, I.A.; Adebayo, O.T.; Oyeleye, A.O. Chemical evaluation of Winged beans (Psophocarpus tetragonolobus), Pitanga cherries (Eugenia uniflora) and Orchid fruit (Orchid fruit myristica). Afr. J. Food Agric. Nutr. Dev. 2006, 6, 1-12. [CrossRef]
- Laosatit, K.; Amkul, K.; Chankaew, S.; Somta, P. Molecular Genetic Diversity of Winged Bean Gene Pool in Thailand Assessed by SSR Markers. Hortic. Plant J. 2022, 10, 9-10. [CrossRef]
- Laido, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic Diversity and Population Structure of Tetraploid Wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree Data. Plos One. 2013, 8, 1-17. [CrossRef]
- Yulianah, I.; Waluyo, B.; Ashari, S.; Kuswanto. Variation in morphological traits of a selection of Indonesian winged bean accessions (Psophocarpus tetragonolobus) and its analysis to assess genetic diversity among accessions. Biodiversitas 2020, 21, 2991-3000. [CrossRef]
- Lawal, B.A.; Azeez, M.A.; Egedegbe, G Raji IA, Omogoye A.M.; Akintola, E.K. Screening Winged Bean (Psophocarpus tetragonolobus (L.) DC.) Accessions Using Agronomic Characters. Asian J. Soil Sci. Plant Nutri. 2019, 4, 1-10. [CrossRef]
- Kant, A.; Nandan, R Performance and variability evaluation in some genotypes of winged bean [Psophocarpus tetragonolobus (L.) DC.]. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2104-2108. [CrossRef]
- Wong, Q.N.; Massawe, F.; Mayes, S. Improving winged bean (Psophocarpus tetragonolobus) productivity: An analysis of the determinants of productivity. In XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes. 2014, 1102, 83-88. [CrossRef]
- Hansen, M.A.E.; Hay, F.R.; Carstensen, J.M. A virtual seed file: the use of multispectral image analysis in the management of genebank seed accessions. Plant Genet. Resour. 2016, 14, 238-241. [CrossRef]
- Bianco, M.L.; Grillo, O.; Cremonini, R.; Sarigu, M.; Venora, V. Characterisation of Italian bean landraces (‘Phaseolus vulgaris’ L.) using seed image analysis and texture descriptors. Aust. J. Crop Sci. 2015, 9, 1022. http://www.cropj.com/search.html?q=Characterisation%20of%20Italian%20bean%20landraces%20(%E2%80%98Phaseolus%20vulgaris%E2%80%99%20L.)%20using%20seed%20image%20analysis%20and%20texture%20descriptors.
- Venora, G.; Grillo, O.; Ravalli, C.; Cremonini, R. ‘’Identification of Italian landraces of bean (Phaseolus vulgaris L.) using an image analysis system.’’ Sci. Hortic. 2009, 121, 410-418. [CrossRef]
- Kılıç, K.; Boyacı, I.H.; Köksel, H.; Küsmenog˘lu, I. A classification system for beans using computer vision system and artificial neural networks. J. Food Eng. 2007, 78, 897-904. [CrossRef]
- Geetha, V.V.; Balamurugan, P.; Bhaskaran, M. Characterization of mustard genotypes through image analysis. Res. J. Seed Sci. 2011, 4, 192–198. https://scialert.net/abstract/?doi=rjss.2011.192.198.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1-48. [CrossRef]
- Ward, J.H.; Jr. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [CrossRef]
- Raj, P.; Rumit, P.; Dinesh, P. Variability: Genetic Variability Analysis for Plant Breeding Research. R package version 0.1.0. 2020. https://cran.r-project.org/package=variability.
- Galletti, P.A.; Carvalho, M.E.; Hirai, W.Y.; Brancaglioni, V.A.; Arthur, V.; Barboza da Silva, C. Integrating optical imaging tools for rapid and non-invasive characterization of seed quality: Tomato (Solanum lycopersicum L.) and carrot (Daucus carota L.) as study cases. Front. Plant Sci. 2020, 11, 577851. [CrossRef]
- Cervantes, E.; Martín, J.; Saadaoui, E. Updated methods for seed shape analysis. Scientifica 2016, 5691825. [CrossRef]
- Daniel, I.O.; Adeboye, K.A.; Oduwaye, O.O.; Porbeni, J. Digital seed morpho-metric characterization of tropical maize. Int. J. Plant Breed. 2012, 6, 245–251. [CrossRef]
- Adewale, B.D.; Kehinde, O.B.; Aremu, C.O.; Popoola, J.O.; Dumet, D.J. Seed metrics for genetic and shape determinations in African yam bean [Fabaceae] (Sphenostylis stenocarpa Hochst. Ex. A. Rich.) harms. Afri. J. Plant Sci. 2010, 4, 107–115. https://www.researchgate.net/publication/266184996_Seed_metrics_for_genetic_and_shape_determinations_in_African_yam_bean_Fabaceae_Sphenostylis_stenocarpa_Hochst_Ex_A_Rich_harms.
- Yadav, R. K. Co-heritability and correlation of yield and associated characters in winged bean (Psophocarpus tetragonolobus L. DC.). JFL, 2021, 34(3), 225-227.
