
Submitted:
23 March 2023
Posted:
27 March 2023
You are already at the latest version
Abstract
Phagosomal escape and intracellular survival, often accompanied by Small Colony Variants (SCVs) formation, are typical features of infections caused by S. aureus. The survival in macro-phages favours S. aureus dissemination and complicates treatment. RAW 264.7 murine macro-phages infected with S. aureus USA300 and treated with erythromycin and 20mM carnosine, alone and in combination, were used as experimental model. SCVs were isolated from all treat-ment conditions, but only those undergoing the pressure of combined erythromycin and carnosine for 48 hours were stable for at least six passages on blood agar. Nucleic acid extraction was car-ried out for S. aureus USA300 wild-type and stable SCVs. Whole Genome Sequencing (WGS) was performed using Illumina DNA Prep and Illumina MiSeq, and quantitative reverse transcription PCR was performed. WGS analysis did not yield mutations pointing to differences between S. au-reus USA300 and stable SCVs, therefore the focus was shifted to evaluating gene expression vari-ations. Genes such as zur, mntR, uhpt, fur, sdrE were shown to be significantly up-regulated in SCVs compared to S. aureus USA300 wild-type, suggesting a global change that allows adapta-tion to intracellular persistence, including protection from inflammatory response and evasion of the immune system.
Keywords:
Staphylococcus aureus
; Small Colony Variants
; Carnosine
; RAW 264.7 murine macrophages
; Host-Pathogen Interaction
1. Introduction
Staphylococcus aureus is widely recognized for its ability to adapt to different environments thanks to an excellent genome plasticity exerted through horizontal gene transfer (HGT) mechanisms [1,2,3].
A large plethora of virulence factors and resistance genes makes S. aureus an opportunistic pathogen that represented and still represents one of the most common causes of human infections [4]. Emerged in the past as a nosocomial pathogen, Methicillin-Resistant S. aureus (MRSA) has quickly spread in the community, following different evolutive paths through the sequential acquisition of mobile genetic elements (MGEs) [5].
A major clinical issue related to S. aureus is represented by persistent infections (e.g., prosthetic joint infections) characterized by an asymptomatic phase with relapses occurring months or even years after the apparent resolution of the infection [6].
The ability of S. aureus to evade the immune response through the activation of the “Immune Evasion Cluster” has extensively been assessed [7,8,9]. Moreover, S. aureus proved capable of surviving phagocytosis by neutrophils and macrophages, performing phagosomal escape, as well as of surviving inside non-phagocytic cells, such as osteoblasts, epithelial and endothelial cells [10,11], with all these mechanisms confirming its persistent behaviour. Intracellular survival may lead to the development of S. aureus reservoirs in humans, contributing to both persistence and dissemination from the infection site to other body areas [7,9,12].
In addition, these kinds of infections have often been associated with a particular S. aureus phenotype called Small Colony Variant (SCV), an alternative bacterial lifestyle generally related to adaptation and persistence in unfavourable environmental conditions [6]. Furthermore, the role of professional phagocytes as “trojan horses” has been largely observed in several infectious diseases, such as those caused by bacteria, fungi, and viruses. Interestingly, they are also involved in the carriage of pathogens across the Blood Brain Barrier (BBB) [13,14,15,16].
Understanding the intracellular survival mechanisms of different S. aureus strains is crucial to developing effective strategies against difficult-to-treat infections. Based on our knowledge, intracellular survival of S. aureus varies depending on the set of virulence factors produced (e.g., PVL toxin, alpha toxin, etc) as well as its genetic background. This finding was already assessed through internalization assays in human MG-63 osteoblast-like cells, which revealed differences in internalization and intracellular persistence of S. aureus based on its sequence type (ST) [17]. The complications related to the treatment of persistent and chronic infections has led to the development of novel therapeutic strategies. These include the use of natural antioxidant compounds to counteract bacterial infections by strengthening the host’s defense mechanisms. In this scenario, the natural dipeptide carnosine may represent a promising choice in addition to the antibiotics currently used [18].
The aim of the study was to evaluate the ability of S. aureus USA300 to survive inside RAW 264.7 murine macrophages, as well as to examine potential genomic and transcriptomic changes involved in the transition from the wild type to the SCV phenotype in the presence of carnosine and erythromycin.
2. Results
2.1. Antimicrobial Susceptibility Test
Carnosine was tested alone in Cation Adjusted Muller Hinton Broth (CA-MHB) and Dulbecco’s Modified Eagle Medium (DMEM), and in a range of concentrations from 0,15 mg/L to 160 mg/L, showing no antimicrobial effects compared to the bacterial growth control.
S. aureus USA300 had an erythromycin MIC value of 32 mg/L in both CA-MH Broth and DMEM, also when tested in combination with carnosine 20 mM, showing no antagonism against the antibiotic.
2.2. Infection Assay and Evaluation of SCV Stability
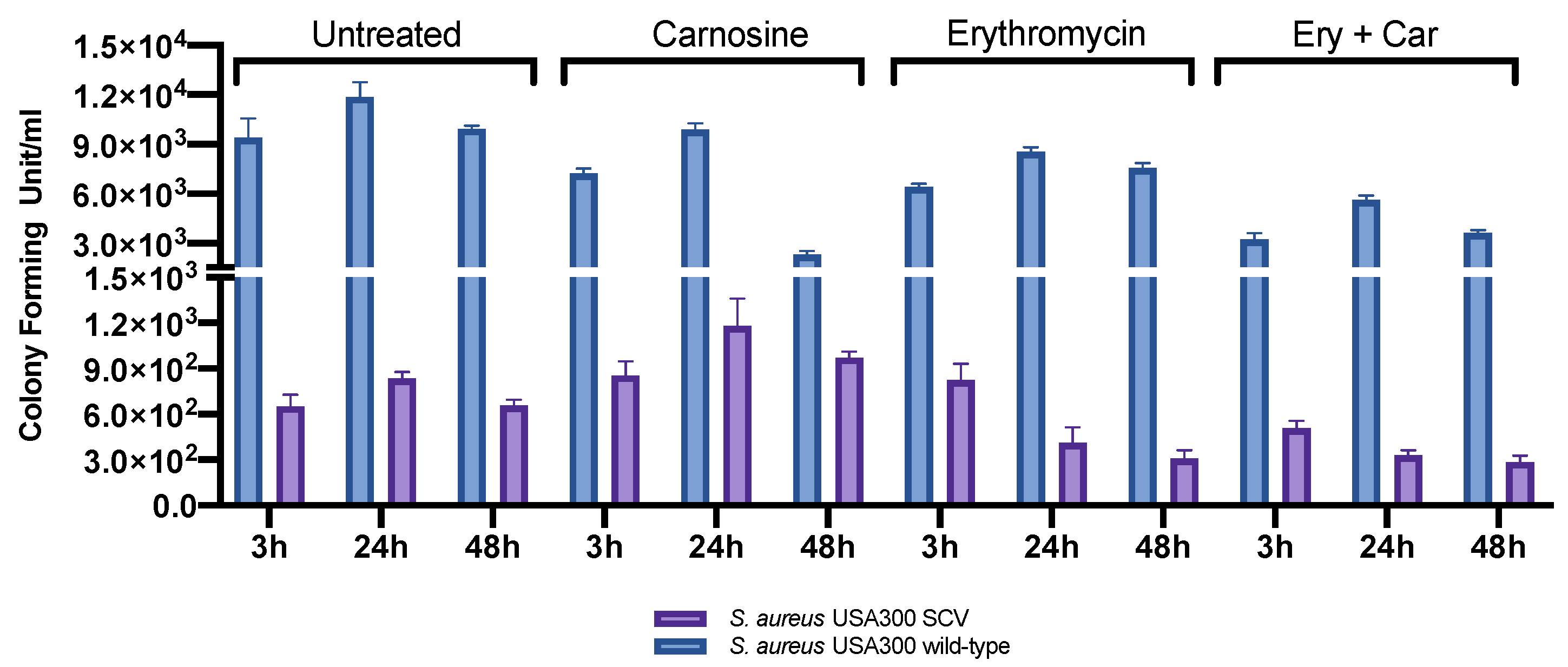
The RAW 264.7 cell line was infected using a Multiplicity of Infection (MOI) of 50:1 (bacterial: eukaryotic cells) for 3, 24 and 48h. The osmotic lysis of infected macrophages was performed at every time point post infection (p.i), lysates were plated on Blood Agar (BA) and bacterial Colony Forming Unit (CFU) were counted after over-night incubation, to assess bacterial recovery compared to the starting inoculum (2,5x106 bacterial cells) as well as to investigate the bacterial phenotype in all experimental conditions (Figure 1).
