
Submitted:
27 March 2023
Posted:
28 March 2023
You are already at the latest version
Abstract
Much evidence suggests a correlation between degeneration and mitochondrial impairment. Typical cases of degeneration can also be observed in physiological phenomena (aging) as well as in neurological neurodegenerative diseases and cancer. All these pathologies have as a common denominator the dyshomeostasis of mitochondrial bioenergy. Even neurodegenerative diseases show a bioenergetics imbalance in their pathogenesis or progression. Huntington's chorea and Parkinson's disease are both neurodegenerative diseases, but while Huntington's disease is a genetic, and progressive disease with early manifestation and severe penetrance, Parkinson's disease is a pathology with a multifactorial aspect. Indeed, there are different types of Parkinson/Parkinsonism. Many forms are early onset diseases linked to gene mutation, others can appear in young adults and senescent only post-injury, and a final group is idiopathic. Huntington's was defined as a hyperkinetic disorder, while Parkinson's is a hypokinetic disorder; but in the middle, there are a lot of similarities as well as neuronal excitability, the loss of striatal function, psychiatric comorbidity, etc. In the review, we would embrace the theories that both diseases start and develop in light of mitochondrial dysfunction. These dysfunctions act on energy metabolism and reduce the vitality of neurons in many different brain areas.
Keywords:
movement disorders
; mitochondria
; energy metabolism
; synaptic plasticity
; basal ganglia
; calcium
Introduction
The Mitochondrion is an evolutionary organelle that originated from the established symbiotic relationship between alphaproteobacteria and eukaryotic cells [1]. This organelle was stabilized in the host through mitotic cell divisions [1,2]. In light of this hypothesis, it is possible that the mitochondria enhanced the development of life as we know it and represents the common factor through which many organisms function or dysfunction [2,3,4]. Mitochondria are well-known energetic units of the eukaryotic cells [2]. They supply energy in the form of ATP to support all cellular functions. In aerobic respiration, through a complex biochemical pathway, which involves the addition of different co-factors, the mitochondria aid the synthesis of ATP molecules from pyruvate [2,3]. In addition, mitochondrial proteomes which are aggregates of more than 1000 proteins, can support a wide variety of critical biochemical processes, including amino acid metabolism, nucleotide metabolism, protein synthesis, and fatty-acid catabolism [3,4].
There is abundant evidence in literature associating mitochondrial disruption with neurological and neurodegenerative diseases [4,5,6]. The brain requires high-energy consumption in the form of ATP. In fact, it was estimated that a cortical neuron in the human brain could utilize 4.7 billion molecules of ATP/sec. Furthermore, the ATP used at rest corresponds to about 5.7 kg per day [7]. The brain’s high-energy demand supports many important functions, such as neurotransmission, cellular activities required for learning and memory, neural plasticity, and synapse development [8]. Since the mitochondrion, through oxidative phosphorylation and the production of ATP, meets all the energy demands of the nervous system, even with a rapid turnover [9], mitochondrial dysfunction with consequent loss in the energetic capacity may represent a neurodegenerative trigger. Severe bioenergetic dysfunction makes neurons very vulnerable to oxidative stress and predisposed to neuronal cell death [10]. Additionally, neurodegeneration can be seen as an energy disorder, which manifests in-different aspects, depending on the severity of the process and the area involved [11]. This energetic disorder can lead to different pathologies, but sometimes with opposite symptoms, such as hypokinetic and hyperkinetic disorders. The purpose of this present review is to emphasize how mitochondrial dysfunctions, which may be associated with phenomena like inflammation and degeneration, are actually a common element in two major neurodegenerative diseases. One is purely caused by genetics, while the other resulting from a combination of genetic and environmental factors: Huntington’s disease (HD) and Parkinson’s disease (PD) respectively.
Role of mitochondria in brain energy metabolism, calcium homeostasis, and signal transduction
Mitochondria are organelles responsible for several critical processes in neuronal function and dysfunction, including energy metabolism, calcium homeostasis, and signal transduction [1]. The brain consumes about 20% of the total oxygen respired to meet the high metabolic demand of neurons that must maintain ionic gradients across membranes, transport molecules from the soma along axons and dendrites, as well as for neurotransmission [12]. The metabolic activities that generate ATP rely mainly on mitochondrial oxidative phosphorylation (OXPHOS) [1]. During the process of OXPHOS, enzymes of the Krebs cycle utilize acetyl-coenzyme A to reduce the cofactors; nicotinamide adenine dinucleotides (NADH) and flavin adenine dinucleotides (FADH2), that aid energy transfer to the electron transport chain (ETC), embedded within the extensive inner mitochondrial membrane [13,14]. The ETC consists of four complexes which transfer electrons from NADH and FADH2 to O2. Ultimately, the energy released by the transfer of electrons is used for the thermodynamically unfavorable pumping of protons against their concentration gradient from the matrix to the intermembrane space, to generate an electrochemical gradient, known as mitochondrial membrane potential (ΔΨm) [1]. This membrane potential is essential for the process of energy storage (Figure 1). The protons accumulated in the intermembrane space are then allowed to move according to a concentration gradient back into the matrix, by passing through one of the domains of the enzyme ATP synthase (or Complex V), which finally harnesses the released energy to phosphorylate ADP molecules into ATP [13]. Proper neuronal functioning is strongly linked to the retention of mitochondrial membrane potential and ATP levels [15] (Figure 1).
Indeed, mitochondrial functions go beyond the primary role of ATP production because these organelles are intimately involved in numerous processes operating within the cell, including calcium homeostasis, generation of free radical species (ROS), steroid synthesis, apoptosis and cell signaling pathways [16]. Particularly, the direction of the mitochondrial membrane potential (negative internal) has important implications, as it elicits the thermodynamic force supporting the accumulation of metal cations, especially the calcium (Ca2+) in the mitochondria [17]. Mitochondrial Ca2+ homeostasis plays a key role in cellular bioenergetics and signaling [18]. Ca2+ passage across the outer mitochondrial membrane (OMM) is mediated by the VDAC channel, whose different selectivity for cations or anions is voltage-dependent [19]. For instance, at low potentials (10 mV), the VDAC channel is highly permeable to anions and able to maintain low Ca2+ flux. Conversely, increases in the membrane potential (20-30 mV), result in the conformational change that allows a 4- to 10-fold increase in Ca2+ influx [20]. In comparison, the inner mitochondrial membrane (IMM) has considerable Ca2+ permeability: the mitochondrial calcium uniporter (MCU) contributes to the potential-dependent Ca2+ influx into the mitochondrial matrix, while the mitochondrial Na+/Ca2+ exchanger (NCLX) is one of the main units involved in Ca2+ extrusion [21]. In general, cytosolic calcium uptake by mitochondria will occur only if they are exposed to elevated Ca2+ concentrations [22,23] (Figure 1). However, within the cell, Ca2+ is compartmentalized and the average resting cytosolic concentration is remarkably low [24,25,26]. One of the main stores of cellular calcium is the endoplasmic reticulum (ER), which contributes to orchestrating cellular Ca2+ homeostasis through its interaction with the mitochondria [27]. Notably, Ca2+ uptake in the mitochondria probably occurs only at close proximity sites between the ER and mitochondria [18]. On the other hand, neuronal mitochondria, unlike those in non-neuronal cells, have a considerably lower threshold for Ca2+ uptake. In addition, ER-mitochondria contacts are critical for Ca2+ uptake in dendritic mitochondria but not in axonic mitochondria [28]. This property of axonal mitochondria is due to the presence of the brain-specific uniporter, MICU3, which causes axons to be less dependent on intracellular Ca2+ storage. In the absence of MICU3, synaptic function is impaired [29] (Figure 1)
Mitochondrial calcium-signaling in neurons regulates metabolism and energy production, which are crucial for neurotransmission and sustaining synaptic plasticity [30]. In addition, mitochondrial Ca2+ also drives the production of reactive oxygen species (ROS) [31]. Mitochondria are the primary sources of ROS in cells and actively participate in cellular redox regulation and ROS signaling [32] (Figure 1). ROS, in the form of superoxide, are natural byproducts of normal mitochondrial activity, naturally converted to H2O2, which are in turn scavenged by the enzyme catalase, to produce water [32]. Under normal conditions, through the generation of ROS and redox signaling, mitochondria can control cell metabolism and physiology, as well as inflammatory response, immune function, autophagy, and stress response [33,34,35]. Indeed, ROS and particularly hydrogen peroxide at low concentrations, act as important signaling molecules in the cell, activating several protein kinases, such as PKA, PKC, PI3K, and p38 [36]. Moreover, in immune cells, mitochondrial metabolites and ROS finely regulate signaling pathways and cell fate, thereby orchestrating the immune response [37].
The immune system comprises a diverse family of cells with multiple roles during homeostasis and inflammation, capable of using distinct metabolic programs to undertake their functions. For instance, during the immune response, effector T cells promote aerobic glycolysis, while memory T cells and regulatory T cells promote fatty acid oxidation [38]. Particularly, the mitochondria can modulate the metabolic and physiological states of different types of immune cells [39]. It can also stimulate the innate immune signaling cascade that can intensify inflammation, following cytotoxic stimuli or microbial infection [40]. For example, the innate immune receptor NLRX1, a member of the Nod-like Receptor (NLR) family, is located in the mitochondria and undertakes an important role in maintaining cellular homeostasis, following acute mitochondrial injury [41,42]. In addition, it has recently been discovered that mitochondria play a central role in initiating and regulating the NLRP3 (nucleotide-binding domain, leucine-rich-repeat containing family, pyrin domain-containing 3) inflammasome [43]. This multiprotein complex activated upon infection or cellular stress, leads to the secretion of proinflammatory cytokines, such as interleukin-1β (IL-1β) and IL-18, that trigger an inflammatory form of cell death called pyroptosis [44,45]. Although the mechanisms of NLRP3 inflammasome activation are still debated, it is widely believed that changes in the mitochondrial membrane potential, permeabilization of the outer mitochondrial membrane and increased formation of mitochondrial ROS are crucial factors in inducing the cytosolic translocation of mitochondrial molecules like cardiolipin and mitochondrial DNA, which are capable of activating the inflammasome [43]. Indeed, the overproduction of ROS and dysregulation of the redox signaling system results in oxidative stress that can lead to mitochondrial damage, induce mitochondrial DNA mutations, damage the respiratory chain, alter membrane permeability and affect the Ca2+ homeostasis and mitochondrial defense systems [46]. Accumulating evidence suggests that oxidative stress and mitochondrial injury can result in cellular DNA damage, degradation of proteins and lipids, as well as in the pathogenesis of neurodegenerative diseases [47,48]. Nevertheless, cells have an accurate endogenous antioxidant defense system that can maintain cellular redox homeostasis between ROS production and elimination to ensure normal cellular signaling and redox regulation (Figure 1) [49].
The nuclear factor erythroid 2–related factor 2 (Nrf2) is an emerging therapeutic target, since it is involved in cellular resistance to oxidants. In detail, Nrf2 controls basal expression of genes coding for enzymes and proteins involved in antioxidant and detoxifying action, repair and removal of damaged proteins and organelles, inflammatory response and mitochondrial bioenergetics [50,51]. Additionally, malfunctioning mitochondria can be selectively removed through a conserved cellular recycling process known as mitochondrial autophagy or mitophagy. In fact, the efficient elimination of damaged mitochondria prevents activation of cell death pathways, protects against ROS overproduction and maintains efficient ATP production [52]. Damaged mitochondria are swallowed into autophagic vesicles that subsequently transport them to lysosomes for destruction. Mitophagy is a strictly regulated process, modulated by mitochondrial fission and fusion proteins, BCL-2 (B-cell lymphoma 2) family proteins [53] and the PINK1/Parkin pathway [54]. As all other defense mechanisms fail, the neuron can orchestrate its self-destruction by activating an intrinsic suicide program, otherwise known as apoptosis [55] (Figure 1). The underlying mechanisms of apoptosis are very sophisticated and engage an energy-dependent cascade of molecular events. In addition, it can follow different molecular pathways, one of which is the intrinsic pathway that involves the mitochondria [56]. Indeed, these organelles represent the site where anti-apoptotic and pro-apoptotic proteins interact and the origin of signals that initiate the activation of caspases, the cysteine proteases capable of cleaving many cellular substrates to disrupt cellular contents [57].
