
Submitted:
24 March 2023
Posted:
27 March 2023
You are already at the latest version
Abstract
Traditional grain vinegar is fermented using multiple acetic acid bacteria (AAB) at various temperatures. A single AAB showed high acid–producing ability at 30 °C with a 5% alcohol concentration and an initial pH adjusted to 4.0. Multiple AAB were similar to single AAB, but the optimal initial pH was 3.0. Acid production ability according to the type of AAB was higher in multiple AAB than that for single AAB. That is, using multiple AAB helped to increase the titratable acidity of traditional grain vinegar. In addition, increasing the titratable acidity and content of volatile flavor compounds was advantageous when two, rather than four, AAB types were mixed and used. Titratable acidity was high at medium temperatures (30 °C), but volatile flavor compounds increased at low temperatures (20 °C) under multiple AAB. A 16S rDNA–based microbiome taxonomic profiling analysis identified differences in beta diversity due to multiple AAB and fermentation temperatures. In particular, beta diversity analysis revealed a specific pattern when a mixture of Acetobacter ascedens GV–8 and Acetobacter pasteurianus GV–22 were fermented at low temperature (20 °C). Therefore, we propose the application of multiple AAB with acidic and flavor–producing properties in traditional grain vinegar.
Keywords:
multiple starter
; acetic acid fermentation
; Acetobacter ascendens
; Acetobacter pasteurianus
1. Introduction
Vinegar is produced industrially in two main ways: a slow process involving a static surface and a fast submerged fermentation process, also known as acetic acid fermentation (AAF) [1]. Traditional vinegar is generally produced via static surface fermentation. This technique is relatively inexpensive, and the time required for complete fermentation is longer than that required for rapid fermentation. Traditional grain vinegar produced through static surface fermentation has various physiological functions, including blood sugar control, lipid metabolism control, weight loss, antibacterial, antioxidant, and anticancer activity due to its organic acid, polyphenol, and melanoidin content [2,3]. Vinegar quality varies greatly depending on the raw material, fermentation method, and manufacturing method used. The acetic acid content, organic acid composition, and free amino acid composition affect taste and influence the quality of vinegar [4].
Acetic acid bacteria (AAB), particularly Acetobacter spp., are commonly used in vinegar production. The Korean Ministry of Food and Drug Safety (https://www.foodsafetykorea.go.kr/) allows limited use of Acetobacter aceti and Acetobacter pasteurianus only in vinegar production. AAB, obligate aerobes, oxidize alcohols Acetobacter and Gluconoacetobacter spp., which can tolerate particularly high concentrations of acetic acid, are used as industrial vinegar producers [5]. A highly complex microbial community and metabolites are involved in traditional AAF; however, the correlation between them is still poorly understood [6]. Species and characterization of dominant strains in static AAF are desirable for stabilizing and improving fermentation [7,8,9,10].
The optimal temperature for traditional static AAF is approximately 30 °C, and a slight temperature increase of 2–3 °C results in significant reductions in both the rate and yield of AAF [11]. Therefore, setting the optimal conditions, including the fermentation temperature, for traditional grain vinegar production is a major task to increase the fermentation rate and yield [8]. In this study, optimal fermentation conditions were used for single and multiple AAF starters.
In our previous study, we investigated the microbiome of Korean traditional grain vinegars using a culture–independent microbiological technique, and correlations between sensory characteristics and fermentation temperature were found [2]. A recent studies reported that the ecological aspects of the microbial community influence metabolic activity and, ultimately, the composition of the fermentation product [12,13]. Fermentation of various foods and beverages, including traditional vinegar, is a complex phenomenon involving microbial consortia, showing the importance of studying the microorganisms in fermented products [14]. Beta diversity analysis was used to quantify the similarity or distance between the microbiome pairs. Overall taxonomic or functional diversity patterns can be linked to environmental features, and ecosystem characteristics can be predicted based on beta diversity analysis [15]. In this study, the microbiological and physicochemical properties of vinegar were investigated based on multiple AAB types and fermentation environments, and a multiple starter with acid–producing ability was presented.
2. Materials and Methods
2.1. Culture–Dependent Isolation and Identification of AAB
Five grain vinegars were collected from three provinces (Gyeongsangnam-do, Gyeongsangbuk-do, and Chungcheongnam-do) in Korea to isolate AAB with acid production and alcohol tolerance (Table 1) [2]. The composition of the solid medium used for isolation of AAB is as follows: 0.5% yeast extract, 3% glucose, 1% CaCO3, 2% agar, and 5% alcohol (v/v). Grain vinegars were spread on solid medium and incubated at 30 °C, and a single colony was isolated [16]. Amplification of the 16S rRNA gene was performed using the universal primers 27F (5’ –AGA GTT TGA TCC TGG CTC AG–3’) and 1492R (5’ –GGT TAC CTT GTT ACG ACT T–3’) in Genocell (Daejeon, Korea) [17]. The PCR mixtures were preheated for 5 min at 95 °C and amplified through 35 cycles of 45 s at 95 °C, 1 min at 55 °C, and 1 min at 72 °C. PCR products were purified and sequenced using the two primers described above. The nucleotide sequence primary analysis was performed using the web hosted BLASTn algorithm with the National Center for Biotechnology Information database. Table 1 shows the grain vinegars and identified AAB strains from the three regions in Korea. Two Acetobacter ascendens and four A. pasteurianus strains were isolated from five grain vinegars.
2.2. Optimal Fermentation Conditions for AAB
To analyze AAB fermentation characteristics, the optimal temperature, alcohol concentration, and initial pH were investigated to evaluate AAF ability. The liquid medium composition was as follows: 0.5% yeast extract, 0.5% glucose, 1% glycerin, 0.02% MgSO4·7H2O, 5% alcohol (v/v), and 1% acetic acid (v/v). AAB adjusted to OD660 nm = 0.5 using a UV spectrophotometer (SYNERGY MX, Biotek, Winooski, Vermont, USA) was inoculated with 0.1% for single AAB and 1% for multiple AAB in a liquid medium. The optimal temperature was evaluated by inoculating single or multiple AAB in a liquid medium supplemented with 5% alcohol and incubating at 20, 30, or 40 °C. To find the optimal value, the initial pH of the liquid medium was adjusted to 2.0, 3.0, and 4.0 at room temperature (20 °C) using an Orion 3 Star pH meter (Thermo Fisher Scientific, Beverly, MA, USA). AAF ability was evaluated by measuring titratable acidity and growth of the samples at intervals of 3 days, and multiple starter candidates were selected.
2.3. Optimal Inoculation Rate and Inoculation Order for Multiple Starters
To evaluate their ratio of inoculation, multiple AAB were mixed at ratios of 1:1, 1:2, and 2:1, inoculated with 1% of the liquid medium, and cultured at 30 °C for 27 days. To evaluate the order of inoculation for multiple starters, simultaneous inoculation, pre–inoculation, and post–inoculation were performed at the previously selected optimal inoculation ratio. In pre–inoculation, the former strain was inoculated on day 0 of fermentation, and the following strain was inoculated on day 12 of fermentation. For post–inoculation, the order of the strains was reversed. After static culture at 30 °C for 27 days, titratable acidity and growth were measured at intervals of 6 days.
2.4. Titratable Acidity and Growth
Titratable acidity was measured as the amount of acetic acid (%) titrated with 0.1 N sodium hydroxide using 1% phenolphthalein as an indicator [18]. Growth was measured at OD660 using a UV spectrophotometer. The optimal alcohol concentration and initial pH of the isolates were evaluated by calculating the fermentation efficiency, which represents the pure acetic acid production amount as a percentage of the amount, according to the initial fermentation acidity [19]. The calculation formula is as follows:
2.5. Microbiome Taxonomic Profiling Analysis
Microbiome taxonomic profiling analysis was performed at CJ Bioscience using cell pellets consisting of 50 mL of the sample obtained via centrifugation at 10,000 rpm for 20 min (CJ Bioscience, Inc., Seoul, Korea). Total DNA was extracted using the FastDNA Spin kit (MP Biomedicals) according to the manufacturer’s instructions [20]. PCR amplification was performed using fusion primers targeting the V3–V4 regions of the 16S rRNA gene with the extracted DNA. The fusion primers 341F (5’-AAT GAT ACG GCG ACC ACC GAG ATC TAC AC – XXX XXX XX – TCG TCG GCA GCG TC – AGA TGT GTA TAA GAG ACA G – CCT ACG GGN GGC WGC AG –3’; the underlined sequence indicates the target region primer) and 805R (5’-CAA GCA GAA GAC GGC ATA CGA GAT – XXX XXX XX – GTC TCG TGG GCT CGG – AGA TGT GTA TAA GAG ACA G – GAC TAC HVG GGT ATC TAA TCC –3’) were used for bacterial amplification. The fusion primers were constructed in the following order: P5 (P7) graft-binding, i5 (i7) index, Nextera consensus, sequencing adaptor, and target region sequence. Amplifications were carried out under the following conditions: initial denaturation at 95 °C for 3 min, followed by 25 cycles of denaturation at 95 °C for 30 s, primer annealing at 55 °C for 30 s, and extension at 72 °C for 30 s, with a final elongation at 72 °C for 5 min. The PCR product was confirmed through 1% agarose gel electrophoresis and visualized using a Gel Doc system (BioRad, Hercules, CA, USA). The amplified products were purified using CleanPCR (CleanNA; Waddinxveen, Netherlands). Equal concentrations of purified products were pooled together, and short fragments (non–target products) were removed using CleanPCR. Quality and product size were assessed on a Bioanalyzer 2100 (Agilent, Palo Alto, CA, USA) using a DNA 7500 chip. Mixed amplicons were pooled, and sequencing was carried out at CJ Bioscience, Inc. using the Illumina MiSeq Sequencing system (Illumina, USA) according to the manufacturer’s instructions.
The secondary analysis which includes diversity calculation and biomarker discovery was conducted by in–house programs of CJ Bioscience, Inc. The alpha diversity indices (ACE, Chao1, Jackknife, Shannon, NPShannon, Simpson, and Phylogenetic diversity), rarefaction curves, rank abundance curves are estimated [21,22,23,24,25,26,27]. To visualize the sample differences, beta diversity distances were calculated by Generalized UniFrac algorithms [29,30,31,32], and exhibited as principal coordinates analysis (PCoA) and dendrogram. With functional profiles that are predicted by PICRUSt and MinPath algorithms [33,34], taxonomic biomarkers and functional biomarkers were discovered by statistical comparison algorithms (LDA Effect Size – LEfSe and Kruskal–Wallis H Test) [35,36]. All analytics mentioned above were performed in EzBioCloud 16S-based MTP, which is a CJ Bioscience’s bioinformatics cloud platform.
2.6. Volatile Compound using SPME Gas Chromatography
The 25 mL sample was collected and centrifuged at 10,000 rpm for 20 min to obtain the supernatant. The supernatant was filtered through a 0.2 μM syringe filter, and GC–MS analysis was performed using EZMASS (Jinju, Korea) [37]. After placing 3 mL of the sample in a headspace glass vial, 2–methyl–1–phentanol was injected as a designated internal standard, and the headspace vial containing the sample was equilibrated at 35 °C for 30 min. After exposure to the SPME fiber (DVB/CAR/PDMS) for 5 min, volatile compounds were collected and desorbed in a GC/MS injector at 200 °C for 5 min for analysis. The GC/MS system was GC–2010 Plus, GCMS–TQ 8030 (Shimadzu, Tokyo, Japan), and DB–WAX (30 mm × 0.25 mm i.d., 0.25 μm film thickness, J&W, CA, USA) was used for the column. After incubation at 40 °C for 3 min, the temperature was increased to 90 °C at a rate of 5 °C/min, heated to 230 °C at a rate of 19 °C/min, and maintained for 5 min. The injection temperature was set to 250 °C, the carrier gas was He, and the flow rate was 1.0 mL/min.