- Alake, C.O.; Porbeni, B.J. Genetic variation and intraspecific diversity for agro-morphological and nutritional traits in African yam bean. J. Crop Improv. 2020, 34, 139-163. [CrossRef]
- Olomitutu, O.E.; Abe, A.; Oyatomi, O.A.; Paliwal, R.; Abberton, M.T. Assessing Intraspecific Variability and Diversity in African Yam Bean Landraces Using Agronomic Traits. Agronomy 2022, 12, 884. https://www.mdpi.com/2073-4395/12/4/884#B31-agronomy-12-00884. [CrossRef]
- Nadeem, M.A.; Karaköy, T.; Yeken, M.Z.; Habyarimana, E.; Hatipoğlu, R.; Çiftçi, V.; Nawaz, M.A.; Sönmez, F.; Shahid, M.Q.; Yang, S.H.; Chung, G.; Baloch, F.S. Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes. Agronomy 2020, 10, 272. [CrossRef]
- Yang, S.Y.; Grall, A.; Chapman, M.A. Origin and diversification of winged bean (Psophocarpus tetragonolobus (L. DC.), a multipurpose underutilized legume. Am. J. Bot. 2018, 105, 888-897. [CrossRef]
Figure 1.
Cluster of the 30 winged bean accessions derived from 17 agro-morphological data of traits studied.
Figure 1.
Cluster of the 30 winged bean accessions derived from 17 agro-morphological data of traits studied.
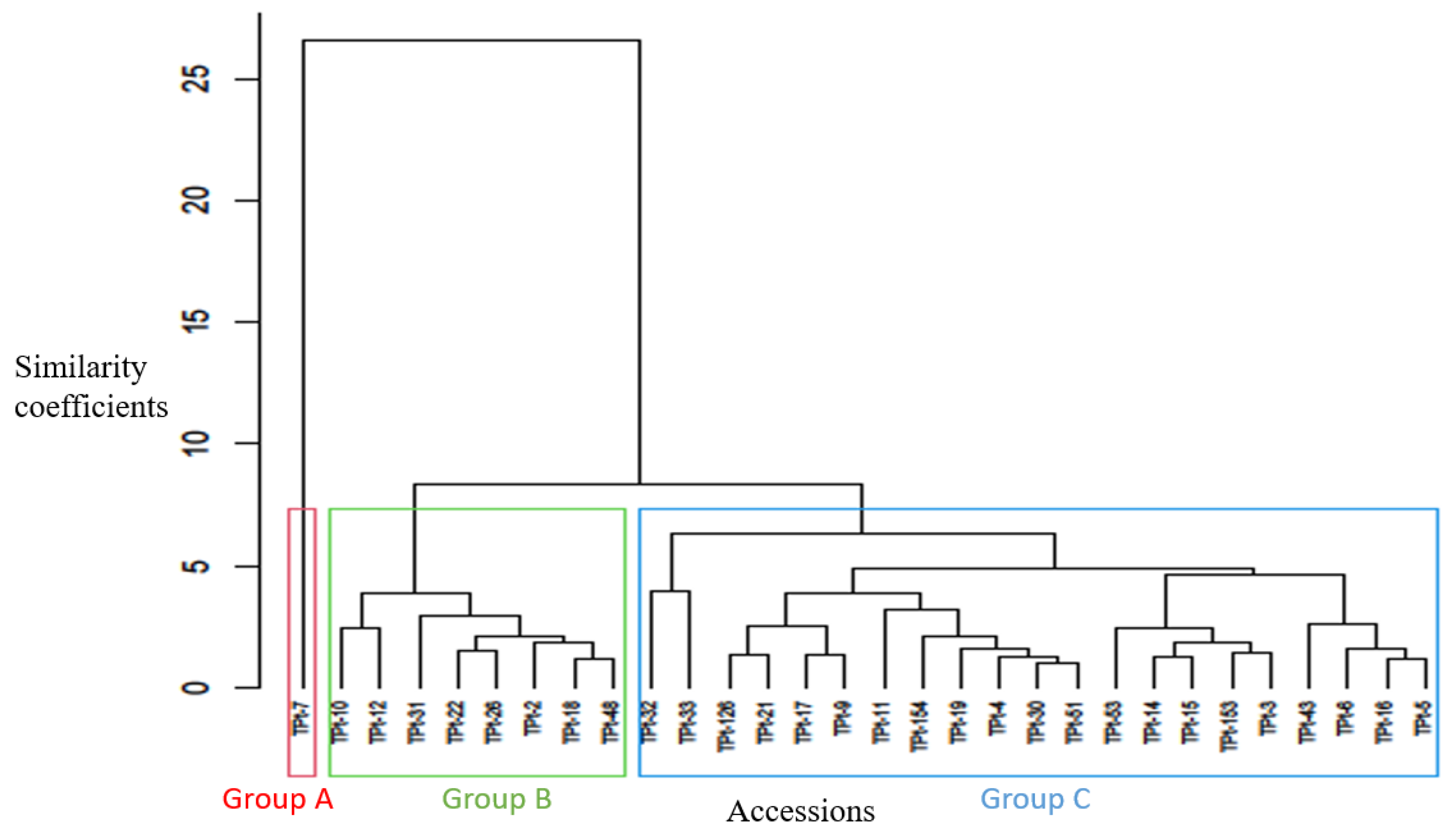
Figure 2.
Dendrogram based on the 11 seed morphometric traits generated from the Videometer MSI system.
Figure 2.
Dendrogram based on the 11 seed morphometric traits generated from the Videometer MSI system.