It is therefore understood that mitochondrial integrity and homeostasis are prerequisites for proper cellular functioning, especially for neurons, which are polarized, complex cells with high energy demand [58]. Not surprisingly, neurons have the highest content of mitochondria compared to other cell populations. Mitochondrial functionality is essential for ensuring membrane excitability and to perform the complex neurotransmission and plasticity processes.
Firstly, the mitochondria provide the energy needed to undertake a wide range of neuronal functions, such as the maintenance of resting membrane potential, the restoration of ionic balance after depolarization, the cycling of synaptic vesicles, as well as the transport of proteins and organelles from the soma to distal sites. Interestingly, the mitochondria in axons and dendrites have different morphologies, some are small and sparsely distributed in the former, whereas some are elongated and densely distributed in the latter. In addition, axonal and dendritic mitochondria differ in movement, metabolism and responses to neuronal activity [59].
Secondly, several lines of evidence support the role of mitochondria in the mobilization and recycling of synaptic vesicles [60]. Neurotransmission is underpinned by endocytosis and the local filling of synaptic vesicles in the presynaptic terminal [61]. Furthermore, the mitochondria support synaptic activity through cytosolic calcium reabsorption, a critical buffering mechanism for establishing and maintaining synaptic activity and preventing neuronal toxicity and excitotoxicity. Moreover, mitochondrial calcium buffering appears to be necessary at the synapse, even when the neuron is at rest: synaptic terminals lacking mitochondria show a higher frequency of spontaneous release of neurotransmitter-containing vesicles [62].
Overall, the importance of the mitochondria in neurons is unequivocal. Therefore, mitochondrial dysfunction leads to a plethora of severe conditions, from impaired neuronal development to various neurodegenerative diseases.
Role of mitochondria in neurodegenerative disease
A great deal of evidence shows the effects of mitochondrial dysfunction in degenerative diseases [63]. Diseases caused by mitochondrial alterations often show a neurodegenerative component involving the nervous system. Similarly, mitochondrial defects are frequently observed in tissue samples taken from patients with a neurodegenerative disorder. This evidence reflected the high-energy requirements for all biological processes [64]. The continuous increase in energy demand, mainly due to excess food consumed daily, constant increase in energy required for thermoregulation or the increase in energy needed to counteract the environmental toxicity caused by humans, leads to an overwork of the mitochondria, resulting in impaired bioenergy efficiency [64,65]. It was demonstrated that the cells exposed to an opulent nutrient environment are inclined to have their mitochondria in a fragmented state. Moreover, the mitochondria observed in cells in malnourished conditions remain for a longer period of time in the associated state [66,67]. This portrays the fact that the mitochondria can change their architecture and therefore, their bioenergy capacity according to external events.
A degenerative disease is regarded as a type of non-physiological condition whose origins in a tissue or organ worsen over time. Energy failure has been linked to degenerative processes as well as cancer, aging, neuroendocrine disease, neurodegenerative disease and inflammatory diseases. Likewise, results from many studies support the involvement of the mitochondria in neurodegenerative diseases, particularly in PD [68,69,70,71,72]. Also, studies relating to the MPTP toxin first highlighted the role of the mitochondrial complex I dysfunction and neurodegeneration in PD [73]. In association with other used toxins, such as rotenone and Paraquat, MPTP offered a new insight into defining the effective role of mitochondrial bioenergy in neurodegeneration [74]. Indeed, more than five thousand manuscripts dealing with the association between Parkinson’s disease and mitochondrial alteration can be found on PubMed (Figure 3). In addition, due to studies on PD, particularly genetic Parkinsonism, all mitochondrial dysfunctions leading to cell death have been well defined [75]. PINK1 gene mutation, responsible for an early onset Parkinsonism, serves as a good example [76]. This gene codes for the mitochondrial protein, phosphatase and tensin homolog serine/threonine-protein kinase 1(PTEN-induced kinase 1) [77]. PTEN- protein can protect cells against oxidative stress, proton chain dysfunction and bioenergy failure [76,78]. The PINK1 gene has also been identified as an oncogene with tumor suppressor properties [77,79,80]. Additionally, the PINK1 protective role has been observed in many disorders characterized by progressive inflammation and neurodegeneration, such as Alzheimer’s disease, multiple sclerosis, amyotrophic lateral sclerosis and HD [77]. In physiological conditions, PINK1 is translocated inside the mitochondria in its mature isoform with the aim to withstand the activity of the mitochondrial chain and to produce (at the level of complex I) the molecules of ATP showing the maximal bioenergetics efficiency (Figure 1). Moreover, PINK1 is also located in the inner mitochondrial membrane and can interact with the chaperone, TRAP1 (recognized also, as an interactor of the type 1 tumor necrosis factor receptor), to maintain important bioenergetics and proteostatic functions [77,81]. Cleaved PINK1 can interact with other chaperon proteins and when it is located in the cytosol, it can activate the m-Tork/Atk pathway. Besides, PINK1 can mediate the phosphorylation of another important gene for PD: Parkin. The coupled action of PINK1 and Parkin may induce mitochondrial fusion in order to cause the elimination of dysfunctional mitochondria. Furthermore, PINK1 is also involved in the formation of an autophagosome complex by the activation of beclin protein and may regulate the apoptotic process [77]. The role of the PINK1 gene in PD was well investigated by the use of animal models and cellular cultures of human fibroblasts [82,83,84,85,86,87] (Table 1). These studies have confirmed the role of bioenergetics efficiency in the maintenance of health state or the progression of neurodegeneration. However, the absence of PINK1 results in low bioenergetic efficiency and programmed cell death after each minor stress [88,89]. It is very interesting to note that PINK1 not only has a key role in degeneration, oncological degenerative processes and neurodegenerative PD, but also in neurodegeneration found in HD. Parkinson’s disease (PD) and HD are two different neurological diseases involving the central nervous system. The symptomatology of both is quite similar (cognitive impairment, limb inflexibility, and problems in walking or talking), but while PD results from a combination of genetic and environmental factors, HD is only an inherited genetic disease. Both pathologies have a common factor, which is the loss of bioenergetics efficiency [77,90].
Huntington disease is an incurable degenerative disorder caused by a mutation in the huntingtin gene, where the CAG sequence is excessively repeated. This mutation alters numerous cellular processes and leads to cell apoptosis. One important alteration is caused by the impairment of the mitochondrial metabolism. In a study conducted on the fly model, the formation of an abnormal ring-shaped mitochondria it was observed; this particular shape was previously identified in mitophagy-blocked cells in which PINK1 overexpression was able to rescue the regular shape and function of the mitochondria. PINK1 over-expression was able to improve bioenergetics efficiency (increasing ATP levels) and rescue neuronal integrity in the adult drosophila model of HD [90]. HD is a degenerative pathology caused by pathological expansion of CAG repeats in the Huntington gene coding for the Huntingtin protein. The gene is located on the chromosome number 4 and is characterized by high levels of polymorphism. Unlike PD, HD has an age range of about 35-44 years and an estimated post-onset life span of 15/18 years. It is also characterized by an advanced motor disability, including chorea [91]. Enough evidence suggests that the mutation may cause alteration in mitochondrial trafficking, increase in oxidative stress, dyshomeostasis in intracellular calcium content, alteration in bioenergetics and alteration in a respiratory mitochondrial chain [92,93,94,95]. Huntington disease, PD, and Alzheimer’s disease are three neurodegenerative diseases that have 37 common genes and about 40% of whose products act at the mitochondrial level [96]. These neurodegenerative diseases are coupled to a physiological degenerative process called aging or senescence that starts at the mitochondrial level and results in reduced bioenergetics efficiency [97]. Cellular senescence is characterized by heavy changes in cellular metabolism and an increase in pyruvate utilization that may produce differences in phosphorylation states, thereby increasing the activity of the mitochondrial pyruvate dehydrogenase complex and ROS production [98,99]. These features may lead to an impairment of cellular function that induce degeneration like in the case of cancer or other neurodegenerative processes [100]. Moreover, recent data have demonstrated that key oncogenes and tumor suppressors modulate mitochondrial metabolism and dynamics. Indeed, different types of cancer result in more or less sensitivity to the modulation of the mitochondrial function in the lesion caused by the tumor [101]. Therefore, the mitochondria play a cardinal role in many degenerative processes, which is mostly demonstrated in animal models.
Mitochondria bioenergy in Parkinson’s disease and Huntington disease in rodents animal models
Parkinson’s and Huntigton’s diseases are both neurodegenerative diseases with multiple mitochondria bioenergy alterations related to metabolism oxidative stress, dynamics biogenesis transport and mitophagy (Table 1). As a result of the close relationship between neurodegeneration and mitochondria bioenergy, one might presume that mitochondria homeostasis changes with the release of calcium and NO as well as reactive oxygen species (ROS), will affect the unfolded protein response modulating specific signalling molecules and reprogramming mitochondria bioenergy [102] (Table 1). The mice models used in the study of PD and HD, serve as valuable tools in defining the role of mitochondria bioenergy in pathogenic mechanisms of these diseases (Table 1).


- Parkinson disease
Neurotoxin-induced and autosomal-dominant PD models: Parkinson’s disease animal models are associated with multiple mitochondrial defects [for review [102,103,104,105] and (Table 1). Indeed, human idiopathic PD patients showed abnormalities in mitochondrial activity with reductions of complex I, NADH: ubiquinone oxidoreductase and strikingly reduced succinate: cytochrome c oxidoreductase, suggesting an etiological role in the pathogenesis of PD [106].
Chemically, the neurotoxin agent is widely used to recapitulate Parkinsonian features in various animal models [107,108]. In fact, the neurotoxin, 1-metil-4-fenil-1,2,3,6-tetraidropiridina (MPTP), rotenone and paraquat, block mitochondrial bioenergetics in dopaminergic neurons and induce Parkinsonian syndrome [for review [109,110,111,112,113,114,115]. However, it is important to consider that overexpression of α-synuclein or agents that generate stress or cytosolic acidification, show the translocation of α-synuclein into the mitochondria or in the mitochondrial membrane [116,117,118]. α-synuclein is a presynaptic neuronal protein that is linked to familial and idiopathic PD [119]. Both the over-expression and loss-of-function of the SNCA gene contribute to the manifestation of the disease [120], according to the “α-synuclein cascade hypothesis” [121].