2.7. Statistical Analysis
One–way analysis of variance, followed by Duncan’s multiple range test, was used to evaluate the significance of the differences in triplicate experiments. Statistical significance was set at P < 0.05. All statistical analyses were performed using SPSS version 25.0 (SPSS Corp., Armonk, NY, USA). Multivariate statistical analyses, including heatmap and partial least squares–discriminant analysis (PLS–DA), were performed using open–source R software (ver. 4.2.1). Metabolite analysis was performed and visualized using the “pheatmap” (ver. 1.0.12) and “plsda” packages, respectively.
3. Results and Discussion
3.1. Changes in Titratable Acidity and AAB Growth According to Fermentation Temperature, Alcohol Concentration, and Initial pH
To evaluate the optimal conditions for the six AAB strains isolated from grain vinegars from various regions in Korea, the acid–producing ability and growth according to the fermentation period were investigated in a liquid medium containing 5% alcohol. Isolates fermented at 20, 30, and 40 °C for 15 days showed the highest acid–producing ability at 30 °C (Table 2). The isolates underwent an induction period until day 3 of fermentation, and the titratable acidity increased from day 6 of fermentation. On day 15 of fermentation at 30 °C, the six isolates showed an average titratable acidity of 5.53 ± 0.13%, and the GV–5, GV–12, GV–17, and GV–16 strains showed an especially high increase. When fermented at 20 °C and 40 °C, there was no increase in titratable acidity, but insignificant increase was observed when GV–12 was fermented at 40 °C. In contrast, growth increased the most when fermented at 20 °C. When fermented at 30 °C, the growth on day 15 of fermentation increased approximately three times, whereas growth increased about 21 times when fermented at 20 °C, compared to the beginning of fermentation (day 0). AAB are known to grow by decomposing alcohol to produce acetic acid [38]; therefore, it is possible that the sample with no increase in titratable acidity fermented at 20 °C showed an increase in contaminating bacteria growth. However, since the initial medium before inoculation with AAB contained 5% ethanol and 1% acetic acid, the probability of bacterial growth contamination was considered to be low. In addition, when fermented at 40 °C, AAB did not grow, similar to the titratable acidity. Therefore, it was determined that the six isolates showed optimal acid production and growth at 30 °C.
To evaluate the optimal alcohol concentration for the six isolates with high acid–producing ability, the isolates were fermented at 30 °C for 15 days in a liquid medium with an alcohol concentration of 5–14%, and then changes in titratable acidity and growth were measured (Table 3). The isolates showed a high increase in titratable acidity at 5% and 8% alcohol, and the growth increased similarly, but they did not grow at 11% or higher alcohol concentrations. At 5 and 8% alcohol, the six isolates showed average titratable acidities of about 5.53 ± 0.13% and 6.30 ± 0.79% after 15 days of fermentation, respectively 4.35 and 4.85 times higher than those at the beginning of fermentation (day 0). In other words, the titratable acidity of the 8% alcohol concentration was higher. On the other hand, the average fermentation efficiency of the isolates at 5% and 8% alcohol concentrations was 65.31% and 47.88%, respectively, exhibiting higher fermentation efficiency at 5% alcohol concentration (Table S1). This suggests that the ability to oxidize alcohol into acetic acid was reduced at 8% alcohol. Therefore, the optimal alcohol concentration for the six isolates was determined to be 5%, where the GV–5, GV–12, and GV–17 strains showed high acid production. At 8% alcohol concentration, the GV–12 strain showed high acid production ability, exhibiting alcohol tolerance.
To identify the optimal pH of the isolates, AAB were inoculated into a liquid medium adjusted to pH 2.0–4.0 and fermented at 30 °C for 15 days. Then, the acid production ability and growth were investigated. The initial acidity was increased by decreasing pH (Acidity as pH decreases: pH 2.0, 5.51%; pH 3.0, 3.24%; pH 4.0, 0.37%) (Table 4). The six isolates produced acetic acid while growing at pH 3.0 and 4.0, but failed to grow at pH 2.0. In particular, isolates showed a high average titratable acidity of 6.92 ± 1.02% at pH 4.0 on day 9, which decreased as the fermentation period elapsed. The growth increased 15.8 times on day 15, compared with day 0. Isolates showed an average titratable acidity of 3.58 ± 1.13% at pH 3.0 on day 9, and the growth increased 8 times on day 15. The average fermentation efficiency on day 9 of fermentation was 53.70 ± 17.50% and 105.80 ± 15.74% at pH 3.0 and 4.0, respectively (Table S1). This suggests that the ability of the isolates to oxidize ethanol to acetic acid increases in an environment of pH 4.0; therefore, pH 4.0 was considered optimal for the strains. In particular, the GV–8, GV–12, GV–22, and GV–5 strains showed high acid–producing ability at pH 4.0.
3.2. Selecting Multiple Starter Candidates for AAF
Evaluating the optimal growth conditions of the six isolates showed the highest acid–producing ability when fermented at 30 °C for 15 days in a liquid medium containing 5% alcohol and adjusted to pH 4.0. GV–12 and GV–17 showed high acid–producing ability and alcohol tolerance in a liquid medium containing 8% alcohol. Moreover, GV–8 and GV–22 showed high acid–producing ability in a liquid medium adjusted to pH 4.0. Therefore, the GV–8 (A), GV–12 (B), GV–17 (C), and GV–22 (D) strains were selected as multiple starter candidates for AAF (Table 1).
3.3. Changes in Titratable Acidity and Growth of Multiple AAB According to Fermentation Temperature and Initial pH
Titratable acidity and growth were measured in a liquid medium containing 5% alcohol and fermented at various temperatures (20, 30, and 40 °C) for 27 days to evaluate the optimal growth conditions and investigate the acid–producing ability of multiple AAB according to the fermentation environment (Figure 1). The seven multiple AAB starters were as follows: AB (A. ascendens GV–8 and A. ascendens GV–12), AC (A. ascendens GV–8 and A. pasteurianus GV–17), AD (A. ascendens GV–8 and A. pasteurianus GV–22), BC (A. ascendens GV–12 and A. pasteurianus GV–17), BD (A. ascendens GV–12 and A. pasteurianus GV–22), CD (A. pasteurianus GV–17 and A. pasteurianus GV–22), and ABCD (mixture of A, B, C, and D starters; control). Multiple AAB showed the highest increase in titratable acidity and growth at 30 °C, similar to single AAB (Figure 1B,E). After 27 days at 30 °C, the average titratable acidity of six AAB was 4.78 ± 0.99%, and the growth increased 10 times compared to day 0. In particular, the AD and AB starters showed high titratable acidity after 27 days of fermentation by oxidizing ethanol to acetic acid as the fermentation period passed after the 9–day induction. In contrast, a single AAB did not produce acid at 20 °C, whereas multiple AAB did. After 27 days at 20 °C, multiple AAB showed an average titratable acidity of 4.01 ± 0.54%, and the growth increased 8.25 times, compared to day 0 (Figure 1A,D). The AC and BD starters showed high titratable acidity on day 27 after a 12–day induction period. In addition, multiple AAB failed to grow and produce acid at 40 °C (Figure 1C,F).
Multiple starters were inoculated into a liquid medium with initial pH adjusted to 2.0–4.0 and fermented at 30 °C for 27 days to measure the titratable acidity and growth of multiple AAB according to the initial pH (Figure 2). The initial acidity was increased by decreasing pH (Acidity as pH decreases: pH 2.0, 5.24%; pH 3.0, 3.24%; pH 4.0, 0.35%) (Figure 2a–c). The six AAB starters, except ABCD starter, produced acetic acid through ethanol oxidation after a 6-day induction period at pH 3.0, and the titratable acidity rapidly increased during days 9–21. At pH 3.0, the titratable acidity increased to 3.94 ± 0.20%, the growth increased 7–fold (Figure 2b,e), and the average fermentation efficiency was 59.35 ± 3.37% (Table S2). On the other hand, at pH 4.0, the titratable acidity gradually increased after an induction period of 6 days, showing a 3.51 ± 0.87% on day 27 (Figure 2c,f). Growth increased 10.5 times, and the average fermentation efficiency was 53.13 ± 13.45% on the 27th day of fermentation (Table S2). Single AAB showed high acetic acid production ability at pH 4.0 (Table 4), whereas multiple AAB showed high activity at pH 3.0. Growth and acid production were not observed at pH 2.0 (Figure 2a,d). In contrast, ABCD showed high acid–producing ability at both pH 3.0 and 4.0.
3.4. Optimal Inoculation Rate and Order of Multiple AAB
To induce high acid production through the interaction of multiple AAB, the inoculation rate of multiple AAB was evaluated. Former and following strains were mixed at ratios of 1:1, 1:2, and 2:1, inoculated into a liquid medium, and fermented at 30 °C. The six AAB were as follows: AB (A. ascendens GV-8 and A. ascendens GV-12), AC (A. ascendens GV-8 and A. pasteurianus GV-17), AD (A. ascendens GV-8 and A. pasteurianus GV-22), BC (A. ascendens GV-12 and A. pasteurianus GV-17), BD (A. ascendens GV-12 and A. pasteurianus GV-22), and CD (A. pasteurianus GV-17 and A. pasteurianus GV-22). The increase in titratable acidity and growth showed a similar trend (Figure 3). On day 27, the titratable acidity of AB, AC, and AD was the highest at a ratio of 1:1 (5.59 ± 0.23%, 5.62 ± 0.14%, and 5.64 ± 0.02%, respectively), and the titratable acidity at a ratio of 1:2 was higher than that at a 2:1 ratio, while both strains showed similar acid–producing effects. BC showed the highest titratable acidity of 5.55 ± 0.37% at a ratio of 2:1, followed by 1:1 and 1:2. In other words, the B (GV-12) strain contribution to the increase in titratable acidity was greater. BD showed the highest titratable acidity of 5.94 ± 0.12% at a ratio of 2:1, followed by 1:2 and 1:1. B and D may inhibit each other and interfere with growth when they are co–cultured. The titratable acidity of the CD starter was the highest at 5.25 ± 0.22% at a ratio of 1:1, followed by 2:1 and 1:2, showing that C and D may have a similar effect on the increase in titratable acidity. The optimal inoculation ratio for each AAB was as follows: AB, 1:1; AC, 1:1; AD, 1:1; BC, 2:1; BD, 2:1; and CD, 1:1.
Multiple AAB strains were inoculated in different succession to identify the optimal inoculation order (Figure 4). For simultaneous inoculation, the former and following strains were simultaneously inoculated on day 0 (Symbol in Figure 4: 0). For pre–inoculation, the former strain was inoculated on day 0, and the following strain was inoculated on day 12 (Symbol in Figure 4: 1st). Post–inoculation reversed the positions of each strain in pre–inoculation (Symbol in Figure 4: 2nd). AB, AC, AD, and CD starters inoculated at a ratio of 1:1 showed the highest titratable acidity post–inoculation (5.73 ± 0.12%, 6.38 ± 0.11%, 6.59 ± 0.11%, 6.82 ± 0.11%, respectively), followed by simultaneous inoculation and pre–inoculation. BC and BD inoculated at a ratio of 2:1 showed the highest titratable acidity post–inoculation (7.01 ± 0.14% and 6.55 ± 0.14%, respectively), followed by pre–inoculation and simultaneous inoculation. This implies that multiple AAB are advantageous when the two strains are inoculated at different times. It can be interpreted that metabolites produced by the first inoculated strain had a beneficial effect on the latter inoculated strain.
