
Submitted:
17 April 2023
Posted:
17 April 2023
Read the latest preprint version here
Preprints on COVID-19 and SARS-CoV-2
Abstract
These three syndromes - long covid (LC), postural orthostatic tachycardia syndrome (POTS), and myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) - have many symptoms in common. The common denominator remains elusive. The blood brain barrier (BBB) has been a barrier not only to microbes and toxins but also to understanding pathogenetic links. There are several areas within the brain that have no BBB. These are known as circumventricular organs (CVOs) and their location relative to CNS nuclei that direct autonomic and neuroendocrine functions is provocative in the quest for pathogenesis. In addition the majority afflicted with LC and ME/CFS appear to be those with two MTHFR polymorphisms, present in over 50% of Americans. These polymorphisms elevate homocysteine. When homocysteine is combined with CVOs, the fog of POTS and its paradox are lifted. POTS may represent the intersection of LC and ME/CFS in those with the MTHFR gene (hypermethylation or 677TT). The gut microbiomes of LC and ME/CFS, deficient in butyrates, GABA, and diversity, are then linked with MTHFR genotype 677TT. Reactivation of neurotropic EBV and VZV, due to loss of surveillance by CD4+/CD8+ T cells, is seen as secondary. The oxidative stress generated by homocysteine, loss of glutathione, low fiber diet, and persistent chronic inflammation exhaust available mitochondria and, assisted by BKN and estrogen, exacerbate all the elements of these post viral fatigue syndromes.
Keywords:
homocysteine
; MTHFR
; circumventricular organ
; magnesium
; vitamin D
; CD147
INTRODUCTION
Covid-19 has unleashed a tsunami of research that has been further energized by the persistent and debilitating aftermath of Long Covid. Explaining the gender reversal from male to female preponderance is challenging and covering the range of symptoms intimidating. Although there have been a few hypotheses incorporating autoantibodies, nutrient deficiencies, reactivation of Epstein-Barr virus (EBV),…, resolution of the paradoxes remains frustratingly out of reach. Recent research has revealed that the BBB is not all inclusive and that some areas of the CNS are not so protected. These are called circumventricular organs (CVOs). Other research has revealed a close association between Covid-19 and a very common enzymatic polymorphism for methylenetetrahydrofolate reductase (MTHFR) [1,2] in the folate cycle. Most with this polymorphism are unaware of it. But it may play a large role in Long Covid and may provide unique insight into its pathogenesis. Incorporating the multiorgan involvement of LC and its bewildering range of symptoms into a single all encompassing model is a complex task. Receptor polymorphisms, enzyme polymorphisms, the impact of methylation on suppressor and promoter genes, …, create a maze. Broad strokes only are presented here.
HYPOTHESIS
The autonomic and neuroendocrine symptoms of LC, POTS, and CFS are primarily determined by CNS nuclei located in areas without a BBB. LC and ME/CFS microbiomes incriminate low fiber diets that support low bacterial diversity with production of minimal short chain fatty acids (SCFAs), e.g., butyrates, GABA, and B vitamins. This collective shortfall drives the symptoms of these syndromes. POTS represents the intersection of LC and ME/CFS that are predominantly undiagnosed MTHFR 677TT, present in over 30-40% of the population. The oxidative stress of Hcy in the MTHFR genotype, the residual spike protein S in LC, and reactivated viruses due to persistent lymphopenia conspire to overwhelm mitochondria lacking an antioxidant shield. Estrogen and BKN play prominent supporting roles.
1. MTHFR and Homocysteine
The MTHFR gene markedly compromises conversion of 5,10 -methylenetetrahydrofolate to 5-methyltetrahydrofolate. This is critical to the recycling of methyl groups (one carbon pathway or folate cycle). This defective gene leads to an increase in homocysteine, which can’t be sufficiently methylated via B12 to return to methionine (see Figure 1). If the active form of B6 (P5P) is not sufficient, then glutathione, the master antioxidant, will not be produced. ROS will increase and mitochondrial function compromised.
The MTHFR 677T allele frequency is high in Europe (24.1-64.3%), and low in Africa (0-35.5%). The MTHFR 1298C allele frequency is approximately 20-70% in Asia, 24-46% in Europe, and 0-15% in America [4]. According to the CDC, there are more people in the United States who have one or two copies of the MTHFR C677T variant than people who do not. About 30 to 40 percent of the American population may have the C677T mutation. Roughly 25 percent of people of Hispanic descent and 10 to 15 percent of Caucasian descent are homozygous for this variant. So this is an exceedingly common polymorphism with incidence that somewhat reflects the percentage of individuals that develop LC.
Hcy is a strong marker for both LC [2], the MTHFR gene [5], and for ME/CFS [6]. High Hcy may be a good marker for the MTHFR gene and low folate may be a good inverse marker for homocysteine [7]. Hcy is a strong marker for thrombosis and may explain the oft encountered thrombosis in Covid-19. Hcy via CVOs causes brain fog in LC (see CVO section) [8].
Wild type and heterozygous MTHFR gene variants characteristically exhibit hypomethylation [9]. But when the genotype is 677TT (homozygous), DNA overmethylation is encountered [10]. Those homozygous for 677T need higher doses of 5-methyltetrahydrofolate (methylfolate) [11].
Methylation status in the body can be determined by measuring whole blood histamine, as histamine and methylation are inversely related. Low blood histamine levels indicate that the individual may be overmethylated; high levels indicate undermethylation [12].
2. POSTURAL ORTHOSTATIC TACHYCARDIA SYNDOME
POTS represents a complex mixture of genetic and epigenetic inputs. It generally exhibits a 80%-90% female predominance, afflicts those 15-45, and is typically precipitated by a viral infection [13]. POTS is considered a disorder of the hypothalamus-pituitary-adrenal (HPA) axis that comes with a paradox - low renin and low aldosterone in the face of low blood volume. This was originally reported in POTS with ME/CFS and later with LC [14]. In addition to low renin and aldosterone ME/CFS patients have lower levels of oxytocin, corticotropin-releasing hormone (CRH), and vasopressin (ADH)[15].
Most POTS patients are hypovolemic. Most are low flow (neuropathic) type. All with low flow POTS have elevated Ang II and low plasma renin activity (PRA). Almost 70% of those with CFS have orthostatic intolerance [16]. These may represent those with concomitant but undiagnosed MTHFR 677TT [17].
The tonic vasoconstriction shrinks blood volume and the juxtaglomerular apparatus (JGA) senses relative normovolemia. Renin is lower than would be otherwise expected. Neuropathic POTS patients have angiotensin II autoantibodies [18] or beta adrenergic autoantibodies [19,20]. Autoantibodies to receptors can block or accentuate their actions [21], but in neuropathic POTS autoantibodies accentuate expression [22]. Angiotensin II antibodies have also been found in LC [23]. Angiotensin receptor blockers (ARBs) can block both angiotensin II type 1 receptors and beta adrenergic receptors [18,24].
Low flow or neuropathic POTS can also be seen in the absence of autoantibodies when the vasodilator nitric oxide (NO) has been oxidized [25]. This can also be blocked by ARBs [26].
Vascular leakage induced by BKN might explain the lower limb cyanosis in neuropathic POTS. A preceding viral respiratory infection can upregulate BKN tenfold [27].
Down regulation of ACE (ACE degrades BKN) by estrogen combined with a viral infection can supercharge BKN. This indirect inhibition of renin by BKN is greater than direct inhibition of renin by the RAS [28]. This might partially explain the decreased renin in the POTS paradox. An increased Ca/Mg might also contribute [29,30].
3. Gut Microbiome
A The neuropathic type of POTS seen in ME/CFS [16] might be due to a CVO mediated glutamate/GABA imbalance that leads to decreased ADH, oxytocin, and CRH (see CVO section). It is both a neuroendocrine and an autonomic disorder. Gut microbiome studies of both LC [31] and ME/CFS [32] report deficiencies in butyrate and GABA producing bacteria, e.g., Bacteroides [33], Bifidobacterium dentium [34], and Lactobacillus brevium [35]. Bacteroides not only drives gut microbiota diversity but also is one of only six known species that produces the active P5P (PLP). B6 deficiency itself reduces the relative abundance of Bacteroides in the microbiota [36].
Without P5P glutathione cannot be synthesized from Hcy (see Figure 1). A gut microbiome that lacks an abundance of histamine degrading Bifidobacteria leaves allergic features unopposed [37]. This may be contributory to MCAS. GABA secreted by histaminergic neurons also downregulates histamine signaling [38]. The endogenous pathways for the degradation of histamine require either methylation (SAMe, Mg2+, and ATP) or mitochondrial ALDH. Alcohol intolerance is a primary complaint in LC, CFS, and MCAS with their dysfunctional mitochondria. Hepatic metabolism of alcohol yields acetaldehyde, but further degradation in mitochondria in such individuals is compromised with symptoms ranging from headache to hangover.
LC, ME/CFS and by extension POTS share gut microbiome features that underscore the criticality of bacterial GABA synthesis to proper functioning of the HPA axis, mediated by CVOs. GABA receptor polymorphisms are linked with restless legs syndrome (RLS) [39]. RLS is a subset of periodic limb movement disorder (PLMD) aka nocturnal myoclonus. This is considered to be a seizure disorder and is increased in LC [40], ME/CFS [41], and POTS [42]. GABA is used to treat seizure disorders. This further underscores the role of GABA deficiency in the pathogenesis of POTS, LC, and CFS.
4. CVOs
Several recent articles have highlighted CVOs [43,44]. There are several areas within the brain that abut ventricles and have no blood brain barrier. Several of these, which are called circumventricular organs (CVOs), are quite relevant to the autonomic and neuroendocrine dysfunctions of LC.
These areas are the pineal gland, the posterior pituitary a.k.a. neurohypothesis, the paraventricular nucleus (PVN) in the median eminence, the area postrema (AP), the organum vasculosum of the lamina terminalis (OVLT) or supraoptic crest, and the subfornical organ (see Figure 2).
The AP is part of the dorsal vagal complex, closely associated with baroreceptors and chemoreceptors, as well as cranial nerves VII and IX (see Figure 3).