Experimental studies in A53T mutant mice, a transgenic animal model that expresses human α- synuclein, show that they develop mitochondrial DNA damage and degeneration with an apoptotic-like death of neocortical, brainstem, and motor neurons [123]. In the same animal model, the mitochondrial complex IV activity was reduced significantly in the spinal cord [122]. Additionally, in-vivo human α-synuclein expression is associated with a decrease of Drp1 (Dynamin-related protein 1), a major player in the regulation of mitochondrial dynamics and the maintenance of their proper function [123]. On the other hand, the dysfunction of Drp1 in the mitochondria is also associated with enlarged neuronal mitochondria [123]. Other proteins, such as Mitofusin1(Mfn1) [123], that facilitate mitochondrial fusion are decreased in these models, correlate with the mitochondria bioenergy dynamic changes. Previous studies in A53T mice suggest that α-synuclein distress the mitochondrial morphology and reduces both Mfn1 and Mfn2 in an age-dependent manner [124]. In addition, an in-vivo study in midbrain dopaminergic neurons suggested that α-synuclein predominantly accumulates in the central mitochondrial membrane and interacts with the complex I, resulting in the impaired activity of the mitochondrial electron transport chain [125]. Consistent with these findings, other experimental analyses show that after post-inoculation of human α-synuclein in rats, its accumulation in striatal dopaminergic terminals from the SNpc and early synapse loss was observed [126]. To explain this early synapse loss, a proteomic analysis using a real-time cell metabolic method and isolated synaptosomes was used to investigate the functional and molecular mechanisms. Upon injection in isolated striatal synapses, multiple dysfunctions of mitochondrial bioenergetics and morphology were detected with the upregulation of PRKAG2 and TTR respectively, which are sensor proteins of the cell energy status and markers of oxidative stress [126]. Moreover, ultrastructural examination of subsequent human α-synuclein accumulation showed an altered expression of Rab5 endocytic and LC3 autophagic proteins that potentially reflect an accumulation of autophagic vesicles [126]. This study allowed the evaluation of an interesting aspect, which is the effect of oxidative stress and the mitochondrial bioenergy alteration in both dopaminergic neuron death and striatal neurons. These could be both responsible for the induction of Parkinsonian syndrome.
The neurons are high-energy consumers and employ most of the energy at the level of the synapse to preserve and restore ionic gradients and for the uptake and recycling of neurotransmitters [127]. Various mouse lines expressing a different type of α-synuclein present impaired dopamine neurotransmission and striatal synaptic plasticity [128]. In rodents, the injection of α-synuclein with an adeno-associated viral vector blocks the induction of long-term potentiation (LTP) and long-term depression (LTD) that are normally expressed in medium spine neurons (SPNs) [115,129,130,131], which are associated with early memory and motor alterations. There is sufficient evidence showing that α-synuclein misfolding and aggregation lead to mitochondrial stress, which appears to be related to the dysfunction of synaptic plasticity. Another interesting aspect is that α-synuclein overexpression via the endoplasmic reticulum (ER), promotes Ca2+ transfer in the mitochondria, while its silencing impairs mitochondrial function by loosening the ER-mitochondria interface [132]. However, the precise mechanism by which α-synuclein accomplished pathological characteristics and specific cell death remain obscure. Nevertheless, it is important to note that α-synuclein plays a pro-apoptotic role in different neuronal cells [133]. The wild-type α-synuclein can defend neurons from apoptosis by the inhibition of caspase-3, after which the mutant α-synuclein loses this activity [133]. Caspases-3 actions then cause general damage and degeneration, aggregates in synapses and persists in neurons without causing acute cell death [134,135,136] and (Table 1).
- Autosomal-recessive PD models
The discovery of many genes related directly or indirectly to the mitochondria in PD, underlines the implication of mitochondria bioenergy in its pathogenesis. For instance, genes linked to early-onset recessive PD include: Parkin [83], PINK1 [76] and DJ-1 [137]. These genes have been strongly implicated in the mitochondria morphology, damage and degradation via mitophagy [138], which is involved in the mitochondrial activity and quality control mechanisms [77,139,140,141,142]. Parkin encodes a protein localized in the cytoplasm that contains an N-terminal ubiquitin-like domain with a function as E3 ubiquitin-protein ligase [143,144,145]. Parkin knockout (KO) mice show alteration in the levels of different proteins involved in detoxification, stress-related chaperones, without neuronal degeneration [146]. Parkin also has neuroprotective effects through the regulation of different cellular processes or pathways such as mitochondrial swelling and cytochrome c release [147,148,149]. Normally, Parkin resides in the cytoplasm but can translocate to depolarized or damaged mitochondria to mediate their removal by mitophagy in cooperation with PINK1 and potentially other factors [150]. In a mouse model, Parkin KO was reported to cause a decrease of subunits of complexes I and IV with reduction of peroxide reductases. In addition, the serum also aids the reduction of antioxidant capacity and increased protein of lipid peroxidation [151]. Other studies on Parkin-deficient mice showed an increase in the content of dopamine in the striatum and the consequent reduction of the excitability of striatal SPNs [152]. Deficits in glutamate neurotransmission and amphetamine-induced dopamine release were also observed in other parkin mutant mice with an increase in the metabolism of dopamine (MAO) [153]. Functional analysis of striatal SPNs showed an impairment of bi-directional corticostriatal synaptic plasticity with the loss of the LTD and LTP in Parkin KO mice, while synaptic plasticity was not altered in the hippocampus of these animals [154]. The dopaminergic defect [152,153] may explain the selectively enhanced sensitivity to the striatal group II metabotropic glutamate receptor in cortically-evoked excitatory post-synaptic potentials recorded from SPNs. This reinforcing effect is an adaptive change [78]. Furthermore, recent evidence in Parkin KO rats suggests that lack of Parkin is also necessary for the maintenance of post-synaptic endocytosis of AMPARs and for the decreased expression of the post-synaptic protein, Homer1, which is essential for coupling the AMPA receptor endocytic zones with the post-synaptic density [155]. On the other hand, changes to the numbers or function of ER-mitochondrial contact sites may also affect mitochondrial calcium homeostasis regulated by Parkin [156]. Calcium uptake into the mitochondrial matrix results in enhanced respiratory function, thereby tuning synaptic activity [60,157]. Recent studies also showed that the conditional Parkin KO in adult animals, expresses the progressive loss of dopamine neurons. Moreover, the overexpression of PGC-1α leads to the selective loss of DA neurons in the substantia nigra [158].
Another gene linked to early-onset recessive PD is PINK1, which encodes a protein with a serine/threonine kinase catalytic domain, whether cytosolic or mitochondrial-associated [77]. PINK KO mice models exhibited significant functional impairment of synaptic plasticity and mitochondria morphology without motor deficits or dopaminergic neuronal loss [83]. Consistent with these findings, PINK KO mice models generated with the silencing of the PINK1 gene did not develop dopaminergic neurodegeneration [159]. The PINK1 heterozygous KO mouse also showed selective impairment of LTP with a normal expression of LTD [160]. Nevertheless, low doses of rotenone was sufficient to induce severe alterations of the corticostriatal LTP and LTD [84]. This led to the proposal that the PINK1 homozygous KO model represents a good model to study the effect of gene- environment interaction. Although PINK1 KO rats show mitochondrial alteration, locomotor deficits and α-synuclein aggregate in several brain regions, such as the cerebral cortex and dorsal striatum, and is responsible for the degeneration of the substantia nigra [161,162,163,164]. However, this model presents clear symptomatic deficits that occur at 4 months and 9 months [163]. In the same model, using magnetic resonance spectroscopy, the mitochondrial metabolomic alterations increased in the cortex and striatum during the asymptomatic period, which coincides with the normal metabolic alterations [163]. Functional, bioenergetics experiments revealed that mitochondrial alterations in PINK1 KO rats is due to impaired complex I respiration, a dysfunction in ATP synthase and reduced substrate oxidation [162]. New scientific research performed in animal models with the deletion of PINK1 mice, provided an indication that lack of PINK1 results in impaired synaptic plasticity, which is caspase-mediated [86,89]. Caspase-3 presents an additional role in neuronal processes, including the regulation of synaptic function and pruning [135,136,165]. Previous studies of PINK1 KO show a loss of LTP and LTD with alterations in dopamine release [83]. Indeed, low activation of the caspase-3 is critical in restoring cortical-striatal LTD [86,89]. However, high activation is involved in degenerative processes [136]. In PINK1 KO mice analysis with electron microscopic and other functional studies of the striatum, revealed significantly enlarged mitochondria shape and impaired activity [166]. Also, the functional examination showed impaired mitochondrial respiration and aconitase activity in the striatum, but not in the cerebral cortex [166]. Additional studies of PINK1 KO mice showed a decreased dopamine release in the dorsal striatum in an age-dependent manner with an impairment of the basal mitochondria respiration [88].
DJ-1 gene mutations cause autosomal recessive early-onset PD. DJ1 encodes a protein of the superfamily, which belongs to the ThiJ/PfpI-like. Under normal conditions, DJ-1 is confined throughout the cytoplasm of neurons with a small amount localized to the mitochondrial matrix and intermembrane space [167], indicating a role in the balance of the mitochondrial physiology [17]. In a study, it was reported that DJ-1 KO mice failed to express PD features, such as substantia nigra degeneration or the formation of protein inclusions similar to PINK1 and Parkin KO mice [140]. Recent experiments in DJ-1 KO mice showed increasing calcium influx into the neuron during its activity, creating basal mitochondrial oxidant stress [168,169]. Functional analysis in isolated mitochondria showed increased ROS and decreased aconitase activity in DJ-1 KO mice [170]. Additionally, DJ-1 gene deletion revealed a deficit in scavenging because the loss DJ-1 pathway, resulted in the loss of atypical peroxiredoxin-like activity [171]. In contrast, an independent study of DJ-1 KO mice reported an increase in mitochondrial respiration-dependent H2O2 consumption, mitochondrial Trx activity, total glutathione (GSH and GSSG respectively) levels, mitochondrial glutaredoxin (GRX) activity and a decrease in mitochondrial glutathione reductase (GR) activity [172]. Furthermore, new research on the semi-quantitative measurement of cerebral metabolites in DJ-1 KO mice, showed significantly increased glutathione (GSH) levels and GSH/glutamate (Glu) ratio in prefrontal cortex [173] (table 1). It is important to note that the GSH system is the more important than the antioxidant system in ROS detoxification in the brain [174,175,176]. Accumulated scientific information hypothesized that the increased sensitivity to oxidative stress in DJ-1 KO mice and dopaminergic neuronal cells is related to a decrease in ROS scavenging arising from deteriorated peroxidase-like scavenging with a deficiency of Nrf2 transcriptional factors and increased mitochondrial dysfunction, due to complex I deficiency [170,177,178,179,180]. Although DJ-1 KO mice had normal corticostriatal LTP, the LTD was absent [152]. Overall, the data from animal models provide a framework that suggests that genes linked to early-onset recessive PD are important for maintaining normal mitochondria bioenergy function (Figure 2) Gradually, the mutation protein described above that is related to the mitochondrial bioenergy, activates the response to mitigate mitochondrial alterations. Indeed, the genetic inactivation of PINK1/Parkin/DJ-1 with a triple KO mouse, exhibits abnormalities in the mitochondrial pathway, activity or morphology without PD-related phenotypes [181].
- Huntington disease
With regards to HD, several transgenic mice have been engineered to address specific pathological characteristics [182]. Multiple scientific information sources implicate that gain and/or loss of function of mutant HTT, leads to impaired mitochondrial bioenergy with alteration of oxidative stress, dysfunction of mitochondrial trafficking, and mitochondrial calcium dyshomeostasis [95]. In fact, malonate and 3-nitropropionic acid (3-NPA) are considered potent neurotoxins and are used to induce experimental pharmacological models of rodents with HD [183]. These oldest models provided the first piece of evidence that mitochondria bioenergy targeting the electron transport chain, is involved in the pathophysiology of HD. However, degenerating mitochondria have been identified in different areas, brain, liver, and muscle in genetic mouse models of HD [184,185,186] (Table 1).