3.5. Volatile Flavor Pattern Profiling According to Fermentation Temperature and Multiple Starters
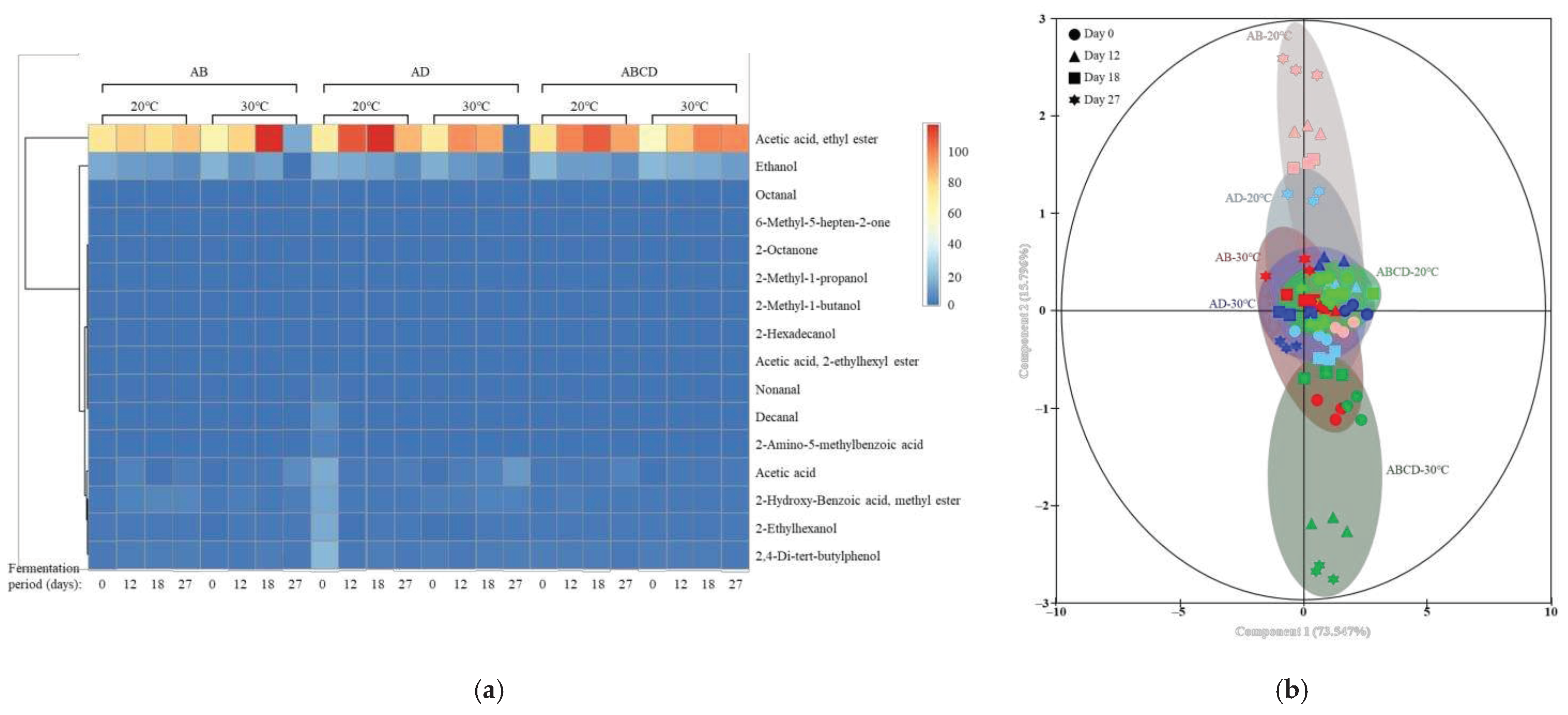
In general, the flavor of vinegar is produced by the complex action of acids, aldehydes, alcohols, ketones, and ester compounds, including acetic acid, owing to the interaction between fermenting microorganisms, resulting in various volatile components via raw material processing, fermentation, and maturation [39]. Analyzing volatile components in vinegar prepared at 20 and 30 °C detected 16 types of volatile components (Figure 5 and Table S3–S5). The main volatile components were acetic acid and alcohol, and the former was most abundant in AD–20 °C. As AAF progresses, AAB consume alcohol to produce acetic acid, so the alcohol content decreases, and the acetic acid content increases. Total volatile compounds were highest when fermented at a low temperature (20 °C), rather than at a medium temperature (30 °C). AB, AD, and ABCD showed the highest titratable acidity on days 12, 18, and 18, respectively. The levels of decanal, which has a citrus flavor, and nonanal (γ-nonalactone), which has a coconut and almond–like flavor, increased as fermentation progressed [40,41]. Decanal and nonanal showed the highest increase at ABCD–30 °C, and AD–30 °C, respectively. Octanal, 6–methyl–5–hepten–2–one, and 2–ethylhexyl acetate (acetic acid, 2–ethylhexyl ester), which have a citrus flavor, and 2–octanone (octanoic acid), which has a sweet flavor, were detected in trace amounts in all samples [42,43,44,45].
A heatmap was used to visualize the detection of volatile compounds according to the fermentation temperature and multiple starters (Figure 5a). Red coloring in the heatmap indicates a large amount detected in GC-MS quantitative analysis, and blue coloring indicates a small amount. A large amount of acetic acid was detected in AB at 30 °C on day 18 compared to day 27 of fermentation, consistent with its acid–producing ability. Acetic acid is produced during the bioconversion of ethanol through two reactions catalyzed by membrane–bound pyrroloquinoline quinone-dependent alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) [46]. ADH oxidizes ethanol to acetaldehyde, which is then converted to acetic acid by ALDH and is released into the environment. These two dehydrogenase complexes are strictly connected to the respiratory chain, which transfers electrons to oxygen through ubiquinone and acts as the final electron acceptor. Acetic acid produced by partial ethanol oxidation can be further oxidized in the cytosol by a series of soluble NaD(P)+-dependent dehydrogenases (ADH and ALDH) via the tricarboxylic acid cycle, resulting in acetate oxidation (peroxidation). Acetobacter spp. are known for their acetate peroxidation activity that can oxidize acetic acid, in addition to its ability to accumulate acetic acid [5]. It seems that volatile acetic acid was lost as the fermentation period elapsed in the AB–30 °C treatment group. For AD, higher acetic acid levels were detected on day 18, which decreased on day 27 at 20 °C. For AB and AD starters, the titratable acidity was slightly higher when fermented at 30 °C, while the measured volatile compounds were higher when fermented at 20 °C. For the ABCD multiple starter, similar amounts of acetic acid were detected at 20 °C and 30 °C, but a lower volatile compounds content was detected, compared to that in the AB and AD starters. Therefore, AB fermented at 30 °C for 18 days and AD fermented at 20 °C for 18 days were judged to have a high flavor component–producing ability.
PLS-DA was used to identify the pattern of volatile compounds according to the fermentation temperature and multiple starters (Figure 5b). Most factors were located in the center of the plot at the beginning of fermentation (days 0–12). For AB, the factors at 20 °C moved to the upper part of the plot as fermentation progressed, and those at 30 °C were located in the 4th quadrant of the plot at the beginning of fermentation and moved to the center of the plot as fermentation progressed. For AD, the factors at 20 °C moved upward in the plot according to the fermentation process, and the factors at 30 °C were in the center of the plot, regardless of the fermentation period. The ABCD factors at 30 °C were in the center of the plot, regardless of the fermentation period, and the factors at 20 °C moved to the lower part of the plot on day 27 of fermentation. Vinegar taste and flavor components originate from the unique characteristics of its raw materials; moreover, differences in volatile compounds appear depending on fermentation and aging methods [47]. In this study, the fermentation temperature and type of AAB affected the flavor composition.
Figure 5.
Figure 5. (a) Heatmap and (b) partial least squares–discriminant analysis (PLS–DA) of vinegar samples fermented by multiple AAB according to the fermentation temperature and period (days). The blue color in the heatmap corresponds to low abundance, and red corresponds to high abundance, as shown in the color key legend on the right side of the figure. Low abundance indicates low detection of volatile compounds. Multivariate PLS–DA displays different colors according to complex AAB and fermentation temperature. The factors indicated by each color are indicated in the figure. Strains: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
Figure 5.
Figure 5. (a) Heatmap and (b) partial least squares–discriminant analysis (PLS–DA) of vinegar samples fermented by multiple AAB according to the fermentation temperature and period (days). The blue color in the heatmap corresponds to low abundance, and red corresponds to high abundance, as shown in the color key legend on the right side of the figure. Low abundance indicates low detection of volatile compounds. Multivariate PLS–DA displays different colors according to complex AAB and fermentation temperature. The factors indicated by each color are indicated in the figure. Strains: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
3.5. Changes in the Beta Diversity of Microbial Communities According to Fermentation Temperature and Multiple Starters
As AAF progresses, dominance between multiple AAB and microorganism diversity change, and microorganisms worsen the fermentation quality can grow. Microbiome taxonomic profiling was performed to confirm the microbial diversity of co–cultured AAB according to fermentation temperature and duration and to control the microbial community of grain vinegar through AAF [48]. The alpha diversity index determines whether sequencing data are sufficient for detecting all microbial species included in the sample [49]. Alpha diversity refers to the diversity “within” a sample; the higher the number of species (abundance) and the more evenly distributed (evenness), the higher the species diversity [50]. In this study, alpha diversity showed similar values, because only the starter was inoculated and fermented in a strictly controlled environment (Table S6).