As reported in Graph 1, a 2-log fold reduction was observed in the untreated control. Carnosine and erythromycin, both alone (20mM carnosine) and in combination (20mM carnosine and 32 mg/L erythromycin), yielded a recovery approximately 3-log lower than the starting inoculum at each time point after infection. As expected, the SCV phenotype was also observed in the untreated control, confirming that S. aureus tends to grow as SCV in order to counteract the cellular stress caused by macrophages. Even though SCVs were isolated in all experimental conditions, several subcultures showed different phenotypic stability based on the treatment. In particular, SCVs induced by either carnosine or erythromycin alone reverted to the wild type form right after the first passage on Blood Agar (BA) (Figure 2a). On the other hand, SCVs isolated 48h p.i. from the combined treatment condition were stable for at least six passages on BA. This observation suggests that the combination of carnosine and erythromycin triggers and supports the mechanisms underlying SCV formation, leading to longer stability compared to other treatment conditions (Figure 2b).
Graph 1. Colony count (CFU/ml) from all experimental conditions at 3, 24 and 48h p.i.
2.3. Genomic Analysis
Illumina Whole Genome Sequencing (WGS), de novo genome assembly, sorting and variants calling were performed. The analysis aimed to test crucial S. aureus genes for one or more point-mutations which may represent a unique genomic signature of S. aureus SCV. Genomic comparison did not reveal any relevant mutations which could differentiate S. aureus USA300 wt and S. aureus SCV. Indeed, all mutations observed are common to both phenotypes and not involved in aminoacidic changes that can affect protein structure and function. The detected mutations are listed in the supplementary materials (Table S1).
2.4. Gene Expression Analysis
Gene expression was evaluated for genes belonging to several categories, so, regardless of their biological functions, they were classified as follows.
Metal Ions Uptake: up-regulation was observed for sirC, zur, mntR, fur, perR mRNA expression levels, with a p value <0.05 for fur, zur and mntR (S. aureus USA300 SCV vs S. aureus USA300 wt).
Ros neutralization: slight, statistically non-significant down-regulation and slight, statistically non-significant up-regulation were observed for katA and for sodM and sodA mRNA expression, respectively (S. aureus USA300 SCV Vs S. aureus USA300 wt).
Metabolic Pathways: slight, statistically non-significant down-regulation and statistically non-significant up-regulation were observed for pdhA and for fumC mRNA expression, respectively; on the other hand, statistically significant up-regulation was observed for uhpt, with a p value <0.05 (S. aureus USA300 SCV Vs S. aureus USA300 wt).
Regulators: slight, statistically non-significant up-regulation and remarkable, statistically non-significant up regulation were observed for sarA and sigB and for agrA mRNA expression, respectively.
Virulence: slight, statistically not significant down-regulation and remarkable, statistically non-significant up regulation were observed for psmA and for hla and hld mRNA expression, respectively.
3. Discussion
S. aureus is a notorious opportunistic pathogen that causes a plethora of diseases. As previously demonstrated, several S. aureus strains exploit their ability to penetrate non-phagocytic cells inducing intracellular toxicity and tissue destruction; on the other hand, bacterial persistence within cells is thought to lead to immune evasion and chronicity of infections. Furthermore, phagocytosis by neutrophils and macrophages allows dissemination of the pathogen. S. aureus host-pathogen interactions, with a focus on intracellular survival, is a matter deserving of serious consideration [9]. Phagosomal escape is another mechanism of persistence in the human body that affects infection dynamics [18].
Synthetized via the ATP-dependent enzyme carnosine synthetase 1 (CARNS1), carnosine is a dipeptide composed of β-alanine and L-histidine and found in several human tissues. The highest concentrations are observed in the skeletal and cardiac muscles (up to 20 mM) as well as in the brain (0.7–2.0 mM) [19,20,21,22]. Carnosine is extensively known for being involved in macrophage activation as well as a ROS and RNS scavenger, enhancing the expression of antioxidant enzymes (Gpx1, SOD-2, Cat) [23].
Previous studies demonstrated a strong increase in IL-6 and TNF-α, two cytokines induced by S. aureus ST239 when infecting non-immune cells such as osteoblasts; the pro-inflammatory cytokines produced during S. aureus infections promote its outgrowth and bacterial intracellular persistence [24].
In the present study, aimed at evaluating the effect of carnosine on macrophages and S. aureus persistence, the RAW 264.7 murine macrophages cell line was infected with the S. aureus USA300 strain and treated both with erythromycin alone and in combination with carnosine 20mM. Experimental conditions were evaluated at 3, 24 and 48 hours post infection. RAW 264.7 murine macrophages infected with S. aureus USA300 and not treated with antibiotics were used as controls, also demonstrating the great ability of this strain to survive in the cells. Small colony variants (SCVs) isolated from all analysed conditions demonstrate S. aureus USA300 adaptation.
Once SCVs were obtained from all experimental conditions, several passages on blood agar were performed in order to test their phenotypical stability. Despite a slight numeric decrement of the SCVs isolated 48 hours p.i. from the combined treatment with erythromycin and carnosine, a stable phenotype was observed for at least six passages on blood agar. These findings support the hypothesis that the use of carnosine as an adjuvant for erythromycin may support the macrophage antioxidant machinery triggering the host defence. On the other hand, this leads S. aureus to acquire a stable SCV phenotype.
As reported in the literature [25,26,27], stable SCV phenotype is usually related to mutations in specific genes related to electron transport chain, such as hemB, menD, ctaA and thyA. Therefore, Whole Genome Sequencing (WGS) was performed in order to compare the wild-type isolate with its SCV. The analysis did not reveal any relevant mutation. Since chromosomal rearrangement might be involved in stable SCV formation [28], the sequencing method employed may represent a limitation. Long-read sequencing is currently ongoing; this will provide greater genomic insights.
Gene expression analysis was performed with the aim of elucidating the differences between the wild-type and SCV phenotypes of S. aureus USA 300, focusing our attention on target genes involved in regulation, adhesion and toxin production, and ROS neutralization. Since increased glycolytic metabolism was reported for stable SCV [29], genes related to glycolysis were also investigated.
Figure 4 reports differentially expressed genes between S. aureus USA300 and SCVs.
The expression of regulatory and virulence genes is reported in Figure 3a,b and Table 1, and summarized in Figure 4.
The over-expression of the agrA gene in SCVs may be explained by the over-expression of the sarA gene; this expression profile is associated with intracellular survival in macrophages [30]. SarA over-expression leads to the over-expression of sdrE and virulence genes (hla and hld genes), thus favoring the mechanism of adhesion (a typical strategy in the initial stages of the internalization process, when S. aureus shows a slowdown in metabolic activity) and persistence in macrophages [31].
The expression of the Ros neutralization and Metal Ione uptake genes is reported in Figure 3d,e and Table 1, and summarized in Figure 5.