Both R6/1 and R6/2 strains express a single copy of a human genomic fragment that contains 116 and 144 CAG repeat HTT under the control of the human HTT promoter [187]. These mice express relatively rapid onset and progression of symptoms that include motor defects and neurodegeneration. Another major contribution to the research was the generation of knock-in mice models where CAG was repeated extensively (CAG94 and CAG140), compared to the human trans gene, for developing HD-associated phenotypes [182]. The R6/2 mice models have been the most widely used because the disease starts earlier with a rapid progression compared to the YAC, BAC, and other knock-in lines [188]. Transgenic animal models expressing the amino (N)-terminal fragment of the mutant form of HTT (R6/2), show an increase in 8-hydroxy-2-deoxyguanosine OH(8)dG during the late stages of the illness [189]. Therefore, oxidative stress occurs in the striatum before the onset of motor symptoms. Also, biochemical analysis performed in the striatum of R6/2 mice at 12 weeks showed a significant reduction of the mitochondrial complex IV activities and a decrease in aconitase in the cerebral cortex [190]. In accordance with these data, new scientific research using synaptosomes isolated from R6/2 mice showed a decrease in the mitochondrial mass with increased ROS production and antioxidant levels in the striatum compared to the cortex [191], indicating oxidative stress. Additionally, in the same model in depolarized conditions, the oxygen consumption rates in synaptosomal were significantly amplified, which was accompanied by a clear increase in the mitochondrial proton leak of the striatal synaptosomes [191], indicating synaptic mitochondrial stress. Indeed, synaptic dysfunction occurs in various mouse models with HD, which appears to be associated with the dysfunction of LTD, but displays a physiological LTP with lost synaptic depotentiation. These have been directly connected with the onset of cognitive deficits in HD models [192,193,194,195,196]. Using the brain slices from R6/1 mice at different ages depicted a significant increase of Ca2+ content after glutamate stimulation. Such alterations have an impact on mitochondrial dysfunction with a decrease in NAD(P)H fluorescence and loss in Mitochondrial membrane potential (ΔΨm) [197]. Several groups characterized the reduction of N-acetyl aspartate in R6/2 mice, similar to symptomatic HD patients [198,199,200,201,202] and noticed fluctuations in some metabolites, which suggest the impairment in cellular metabolism and mitochondrial bioenergy [203].
Another interesting contribution to assessing the deficits of mitochondrial function and redox deregulation is the experimental data performed with PET in YAC128 transgenic mice [204]. PET analysis displays a specific accumulation of [64Cu]-ATSM in the striatum with concomitant alterations in mitochondrial respiration and ATP production with increased complex II and III activities. In turn, an increase in mitochondrial H2O2 levels in YAC128 mice and defects in Ca2+ handling [204] support an early increased striatal susceptibility of mitochondrial. Furthermore, functional assays revealed an impairment of the mitochondrial respiratory capacity in the striatum and cortex of R6/1 mice. It is important to note that N-acetylcysteine administration delayed the onset and progression of motor deficits in R6/1 mice [205], which may have reduced both excitotoxicity and oxidative stress in the striatum. A recent study established that voluntary wheel running reduces hindlimb clasping in the R6/1 mouse model. Furthermore, chronic exercise in the R6/1 showed the clasping phenotype with normal mitochondrial respiration in the cortex and striatum, suggesting that mitochondrial dysfunction may not necessary for the progression of symptoms [206]. At the molecular level, different studies explored the role of the mitochondria in apoptotic cell death processes with an elevated caspase activity in HD mice models [184,207]. Interestingly, HTT has been shown to be involved in-vitro with caspases-1,2 and 3 [208,209,210]. In fact, immunostaining analysis of YAC72 mice displays an increase of caspase-2 in the SPNs within the striatum with concomitantly decreased levels of a brain-derived neurotrophic factor in the cortex and striatum at 3 months [208]. This indicates that caspase-2 participates selectively on the neurodegeneration of SNPs in the striatum. In contrast, in YAC128 mice models, the lack of caspase-2, results in protection from the well-validated motor and cognitive features of HD, [211]. Moreover, previous studies in transgenic mice expressing exon 1 of the human HTT demonstrate that intracerebroventricular administration of a caspase inhibitor 1 delays the disease progression of the disease [211] (table 1).
In conclusion, while HD animal models have not yet provided definitive evidence whether or not the mitochondria bioenergy is critically involved in HTT-induced disease symptoms compared to PD animals models’ genes, mitochondrial bioenergy defects are nonetheless, a major component for both diseases progression. On the other hand, the creation of HD mouse models using different strain backgrounds and expressing only a portion of the HTT protein leads to increased difficulty in understanding compelling findings that were obtained with the genetic models of PD.
Mitochondria Bioenergy in Parkinson’s disease and Huntington disease, based on human evidences
Assessment of mitochondrial activity (or dysfunction) is not easy in the CNS of living patients. However, analyzing peripheral tissues and fluids can offer a reliable measure. Although the core pathology in PD and HD is central, a number of substantial abnormalities occur at the systemic or peripheral level. Peripheral blood mononucleate cells (PBMCs) are one of the most reliable “in-vivo PD models”. Peripheral blood mononucleate cells present several alterations in critical metabolic pathways and accumulate α- synuclein pathological forms, well recapitulating PD-related neuropathology [212,213]. In a recent work, PBMCs mitochondrial bioenergetics (Figure 2) from PD patients was assessed by the Seahorse Bioscience technology. The results showed a peculiar pattern of mitochondrial respiration, including normal basal respiration, significant augmentation of the maximal respiratory and spare respiratory capacity, and a tendency to increase ATP production. The increased spare respiratory capacity was found to follow the disease duration and the severity of motor disturbances, revealing some mitochondrial adaptations to the higher bioenergetics requirements occurring at later disease stages [214]. Similarly, consistent findings of greater mitochondrial respiration were also observed in different cell lines obtained from PD patients, such as lymphoblasts (immortalized blood lymphocyte-derived cells) [215] and fibroblasts [216,217]. Therefore, it is reasonable to assume that in PD patients, unlike animal models, the mitochondrial activity may increase in a compensatory manner. Indeed, the “nuclear factor erythroid 2-related factor 2” (Nrf2) pathway, a master regulator of the cellular defence and mitochondrial activity, can be overexpressed in PD patients’ PBMCs in proportion to the disease duration, suggesting a systemic defensive response to the PD clinical-pathological progression [213]. Otherwise, changes in respiratory activity could reflect defects in mitochondrial structures, metabolism, or respiratory proteins functioning [218]. In another study, PBMCs increased glycolysis and deficits in superoxide dismutase, together with a peculiar mitochondrial vulnerability in monocyte subpopulation in PD patients’ [219]. Substantial evidence can also be obtained from the assay of mitochondrial dysfunction biomarkers in PD patients’ fluids. Indeed, metabolomic studies performed in blood and cerebrospinal fluid (CSF), disclosed various abnormalities in metabolic pathways related to the mitochondria (in alanine, branched-chain amino acids, fatty acids and acylcarnitines levels; in steroidogenesis; in glutathione cycle) [220,221,222]. Likewise, the redox balance-related biomarker levels, such as uric acid [223,224,225,226], catalase, total glutathione [227], nonmercaptalbumin (oxidized form of albumin) [228], oxidized glutathione [213], oxidized DJ-1 [229], α-klotho [230], lactoperoxidase [231], and heme-oxygenase-1 [232], were altered in the same fluids. Genomic studies performed on peripheral blood showed the down-regulation of genes critical for mitochondrial functions (COX4I1, ATP5A1, and VDAC3) [233], while the analysis of blood-circulating extracellular vesicles demonstrated the reduction of mitochondrial components (i.e., ATP5A, NDUFS3, and SDHB) in PD patients [234]. Furthermore, analysis of mitochondrial bioenergetics has also been performed in peripheral tissues of HD patients with some controversial findings, depending on the matrix or the experimental technique used. Indeed, mutant huntingtin is expressed out of the CNS as well, accounting for the substantial impairment in multiple cellular pathways related to the mitochondria and redox balance [235] (Figure 2). It has also been observed that skin fibroblast may show some energetic, respiratory, redox, and morphological abnormalities [236,237]. Likewise, several functional or structural mitochondrial defects have been noticed in HD patients-derived lymphobasts [238]. Finally, increased oxidative damage, reduced antioxidant capacity, and mitochondrial abnormalities have been tracked even in HD patients’ blood through different fluid markers [239].
Figure 2.
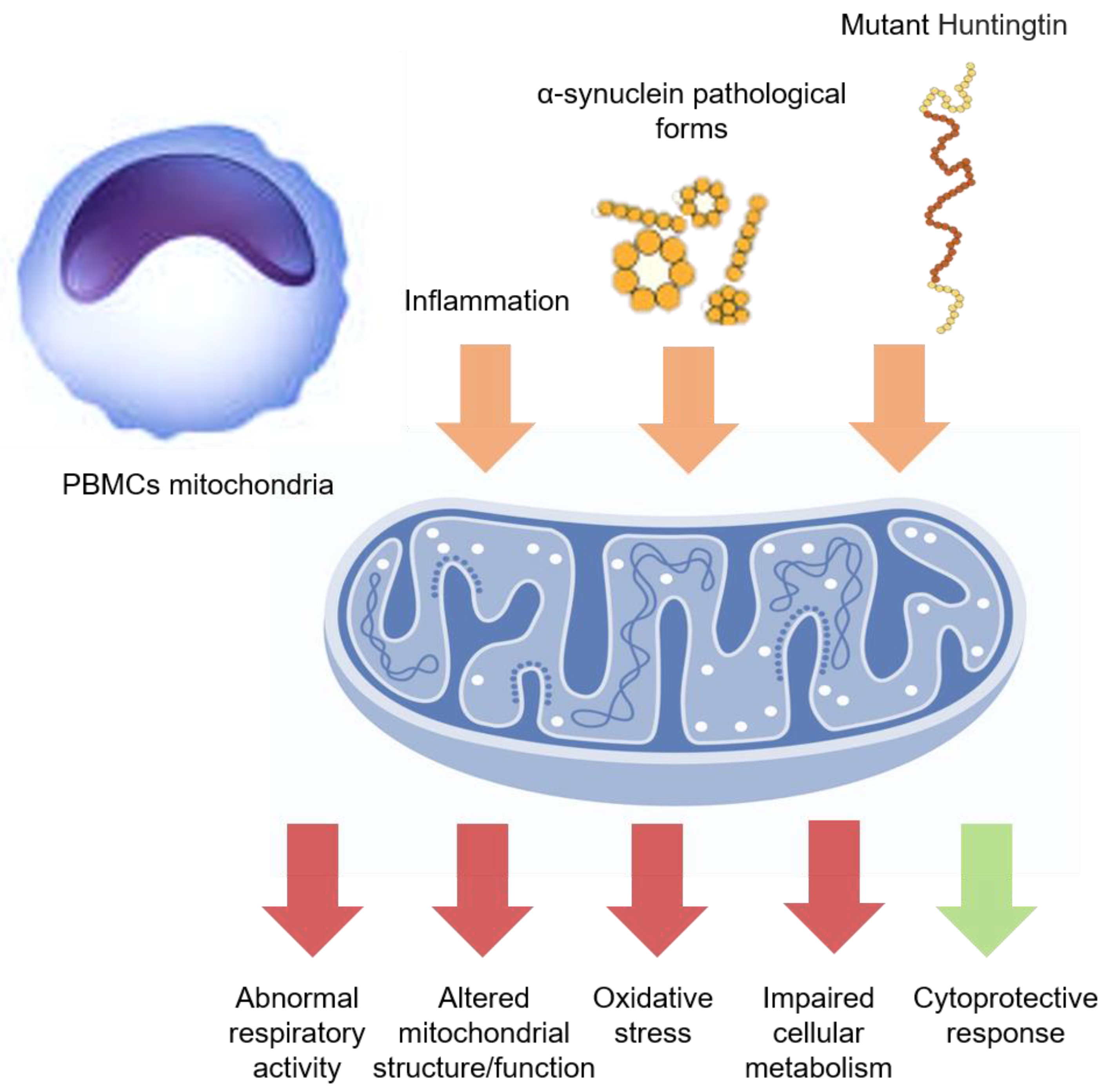
PBMCs mitochondrial function. In PD and HD, the systemic inflammation and the circulating neurodegeneration-related peptides (orange arrows) might affect the PBMCs mitochondrial function, inducing altered function or morphology, abnormal respiratory activity, increased oxidative stress, and impaired cellular metabolism (red arrows). However, some protective mechanisms could be also triggered (green arrow), especially at early disease stages, although later overwhelmed by clinical-pathological progression.