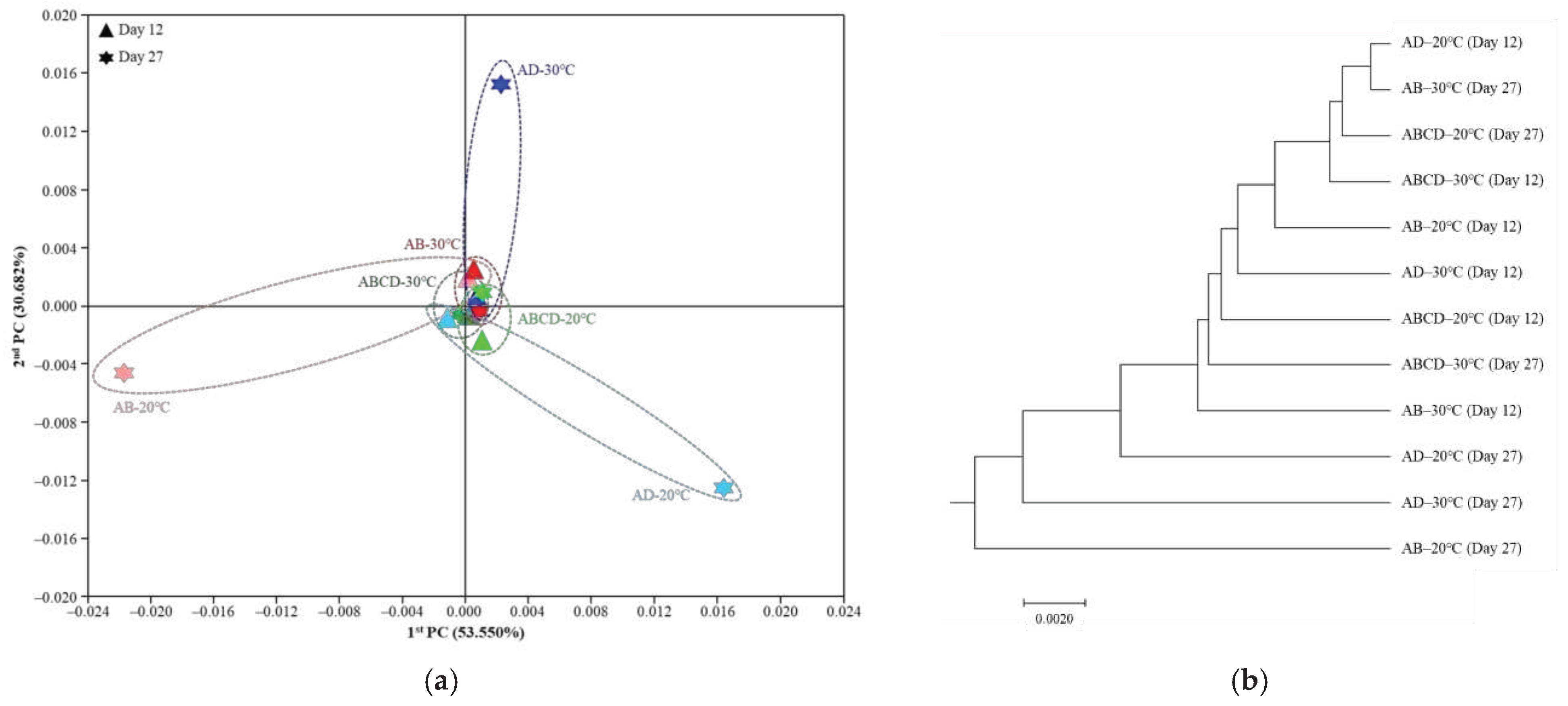
Alpha diversity analysis revealed no significant difference in the diversity within the sample. Therefore, beta diversity was analyzed to investigate the diversity of the samples. The distance metric of beta diversity measures the diversity between samples using the Generalized UniFrac method, where higher values indicate a more diverse microbiome composition [51,52]. When the samples on days 12 and 27 of fermentation were compared, PC1 and PC2 had explanatory powers of 56.550% and 30.682%, respectively, in Generalized UniFrac (Figure 5a). All samples collected at the beginning of fermentation were located at the center of the PCoA. AD–20 °C moved to the 1st quadrant of the PCoA as fermentation progressed, and AD–30 °C moved to the 4th quadrant. In other words, AD results suggest that microbial diversity changes with the fermentation temperature. In addition, AB–20 °C, which was located in the center of the PCoA at the beginning of fermentation, moved to the third quadrant as fermentation progressed, and AB–30 °C was located in the center without change. Differences in microbial diversity according to the fermentation temperature of AB were suggested. Similarly, the day 27 samples of AB–20 °C, AD–20 °C, and AD–30 °C appeared to have a higher scale in the dendrogram, compared to the sample located in the center of the PCoA (Figure 5b). Therefore, a significant difference in beta diversity was observed according to the fermentation temperature and period, and AB–20 °C, AD–20 °C, and AD–30 °C were considered to have a different AAB community composition, compared to other samples. This implies that fermentation temperature and duration must be considered in the production of fermented vinegar.
Figure 6.
Principal coordinates analysis (PCoA, a) and dendrogram (b) of the Generalized UniFrac of multiple starters fermented for 12 and 27 days. Strains: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, and A. pasteurianus GV–22.
Figure 6.
Principal coordinates analysis (PCoA, a) and dendrogram (b) of the Generalized UniFrac of multiple starters fermented for 12 and 27 days. Strains: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, and A. pasteurianus GV–22.
4. Conclusions
In conclusion, this study evaluated the effect of multiple AAB on the quality of traditional vinegar. The use of starters for AAF can help standardize the taste and quality of the final product and control microorganism community diversity [52]. In this study, the acid–producing ability and amount of volatile aroma components were measured according to the fermentation temperature and type of AAB, which showed a correlation with the beta diversity of the microbial community. The titratable acidity of vinegar measured when using a single AAB was 5.53 ± 0.13% and 6.51 ± 0.44% when using multiple AAB. This suggests that using multiple AAB to prepare traditional vinegar may help increase titratable acidity. AAF using two AAB types in combination, rather than four, was advantageous for increasing titratable acidity and volatile compound content. When fermented at the optimal fermentation temperature for AAF (30 °C), the titratable acidity increased significantly due to AAB oxidation. When fermented at a low temperature (20 °C), the volatile compounds of multiple AAB tended to increase. AD (A. ascendens GV–8 and A. pasteurianus GV–22) fermented at 20 °C had the most volatile components, showing a specific pattern in beta diversity analysis. Therefore, we propose the application of multiple AAB with acid–producing and flavor–producing abilities to prepare traditional vinegar. Meanwhile, this study has a limitation in that the alcohol fermentation process by yeast was omitted. Therefore, several studies have been required on traditional vinegar using Makgeolli produced by yeast and profiling its metabolites.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Fermentation efficiency according to concentration of ethanol and initial pH. Table S2: Fermentation efficiency under multiple starters according to initial pH and fermentation period. Table S3: Volatile compounds according to fermentation period using Acetobacter ascendens GV-8 (A) and A. ascendens GV-12 (B) (Unit: Area%). Table S4: Volatile compounds according to fermentation period using Acetobacter ascendens GV-8 (A) and Acetobacter pasteurianus GV-22 (D) (Unit: Area%). Table S5: Volatile compounds according to fermentation period using A. ascendens GV-8 (A), A. ascendens GV-12 (B), A. pasteurianus GV-17 (C), and A. pasteurianus GV-22 (D) (Unit: Area%). Table S6: Alpha diversity analysis under multiple starters and fermentation temperature.
Author Contributions
This work was carried out in collaboration with all authors. S.-H.Y. designed the study. H.K. performed the statistical analysis, wrote the protocol, and wrote the first draft of the manuscript. Authors S.H.K., W.S.J., and S.-Y.K. managed the analyses of the study and literature searches. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Program for Agricultural Science and Technology Development (Project No. PJ01600601) and the National Institute of Agricultural Science, Rural Development Administration, Republic of Korea.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The financial support of the Rural Development Administration is gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nanda, K.; Taniguchi, M.; Ujike, S.; Ishihara, N.; Mori, H.; Ono, H.; Murooka, Y. Characterization of acetic acid bacteria in traditional acetic acid fermentation of rice vinegar (Komesu) and unpolished rice vinegar (Kurosu) produced in Japan. Appl. Environ. Microbiol. 2001, 67(2), 986–990. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Kim, S. H.; Jeong, W. S.; Kim, S. Y.; Yeo, S. H. Microbiome analysis of traditional grain vinegar produced under different fermentation conditions in various regions in Korea. Foods 2022, 11(22), 3573. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, T.; Giudici, P.; Chen, F. Vinegar functions on health: Constituents, sources, and formation mechanisms. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1124–1138. [Google Scholar] [CrossRef]
- Kim, K. O.; Kim, S. M.; Kim, S. M.; Kim, D. Y.; Jo, D.; Yeo, S. H.; Jeong, Y. J.; Kwon, J. H. Physicochemical properties of commercial fruit vinegars with different fermentation methods. J. Korean Soc. Food Sci. Nutr. 2013, 42(5), 736–742. [Google Scholar] [CrossRef]
- Kanchanarach, W.; Theeragool, G.; Inoue, T.; Yakushi, T.; Adachi, O.; Matsushita, K. Acetic acid fermentation of Acetobacter pasteurianus: relationship between acetic acid resistance and pellicle polysaccharide formation. Biosci. Biotechnol. Biochem. 2010, 74(8), 1591–1597. [Google Scholar] [CrossRef]
- Li, S.; Li, P.; Liu, X.; Luo, L.; Lin, W. Bacterial dynamics and metabolite changes in solid–state acetic acid fermentation of Shanxi aged vinegar. Appl. Microbiol. Biotechnol. 2016, 100, 4395–4411. [Google Scholar] [CrossRef]
- Fleet, G. H. Microorganisms in food ecosystems. Int. J. Food Microbiol. 1999, 50(1), 101–117. [Google Scholar] [CrossRef] [PubMed]
- Gullo, M.; Giudici, P. Acetic acid bacteria in traditional balsamic vinegar: phenotypic traits relevant for starter cultures selection. Int. J. Food Microbiol 2008, 125, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Vegas, C.; Mateo, E.; Tesfaye, W.; Cerezo, A.; Callejón, R.; Poblet, M.; Guillamón, J.; Mas, A.; Torija, M. Effect of barrel design and the inoculation of Acetobacter pasteurianus in wine vinegar production. Int. J. Food Microbiol 2010, 141, 56–62. [Google Scholar] [CrossRef]
- Wu, J. J.; Ma, Y. K.; Zhang, F. F.; Chen, F. S. Biodiversity of yeasts, lactic acid bacteria and acetic acid bacteria in the fermentation of “Shanxi aged vinegar”, a traditional Chinese vinegar. Food Microbiol 2012, 30, 289–297. [Google Scholar] [CrossRef]
- Ohmori, S.; Masai, H.; Arima, K.; Beppu, T. Isolation and identification of acetic acid bacteria for submerged acetic acid fermentation at high temperature. Agric. Biol. Chem. 1980, 44(12), 2901–2906. [Google Scholar] [CrossRef]
- Astudillo–Melgar, F.; Ochoa–Leyva, A.; Utrilla, J.; Huerta–Beristain, G. Bacterial diversity and population dynamics during the fermentation of palm wine from Guerrero Mexico. Front. Microbiol. 2019, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A. E.; Rebolleda–Gómez, M.; Benítez, M.; Travisano, M. Ecological perspectives on synthetic biology: insights from microbial population biology. Front. Microbiol. 2015, 6, 143. [Google Scholar] [CrossRef]
- Prathiviraj, R.; Rajeev, R.; Jose, C. M.; Begum, A.; Selvin, J.; Kiran, G. S. Fermentation microbiome and metabolic profiles of Indian palm wine. Gene Rep. 2022, 27, 101543. [Google Scholar] [CrossRef]