CVOs enable a diet induced CNS glutamate (excitatory)/GABA (inhibitory) imbalance to precipitate not only POTS but also symptoms that involve decreased oxytocin, vasopressin (ADH), corticotropin releasing hormone (CRH for ACTH), and gonadotropin releasing hormone (GnRH for LH, FSH).
CVOs can conveniently explain the POTS paradox in the neuropathic hypovolemic low flow subtype (majority). The PVN normally inhibits the baroreflex via tonic GABAergic inputs [47]. Just as sympathetic inhibition and parasympathetic activation respond to carotid and aortic stretch receptors to decrease BP, sympathetic activation and parasympathetic inhibition are required to increase it upon standing. An imbalanced (elevated) glutamate/GABA opposes the latter, causing postural hypotension. GABAergic signals also dictate secretion of neurohypophyseal hormones, including vasopressin (ADH) [48], oxytocin [49] (see Figure 4), CRH [51], and GnRH [52]. On the other hand GABA inhibits the secretion of TRH [53].
The lamina terminalis controls renal secretion of renin and angiotensin [54]. ME/CFS patients have experienced relief from desmopressin [55] and CRF [56]. POTS and ME/CFS have benefited from exogenous CRF and vasopressin (ADH) [57]. Oxytocin in CFS increased stamina, decreased pain, improved cognitive function, decreased fibromyalgia pain, anxiety, and depression. Some LC patients have experienced relief with oxytocin [58]. Many POTS patients claim heat intolerance and this may be mediated by unopposed glutamate [59].
5. Methylation and Shingles
In a recent review compared to controls, patients that experienced mild to moderate COVID-19 or LC exhibited hypomethylation and hypermethylation [60] respectively. This reflects the methylation disposition of MTHFR 677CT and 677TT respectively, i.e., 677CT quickly recovered and 677TT devolved into LC. Hypermethylation in LC persisted for one year after hospitalization [61]. The hypermethylated have higher Hcy levels, which portend more serious Covid-19 and LC [62].
Alcohol intolerance is almost universal among ME/CFS patients [63]. Acetaldehyde, produced by the liver, must then be metabolized in mitochondria. Post exertional malaise may then be joined by a persistent post alcohol hangover.
Shingles serves as a good example for modeling the pathogenesis of LC. VZV can emerge from its dorsal root ganglion (DRG) and invade its designated peripheral sensory nerve (shingles) or it can invade sensory fibers of the facial nerve (VII). These facial nerve sensory fibers emerge from the NTS and the AP (see Figure 3). The AP is the first CVO encountered by any VZV migrating upstream through the subarachnoid CSF and the foramen of Magendie and into the fourth ventricle from its DRG origin. Justin Bieber encountered herpes zoster oticus affiliated with cranial nerve VII (Ramsay Hunt Syndrome) post vaccine. Shingles risk is definitely increased in those with LC or CFS [64]. This may be due to hypermethylation that characterizes LC [60], as lysine is hypermethylated initiating cold sores [65]. Whether the culprit virus is VZV, human herpes virus 6 or 7 (HHV6,7) or is a recent (migrating) or remote (dormant) resident, e.g., geniculate ganglion, seems moot.
6. Autoimmunity in Females
Increased autoimmune disease in females may be due to their enhanced innate immunity and production of pleiotropic IFN gamma. When combined with chronic low grade inflammation, this pro-inflammatory pleiotropic cytokine may switch from self recognition to autoimmune status [66].
IFN gamma is primarily secreted by CD4+ and CD8+ T cells. These cells also induce production of C1 esterase inhibitors (C1INH) by hepatocytes. Lymphopenia is associated with persistence of COVID-19 symptoms [67]. CD147 (basigin) receptors but not ACE2 receptors are found on peripheral blood mononuclear cells (PBMCs) [68]. This receptor may mediate the lymphopenia. CD147 epitopes are also found on the spike protein S, as Justin Bieber discovered, and are responsible for this T cell lymphopenia and drop in surveillance. Any subsequent viral infection that challenges lymphocytes opens the door to the classic complement cascade, which cross talks with the kallikrein kinin system (KKS) and increases BKN [69].
Estrogen affects more than BKN. It may play a role in neurogenic POTS by decreasing renin levels, ACE activity (increasing BKN), AT1 receptor density, and aldosterone [70].
In addition estrogen appears to lower cortisol [71] and ACE2, at least in the lungs [72]. Perhaps lower pulmonary ACE2 is Covid 19 protective, but the downregulated ACE2 may be associated with higher levels of des-Arg9-bradykinin (see Figure 5). Des-arg-BKN appears to be a prominent kinin in Covid-19 [73].
7. A Mitochondria and Oxidative Stress
Mitochondrial health is especially important in maintaining optimal levels of active vitamin D. Men with the highest compared to lowest 1,25(OH)2D and activation ratios, i.e., active form/storage form, (1,25(OH)2D/25(OH)D) are more likely to possess butyrate-producing bacteria that are associated with favorable gut microbial health. Sun exposure may affect the storage form of vitamin D, but it exerts little influence on the active hormone [75]. Oxidative stress (ROS due to inflammation, smoking, toxins, …) compromises mitochondrial oxidative phosphorylation. If these ROS are not quenched by onboard antioxidants, the cell risks lysis [76]. Many mitochondria are shut down to avoid this, diminishing ATP production. The Krebs or TCA cycle with its electron transport chain ETC also requires activated B2 and B3. ATP is required to synthesize glutamine (Gln) from glutamate (Glu) and to recycle GABA in astrocytes (see Figure 6). Recycling of Glu is not so limited. Any shortage of active B6 (P-5-P), required by glutamate decarboxylase (GAD), further compromises production of GABA from glutamate.
7. B Magnesium and Vitamins
Magnesium deficiency is an additional plausible explanation for low aldosterone and low cortisol. Synthesis of aldosterone requires aldosterone synthase and synthesis of cortisol requires 11beta hydroxylase. These two enzymes are part of the CYP450 system, all of which require Mg++. Furthermore these two reactions occur in the mitochondria [78], i.e., they also require ATP. If oxidative stress is sufficient and antioxidants are not, mitochondria are shut down and ATP down regulated, limiting synthesis of aldosterone and cortisol [79]. Females are more magnesium deficient than males before age 50 (see Figure 7). An elevated Ca:Mg may also compromise synthesis of aldosterone and cortisol [29,30].
Magnesium is also required for the synthesis of all eight B vitamins, except biotin (B7). Mitochondrial function is dependent upon adequacy of all the B vitamins[80].
If mitochondrial function is marginal, production of the active form of vitamin D may be marginal. Mitochondrial CYP27B1 is the only enzyme that hydroxylates C1 (activates vitamin D)[81].
Thiamine (B1), riboflavin (B2), niacin (B3), pantothenate (B5), pyridoxine (B6) require phosphorylation for activation. Cyanocobalamin and folate require methylation (SAMe) for activation. Magnesium is required for both phosphorylation and methylation. Hcy is highly influenced by B vitamin status [82].
The last enzymatic step to produce serotonin, dopamine, and GABA are decarboxylases that require active B6 (P5P), as a cofactor. Not surprisingly B6 enhances libido. Melatonin and glutathione synthesis also requires B6.
Norepinephrine synthesis requires magnesium. A shortfall in magnesium or B6 compromises monoamine neurotransmitter synthesis and leads to depression, a symptom shared by LC, POTS, ME/CFS, MCAS, and EDS. High doses of pyridoxine are associated with peripheral neuropathy, whereas high doses of the active form (P5P) are not[83]. Estrogens compete with B6 for some receptor sites including that for serotonin synthesis[84] and may increase risk for depression.
8. Thoughts on Therapy
Changing one's diet is more difficult than changing one's religion. But adding a little supplemental GABA might help in the meantime. Attaining a blood Ca/Mg around 2.0 can also be challenging. But maintaining a 25(OH)D3 of 50 ng/mL with abundant antioxidants is critical to its activation to 1,25(OH)2D3. Other recommended epigenetic actions include supplementing with the methylated or phosphorylated forms of all the B vitamins, especially methyl folate. Absorption of magnesium and B6 is enhanced when taken together. A good supplementary probiotic (with some GABA) might be worth considering. However, under professional supervision angiotensin receptor blockers (ARBs) might be worth pursuing, but even a small dose can elicit hypoglycemia and/or hypotension.
The potential benefits of ARBs for LC include:
1 ARBs upregulate GABA, which displays anti-hypertension, anti-senescence, anti-diabetes, antioxidant, and anti-inflammatory properties[85].
2 ARBs block AT1Rs (and hypertension induced dementia) and increase ACE[86].
5 ARBs down regulate TGF-β1, increased in LC[90], CFS[91], AD[92], Marfan Syndrome, and Ehlers–Danlos Syndrome (EDS).
8 ARBs are neuroprotective[95].
CONCLUSION
In summary LC reveals undiagnosed MTHFR 677TT present in 30-40% of the population. These patients comprise the majority of those with LC and POTS. The integration of CVOs with the gut microbiome opens new insight into the POTS paradox and links LC, POTS, and ME/CFS. Homocysteine, a marker for MTHFR polymorphisms, is elevated in LC and ME/CFS. It can directly access the BBB less CNS and is directly proportional to brain fog severity and oxidative stress. In the absence of critical cofactors synthesis of the master antioxidant glutathione from Hcy cannot proceed (see Figure 1). HIV (low CD4/CD8) and SARS CoV2 (high CD4/CD8) specifically target CD4+ and CD8+ T cells respectively, disabling surveillance of intracellular pathogens and enabling the emergence of EBV and VZV, previously dormant within dorsal root ganglia. VZV also targets CD8+ T cells [98].
Loss of these T cells translates to a decrease in IFN gamma and with it a decrease in hepatic C1INH. The classic complement pathway is triggered and the KKS is activated. BKN is released. Any recent viral infection induces additional release of BKN. Estrogen, an ACE inhibitor, prolongs BKN half life. BKN combines with histamine to produce low volume low flow POTS. The LC and ME/CFS gut microbiomes lack biodiversity (insufficient bifidobacteria, lactobacilli, ..). These latter bacteria produce butyrates (SCFAs), GABA, B vitamins, and can degrade histamine.
Subsequent inflammation induced oxidative stress from homocysteine, residual spike S protein, and reactivated viruses conspire to overwhelm those with insufficient on board antioxidants. An additional magnesium shortfall in 15-50 year old females leads to fatigue and post exertional dyspnea.
