
Submitted:
14 September 2023
Posted:
15 September 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
These three syndromes - long covid (LC), postural orthostatic tachycardia syndrome (POTS), and myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) - have many symptoms in common. The common denominator remains elusive. The blood brain barrier (BBB) has been a barrier not only to microbes and toxins but also to understanding pathogenetic links. There are several areas within the brain that have no BBB. These are known as circumventricular organs (CVOs) and their location relative to CNS nuclei that direct autonomic and neuroendocrine functions is provocative in the quest for pathogenesis. In addition the majority afflicted with LC and ME/CFS appear to be those with two MTHFR polymorphisms, present in over 50% of Americans. These polymorphisms elevate homocysteine. When homocysteine is combined with CVOs, the fog of POTS and its paradox are lifted. POTS may represent the intersection of LC and ME/CFS in those with the MTHFR gene (hypermethylation or 677TT). The gut microbiomes of LC and ME/CFS, deficient in butyrates, GABA, and diversity, are then linked with MTHFR genotype 677TT. Reactivation of neurotropic EBV and VZV, due to loss of surveillance by CD4+/CD8+ T cells, is seen as secondary. The oxidative stress generated by homocysteine, loss of glutathione, low fiber diet, and persistent chronic inflammation exhaust available mitochondria and, assisted by BKN and estrogen, exacerbate all the elements of these post viral fatigue syndromes.
Keywords:
homocysteine
; MTHFR
; circumventricular organ
; magnesium
; vitamin D
; CD147
Introduction
COVID-19 has unleashed a tsunami of research that has been further energized by the persistent and debilitating aftermath of Long COVID. Explaining the gender reversal from male preponderance in COVID-19 to female in LC is challenging and covering the range of symptoms intimidating. Although there have been a few hypotheses incorporating autoantibodies, nutrient deficiencies, and reactivation of EBV, resolution of the paradoxes remains frustratingly out of reach. Recent research has revealed that the BBB is not all-inclusive and that some areas of the CNS are not so protected. These are called circumventricular organs (CVOs). Other research has revealed a close association between Long Covid and Hcy levels, often elevated in those with two very common enzymatic polymorphisms for MTHFR (Ponti et al., 2021). Most with these polymorphisms are unaware. But serum Hcy may play a large role in Long COVID and may provide unique insight into its pathogenesis. Receptor polymorphisms, enzyme polymorphisms, and the impact of methylation on suppressor and promoter genes, create a maze. A hypothesis incorporating the gut microbiome, gender predilection, and differential methylation is presented.
Hypothesis
The autonomic and neuroendocrine symptoms of LC, POTS, and CFS appear to be primarily determined by CNS nuclei located in areas lacking a BBB. Similar LC and CFS gut microbiomes incriminate low fiber diets that dictate low bacterial diversity. Dietary glutamate (Glu), an excitatory neurotransmitter, permeates CVOs and overwhelms inhibitory GABA neurotransmission, impeding release of specific hypothalamic hormones, including those causing low cortisol, low aldosterone, and low ADH with subsequent hypovolemia. The hypovolemia triggers an increase in norepinephrine. The baroreflex is also compromised. POTS is often associated with MCAS, EDS, and histamine intolerance. Metabolism of histamine requires methylation and intestinal bacteria are primarily responsible for this. All one carbon metabolism (methylation) reactions occur within mitochondria. Both hypomethylation (COVID-19, POTS, 1298C/677T, FM, and CFS) and hypermethylation (LC, 677TT) increase Hcy. The oxidative stress induced by Hcy due to 1) gut microbiome lacking diversity, 2) compromised methylation (genetic or epigenetic), 3) chronic inflammation elicited by residual spike protein S in LC, or 4) persistent SARS-CoV-2 induced T cell lymphopenia reactivating dormant viruses all conspire to overwhelm mitochondria lacking an antioxidant shield. Estrogen and BKN (and histamine) enhance endothelial permeability and may play prominent supporting roles.
Postural Orthostatic Tachycardia Syndrome
POTS represents a complex mixture of genetic and epigenetic inputs. It generally exhibits an 80%-90% female predominance, afflicts those 15-45, and is typically precipitated by a preceding viral infection (Fedorowski, 2018). In some POTS is felt due to a dysfunctional hypothalamus-pituitary-adrenal (HPA) axis that comes with a paradox – low renin and low aldosterone in the face of low blood volume (POTS paradox). This was originally reported in POTS with CFS and later in POTS with LC (Chadda et al., 2022). In addition to low renin and aldosterone CFS patients have lower levels of oxytocin, antidiuretic hormone (ADH), and corticotropin-releasing hormone (CRH) => less ACTH => less aldosterone, less cortisol in combination with less ADH => hypovolemia with reflexive increase in norepinephrine (Goldstein, 1996). POTS has often been partitioned into subsets, e.g., neuropathic, hyperadrenergic, low flow, high flow, and these can overlap. A dysfunctional HPA axis is felt causative for neuropathic POTS. Most POTS exhibit hypovolemia and low flow. Those with low flow POTS often report low plasma renin activity and elevated Ang II, resulting in vasoconstriction (Stewart et al., 2006). Cortisol and aldosterone are low in both LC (neuropathic) and CFS hyperadrenergic POTS (Okamoto et al., 2012). POTS is also directly related to Hcy levels, at least in the young (Li et al., 2018).
Many with low flow POTS have angiotensin II autoantibodies and/or beta adrenergic autoantibodies (Yu et al., 2018; Fedorowski et al., 2017; Badiudeen et al., 2019). In hyperadrenergic POTS autoantibodies accentuate expression (Wirth & Scheibenbogen, 2020). Angiotensin II antibodies have also been found in LC (Briquez et al., 2022). Angiotensin receptor blockers (ARBs) can block both autoantibodies (Yu et al., 2018; Barki-Harrington et al., 2003). Elevated Angiotensin II, due to autoantibodies and well described in low flow POTS, might provide negative feedback inhibition of renin.
High flow POTS may be due to vasodilation instead of vasoconstriction. Histamine dilates blood vessels and increases their permeability. POTS in MCAS and EDS may represent high flow type.
Elevated Hcy is prevalent in the MTHFR variants and may provide one link between some forms of POTS and some of these polymorphisms. Some MTHFR variants that yield hypomethylation appear to be predisposing factors for MCAS, providing another link to POTS. Whether these links are cause or effect is not yet clear. However, methylation is required to degrade histamine, suggesting causation.
Like autoantibodies, Hcy can also activate AT1Rs (angiotensin II type 1 receptors) (Li et al., 2018). The Hcy induced redox imbalance can compromise aldosterone synthase (low aldosterone) and 11beta hydroxylase (low cortisol), both mitochondrial based (Häggström & Richfield, 2014).
Some MTHFR variants with significant hypomethylation are theoretically capable of developing either low flow POTS or high flow MCAS type POTS. The former could be caused by elevated norepinephrine, as catechol-O-methyltransferase (COMT) degrades monoamines (Hall et al., 2019). The latter could be caused by elevated histamine, degradation of which is dependent upon histamine N-methyltransferase (HNMT). Both require S-adenosylmethionine (SAMe) and both reactions occur in mitochondria (Malecki et al., 2022).
Gut Microbiome
The neuropathic type of POTS seen in LC might be due to a CVO mediated Glu/GABA imbalance that leads to decreased ADH, oxytocin, and CRH. It is both a neuroendocrine and an autonomic disorder. LC and CFS are excellent models for the gut-brain axis. Gut microbiome studies of both LC (Yeoh et al., 2021) and CFS (König et al., 2021) report deficiencies in butyrate and GABA producing bacteria, e.g., Bacteroides (Strandwitz et al., 2018), Bifidobacterium dentium (Pokusaeva et al., 2017), and Lactobacillus brevium (Ko et al., 2013). Bacteroides not only drives gut microbiota diversity but also is one of only six known species that produces active B6 or pyridoxal-5-phosphate (P5P). B6 deficiency itself reduces the relative abundance of Bacteroides in the microbiota (Mayengbam et al., 2020). Without P5P glutathione cannot be synthesized from Hcy. A gut microbiome that lacks an abundance of histamine degrading Bifidobacteria leaves allergic features unopposed (Dev et al., 2008). This may contribute to MCAS. Furthermore, GABA secreted by histaminergic neurons also downregulates histamine signaling (Yu et al., 2015). The endogenous pathways for the degradation of histamine require either methylation (histamine-N-methyl transferase) or intestinal DAO. Both require P5P. A suboptimal gut microbiome might compromise the function of DAO. All methyltransferase reactions require SAMe, P5P and magnesium. Many such reactions require functioning mitochondria.
LC, CFS, and by extension the oft accompanying POTS may share gut microbiome features that underscore the criticality of bacterial GABA synthesis to proper functioning of the HPA axis, accentuated by CVOs.
GABA receptor polymorphisms are linked with restless legs syndrome (RLS) (Jiménez-Jiménez et al., 2018). RLS is a subset of periodic limb movement disorder (PLMD) aka nocturnal myoclonus. This is considered to be a seizure disorder and is increased in LC (Weinstock et al., 2022), CFS (Civelek et al., 2014), and POTS (Dodson, et al., 2021). RLS is also associated with magnesium deficiency, especially in those on proton pump inhibitors. GABA and magnesium display anti-seizure properties. This further underscores the probable role of a Glu/GABA imbalance in the pathogenesis of POTS, LC, and CFS. RLS may also be linked to SIBO (Blum et al., 2019), further evidence of the gut-brain axis at work (see Figure 1). RLS is associated with MCAS and histamine intolerance. EDS is also linked to MCAS. RLS is increased in POTS. Hypomethylation, GABA/P5P/Mg++ deficiency, and suboptimal gut microbiome might be at the root of all these syndromes.
Circumventricular Organs
Several recent articles have highlighted CVOs (Jammoul et al., 2023; Ganong, 2000). There are several areas within the brain that abut ventricles and have no BBB. Several of these CVOs contain nuclei that are quite relevant to the autonomic and neuroendocrine dysfunctions of LC. These CVOs are the pineal gland, the posterior pituitary a.k.a. neurohypothesis, the paraventricular nucleus (PVN) in the median eminence, the area postrema, the organum vasculosum of the lamina terminalis or supraoptic crest, and the subfornical organ (see Figure 2). The AP is part of the dorsal vagal complex, closely associated with baroreceptors and chemoreceptors, as well as cranial nerves VII and IX (see Figure 3).
CVOs enable diet induced excess of Glu to enter the CNS and create a Glu (excitatory)/GABA (inhibitory) imbalance to precipitate not only POTS but also symptoms that involve decreased oxytocin, vasopressin (ADH), CRH for ACTH, and gonadotropin releasing hormone (GnRH for LH, FSH).
CVOs can conveniently explain the POTS paradox in the hypovolemic low flow subtype (majority). The PVN normally inhibits the baroreflex via tonic GABAergic inputs (Page et al., 2011). Just as sympathetic inhibition and parasympathetic activation respond to carotid and aortic stretch receptors to decrease BP, so sympathetic activation and parasympathetic inhibition are required to increase it upon standing. An imbalanced Glu (elevated)/GABA(depressed) impedes both, causing postural hypotension. GABAergic signals also positively dictate secretion of neurohypophyseal hormones, including ADH (Haam et al., 2012), oxytocin (Lee et al., 2015), CRH (Kakizawa et al., 2016) and GnRH (Moore et al., 2018). On the other hand GABA inhibits the secretion of TRH (Wiens & Trudeau, 2006)
CFS patients have experienced relief from desmopressin (Miwa, 2017) and CRH (Pereira et al., 2021). POTS and CFS patients have benefited from exogenous CRH and ADH (Coffin et al., 2012). Oxytocin in CFS increased stamina, decreased pain, improved cognitive function, decreased FM pain, anxiety, and depression (Diep et al., 2022). Some LC patients have experienced relief with oxytocin (Goldstein, 2012). Many POTS patients claim heat intolerance and this may be mediated by unopposed Glu (Amir, 1990).
MTHFR and Homocysteine
Polymorphisms in the MTHFR gene markedly compromise conversion of 5,10 -methylenetetrahydrofolate to 5-methyltetrahydrofolate. The 1298C/677T (compound heterozygous) and 677TT MTHFR variants in order pose escalating bottlenecks, 35% and 70% respectively, in recycling Hcy to methionine (Frosst et al., 1995). MTHFR is critical to methylation. A fully functional MTHFR with the assistance of B2, B6, and B12 (methylcyanocobalamin) helps methylate Hcy, recycling methionine (see Figure 4). If the active form of B6 (P5P) is insufficient, then glutathione, the master antioxidant, will not be produced with additional buildup in Hcy. ROS, induced by Hcy, will increase and mitochondrial function is compromised, especially in endothelial cells.