- Su, X. Elucidating the beta–diversity of the microbiome: from global alignment to local alignment. Msystems 2021, 6(4), e00363–21. [Google Scholar] [CrossRef] [PubMed]
- Baek, S. Y.; Park, H. Y.; Lee, C. H.; Yeo, S. H. Comparison of the fermented property and isolation of acetic–acid bacteria from traditional Korean vinegar. Korean J. Food Preserv. 2014, 21(6), 1738–7248. [Google Scholar] [CrossRef]
- Baek, C. H.; Baek, S. Y.; Lee, S. H.; Kang, J. E.; Choi, H. S.; Kim, J. H.; Yeo, S. H. Characterization of Acetobacter sp. strain CV1 isolated from a fermented vinegar. Microbiol. Biotechnol. Lett. 2015, 43(2), 126–133. [Google Scholar] [CrossRef]
- Gil, N. Y.; Jang, Y. J.; Gwon, H. M.; Jeong, W. S.; Yeo, S. H.; Kim, S. Y. Comparative evaluation of quality and metabolite profiles in Meju using starter cultures of Bacillus velezensis and Aspergillus oryzae. Foods 2022, 11(1), 68. [Google Scholar] [CrossRef]
- Kang, S. K.; Jang, M. J.; Kim, Y. D. Isolation and culture conditions of Acetobacter sp. for the production of citron (Citrus junos) vinegar. Korean J. Food Preserv 2006, 13(3), 357–362. [Google Scholar]
- Burbach, K.; Seifert, J.; Pieper, D. H.; Camarinha-Silva, A. Evaluation of DNA extraction kits and phylogenetic diversity of the porcine gastrointestinal tract based on Illumina sequencing of two hypervariable regions. MicrobiologyOpen 2016, 5(1), 70–82. [Google Scholar] [CrossRef]
- Chao, A.; Lee, S. M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87(417), 210–217. [Google Scholar] [CrossRef]
- Chao, A. Estimating the population size for capture–recapture data with unequal catchability. Biometrics 1987, 783–791. [Google Scholar] [CrossRef]
- Burnham, K. P.; Overton, W. S. Robust estimation of population size when capture probabilities vary among animals. Ecology 1979, 60(5), 927–936. [Google Scholar] [CrossRef]
- Magurran, A. E. Measuring biological diversity. Curr. Biol. 2021, 31(19), R1174–R1177. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Shen, T. J. Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample. Environ. Ecol. Stat 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Faith, D. P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61(1), 1–10. [Google Scholar] [CrossRef]
- Heck, K. L.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56(6), 1459–1461. [Google Scholar] [CrossRef]
- Whittaker, R. H. Dominance and diversity in land plant communities. Science 1965, 147, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Lin, J. Divergence measures based on the Shannon entropy. IEEE Trans. Inf. Theory 1991, 37, 145–151. [Google Scholar] [CrossRef]
- Beals, E. W. Bray–Curtis ordination: an effective strategy for analysis of multivariate ecological data. Adv. Ecol. Res. 1984, 14, 1–55. [Google Scholar] [CrossRef]
- Chen, J.; Bittinger, K.; Charlson, E. S.; Hoffmann, C.; Lewis, J.; Wu, G. D.; Li, H. (2012). Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- Hamady, M.; Lozupone, C.; Knight, R. Fast UniFrac: facilitating high–throughput phylogenetic analyses of microbial communities including analysis of pyrosequencing and PhyloChip data. ISME J. 2010, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Doak, T. G. A parsimony approach to biological pathway reconstruction/inference for genomes and metagenomes. PLoS Comput. Biol. 2009, 5, e1000465. [Google Scholar] [CrossRef] [PubMed]
- Langille, M. G.; Zaneveld, J.; Caporaso, J. G.; McDonald, D.; Knights, D.; Reyes, J. A.; Beiko, R. G. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W. S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Kruskal, W. H.; Wallis, W. A. Use of ranks in one–criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Lee, S. J.; Kim, S. H.; Kim, S. Y.; Yeo, S. H. Quality characteristics of Kujippong (Cudrania tricuspidata) vinegar fermented by various acetic acid bacteria. Korean J. Food Preserv. 2019, 26(7), 766–776. [Google Scholar] [CrossRef]
- Lynch, K. M.; Zannini, E.; Wilkinson, S.; Daenen, L.; Arendt, E. K. Physiology of acetic acid bacteria and their role in vinegar and fermented beverages. Compr. Rev. Food Sci. Food Saf. 2019, 18(3), 587–625. [Google Scholar] [CrossRef] [PubMed]
- Longo, M. A.; Sanromán, M. A. Production of food aroma compounds: microbial and enzymatic methodologies. Food Technol. Biotechnol. 2006, 44(3), 335–353. [Google Scholar] [CrossRef]
- Sawamura, M.; Thi Minh Tu, N.; Onishi, Y.; Ogawa, E.; Choi, H. S. Characteristic odor components of Citrus reticulata Blanco (Ponkan) cold–pressed oil. Biosci. Biotechnol. Biochem. 2004, 68(8), 1690–1697. [Google Scholar] [CrossRef]
- Rocha, S. M.; Rodrigues, F.; Coutinho, P.; Delgadillo, I.; Coimbra, M. A. Volatile composition of Baga red wine: Assessment of the identification of the would–be impact odourants. Anal. Chim. Acta. 2004, 513(1), 257–262. [Google Scholar] [CrossRef]
- Feng, S.; Suh, J. H.; Gmitter, F. G.; Wang, Y. Differentiation between flavors of sweet orange (Citrus sinensis) and mandarin (Citrus reticulata). J. Agric. Food Chem. 2018, 66(1), 203–211. [Google Scholar] [CrossRef]
- Tandon, K. S.; Baldwin, E. A.; Shewfelt, R. L. Aroma perception of individual volatile compounds in fresh tomatoes (Lycopersicon esculentum, Mill.) as affected by the medium of evaluation. Postharvest Biol. Technol, 2000; 20, 3, 261–268. [Google Scholar] [CrossRef]
- Bett-Garber, K. L.; Bryant, R. J.; Grimm, C. C.; Chen, M. H.; Lea, J. M.; McClung, A. M. Physicochemical and sensory analysis of US rice varieties developed for the basmati and jasmine markets. Cereal Chem. 2017, 94(3), 602–610. [Google Scholar] [CrossRef]
- Su, M. S.; Chien, P. J. Aroma impact components of rabbiteye blueberry (Vaccinium ashei) vinegars. Food Chem. 2010, 119(3), 923–928. [Google Scholar] [CrossRef]
- Gullo, M.; Verzelloni, E.; Canonico, M. Aerobic submerged fermentation by acetic acid bacteria for vinegar production: Process and biotechnological aspects. Process Biochem. 2014, 49(10), 1571–1579. [Google Scholar] [CrossRef]
- Jo, Y.; Gu, S. Y.; Chung, N.; Gao, Y.; Kim, H. J.; Jeong, M. H.; Jeong, Y. J.; Kwon, J. H. Comparative analysis of sensory profiles of commercial cider vinegars from Korea, China, Japan, and US by SPME/GC–MS, E-nose, and E-tongue. Korean J. Food Sci. Technol. 2016, 48, 430–436. [Google Scholar] [CrossRef]
- Seo, H.; Lee, S.; Park, H.; Jo, S.; Kim, S.; Rahim, M. A.; Ul-Haq, A.; Barman, I.; Lee, Y.; Seo, A.; Kim, M.; Jung, I. Y.; Song, H. Y. Characteristics and microbiome profiling of Korean Gochang bokbunja vinegar by the fermentation process. Foods 2022, 11(20), 3308. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, L.; Osman, O. A.; Bertilsson, S.; Eiler, A. Microbial community composition and diversity via 16S rRNA gene amplicons: evaluating the illumina platform. PloS one 2015, 10(2), e0116955. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, T. D. A unified mathematical framework for the measurement of richness and evenness within and among multiple communities. Oikos 2004, 104(2), 377–387. [Google Scholar] [CrossRef]
- Tawfik, S. A.; Azab, M. M.; Ahmed, A. A. A.; Fayyad, D. M. Illumina MiSeq sequencing for preliminary analysis of microbiome causing primary endodontic infections in Egypt. Int. J. Microbial. 2018, 2018, 2837328. [Google Scholar] [CrossRef]
- Dai, Y.; Tian, Z.; Meng, W.; Li, Z. Microbial diversity and physicochemical characteristics of the Maotai-flavored liquor fermentation process. J. Nanosci. Nanotechnol. 2020, 20(7), 4097–4109. [Google Scholar] [CrossRef]
- Wang, Z. M.; Lu, Z. M.; Yu, Y. J.; Li, G. Q.; Shi, J. S.; Xu, Z. H. Batch-to-batch uniformity of bacterial community succession and flavor formation in the fermentation of Zhenjiang aromatic vinegar. Food Microbiol. 2015, 50, 64–69. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Changes in titratable acidity (a–c) and growth (d–f) under multiple starters according to fermentation temperature and period. Fermentation temperature: (a) and (d) 20 °C; (b) and (e) 30 °C; (c) and (f) 40 °C. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
Figure 1.
Changes in titratable acidity (a–c) and growth (d–f) under multiple starters according to fermentation temperature and period. Fermentation temperature: (a) and (d) 20 °C; (b) and (e) 30 °C; (c) and (f) 40 °C. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
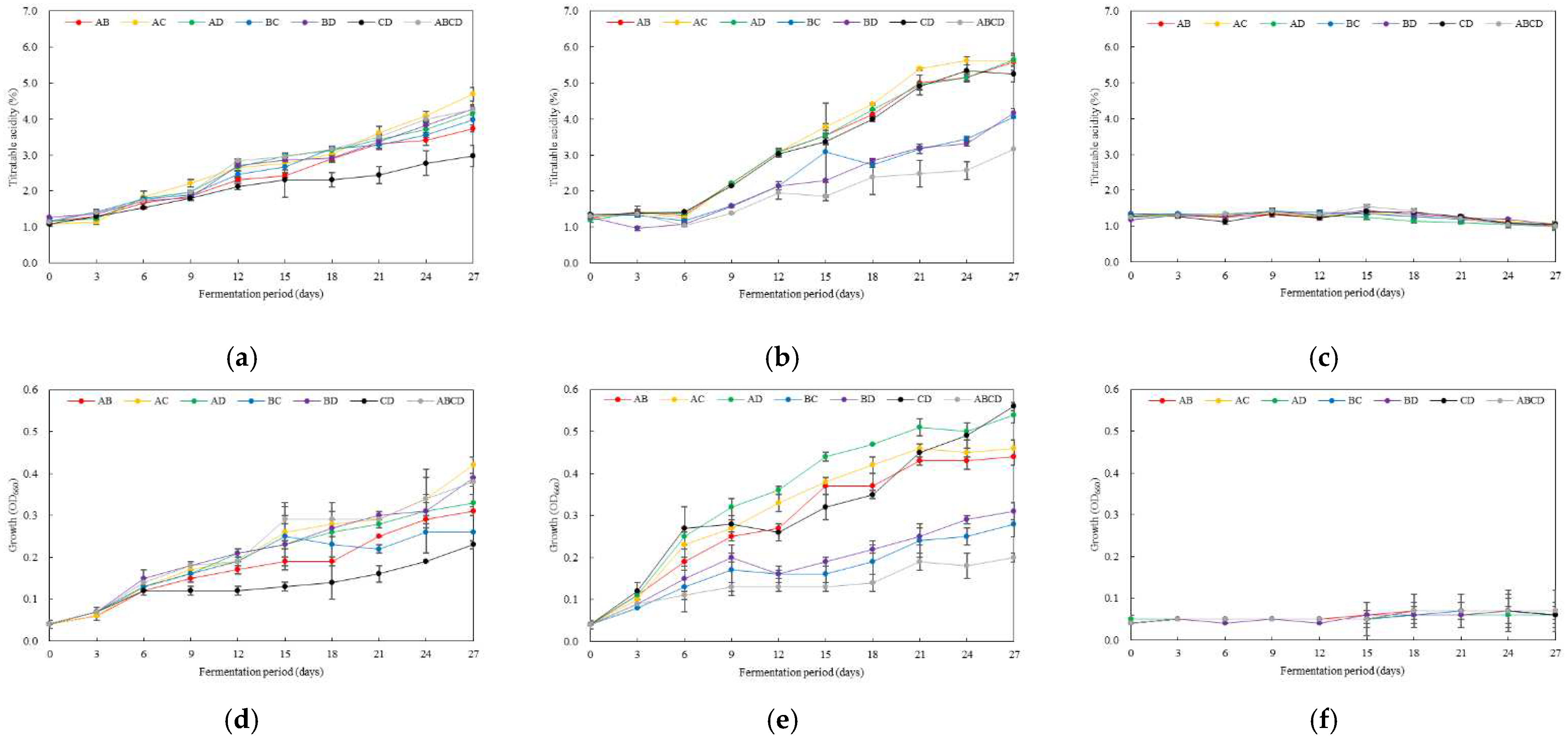
Figure 2.
Changes in titratable acidity (a–c) and growth (d–f) under multiple starters according to initial pH and fermentation period. Initial pH: (a) and (d) pH 2.0; (b) and (e) pH 3.0; (c) and (f) pH 4.0. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22. The initial acidity was increased by decreasing pH (Acidity as pH decreases: pH 2.0, 5.24%; pH 3.0, 3.24%; pH 4.0, 0.35%). For example, when the pH 2.0 treatment group showed titratable acidity of 5.53% and 5.60% on the 0th and 15th days of fermentation, Table 4 showed them as 0.02% and 0.09%, respectively.