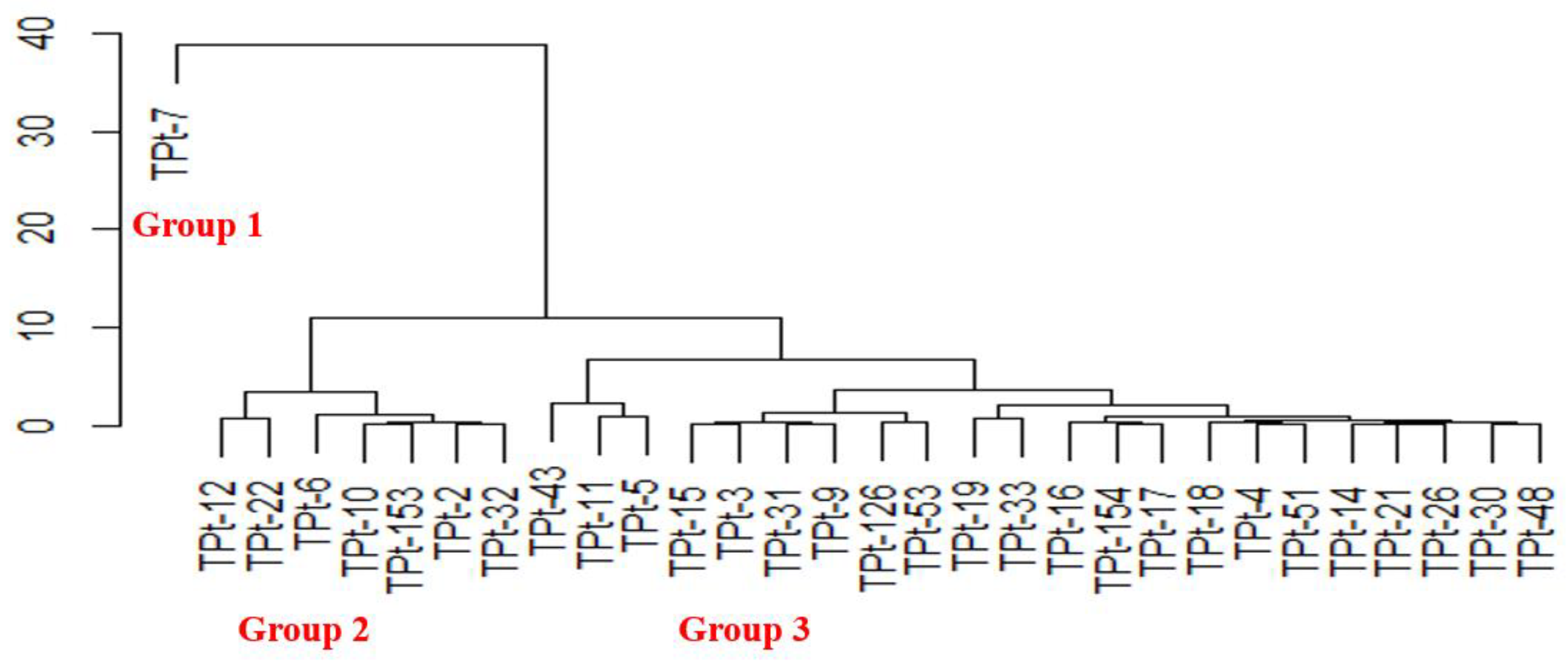
Figure 3.
Correlation coefficients among seed morphometric traits of the winged bean accessions. LNTH-seed length, WDTH-seed width, HUE- seed hue, PER- seed perimeter, FORFACT- seed form factor, CMPTNESS-seed compactness, ECC-seed eccentricity, RECTY-seed rectangularity, CI-seed circularity index, EI- seed eccentricity index.
Figure 3.
Correlation coefficients among seed morphometric traits of the winged bean accessions. LNTH-seed length, WDTH-seed width, HUE- seed hue, PER- seed perimeter, FORFACT- seed form factor, CMPTNESS-seed compactness, ECC-seed eccentricity, RECTY-seed rectangularity, CI-seed circularity index, EI- seed eccentricity index.