Over-expression of the genes encoding for zinc, iron and manganese transporters (zur, sirC and mntR) was reported, highlighting the importance of metal ions as cofactors of several enzymes involved in ROS neutralization, such as catalase and superoxide dismutase [32,33,34] and their centrality for macrophage survival [7,30].
These regulators act on PerR, which affects the oxidative stress acting on catalase and superoxide dismutase encoded by katA, sodM, sodA [35,36]. The latter was found to be over-expressed. These genes were involved in the detoxification of vacuolar reactive oxygen species, contributing to intracellular survival, which in this contest is not yet required [7].
As previously demonstrated, the glycolytic and fermentative pathways are up-regulated in S. aureus SCVs, even in the presence of oxygen, thus exhibiting a type of anaerobic metabolism [37]. Indeed, statistically significant over-expression of the uhpt gene (a hexose phosphate transporter) was observed. Moreover, statistically non-significant over-expression was shown for the fumC gene. This gene encodes for fumarate dehydrogenase, responsible for the conversion of fumarate into malate. Fumarate is known to be essential for the epigenetic changes associated with trained immunity, therefore its depletion, due to fumC up-regulation, was shown to be crucial for the intracellular survival of SCVs [29].
Since chronic and persistent infections are a major clinical burden due to failure of prolonged antibiotic treatment, our findings allow to improve the knowledge of antibiotic resistance mechanisms, sometimes related to the most difficult-to-treat SCV phenotype.
In order to clarify the mechanisms involved in host-pathogen interaction, this experimental model should be improved to include the impairment of the host’s inflammation pathways that lead to the failure of macrophage immune activity.
4. Materials and Methods
4.1. Sample Description
S. aureus USA300 subspecies aureus Rosenbach (BAA-1556TM) was chosen for the internalization study.
Molecular analysis revealed that S. aureus USA300 is a Panton-Valentine leucocidin (PVL) producer, and harbors Staphylococcal Cassette Chromosome mec (SCCmec) type IV, spa type t008 and MLST type ST8, ACME locus [38].
4.2. Broth Microdilution
To test erythromycin and carnosine activity, both alone and in combination, minimum inhibitory concentrations (MICs) were assessed by broth microdilution (BMD) as recommended by the CLSI guidelines [39]. MIC values were evaluated based on the EUCAST v 12.0 breakpoints [40].
BMDs using a fixed concentration of carnosine (Cat. No C9625, Sigma-Aldrich-Merck,) (20 mM) combined with increasing erythromycin (Cat. No E5389-1G, Sigma-Aldrich-Merck) concentrations (from 0.25 to 256 mg/L) were performed both in the CA-MHB (Cat. No 11703503, BD Difco™ Mueller Hinton Broth) and DMEM (Cat. No 11965092, Gibco™ DMEM, high glucose) media used to grow RAW 264.7 (ATCC® TIB-71™) eukaryotic cells. All plates were incubated overnight at 37°C.
4.3. Infection Assay
4.3.1. Eukaryotic Cell Culture Preparation
RAW 264.7 cells were selected to perform the infection assay. Cells were grown in 75 cm2 flasks with DMEM, HEPES, GlutaMAX™ Supplement DMEM (Cat. No 11965092, Gibco™ DMEM, high glucose DRAFT) supplemented with Fetal Bovine Serum (FBS) 10% (Cat. No F7524, Sigma-Aldrich-Merck) and 100 U/ml of Penicillin/Streptomycin (Cat. No 15140148, Gibco™, ThermoFisher Scientific). The cell culture was incubated in humid atmosphere at 37° C and CO2 5%. The medium was changed twice weekly.
5x104 RAW 264.7 cells were plated in a 24-well plate, 24 hours before infection, in DMEM without Pen/Strep, supplemented with FBS 10% and treated with 20mM of carnosine.
4.3.2. Infection of RAW 264.7 cells
RAW 264.7 cells were infected with S. aureus USA 300 with a Multiplicity of Infection (MOI) of 50:1 (2,5x106 bacterial cells). Treatment with lysostaphin (100 mg/ml) (Cat. No SAE0091, Sigma-Aldrich-Merck) was performed at 37°C for 1 hour to remove all bacterial cells in the extracellular environment. To confirm the absence of extracellular bacteria, 100µl of each well were plated in Tryptic Soy Agar (TSA) (Cat. No 11973752, Oxoid™, Thermo Scientific™) plates and incubated overnight at 35/37°C.
Different treatments were performed: untreated, carnosine alone (20 mM), erythromycin alone (32 mg/L), carnosine and erythromycin in combination. Osmotic lysis, 3h, 24h and 48h post-infection, was performed by incubating cells in sterile water at 37°C for 30 minutes. Cellular lysates, containing internalized bacteria, were suitably diluted, plated on Blood Agar (BA) plates and incubated overnight at 35/37°C to evaluate bacterial recovery in all different experimental conditions.
4.3.3. Evaluation of SCV Stability
Since coexistence of the wild type and SCV phenotypes was reported in all of the experimental conditions, SCVs were isolated and sub-cultured six times on BA plates to evaluate their phenotypic stability. SCV stability was also tested after freezing in TSB with glycerol 15% (Cat. No 27002, LIOFILCHEM® S.r.l., Teramo, Italy) and subsequently cultured in BA. In this work, SCVs were defined as stable if no transition to the wild type phenotype took place after at least six passages on BA.
4.4. Genomic Analysis
4.4.1. DNA Extraction
S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin were selected for DNA isolation.
DNA extraction was carried out following the manufacturer’s instructions provided by the QIAGEN QIAamp® DNA Mini Kit (Cat. No 51304, QIAGEN, Hilden, Germany). DNA was quantified using both the Eppendorf BioPhotometer® D30 and the fluorimeter Qubit dsDNA BR Assay Kit to evaluate purity and quantity of the initial sample, respectively (Cat. No 32850, Invitrogen, 92008 Carlsbad, CA, USA).
4.4.2. Whole Genome Sequencing
A starting amount of 10 ng from the two selected samples was used for Next Generation Sequencing (NGS). This was performed on the Illumina MiSeq platform according to the manufacturer’s instructions provided by the QIAseq® FX DNA Library Core Kit (Cat. No 1120146, QIAGEN, Hilden, Germany). Bead-based library purifications were performed using Agencourt® AMPure® XP Beads (Cat. No A63880, Beckman Coulter®, Indianapolis, IN, USA).
Libraries were quantified and their quality evaluated using both the fluorometric Qubit dsDNA HS Assay Kit (Cat. No Q32851, Invitrogen, Carlsbad, CA 92008, USA) and the Agilent® High Sensitivity DNA Kit (Cat. No. 5067-4626, Agilent Technologies, California, USA).
Denature and dilute libraries were performed following the “Denature and Dilute Libraries Guide” protocol provided by Illumina®, choosing 8,5 pM as the loading concentration. [41] Finally, sequencing was performed using the MiSeq Reagent Kits v3 (Ref. 15043895, Illumina, Inc., 92122, San Diego, CA, USA).
4.4.3. Data Analysis
Data were analyzed using the QIAGEN CLC Genomics Workbench software and following the User Manual for the CLC Microbial Genomics Module v22.0, released on January 4, 2022 (QIAGEN, Aarhus, 8000, Denmark), that uses the CARD database to assign resistance, virulence and MLST genes (https://card.mcmaster.ca/).
4.4.4. Bioinformatic Analysis
Two paired-end bacterial raw reads were firstly trimmed with TrimGalore (v0.5.0) [42,43] to remove the adapter sequence. Thereafter, the bacterial genome was assembled de novo using Unicycler (v0.4.8) [44] with the Illumina-only assembly. Unicycler output assemblies were then aligned with several protein sequences in order to identify punctual mutations. Firstly, the alignment was carried out using bwa mem [45]. After that, files were sorted by SAMtools, variants were called and consensus sequences were generated using BCFtools [46,47]. The research of aminoacidic mutations was done using transeq (EMBOSS:v.6.6.0.0) and blastp (2.12.0+) [48].