Figure 2.
Changes in titratable acidity (a–c) and growth (d–f) under multiple starters according to initial pH and fermentation period. Initial pH: (a) and (d) pH 2.0; (b) and (e) pH 3.0; (c) and (f) pH 4.0. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22. The initial acidity was increased by decreasing pH (Acidity as pH decreases: pH 2.0, 5.24%; pH 3.0, 3.24%; pH 4.0, 0.35%). For example, when the pH 2.0 treatment group showed titratable acidity of 5.53% and 5.60% on the 0th and 15th days of fermentation, Table 4 showed them as 0.02% and 0.09%, respectively.
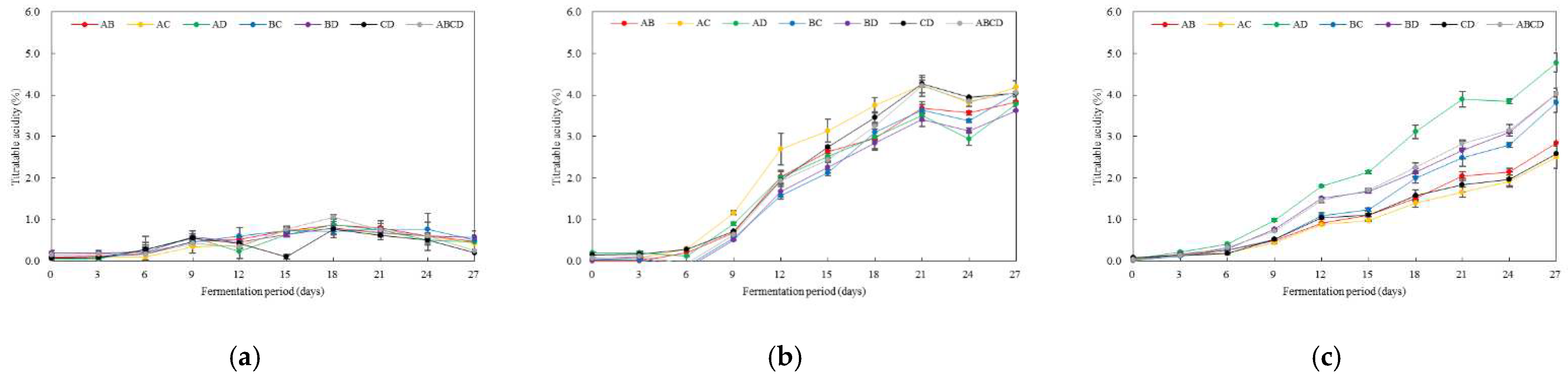
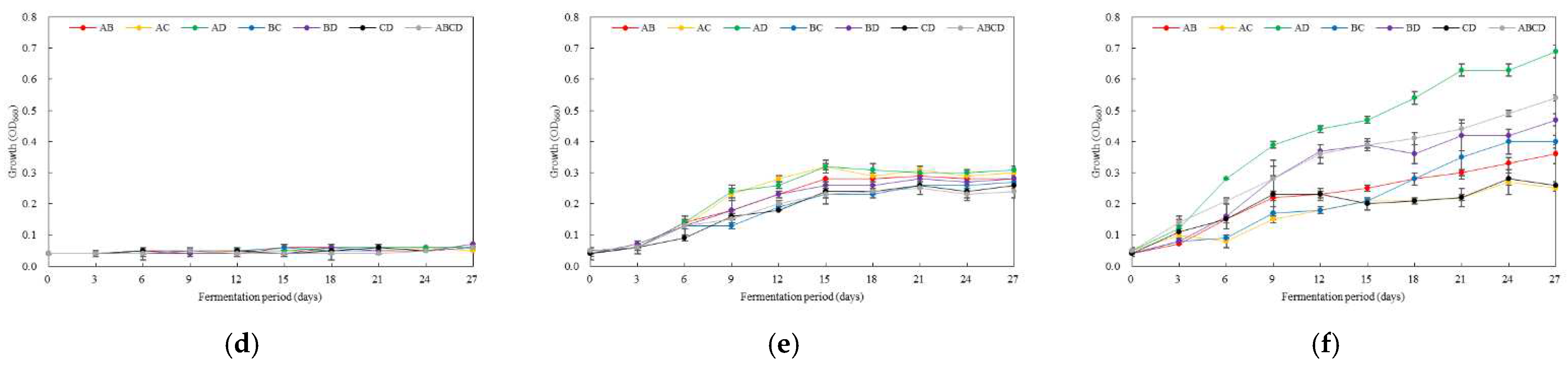
Figure 3.
Changes in titratable acidity (solid line) and growth (dotted line) under multiple starters according to the AAB inoculation ratio and fermentation period. Inoculation ratios: 1:1, 1:2, and 2:1. Multiple starters: (a) AB; (b) AC; (c) AD; (d) BC; (e) BD; (f) CD. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
Figure 3.
Changes in titratable acidity (solid line) and growth (dotted line) under multiple starters according to the AAB inoculation ratio and fermentation period. Inoculation ratios: 1:1, 1:2, and 2:1. Multiple starters: (a) AB; (b) AC; (c) AD; (d) BC; (e) BD; (f) CD. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
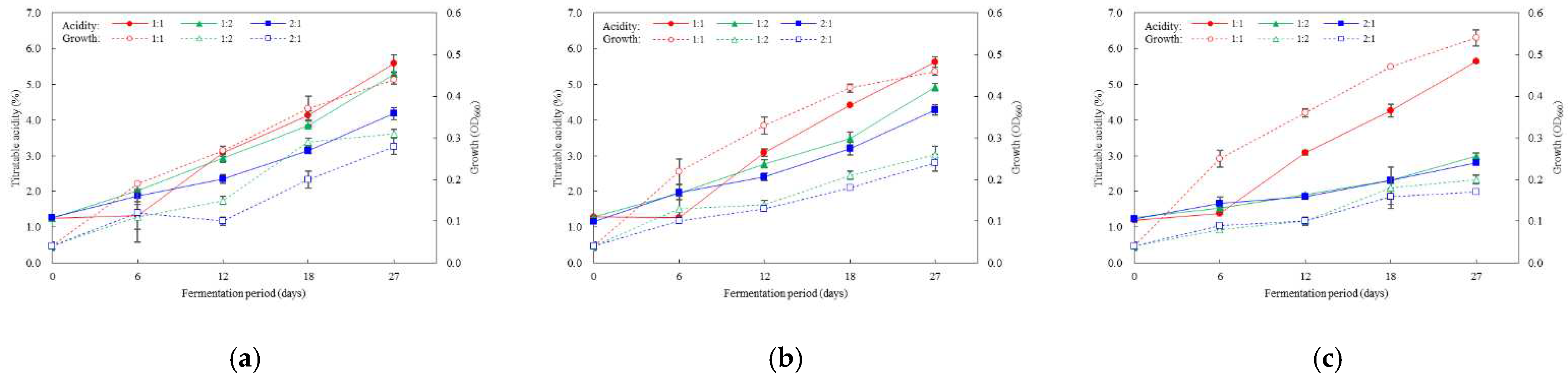
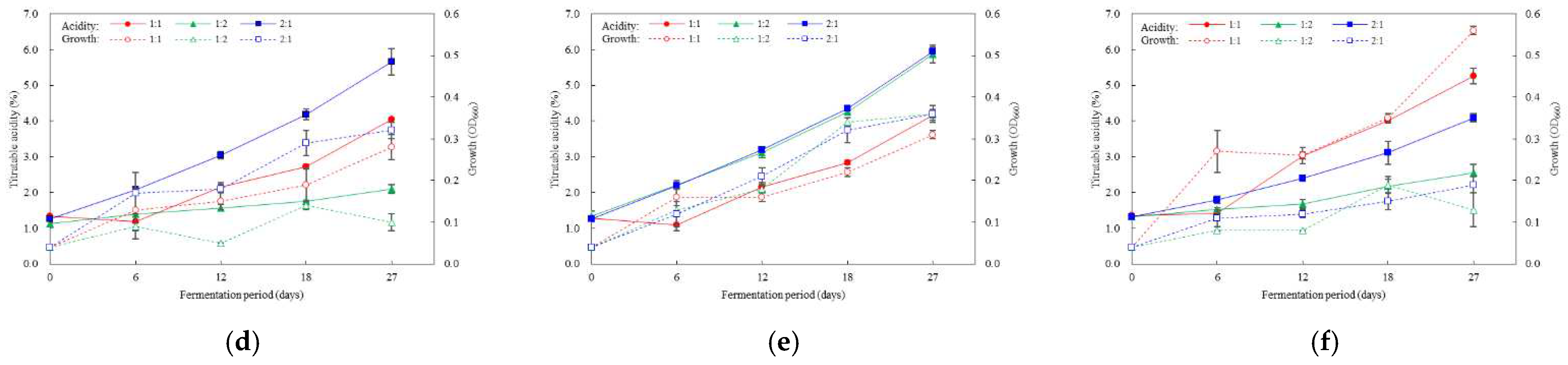
Figure 4.
Changes in titratable acidity (solid line) and growth (dotted line) under multiple starters according to the AAB inoculation order and fermentation period. Inoculation order: 0, simultaneous inoculation; 1st, former inoculation; 2nd, following inoculation. Multiple starters: (a) AB; (b) AC; (c) AD; (d) BC; (e) BD; (f) CD. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
Figure 4.
Changes in titratable acidity (solid line) and growth (dotted line) under multiple starters according to the AAB inoculation order and fermentation period. Inoculation order: 0, simultaneous inoculation; 1st, former inoculation; 2nd, following inoculation. Multiple starters: (a) AB; (b) AC; (c) AD; (d) BC; (e) BD; (f) CD. Symbols: A, Acetobacter ascendens GV–8; B, A. ascendens GV–12; C, A. pasteurianus GV–17; D, A. pasteurianus GV–22.