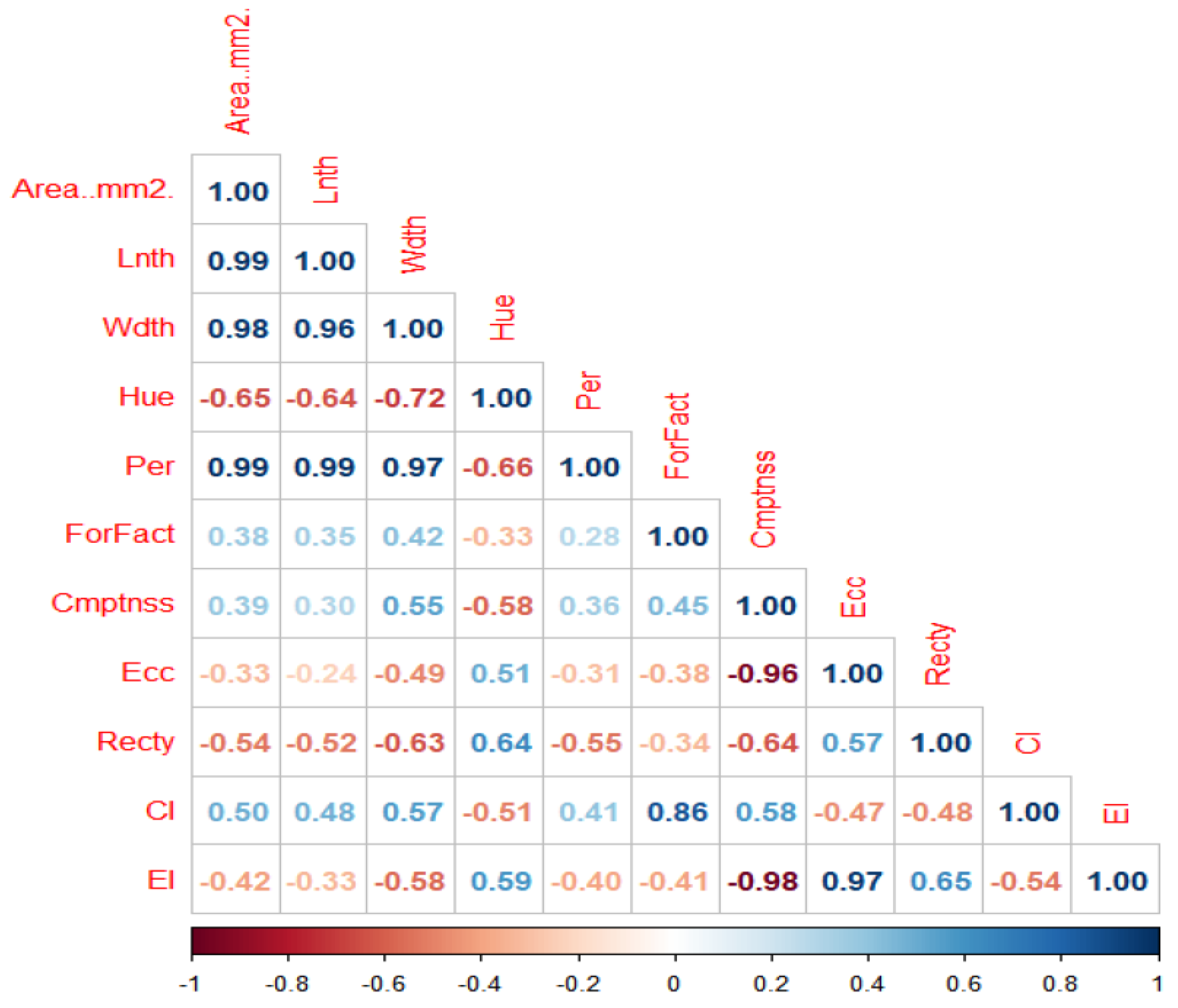

Table 1.
List of winged bean accessions used for the study and their countries of origin.
| S/N | Accession number | Sources |
|---|---|---|
| 1 | TPt-2 | Nigeria |
| 2 | TPt-3 | Nigeria |
| 3 | TPt-4 | Trinidad and Tobago |
| 4 | TPt-5 | Nigeria |
| 5 | TPt-6 | Nigeria |
| 6 | TPt-7 | Unknown |
| 7 | TPt-9 | Unknown |
| 8 | TPt-10 | Papua New Guinea |
| 9 | TPt-11 | Costa Rica |
| 10 | TPt-12 | Liberia |
| 11 | TPt-14 | Indonesia |
| 12 | TPt-15 | Indonesia |
| 13 | TPt-16 | Indonesia |
| 14 | TPt-17 | Indonesia |
| 15 | TPt-18 | Unknown |
| 16 | TPt-19 | Nigeria |
| 17 | TPt-21 | Papua New Guinea |
| 18 | TPt-22 | Papua New Guinea |
| 19 | TPt-26 | Nigeria |
| 20 | TPt-30 | Unknown |
| 21 | TPt-31 | Indonesia |
| 22 | TPt-32 | Unknown |
| 23 | TPt-33 | Unknown |
| 24 | TPt-43 | Unknown |
| 25 | TPt-48 | Unknown |
| 26 | TPt-51 | Bangladesh |
| 27 | TPt-53 | Bangladesh |
| 28 | TPt-126 | Unknown |
| 29 | TPt-153 | Unknown |
| 30 | TPt-154 | Unknown |
Table 2.
The descriptions of the seed morphometric features measured.
| S/N | Parameters | Descriptions/mode of estimation |
|---|---|---|
| 1 | Seed length (mm) | Longest dimension on the seed taken at base of the seed. |
| 2 | Seed width (mm) | Cross section taken at right angle to the length of the seed. |
| 3 | Seed area (mm2) | Product of seed length and seed width. |
| 4 | Seed Perimeter (mm) | the number of pixels in the boundary of the object. |
| 5 | Seed Rectangularity (mm) | the ratio of the object to the area of the minimum bounding rectangle. |
| 6 | Seed hue | Color appearance parameter. |
| 7 | Seed compactness | the ratio of the area of an object to the area of a circle with the same perimeter. |
| 8 | Seed circularity index (CI) | This is how alike a circle the shape of the seed is. CI = 4πArea/(Perimeter)2 |
| 9 | Seed eccentricity index-ratio (EI) | the measure of aspect ratio. EI = SL/SW |
mm-millimeter, mm2-square millimeters.
Table 3.
Combined mean squares for yield traits of thirty winged bean accessions evaluated across the three environments.
Table 3.