Figure 2.
PBMCs mitochondrial function. In PD and HD, the systemic inflammation and the circulating neurodegeneration-related peptides (orange arrows) might affect the PBMCs mitochondrial function, inducing altered function or morphology, abnormal respiratory activity, increased oxidative stress, and impaired cellular metabolism (red arrows). However, some protective mechanisms could be also triggered (green arrow), especially at early disease stages, although later overwhelmed by clinical-pathological progression.
Discussion
Degenerative diseases commonly entail mitochondrial impairment. The mitochondria represent the major source of bioenergy for the cell and as a consequence, their putative failure leads to a lack of energy. Daily energy metabolism for both humans and organisms is the sum of the daily energy expenditure and daily energy intake. Daily Energy Expenditure may be divided into different categories: 1) energy spent on basal metabolism; 2) energy by thermic increment following food intake; 3) energy spent on thermoregulation; 4) energy spent during daily activities [240]. In particular, energy metabolism is the process of generating energy (ATP) from nutrients and mitochondria represent the main powerhouse of the cell. To produce energy in the form of ATP and GTP, all the cells especially the neurons, consume glucose, amino acids, and fatty acids [240]. These nutrients are processed and transferred into the tricarboxylic acid (TCA) cycle and electrons are stored in their reducing equivalents, NADH and FADH2 through iterative oxidations. NADH and FADH2 serve as carrier molecules that transport electrons into the electron transport chain (ETC), while protons flow down to generate ATP [241]. The capacity of the mitochondria to supply energy in form of ATP when required, could be referred to as bioenergetics efficiency, which may change due to the effect of environmental stress, aging, or pathologies. For example, during aging, the mitochondria undergo alterations in their capacity to produce ATP [241,242]. This is due to the release of a great number of reactive oxygen species (ROS). ROS are able to increase spontaneous DNA mutations and can start the processes that lead to cancer [242,243].
Mitochondrial biogenesis is a process that continuously balances the degradation of dysfunctional mitochondria inside the cells. The disruption of this homeostasis, which may be guaranteed in all functioning cells, represents an additional point that focalizes the loss of energetic function, which may precede cellular loss. This critical balance is called Mitochondrial dynamics. During these processes, mitochondrial fusion and fission regulate the number and size of the mitochondria; while mitochondrial biogenesis increases the mitochondrial mass inside the cells, by producing ATP as a response to greater energy demand. Mitochondrial biogenesis requires a very multifaceted process that starts with the coordination of the nuclear and mitochondrial expression programs. The regulator for the cited process is the protein PGC-1α, which is able to interact with many transcription factors, such as myocyte enhancer factor 2C (MEF2C), nuclear respiratory factors 1 and 2 (NRF1 and NRF2), peroxisome proliferator-activated receptors (PPAR), estrogen-related receptor alpha (ERRα) and many others involved in coordinating mitochondrial biogenesis and oxidative metabolism [244,245,246]. Moreover, NRF1 and NRF2 cooperate to upregulate the transcription of several nuclear-encoded genes that act in the negative feedback effect on the mitochondria production [245,247]. Selective autophagic degradation called mitophagy, recruits the damaged mitochondria into a pre-autophagosome structure via a PINK1/Parkin-dependent process. Indeed, the mitochondria transport essential for the distribution of the mitochondria into the cells and in particular, into the different of neuronal structures, require the synthesis of the mitochondria and its proteins [248,249,250,251].
A consistent number of mitochondria are localized at the presynaptic terminals, to provide ATP for intense synaptic activity [252]. In fact, it was hypothesized that the loss of synaptic plasticity found in PINK1 KO mice, was as a result of the lack of ATP at the presynaptic terminal, thereby confirming the essential role of mitochondria in this area [253].
Previously, we have referred to the role of PINK1 gene in early inherited parkinsonism and these findings are in support of the theory that link mitochondrial dysfunction to PD.
In addition, the mitochondria in aging people show different features [254]; they swell while their numbers dwindleand are unable to replace themselves as quickly in their dysfunctional state [255]. Parkinson’s disease is also an aging disease, in the idiopathic form and this suggests a link with the common non-genetic form of PD. Therefore, failure in the mitochondria is the starting phase of the all neurodegenerative process [256].
It was demonstrated in both PD and HD that the presence of environmental stress can increase the production of oxidative species and can also modify the structure of proteins and DNA inside the mitochondria and its nuclei [257]. Usually, HD is considered a hyperkinetic disorder despite the presence of hypokinetic features in the motor symptoms [251]. Conversely, PD is considered as a hypokinetic disorder, in which the resultant speech motion disease may be classified as hyperkinetic or hypokinetic dysarthria [258,259]. Both are defined as neurodegenerative disorders causing the gradual loss of neurons, and we can explain why. Concerning HD, the mutant HTT protein was shown to be able to directly impair the ability of PGC-1α to activate target genes related to mitochondrial biogenesis and normal mitochondrial function. The amplification of mHTT proteins enhances mitochondrial depolarization and mitochondrial swelling, which is calcium-dependent. In addition, it has been demonstrated that mutant HTT’s ability to combine specifically with the beta-tubulin subunit can adversely affect mitochondrial transport. Moreover, both disorders result in a deficit at the level of the basal ganglia, with peculiar references to striatal [260] and hippocampal areas [261].
In rodent models of HD, only the progression of the disease conduces to neuronal loss. Cell dysfunctions may generate symptoms and in particular, abnormal synaptic statements due to a lack of ATP [262]. In contrast, in human patients, clinical symptoms occur as a combination of functional and structural changes, that may result in a reduction in lactate levels in the brain area, as a consequence of an altered mitochondrial metabolism [251].
In PD models, dysfunction in striatal plasticity are described in details [115] and are strictly connected to energy failure [112,115].
These areas are linked to movement and cognitive ability. For this reason, it is not a surprise that both diseases can affect cognitive or thinking abilities and motor functions. Studies on rodent models of PD show the impairment in the bioenergetics pathway and a block of function in mitochondrial complexes I and III. Also in genetics, phenotypic models were induced by toxin exposure, in the early stages of the disease as well as in the developmental stage [86,102,104,105,169,239]. On the other hand, studies on animal models of HD show an evident alteration in mitochondrial complexes II and III and describe a reduction of bioenergetics efficiency that was described essentially in the early pathogenic mechanism [263].
Interestingly, this evidence has been confirmed by translational studies in man, in which the association between mitochondrial deficit and biomarkers of PD and HD was shown [253]. Actually, there is still no cure for both pathologies and neither do drugs that hinder the development and gravity of the diseases exist. Consequently, the knowledge of pathogenetic mechanisms and the similarities between these diseases can help us while reviewing the traditional approach, in order to develop novel approaches. In fact, new pharmacological approaches focused on mitochondria target agents could be used in the future to cure degenerative and in particular, neurodegenerative pathologies.
Future perspectives
Numerous evidences show the role of mitochondria involvement in neurodegenerative diseases. Approximately 1209 papers described the mitochondria involvement in non-neurological degenerative processes (cancer, aging, other pathologies) (Figure 3 A, B). Among the published works on PD, 53% (of 5185 papers) talked about mitochondria involvement in the pathogenesis of the disease, while only 5% focused on the bioenergetic role of the mitochondria and the regulatory and protective role of the PINK1 gene in the mitochondria machinery (Figure 3C). In HD, on the other hand, 79% (of 1031 works) indicated the involvement of the mitochondria in the progression of symptoms; and only 1% highlighted the protective role of the PINK1 gene in the mitochondria machinery (Figure 3D).
Conversely, many questions remain unresolved, especially about how mitochondrial failures progress and the loss of energy efficiency triggered by environmental events. However, it is clear that in these two illnesses, the role of mitochondria in providing the energetic functions of the neuron is still lacking [4].
Therefore, in-depth studies in this field could provide more insight and knowledge about movement disorders and could offer new therapeutic solutions in pathologies, such as PD and HD, in which only symptomatic cures currently exist.
Author Contributions
Conceptualization: A.T, M.M, T.S, and G.M, bibliography search & collecting: A.T, M.M, T.S, and G.M, resources: P.B; supervision: P.B; visualization: G.P; writing—original draft: A.T, M.M, T.S and G.M; writing—review and editing: A.T, M.M, T.S, G.P and P.B; table and figure A.T, M.M, T.S, and G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work is partially supported by Italian Ministry of Health “Ricerca Finalizzata” grants RF-2019-12370182 to P.B. The funding sources had no involvement in the design or writing of the report and in the decision to submit the article for publication.
Data Availability Statement
The data presented in this study are available in Pub-Med library.
Acknowledgments
All the authors wish to thank Massimo Tolu for his excellent technical assistance. We thank Dr Diepriye Charls-Davies for skilled editorial assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the revision, in the collection, interpretation of data, or in the writing of the manuscript.
Abbreviations
| A53T | α-synuclein with a PD-associated mutation |
| ADP | Adenosine diphosphate |
| AMPAR | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor |
| ATP | Adenosine triphosphate |
| ATP5A | ATP synthase alpha-subunit gene |
| ATP5A1 | encodes for a subunit of ATP synthase (complex V) |
| BAC | bacterial artificial chromosome |
| BCL-2 | B-cell lymphoma 2 |
| Ca | calcium |
| COX4I1 | Cytochrome C Oxidase Subunit 4I1 |
| CSF | cerebrospinal fluid |
| DJ-1 | protein of the peptidase C56 family |
| DNA | deoxyribonucleic acid |
| Drp1 | dynamin-related protein 1 |
| ER | endoplasmic reticulum |
| ETC | electron transport chain |
| ETC | electron transport chain |
| FADH2 | flavin adenine dinucleotides |
| Glu | glutamate |
| GR | glutathione reductase |
| GRX | glutaredoxin |
| GSH | glutathione reduced |
| GSSG | Glutathione oxidized |
| GTP | Guanosine-5’-triphosphate |
| HD | Huntington’s disease |
| Homer1 | Homer Scaffold Protein 1 |
| HTT | huntingtin |
| IL-18 | interleukin-18 |
| IL-1β | interleukin-1β |
| IMM | inner mitochondrial membrane |
| KO | knockout |
| LC3 | Microtubule-associated protein 1A/1B-light chain 3 |
| LTD | long-term depression |
| LTP | long-term potentiation |
| MAO | Monoamine oxidases |
| MCU | mitochondrial calcium uniporter |
| Mfn1 | Mitofusin1 |
| Mfn2 | Mitofusin2 |
| MICU3 | mitochondrial calcium uptake family member3 |
| MPTP | 1-metil-4-fenil-1,2,3,6-tetraidropiridina |
| NADH | nicotinamide adenine dinucleotides |
| NCLX | mitochondrial Na+/Ca2+ exchanger |
| NDUFS3 | NADH Ubiquinone Oxidoreductase Core Subunit S3 |
| NLR | Nod-like Receptor |
| NLRP3 | nucleotide-binding domain, leucine-rich-repeat containing family, pyrin domain-containing 3 |
| NLRX1 | NLR Family Member X1 |
| NO | Nitric oxide |
| 3-NPA | 3-nitropropionic acid |
| Nrf2 | nuclear factor erythroid 2–related factor 2 |
| OMM | outer mitochondrial membrane |
| OXPHOS | mitochondrial oxidative phosphorylation |
| PBMCs | Peripheral blood mononucleate cells |
| PD | Parkinson’s disease |
| PGC-1α | eroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PINK1 | PTEN-induced kinase 1 |
| PRKAG2 | protein kinase AMP-activated non-catalytic subunit gamma 2 |
| Rab5 | Ras-related protein5 |
| ROS | generation of free radical species |
| SDHB | Succinate Dehydrogenase Complex, Subunit B |
| SNCA | alpha-synuclein gene |
| SNpc | Substantia nigra pars compacta |
| SPN | medium spine neuron |
| TCA | tricarboxylic acid |
| Trx | Thioredoxin |
| TTR | Transthyretin |
| VDAC | Voltage-dependent anion channel |
| VDAC3 | oltage-dependent anion-selective channel protein 3 |
| YAC | yeast artificial chromosome |
| ΔΨm | Mitochondrial membrane potential |
References
- Osellame, L.D.; Blacker, T.S.; Duchen, M.R. Cellular and molecular mechanisms of mitochondrial function. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 711–723. [CrossRef]
- Pizzorno J Mitochondria-Fundamental to Life and Health.