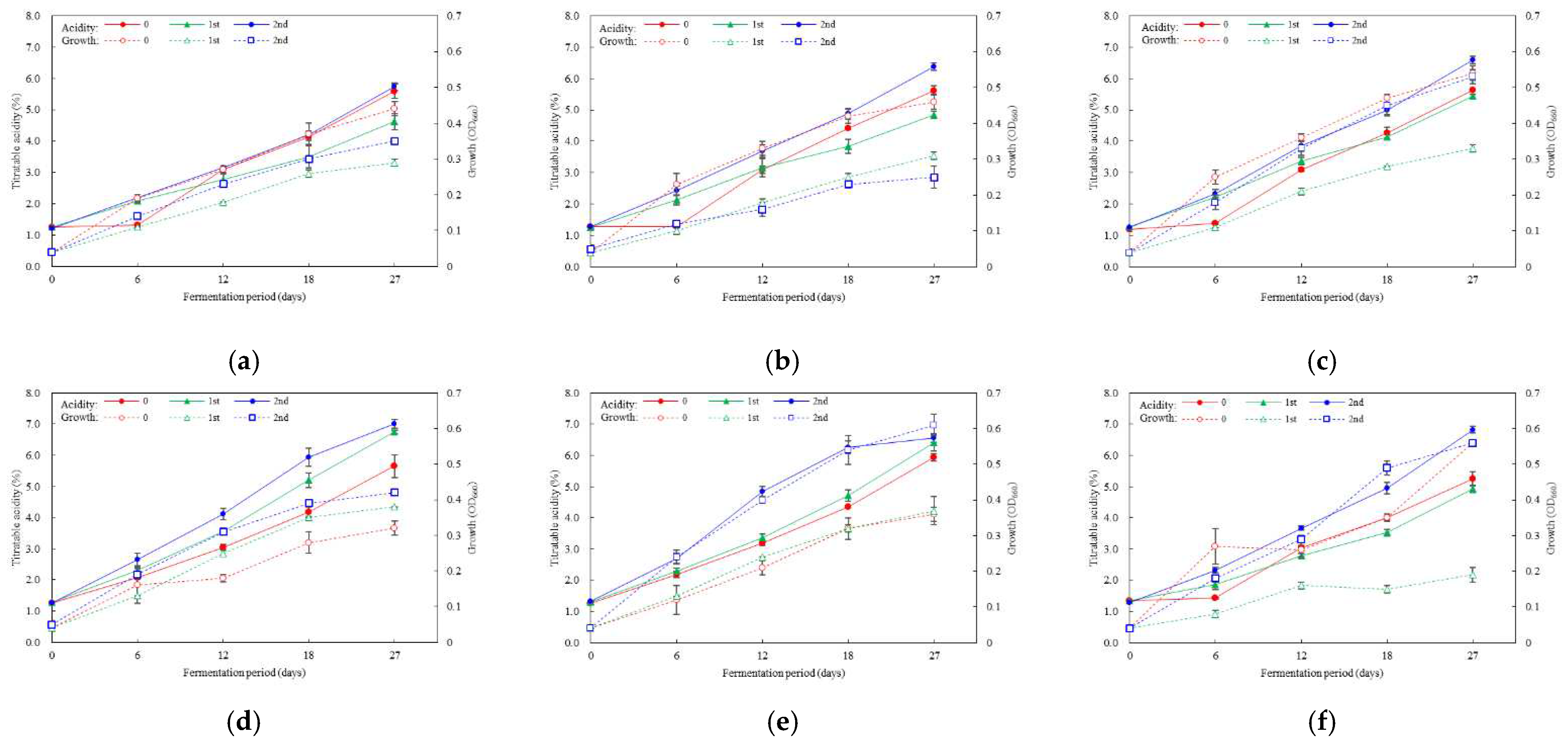

Table 1.
Acetic acid bacteria (AAB) isolated from traditional grain vinegar in various regions in Korea.
Table 1.
Acetic acid bacteria (AAB) isolated from traditional grain vinegar in various regions in Korea.
| Region (Province) | Origin | Sample Name | Species | Strain |
|---|---|---|---|---|
| Gyeongsangnam–do | Brown rice | GN_BWR | Acetobacter pasteurianus | GV–5 |
| Black rice | GN_BR | Acetobacter ascendens | GV–8 (A)1 | |
| Gyeongsangbuk–do | Five grains2 | GB_FG | Acetobacter ascendens | GV–12 (B) |
| Brown rice | GB_UR | Acetobacter pasteurianus | GV–22 (D) | |
| Chungcheongnam–do | Brown rice | CN_UR | Acetobacter pasteurianus | GV–16 |
| Acetobacter pasteurianus | GV–17 (C) |
1 The strains selected as multiple starter candidate are indicated using letters (A–D). 2 Mixture of brown rice, barley, sorghum, millet, and glutinous millet.
Table 2.
Change of titratable acidity and growth under a single AAB according to fermentation temperature and period.
Table 2.
Change of titratable acidity and growth under a single AAB according to fermentation temperature and period.
| Temp. | Strains | Titratable acidity according to fermentation period (days) | Growth (OD660) according to fermentation period (days) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 6 | 9 | 12 | 15 | 0 | 3 | 6 | 9 | 12 | 15 | |||
| 20 °C | GV–5 | 1.25±0.001,c | 1.43±0.03b | 2.14±0.10de | 1.26±0.06d | 0.75±0.18d | 0.57±0.03d | 0.05±0.00c | 0.06±0.00cd | 0.10±0.03bc | 0.38±0.38a | 0.83±0.40a | 1.44±0.29a | |
| GV–8 | 1.30±0.01b | 1.41±0.03b | 2.01±0.02e | 1.11±0.12d | 0.73±0.20d | 0.48±0.08d | 0.06±0.00b | 0.06±0.00cd | 0.06±0.00c | 0.14±0.02b | 0.47±0.38b | 1.49±0.06a | ||
| GV–12 | 1.40±0.00a | 1.40±0.07b | 2.66±0.13bcd | 1.85±0.12d | 1.08±0.15cd | 0.44±0.02d | 0.06±0.00b | 0.06±0.00cd | 0.22±0.03a | 0.25±0.11ab | 0.79±0.20a | 1.13±0.20b | ||
| GV–16 | 1.38±0.00a | 1.40±0.06b | 2.53±0.36cde | 1.54±0.04d | 1.14±0.08cd | 0.47±0.08d | 0.07±0.01a | 0.06±0.00cd | 0.07±0.01bc | 0.22±0.07ab | 0.48±0.12b | 1.05±0.08b | ||
| GV–17 | 1.38±0.00a | 1.39±0.08b | 2.46±0.14de | 1.74±0.11d | 1.41±0.36c | 0.98±0.48c | 0.06±0.00b | 0.06±0.00cd | 0.08±0.00bc | 0.19±0.05ab | 0.45±0.11b | 1.13±0.24b | ||
| GV–22 | 1.41±0.00a | 1.41±0.12b | 2.69±0.31bcd | 1.74±0.19d | 1.07±0.23cd | 0.51±0.14d | 0.05±0.00c | 0.06±0.00cd | 0.21±0.07a | 0.20±0.08ab | 0.34±0.20bc | 1.39±0.06a | ||
| 30 °C | GV–5 | 1.27±0.03bc | 2.23±0.20a | 3.56±0.51a | 4.57±0.60ab | 5.19±0.29a | 5.64±0.16a | 0.06±0.01b | 0.06±0.02cd | 0.10±0.09bc | 0.11±0.11b | 0.13±0.07c | 0.16±0.07c | |
| GV–8 | 1.28±0.05bc | 2.19±0.14a | 3.41±0.33a | 4.77±0.72a | 5.17±0.07a | 5.33±0.19a | 0.07±0.01a | 0.06±0.02cd | 0.08±0.06bc | 0.10±0.08b | 0.10±0.05c | 0.18±0.06c | ||
| GV–12 | 1.20±0.04d | 2.14±0.14a | 3.12±0.27abc | 4.03±0.21b | 5.28±0.16a | 5.62±0.03a | 0.05±0.00c | 0.10±0.04bc | 0.11±0.07bc | 0.10±0.02b | 0.12±0.07c | 0.12±0.04c | ||
| GV–16 | 1.30±0.03b | 2.34±0.19a | 3.27±0.72ab | 4.35±1.00ab | 5.17±0.46a | 5.56±0.25a | 0.05±0.00c | 0.12±0.01ab | 0.22±0.03a | 0.22±0.03ab | 0.23±0.05bc | 0.23±0.03c | ||
| GV–17 | 1.28±0.02bc | 2.21±0.25a | 3.18±0.60ab | 4.19±0.64ab | 4.81±0.55a | 5.60±0.17a | 0.05±0.00c | 0.09±0.03bcd | 0.15±0.06ab | 0.18±0.08ab | 0.22±0.06bc | 0.22±0.04c | ||
| GV–22 | 1.27±0.03bc | 2.25±0.11a | 3.40±0.18a | 4.85±0.24a | 5.27±0.14a | 5.40±0.01a | 0.05±0.00c | 0.15±0.08a | 0.15±0.08ab | 0.16±0.07b | 0.19±0.06bc | 0.23±0.04c | ||
| 40 °C | GV–5 | 1.19±0.00d | 1.18±0.01c | 1.19±0.02f | 1.16±0.01d | 1.16±0.04cd | 1.15±0.02c | 0.05±0.00c | 0.05±0.00d | 0.05±0.00c | 0.05±0.00b | 0.05±0.00c | 0.05±0.00c | |
| GV–8 | 1.17±0.00d | 1.17±0.01c | 1.19±0.01f | 1.17±0.03d | 1.19±0.07cd | 1.16±0.04c | 0.05±0.00c | 0.05±0.00d | 0.05±0.00c | 0.05±0.00b | 0.05±0.00c | 0.05±0.00c | ||
| GV–12 | 1.25±0.05c | 1.23±0.02bc | 2.42±0.62de | 2.61±0.53c | 3.10±0.54b | 2.09±0.68b | 0.05±0.00c | 0.05±0.00d | 0.12±0.05bc | 0.09±0.03b | 0.09±0.03c | 0.09±0.03c | ||
| GV–16 | 1.30±0.00b | 1.24±0.02bc | 1.12±0.02f | 1.18±0.04d | 1.23±0.02cd | 1.20±0.08c | 0.05±0.00c | 0.05±0.00d | 0.05±0.00c | 0.05±0.00b | 0.05±0.00c | 0.05±0.00c | ||
| GV–17 | 1.26±0.01bc | 1.18±0.03c | 1.19±0.02f | 1.16±0.03d | 1.26±0.02c | 1.09±0.07c | 0.05±0.00c | 0.05±0.00d | 0.05±0.00c | 0.05±0.00b | 0.05±0.00c | 0.05±0.00c | ||
| GV–22 | 1.28±0.00bc | 1.15±0.02c | 1.15±0.02f | 1.14±0.02d | 1.20±0.02cd | 1.13±0.06c | 0.05±0.00c | 0.05±0.00d | 0.05±0.00c | 0.05±0.00b | 0.05±0.00c | 0.05±0.00c | ||
1 Values are presented as means ± standard deviation (number of replicates = 3). Different superscripts in the same column indicate a significant difference at P < 0.05, as determined by Duncan’s multiple range test. Different lowercase letters between columns represent significant differences between the cultivars (P < 0.05).
Table 3.
Changes in titratable acidity and growth under a single starter according to ethanol concentration (%) and fermentation period.
Table 3.
Changes in titratable acidity and growth under a single starter according to ethanol concentration (%) and fermentation period.