The MTHFR 677T and 1298C alleles (variants) vary ethnically, but are present in about 33% and 38% of Caucasians respectively (Nefic et al., 2018). However, according to the CDC, there are more people in the United States who have one or two copies of the MTHFR C677T variant than people who do not. Perhaps 10-20% of the American population have 677TT. So this is an exceedingly common polymorphism with incidence that roughly somewhat reflects the incidence of LC.
Hcy is a strong marker for LC (Ponti et al., 2021), MTHFR variants (Carpenè et al., 2022), and for CFS (Regland et al., 1997). Both histamine and Hcy require methylation for their metabolism, which suggests that hypomethylation may cause histamine intolerance in MCAS and EDS. High Hcy may be a good marker for MTHFR variants and low folate a good inverse marker for Hcy (Bottiglieri et al., 2000). Hcy is a strong marker for thrombosis. CNS Hcy is associated with brain fog that may be mediated by CVOs (Oner et al., 2023).
Compound heterozygous MTHFR gene variants (677T/1298C) characteristically exhibit hypomethylation (Ren et al., 2018). In a recent review compared to controls, patients that experienced mild/moderate COVID-19 or LC exhibited hypomethylation and hypermethylation respectively (Nikesjö et al., 2022). This reflects the hypomethylation disposition of MTHFR 1298C/677CT (slightly elevated Hcy) and the hypermethylation disposition of MTHFR 677TT (more pronounced elevation of Hcy). Hcy associated with the 677TT variant appears to trigger hypermethylation via an alternative B2 dependent “rescue” pathway (Fryar-Williams, 2016).
Hypermethylation of the promoter region for the MTHFR gene silences it, further compromising transmethylation (Shaker et al., 2021). Those with 677TT usually need higher doses of 5-methyltetrahydrofolate (methylfolate). Methylation status in the body can be determined by measuring whole blood histamine, as histamine and methylation are inversely related. Low blood histamine levels indicate overmethylation; high levels indicate undermethylation (Walsh, 2012) 61. Histamine is predominantly metabolized via methylation and is upregulated by estrogen (Bonds & Midoro-Horiuti, 2013).
This suggests that mild to moderate Covid 19 might include many with hypomethylated 1298C/677CT and LC might include many with hypermethylated 677TT. Epigenetic hypermethylation can persist for one year after hospitalization (Balnis et al., 2022). The hypermethylated have higher Hcy levels, which portend more serious COVID-19 and LC (Kalan Sari et al., 2023). Indeed those without the MTHFR variants might comprise a large portion of the asymptomatic.
Hcy may be the primary common denominator linking LC, CFS, POTS, and MTHFR variants. Hcy is increased in LC, CFS, and MTHFR 1298C/677T and 677TT. Some with low flow POTS exhibit oxidation of the vasodilator NO with microvascular endothelial dysfunction (Medow et al., 2005). Hcy promotes oxidant injury to endothelial cells and microthrombosis, prominent features of LC and CFS. Hcy is directly related to oxidative stress and inversely reflects B vitamin sufficiency, especially that of B6. Two steps in the metabolism of Hcy to glutathione, the master antioxidant, require B6 (see Figure 4).
A suboptimal diet and gut microbiome (low microbiota diversity) might compromise absorption of exogenous antioxidants, enhancing the pro-oxidative damage wreaked by Hcy. Many ROS producing mitochondria are shut down to enable cell survival and avoid lysis. Most endogenous antioxidants require methylation and SAMe, the universal methyl donor, i.e., cofactor for all methyltransferases. All SAMe methylations require ATP and occur in mitochondria (Malecki et al., 2022; Rhein et al., 2014). Mg++ is a required chelate for all reactions requiring ATP and a required cofactor for all SAMe reactions. Methylation is also critical to the degradation of neurotransmitters and hormones, including serotonin, dopamine, epinephrine, norepinephrine and melatonin. A suboptimal gut microbiome in combination with mitochondrial dysfunction retards synthesis of 1,25(OH)2D3.
Ultimately LC, CFS, and low flow POTS seem to stem from antioxidant insufficiency (redox imbalance) and mitochondrial dysfunction. Elevated Hcy of endogenous origin, whether genetic (MTHFR) or epigenetic (SAMe related), determined in part by nutrition, assisted by CVO/BKN induced permeability, and characterized by altered 1) mitochondrial function, 2) neurotransmission, 3) enzyme activity, may represent the final common pathway. Elevated histamine and BKN in females also appear to be major players in the hypomethylated. MTHFR variants with differential methylation may be more susceptible to some, e.g., CFS (de Vega et al., 2014), or all of these.
As a corollary, chronic inflammation and age increase differentially methylated DNA and subsequent cancer risk, especially breast and colorectal cancers (Benelli et al., 2021). An elevated Hcy is associated with cardiovascular disease, depression, dementia, AD, and cancer (McKinney et al., 2019).
Female Preponderance
Females exhibit more robust T cell activation than males (Takahashi et al., 2020). Females have higher levels of type I IFN alpha and beta (Pujantell & Altfeld, 2022). T cell production of IFN gamma (IFN type II) is triggered by IFN alpha and beta. It is a secondary release, STAT dependent IFN (Chambers, 2022). T cell population decreases with age in males but not females. T cell production of IFN-γ persists in females (Goetzl et al., 2010). Increased autoimmune disease in females may be due to their enhanced innate immunity and production of pleiotropic IFN-γ, secreted by CD4+ and CD8+ T cells. When combined with chronic low grade inflammation, this proinflammatory pleiotropic cytokine might switch from self recognition to loss of self-recognition or autoimmune status (De Benedetti et al., 2021).
IFN-γ produced by these CD4+ and CD8+ T cells also induces hepatic synthesis of C1 esterase inhibitors (C1INH). Loss of complement inhibition initiates the classic complement pathway, which cross talks with the kallikrein kinin system (KKS). BKN is then also upregulated (Bossi, 2011). Lymphopenia (primarily of T cells) is associated with persistence of COVID-19 symptoms. CD147 (basigin) receptors but not ACE2 receptors are found on peripheral blood mononuclear cells (Kuklina, 2022). This receptor when bound to the CD147 epitope on the spike protein S (Chambers, 2021) may trigger the complement cascade and mediate lymphopenia. Any viral infection that challenges T lymphocytes increases BKN.
Estrogen may play a role in low flow POTS by decreasing renin levels, decreasing ACE activity and thereby increasing BKN, decreasing AT1 receptor density and decreasing aldosterone (Ma et al., 2021). In addition estrogen appears to lower cortisol (Herrera et al., 2017) and ACE2, at least in the lungs (Stelzig et al., 2020). Perhaps lower pulmonary ACE2 is Covid 19 protective, but downregulated ACE2 may be associated with higher systemic levels of des-Arg9-bradykinin (see Figure 5). Des-arg-BKN appears to be a prominent kinin in COVID-19 (Mendes et al., 2022).
Additionally vascular dilation and leakage induced by BKN might aggravate hypovolemia in high flow POTS, e.g., POTS in MCAS. A preceding viral respiratory infection markedly upregulates BKN (Stewart et al., 2008). Down regulation of ACE (ACE degrades BKN) by estrogen combined with a viral infection can supercharge BKN. BKN indirectly inhibits renin to a greater than RAS directly inhibits renin (Kiowski et al., 1992). This might in part explain the decreased renin in predominantly female POTS and its paradox. An increased Ca/Mg might also impact this, as Mg++ is required to synthesize cAMP and cGMP that mediate the release of renin from the juxtaglomerular apparatus (Persson, 2003).
Upregulation of BKN and histamine by estrogen in combination with hypomethylation (mild increase in homocysteine) may be linked to the pathogenesis of high flow in female dominated POTS, SIBO, MCAS, and EDS.
A more significant increase in Hcy might elicit hypermethylation by an alternative “rescue” pathway (Fryar-Wiliams, 2016). The associated endothelial redox imbalance diminishes arterial sensitivity to norepinephrine, possibly driving a reflexive increase, as seen in CFS with hyperadrenergic POTS.
Vitamin D and Oxidative Stress in Mitochondria
Men with the highest compared to lowest 1,25(OH)2D and activation ratios, i.e., active form/storage form (1,25(OH)2D/25(OH)D), are more likely to possess butyrate-producing bacteria associated with favorable gut microbial health. Sun exposure may affect the storage form of vitamin D, but it exerts little influence on the active hormone (Thomas et al., 2020). However, endothelium, targeted by Hcy, may benefit from D3 alone (Gibson et al., 2015).
Mitochondrial health is especially important in maintaining optimal levels of active vitamin D. Oxidative stress (ROS due to inflammation, smoking, toxins, …) compromises mitochondrial oxidative phosphorylation. C-reactive protein reflects oxidative stress (Moreira-Rosário et al., 2021) and is a marker for LC, CFS, and vitamin D deficiency. Mitochondrial CYP27B1 is the only enzyme (requires Mg++) that hydroxylates carbon number one in cholecalciferol, activating 25(OH)D3 (Bikle, 2014; Warren et al., 2021).
Once chronic inflammation becomes entrenched and mitochondria oxidatively stressed (less CYP27B1) exogenous 1,25(OH)2D3 is needed.
Most with POTS have 25(OH)D3 levels below 20 nm/mL (Ashangari & Suleman, 2015). There is a seasonal variation to the gut microbiome, as there is to 25(OH)D3. Viral respiratory illness increases in the winter and induces robust depletion of the gut microbiome (Yildiz et al., 2018). Although the gut microbiota in females is larger and more diverse than that in males, the greater incidence of URIs in females (Groeneveld et al., 2020) with concomitant microbiota depletion may explain the female gender disparity in these syndromes. A suboptimal diet, less sun exposure, and greater risk of a viral respiratory illness may conspire to produce this suboptimal gut microbiome (Al-Khaldy et al., 2023; Waterhouse et al., 2019; Singh et al., 2020), enhancing susceptibility to not only LC and CFS but also post viral fatigue syndrome in general, POTS (Blitshteyn, 2017), SIBO, MCAS, EDS, and some MTHFR variants (Okamot et al, 2012).
Antioxidants in Post Viral Fatigue Syndrome
The symptom list for CFS is long and recapitulates many of the symptoms of LC, FM, CMV, and EBV. Indeed these are all probably best classified under the umbrella of post viral fatigue syndrome. There are numerous articles and at least one book that extoll the value of antioxidants aka mitochondrial optimizers or enhancers in the treatment of CFS98. Many that develop CFS can retrospectively recall an episode of viral illness that preceded the onset of initial symptoms. Like those with POTS, many with CFS and LC are vitamin D deficient.
Low gut microbiota diversity is associated with CFS, LC, and deficiencies of vitamin D (Thomas et al., 2020), magnesium, zinc, selenium, and other micronutrients. Vitamin D deficiency is often accompanied by magnesium deficiency, as synthesis of the active form of vitamin D from cholesterol requires the cofactor Mg++ in half a dozen enzymatic reactions. B2 and Mg++ are required cofactors for the synthesis of 7-dehydrocholesterol from which D3 and cholesterol are produced.
Synthesis of aldosterone requires aldosterone synthase and synthesis of cortisol requires 11beta hydroxylase. These two enzymes are part of the CYP450 system, all of which require Mg++. Furthermore these two reactions occur in the mitochondria, i.e., they also require Mg++ chelated to ATP. So magnesium deficiency might also contribute to the low aldosterone and cortisol in the POTS paradox. If oxidative stress is sufficient and antioxidants are not, mitochondria are shut down to avoid cell lysis and ATP is down regulated, limiting synthesis of aldosterone and cortisol (Atanassova & Koeva, 2012). Females tend to be more magnesium deficient than males before age 50 (see Figure 6). An elevated Ca:Mg may also compromise synthesis of aldosterone and cortisol (Kiowski, 1992; Persson, 2003), as calcium and magnesium both compete for the same receptor, the calcium sensing receptor.
Magnesium and the B Vitamins
Magnesium is also required for the synthesis of all B vitamins, except biotin (B7). Mitochondrial function is dependent upon adequacy of all the B vitamins (Depeint et al., 2006). If mitochondrial function is marginal, production of the active form of vitamin D may be marginal. Mitochondrial CYP27B1 requires B2 and Mg++ as cofactors.