- Bruce Alberts Molecular Biology of the Cell; ISBN ISBN-10: 0-8153-4072-9.
- Wang, Y.; Xu, E.; Musich, P.R.; Lin, F. Mitochondrial dysfunction in neurodegenerative diseases and the potential countermeasure. CNS Neurosci. Ther. 2019, 25, 816–824. [CrossRef]
- Golpich, M.; Amini, E.; Mohamed, Z.; Azman Ali, R.; Mohamed Ibrahim, N.; Ahmadiani, A. Mitochondrial Dysfunction and Biogenesis in Neurodegenerative diseases: Pathogenesis and Treatment. CNS Neurosci. Ther. 2017, 23, 5–22. [CrossRef]
- Jadiya, P.; Garbincius, J.F.; Elrod, J.W. Reappraisal of metabolic dysfunction in neurodegeneration: Focus on mitochondrial function and calcium signaling. Acta Neuropathol. Commun. 2021, 9, 124. [CrossRef]
- Zhu, X.-H.; Qiao, H.; Du, F.; Xiong, Q.; Liu, X.; Zhang, X.; Ugurbil, K.; Chen, W. Quantitative imaging of energy expenditure in human brain. NeuroImage 2012, 60, 2107–2117. [CrossRef]
- Vergara, R.C.; Jaramillo-Riveri, S.; Luarte, A.; Moënne-Loccoz, C.; Fuentes, R.; Couve, A.; Maldonado, P.E. The Energy Homeostasis Principle: Neuronal Energy Regulation Drives Local Network Dynamics Generating Behavior. Front. Comput. Neurosci. 2019, 13. [CrossRef]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [CrossRef]
- Akbar, M.; Essa, M.M.; Daradkeh, G.; Abdelmegeed, M.A.; Choi, Y.; Mahmood, L.; Song, B.-J. Mitochondrial dysfunction and cell death in neurodegenerative diseases through nitroxidative stress. Brain Res. 2016, 1637, 34–55. [CrossRef]
- Jellinger, K.A. Basic mechanisms of neurodegeneration: a critical update. J. Cell. Mol. Med. 2010. [CrossRef]
- Murali Mahadevan, H.; Hashemiaghdam, A.; Ashrafi, G.; Harbauer, A.B. Mitochondria in Neuronal Health: From Energy Metabolism to Parkinson’s Disease. Adv. Biol. 2021, 5, 2100663. [CrossRef]
- Zhao, R.; Jiang, S.; Zhang, L.; Yu, Z. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019. [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [CrossRef]
- Trigo, D.; Avelar, C.; Fernandes, M.; Sá, J.; Cruz e Silva, O. Mitochondria, energy, and metabolism in neuronal health and disease. FEBS Lett. 2022, 596, 1095–1110. [CrossRef]
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More Than Just a Powerhouse. Curr. Biol. 2006, 16, R551–R560. [CrossRef]
- Giorgi, C.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Marchi, S.; Missiroli, S.; Patergnani, S.; Poletti, F.; Rimessi, A.; et al. Mitochondrial calcium homeostasis as potential target for mitochondrial medicine. Mitochondrion 2012, 12, 77–85. [CrossRef]
- Pathak, T.; Trebak, M. Mitochondrial Ca2+ signaling. Pharmacol. Ther. 2018, 192, 112–123. [CrossRef]
- Shoshan-Barmatz, V.; Ben-Hail, D. VDAC, a multi-functional mitochondrial protein as a pharmacological target. Mitochondrion 2012, 12, 24–34. [CrossRef]
- Rostovtseva, T.K.; Bezrukov, S.M.; Hoogerheide, D.P. Regulation of Mitochondrial Respiration by VDAC Is Enhanced by Membrane-Bound Inhibitors with Disordered Polyanionic C-Terminal Domains. Int. J. Mol. Sci. 2021, 22, 7358. [CrossRef]
- Jung, H.; Kim, S.Y.; Canbakis Cecen, F.S.; Cho, Y.; Kwon, S.-K. Dysfunction of Mitochondrial Ca2+ Regulatory Machineries in Brain Aging and Neurodegenerative Diseases. Front. Cell Dev. Biol. 2020, 8, 599792. [CrossRef]
- Sparagna, G.C.; Gunter, K.K.; Gunter, T.E. A System for Producing and Monitoring in Vitro Calcium Pulses Similar to Those Observed in Vivo. Anal. Biochem. 1994, 219, 96–103. [CrossRef]
- Sparagna, G.C.; Gunter, K.K.; Sheu, S.-S.; Gunter, T.E. Mitochondrial Calcium Uptake from Physiological-type Pulses of Calcium: A DESCRIPTION OF THE RAPID UPTAKE MODE (*). J. Biol. Chem. 1995, 270, 27510–27515. [CrossRef]
- Grienberger, C.; Konnerth, A. Imaging Calcium in Neurons. Neuron 2012, 73, 862–885. [CrossRef]
- Petersen, O.H. Calcium signal compartmentalization. Biol. Res. 2002, 35. [CrossRef]
- Laude, A.J.; Simpson, A.W.M. Compartmentalized signalling: Ca2+ compartments, microdomains and the many facets of Ca2+ signalling: Ca2+ microdomains. FEBS J. 2009, 276, 1800–1816. [CrossRef]
- Panda, S.; Behera, S.; Alam, M.F.; Syed, G.H. Endoplasmic reticulum & mitochondrial calcium homeostasis: The interplay with viruses. Mitochondrion 2021, 58, 227–242. [CrossRef]
- Hirabayashi, Y.; Kwon, S.-K.; Paek, H.; Pernice, W.M.; Paul, M.A.; Lee, J.; Erfani, P.; Raczkowski, A.; Petrey, D.S.; Pon, L.A.; et al. ER-mitochondria tethering by PDZD8 regulates Ca2+ dynamics in mammalian neurons. Science 2017, 358, 623–630. [CrossRef]
- Ashrafi, G.; de Juan-Sanz, J.; Farrell, R.J.; Ryan, T.A. Molecular Tuning of the Axonal Mitochondrial Ca2+ Uniporter Ensures Metabolic Flexibility of Neurotransmission. Neuron 2020, 105, 678-687.e5. [CrossRef]
- Ryan, K.C.; Ashkavand, Z.; Norman, K.R. The Role of Mitochondrial Calcium Homeostasis in Alzheimer’s and Related Diseases. Int. J. Mol. Sci. 2020, 21, 9153. [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [CrossRef]
- Kuznetsov, A.V.; Margreiter, R.; Ausserlechner, M.J.; Hagenbuchner, J. The Complex Interplay between Mitochondria, ROS and Entire Cellular Metabolism. Antioxidants 2022, 11, 1995. [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Functional role of mitochondrial reactive oxygen species in physiology. Free Radic. Biol. Med. 2016, 100, 81–85. [CrossRef]
- Angelova, P.R. Sources and triggers of oxidative damage in neurodegeneration. Free Radic. Biol. Med. 2021, 173, 52–63. [CrossRef]
- Tait, S.W.G.; Green, D.R. Mitochondria and cell signalling. J. Cell Sci. 2012, 125, 807–815. [CrossRef]
- Hordijk, P.L. Regulation of NADPH Oxidases: The Role of Rac Proteins. Circ. Res. 2006, 98, 453–462. [CrossRef]
- Angajala, A.; Lim, S.; Phillips, J.B.; Kim, J.-H.; Yates, C.; You, Z.; Tan, M. Diverse Roles of Mitochondria in Immune Responses: Novel Insights Into Immuno-Metabolism. Front. Immunol. 2018, 9, 1605. [CrossRef]
- Pearce, E.L.; Pearce, E.J. Metabolic Pathways in Immune Cell Activation and Quiescence. Immunity 2013, 38, 633–643. [CrossRef]
- Breda, C.N. de S.; Davanzo, G.G.; Basso, P.J.; Saraiva Câmara, N.O.; Moraes-Vieira, P.M.M. Mitochondria as central hub of the immune system. Redox Biol. 2019, 26, 101255. [CrossRef]
- Chen, Y.; Zhou, Z.; Min, W. Mitochondria, Oxidative Stress and Innate Immunity. Front. Physiol. 2018, 9, 1487. [CrossRef]
- Chu, X.; Wu, S.; Raju, R. NLRX1 Regulation Following Acute Mitochondrial Injury. Front. Immunol. 2019, 10, 2431. [CrossRef]
- Stokman, G.; Kors, L.; Bakker, P.J.; Rampanelli, E.; Claessen, N.; Teske, G.J.D.; Butter, L.; van Andel, H.; van den Bergh Weerman, M.A.; Larsen, P.W.B.; et al. NLRX1 dampens oxidative stress and apoptosis in tissue injury via control of mitochondrial activity. J. Exp. Med. 2017, 214, 2405–2420. [CrossRef]
- Liu, Q.; Zhang, D.; Hu, D.; Zhou, X.; Zhou, Y. The role of mitochondria in NLRP3 inflammasome activation. Mol. Immunol. 2018, 103, 115–124. [CrossRef]
- Lamkanfi, M.; Kanneganti, T.-D. Nlrp3: An immune sensor of cellular stress and infection. Int. J. Biochem. Cell Biol. 2010, 42, 792–795. [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [CrossRef]
- Behrouzi, A.; Kelley, M.R.; Fehrenbacher, J.C. Oxidative DNA Damage: A Role in Altering Neuronal Function. J. Cell. Signal. 2022, 3, 160–166. [CrossRef]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: when to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V.; Kazantsev, A.G. The role of Nrf2 signaling in counteracting neurodegenerative diseases. FEBS J. 2018, 285, 3576–3590. [CrossRef]