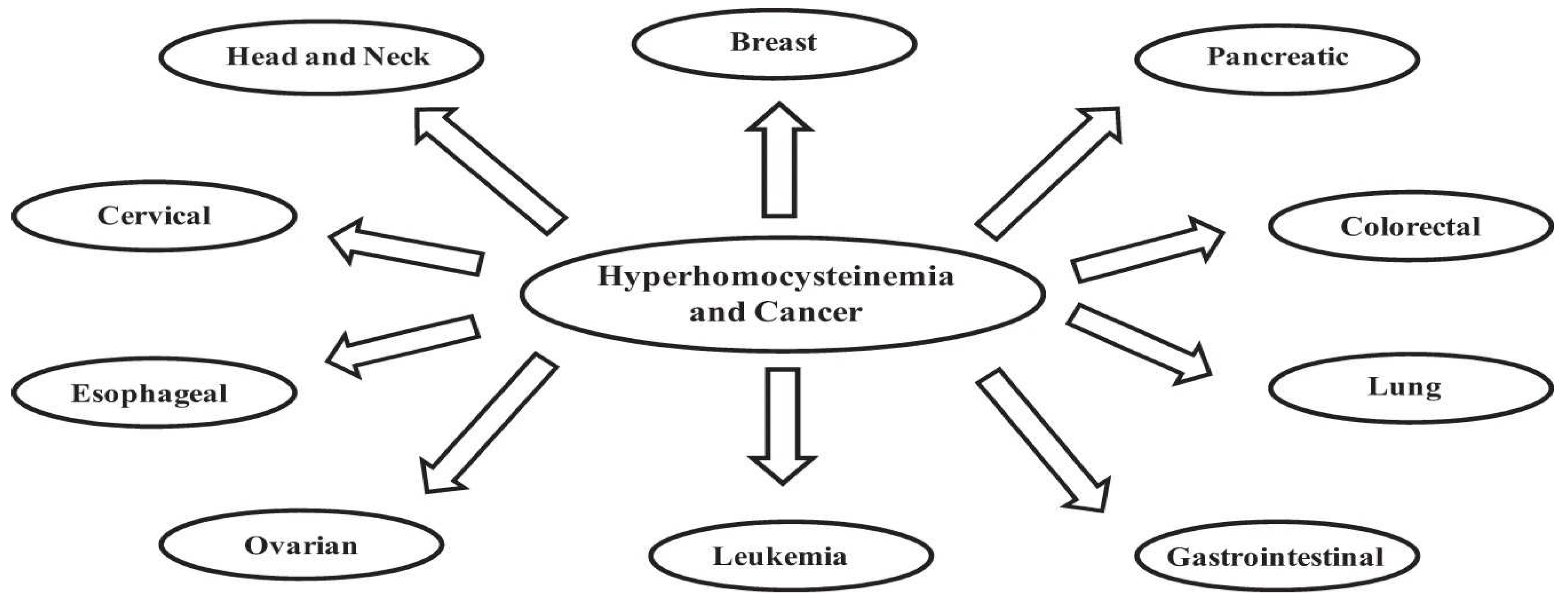
In the presented hypothesis methylation plays a prominent role. It is at the center of the MTHFR gene, afflicting undiagnosed millions, now suffering from LC and/or POTS. This information is easily available via 23andme or a blood test for homocysteine. This should be job one. If the latter is within normal limits, then MTHFR is off the table. 677TT creates a 70% bottleneck and 1298C/677T a 30% bottleneck in recycling/removing Hcy. Elevated Hcy carries many short term (thrombosis) and long-term problems, best addressed ASAP. Although the primary threat is vascular, the hypermethylation seen in LC is also a marker for many cancers (see Figure 8).
Hypomethylation (methylation decreases with age) is seen in hepatocellular carcinoma, breast cancer, lung cancer, nasopharyngeal cancer, smooth muscle sarcoma, and neuroendocrine tumor [100]. Note some are increased in both hyper- and hypomethylation.
Methylation is only half the story. Sufficient antioxidants to protect mitochondria are vital. Heart rate variability (HRV), which varies inversely with high sensitivity C reactive protein (hs-CRP), should reflect oxidative stress.
Given the high frequency of this polymorphism in the general population and its potential devastation, everyone should be privy to their risks and those of their offspring, especially in the post Covid era.
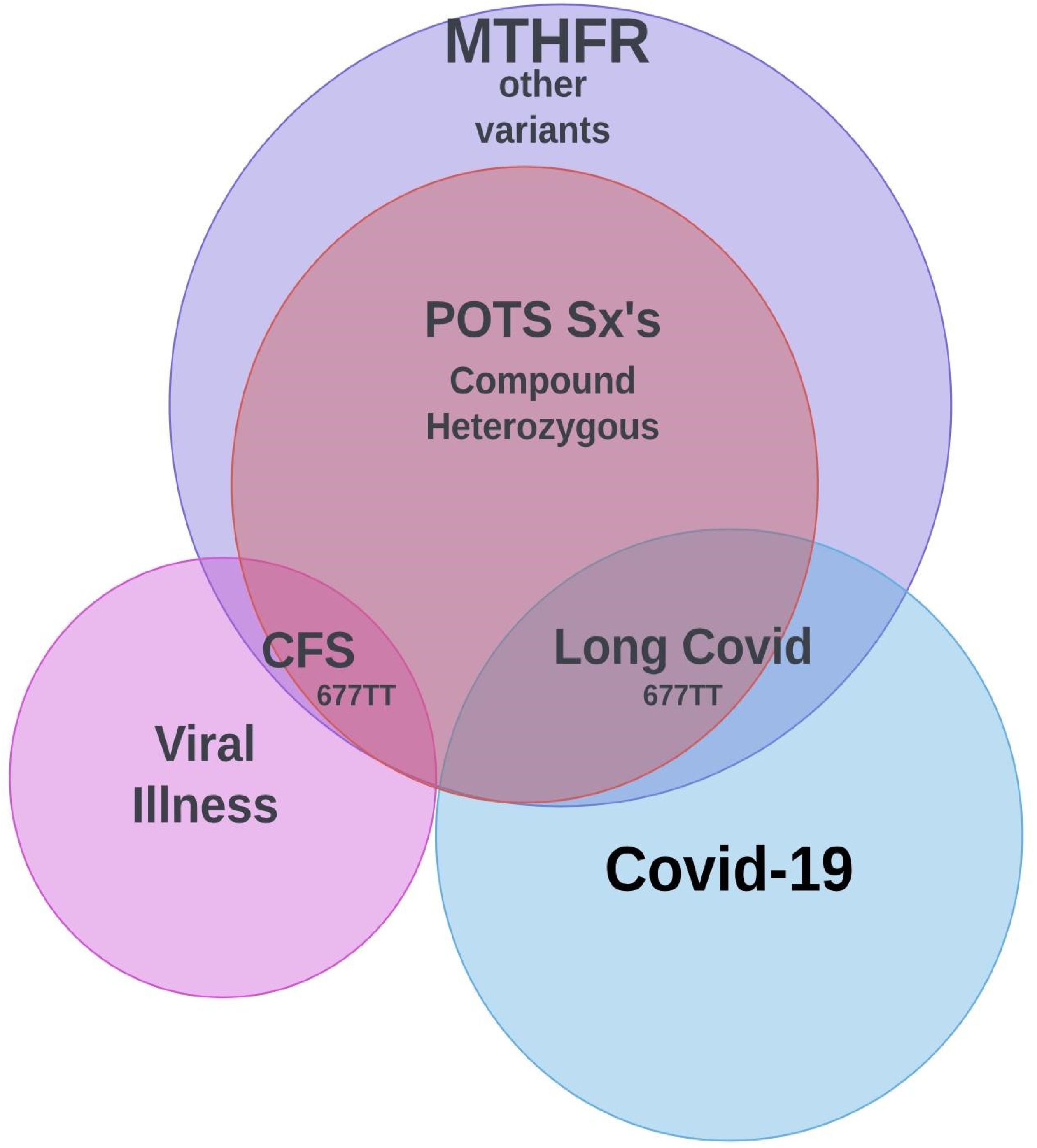
Figure 9.
Venn view of hypothesis. Circle sizes not to scale. The LC/CFS microbiome of decreased bacterial diversity and decreased butyrate and GABA producing bacteria[1,2,67] aggravates symptoms in those with the MTHFR 677T genotype. The vitamin D deficient and those with POTS also lack these gut microbiota [75, 101,102].
Figure 9.
Venn view of hypothesis. Circle sizes not to scale. The LC/CFS microbiome of decreased bacterial diversity and decreased butyrate and GABA producing bacteria[1,2,67] aggravates symptoms in those with the MTHFR 677T genotype. The vitamin D deficient and those with POTS also lack these gut microbiota [75, 101,102].