Alcohol intolerance is a primary complaint in LC (alcohol tastes different), CFS, and MCAS. Hepatic metabolism of alcohol yields acetaldehyde, but further degradation in the oxidatively stressed mitochondria is compromised with symptoms ranging from headache to hangover, both acetaldehyde induced.
Thiamine (B1), riboflavin (B2), niacin (B3), pantothenate (B5), and pyridoxine (B6) require phosphorylation for activation. Cyanocobalamin (B12) and folate (B9) require methylation (SAMe) for activation. Mg++ is a required chelate for phosphorylation and a required cofactor for methylation. Hcy is highly influenced by B vitamin status (Azzini et al., 2020).
The last enzymatic steps to produce serotonin, dopamine, and GABA are decarboxylases that require active B6 (P5P), as a cofactor. Melatonin and glutathione synthesis also require B6. Norepinephrine synthesis requires Mg++. A shortfall in magnesium or B6 compromises monoamine neurotransmitter synthesis and leads to depression, a symptom shared by LC, POTS, CFS, MCAS, and EDS.
Genetic (MTHFR) and/or epigenetic hypomethylation appears to link CFS, MCAS, POTS, and EDS. All syndromes predominantly affect females and all may in part be due to inability to methylate B9 and B12. Indeed P5P is required to synthesize P5P from pyridoxine. Histamine is degraded either via the methyltransferase pathway or via DAO. Both pathways require P5P. All methyltransferases require Mg++ as a cofactor (Doctor’s Data, Inc). Furthermore, estrogen upregulates histamine and vice versa (Bonds & Midoro-Horiuti, 2013). Histamine also upregulates NE.
In a clinical study of 33 POTS patients with GI symptoms 27 had SIBO, 9 had MCAS and 14 had EDS (Weinstock et al., 2018). SIBO has been linked to EDS (Uy et al., 2021) and is more common in females (Rao & Bhagatwala, 2019). Gut dysbiosis, including SIBO, is associated with MCAS due to diamine oxidase (DAO) deficiency (Sánchez-Pérez et al., 2022).
Histamine is processed primarily in the intestines by methyltransferase or DAO. An April 2023 review suggested that MTHFR polymorphisms play a large role in EDS (Courseault et al., 2023).
High doses of pyridoxine are associated with peripheral neuropathy, whereas high doses of the active form, pyridoxal-5-phosphate, are not (Hadtstein & Vrolijk, 2021). Unfortunately a nutritious diet is becoming less available due to soil mineral depletion, enhancing the need for supplementation.
Thoughts on Therapy
Adding some fiber (prebiotic), e.g., sauerkraut, kimchi, with judicious supplementation, especially magnesium and D3, seems a prudent first step to cultivate good gut microbiota, e.g., bifidobacteria, lactobacillus. Probiotics or live bacteria in a pill also help in the search for the optimal gut microbiome. The heavy emphasis of dairy in the Western diet has resulted in an ever increasing Ca/Mg ratio. The 1977 NHANES (National Health and Nutrition Examination Survey) data indicated a ratio of 2.6. This ratio first exceeded 3.0 in the 2000 survey. When elevated, sustaining a 25(OH)D3 level of 50 ng/mL (requires about 4000 IUs/day) becomes difficult, as these divalent cations compete for the same receptor. All post viral fatigue syndromes are due to ROS induced loss of mitochondria due to a shortage of onboard antioxidants. Furthermore, any antioxidant or mitochondrial shortfall impedes activation to 1,25(OH)2D3. Selenium, zinc, vitamin C, all fat soluble vitamins (A, D, E, K) are excellent antioxidants (Chambers, 2022).
Methylated forms of B9 and B12 are vital, if an MTHFR variant and/or elevated Hcy is discovered. Any tolerable (avoid laxative effect) increase in magnesium intake is especially recommended. If truly desperate, an ARB, under supervision, might be worth exploring.
The potential benefits of ARBs for LC include:
- 1. ARBs upregulate GABA, which displays anti-hypertension, anti-senescence, anti-diabetes, antioxidant, and anti-inflammatory properties (Sfera et al., 2022).
- 2. ARBs increase ACE, which degrades BKN, but block AT1Rs and hypertension induced dementia (Campbell et al., 1995)
- 3. ARBs alleviate POTS (Medow et al., 2005) and degrade BKN otherwise associated with AD (Singh et al., 2020).
- 4. ARBs increase ACE2 (Zaheer et al., 2021) that degrades amyloid β-peptide (Aβ) in AD (Kehoe et al., 2016). The benefits of ACE2 in COVID-19 exceed its risks as a receptor for SARS-CoV-2 (Bastolla et al., 2022).
- 5. ARBs down regulate pro-inflammatory transforming growth factor-beta (TGF-β), increased in LC (Mahudpour et al., 2020), CFS (Yang et al., 2019), AD (Zhang et al., 2016), Marfan Syndrome, and EDS.
- 6. ARBs improve insulin sensitivity (Zhang et al., 2013) and are antidiabetic (Chang et al., 2014).
- 7. ARBs are neuroprotective (Villapol & Saavedra, 2015).
Indeed, a post mortem study of brains demonstrated more neuropathology in non-hypertensives, than in hypertensives on ARBs (Hoffman et al., 2009). Furthermore, angiotensin II–stimulating antihypertensives, e.g., ARBs, exhibited lower dementia risk than angiotensin II inhibiting antihypertensives, e.g., ACE inhibitors (van Dalen et al., 2021).
Conclusion
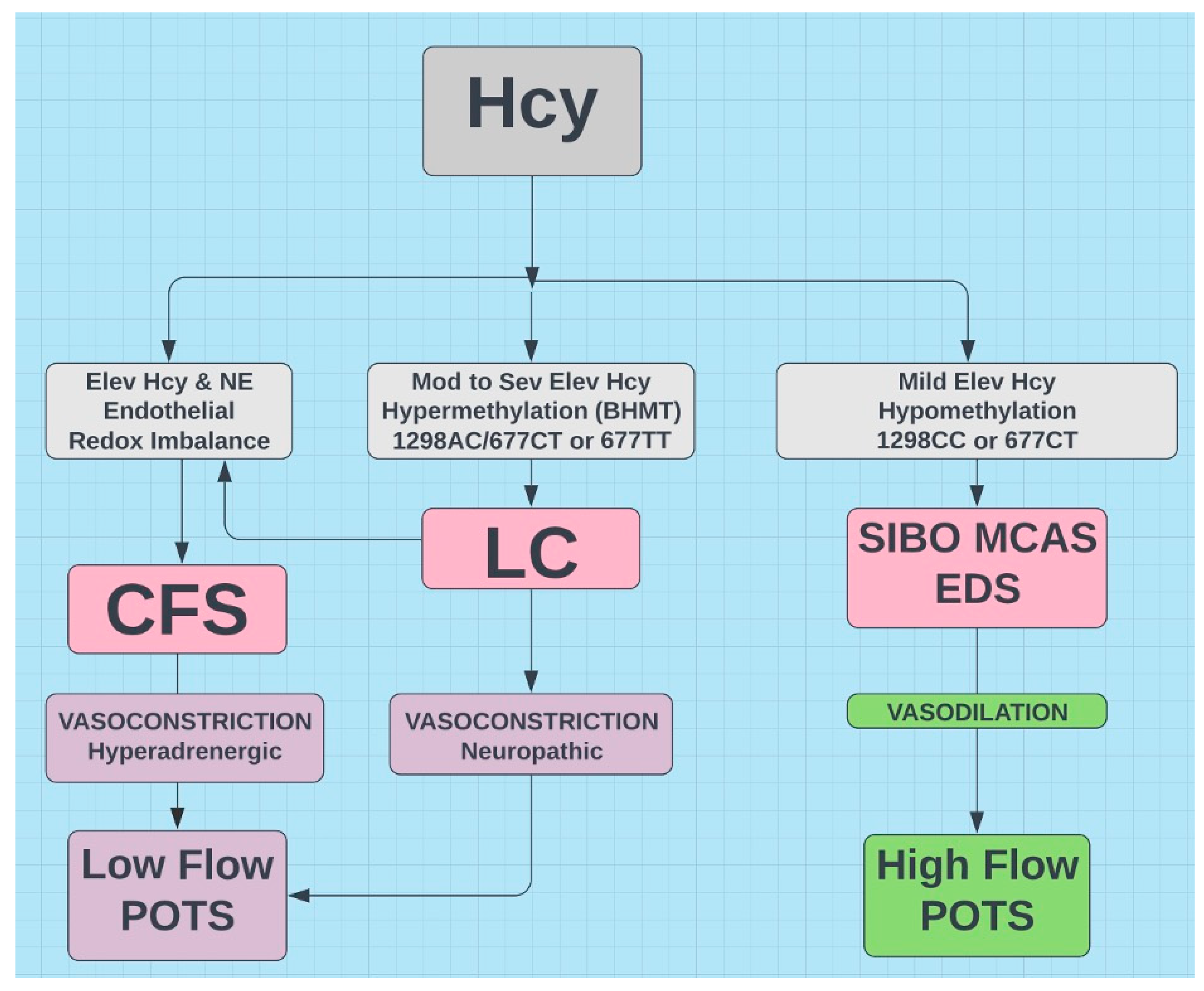
Three hypothetical conclusions involving the gut microbiome, female gender propensity, and transmethylation appear to link LC, POTS, CFS, and some MTHFR variants. Hcy appears to play a vital role (see Figure 7) and its elevation in some MTHFR variants suggests increased risk.
- Lack of gut microbiota diversity and elevated C-reactive protein, indicative of chronic information, are independent prognostic biomarkers for LC, CFS, and vitamin D deficiency (Moreira-Rosário et al., 2021). MTHFR mutations do not directly compromise gut microbiota diversity. The absence of the BBB in CVOs with nuclei that control baroreceptor function (area postrema and PVN) and neuroendocrine function (PVN), combined with increased Glu, histamine and decreased GABA producing bacteria, may in part explain pathogenesis of the myriad symptoms in these syndromes. Glu permeates CVOs and may overwhelm GABA neurotransmission. This suboptimal microbiome aggravates symptoms in those with the MTHFR 677T genotype (Regland et al., 2015; Ponti et al., 2021) and is seen in the vitamin D deficient (Thomas et al., 2020). Most with POTS have low 25(OH)D3 levels (Ashangari & Suoleman, 2015). Seasonal variation in the gut microbiome (Machado, 2023) may reflect the cold and flu season due to low vitamin D. These URIs cause and are caused by a sub optimal gut microbiome (Yildiz et al., 2018; Al-Khaldy et al., 2023; Waterhouse et al., 2019; Singh et al., 2020), increasing susceptibility to post viral fatigue syndrome, LC, CFS, POTS (Blitshteyn et alo., 2017), and some MTHFR variants (Okamoto et al., 2012). Unfortunately once chronic inflammation becomes entrenched and mitochondria oxidatively stressed (less CYP27B1) exogenous 1,25(OH)2D3 is needed (Warren et al., 2021). SIBO is an issue separate from gut microbiome diversity. Colon bacteria gain a foothold in the small bowel. This is associated with low volume high flow POTS seen in MCAS and EDS with histamine intolerance.
- Females in their reproductive years appear to be more susceptible to LC. This group is also more likely to be magnesium deficient (see Figure 7), probably due to the Western diet, high in calcium and low in magnesium. Estrogen, a mild ACE inhibitor, promotes BKN, linked to brain fog, cognitive decline, and AD. Estrogen also upregulates IFN-γ, produced by CD4+ and CD8+ T cells. Decreased IFN-γ translates to decreased C1INH and activation of the classic complement pathway and the KKS. Respiratory viral infections, increased in females, induce additional release of BKN (Stewart et al., 2008) 82. Estrogen induced upregulation of histamine (degradation requires transmethylation) enhances endothelial permeability and MCAS type low volume high flow POTS. The “post viral fatigue syndrome” in general may stem from a discriminating viral assault on T cells or from a URI induced depletion of gut microbiota. The loss of IFN-γ and the presence of estrogen potentiate chronic inflammation and endothelial permeability. Subsequent chronic inflammation induced oxidative stress from Hcy, residual spike protein S, and reactivated viruses conspire to overwhelm those with insufficient onboard antioxidants. In the absence of critical cofactors synthesis of the master antioxidant glutathione from Hcy cannot proceed (see Figure 5). This downregulates mitochondrial activity and upregulates fatigue (Wood et al., 2021). This also compromises the synthesis of 1,25(OH)2D3, the active form of vitamin D, and with it immune function.