| Conc. EtOH | Strains | Titratable acidity according to fermentation period (days) | Growth (OD660) according to fermentation period (days) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 6 | 9 | 12 | 15 | 0 | 3 | 6 | 9 | 12 | 15 | |||
| 5% | GV–5 | 1.27±0.031,f | 2.23±0.20a | 3.56±0.51a | 4.57±0.40ab | 5.19±0.29abc | 5.64±0.16cd | 0.06±0.01ab | 0.06±0.02de | 0.10±0.09bcd | 0.11±0.11bcd | 0.13±0.07cdef | 0.16±0.07bcd | |
| GV–8 | 1.28±0.05ef | 2.19±0.14a | 3.41±0.33ab | 4.77±0.38a | 5.17±0.07abc | 5.33±0.19d | 0.07±0.01a | 0.06±0.02de | 0.08±0.06bcd | 0.10±0.08bcd | 0.10±0.05efg | 0.18±0.06abc | ||
| GV–12 | 1.20±0.04g | 2.14±0.14a | 3.12±0.27ab | 4.03±0.21b | 5.28±0.16ab | 5.62±0.03cd | 0.05±0.00b | 0.10±0.04bc | 0.11±0.07bcd | 0.10±0.02bcd | 0.12±0.07def | 0.12±0.04cde | ||
| GV–16 | 1.30±0.03ef | 2.34±0.19a | 3.27±0.72ab | 4.35±0.42ab | 5.17±0.46abc | 5.56±0.25cd | 0.05±0.00b | 0.12±0.01ab | 0.22±0.03a | 0.22±0.03a | 0.23±0.05a | 0.23±0.03a | ||
| GV–17 | 1.28±0.02ef | 2.21±0.25a | 3.18±0.60ab | 4.19±0.64ab | 4.81±0.55bc | 5.60±0.17cd | 0.05±0.00b | 0.09±0.03bcd | 0.15±0.06b | 0.18±0.08ab | 0.22±0.06ab | 0.22±0.04ab | ||
| GV–22 | 1.27±0.03f | 2.25±0.11a | 3.4±0.18ab | 4.85±0.24a | 5.27±0.14ab | 5.40±0.01d | 0.05±0.00b | 0.15±0.08a | 0.15±0.08b | 0.16±0.07ab | 0.19±0.06abc | 0.23±0.04a | ||
| 8% | GV–5 | 1.28±0.03ef | 1.42±0.14c | 2.05±0.25c | 3.17±0.51c | 4.13±0.30d | 5.51±0.38cd | 0.05±0.01b | 0.05±0.00de | 0.07±0.02cd | 0.07±0.02cd | 0.08±0.01fg | 0.08±0.01ef | |
| GV–8 | 1.34±0.00cdef | 1.44±0.08c | 2.06±0.24c | 3.13±0.36c | 4.09±0.28d | 5.30±0.50d | 0.06±0.01ab | 0.05±0.00de | 0.10±0.04bcd | 0.13±0.04bcd | 0.15±0.04cde | 0.16±0.02bcd | ||
| GV–12 | 1.29±0.05ef | 1.76±0.12b | 2.89±0.29b | 4.16±0.64ab | 5.7±0.60a | 7.27±0.46a | 0.06±0.01ab | 0.07±0.01cde | 0.09±0.02bcd | 0.11±0.07bcd | 0.12±0.01def | 0.11±0.06de | ||
| GV–16 | 1.30±0.03ef | 1.77±0.06b | 2.86±0.21b | 3.99±0.42b | 4.70±0.17c | 6.10±0.34c | 0.06±0.01ab | 0.06±0.02de | 0.13±0.00bc | 0.14±0.02abc | 0.17±0.03abcd | 0.21±0.09ab | ||
| GV–17 | 1.29±0.06ef | 1.88±0.23b | 3.30±0.67ab | 4.64±1.12ab | 5.55±0.92a | 6.95±1.14ab | 0.05±0.00b | 0.07±0.02cde | 0.14±0.07bc | 0.16±0.05ab | 0.17±0.05abcd | 0.20±0.04ab | ||
| GV–22 | 1.30±0.02ef | 1.47±0.12c | 1.96±0.06cd | 3.35±0.23c | 4.69±0.22c | 6.64±0.50b | 0.05±0.00b | 0.05±0.00de | 0.08±0.04bcd | 0.10±0.10bcd | 0.16±0.01bcde | 0.22±0.02ab | ||
| 11% | GV–5 | 1.38±0.00abcd | 1.48±0.01c | 1.48±0.14de | 1.34±0.11d | 1.37±0.09e | 1.37±0.08e | 0.05±0.00b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | |
| GV–8 | 1.40±0.01abc | 1.45±0.10c | 1.42±0.04e | 1.36±0.16d | 1.32±0.03e | 1.31±0.02e | 0.05±0.00b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–12 | 1.41±0.05ab | 1.55±0.08c | 1.36±0.03e | 1.28±0.01d | 1.28±0.07e | 1.28±0.03e | 0.06±0.01ab | 0.04±0.00e | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–16 | 1.35±0.04bcde | 1.46±0.02c | 1.37±0.04e | 1.28±0.03d | 1.21±0.05e | 1.28±0.09e | 0.05±0.01b | 0.04±0.00e | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–17 | 1.40±0.00abc | 1.45±0.05c | 1.40±0.04e | 1.27±0.02d | 1.21±0.03e | 1.28±0.04e | 0.05±0.00b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–22 | 1.42±0.00a | 1.44±0.07c | 1.35±0.07e | 1.25±0.03d | 1.18±0.01e | 1.26±0.04e | 0.05±0.00b | 0.04±0.00e | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| 14% | GV–5 | 1.30±0.05ef | 1.47±0.02c | 1.40±0.08e | 1.28±0.01d | 1.38±0.05e | 1.28±0.03e | 0.05±0.00b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | |
| GV–8 | 1.27±0.02f | 1.42±0.06c | 1.31±0.02e | 1.35±0.16d | 1.28±0.03e | 1.39±0.08e | 0.05±0.00b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–12 | 1.27±0.03f | 1.38±0.05c | 1.35±0.08e | 1.27±0.05d | 1.32±0.06e | 1.20±0.18e | 0.06±0.01ab | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–16 | 1.28±0.00ef | 1.38±0.02c | 1.32±0.01e | 1.28±0.01d | 1.38±0.02e | 1.44±0.11e | 0.05±0.00b | 0.04±0.00e | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–17 | 1.29±0.10ef | 1.40±0.04c | 1.41±0.12e | 1.33±0.03d | 1.34±0.01e | 1.30±0.03e | 0.05±0.00b | 0.04±0.00e | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
| GV–22 | 1.32±0.00def | 1.44±0.08c | 1.30±0.01e | 1.28±0.02d | 1.30±0.03e | 1.32±0.02e | 0.05±0.01b | 0.05±0.00de | 0.04±0.00d | 0.04±0.00d | 0.04±0.00g | 0.04±0.00f | ||
1 Values are presented as means ± standard deviation. Different letters in the same row show significant differences (P < 0.05) in Duncan’s multiple range test.
Table 4.
Changes in titratable acidity and growth under a single starter according to initial pH and fermentation period.
Table 4.
Changes in titratable acidity and growth under a single starter according to initial pH and fermentation period.
| Initial pH1 | Strains | Titratable acidity according to fermentation period (days) | Growth (OD660) according to fermentation period (days) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 6 | 9 | 12 | 15 | 0 | 3 | 6 | 9 | 12 | 15 | |||
| pH 2.0 | GV–5 | 0.02±0.032,bc | 0.03±0.01e | 0.09±0.10g | 0.06±0.04h | 0.13±0.14h | 0.09±0.04h | 0.06±0.00a | 0.06±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.00g | 0.05±0.00e | |
| GV–8 | 0.04±0.03bc | 0.04±0.03e | 0.03±0.05g | 0.16±0.20h | 0.22±0.35h | 0.16±0.20h | 0.06±0.00a | 0.06±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.00g | 0.05±0.01e | ||
| GV–12 | 0.09±0.10abc | 0.06±0.03e | 0.07±0.09g | 0.04±0.08h | 0.07±0.09h | 0.11±0.07h | 0.05±0.01b | 0.06±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.01g | 0.05±0.00e | ||
| GV–16 | 0.12±0.11ab | 0.07±0.03e | 0.06±0.03g | 0.20±0.22h | 0.17±0.08h | 0.40±0.36h | 0.06±0.00a | 0.05±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.00g | 0.05±0.00e | ||
| GV–17 | 0.04±0.02bc | 0.05±0.03e | 0.03±0.03g | 0.05±0.03h | 0.06±0.05h | 0.13±0.02h | 0.05±0.00b | 0.06±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.00g | 0.05±0.00e | ||
| GV–22 | 0.03±0.02bc | 0.07±0.03e | 0.15±0.06g | 0.19±0.10h | 0.25±0.16h | 0.31±0.16h | 0.05±0.00b | 0.07±0.00e | 0.05±0.00g | 0.05±0.00e | 0.05±0.00g | 0.05±0.00e | ||
| pH 3.0 | GV–5 | 0.08±0.05abc | 0.09±0.06e | 0.32±0.06g | 2.06±0.47g | 2.19±0.47g | 2.04±0.50g | 0.05±0.00b | 0.06±0.00e | 0.09±0.00g | 0.34±0.05cd | 0.40±0.06ef | 0.43±0.09d | |
| GV–8 | 0.04±0.05bc | 0.11±0.03e | 4.30±0.16c | 5.13±0.24d | 4.90±0.32d | 4.39±0.15de | 0.05±0.00b | 0.06±0.01e | 0.23±0.05def | 0.29±0.02cde | 0.70±0.35bc | 0.88±0.09ab | ||
| GV–12 | 0.06±0.07abc | 0.08±0.03e | 3.41±0.70d | 4.35±0.77e | 4.12±0.43e | 3.78±0.38ef | 0.06±0.00a | 0.07±0.00e | 0.29±0.02d | 0.28±0.06cde | 0.27±0.17fg | 0.48±0.05d | ||
| GV–16 | 0.15±0.09a | 0.10±0.03e | 2.69±0.66de | 3.79±0.47ef | 3.99±0.03ef | 3.81±0.66ef | 0.06±0.01a | 0.06±0.00e | 0.17±0.03f | 0.16±0.00de | 0.46±0.26cdef | 0.44±0.03d | ||
| GV–17 | 0.05±0.09abc | 0.10±0.03e | 2.04±0.77ef | 3.57±0.75f | 3.55±0.22f | 3.44±0.96f | 0.05±0.00b | 0.06±0.00e | 0.10±0.05g | 0.15±0.01de | 0.15±0.01g | 0.17±0.08e | ||
| GV–22 | 0.08±0.06abc | 0.10±0.06e | 1.49±0.23f | 2.57±0.09g | 2.47±0.23g | 2.07±0.78g | 0.06±0.01a | 0.06±0.01e | 0.19±0.01df | 0.32±0.12cde | 0.41±0.09def | 0.49±0.16d | ||
| pH 4.0 | GV–5 | 0.03±0.02bc | 1.60±0.04c | 6.78±0.07a | 7.05±0.02c | 6.99±0.24b | 6.79±0.07a | 0.05±0.00b | 0.22±0.09c | 0.35±0.04c | 0.87±0.04a | 0.94±0.12a | 1.07±0.02a | |
| GV–8 | 0.01±0.02c | 2.99±0.19a | 6.88±1.44a | 8.04±0.19a | 7.55±0.40a | 5.81±0.31b | 0.06±0.00a | 0.53±0.06a | 0.62±0.03a | 0.91±0.00a | 0.90±0.13ab | 1.03±0.23a | ||
| GV–12 | 0.01±0.01c | 2.19±0.10b | 7.16±0.27a | 7.69±0.45ab | 7.51±0.51a | 7.21±0.54a | 0.05±0.00b | 0.28±0.02b | 0.41±0.10b | 0.49±0.13bc | 0.65±0.18cd | 0.78±0.12bc | ||
| GV–16 | 0.02±0.01bc | 1.12±0.11d | 4.60±0.68c | 5.64±0.18d | 5.62±0.15c | 5.41±0.12bc | 0.06±0.01a | 0.15±0.05d | 0.25±0.05de | 0.35±0.07cd | 0.54±0.05cde | 0.61±0.29cd | ||
| GV–17 | 0.02±0.01bc | 1.06±0.11d | 4.54±0.19c | 5.69±0.29d | 5.56±0.13c | 4.80±0.26cd | 0.05±0.00b | 0.11±0.04de | 0.23±0.03def | 0.31±0.22cde | 0.51±0.05cdef | 0.52±0.08d | ||
| GV–22 | 0.03±0.01bc | 1.19±0.21d | 5.71±0.15b | 7.40±0.10bc | 7.13±0.23ab | 7.04±0.36a | 0.05±0.00b | 0.10±0.03de | 0.24±0.01de | 0.66±0.53ab | 0.70±0.15bc | 0.75±0.13bc | ||
1 The initial acidity was increased by decreasing pH (Acidity as pH decreases: pH 2.0, 5.51%; pH 3.0, 3.24%; pH 4.0, 0.37%). For example, when the pH 2.0 treatment group showed titratable acidity of 5.53% and 5.60% on the 0th and 15th days of fermentation, Table 4 showed them as 0.02% and 0.09%, respectively. 2 Values are presented as means ± standard deviation. Different letters in the same row indicate significant differences (P < 0.05) in Duncan’s multiple range test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