4.4.5. Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: http://www.ncbi.nlm.nih.gov/bioproject/ PRJNA912391.
4.5. Expression Study
4.5.1. RNA Extraction
S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin at 48 p.i were selected for RNA isolation.
RNA extraction was performed using the RNeasy Mini Kit (Cat. No 74104, Qiagen, Hilden, Germany) following the manufacturer's instructions, with some modifications as previously described [17] RNA quality was tested with a Qubit® 3.0 Fluorometer (Cat. No Q33216, Life Technologies, Thermo Fisher Scientific, Waltham, Massachusetts, USA) using the Qubit RNA HS Assay Kit (250 pg/µl and 100 ng/µl).
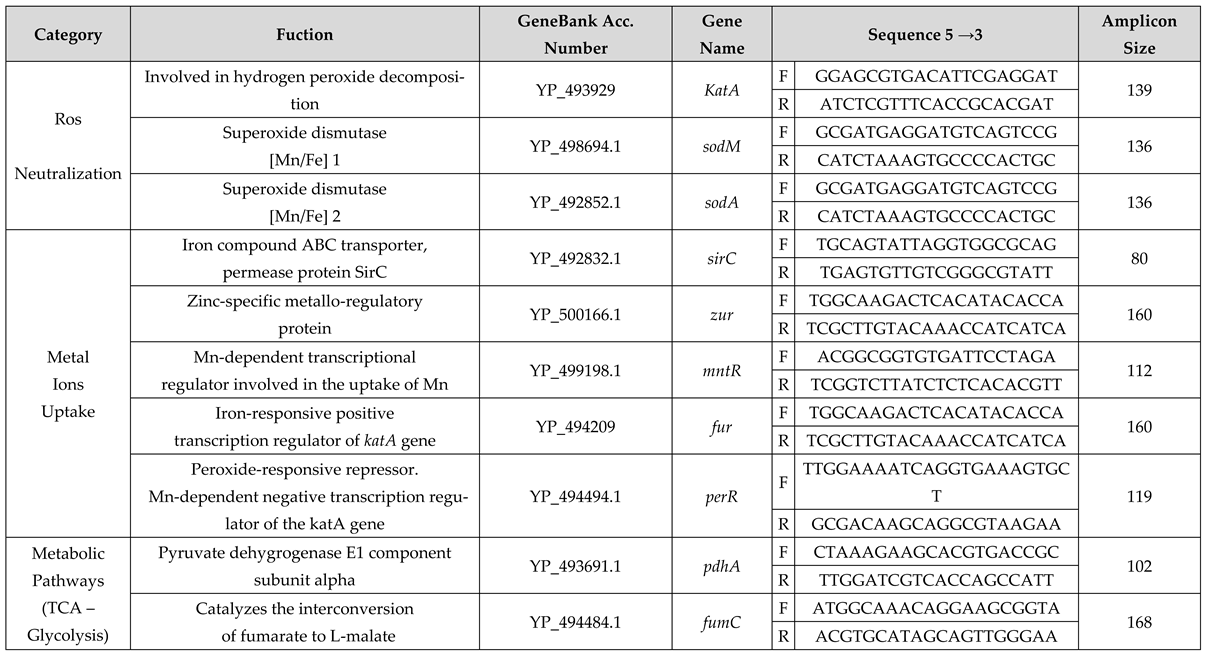
4.5.2. Primer Design
The primers used, as reported in Table 2, were designed with the Primer designing tool – NCBI (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) using S. aureus USA300_FPR3757; NC_007793 as the reference genome.
Furthermore, the agrA, sdrE, psmA, hla, hld, uhpt and SigB genes were amplified using the primers previously reported [17].
4.5.3. RT-qPCR
RNA was normalized at 50 ng/µl using the QuantiTect Reverse Transcription Kit (Cat. No 205311, Qiagen, Hilden, Germany) to obtain cDNA, and amplifications were performed using the QuantiNova SYBR Green PCR Kit (Cat. No 208052, Qiagen, Hilden, Germany) at a final cDNA concentration of 8 ng/µl. The negative control consisted in a reaction in the absence of cDNA, indicated as NTC (No Template Control). The annealing temperature was 60°C for all customized primers. qPCRs were performed in a LightCycler® 480 Real-Time PCR System (Roche, Monza, Italy). PCR efficiencies, melting curve analysis, and expression rate were calculated using the LightCycler® 480 software (Roche, Monza, Italy).
4.5.4. Statistical Analysis
Statistical analysis was performed and the related graphs were prepared using GraphPad Prism Software, Version 9.4.0, June 3, 2022 (GraphPad Software, San Diego, CA, USA). The relative RNA expression level for each sample was calculated using the 2−ΔΔCT method (threshold cycle (CT) value of the gene of interest vs. CT value of the housekeeping gene). [49]
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: mutations annotated by comparing S. aureus USA300 and its SCVs genomes; Table S2: statistical insights of gene expression levels detected through qRT-PCR.
Author Contributions
Conceptualization, investigation, data curation, formal analysis, writing—original draft preparation D.A.B. and C.B.; formal analysis, investigation and data curation A.M., P.G.B.; software and data analysis G.P. and N.M.; data curation, validation, visualization N.M. and G.C; resources, supervision, funding acquisition, writing—review F.C., D.B., S.S.; methodology, investigation, supervision and writing-review and editing N.M. and D.B. All authors have read and agreed to the published version of the manuscript.
Funding
The manuscript was partially supported by: a research project grant, PRIN2020 CODICO PROGETTO 202089LLEH_002 from the Ministry of Research (MIUR)Italy, and by the research project Grant of European Union -FSE-REACT-EU, PON Ricerca e Innovazione 2014-2020 DM1062/2021 and the MUR PNRR Extended Partnership initiative on Emerging Infectious Diseases (Project no. PE00000007, INF-ACT), and by Italian Ministry of Health Research Program (grant number RC2022-N4).
Data Availability Statement
not available.