- In the presented hypothesis protein methylation plays a prominent role. It is at the center of the MTHFR gene variants. It manifests in LC (Nikesjö et al., 2022) and POTS (Li et al., 2018) high flow type. MTHFR status is easily available via 23andme or a blood test for Hcy. However, given the prevalence of the MTHFR variants in the general population and from which the laboratory reference values for Hcy are determined, any value near the upper limit of normal may be cause for concern. Elevated Hcy carries many short term (thrombosis) and long-term (neurodegenerative disease) problems, best addressed in a timely manner. Mild elevation of Hcy is associated with hypomethylation. Significantly elevated Hcy associated with the 677TT variant appears to trigger hypermethylation via an alternative B2 dependent “rescue” pathway (Fryar-Williams, 2016). Another protein impacted by hypomethylation is histamine linked to SIBO. DNA can also be differentially methylated (hypo and hyper methylated regions) and is associated with chronic inflammation, cancer, and increasing age. Given the high frequency of this polymorphism in the general population and its potential devastating effects, everyone should be privy to their risks and those of their offspring, especially in the post Covid era.
Abbreviation
| ADH | Antidiuretic hormone (vasopressin) |
| ARBs | Angiotensin receptor blockers |
| BBB | Blood brain barrier |
| BKN | Bradykinin |
| CFS | Chronic Fatigue Syndrome |
| COMT | Catechol-O-methyltransferase |
| CRH | Corticotropin-releasing hormone |
| CVOs | Circumventricular organs |
| DAO | Diamine oxidase |
| EDS | Ehlers Danlos syndrome |
| FB | Fibromyalgia |
| Glu | Glutamate |
| GnRH | Gonadotropin releasing hormone |
| Hcy | Homocysteine |
| HNMT | Histamine N-methyltransferase |
| HPA | Hypothalamus-pituitary-adrenal |
| LC | Long COVID |
| MCAS | Mast cell activation syndrome |
| MTHFR | Methylenetetrahydrofolate reductase |
| P5P | Pyridoxal-5-phosphate |
| POTS | Postural orthostatic tachycardia syndrome |
| PVN | Paraventricular nucleus |
| RLS | Restless legs syndrome |
| SAMe | S-adenosylmethionine |
| SIBO | Small intestine bacterial overgrowth |
Acknowledgements
The author would like to thank Dr. Thomas-Gabriel Schreiner and Dr. Camila Malta Romano for reviewing this article.
Disclosures
A preliminary version of this manuscript was published in the Journal of Neurology and Neurophysiology.
References
- Regland, B.; Forsmark, S.; Halaouate, L.; Matousek, M.; Peilot, B.; Zachrisson, O.; Gottfries, C.-G. Response to Vitamin B12 and Folic Acid in Myalgic Encephalomyelitis and Fibromyalgia. PLOS ONE 2015, 10, e0124648. [Google Scholar] [CrossRef]
- Ponti, G.; Pastorino, L.; Manfredini, M.; Ozben, T.; Oliva, G.; Kaleci, S.; Iannella, R.; Tomasi, A. COVID-19 spreading across world correlates with C677T allele of the methylenetetrahydrofolate reductase (MTHFR) gene prevalence. J. Clin. Lab. Anal. 2021, 35, e23798. [Google Scholar] [CrossRef]
- Fedorowski, A. Postural orthostatic tachycardia syndrome: clinical presentation, aetiology and management. J. Intern. Med. 2018, 285, 352–366. [Google Scholar] [CrossRef]
- Chadda, K.R.; Blakey, E.E.; Huang, C.L.-. .-H.; Jeevaratnam, K. Long COVID-19 and Postural Orthostatic Tachycardia Syndrome- Is Dysautonomia to Be Blamed? Front. Cardiovasc. Med. 2022, 9, 860198. [Google Scholar] [CrossRef]
- Goldstein, J. Betrayal by the Brain: The Neurologic Basis of Chronic Fatigue Syndrome, Fibromyalgia Syndrome, and Related Neural Network. 1st edition Routledge 1996 USA.
- Stewart, J.M.; Glover, J.L.; Medow, M.S. Increased plasma angiotensin II in postural tachycardia syndrome (POTS) is related to reduced blood flow and blood volume. 110. [CrossRef]
- Okamoto, L.E.; Raj, S.R.; Peltier, A.; Gamboa, A.; Shibao, C.; Diedrich, A.; Black, B.K.; Robertson, D.; Biaggioni, I. Neurohumoral and haemodynamic profile in postural tachycardia and chronic fatigue syndromes. 122. [CrossRef]
- Li, Y.; He, B.; Li, H.; Zhang, Q.; Tang, C.; Du, J.; Jin, H. Plasma Homocysteine Level in Children With Postural Tachycardia Syndrome. Front. Pediatr. 2018, 6, 375. [Google Scholar] [CrossRef]
- Yu, X.; Li, H.; Murphy, T.A.; Nuss, Z.; Liles, J.; Liles, C.; Aston, C.E.; Raj, S.R.; Fedorowski, A.; Kem, D.C. Angiotensin II Type 1 Receptor Autoantibodies in Postural Tachycardia Syndrome. J. Am. Hear. Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Fedorowski, A.; Li, H.; Yu, X.; Koelsch, K.A.; Harris, V.M.; Liles, C.; Murphy, T.A.; Quadri, S.M.S.; Scofield, R.H.; Sutton, R.; et al. Antiadrenergic autoimmunity in postural tachycardia syndrome. Eur. 2016, 19, 1211–1219. [Google Scholar] [CrossRef]
- Badiudeen, T.; Forsythe, E.A.; Bennett, G.; Li, H.; Yu, X.; Beel, M.; Nuss, Z.; Blick, K.E.; Okamoto, L.E.; Arnold, A.C.; et al. A functional cell-based bioassay for assessing adrenergic autoantibody activity in postural tachycardia syndrome. J. Transl. Autoimmun. 2019, 2, 100006. [Google Scholar] [CrossRef]
- Wirth, K.; Scheibenbogen, C. A Unifying Hypothesis of the Pathophysiology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Recognitions from the finding of autoantibodies against ß2-adrenergic receptors. Autoimmun. Rev. 2020, 19, 102527. [Google Scholar] [CrossRef]
- Briquez, P.S.; Rouhani, S.J.; Yu, J.; Pyzer, A.R.; Trujillo, J.; Dugan, H.L.; Stamper, C.T.; Changrob, S.; Sperling, A.I.; Wilson, P.C.; et al. Severe COVID-19 induces autoantibodies against angiotensin II that correlate with blood pressure dysregulation and disease severity. Sci. Adv. 2022, 8, eabn3777. [Google Scholar] [CrossRef]
- Barki-Harrington, L.; Luttrell, L.M.; Rockman, H.A. Dual Inhibition of β-Adrenergic and Angiotensin II Receptors by a Single Antagonist. Circulation 2003, 108, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yu, B.; Liu, Z.; Li, J.; Ma, M.; Wang, Y.; Zhu, M.; Yin, H.; Wang, X.; Fu, Y.; et al. Homocysteine directly interacts and activates the angiotensin II type I receptor to aggravate vascular injury. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Häggström, M.; Richfield, D. Diagram of the pathways of human steroidogenesis. WikiJournal Med. 2014, 1. [Google Scholar] [CrossRef]
- Hall, K.T.; Battinelli, E.; Chasman, D.I.; Ridker, P.M.; Psaty, B.M.; Rotter, J.I.; Kaptchuk, T.J.; Tracy, R.P.; Wassel, C.L.; Mukamal, K.J. Catechol-O-Methyltransferase and Cardiovascular Disease: MESA. J. Am. Hear. Assoc. 2019, 8, e014986. [Google Scholar] [CrossRef] [PubMed]
- Małecki, J.M.; Davydova, E.; Falnes, P. . Protein methylation in mitochondria. J. Biol. Chem. 2022, 298, 101791. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, YK, et al “Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19”. Gut 70 (2021):698-706 https://gut.bmj.
- König, R.S.; Albrich, W.C.; Kahlert, C.R.; Bahr, L.S.; Löber, U.; Vernazza, P.; Scheibenbogen, C.; Forslund, S.K. The Gut Microbiome in Myalgic Encephalomyelitis (ME)/Chronic Fatigue Syndrome (CFS). Front. Immunol. 2022, 12, 628741. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y.; et al. GABA-producing Bifidobacterium dentium modulates visceral sensitivity in the intestine. Neurogastroenterol. Motil. 2016, 29. [Google Scholar] [CrossRef]
- Ko, C.Y.; Lin, H.-T.V.; Tsai, G.J. Gamma-aminobutyric acid production in black soybean milk by Lactobacillus brevis FPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model. Process. Biochem. 2013, 48, 559–568. [Google Scholar] [CrossRef]
- Mayengbam, S.; Chleilat, F.; Reimer, R.A. Dietary Vitamin B6 Deficiency Impairs Gut Microbiota and Host and Microbial Metabolites in Rats. Biomedicines 2020, 8, 469. [Google Scholar] [CrossRef]
- Dev, S.; Mizuguchi, H.; Das, A.K.; Matsushita, C.; Maeyama, K.; Umehara, H.; Ohtoshi, T.; Kojima, J.; Nishida, K.; Takahashi, K.; et al. Suppression of Histamine Signaling by Probiotic Lac-B: a Possible Mechanism of Its Anti-allergic Effect. J. Pharmacol. Sci. 2008, 107, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ye, Z.; Houston, C.M.; Zecharia, A.Y.; Ma, Y.; Zhang, Z.; Uygun, D.S.; Parker, S.; Vyssotski, A.L.; Yustos, R.; et al. Wakefulness Is Governed by GABA and Histamine Cotransmission. Neuron 2015, 87, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Jiménez, F.J.; Esguevillas, G.; Alonso-Navarro, H.; Zurdo, M.; Turpín-Fenoll, L.; Millán-Pascual, J.; Adeva-Bartolomé, T.; Cubo, E.; Navacerrada, F.; Amo, G.; et al. Gamma-aminobutyric acid (GABA) receptors genes polymorphisms and risk for restless legs syndrome. Pharmacogenomics J. 2018, 18, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, L.B.; Brook, J.B.; Walters, A.S.; Goris, A.; Afrin, L.B.; Molderings, G.J. Restless legs syndrome is associated with long-COVID in women. Sleep Med. 2022, 18, 1413–1418. [Google Scholar] [CrossRef]
- Civelek, G.M.; Ciftkaya, P.O.; Karatas, M. Evaluation of restless legs syndrome in fibromyalgia syndrome: An analysis of quality of sleep and life. J. Back Musculoskelet. Rehabilitation 2014, 27, 537–544. [Google Scholar] [CrossRef]
- Dodson, C.; Bagai, K.; Weinstock, L.B.; Thompson, E.; Okamoto, L.E.; Peltier, A.; Raj, S.R.; Walters, A.S. Restless legs syndrome is increased in postural orthostatic tachycardia syndrome. Sleep Med. 2021, 17, 791–795. [Google Scholar] [CrossRef]
- Blum, D.J.; During, E.; Barwick, F.; Davidenko, P.; Zeitzer, J.M. 0009 Restless Leg Syndrome: Does It Start With A Gut Feeling? Sleep 2019, 42, A4–A4. [Google Scholar] [CrossRef]
- Riera, J.J.; Schousboe, A.; Waagepetersen, H.S.; Howarth, C.; Hyder, F. The micro-architecture of the cerebral cortex: Functional neuroimaging models and metabolism. NeuroImage 2008, 40, 1436–1459. [Google Scholar] [CrossRef]
- Jammoul, M.; Naddour, J.; Madi, A.; Reslan, M.A.; Hatoum, F.; Zeineddine, J.; Abou-Kheir, W.; Lawand, N. Investigating the possible mechanisms of autonomic dysfunction post-COVID-19. Auton. Neurosci. 2022, 245, 103071–103071. [Google Scholar] [CrossRef]
- Kaur, C.; Ling, E.-A. The circumventricular organs. 32. [CrossRef]
- Cytokines Driving Sympathetic Nervous System Activation In The Subfornical Organ: Implications For Heart Failure And Hypertension (2015) Gabriel Bassi https://brainimmune.