References
- Karst, M.; Hollenhorst, J.; Achenbach, J. Life-threatening course in coronavirus disease 2019 (COVID-19): Is there a link to methylenetetrahydrofolic acid reductase (MTHFR) polymorphism and hyperhomocysteinemia? Med Hypotheses 2020, 144, 110234–110234. [Google Scholar] [CrossRef] [PubMed]
- Ponti, G.; Pastorino, L.; Manfredini, M.; Ozben, T.; Oliva, G.; Kaleci, S.; Iannella, R.; Tomasi, A. COVID-19 spreading across world correlates with C677T allele of the methylenetetrahydrofolate reductase (MTHFR) gene prevalence. J. Clin. Lab. Anal. 2021, 35, e23798. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadeq, D.W.; Nasrallah, G.K. The Spectrum of Mutations of Homocystinuria in the MENA Region. Genes 2020, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liu, Y.; Li, Y.; Fan, S.; Zhi, X.; Lu, X.; Wang, D.; Zheng, Q.; Wang, Y.; Wang, Y.; et al. Geographical Distribution of MTHFR C677T, A1298C and MTRR A66G Gene Polymorphisms in China: Findings from 15357 Adults of Han Nationality. PLoS ONE 2013, 8, e57917. [Google Scholar] [CrossRef] [PubMed]
- Carpenè, G.; Negrini, D.; Henry, B.M.; Montagnana, M.; Lippi, G. Homocysteine in coronavirus disease (COVID-19): A systematic literature review. Diagnosis 2022, 9, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Regland, B.; Andersson, M.; Abrahamsson, L.; Bagby, J.; Dyrehag, L.E.; Gottfries, C.G. Increased Concentrations of Homocysteine in the Cerebrospinal Fluid in Patients with Fibromyalgia and Chronic Fatigue Syndrome. Scand. J. Rheumatol. 1997, 26, 301–307. [Google Scholar] [CrossRef]
- Bottiglieri, T.; Laundy, M.; Crellin, R.; Toone, B.K.; Carney, M.W.P.; Reynolds, E.H. Homocysteine, folate, methylation, and monoamine metabolism in depression. J. Neurol. Neurosurg. Psychiatry 2000, 69, 228–232. [Google Scholar] [CrossRef]
- Oner, P.; Yilmaz, S.; Doğan, S. High Homocysteine Levels Are Associated with Cognitive Impairment in Patients Who Recovered from COVID-19 in the Long Term. J. Pers. Med. 2023, 13, 503. [Google Scholar] [CrossRef]
- Ren, J.-C.; Wu, Y.-X.; Wu, Z.B.; Zhang, G.-H.; Wang, H.B.; Liu, H.B.; Cui, J.-P.B.; Chen, Q.; Liu, J.; Frank, A.; et al. MTHFR Gene Polymorphism Is Associated With DNA Hypomethylation and Genetic Damage Among Benzene-Exposed Workers in Southeast China. J. Occup. Environ. Med. 2018, 60, e188–e192. [Google Scholar] [CrossRef]
- Fryar-Williams, S. Fundamental Role of Methylenetetrahydrofolate Reductase 677 C → T Genotype and Flavin Compounds in Biochemical Phenotypes for Schizophrenia and Schizoaffective Psychosis. Front. Psychiatry 2016, 7, 172. [Google Scholar] [CrossRef]
- Ashfield-Watt, P.A.; et al. “Methylenetetrahydrofolate reductase 677C-->T genotype modulates homocysteine responses to a folate-rich diet or a low-dose folic acid supplement: A randomized controlled trial”. Am. J. Clin. Nutr. 2002, 76, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Walsh, W.J. Nutrient Power: Heal Your Biochemistry and Heal Your Brain. Skyhorse Publishing 2012 USA.
- Fedorowski, A. Postural orthostatic tachycardia syndrome: Clinical presentation, aetiology and management. J. Intern. Med. 2018, 285, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Chadda, K.R.; Blakey, E.E.; Huang, C.L.H.; Jeevaratnam, K. Long COVID-19 and Postural Orthostatic Tachycardia Syndrome- Is Dysautonomia to Be Blamed? Front. Cardiovasc. Med. 2022, 9, 860198. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J. Betrayal by the Brain: The Neurologic Basis of Chronic Fatigue Syndrome, Fibromyalgia Syndrome, and Related Neural Network. 1st edition Routledge 1996 USA.
- Okamoto, L.E.; Raj, S.R.; Peltier, A.; Gamboa, A.; Shibao, C.; Diedrich, A.; Black, B.K.; Robertson, D.; Biaggioni, I. Neurohumoral and haemodynamic profile in postural tachycardia and chronic fatigue syndromes. Clin. Sci. 2011, 122, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.M.; Glover, J.L.; Medow, M.S. Increased plasma angiotensin II in postural tachycardia syndrome (POTS) is related to reduced blood flow and blood volume. Clin. Sci. 2006, 110, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, H.; Murphy, T.A.; Nuss, Z.; Liles, J.; Liles, C.; Aston, C.E.; Raj, S.R.; Fedorowski, A.; Kem, D.C. Angiotensin II Type 1 Receptor Autoantibodies in Postural Tachycardia Syndrome. J. Am. Hear. Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Fedorowski, A.; Li, H.; Yu, X.; Koelsch, K.A.; Harris, V.M.; Liles, C.; Murphy, T.A.; Quadri, S.M.S.; Scofield, R.H.; Sutton, R.; et al. Antiadrenergic autoimmunity in postural tachycardia syndrome. Europace 2016, 19, 1211–1219. [Google Scholar] [CrossRef]
- Badiudeen, T.; Forsythe, E.A.; Bennett, G.; Li, H.; Yu, X.; Beel, M.; Nuss, Z.; Blick, K.E.; Okamoto, L.E.; Arnold, A.C.; et al. A functional cell-based bioassay for assessing adrenergic autoantibody activity in postural tachycardia syndrome. J. Transl. Autoimmun. 2019, 2, 100006. [Google Scholar] [CrossRef]
- Blecher, M. “Receptors, antibodies, and disease”. Clin Chem. 1984, 30, 1137–1356. [Google Scholar] [CrossRef]
- Wirth, K.; Scheibenbogen, C. A Unifying Hypothesis of the Pathophysiology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Recognitions from the finding of autoantibodies against ß2-adrenergic receptors. Autoimmun. Rev. 2020, 19, 102527. [Google Scholar] [CrossRef]
- Briquez, P.S.; Rouhani, S.J.; Yu, J.; Pyzer, A.R.; Trujillo, J.; Dugan, H.L.; Stamper, C.T.; Changrob, S.; Sperling, A.I.; Wilson, P.C.; et al. Severe COVID-19 induces autoantibodies against angiotensin II that correlate with blood pressure dysregulation and disease severity. Sci. Adv. 2022, 8, eabn3777. [Google Scholar] [CrossRef]
- Barki-Harrington, L.; Luttrell, L.M.; Rockman, H.A. Dual Inhibition of β-Adrenergic and Angiotensin II Receptors by a Single Antagonist. Circulation 2003, 108, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Medow, M.S.; et al. Decreased Microvascular Nitric Oxide–Dependent Vasodilation in Postural Tachycardia Syndrome. Circulation 2005, 112, 2611–2618. [Google Scholar] [CrossRef]
- Stewart, J.M.; Taneja, I.; Glover, J.; Medow, M.S.; Lang, J.A.; Krajek, A.C.; McNeely, B.D.; Meade, R.D.; Fujii, N.; Seely, A.J.E.; et al. Angiotensin II type 1 receptor blockade corrects cutaneous nitric oxide deficit in postural tachycardia syndrome. Am. J. Physiol. Circ. Physiol. 2008, 294, H466–H473. [Google Scholar] [CrossRef] [PubMed]
- Laith, AIK; et al. “Association between the level of Bradykinin and viral infection in patient suffering from respiratory infection, renal transplant, and renal failure”. Ann. Trop. Med. Public Health 22020, 3, SP231203. [Google Scholar]
- Kiowski, W.; Linder, L.; Kleinbloesem, C.; van Brummelen, P.; Bühler, F.R. Blood pressure control by the renin-angiotensin system in normotensive subjects. Assessment by angiotensin converting enzyme and renin inhibition. Circulation 1992, 85, 1–8. [Google Scholar] [CrossRef]
- Persson, P.B. Renin: Origin, secretion and synthesis. J. Physiol. 2003, 552, 667–671. [Google Scholar] [CrossRef]
- Ichihara, A.; Suzuki, H.; Saruta, T. Effects of magnesium on the renin-angiotensin-aldosterone system in human subjects. J. Lab. Clin. Med. 1993, 122, 432–440. [Google Scholar]
- Yeoh, YK, et al “Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19”. Gut 2021, 70, 698–706. [CrossRef]
- König, R.S.; Albrich, W.C.; Kahlert, C.R.; Bahr, L.S.; Löber, U.; Vernazza, P.; Scheibenbogen, C.; Forslund, S.K. The Gut Microbiome in Myalgic Encephalomyelitis (ME)/Chronic Fatigue Syndrome (CFS). Front. Immunol. 2022, 12, 628741. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y.; et al. GABA-producing Bifidobacterium dentium modulates visceral sensitivity in the intestine. Neurogastroenterol. Motil. 2016, 29. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.Y.; Lin, H.-T.V.; Tsai, G.J. Gamma-aminobutyric acid production in black soybean milk by Lactobacillus brevis FPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model. Process. Biochem. 2013, 48, 559–568. [Google Scholar] [CrossRef]
- Mayengbam, S.; Chleilat, F.; Reimer, R.A. Dietary Vitamin B6 Deficiency Impairs Gut Microbiota and Host and Microbial Metabolites in Rats. Biomedicines 2020, 8, 469. [Google Scholar] [CrossRef]
- Dev, S.; Mizuguchi, H.; Das, A.K.; Matsushita, C.; Maeyama, K.; Umehara, H.; Ohtoshi, T.; Kojima, J.; Nishida, K.; Takahashi, K.; et al. Suppression of Histamine Signaling by Probiotic Lac-B: A Possible Mechanism of Its Anti-allergic Effect. J. Pharmacol. Sci. 2008, 107, 159–166. [Google Scholar] [CrossRef]
- Yu, X.; Ye, Z.; Houston, C.M.; Zecharia, A.Y.; Ma, Y.; Zhang, Z.; Uygun, D.S.; Parker, S.; Vyssotski, A.L.; Yustos, R.; et al. Wakefulness Is Governed by GABA and Histamine Cotransmission. Neuron 2015, 87, 164–178. [Google Scholar] [CrossRef]
- Jiménez-Jiménez, F.J.; Esguevillas, G.; Alonso-Navarro, H.; Zurdo, M.; Turpín-Fenoll, L.; Millán-Pascual, J.; Adeva-Bartolomé, T.; Cubo, E.; Navacerrada, F.; Amo, G.; et al. Gamma-aminobutyric acid (GABA) receptors genes polymorphisms and risk for restless legs syndrome. Pharmacogenomics J. 2018, 18, 565–577. [Google Scholar] [CrossRef]
- Weinstock, L.B.; Brook, J.B.; Walters, A.S.; Goris, A.; Afrin, L.B.; Molderings, G.J. Restless legs syndrome is associated with long-COVID in women. Sleep Med. 2022, 18, 1413–1418. [Google Scholar] [CrossRef]
- Civelek, G.M.; Ciftkaya, P.O.; Karatas, M. Evaluation of restless legs syndrome in fibromyalgia syndrome: An analysis of quality of sleep and life. J. Back Musculoskelet. Rehabilitation 2014, 27, 537–544. [Google Scholar] [CrossRef]
- Dodson, C.; Bagai, K.; Weinstock, L.B.; Thompson, E.; Okamoto, L.E.; Peltier, A.; Raj, S.R.; Walters, A.S. Restless legs syndrome is increased in postural orthostatic tachycardia syndrome. Sleep Med. 2021, 17, 791–795. [Google Scholar] [CrossRef]
- Jammoul, M.; Naddour, J.; Madi, A.; Reslan, M.A.; Hatoum, F.; Zeineddine, J.; Abou-Kheir, W.; Lawand, N. Investigating the possible mechanisms of autonomic dysfunction post-COVID-19. Auton. Neurosci. 2022, 245, 103071–103071. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Ling, E.-A. The circumventricular organs. Clin Exp Pharmacol Physiol. 2017, 32, 879–892. [Google Scholar] [CrossRef]
- Cytokines Driving Sympathetic Nervous System Activation In The Subfornical Organ: Implications For Heart Failure And Hypertension (2015) Gabriel Bassi https://brainimmune.com/subfornical-organ-heart-failure-hypertension/).