Combined mean squares for yield traits of thirty winged bean accessions evaluated across the three environments.
| SOV | Env | Accn | Rep (Env) | Accn*Env | Error | CV (%) |
|---|---|---|---|---|---|---|
| DF | 2 | 29 | 6 | 58 | 174.00 | |
| SYD (g) | 27165.29** | 2676.63** | 6658.00** | 612.77 | 521.04 | 43.8 |
| HSW (g) | 301.32** | 102.06** | 24.07 | 28.48* | 23.42 | 15.5 |
| PWT | 112784.02** | 9480.81** | 31057.73** | 2339.60** | 2438.09 | 42.4 |
| NLP | 52.34** | 22.57** | 7.05** | 2.60 | 1.98 | 9.0 |
| NSP | 1.11** | 0.58** | 0.13** | 0.05 | 0.03 | 12.5 |
| D50P | 8017.31** | 150.30** | 24.68 | 27.38* | 18.40 | 7.5 |
| DFF | 5987.73** | 223.16** | 3.24 | 20.34 | 19.93 | 6.7 |
| D50F | 8782.07** | 302.19** | 43.16 | 18.35 | 23.84 | 6.6 |
| DPM | 11124.58** | 815.00** | 154.82* | 86.44* | 60.84 | 8.5 |
| D50M | 13027.07** | 867.75** | 389.78** | 144.00** | 72.26 | 8.6 |
| NPd | 1.74** | 3.36** | 0.42 | 0.44** | 0.20 | 30.8 |
| NPP | 2164.70** | 334.32** | 845.66** | 69.88 | 75.81 | 39.3 |
| PL (cm) | 5.67* | 78.63** | 4.19* | 1.51 | 1.45 | 6.4 |
| PW (mm) | 47.58** | 24.24** | 14.14** | 1.35 | 1.83 | 9.5 |
| SL (mm) | 3.70** | 3.36** | 0.11 | 0.159* | 0.11 | 3.7 |
| SW (mm) | 8.02** | 3.19** | 0.11 | 0.07 | 0.07 | 3.3 |
| ST (mm) | 9.47** | 1.66** | 0.26* | 0.12 | 0.09 | 4.5 |
*, ** significant at 5% and 1% levels respectively. SOV-source of variation, Env-environment, Accn-accession, Rep-replicates, CV-co-efficient of variation, DF-degree of freedom, g-grams, cm-centimeter, mm-millimeter, SYD-seed yield per plot, HSW-100 seed weight, PWT-pod weight per plant, NLP-number of locules per pod , NSP-number of seed per pod, D50P- days to fifty percent peduncle initiation, DFF-days to first flowering, D50F-days to fifty percent flowering, DPM-days to first pod maturity, D50M-days to fifty percent maturity, NPd-number of pod per peduncle, NPP-number of pod per plant, PL-pod length, PW-pod width, SL- seed length, SW-seed width, ST-seed thickness.
Table 4.
Mean performance of the 30 winged bean accessions with respect to the traits evaluated across three environments.
Table 4.
Mean performance of the 30 winged bean accessions with respect to the traits evaluated across three environments.
| ACCN | SYD | HSW | PWT | NLP | NSP | D50P | DFF | D50F | DPM | D50M | NPd | NPP | PL(cm) | PW(mm) | SL(mm) | SW(mm) | ST(mm) |
| TPt-10 | 83.22 | 35.52 | 173.75 | 12.44 | 12.33 | 60.56 | 69.11 | 77.22 | 95.56 | 105.33 | 1.44 | 32.44 | 19.66 | 16.06 | 8.98 | 8.14 | 7.01 |
| TPt-11 | 76.96 | 28.02 | 139.30 | 10.56 | 10.33 | 57.33 | 65.67 | 70.56 | 94.22 | 107.00 | 1.56 | 31.11 | 17.32 | 15.75 | 8.62 | 7.91 | 6.83 |
| TPt-12 | 72.47 | 37.62 | 157.77 | 11.00 | 10.89 | 57.33 | 65.44 | 72.33 | 95.11 | 106.00 | 1.56 | 30.00 | 18.38 | 17.15 | 9.36 | 8.32 | 6.87 |
| TPt-126 | 49.82 | 31.41 | 123.86 | 11.78 | 11.33 | 53.22 | 58.00 | 69.67 | 87.78 | 98.78 | 1.44 | 21.33 | 20.50 | 15.66 | 8.81 | 8.16 | 6.86 |
| TPt-14 | 65.60 | 31.19 | 141.08 | 12.89 | 12.67 | 57.89 | 66.22 | 72.22 | 91.78 | 102.22 | 1.11 | 23.00 | 21.01 | 17.31 | 8.88 | 8.16 | 6.89 |
| TPt-15 | 70.55 | 29.54 | 146.30 | 13.22 | 12.78 | 59.22 | 67.33 | 74.89 | 96.67 | 107.67 | 1.22 | 24.78 | 21.32 | 16.96 | 8.90 | 8.28 | 6.96 |
| TPt-153 | 59.73 | 32.09 | 137.15 | 12.89 | 12.33 | 56.33 | 64.00 | 73.78 | 90.22 | 102.22 | 1.56 | 21.67 | 22.55 | 16.21 | 9.03 | 8.18 | 6.78 |