- Cutsforth-Gregory, J.K.; Benarroch, E.E. Nucleus of the solitary tract, medullary reflexes, and clinical implications. Neurology 2017, 88, 1187–1196. [Google Scholar] [CrossRef]
- Page, M.C.; Cassaglia, P.A.; Brooks, V.L.; Erdos, B.; Clifton, R.R.; Liu, M.; Li, H.; McCowan, M.L.; Sumners, C.; Scheuer, D.A.; et al. GABA in the paraventricular nucleus tonically suppresses baroreflex function: alterations during pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R1452–R1458. [Google Scholar] [CrossRef]
- Haam, J.; Popescu, I.R.; Morton, L.A.; Halmos, K.C.; Teruyama, R.; Ueta, Y.; Tasker, J.G. GABA Is Excitatory in Adult Vasopressinergic Neuroendocrine Cells. J. Neurosci. 2012, 32, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Kim, Y.-B.; Kim, J.S.; Bin Kim, W.; Kim, Y.S.; Han, H.C.; Colwell, C.S.; Cho, Y.-W.; Kim, Y.I. GABAergic inhibition is weakened or converted into excitation in the oxytocin and vasopressin neurons of the lactating rat. Mol. Brain 2015, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kakizawa, K.; Watanabe, M.; Mutoh, H.; Okawa, Y.; Yamashita, M.; Yanagawa, Y.; Itoi, K.; Suda, T.; Oki, Y.; Fukuda, A. A novel GABA-mediated corticotropin-releasing hormone secretory mechanism in the median eminence. Sci. Adv. 2016, 2, e1501723–1501723. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.M.; Abbott, G.; Mair, J.; Prescott, M.; Campbell, R.E. Mapping GABA and glutamate inputs to gonadotrophin-releasing hormone neurones in male and female mice. J. Neuroendocr. 2018, 30, e12657. [Google Scholar] [CrossRef]
- Wiens, S.C.; Trudeau, V.L. Thyroid hormone and γ-aminobutyric acid (GABA) interactions in neuroendocrine systems. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2006, 144, 332–344. [Google Scholar] [CrossRef]
- Unnikrishnan P, Diabetes Insipidus, Dept of Neuroanesthesia SCTIMST, Trivandrum, Kerala, India https://www.uzhnu.edu. 2398.
- Miwa, K. Down-regulation of renin–aldosterone and antidiuretic hormone systems in patients with myalgic encephalomyelitis/chronic fatigue syndrome. J. Cardiol. 2017, 69, 684–688. [Google Scholar] [CrossRef]
- Pereira, G.; Gillies, H.; Chanda, S.; Corbett, M.; Vernon, S.D.; Milani, T.; Bateman, L. Acute Corticotropin-Releasing Factor Receptor Type 2 Agonism Results in Sustained Symptom Improvement in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Syst. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Coffin, S.T.; Black, B.K.; Biaggioni, I.; Paranjape, S.Y.; Orozco, C.; Black, P.W.; Dupont, W.D.; Robertson, D.; Raj, S.R. Desmopressin acutely decreases tachycardia and improves symptoms in the postural tachycardia syndrome. Hear. Rhythm. 2012, 9, 1484–1490. [Google Scholar] [CrossRef]
- Diep, P.-T.; Chaudry, M.; Dixon, A.; Chaudry, F.; Kasabri, V. Oxytocin, the panacea for long-COVID? a review. Horm. Mol. Biol. Clin. Investig. 2022, 43, 363–371. [Google Scholar] [CrossRef]
- Goldstein, JA. , Betrayal by the Brain: The Neurologic Basis of Chronic Fatigue Syndrome, Fibromyalgia Syndrome, and Related Neural Network 2012 Haworth Medical Press.
- Amir, S. Stimulation of the paraventricular nucleus with glutamate activates interscapular brown adipose tissue thermogenesis in rats. Brain Res. 1990, 508, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Frosst, P.; Blom, H.J.; Milos, R.; Goyette, P.; Sheppard, C.A.; Matthews, R.G.; Boers, G.J.H.; den Heijer, M.; Kluijtmans, L.A.J.; van den Heuve, L.P.; et al. A candidate genetic risk factor for vascular disease: a common mutation in methylenetetrahydrofolate reductase. Nat. Genet. 1995, 10, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadeq, D.W.; Nasrallah, G.K. The Spectrum of Mutations of Homocystinuria in the MENA Region. Genes 2020, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Nefic, H, et al. “The Frequency of the 677C>T and 1298A>C Polymorphisms in the Methylenetetrahydrofolate Reductase (MTHFR) Gene in the Population” Med Arch. 2018 Jun;72(3):164-169.
- Carpenè, G.; Negrini, D.; Henry, B.M.; Montagnana, M.; Lippi, G. Homocysteine in coronavirus disease (COVID-19): a systematic literature review. Diagnosis 2022, 9, 306–310. [Google Scholar] [CrossRef]
- Regland, B.; Andersson, M.; Abrahamsson, L.; Bagby, J.; Dyrehag, L.E.; Gottfries, C.G. Increased Concentrations of Homocysteine in the Cerebrospinal Fluid in Patients with Fibromyalgia and Chronic Fatigue Syndrome. Scand. J. Rheumatol. 1997, 26, 301–307. [Google Scholar] [CrossRef]
- Bottiglieri, T.; Laundy, M.; Crellin, R.; Toone, B.K.; Carney, M.W.P.; Reynolds, E.H. Homocysteine, folate, methylation, and monoamine metabolism in depression. J. Neurol. Neurosurg. Psychiatry 2000, 69, 228–232. [Google Scholar] [CrossRef]
- Oner, P.; Yilmaz, S.; Doğan, S. High Homocysteine Levels Are Associated with Cognitive Impairment in Patients Who Recovered from COVID-19 in the Long Term. J. Pers. Med. 2023, 13, 503. [Google Scholar] [CrossRef]
- Ren, J.-C.; Wu, Y.-X.; Wu, Z.B.; Zhang, G.-H.; Wang, H.B.; Liu, H.B.; Cui, J.-P.B.; Chen, Q.; Liu, J.; Frank, A.; et al. MTHFR Gene Polymorphism Is Associated With DNA Hypomethylation and Genetic Damage Among Benzene-Exposed Workers in Southeast China. J. Occup. Environ. Med. 2018, 60, e188–e192. [Google Scholar] [CrossRef]
- Nikesjö, F.; Sayyab, S.; Karlsson, L.; Apostolou, E.; Rosén, A.; Hedman, K.; Lerm, M. Defining post-acute COVID-19 syndrome (PACS) by an epigenetic biosignature in peripheral blood mononuclear cells. Clin. Epigenetics 2022, 14, 1–6. [Google Scholar] [CrossRef]
- Fryar-Williams, S. Fundamental Role of Methylenetetrahydrofolate Reductase 677 C → T Genotype and Flavin Compounds in Biochemical Phenotypes for Schizophrenia and Schizoaffective Psychosis. Front. Psychiatry 2016, 7, 172. [Google Scholar] [CrossRef]
- Shaker, MM, et al. “Correlation of methylation status in MTHFR promoter region with recurrent pregnancy loss” J Genet Eng Biotechnol. 2021 Mar 22;19(1):44.
- Walsh, WJ. “Nutrient Power: Heal Your Biochemistry and Heal Your Brain”. 2012. [Google Scholar]
- Bonds, R.S.; Midoro-Horiuti, T. Estrogen effects in allergy and asthma. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Balnis, J.; Madrid, A.; Hogan, K.J.; Drake, L.A.; Adhikari, A.; Vancavage, R.; Singer, H.A.; Alisch, R.S.; Jaitovich, A. Persistent blood DNA methylation changes one year after SARS-CoV-2 infection. Clin. Epigenetics 2022, 14, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sarı, I.K.; Keskin, O.; Keskin, A.S.; Elli̇dağ, H.Y.; Harmandar, O. Is Homocysteine Associated with the Prognosis of Covid-19 Pneumonia. Int. J. Clin. Pr. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- Rhein, V.F.; Carroll, J.; He, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Human METTL20 Methylates Lysine Residues Adjacent to the Recognition Loop of the Electron Transfer Flavoprotein in Mitochondria. J. Biol. Chem. 2014, 289, 24640–24651. [Google Scholar] [CrossRef] [PubMed]
- de Vega, W.C.; Vernon, S.D.; McGowan, P.O. DNA Methylation Modifications Associated with Chronic Fatigue Syndrome. PLOS ONE 2014, 9, e104757. [Google Scholar] [CrossRef] [PubMed]
- Benelli, M.; Franceschini, G.M.; Magi, A.; Romagnoli, D.; Biagioni, C.; Migliaccio, I.; Malorni, L.; Demichelis, F. Charting differentially methylated regions in cancer with Rocker-meth. Commun. Biol. 2021, 4, 1–15. [Google Scholar] [CrossRef]
- McKinney, B.C.; Lin, C.-W.; Rahman, T.; Oh, H.; Lewis, D.A.; Tseng, G.; Sibille, E. DNA methylation in the human frontal cortex reveals a putative mechanism for age-by-disease interactions. Transl. Psychiatry 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Takahashi, T.; Ellingson, M.K.; Wong, P.; Israelow, B.; Lucas, C.; Klein, J.; Silva, J.; Mao, T.; Oh, J.E.; Tokuyama, M.; et al. Sex differences in immune responses that underlie COVID-19 disease outcomes. Nature 2020, 588, 315–320. [Google Scholar] [CrossRef]
- Pujantell, M.; Altfeld, M. Consequences of sex differences in Type I IFN responses for the regulation of antiviral immunity. Front. Immunol. 2022, 13, 986840. [Google Scholar] [CrossRef]
- Chambers, P.W. (2022) Long Covid, Short Magnesium. Open Access Library Journal, 9, e8736. https://www.scirp.org/journal/paperinformation.aspx? 1174. [Google Scholar]
- Goetzl, EJ, et al. “Gender specificity of altered human immune cytokine profiles in aging” FASEB J. 2010 Sep;24(9):3580-9.
- De Benedetti, F.; Prencipe, G.; Bracaglia, C.; Marasco, E.; Grom, A.A. Targeting interferon-γ in hyperinflammation: opportunities and challenges. Nat. Rev. Rheumatol. 2021, 17, 678–691. [Google Scholar] [CrossRef]
- Bossi, F.; Peerschke, E.I.; Ghebrehiwet, B.; Tedesco, F. Cross-talk between the complement and the kinin system in vascular permeability. Immunol. Lett. 2011, 140, 7–13. [Google Scholar] [CrossRef]
- Kuklina, E.M. T Lymphocytes as Targets for SARS-CoV-2. Biochem. (Moscow) 2022, 87, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Chambers, P.W. Basigin Binds Spike S on SARS-CoV2. OALib 2021, 08, 1–7. [Google Scholar] [CrossRef]
- Ma, Q.; Hao, Z.-W.; Wang, Y.-F. The effect of estrogen in coronavirus disease 2019. Am. J. Physiol. Cell. Mol. Physiol. 2021, 321, L219–L227. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.Y.; Hodis, H.N.; Mack, W.J.; Mather, M. Estradiol Therapy After Menopause Mitigates Effects of Stress on Cortisol and Working Memory. J. Clin. Endocrinol. Metab. 2017, 102, 4457–4466. [Google Scholar] [CrossRef] [PubMed]
- Stelzig, K.E.; Canepa-Escaro, F.; Schiliro, M.; Berdnikovs, S.; Prakash, Y.S.; Chiarella, S.E. Estrogen regulates the expression of SARS-CoV-2 receptor ACE2 in differentiated airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1280–L1281. [Google Scholar] [CrossRef] [PubMed]
- Mendes, G.M.d.M.; Nascimento, I.J.B.D.; Marazzi-Diniz, P.H.; Da Silveira, I.B.; Itaborahy, M.F.; Viana, L.E.; Silva, F.A.; Santana, M.F.; Pinto, R.A.; Dutra, B.G.; et al. The des-Arg9-bradykinin/B1R axis: Hepatic damage in COVID-19. Front. Physiol. 2022, 13, 1080837. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Netea, M.G.; van Deuren, M.; van der Meer, J.W.; de Mast, Q.; Brüggemann, R.J.; van der Hoeven, H. Kallikrein-kinin blockade in patients with COVID-19 to prevent acute respiratory distress syndrome. eLife 2020, 9. [Google Scholar] [CrossRef]
- Stewart, J.M.; Taneja, I.; Glover, J.; Medow, M.S.; Lang, J.A.; Krajek, A.C.; McNeely, B.D.; Meade, R.D.; Fujii, N.; Seely, A.J.E.; et al. Angiotensin II type 1 receptor blockade corrects cutaneous nitric oxide deficit in postural tachycardia syndrome. Am. J. Physiol. Circ. Physiol. 2008, 294, H466–H473. [Google Scholar] [CrossRef]
- Kiowski, W.; Linder, L.; Kleinbloesem, C.; van Brummelen, P.; Bühler, F.R. Blood pressure control by the renin-angiotensin system in normotensive subjects. Assessment by angiotensin converting enzyme and renin inhibition. Circulation 1992, 85, 1–8. [Google Scholar] [CrossRef]
- Persson, P.B. Renin: origin, secretion and synthesis. J. Physiol. 2003, 552, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.L.; Jiang, L.; Adams, J.S.; Xu, Z.Z.; Shen, J.; Janssen, S.; Ackermann, G.; Vanderschueren, D.; Pauwels, S.; Knight, R.; et al. Vitamin D metabolites and the gut microbiome in older men. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.C.; Davis, C.T.; Zhu, W.; Bowman-Kirigin, J.A.; Walker, A.E.; Tai, Z.; Thomas, K.R.; Donato, A.J.; Lesniewski, L.A.; Li, D.Y. Dietary Vitamin D and Its Metabolites Non-Genomically Stabilize the Endothelium. PLOS ONE 2015, 10, e0140370–e0140370. [Google Scholar] [CrossRef]
- Moreira-Rosário, A.; Marques, C.; Pinheiro, H.; Araújo, J.R.; Ribeiro, P.; Rocha, R.; Mota, I.; Pestana, D.; Ribeiro, R.; Pereira, A.; et al. Gut Microbiota Diversity and C-Reactive Protein Are Predictors of Disease Severity in COVID-19 Patients. Front. Microbiol. 2021, 12, 705020. [Google Scholar] [CrossRef] [PubMed]
- Bikle, DD. “Vitamin D metabolism, mechanism of action, and clinical applications”. Chem Biol. 21.3 (2014): 319-29.