- Thomas, R.L.; Gustafsson, A.B. Mitochondrial autophagy--an essential quality control mechanism for myocardial homeostasis. Circ. J. Off. J. Jpn. Circ. Soc. 2013, 77, 2449–2454. [CrossRef]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [CrossRef]
- Narendra, D.P.; Jin, S.M.; Tanaka, A.; Suen, D.-F.; Gautier, C.A.; Shen, J.; Cookson, M.R.; Youle, R.J. PINK1 Is Selectively Stabilized on Impaired Mitochondria to Activate Parkin. PLoS Biol. 2010, 8, e1000298. [CrossRef]
- Elmore, S. Apoptosis: a review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [CrossRef]
- Suen, D.-F.; Norris, K.L.; Youle, R.J. Mitochondrial dynamics and apoptosis. Genes Dev. 2008, 22, 1577–1590. [CrossRef]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis*. Annu. Rev. Genet. 2009, 43, 95–118. [CrossRef]
- Sheng, Z.-H. The Interplay of Axonal Energy Homeostasis and Mitochondrial Trafficking and Anchoring. Trends Cell Biol. 2017, 27, 403–416. [CrossRef]
- Seager, R.; Lee, L.; Henley, J.M.; Wilkinson, K.A. Mechanisms and roles of mitochondrial localisation and dynamics in neuronal function. Neuronal Signal. 2020, 4, NS20200008. [CrossRef]
- Vos, M.; Lauwers, E.; Verstreken, P. Synaptic mitochondria in synaptic transmission and organization of vesicle pools in health and disease. Front. Synaptic Neurosci. 2010, 2, 139. [CrossRef]
- Chanaday, N.L.; Cousin, M.A.; Milosevic, I.; Watanabe, S.; Morgan, J.R. The Synaptic Vesicle Cycle Revisited: New Insights into the Modes and Mechanisms. J. Neurosci. 2019, 39, 8209–8216. [CrossRef]
- Datta, S.; Jaiswal, M. Mitochondrial calcium at the synapse. Mitochondrion 2021, 59, 135–153. [CrossRef]
- Orth, M.; Schapira, A.H. Mitochondria and degenerative disorders. Am. J. Med. Genet. 2001, 106, 27–36. [CrossRef]
- Liesa, M.; Shirihai, O.S. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. 2013, 17, 491–506. [CrossRef]
- Zhang, P.N.; Zhou, M.Q.; Guo, J.; Zheng, H.J.; Tang, J.; Zhang, C.; Liu, Y.N.; Liu, W.J.; Wang, Y.X. Mitochondrial Dysfunction and Diabetic Nephropathy: Nontraditional Therapeutic Opportunities. J. Diabetes Res. 2021, 2021, 1010268. [CrossRef]
- Molina, A.J.A.; Wikstrom, J.D.; Stiles, L.; Las, G.; Mohamed, H.; Elorza, A.; Walzer, G.; Twig, G.; Katz, S.; Corkey, B.E.; et al. Mitochondrial networking protects beta-cells from nutrient-induced apoptosis. Diabetes 2009, 58, 2303–2315. [CrossRef]
- Gomes, L.C.; Di Benedetto, G.; Scorrano, L. During autophagy mitochondria elongate, are spared from degradation and sustain cell viability. Nat. Cell Biol. 2011, 13, 589–598. [CrossRef]
- Lewin, R. Trail of ironies to Parkinson’s disease. Science 1984, 224, 1083–1085. [CrossRef]
- Hauser, D.N.; Hastings, T.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease and monogenic parkinsonism. Neurobiol. Dis. 2013, 51, 35–42. [CrossRef]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [CrossRef]
- Park, J.-S.; Davis, R.L.; Sue, C.M. Mitochondrial Dysfunction in Parkinson’s Disease: New Mechanistic Insights and Therapeutic Perspectives. Curr. Neurol. Neurosci. Rep. 2018, 18, 21. [CrossRef]
- Magalhães, J.D.; Cardoso, S.M. Mitochondrial signaling on innate immunity activation in Parkinson disease. Curr. Opin. Neurobiol. 2023, 78, 102664. [CrossRef]
- Sagan, L. On the origin of mitosing cells. J. Theor. Biol. 1967, 14, 225-IN6. [CrossRef]
- Rossi, A.; Pizzo, P. Mitochondrial bioenergetics and neurodegeneration: a paso doble. Neural Regen. Res. 2021, 16, 686–687. [CrossRef]
- Zhang, C.; Chen, S.; Li, X.; Xu, Q.; Lin, Y.; Lin, F.; Yuan, M.; Zi, Y.; Cai, J. Progress in Parkinson’s disease animal models of genetic defects: Characteristics and application. Biomed. Pharmacother. Biomedecine Pharmacother. 2022, 155, 113768. [CrossRef]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.K.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [CrossRef]
- Arena, G.; Valente, E.M. PINK1 in the limelight: multiple functions of an eclectic protein in human health and disease. J. Pathol. 2017, 241, 251–263. [CrossRef]
- Silvestri, L.; Caputo, V.; Bellacchio, E.; Atorino, L.; Dallapiccola, B.; Valente, E.M.; Casari, G. Mitochondrial import and enzymatic activity of PINK1 mutants associated to recessive parkinsonism. Hum. Mol. Genet. 2005, 14, 3477–3492. [CrossRef]
- Unoki, M.; Nakamura, Y. Growth-suppressive effects of BPOZ and EGR2, two genes involved in the PTEN signaling pathway. Oncogene 2001, 20, 4457–4465. [CrossRef]
- Garber, K. Parkinson’s disease and cancer: the unexplored connection. J. Natl. Cancer Inst. 2010, 102, 371–374. [CrossRef]
- Masgras, I.; Laquatra, C.; Cannino, G.; Serapian, S.A.; Colombo, G.; Rasola, A. The molecular chaperone TRAP1 in cancer: From the basics of biology to pharmacological targeting. Semin. Cancer Biol. 2021, 76, 45–53. [CrossRef]
- Exner, N.; Treske, B.; Paquet, D.; Holmström, K.; Schiesling, C.; Gispert, S.; Carballo-Carbajal, I.; Berg, D.; Hoepken, H.-H.; Gasser, T.; et al. Loss-of-function of human PINK1 results in mitochondrial pathology and can be rescued by parkin. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 12413–12418. [CrossRef]
- Kitada, T.; Pisani, A.; Porter, D.R.; Yamaguchi, H.; Tscherter, A.; Martella, G.; Bonsi, P.; Zhang, C.; Pothos, E.N.; Shen, J. Impaired dopamine release and synaptic plasticity in the striatum of PINK1 -deficient mice. Proc. Natl. Acad. Sci. 2007, 104, 11441–11446. [CrossRef]
- Martella, G.; Madeo, G.; Maltese, M.; Vanni, V.; Puglisi, F.; Ferraro, E.; Schirinzi, T.; Valente, E.M.; Bonanni, L.; Shen, J.; et al. Exposure to low-dose rotenone precipitates synaptic plasticity alterations in PINK1 heterozygous knockout mice. Neurobiol. Dis. 2016, 91, 21–36. [CrossRef]
- Imbriani, P.; D’Angelo, V.; Platania, P.; Di Lazzaro, G.; Scalise, S.; Salimei, C.; El Atiallah, I.; Colona, V.L.; Mercuri, N.B.; Bonsi, P.; et al. Ischemic injury precipitates neuronal vulnerability in Parkinson’s disease: Insights from PINK1 mouse model study and clinical retrospective data. Parkinsonism Relat. Disord. 2020, 74, 57–63. [CrossRef]
- Imbriani, P.; Tassone, A.; Meringolo, M.; Ponterio, G.; Madeo, G.; Pisani, A.; Bonsi, P.; Martella, G. Loss of Non-Apoptotic Role of Caspase-3 in the PINK1 Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 3407. [CrossRef]
- Brunelli, F.; Valente, E.M.; Arena, G. Mechanisms of neurodegeneration in Parkinson’s disease: keep neurons in the PINK1. Mech. Ageing Dev. 2020, 189, 111277. [CrossRef]
- Zhi, L.; Qin, Q.; Muqeem, T.; Seifert, E.L.; Liu, W.; Zheng, S.; Li, C.; Zhang, H. Loss of PINK1 causes age-dependent decrease of dopamine release and mitochondrial dysfunction. Neurobiol. Aging 2019, 75, 1–10. [CrossRef]
- Imbriani, P.; Tassone, A.; Meringolo, M.; Ponterio, G.; Madeo, G.; Pisani, A.; Bonsi, P.; Martella, G. Loss of Non-Apoptotic Role of Caspase-3 in the PINK1 Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 3407. [CrossRef]
- Onyango, I.G.; Bennett, J.P.; Stokin, G.B. Regulation of neuronal bioenergetics as a therapeutic strategy in neurodegenerative diseases. Neural Regen. Res. 2021, 16, 1467–1482. [CrossRef]
- Anderson, K.E.; Marshall, F.J. Behavioral symptoms associated with Huntington’s disease. Adv. Neurol. 2005, 96, 197–208.
- Caron, N.S.; Wright, G.E.; Hayden, M.R. Huntington Disease. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; University of Washington, Seattle: Seattle (WA), 1993.