| TPt-154 | 66.47 | 31.82 | 154.19 | 11.33 | 11.11 | 56.78 | 62.22 | 70.33 | 92.00 | 103.00 | 1.22 | 27.67 | 17.96 | 15.78 | 8.66 | 8.19 | 7.24 |
| TPt-16 | 53.09 | 30.04 | 122.91 | 11.56 | 11.22 | 60.89 | 68.33 | 74.89 | 95.33 | 107.56 | 1.44 | 25.00 | 19.32 | 16.29 | 8.89 | 7.96 | 6.64 |
| TPt-17 | 55.90 | 27.37 | 120.83 | 11.78 | 11.67 | 55.44 | 64.67 | 73.78 | 91.56 | 102.11 | 1.11 | 23.44 | 18.80 | 14.88 | 8.86 | 8.21 | 6.98 |
| TPt-18 | 71.88 | 31.86 | 157.12 | 12.89 | 12.56 | 55.11 | 63.78 | 72.11 | 90.44 | 101.56 | 1.22 | 26.44 | 19.53 | 16.82 | 9.00 | 8.03 | 6.78 |
| TPt-19 | 69.07 | 31.24 | 152.14 | 12.11 | 11.89 | 56.89 | 61.89 | 75.00 | 88.67 | 102.33 | 1.22 | 26.00 | 20.50 | 16.37 | 8.78 | 8.23 | 7.04 |
| TPt-2 | 77.79 | 33.03 | 190.41 | 12.22 | 12.33 | 55.44 | 63.78 | 74.33 | 94.67 | 105.56 | 1.67 | 31.11 | 21.22 | 16.35 | 9.14 | 8.30 | 6.72 |
| TPt-21 | 50.45 | 30.60 | 105.53 | 12.22 | 12.11 | 55.33 | 60.44 | 70.67 | 89.22 | 99.67 | 1.44 | 18.78 | 20.19 | 15.79 | 8.84 | 8.25 | 7.09 |
| TPt-22 | 76.77 | 31.34 | 172.01 | 13.00 | 12.78 | 57.11 | 62.33 | 70.56 | 83.67 | 97.00 | 1.44 | 27.56 | 20.92 | 15.89 | 9.25 | 8.53 | 7.09 |
| TPt-26 | 70.18 | 34.47 | 174.99 | 13.00 | 12.67 | 57.56 | 63.11 | 72.33 | 89.33 | 101.56 | 1.33 | 28.33 | 21.59 | 16.67 | 8.97 | 8.29 | 6.90 |
| TPt-3 | 60.97 | 29.64 | 134.73 | 13.33 | 13.11 | 58.33 | 64.67 | 74.00 | 94.44 | 105.78 | 1.22 | 21.89 | 21.48 | 15.90 | 8.79 | 8.07 | 6.65 |
| TPt-30 | 61.89 | 31.73 | 131.66 | 11.67 | 11.44 | 56.33 | 63.44 | 73.22 | 88.78 | 100.78 | 1.22 | 23.89 | 19.16 | 15.89 | 8.83 | 8.13 | 6.83 |
| TPt-31 | 99.76 | 30.90 | 197.50 | 13.44 | 13.22 | 55.00 | 63.33 | 72.00 | 86.11 | 98.33 | 1.56 | 34.22 | 20.87 | 15.45 | 8.70 | 8.16 | 6.86 |
| TPt-32 | 27.65 | 33.68 | 81.89 | 9.11 | 8.89 | 56.78 | 64.78 | 72.89 | 92.33 | 104.22 | 1.11 | 20.22 | 18.27 | 15.94 | 9.18 | 8.57 | 7.26 |
| TPt-33 | 30.58 | 30.90 | 79.89 | 11.89 | 11.67 | 60.00 | 68.78 | 78.89 | 93.44 | 104.78 | 1.11 | 13.56 | 21.45 | 18.43 | 8.93 | 8.30 | 6.78 |
| TPt-4 | 63.79 | 32.43 | 136.32 | 12.11 | 11.78 | 55.78 | 65.22 | 71.78 | 93.44 | 102.44 | 1.11 | 22.78 | 20.96 | 16.74 | 9.09 | 8.10 | 6.66 |
| TPt-43 | 56.91 | 30.59 | 123.84 | 12.22 | 12.11 | 58.56 | 69.89 | 80.67 | 98.56 | 113.44 | 1.22 | 23.00 | 19.40 | 13.94 | 8.55 | 7.84 | 6.67 |
| TPt-48 | 79.19 | 30.88 | 165.51 | 12.67 | 12.67 | 54.78 | 64.78 | 72.22 | 91.78 | 104.00 | 1.67 | 28.67 | 20.78 | 16.53 | 9.16 | 8.02 | 6.88 |
| TPt-5 | 45.19 | 30.50 | 120.28 | 12.00 | 11.67 | 59.78 | 67.89 | 77.00 | 94.78 | 107.11 | 1.56 | 22.11 | 18.70 | 16.01 | 8.50 | 8.02 | 6.60 |
| TPt-51 | 65.13 | 34.14 | 144.93 | 11.89 | 11.78 | 56.67 | 63.67 | 72.11 | 90.56 | 104.00 | 1.22 | 24.56 | 20.03 | 15.84 | 9.05 | 8.14 | 6.88 |
| TPt-53 | 70.61 | 31.10 | 153.43 | 13.22 | 14.56 | 59.22 | 64.56 | 77.56 | 96.44 | 107.00 | 1.56 | 24.00 | 20.19 | 15.33 | 8.59 | 8.08 | 6.95 |
| TPt-6 | 49.56 | 32.04 | 125.89 | 11.00 | 10.78 | 57.78 | 65.44 | 74.44 | 94.67 | 107.89 | 1.56 | 23.00 | 19.69 | 15.62 | 9.00 | 8.12 | 6.67 |
| TPt-7 | 16.72 | 14.07 | 47.09 | 5.44 | 5.33 | 77.22 | 88.11 | 102.00 | 140.78 | 154.22 | 4.56 | 48.00 | 5.33 | 8.27 | 5.76 | 5.02 | 4.71 |
| TPt-9 | 49.16 | 29.53 | 108.86 | 11.89 | 11.22 | 57.11 | 64.11 | 74.67 | 88.56 | 101.56 | 1.33 | 20.33 | 18.84 | 15.49 | 8.73 | 8.09 | 6.73 |
| mean | 61.57 | 30.98 | 137.37 | 11.89 | 11.71 | 57.86 | 65.50 | 74.60 | 93.70 | 105.50 | 1.47 | 25.63 | 19.53 | 15.84 | 8.80 | 8.06 | 6.79 |