- Cutsforth-Gregory, J.K.; Benarroch, E.E. Nucleus of the solitary tract, medullary reflexes, and clinical implications. Neurology 2017, 88, 1187–1196. [Google Scholar] [CrossRef]
- Page, M.C.; Cassaglia, P.A.; Brooks, V.L.; Erdos, B.; Clifton, R.R.; Liu, M.; Li, H.; McCowan, M.L.; Sumners, C.; Scheuer, D.A.; et al. GABA in the paraventricular nucleus tonically suppresses baroreflex function: Alterations during pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R1452–R1458. [Google Scholar] [CrossRef] [PubMed]
- Haam, J.; Popescu, I.R.; Morton, L.A.; Halmos, K.C.; Teruyama, R.; Ueta, Y.; Tasker, J.G. GABA Is Excitatory in Adult Vasopressinergic Neuroendocrine Cells. J. Neurosci. 2012, 32, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Kim, Y.-B.; Kim, J.S.; Bin Kim, W.; Kim, Y.S.; Han, H.C.; Colwell, C.S.; Cho, Y.-W.; Kim, Y.I. GABAergic inhibition is weakened or converted into excitation in the oxytocin and vasopressin neurons of the lactating rat. Mol. Brain 2015, 8, 1–9. [Google Scholar] [CrossRef]
- Unnikrishnan P, Diabetes Insipidus, Dept of Neuroanesthesia SCTIMST, Trivandrum, Kerala, India https://www.uzhnu.edu.ua/uk/infocentre/get/23985.
- Kakizawa, K.; Watanabe, M.; Mutoh, H.; Okawa, Y.; Yamashita, M.; Yanagawa, Y.; Itoi, K.; Suda, T.; Oki, Y.; Fukuda, A. A novel GABA-mediated corticotropin-releasing hormone secretory mechanism in the median eminence. Sci. Adv. 2016, 2, e1501723–1501723. [Google Scholar] [CrossRef]
- Moore, A.M.; Abbott, G.; Mair, J.; Prescott, M.; Campbell, R.E. Mapping GABA and glutamate inputs to gonadotrophin-releasing hormone neurones in male and female mice. J. Neuroendocr. 2018, 30, e12657. [Google Scholar] [CrossRef]
- Wiens, S.C.; Trudeau, V.L. Thyroid hormone and γ-aminobutyric acid (GABA) interactions in neuroendocrine systems. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 332–344. [Google Scholar] [CrossRef]
- McKinley, M.J.; Gerstberger, R.; Mathai, M.L.; Oldfield, B.J.; Schmid, H. The lamina terminalis and its role in fluid and electrolyte homeostasis. J. Clin. Neurosci. 1999, 6, 289–301. [Google Scholar] [CrossRef]
- Miwa, K. Down-regulation of renin–aldosterone and antidiuretic hormone systems in patients with myalgic encephalomyelitis/chronic fatigue syndrome. J. Cardiol. 2017, 69, 684–688. [Google Scholar] [CrossRef]
- Pereira, G.; Gillies, H.; Chanda, S.; Corbett, M.; Vernon, S.D.; Milani, T.; Bateman, L. Acute Corticotropin-Releasing Factor Receptor Type 2 Agonism Results in Sustained Symptom Improvement in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Syst. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Coffin, S.T.; Black, B.K.; Biaggioni, I.; Paranjape, S.Y.; Orozco, C.; Black, P.W.; Dupont, W.D.; Robertson, D.; Raj, S.R. Desmopressin acutely decreases tachycardia and improves symptoms in the postural tachycardia syndrome. Hear. Rhythm. 2012, 9, 1484–1490. [Google Scholar] [CrossRef]
- Diep, P.-T.; Chaudry, M.; Dixon, A.; Chaudry, F.; Kasabri, V. Oxytocin, the panacea for long-COVID? a review. Horm. Mol. Biol. Clin. Investig. 2022, 43, 363–371. [Google Scholar] [CrossRef]
- Amir, S. Stimulation of the paraventricular nucleus with glutamate activates interscapular brown adipose tissue thermogenesis in rats. Brain Res. 1990, 508, 152–155. [Google Scholar] [CrossRef]
- Nikesjö, F.; Sayyab, S.; Karlsson, L.; Apostolou, E.; Rosén, A.; Hedman, K.; Lerm, M. Defining post-acute COVID-19 syndrome (PACS) by an epigenetic biosignature in peripheral blood mononuclear cells. Clin. Epigenetics 2022, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Balnis, J.; Madrid, A.; Hogan, K.J.; Drake, L.A.; Adhikari, A.; Vancavage, R.; Singer, H.A.; Alisch, R.S.; Jaitovich, A. Persistent blood DNA methylation changes one year after SARS-CoV-2 infection. Clin. Epigenetics 2022, 14, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sarı, I.K.; Keskin, O.; Keskin, A.S.; Ellidağ, H.Y.; Harmandar, O. Is Homocysteine Associated with the Prognosis of Covid-19 Pneumonia. Int. J. Clin. Pr. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- American Myalgic Encephalomyelitis and Chronic Fatigue Syndrome Society https://ammes.org/diet/.
- Waltz, E. Could long COVID be linked to herpes viruses? Early data offer a hint. Nature 2022. [Google Scholar] [CrossRef]
- Cruz, C.; Della Rosa, M.; Krueger, C.; Gao, Q.; Horkai, D.; King, M.; Field, L.; Houseley, J.; The Babraham Institute; Kingdom, U. Tri-methylation of histone H3 lysine 4 facilitates gene expression in ageing cells. eLife 2018, 7. [Google Scholar] [CrossRef]
- De Benedetti, F.; Prencipe, G.; Bracaglia, C.; Marasco, E.; Grom, A.A. Targeting interferon-γ in hyperinflammation: Opportunities and challenges. Nat. Rev. Rheumatol. 2021, 17, 678–691. [Google Scholar] [CrossRef]
- Varghese, J.; Sandmann, S.; Ochs, K.; Schrempf, I.-M.; Frömmel, C.; Dugas, M.; Schmidt, H.H.; Vollenberg, R.; Tepasse, P.-R. Persistent symptoms and lab abnormalities in patients who recovered from COVID-19. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kuklina, E.M. T Lymphocytes as Targets for SARS-CoV-2. Biochem. (Moscow) 2022, 87, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Bossi, F.; Peerschke, E.I.; Ghebrehiwet, B.; Tedesco, F. Cross-talk between the complement and the kinin system in vascular permeability. Immunol. Lett. 2011, 140, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Hao, Z.-W.; Wang, Y.-F. The effect of estrogen in coronavirus disease 2019. Am. J. Physiol. Cell. Mol. Physiol. 2021, 321, L219–L227. [Google Scholar] [CrossRef]
- Herrera, A.Y.; Hodis, H.N.; Mack, W.J.; Mather, M. Estradiol Therapy After Menopause Mitigates Effects of Stress on Cortisol and Working Memory. J. Clin. Endocrinol. Metab. 2017, 102, 4457–4466. [Google Scholar] [CrossRef]
- Stelzig, K.E.; Canepa-Escaro, F.; Schiliro, M.; Berdnikovs, S.; Prakash, Y.S.; Chiarella, S.E. Estrogen regulates the expression of SARS-CoV-2 receptor ACE2 in differentiated airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1280–L1281. [Google Scholar] [CrossRef]
- Mendes, G.M.d.M.; Nascimento, I.J.B.D.; Marazzi-Diniz, P.H.; Da Silveira, I.B.; Itaborahy, M.F.; Viana, L.E.; Silva, F.A.; Santana, M.F.; Pinto, R.A.; Dutra, B.G.; et al. The des-Arg9-bradykinin/B1R axis: Hepatic damage in COVID-19. Front. Physiol. 2022, 13, 1080837. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Netea, M.G.; van Deuren, M.; van der Meer, J.W.; de Mast, Q.; Brüggemann, R.J.; van der Hoeven, H. Kallikrein-kinin blockade in patients with COVID-19 to prevent acute respiratory distress syndrome. eLife 2020, 9. [Google Scholar] [CrossRef]
- Thomas, R.L.; Jiang, L.; Adams, J.S.; Xu, Z.Z.; Shen, J.; Janssen, S.; Ackermann, G.; Vanderschueren, D.; Pauwels, S.; Knight, R.; et al. Vitamin D metabolites and the gut microbiome in older men. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Wood, E.; et al. “Role of mitochondria, oxidative stress and the response to antioxidants in myalgic encephalomyelitis/chronic fatigue syndrome: A possible approach to SARS-CoV-2 'long-haulers'? ” Chronic Dis Transl Med. 2021, 7, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Riera, J.J.; Schousboe, A.; Waagepetersen, H.S.; Howarth, C.; Hyder, F. The micro-architecture of the cerebral cortex: Functional neuroimaging models and metabolism. NeuroImage 2008, 40, 1436–1459. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, N.; Koeva, Y. Hydrohysteroid Dehydrogenases—Biological Role and Clinical Importance—Review; Canuto, R.A., Ed.; Dehydrogenases IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Chambers, P. Antioxidants and Long Covid. OALib 2022, 9, 1–19. [Google Scholar] [CrossRef]
- Depeint, F.; Bruce, W.R.; Shangari, N.; Mehta, R.; O’Brien, P.J. Mitochondrial function and toxicity: Role of the B vitamin family on mitochondrial energy metabolism. Chem. Biol. Interact. 2006, 163, 94–112. [Google Scholar] [CrossRef]
- Bikle, DD. “Vitamin D metabolism, mechanism of action, and clinical applications”. Chem Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef]
- Azzini, E.; Ruggeri, S.; Polito, A. Homocysteine: Its Possible Emerging Role in At-Risk Population Groups. Int. J. Mol. Sci. 2020, 21, 1421. [Google Scholar] [CrossRef] [PubMed]
- Hadtstein, F.; Vrolijk, M. Vitamin B-6-Induced Neuropathy: Exploring the Mechanisms of Pyridoxine Toxicity. Adv. Nutr. Int. Rev. J. 2021, 12, 1911–1929. [Google Scholar] [CrossRef]
- Rose, DP. “The interactions between vitamin B6 and hormones”. Vitam Horm. 1978, 36, 53–99. [Google Scholar] [CrossRef]
- Sfera, A.; Thomas, K.G.; Sasannia, S.; Anton, J.J.; Andronescu, C.V.; Garcia, M.; Sfera, D.O.; Cummings, M.A.; Kozlakidis, Z. Neuronal and Non-Neuronal GABA in COVID-19: Relevance for Psychiatry. Reports 2022, 5, 22. [Google Scholar] [CrossRef]
- Campbell, D.J.; Kladis, A.; Valentijn, A.J. Effects of Losartan on Angiotensin and Bradykinin Peptides and Angiotensin-Converting Enzyme. J. Cardiovasc. Pharmacol. 1995, 26, 233–240. [Google Scholar] [CrossRef]
- Singh, PK; et al. “Increased plasma bradykinin level is associated with cognitive impairment in Alzheimer's patients”. Neurobiol Dis. 2020, 139, 104833. [Google Scholar] [CrossRef]
- Zaheer, J.; Kim, H.; Kim, J.S. Correlation of ACE2 with RAS components after Losartan treatment in light of COVID-19. Sci. Rep. 2021, 11, 1–7. [Google Scholar] [CrossRef]
- Kehoe, P.G.; Wong, S.; AL Mulhim, N.; Palmer, L.E.; Miners, J.S. Angiotensin-converting enzyme 2 is reduced in Alzheimer’s disease in association with increasing amyloid-β and tau pathology. Alzheimer's Res. Ther. 2016, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chambers, P. “Long Covid and Neurodegenerative Disease”. J Path Lab Med, /: https, 3673. [Google Scholar]
- Yang, T.; Yang, Y.; Wang, D.; Li, C.; Qu, Y.; Guo, J.; Shi, T.; Bo, W.; Sun, Z.; Asakawa, T. The clinical value of cytokines in chronic fatigue syndrome. J. Transl. Med. 2019, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X; et al. “TGF-β1 factor in the cerebrovascular diseases of Alzheimer's disease”. Eur Rev Med Pharmacol Sci. 2016, 20, 5178–5185.