- Warren, T.; McAllister, R.; Morgan, A.; Rai, T.S.; McGilligan, V.; Ennis, M.; Page, C.; Kelly, C.; Peace, A.; Corfe, B.M.; et al. The Interdependency and Co-Regulation of the Vitamin D and Cholesterol Metabolism. Cells 2021, 10, 2007. [Google Scholar] [CrossRef]
- Ashangari, C.; Suleman, A. Abstract 121: Vitamin D Deficiency Study in Postural Orthostatic Tachycardia Syndrome. Circ. Cardiovasc. Qual. Outcomes 2015, 8. [Google Scholar] [CrossRef]
- Machado, CD. “The human gut microbiome displays diurnal and seasonal rhythmic patterns,” abstract 395, on Sunday, , at 11:20 am, Digestive Disease Week (2023) in Chicago, IL https://news.ddw. 7 May.
- Yildiz, S.; Mazel-Sanchez, B.; Kandasamy, M.; Manicassamy, B.; Schmolke, M. Influenza A virus infection impacts systemic microbiota dynamics and causes quantitative enteric dysbiosis. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Groeneveld, J.M.; Ballering, A.V.; van Boven, K.; Akkermans, R.P.; Hartman, T.C.O.; A Uijen, A. Sex differences in incidence of respiratory symptoms and management by general practitioners. Fam. Pr. 2020, 37, 631–636. [Google Scholar] [CrossRef]
- Al-Khaldy, N.S.; Al-Musharaf, S.; Aljazairy, E.A.; Hussain, S.D.; Alnaami, A.M.; Al-Daghri, N.; Aljuraiban, G. Serum Vitamin D Level and Gut Microbiota in Women. Healthcare 2023, 11, 351. [Google Scholar] [CrossRef]
- Waterhouse, M.; Hope, B.; Krause, L.; Morrison, M.; Protani, M.M.; Zakrzewski, M.; Neale, R.E. Vitamin D and the gut microbiome: a systematic review of in vivo studies. Eur. J. Nutr. 2018, 58, 2895–2910. [Google Scholar] [CrossRef]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Blitshteyn, S. Vitamin B1 deficiency in patients with postural tachycardia syndrome (POTS). Neurol. Res. 2017, 39, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Know, Lee. T: Mitochondria and the Future of Medicine, 2018.
- Atanassova, N. and Koeva, Y. (2012) Hydrohysteroid Dehydrogenases—Biological Role and Clinical Importance—Review. In: Canuto, R.A., Ed., Dehydrogenases, IntechOpen, London. [CrossRef]
- Depeint, F.; Bruce, W.R.; Shangari, N.; Mehta, R.; O’Brien, P.J. Mitochondrial function and toxicity: Role of the B vitamin family on mitochondrial energy metabolism. Chem. Biol. Interact. 2006, 163, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Azzini, E.; Ruggeri, S.; Polito, A. Homocysteine: Its Possible Emerging Role in At-Risk Population Groups. Int. J. Mol. Sci. 2020, 21, 1421. [Google Scholar] [CrossRef]
- Plasma Methylation Enzyme and Nutrition Guide, Doctor’s Data, Inc., St. Charles, Illinois https://site-akiajqrf22xmaqzsiz6q.s3.amazonaws.com/DDI+Website/Resource+Guides/78338+DDI+PlasmaMethylationEnzyme+WP_R4.
- Weinstock, L.B.; Brook, J.B.; Myers, T.L.; Goodman, B. Successful treatment of postural orthostatic tachycardia and mast cell activation syndromes using naltrexone, immunoglobulin and antibiotic treatment. BMJ Case Rep. 2018, 2018. [Google Scholar] [CrossRef]
- Uy, P.; Jehangir, A.; Yap, J.E.; Rao, S. S493 SIBO and SIFO Prevalence in Patients With Ehlers-Danlos Syndrome Based on Duodenal Aspirates/Culture. Am. J. Gastroenterol. 2021, 116, S218–S219. [Google Scholar] [CrossRef]
- Rao, S.S.C.; Bhagatwala, J. Small Intestinal Bacterial Overgrowth. Clin. Transl. Gastroenterol. 2019, 10, e00078. [Google Scholar] [CrossRef]
- Sánchez-Pérez, S.; Comas-Basté, O.; Duelo, A.; Veciana-Nogués, M.T.; Berlanga, M.; Latorre-Moratalla, M.L.; Vidal-Carou, M.C. Intestinal Dysbiosis in Patients with Histamine Intolerance. Nutrients 2022, 14, 1774. [Google Scholar] [CrossRef]
- Courseault, J.; Kingry, C.; Morrison, V.; Edstrom, C.; Morrell, K.; Jaubert, L.; Elia, V.; Bix, G. Folate-dependent hypermobility syndrome: A proposed mechanism and diagnosis. Heliyon 2023, 9, e15387. [Google Scholar] [CrossRef]
- Hadtstein, F.; Vrolijk, M. Vitamin B-6-Induced Neuropathy: Exploring the Mechanisms of Pyridoxine Toxicity. Adv. Nutr. Int. Rev. J. 2021, 12, 1911–1929. [Google Scholar] [CrossRef]
- Chambers P, Antioxidants and Long Covid. Open Access Library Journal (2022) 9:e9414. https://www.scirp.org/journal/paperinformation.aspx? 1208.
- Sfera, A.; Thomas, K.G.; Sasannia, S.; Anton, J.J.; Andronescu, C.V.; Garcia, M.; Sfera, D.O.; Cummings, M.A.; Kozlakidis, Z. Neuronal and Non-Neuronal GABA in COVID-19: Relevance for Psychiatry. Reports 2022, 5, 22. [Google Scholar] [CrossRef]
- Campbell, D.J.; Kladis, A.; Valentijn, A.J. Effects of Losartan on Angiotensin and Bradykinin Peptides and Angiotensin-Converting Enzyme. J. Cardiovasc. Pharmacol. 1995, 26, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Chen, Z.-L.; Ghosh, D.; Strickland, S.; Norris, E.H. Increased plasma bradykinin level is associated with cognitive impairment in Alzheimer's patients. Neurobiol. Dis. 2020, 139, 104833–104833. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, J.; Kim, H.; Kim, J.S. Correlation of ACE2 with RAS components after Losartan treatment in light of COVID-19. Sci. Rep. 2021, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kehoe, P.G.; Wong, S.; AL Mulhim, N.; Palmer, L.E.; Miners, J.S. Angiotensin-converting enzyme 2 is reduced in Alzheimer’s disease in association with increasing amyloid-β and tau pathology. Alzheimer's Res. Ther. 2016, 8, 1–10. [Google Scholar] [CrossRef]
- Bastolla, U.; Chambers, P.; Abia, D.; Garcia-Bermejo, M.-L.; Fresno, M. Is Covid-19 Severity Associated With ACE2 Degradation? Front. Drug Discov. 2022, 1. [Google Scholar] [CrossRef]
- Mahmudpour, M.; Roozbeh, J.; Keshavarz, M.; Farrokhi, S.; Nabipour, I. COVID-19 cytokine storm: The anger of inflammation. Cytokine 2020, 133, 155151–155151. [Google Scholar] [CrossRef]
- Yang, T.; Yang, Y.; Wang, D.; Li, C.; Qu, Y.; Guo, J.; Shi, T.; Bo, W.; Sun, Z.; Asakawa, T. The clinical value of cytokines in chronic fatigue syndrome. J. Transl. Med. 2019, 17, 1–12. [Google Scholar] [CrossRef]
- Zhang, X, et al. “TGF-β1 factor in the cerebrovascular diseases of Alzheimer’s disease”. Eur Rev Med Pharmacol Sci. 20.24 (2016): 5178-5185. https://pubmed.ncbi.nlm.nih. 2805.
- Zhang, Z.; Liu, C.; Gan, Z.; Wang, X.; Yi, Q.; Liu, Y.; Wang, Y.; Lu, B.; Du, H.; Shao, J.; et al. Improved Glucose-Stimulated Insulin Secretion by Selective Intraislet Inhibition of Angiotensin II Type 1 Receptor Expression in Isolated Islets of db/db Mice. Int. J. Endocrinol. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Chang, C.-H.; Chang, Y.-C.; Wu, L.-C.; Lin, J.-W.; Chuang, L.-M.; Lai, M.-S. Different angiotensin receptor blockers and incidence of diabetes: a nationwide population-based cohort study. Cardiovasc. Diabetol. 2014, 13, 91–91. [Google Scholar] [CrossRef]
- Villapol, S.; Saavedra, J.M. Neuroprotective Effects of Angiotensin Receptor Blockers. Am. J. Hypertens. 2014, 28, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.B.; Schmeidler, J.; Lesser, G.T.; Beeri, M.S.; Purohit, D.P.; Grossman, H.T.; Haroutunian, V. Less Alzheimer disease neuropathology in medicated hypertensive than nonhypertensive persons. Neurology 2009, 72, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- van Dalen, J.W.; Marcum, Z.A.; Gray, S.L.; Barthold, D.; van Charante, E.P.M.; van Gool, W.A.; Crane, P.K.; Larson, E.B.; Richard, E. Association of Angiotensin II–Stimulating Antihypertensive Use and Dementia Risk. Neurology 2020, 96, e67–e80. [Google Scholar] [CrossRef] [PubMed]
- Wood, E, et al. “Role of mitochondria, oxidative stress and the response to antioxidants in myalgic encephalomyelitis/chronic fatigue syndrome: A possible approach to SARS-CoV-2 ‘long-haulers’?” Chronic Dis Transl Med. 7.1 (2021): 14-26.
Figure 1.
Displayed are glutamatergic and GABAergic neurons. Note the magnitudes of the arrows (Riera et al., 2008). Note the need for TCA recycling of GABA within astrocytes.
Figure 1.
Displayed are glutamatergic and GABAergic neurons. Note the magnitudes of the arrows (Riera et al., 2008). Note the need for TCA recycling of GABA within astrocytes.