| LSD | 25.06 | 7.80 | 54.18 | 0.99 | 1.36 | 4.01 | 4.09 | 4.60 | 7.44 | 8.47 | 0.42 | 9.37 | 1.15 | 1.39 | 0.31 | 0.25 | 0.29 |
SYD-seed yield per plot, HSW-100 seed weight, PWT-pod weight per plant, NLP-number of locules per pod , NSP-number of seed per pod, D50P- days to fifty percent peduncle initiation, DFF-days to first flowering, D50F-days to fifty percent flowering, DPM-days to first pod maturity, D50M-days to fifty percent maturity, NPd-number of pod per peduncle, NPP-number of pod per plant, PL-pod length, PW-pod width, SL- seed length, SW-seed width, ST-seed thickness, LSD- least significant difference.
Table 5.
Genotypic and phenotypic correlation coefficients among 17 agro-morphological traits of 30 winged bean accessions evaluated across the three environments in Nigeria.
Table 5.
Genotypic and phenotypic correlation coefficients among 17 agro-morphological traits of 30 winged bean accessions evaluated across the three environments in Nigeria.
| Traits | SYD | HSW | PWT | NLP | NSP | D50P | DFF | D50F | DPM | D50M | NPd | NPP | PL | PW | SL | SW | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HSW | rg | 0.52* | |||||||||||||||
| rp | 0.45* | ||||||||||||||||
| PWT | rg | 0.99** | 0.60** | ||||||||||||||
| rp | 0.95** | 0.56** | |||||||||||||||
| NLP | rg | 0.76** | 0.81** | 0.79** | |||||||||||||
| rp | 0.68** | 0.62** | 0.70** | ||||||||||||||
| NSP | rg | 0.76** | 0.75** | 0.81** | 1.00** | ||||||||||||
| rp | 0.69** | 0.60** | 0.71** | 0.98** | |||||||||||||
| D50P | rg | -0.60** | -0.81** | -0.62** | -0.81** | -0.77** | |||||||||||
| rp | -0.56** | -0.8** | -0.59** | -0.72** | -0.68** | ||||||||||||
| DFF | rg | -0.56** | -0.82** | -0.64** | -0.77** | -0.75** | 1.00** | ||||||||||
| rp | -0.52* | -0.79** | -0.57** | -0.71** | -0.68** | 0.95** | |||||||||||
| D50F | rg | -0.62** | -0.86** | -0.65** | -0.72** | -0.67** | 1.00** | 0.99** | |||||||||
| rp | -0.58** | -0.81** | -0.59** | -0.69** | -0.63** | 0.94** | 0.94** | ||||||||||
| DPM | rg | -0.61** | -0.93** | -0.69** | -0.85** | -0.81** | 1.00** | 1.00** | 1.00** | ||||||||
| rp | -0.53** | -0.81** | -0.56** | -0.78** | -0.73** | 0.93** | 0.94** | 0.93** | |||||||||
| D50M | rg | -0.63** | -1.00** | -0.71** | -0.89** | -0.84** | 1.00** | 1.00** | 1.00** | 1.00** | |||||||
| rp | -0.53* | -0.81** | -0.56** | -0.79** | -0.74** | 0.94** | 0.95** | 0.95** | 0.99** | ||||||||
| NPd | rg | -0.48** | -0.94** | -0.50** | -0.84** | -0.82** | 0.94** | 0.89** | 0.95** | 1.00** | 1.00** | ||||||
| rp | -0.38* | -0.80** | -0.40* | -0.74** | -0.69** | 0.85** | 0.81** | 0.85** | 0.9** | 0.90** | |||||||
| NPP | rg | 0.11 | -0.57** | -0.03 | -0.58 | -0.55* | 0.79** | 0.71** | 0.63** | 0.77** | 0.80** | 0.91** | |||||
| rp | 0.17 | -0.55* | 0.14** | -0.51** | -0.46** | 0.57** | 0.57* | 0.53** | 0.64** | 0.66** | 0.78** | ||||||
| PL | rg | 0.57** | 0.91** | 0.63** | 0.92** | 0.90** | -0.92** | -0.86** | -0.83** | -0.94** | -0.98** | -0.93** | -0.79** | ||||
| rp | 0.52* | 0.80** | 0.58** | 0.91** | 0.87** | -0.83** | -0.81** | -0.8** | -0.88** | -0.89** | -0.86** | -0.69** | |||||
| PW | rg | 0.45** | 1.00** | 0.51** | 0.76** | 0.73** | -0.85** | -0.77** | -0.86** | -0.88** | -0.94** | -0.94** | -0.78** | 0.89** | |||
| rp | 0.41* | 0.83** | 0.46* | 0.72** | 0.67** | -0.75** | -0.72** | -0.8** | -0.82** | -0.84** | -0.85** | -0.67** | 0.87** | ||||