- Zhang, Z.; Liu, C.; Gan, Z.; Wang, X.; Yi, Q.; Liu, Y.; Wang, Y.; Lu, B.; Du, H.; Shao, J.; et al. Improved Glucose-Stimulated Insulin Secretion by Selective Intraislet Inhibition of Angiotensin II Type 1 Receptor Expression in Isolated Islets of db/db Mice. Int. J. Endocrinol. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Chang, Y.-C.; Wu, L.-C.; Lin, J.-W.; Chuang, L.-M.; Lai, M.-S. Different angiotensin receptor blockers and incidence of diabetes: A nationwide population-based cohort study. Cardiovasc. Diabetol. 2014, 13, 91–91. [Google Scholar] [CrossRef]
- Villapol, S.; Saavedra, J.M. Neuroprotective Effects of Angiotensin Receptor Blockers. Am. J. Hypertens. 2014, 28, 289–299. [Google Scholar] [CrossRef]
- Hoffman, L.B.; Schmeidler, J.; Lesser, G.T.; Beeri, M.S.; Purohit, D.P.; Grossman, H.T.; Haroutunian, V. Less Alzheimer disease neuropathology in medicated hypertensive than nonhypertensive persons. Neurology 2009, 72, 1720–1726. [Google Scholar] [CrossRef]
- van Dalen, J.W.; Marcum, Z.A.; Gray, S.L.; Barthold, D.; van Charante, E.P.M.; van Gool, W.A.; Crane, P.K.; Larson, E.B.; Richard, E. Association of Angiotensin II–Stimulating Antihypertensive Use and Dementia Risk. Neurology 2020, 96, e67–e80. [Google Scholar] [CrossRef]
- Arvin, AM; et al. “Varicella-zoster virus: Aspects of pathogenesis and host response to natural infection and varicella vaccine”. Adv Virus Res. 1996, 46, 263–309. [Google Scholar] [CrossRef]
- Hasan, T.; Arora, R.; Bansal, A.K.; Bhattacharya, R.; Sharma, G.S.; Singh, L.R. Disturbed homocysteine metabolism is associated with cancer. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.C.A.; Jiang, P.; Chan, C.W.M.; Sun, K.; Wong, J.; Hui, E.P.; Chan, S.L.; Chan, W.C.; Hui, D.S.C.; Ng, S.S.M.; et al. Noninvasive detection of cancer-associated genome-wide hypomethylation and copy number aberrations by plasma DNA bisulfite sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 18761–18768. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ashangari, C.; Suleman, A. Abstract 121: Vitamin D Deficiency Study in Postural Orthostatic Tachycardia Syndrome. Circ. Cardiovasc. Qual. Outcomes 2015, 8. [Google Scholar] [CrossRef]
Figure 1.
Recycling of Hcy in the methionine cycle is enabled by methyl THF, otherwise compromised by the MTHFR gene. Activated B2, B6, and B9 are required in the folate cycle. Folate is vitamin B9. PLP is B6 [3].
Figure 1.
Recycling of Hcy in the methionine cycle is enabled by methyl THF, otherwise compromised by the MTHFR gene. Activated B2, B6, and B9 are required in the folate cycle. Folate is vitamin B9. PLP is B6 [3].
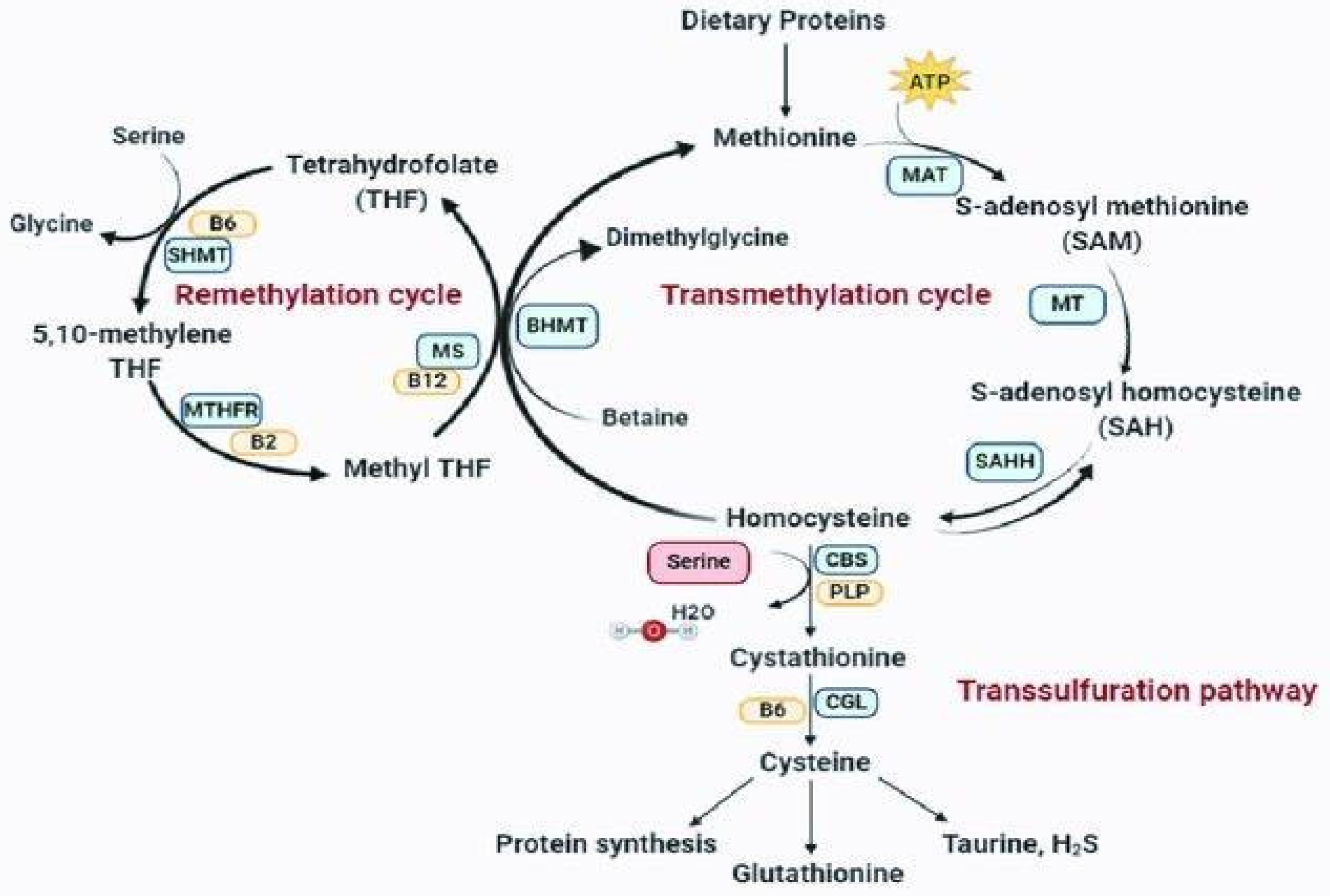
Figure 2.
Red structures designate CVOs [45]. The inferior area postrema is at the foramen of Magendie.
Figure 2.
Red structures designate CVOs [45]. The inferior area postrema is at the foramen of Magendie.