Acknowledgments
We would like to thank the BRIT laboratory at the University of Catania (Italy) for the valuable technical assistance and the use of its facility. The authors wish to thank PharmaTranslated (http://www.pharmatranslated.com/, accessed data 10 May 2022) and, in particular, Silvia Montanari. Francesco Iannelli and Francesco Santoro for bioinformatic support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Lencastre, H.; Oliveira, D.; Tomasz, A. Antibiotic Resistant Staphylococcus Aureus: A Paradigm of Adaptive Power. Curr Opin Microbiol 2007, 10, 428–435. [CrossRef]
- Boucher, H.W.; Corey, G.R. Epidemiology of Methicillin-Resistant Staphylococcus Aureus. Clin Infect Dis 2008, 46 Suppl 5, S344-349. [CrossRef]
- Filippova, E.V.; Zemaitaitis, B.; Aung, T.; Wolfe, A.J.; Anderson, W.F. Structural Basis for DNA Recognition by the Two-Component Response Regulator RcsB. mBio 2018, 9, e01993-17. [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin Microbiol Rev 2015, 28, 603–661. [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus Aureus: Molecular Characterization, Evolution, and Epidemiology. Clin Microbiol Rev 2018, 31, e00020-18. [CrossRef]
- Tuchscherr, L.; Medina, E.; Hussain, M.; Völker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Roth, J.; Proctor, R.A.; Becker, K.; et al. Staphylococcus Aureus Phenotype Switching: An Effective Bacterial Strategy to Escape Host Immune Response and Establish a Chronic Infection. EMBO Mol Med 2011, 3, 129–141. [CrossRef]
- Flannagan, R.S.; Heit, B.; Heinrichs, D.E. Intracellular Replication of Staphylococcus Aureus in Mature Phagolysosomes in Macrophages Precedes Host Cell Death, and Bacterial Escape and Dissemination. Cell Microbiol 2016, 18, 514–535. [CrossRef]
- Fraunholz, M.; Sinha, B. Intracellular Staphylococcus Aureus: Live-in and Let Die. Front Cell Infect Microbiol 2012, 2, 43. [CrossRef]
- Horn, J.; Stelzner, K.; Rudel, T.; Fraunholz, M. Inside Job: Staphylococcus Aureus Host-Pathogen Interactions. Int J Med Microbiol 2018, 308, 607–624. [CrossRef]
- Bongiorno, D.; Musso, N.; Lazzaro, L.M.; Mongelli, G.; Stefani, S.; Campanile, F. Detection of Methicillin-Resistant Staphylococcus Aureus Persistence in Osteoblasts Using Imaging Flow Cytometry. Microbiologyopen 2020, 9, e1017. [CrossRef]
- Stracquadanio, S.; Musso, N.; Costantino, A.; Lazzaro, L.M.; Stefani, S.; Bongiorno, D. Staphylococcus Aureus Internalization in Osteoblast Cells: Mechanisms, Interactions and Biochemical Processes. What Did We Learn from Experimental Models? Pathogens 2021, 10, 239. [CrossRef]
- Kubica, M.; Guzik, K.; Koziel, J.; Zarebski, M.; Richter, W.; Gajkowska, B.; Golda, A.; Maciag-Gudowska, A.; Brix, K.; Shaw, L.; et al. A Potential New Pathway for Staphylococcus Aureus Dissemination: The Silent Survival of S. Aureus Phagocytosed by Human Monocyte-Derived Macrophages. PLoS One 2008, 3, e1409. [CrossRef]
- Santiago-Tirado, F.H.; Onken, M.D.; Cooper, J.A.; Klein, R.S.; Doering, T.L. Trojan Horse Transit Contributes to Blood-Brain Barrier Crossing of a Eukaryotic Pathogen. mBio 2017, 8, e02183-16. [CrossRef]
- Santiago-Tirado, F.H.; Doering, T.L. False Friends: Phagocytes as Trojan Horses in Microbial Brain Infections. PLoS Pathog 2017, 13, e1006680. [CrossRef]
- Cafruny, W.A.; Bradley, S.E. Trojan Horse Macrophages: Studies with the Murine Lactate Dehydrogenase-Elevating Virus and Implications for Sexually Transmitted Virus Infection. J Gen Virol 1996, 77 ( Pt 12), 3005–3012. [CrossRef]
- Percivalle, E.; Sammartino, J.C.; Cassaniti, I.; Arbustini, E.; Urtis, M.; Smirnova, A.; Concardi, M.; Belgiovine, C.; Ferrari, A.; Lilleri, D.; et al. Macrophages and Monocytes: “Trojan Horses” in COVID-19. Viruses 2021, 13, 2178. [CrossRef]
- Bongiorno, D.; Musso, N.; Caruso, G.; Lazzaro, L.M.; Caraci, F.; Stefani, S.; Campanile, F. Staphylococcus Aureus ST228 and ST239 as Models for Expression Studies of Diverse Markers during Osteoblast Infection and Persistence. Microbiologyopen 2021, 10, e1178. [CrossRef]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus Aureus Infection. Frontiers in Immunology 2021, 11. [CrossRef]
- Hipkiss, A.R.; Preston, J.E.; Himsworth, D.T.; Worthington, V.C.; Keown, M.; Michaelis, J.; Lawrence, J.; Mateen, A.; Allende, L.; Eagles, P.A.; et al. Pluripotent Protective Effects of Carnosine, a Naturally Occurring Dipeptide. Ann N Y Acad Sci 1998, 854, 37–53. [CrossRef]
- Gariballa, S.E.; Sinclair, A.J. Carnosine: Physiological Properties and Therapeutic Potential. Age Ageing 2000, 29, 207–210. [CrossRef]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of Beta-Alanine Supplementation on Skeletal Muscle Carnosine Concentrations and High Intensity Cycling Capacity. Amino Acids 2007, 32, 225–233. [CrossRef]
- Kalyankar, G.D.; Meister, A. Enzymatic Synthesis of Carnosine and Related Beta-Alanyl and Gamma-Aminobutyryl Peptides. J Biol Chem 1959, 234, 3210–3218. [CrossRef]
- Aldini, G.; Piccoli, A.; Beretta, G.; Morazzoni, P.; Riva, A.; Marinello, C.; Maffei Facino, R. Antioxidant Activity of Polyphenols from Solid Olive Residues of c.v. Coratina. Fitoterapia 2006, 77, 121–128. [CrossRef]
- Rokicki, J.; Li, L.; Imabayashi, E.; Kaneko, J.; Hisatsune, T.; Matsuda, H. Daily Carnosine and Anserine Supplementation Alters Verbal Episodic Memory and Resting State Network Connectivity in Healthy Elderly Adults. Front Aging Neurosci 2015, 7, 219. [CrossRef]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small Colony Variants: A Pathogenic Form of Bacteria That Facilitates Persistent and Recurrent Infections. Nat Rev Microbiol 2006, 4, 295–305. [CrossRef]
- Dean, M.A.; Olsen, R.J.; Long, S.W.; Rosato, A.E.; Musser, J.M. Identification of Point Mutations in Clinical Staphylococcus Aureus Strains That Produce Small-Colony Variants Auxotrophic for Menadione. Infect Immun 2014, 82, 1600–1605. [CrossRef]
- Besier, S.; Ludwig, A.; Ohlsen, K.; Brade, V.; Wichelhaus, T.A. Molecular Analysis of the Thymidine-Auxotrophic Small Colony Variant Phenotype of Staphylococcus Aureus. Int J Med Microbiol 2007, 297, 217–225. [CrossRef]
- Guérillot, R.; Kostoulias, X.; Donovan, L.; Li, L.; Carter, G.P.; Hachani, A.; Vandelannoote, K.; Giulieri, S.; Monk, I.R.; Kunimoto, M.; et al. Unstable Chromosome Rearrangements in Staphylococcus Aureus Cause Phenotype Switching Associated with Persistent Infections. Proceedings of the National Academy of Sciences 2019, 116, 20135–20140. [CrossRef]
- Wong Fok Lung, T.; Monk, I.R.; Acker, K.P.; Mu, A.; Wang, N.; Riquelme, S.A.; Pires, S.; Noguera, L.P.; Dach, F.; Gabryszewski, S.J.; et al. Staphylococcus Aureus Small Colony Variants Impair Host Immunity by Activating Host Cell Glycolysis and Inducing Necroptosis. Nat Microbiol 2020, 5, 141–153. [CrossRef]
- Moldovan, A.; Fraunholz, M.J. In or out: Phagosomal Escape of Staphylococcus Aureus. Cell Microbiol 2019, 21, e12997. [CrossRef]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pförtner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus Aureus Adaptation during Chronic Infections. PLoS Pathog 2015, 11, e1004870. [CrossRef]
- Horsburgh, M.J.; Clements, M.O.; Crossley, H.; Ingham, E.; Foster, S.J. PerR Controls Oxidative Stress Resistance and Iron Storage Proteins and Is Required for Virulence in Staphylococcus Aureus. Infect Immun 2001, 69, 3744–3754. [CrossRef]
- Horsburgh, M.J.; Ingham, E.; Foster, S.J. In Staphylococcus Aureus, Fur Is an Interactive Regulator with PerR, Contributes to Virulence, and Is Necessary for Oxidative Stress Resistance through Positive Regulation of Catalase and Iron Homeostasis. J Bacteriol 2001, 183, 468–475. [CrossRef]
- Xiong, A.; Singh, V.K.; Cabrera, G.; Jayaswal, R.K. Molecular Characterization of the Ferric-Uptake Regulator, Fur, from Staphylococcus Aureus. Microbiology (Reading) 2000, 146 ( Pt 3), 659–668. [CrossRef]
- Nelson, N. Metal Ion Transporters and Homeostasis. EMBO J 1999, 18, 4361–4371. [CrossRef]
- Agranoff, D.D.; Krishna, S. Metal Ion Homeostasis and Intracellular Parasitism. Mol Microbiol 1998, 28, 403–412. [CrossRef]
- Kriegeskorte, A.; König, S.; Sander, G.; Pirkl, A.; Mahabir, E.; Proctor, R.A.; von Eiff, C.; Peters, G.; Becker, K. Small Colony Variants of Staphylococcus Aureus Reveal Distinct Protein Profiles. Proteomics 2011, 11, 2476–2490. [CrossRef]
- Wu, K.; Conly, J.; McClure, J.-A.; Kurwa, H.A.; Zhang, K. Arginine Catabolic Mobile Element in Evolution and Pathogenicity of the Community-Associated Methicillin-Resistant Staphylococcus Aureus Strain USA300. Microorganisms 2020, 8, 275. [CrossRef]
- M100Ed32 | Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 20 December 2022).