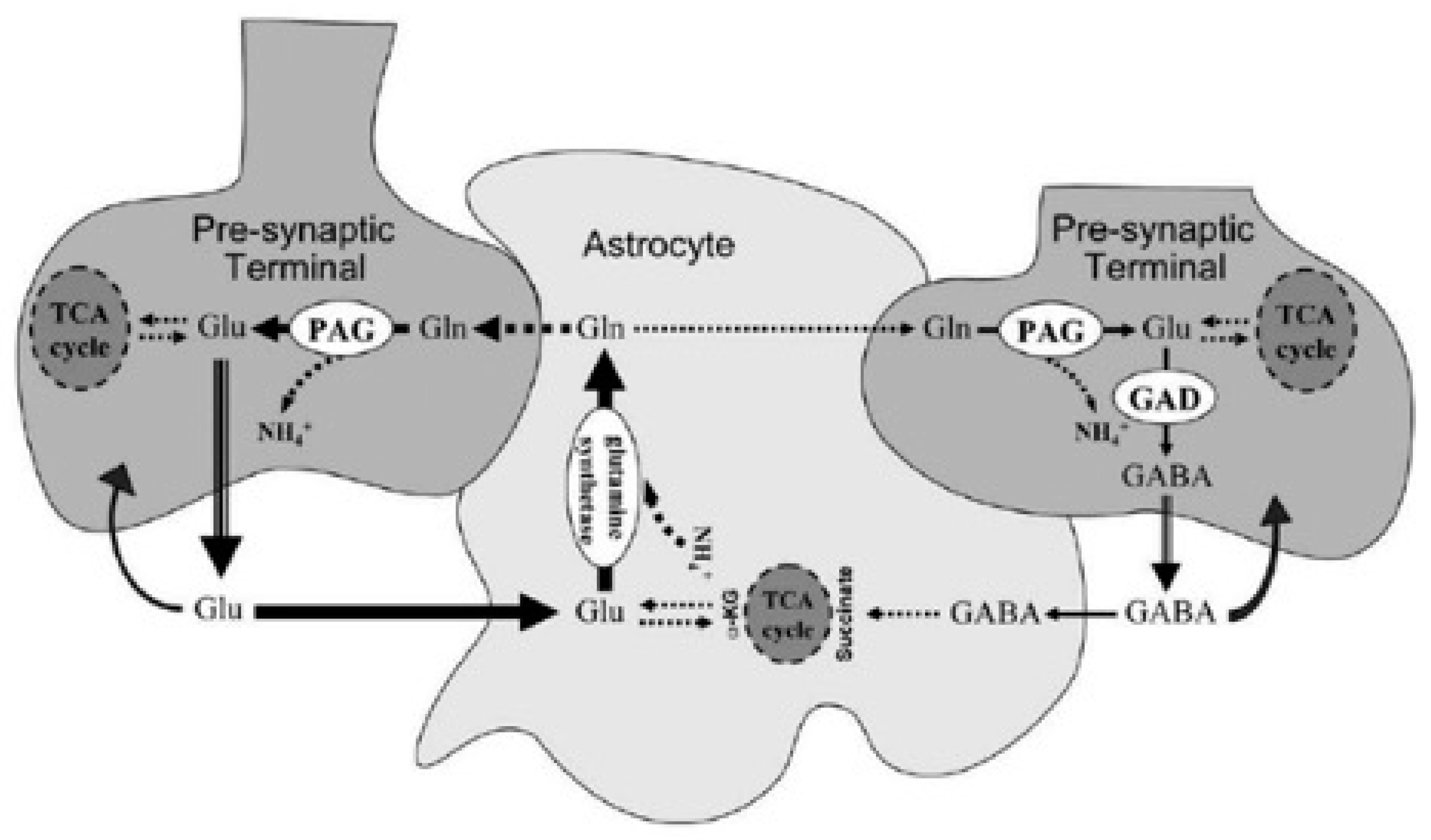
Figure 2.
Red structures designate CVOs (Bassi, 2015). The inferior area postrema is adjacent to the foramen of Magendie, a CSF passageway.
Figure 2.
Red structures designate CVOs (Bassi, 2015). The inferior area postrema is adjacent to the foramen of Magendie, a CSF passageway.
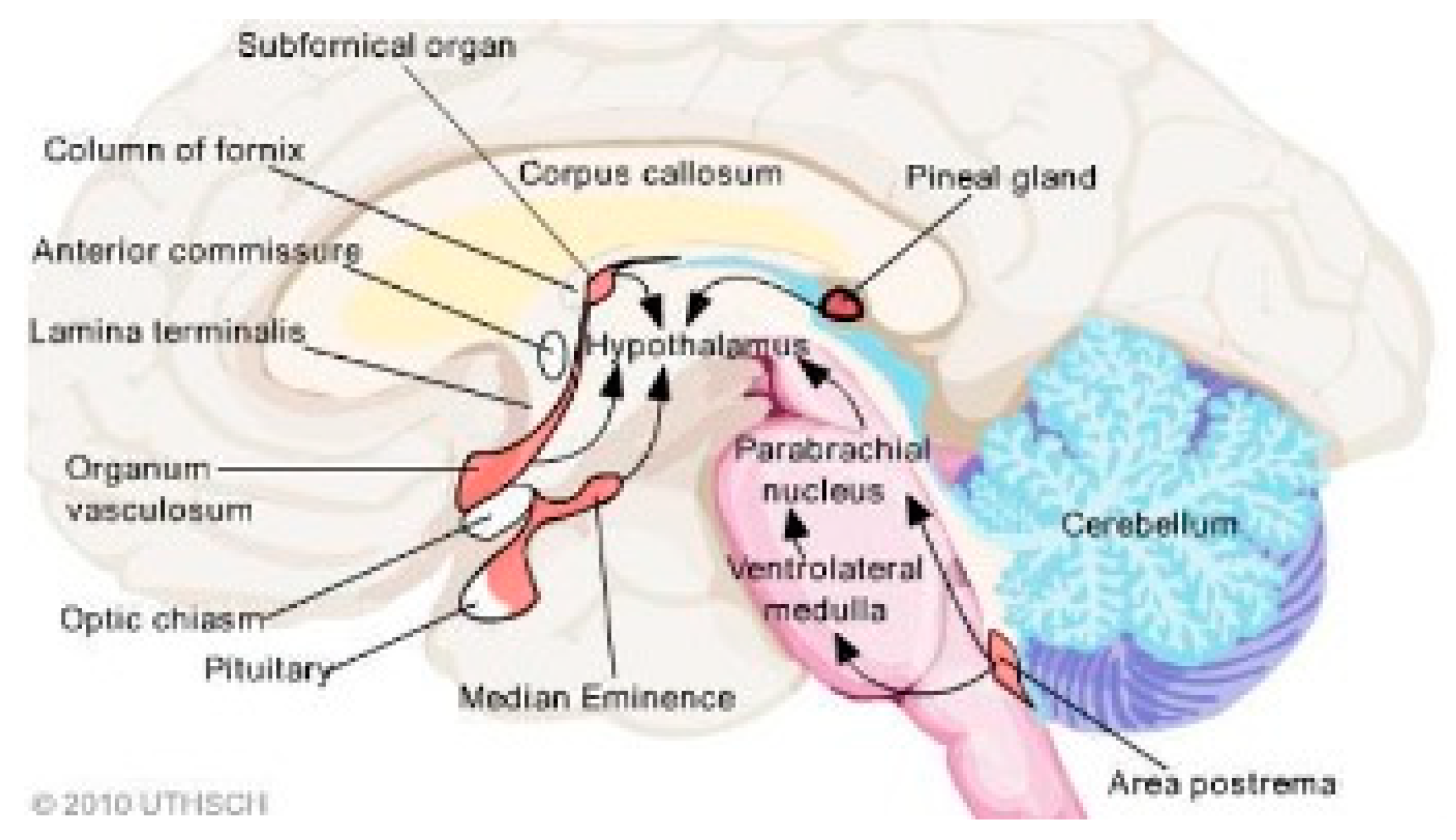
Figure 3.
The AP, the dorsal motor nucleus of X, and the nucleus of the solitary tract (NTS) form the dorsal vagal complex (Cutsforth-Gregory & Benarroch, 2017).
Figure 3.
The AP, the dorsal motor nucleus of X, and the nucleus of the solitary tract (NTS) form the dorsal vagal complex (Cutsforth-Gregory & Benarroch, 2017).
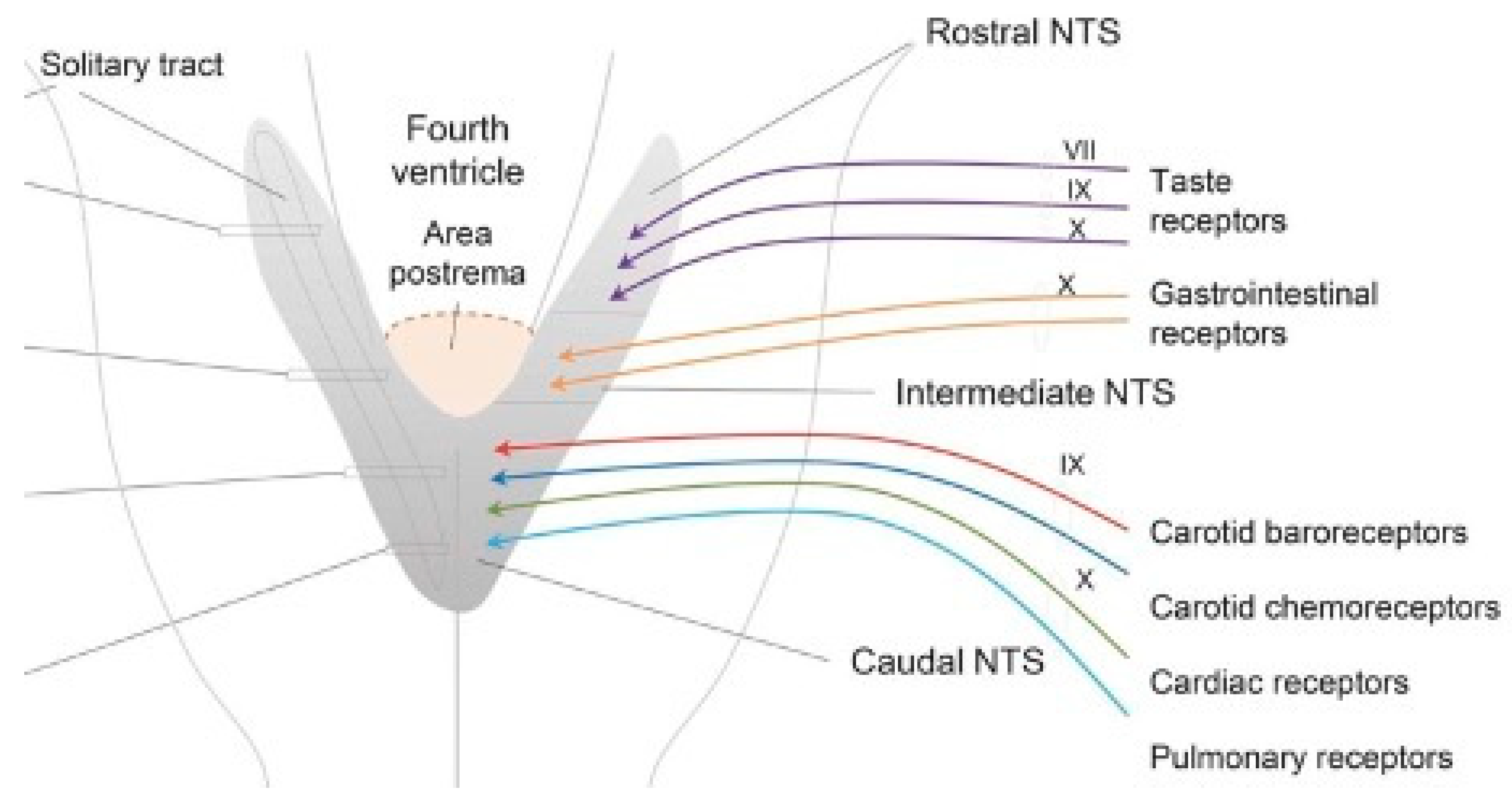
Figure 4.
Recycling of Hcy in the methionine cycle is enabled by methyl THF, otherwise compromised by MTHFR gene variants. Activated B2, B6, and B12 are required in the folate or re-methylation cycle. Pyridoxal phosphate (P5P) is B6 (Al-Sadeq & Nasrallah, 2020). B2 is also required to recycle oxidized glutathione back to its reduced antioxidant state.
Figure 4.
Recycling of Hcy in the methionine cycle is enabled by methyl THF, otherwise compromised by MTHFR gene variants. Activated B2, B6, and B12 are required in the folate or re-methylation cycle. Pyridoxal phosphate (P5P) is B6 (Al-Sadeq & Nasrallah, 2020). B2 is also required to recycle oxidized glutathione back to its reduced antioxidant state.