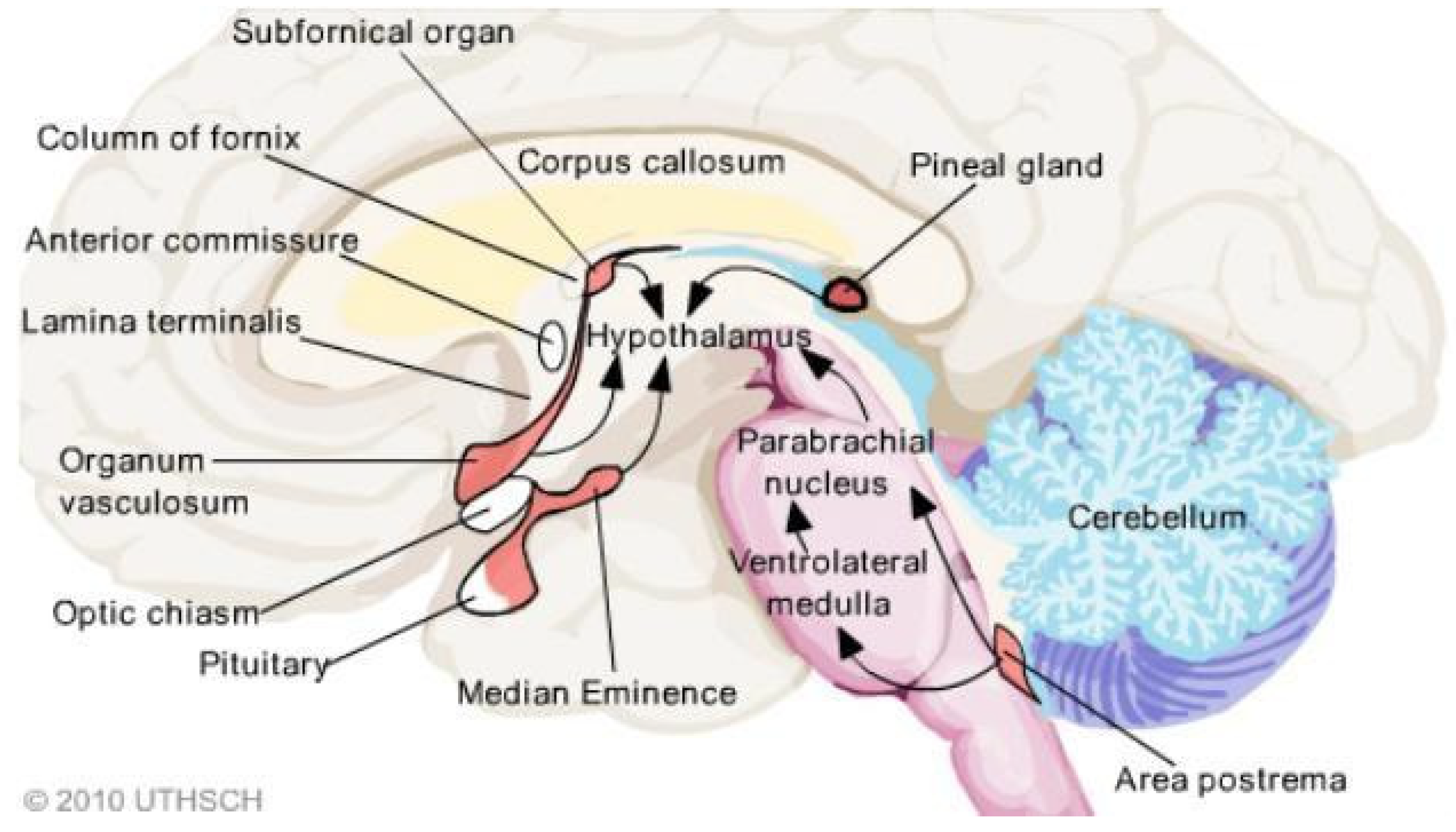
Figure 3.
The AP, the dorsal motor nucleus of X, and the nucleus of the solitary tract (NTS) form the dorsal vagal complex [46].
Figure 3.
The AP, the dorsal motor nucleus of X, and the nucleus of the solitary tract (NTS) form the dorsal vagal complex [46].
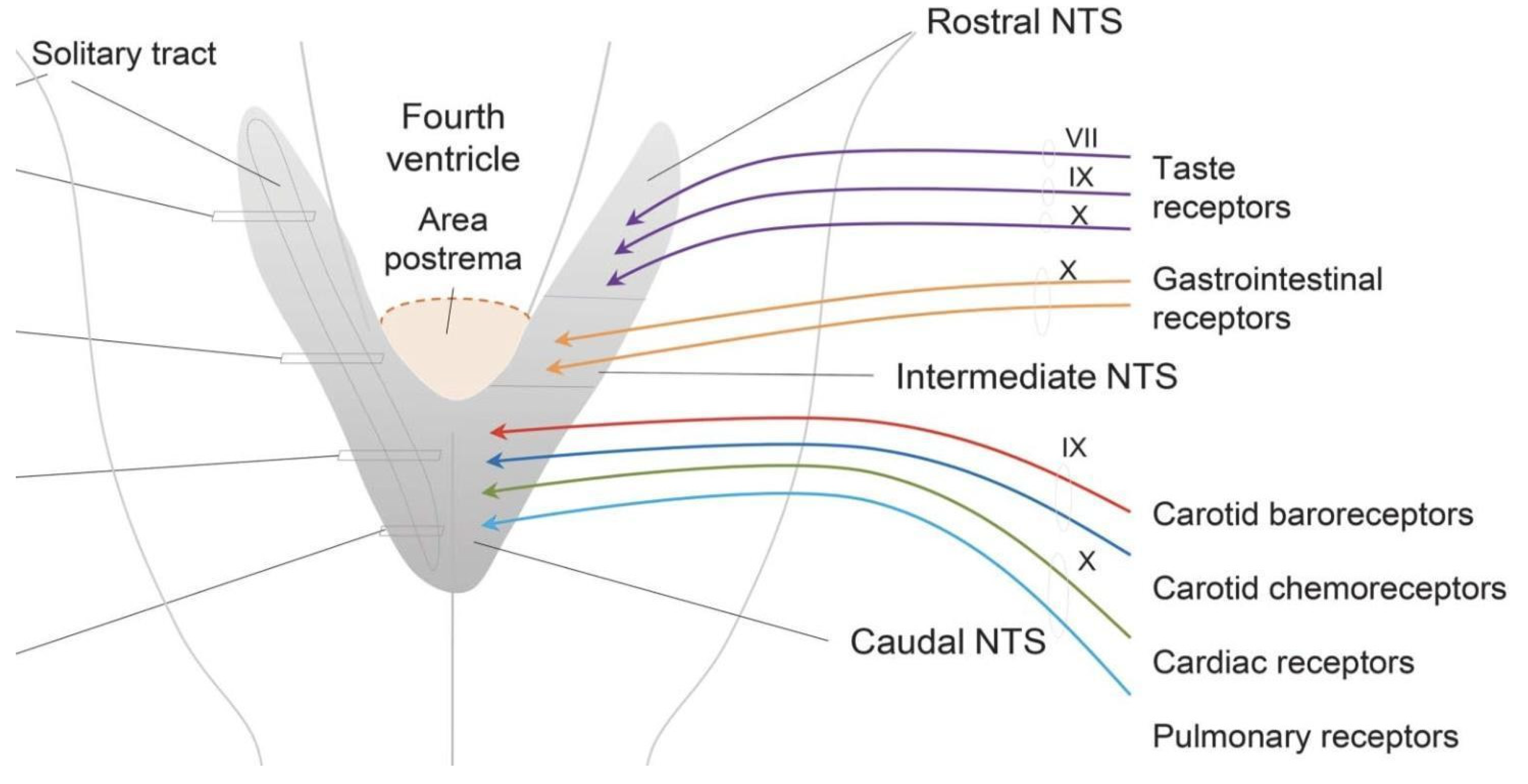
Figure 4.
Neuroanatomy and function of the PVN and posterior pituitary [50].
Figure 4.
Neuroanatomy and function of the PVN and posterior pituitary [50].
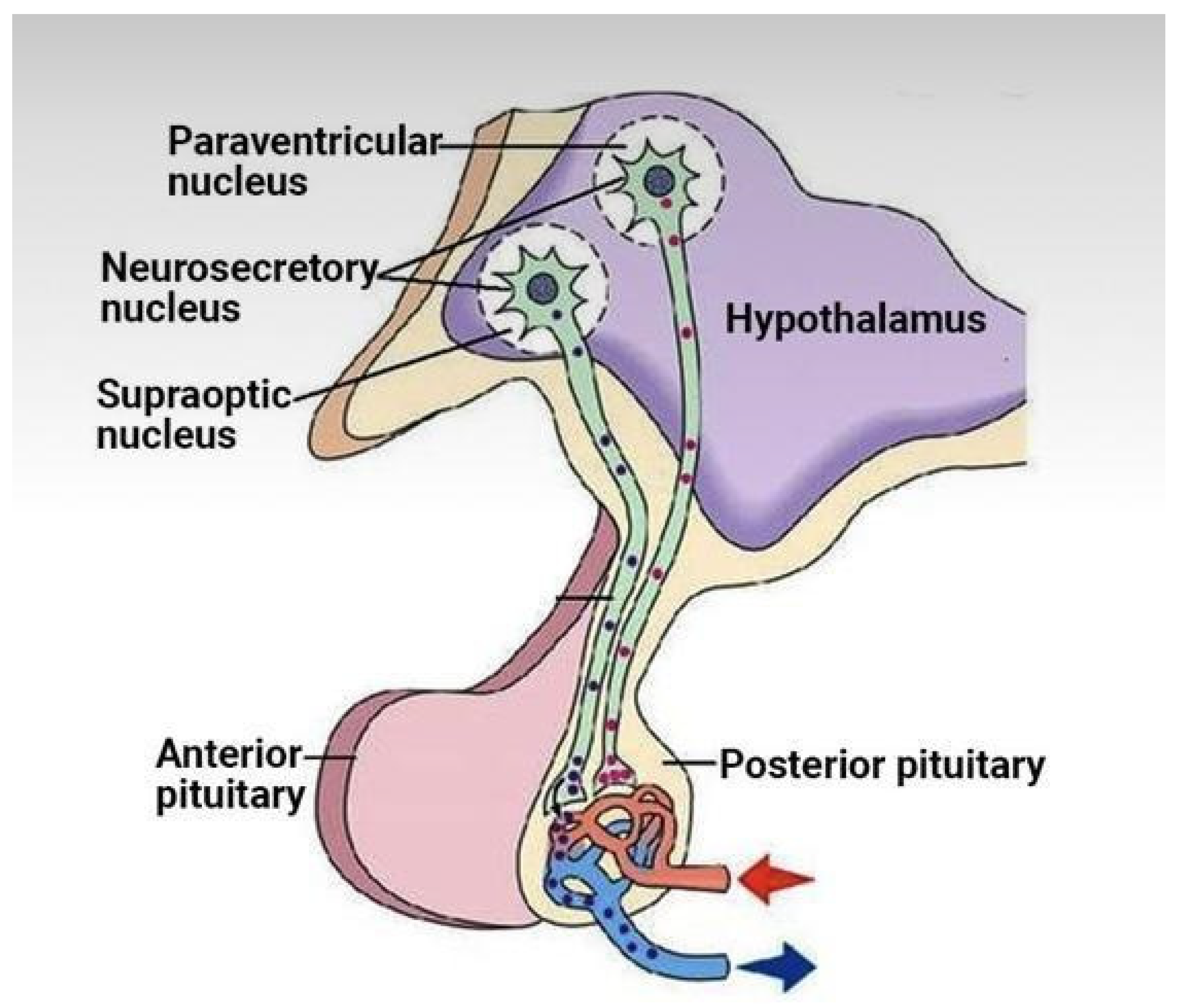
Figure 5.