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 12 July 2022).
- MiSeq System Denature and Dilute Libraries Guide (15039740). 15.
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 2011, 17, 10–12. [CrossRef]
- Krueger, F. Trim Galore 2022.
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLOS Computational Biology 2017, 13, e1005595. [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [CrossRef]
- Li, H. A Statistical Framework for SNP Calling, Mutation Discovery, Association Mapping and Population Genetical Parameter Estimation from Sequencing Data. Bioinformatics 2011, 27, 2987–2993. [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet 2000, 16, 276–277. [CrossRef]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of Pro-Oxidant and Pro-Inflammatory Activities of M1 Macrophages by the Natural Dipeptide Carnosine. Int J Mol Sci 2020, 21, 776. [CrossRef]
- Cafiso, V.; Bertuccio, T.; Purrello, S.; Campanile, F.; Mammina, C.; Sartor, A.; Raglio, A.; Stefani, S. DltA Overexpression: A Strain-Independent Keystone of Daptomycin Resistance in Methicillin-Resistant Staphylococcus Aureus. Int J Antimicrob Agents 2014, 43, 26–31. [CrossRef]
- Chen, L.; Shopsin, B.; Zhao, Y.; Smyth, D.; Wasserman, G.A.; Fang, C.; Liu, L.; Kreiswirth, B.N. Real-Time Nucleic Acid Sequence-Based Amplification Assay for Rapid Detection and Quantification of Agr Functionality in Clinical Staphylococcus Aureus Isolates. J Clin Microbiol 2012, 50, 657–661. [CrossRef]
Figure 1.
RAW 264.7 murine macrophages infected with S. aureus USA300 at 24 h p.i., captured with Leica DMI 4000B inverted microscope, 20x enlargement.
Figure 1.
RAW 264.7 murine macrophages infected with S. aureus USA300 at 24 h p.i., captured with Leica DMI 4000B inverted microscope, 20x enlargement.
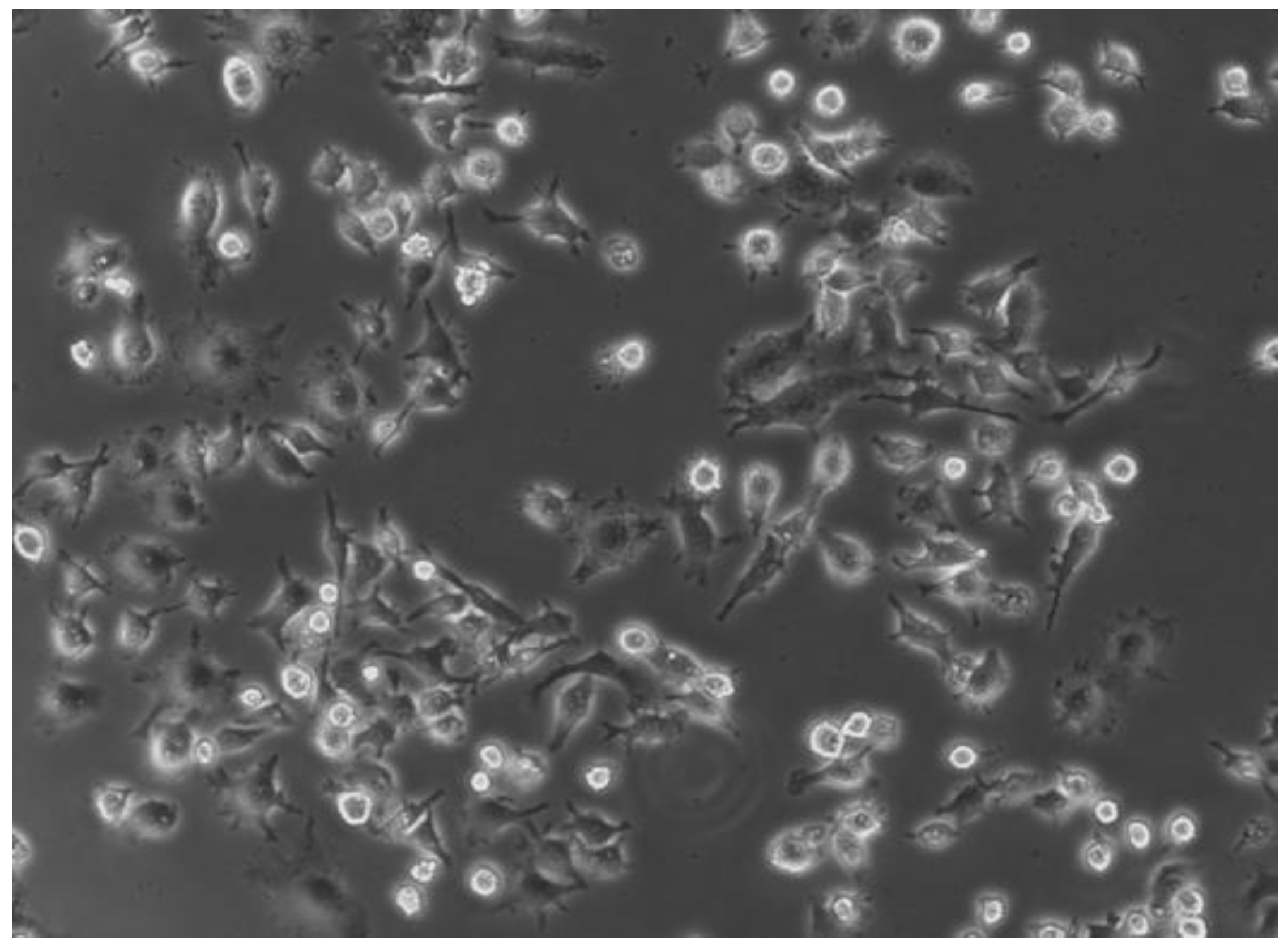
Figure 2.
a) Coexistence of S.sureus wild-type and SCV in the presence of carnosine and erythromycin used alone; b) Stable SCVs isolated from erythromycin/carnosine obtained 48h p.i. and after six passages on BA.
Figure 2.
a) Coexistence of S.sureus wild-type and SCV in the presence of carnosine and erythromycin used alone; b) Stable SCVs isolated from erythromycin/carnosine obtained 48h p.i. and after six passages on BA.