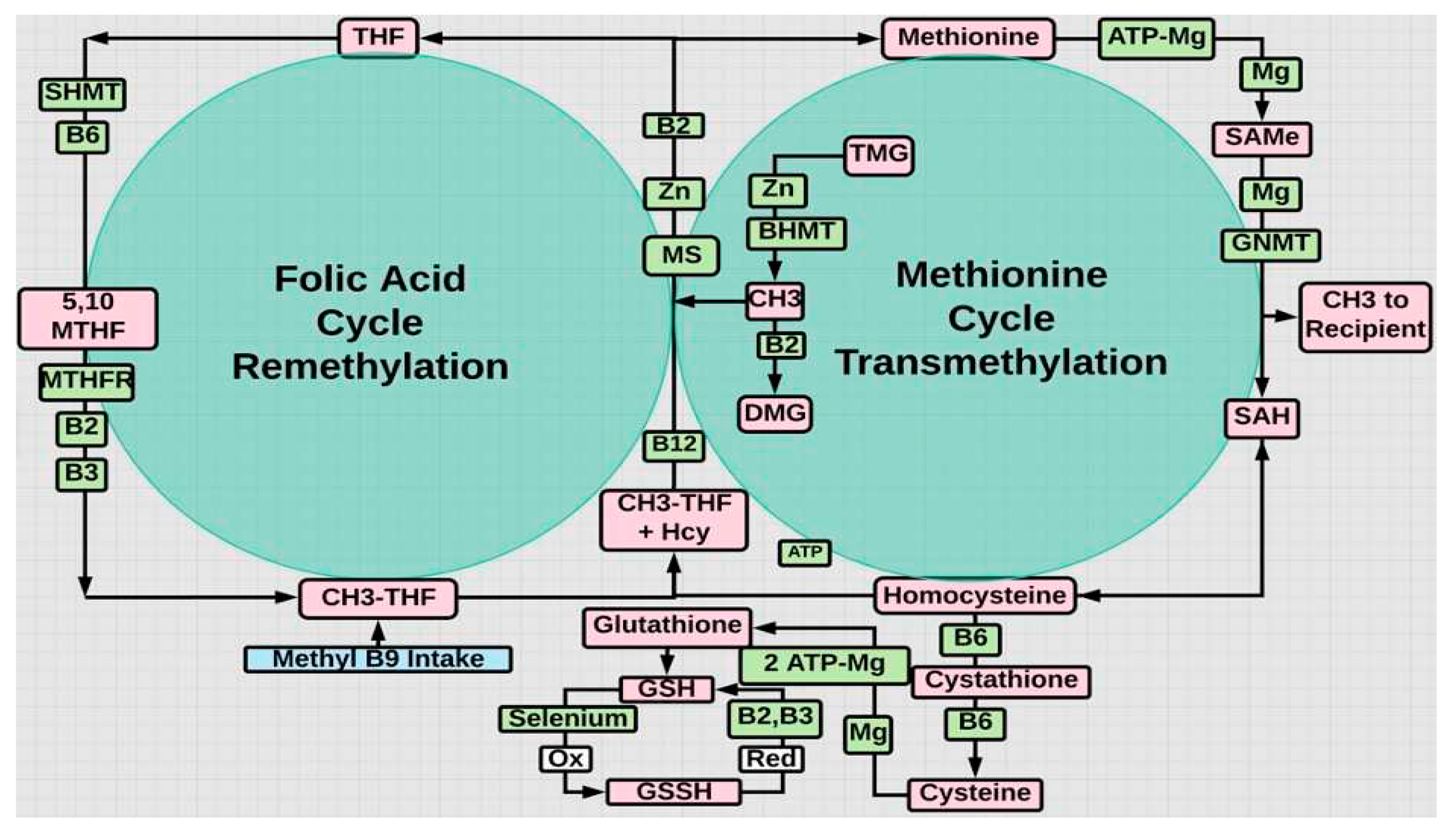
Figure 5.
ACE2 degrades des-argBKN (BK1R) and ACE degrades BKN (BK2R) => ACE and ACE2 are neuroprotective (van de Veerdonk, 2020).
Figure 5.
ACE2 degrades des-argBKN (BK1R) and ACE degrades BKN (BK2R) => ACE and ACE2 are neuroprotective (van de Veerdonk, 2020).
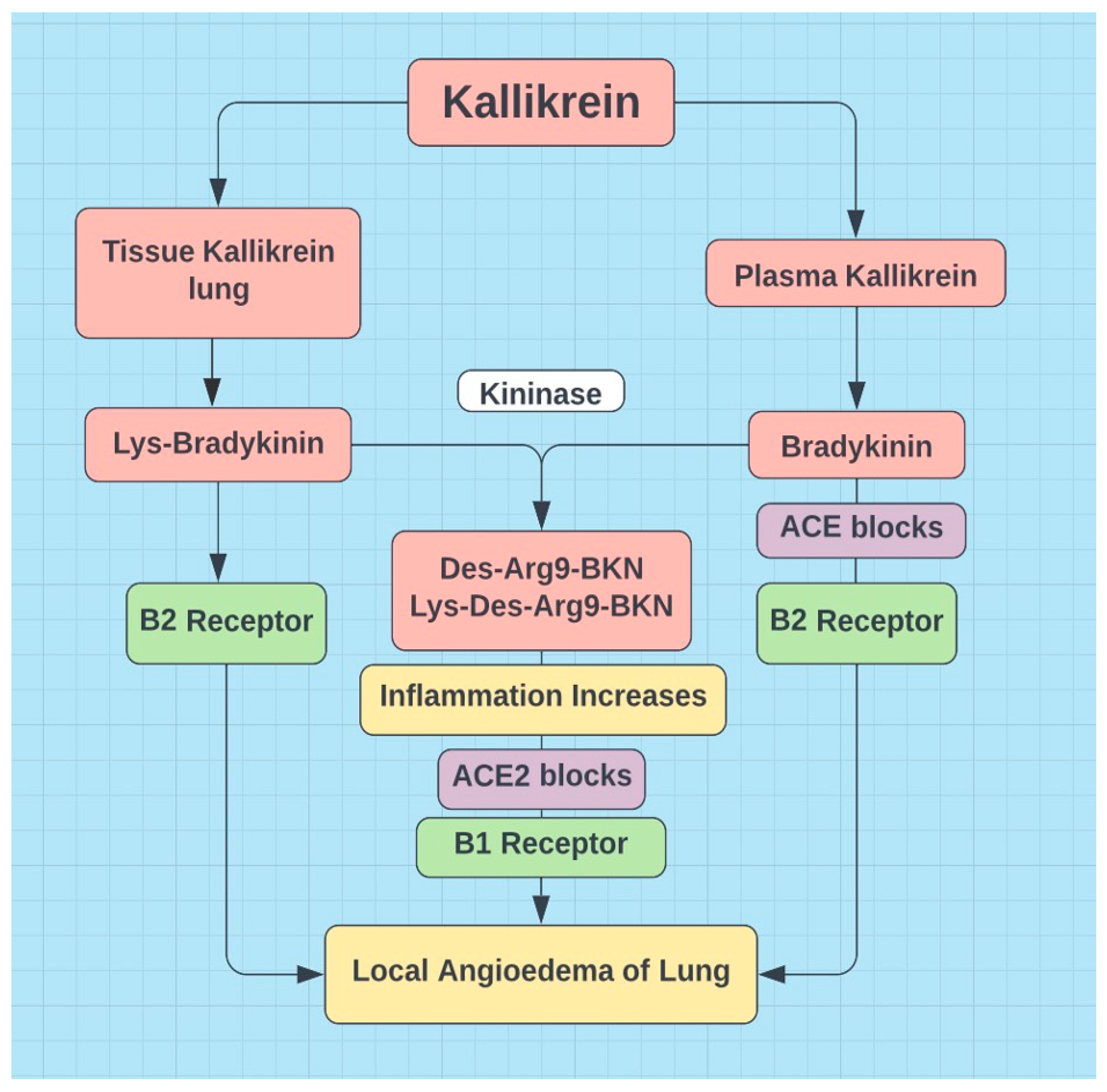
Figure 6.
Females less than 50, the demographic representing the vast majority of individuals diagnosed with POTS, are more Mg++ deficient than their male counterparts.
Figure 6.
Females less than 50, the demographic representing the vast majority of individuals diagnosed with POTS, are more Mg++ deficient than their male counterparts.
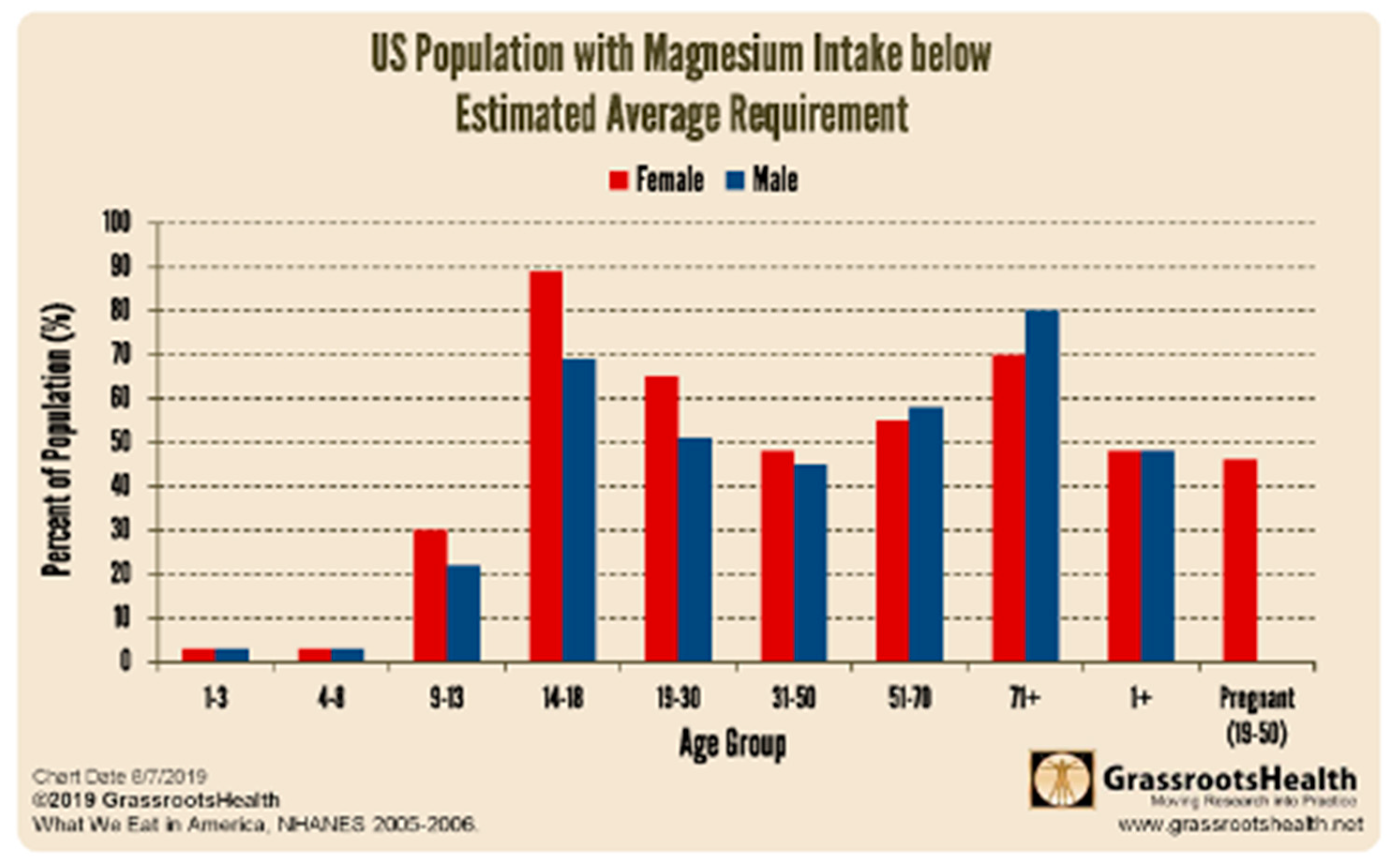
Figure 7.
Proposed flow chart indicating hypothetical relationships for LC, CFS, SIBO, MCAS, EDS, POTS, and MTHFR with Hcy as common denominator.
Figure 7.
Proposed flow chart indicating hypothetical relationships for LC, CFS, SIBO, MCAS, EDS, POTS, and MTHFR with Hcy as common denominator.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