ACE2 degrades des-argBKN (BK1R) and ACE degrades BKN (BK2R) => ACE and ACE2 are neuroprotective [74].
Figure 5.
ACE2 degrades des-argBKN (BK1R) and ACE degrades BKN (BK2R) => ACE and ACE2 are neuroprotective [74].
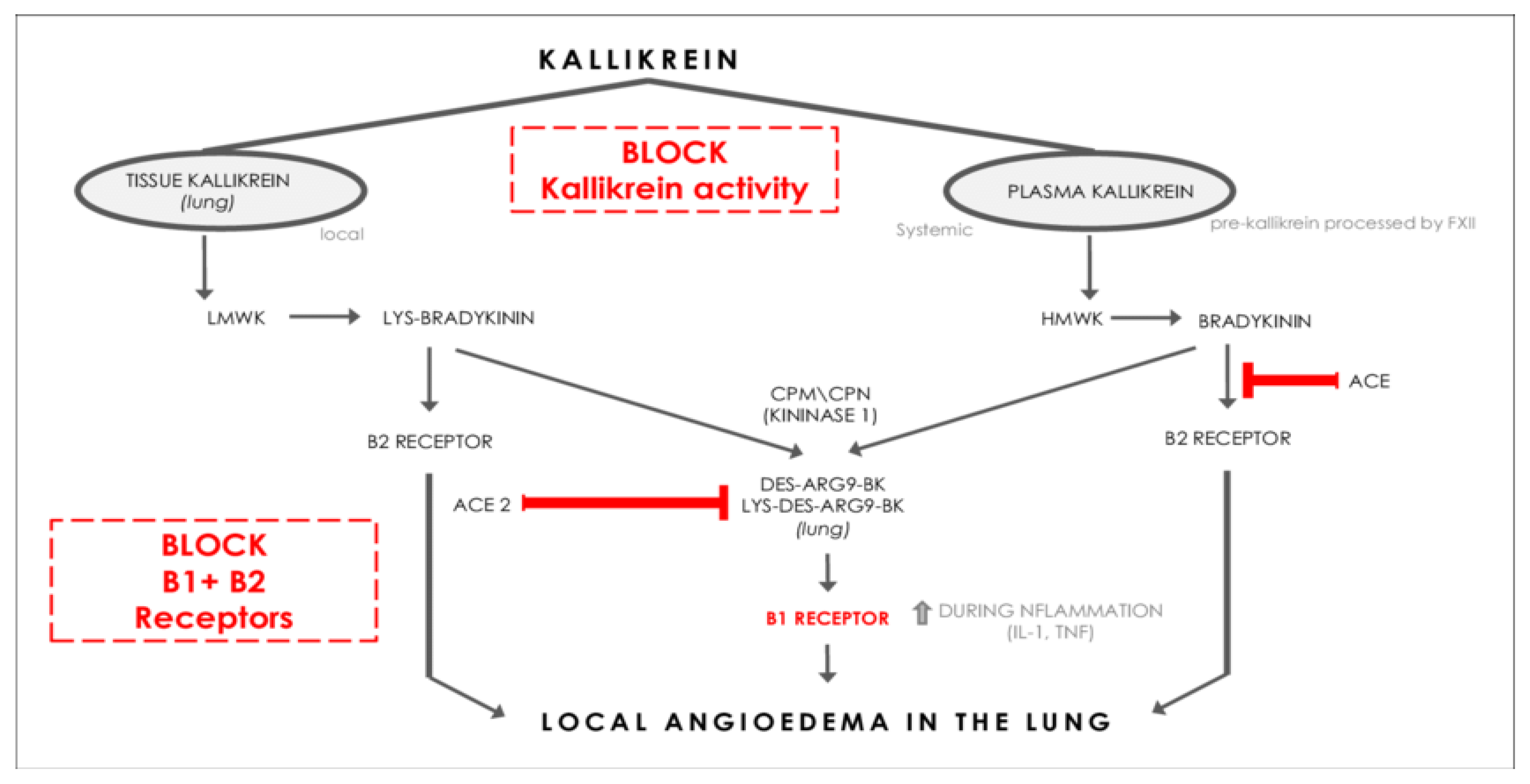
Figure 6.
Displayed are glutamatergic and GABAergic neurons. Note the magnitudes of the arrows [77]. Note the need for TCA recycling of GABA within astrocytes.
Figure 6.
Displayed are glutamatergic and GABAergic neurons. Note the magnitudes of the arrows [77]. Note the need for TCA recycling of GABA within astrocytes.
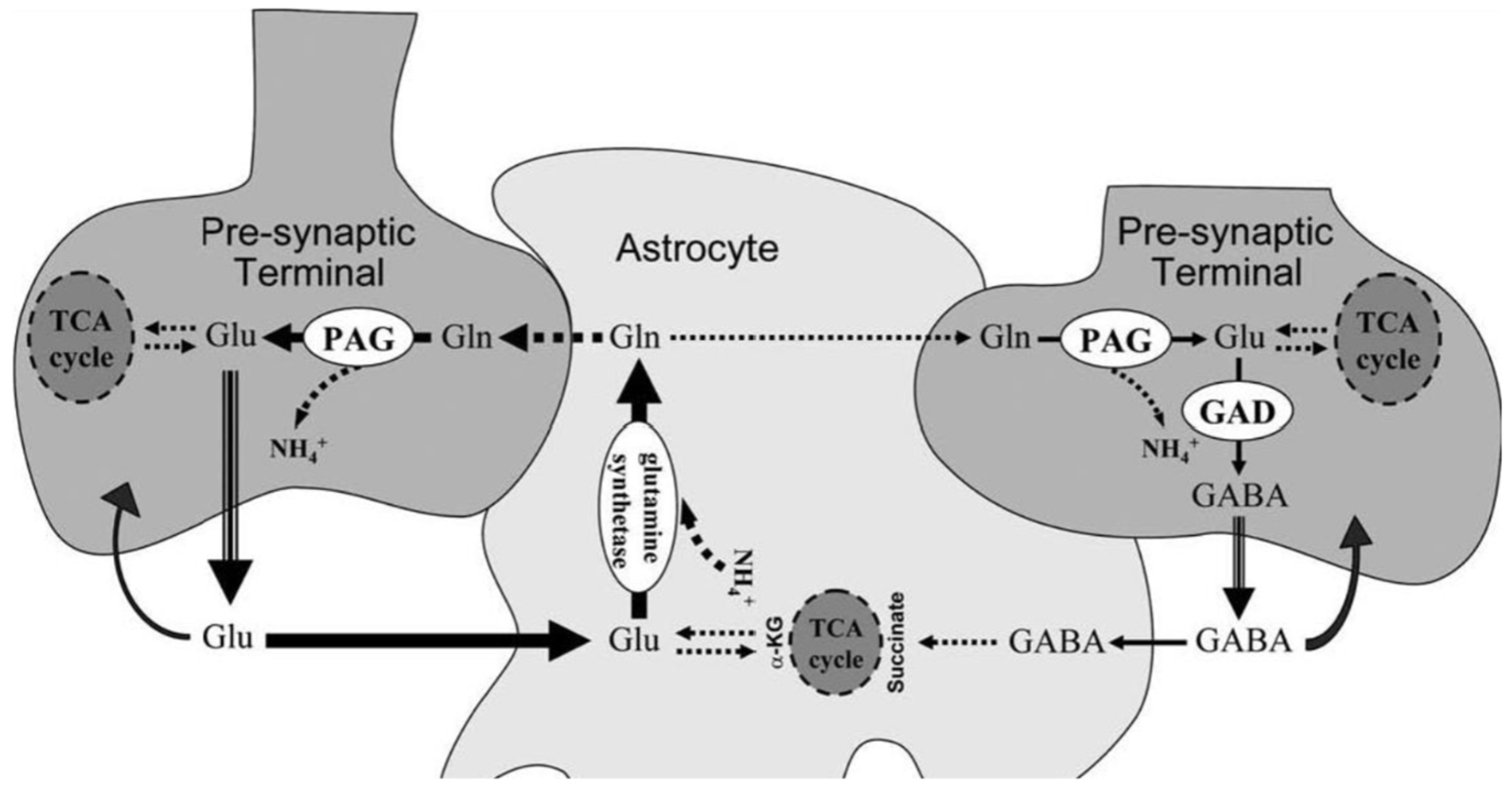
Figure 7.
Females less than 50, the vast majority of those with POTS, are more deficient in Mg++ than their male counterparts.
Figure 7.
Females less than 50, the vast majority of those with POTS, are more deficient in Mg++ than their male counterparts.
Figure 8.
Lung and breast cancer are increased in hyper- and hypomethylation[99].
Figure 8.
Lung and breast cancer are increased in hyper- and hypomethylation[99].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