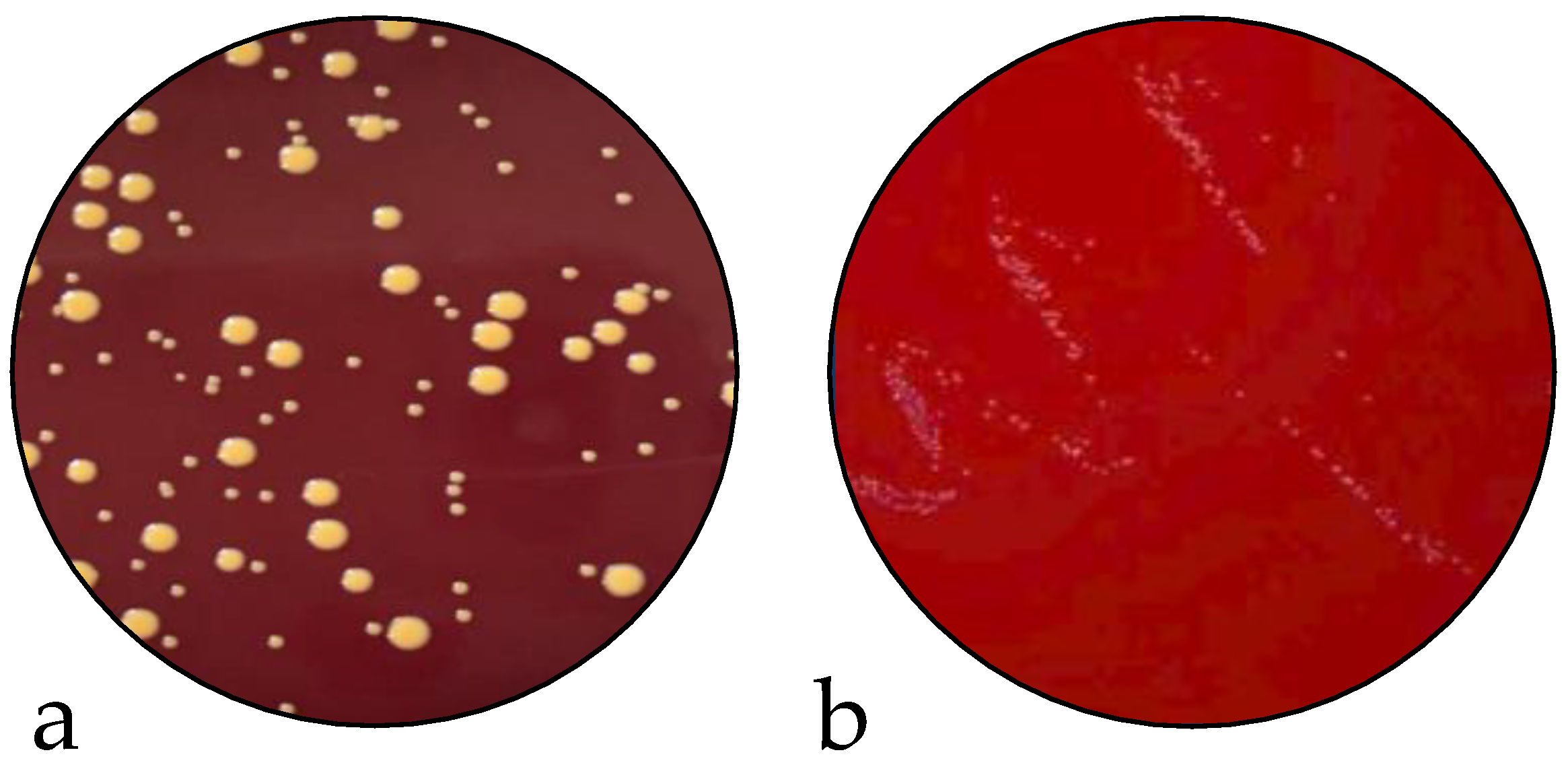
Figure 3.
S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin at 48 p.i were selected for the evaluation of mRNA expression in genes belonging to the categories a) Regulator, b) Virulence, c) Surface Protein, d) Ros Neutralization, e) Metal Ions Uptake, f) Metabolic Pathways.
Figure 3.
S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin at 48 p.i were selected for the evaluation of mRNA expression in genes belonging to the categories a) Regulator, b) Virulence, c) Surface Protein, d) Ros Neutralization, e) Metal Ions Uptake, f) Metabolic Pathways.
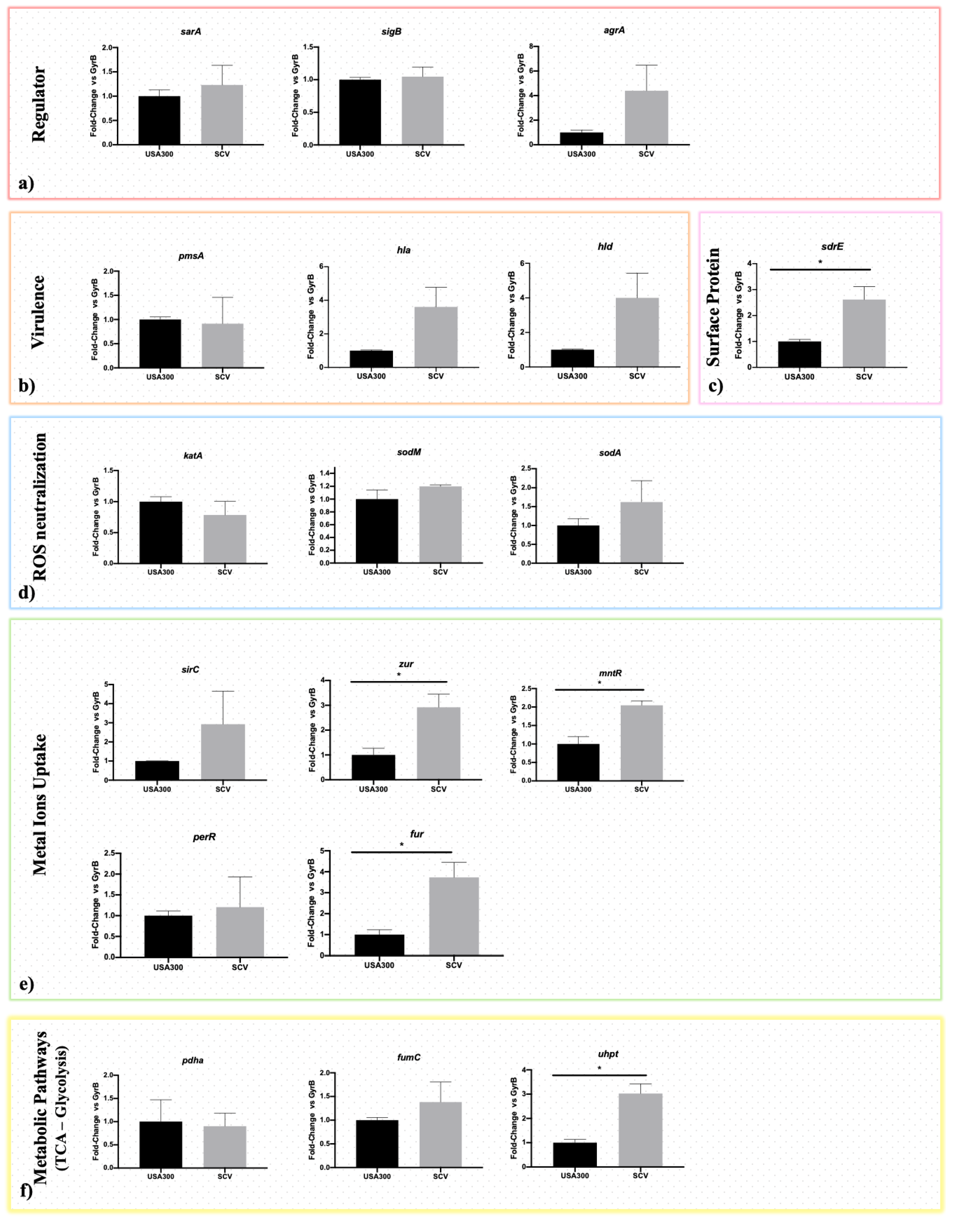
Figure 4.