| SL | rg | 0.55** | 0.99** | 0.63** | 0.77** | 0.74** | -0.99** | -0.88** | -0.94** | -0.97** | -1.00** | -0.97** | -0.71** | 0.91** | 0.95** | ||
| rp | 0.48** | 0.91** | 0.55** | 0.72** | 0.68** | -0.89** | -0.84** | -0.90** | -0.90** | -0.91** | -0.89** | -0.64** | 0.89** | 0.90** | |||
| SW | rg | 0.49** | 0.95** | 0.56** | 0.78** | 0.76** | -0.98** | -0.93** | -0.93** | -1.00** | -1.00** | -1.00** | -0.80** | 0.91** | 0.91** | 0.97** | |
| rp | 0.45* | 0.88** | 0.51** | 0.74** | 0.71** | -0.89** | -0.88** | -0.9** | -0.94** | -0.95** | -0.93** | -0.70** | 0.90** | 0.88** | 0.96** | ||
| ST | rg | 0.52** | 0.78** | 0.56** | 0.73** | 0.72** | -0.97** | -0.94** | -0.94** | -0.99** | -1.00** | -0.97** | -0.73** | 0.83** | 0.84** | 0.91** | 0.97** |
| rp | 0.47** | 0.81** | 0.49** | 0.68** | 0.66** | -0.88** | -0.89** | -0.9** | -0.91** | -0.92** | -0.9** | -0.64** | 0.81** | 0.8** | 0.89** | 0.95** |
*, ** significant at 0.05, 0.01 levels of probability, respectively. SYD-seed yield per plot, HSW-100 seed weight, PWT-pod weight per plant, NLP-number of locules per pod , NSP-number of seed per pod, D50P- days to fifty percent peduncle initiation, DFF-days to first flowering, D50F-days to fifty percent flowering, DPM-days to first pod maturity, D50M-days to fifty percent maturity, NPd-number of pod per peduncle, NPP-number of pod per plant, PL-pod length, PW-pod width, SL- seed length, SW-seed width, ST-seed thickness.
Table 6.
Principal components of the 30 winged bean accessions based on agro-morphological traits.
| Traits | PC1 | PC2 | PC3 |
| Seed yield (g) | -0.16 | -0.56 | 0.06 |
| 100 seed weight (g) | -0.24 | 0.00 | 0.20 |
| Pod weight per plant (g) | -0.17 | -0.54 | 0.08 |
| Number of locules per pod | -0.24 | -0.18 | -0.49 |
| Number of seed per pod | -0.23 | -0.22 | -0.52 |
| Days to 50% peduncle initiation | 0.26 | -0.04 | -0.21 |
| Days to first flowering | 0.25 | -0.05 | -0.21 |
| Days to 50% flowering | 0.26 | -0.02 | -0.30 |
| Days to first pod maturity | 0.27 | -0.05 | -0.09 |
| Days to 50% maturity | 0.27 | -0.05 | -0.08 |
| Number of pods per peduncle | 0.26 | -0.17 | 0.06 |
| Number of pods per plant | 0.17 | -0.50 | 0.33 |
| Pod length (cm) | -0.26 | 0.01 | -0.29 |
| Pod width (mm) | -0.25 | 0.10 | -0.08 |
| Seed length (mm) | -0.26 | 0.06 | 0.12 |
| Seed width (mm) | -0.27 | 0.10 | 0.07 |
| Seed thickness(mm) Eigen values |
-0.26 3.59 |
0.07 1.42 |
0.18 0.91 |
| Percent variance (%) | 0.76 | 0.12 | 0.05 |
| Cumulative variance (%) | 0.76 | 0.88 | 0.93 |
Table 7.
Descriptive statistics of eleven seed morphometric traits of winged bean.
|
Parameters |
Mean |
Minimum |
Maximum |
Standard deviation |
CV |
|---|---|---|---|---|---|
| Seed area (mm2) | 56.4 | 25.14 | 62.75 | 6.31 | 0.11 |
| Seed circularity index | 1.18 | 1.08 | 1.19 | 0.02 | 0.02 |
| Seed compactness | 0.90 | 0.83 | 0.92 | 0.02 | 0.02 |
| Seed eccentricity | 0.42 | 0.36 | 0.52 | 0.03 | 0.07 |
| Seed eccentricity index | 1.11 | 1.08 | 1.19 | 0.02 | 0.02 |
| Seed form factor | 0.91 | 0.88 | 0.92 | 0.01 | 0.01 |
| Seed hue | 0.99 | 0.94 | 1.25 | 0.06 | 0.06 |
| Seed length (mm) | 8.94 | 6.18 | 9.63 | 0.56 | 0.06 |
| Seed perimeter (mm) | 24.4 | 17.11 | 25.97 | 1.48 | 0.06 |
| Seed rectangularity | 0.80 | 0.79 | 0.83 | 0.01 | 0.01 |
| Seed width (mm) | 8.07 | 5.18 | 8.53 | 0.57 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