Regulation of adhesion and virulence determinants in S. aureus by global regulatory loci. Arrows stand for activation; bars for repression. The molecule that acts as activator or repressor (SarA protein) is represented in the orange oval; the alternative sigma factor (SigB) in the light green rectangle; the agr quorum-sensing system (AgrA) in the light blue octagon; toxins (Hla, Hld, SdrE, and PsmA) in the yellow circles; and RNAIII in the blue rectangle. The asterisk indicates significant expressed genes. Modified from Bongiorno et al, 2020 [17].
Figure 4.
Regulation of adhesion and virulence determinants in S. aureus by global regulatory loci. Arrows stand for activation; bars for repression. The molecule that acts as activator or repressor (SarA protein) is represented in the orange oval; the alternative sigma factor (SigB) in the light green rectangle; the agr quorum-sensing system (AgrA) in the light blue octagon; toxins (Hla, Hld, SdrE, and PsmA) in the yellow circles; and RNAIII in the blue rectangle. The asterisk indicates significant expressed genes. Modified from Bongiorno et al, 2020 [17].
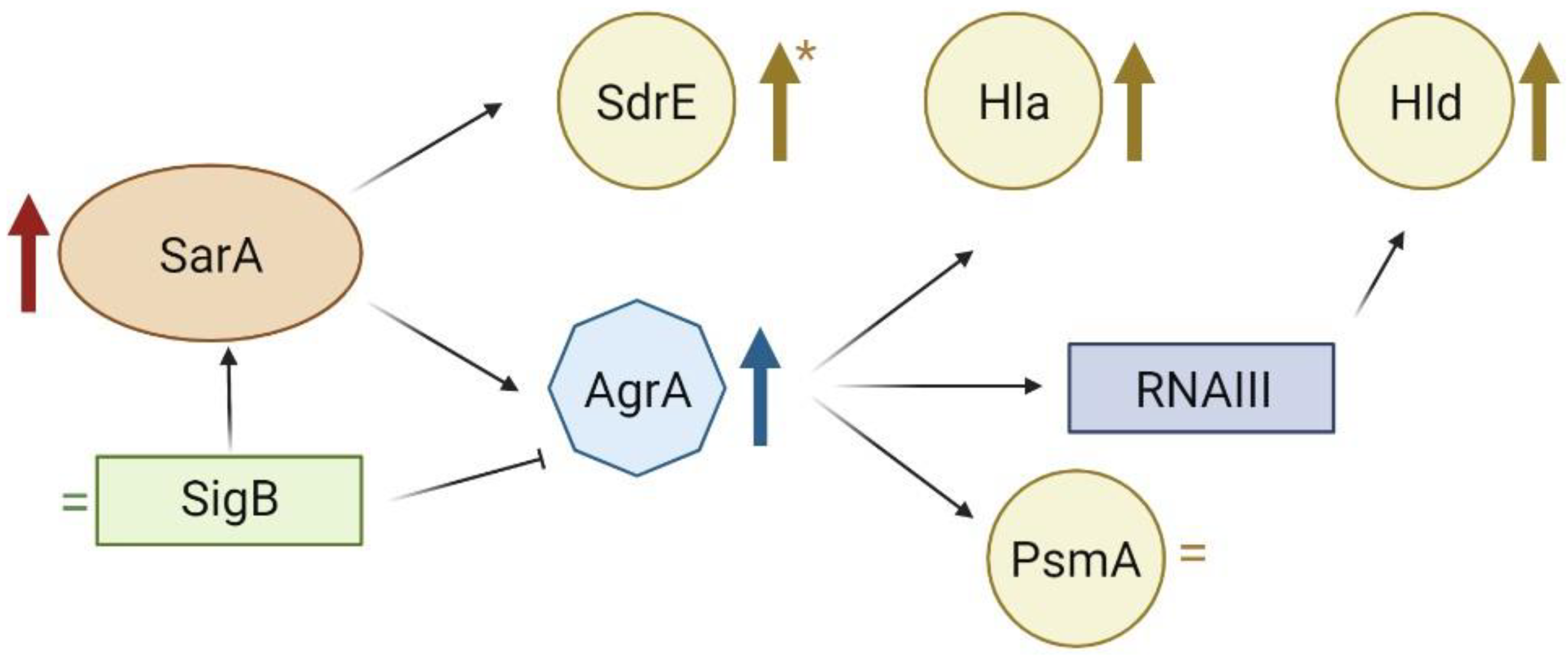
Figure 5.
Different expression of SirC, Zur, Fur and MntR affects PerR modulatory effect on KatA expression in S. aureus, managing metal ions uptake. Arrows stand for activation, bars for repression. The asterisk indicates significant expressed genes.
Figure 5.
Different expression of SirC, Zur, Fur and MntR affects PerR modulatory effect on KatA expression in S. aureus, managing metal ions uptake. Arrows stand for activation, bars for repression. The asterisk indicates significant expressed genes.


Table 1.
The statistically significant values reported are the means of the expression study conducted in triplicate for each single target. S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin at 48 p.i were selected for the analysis. ↑ up-regulated, p-value > 0,05, statistically non-significant; ↑* up-regulated, p-value < 0,05, statistically significant; = no difference in level expression
Table 1.
The statistically significant values reported are the means of the expression study conducted in triplicate for each single target. S. aureus USA300 wild-type and S. aureus USA300 SCVs isolated from the treatment with fixed concentration (20mM) of carnosine and erythromycin at 48 p.i were selected for the analysis. ↑ up-regulated, p-value > 0,05, statistically non-significant; ↑* up-regulated, p-value < 0,05, statistically significant; = no difference in level expression
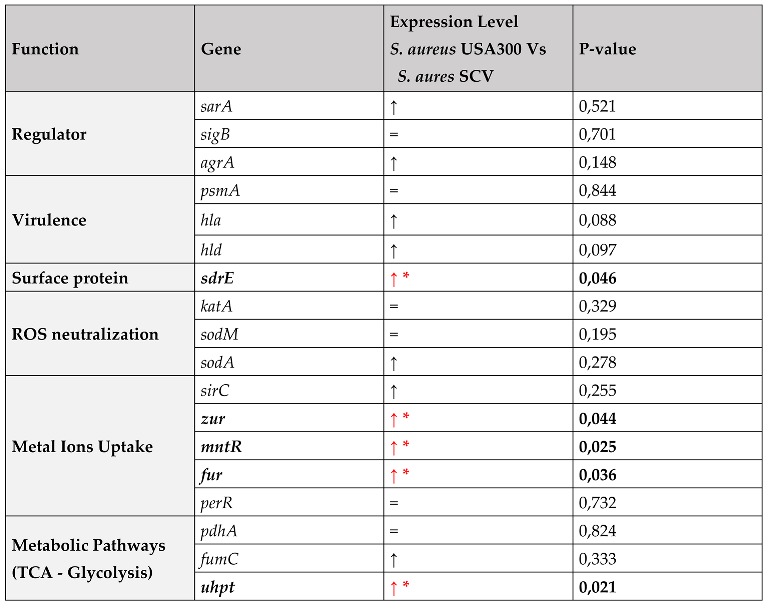
Table 2.
Primer features used to evaluate gene expression in qRT-PCR.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