- Damiano, M.; Galvan, L.; Déglon, N.; Brouillet, E. Mitochondria in Huntington’s disease. Biochim. Biophys. Acta 2010, 1802, 52–61. [CrossRef]
- Rehman, M.U.; Sehar, N.; Dar, N.J.; Khan, A.; Arafah, A.; Rashid, S.; Rashid, S.M.; Ganaie, M.A. Mitochondrial dysfunctions, oxidative stress and neuroinflammation as therapeutic targets for neurodegenerative diseases: An update on current advances and impediments. Neurosci. Biobehav. Rev. 2023, 144, 104961. [CrossRef]
- Jurcau, A.; Jurcau, C.M. Mitochondria in Huntington’s disease: implications in pathogenesis and mitochondrial-targeted therapeutic strategies. Neural Regen. Res. 2023, 18, 1472–1477. [CrossRef]
- Kawsar, Md.; Taz, T.A.; Paul, B.K.; Ahmed, K.; Habib, Md.A.; Bhuyian, T. Identification of vital regulatory genes with network pathways among Huntington’s, Parkinson’s, and Alzheimer’s diseases. Netw. Model. Anal. Health Inform. Bioinforma. 2020, 9, 50. [CrossRef]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: from mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [CrossRef]
- Quijano, C.; Cao, L.; Fergusson, M.M.; Romero, H.; Liu, J.; Gutkind, S.; Rovira, I.I.; Mohney, R.P.; Karoly, E.D.; Finkel, T. Oncogene-induced senescence results in marked metabolic and bioenergetic alterations. Cell Cycle Georget. Tex 2012, 11, 1383–1392. [CrossRef]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell 2016, 61, 654–666. [CrossRef]
- Moro, L. Mitochondrial Dysfunction in Aging and Cancer. J. Clin. Med. 2019, 8, 1983. [CrossRef]
- Boland, M.L.; Chourasia, A.H.; Macleod, K.F. Mitochondrial dysfunction in cancer. Front. Oncol. 2013, 3, 292. [CrossRef]
- Wang, X.-L.; Feng, S.-T.; Wang, Y.-T.; Yuan, Y.-H.; Li, Z.-P.; Chen, N.-H.; Wang, Z.-Z.; Zhang, Y. Mitophagy, a Form of Selective Autophagy, Plays an Essential Role in Mitochondrial Dynamics of Parkinson’s Disease. Cell. Mol. Neurobiol. 2022, 42, 1321–1339. [CrossRef]
- Ryan, B.J.; Hoek, S.; Fon, E.A.; Wade-Martins, R. Mitochondrial dysfunction and mitophagy in Parkinson’s: from familial to sporadic disease. Trends Biochem. Sci. 2015, 40, 200–210. [CrossRef]
- Büeler, H. Impaired mitochondrial dynamics and function in the pathogenesis of Parkinson’s disease. Exp. Neurol. 2009, 218, 235–246. [CrossRef]
- Chen, C.; Turnbull, D.M.; Reeve, A.K. Mitochondrial Dysfunction in Parkinson’s Disease-Cause or Consequence? Biology 2019, 8, 38. [CrossRef]
- Parker, W.D.; Boyson, S.J.; Parks, J.K. Abnormalities of the electron transport chain in idiopathic Parkinson’s disease. Ann. Neurol. 1989, 26, 719–723. [CrossRef]
- Thirugnanam, T.; Santhakumar, K. Chemically induced models of Parkinson’s disease. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2022, 252, 109213. [CrossRef]
- Bové, J.; Perier, C. Neurotoxin-based models of Parkinson’s disease. Neuroscience 2012, 211, 51–76. [CrossRef]
- Exner, N.; Lutz, A.K.; Haass, C.; Winklhofer, K.F. Mitochondrial dysfunction in Parkinson’s disease: molecular mechanisms and pathophysiological consequences. EMBO J. 2012, 31, 3038–3062. [CrossRef]
- Meredith, G.E.; Rademacher, D.J. MPTP mouse models of Parkinson’s disease: an update. J. Park. Dis. 2011, 1, 19–33. [CrossRef]
- Zeng, X.-S.; Geng, W.-S.; Jia, J.-J. Neurotoxin-Induced Animal Models of Parkinson Disease: Pathogenic Mechanism and Assessment. ASN Neuro 2018, 10, 1759091418777438. [CrossRef]
- Imbriani, P.; Schirinzi, T.; Meringolo, M.; Mercuri, N.B.; Pisani, A. Centrality of Early Synaptopathy in Parkinson’s Disease. Front. Neurol. 2018, 9, 103. [CrossRef]
- Imbriani, P.; Sciamanna, G.; Santoro, M.; Schirinzi, T.; Pisani, A. Promising rodent models in Parkinson’s disease. Parkinsonism Relat. Disord. 2018, 46 Suppl 1, S10–S14. [CrossRef]
- Innos, J.; Hickey, M.A. Using Rotenone to Model Parkinson’s Disease in Mice: A Review of the Role of Pharmacokinetics. Chem. Res. Toxicol. 2021, 34, 1223–1239. [CrossRef]
- Imbriani, P.; Martella, G.; Bonsi, P.; Pisani, A. Oxidative stress and synaptic dysfunction in rodent models of Parkinson’s disease. Neurobiol. Dis. 2022, 173, 105851. [CrossRef]
- McLean, P.J.; Ribich, S.; Hyman, B.T. Subcellular localization of alpha-synuclein in primary neuronal cultures: effect of missense mutations. J. Neural Transm. Suppl. 2000, 53–63. [CrossRef]
- Zhang, L.; Zhang, C.; Zhu, Y.; Cai, Q.; Chan, P.; Uéda, K.; Yu, S.; Yang, H. Semi-quantitative analysis of alpha-synuclein in subcellular pools of rat brain neurons: an immunogold electron microscopic study using a C-terminal specific monoclonal antibody. Brain Res. 2008, 1244, 40–52. [CrossRef]
- Cole, N.B.; Dieuliis, D.; Leo, P.; Mitchell, D.C.; Nussbaum, R.L. Mitochondrial translocation of alpha-synuclein is promoted by intracellular acidification. Exp. Cell Res. 2008, 314, 2076–2089. [CrossRef]
- Polymeropoulos, M.H.; Higgins, J.J.; Golbe, L.I.; Johnson, W.G.; Ide, S.E.; Di Iorio, G.; Sanges, G.; Stenroos, E.S.; Pho, L.T.; Schaffer, A.A.; et al. Mapping of a gene for Parkinson’s disease to chromosome 4q21-q23. Science 1996, 274, 1197–1199. [CrossRef]
- Koprich, J.B.; Kalia, L.V.; Brotchie, J.M. Animal models of α-synucleinopathy for Parkinson disease drug development. Nat. Rev. Neurosci. 2017, 18, 515–529. [CrossRef]
- Ingelsson, M. Alpha-Synuclein Oligomers-Neurotoxic Molecules in Parkinson’s Disease and Other Lewy Body Disorders. Front. Neurosci. 2016, 10, 408. [CrossRef]
- Martin, L.J.; Pan, Y.; Price, A.C.; Sterling, W.; Copeland, N.G.; Jenkins, N.A.; Price, D.L.; Lee, M.K. Parkinson’s disease alpha-synuclein transgenic mice develop neuronal mitochondrial degeneration and cell death. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 41–50. [CrossRef]
- Portz, P.; Lee, M.K. Changes in Drp1 Function and Mitochondrial Morphology Are Associated with the α-Synuclein Pathology in a Transgenic Mouse Model of Parkinson’s Disease. Cells 2021, 10, 885. [CrossRef]
- Xie, W.; Chung, K.K.K. Alpha-synuclein impairs normal dynamics of mitochondria in cell and animal models of Parkinson’s disease. J. Neurochem. 2012, 122, 404–414. [CrossRef]
- Chinta, S.J.; Mallajosyula, J.K.; Rane, A.; Andersen, J.K. Mitochondrial α-synuclein accumulation impairs complex I function in dopaminergic neurons and results in increased mitophagy in vivo. Neurosci. Lett. 2010, 486, 235–239. [CrossRef]
- Merino-Galán, L.; Jimenez-Urbieta, H.; Zamarbide, M.; Rodríguez-Chinchilla, T.; Belloso-Iguerategui, A.; Santamaria, E.; Fernández-Irigoyen, J.; Aiastui, A.; Doudnikoff, E.; Bézard, E.; et al. Striatal synaptic bioenergetic and autophagic decline in premotor experimental parkinsonism. Brain J. Neurol. 2022, 145, 2092–2107. [CrossRef]
- Available online: https://pubmed.ncbi.nlm.nih.gov/28341600/ (accessed on Mar 6, 2023).
- Kurz, A.; Double, K.L.; Lastres-Becker, I.; Tozzi, A.; Tantucci, M.; Bockhart, V.; Bonin, M.; García-Arencibia, M.; Nuber, S.; Schlaudraff, F.; et al. A53T-alpha-synuclein overexpression impairs dopamine signaling and striatal synaptic plasticity in old mice. PloS One 2010, 5, e11464. [CrossRef]
- Durante, V.; de Iure, A.; Loffredo, V.; Vaikath, N.; De Risi, M.; Paciotti, S.; Quiroga-Varela, A.; Chiasserini, D.; Mellone, M.; Mazzocchetti, P.; et al. Alpha-synuclein targets GluN2A NMDA receptor subunit causing striatal synaptic dysfunction and visuospatial memory alteration. Brain J. Neurol. 2019, 142, 1365–1385. [CrossRef]
- Tozzi, A.; de Iure, A.; Bagetta, V.; Tantucci, M.; Durante, V.; Quiroga-Varela, A.; Costa, C.; Di Filippo, M.; Ghiglieri, V.; Latagliata, E.C.; et al. Alpha-Synuclein Produces Early Behavioral Alterations via Striatal Cholinergic Synaptic Dysfunction by Interacting With GluN2D N-Methyl-D-Aspartate Receptor Subunit. Biol. Psychiatry 2016, 79, 402–414. [CrossRef]
- Tozzi, A.; Sciaccaluga, M.; Loffredo, V.; Megaro, A.; Ledonne, A.; Cardinale, A.; Federici, M.; Bellingacci, L.; Paciotti, S.; Ferrari, E.; et al. Dopamine-dependent early synaptic and motor dysfunctions induced by α-synuclein in the nigrostriatal circuit. Brain J. Neurol. 2021, 144, 3477–3491. [CrossRef]
- Calì, T.; Ottolini, D.; Negro, A.; Brini, M. α-Synuclein controls mitochondrial calcium homeostasis by enhancing endoplasmic reticulum-mitochondria interactions. J. Biol. Chem. 2012, 287, 17914–17929. [CrossRef]
- Alves Da Costa, C.; Paitel, E.; Vincent, B.; Checler, F. Alpha-synuclein lowers p53-dependent apoptotic response of neuronal cells. Abolishment by 6-hydroxydopamine and implication for Parkinson’s disease. J. Biol. Chem. 2002, 277, 50980–50984. [CrossRef]
- Chan, S.L.; Mattson, M.P. Caspase and calpain substrates: roles in synaptic plasticity and cell death. J. Neurosci. Res. 1999, 58, 167–190. [CrossRef]
- D’Amelio, M.; Cavallucci, V.; Cecconi, F. Neuronal caspase-3 signaling: not only cell death. Cell Death Differ. 2010, 17, 1104–1114. [CrossRef]
- Snigdha, S.; Smith, E.D.; Prieto, G.A.; Cotman, C.W. Caspase-3 activation as a bifurcation point between plasticity and cell death. Neurosci. Bull. 2012, 28, 14–24. [CrossRef]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.J.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [CrossRef]
- Thomas, K.J.; McCoy, M.K.; Blackinton, J.; Beilina, A.; van der Brug, M.; Sandebring, A.; Miller, D.; Maric, D.; Cedazo-Minguez, A.; Cookson, M.R. DJ-1 acts in parallel to the PINK1/parkin pathway to control mitochondrial function and autophagy. Hum. Mol. Genet. 2011, 20, 40–50. [CrossRef]
- Trancikova, A.; Tsika, E.; Moore, D.J. Mitochondrial dysfunction in genetic animal models of Parkinson’s disease. Antioxid. Redox Signal. 2012, 16, 896–919. [CrossRef]
- Dzamko, N.; Zhou, J.; Huang, Y.; Halliday, G.M. Parkinson’s disease-implicated kinases in the brain; insights into disease pathogenesis. Front. Mol. Neurosci. 2014, 7, 57. [CrossRef]
- Du, F.; Yu, Q.; Yan, S.; Hu, G.; Lue, L.-F.; Walker, D.G.; Wu, L.; Yan, S.F.; Tieu, K.; Yan, S.S. PINK1 signalling rescues amyloid pathology and mitochondrial dysfunction in Alzheimer’s disease. Brain J. Neurol. 2017, 140, 3233–3251. [CrossRef]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314. [CrossRef]
- Lücking, C.B.; Dürr, A.; Bonifati, V.; Vaughan, J.; De Michele, G.; Gasser, T.; Harhangi, B.S.; Meco, G.; Denèfle, P.; Wood, N.W.; et al. Association between early-onset Parkinson’s disease and mutations in the parkin gene. N. Engl. J. Med. 2000, 342, 1560–1567. [CrossRef]
- Moore, D.J. Parkin: a multifaceted ubiquitin ligase. Biochem. Soc. Trans. 2006, 34, 749–753. [CrossRef]
- Xiong, H.; Wang, D.; Chen, L.; Choo, Y.S.; Ma, H.; Tang, C.; Xia, K.; Jiang, W.; Ronai, Z.; Zhuang, X.; et al. Parkin, PINK1, and DJ-1 form a ubiquitin E3 ligase complex promoting unfolded protein degradation. J. Clin. Invest. 2009, 119, 650–660. [CrossRef]
- Periquet, M.; Corti, O.; Jacquier, S.; Brice, A. Proteomic analysis of parkin knockout mice: alterations in energy metabolism, protein handling and synaptic function. J. Neurochem. 2005, 95, 1259–1276. [CrossRef]
- Darios, F. Parkin prevents mitochondrial swelling and cytochrome c release in mitochondria-dependent cell death. Hum. Mol. Genet. 2003, 12, 517–526. [CrossRef]
- Shin, J.-H.; Ko, H.S.; Kang, H.; Lee, Y.; Lee, Y.-I.; Pletinkova, O.; Troconso, J.C.; Dawson, V.L.; Dawson, T.M. PARIS (ZNF746) repression of PGC-1α contributes to neurodegeneration in Parkinson’s disease. Cell 2011, 144, 689–702. [CrossRef]
- Jo, A.; Lee, Y.; Kam, T.-I.; Kang, S.-U.; Neifert, S.; Karuppagounder, S.S.; Khang, R.; Kang, H.; Park, H.; Chou, S.-C.; et al. PARIS farnesylation prevents neurodegeneration in models of Parkinson’s disease. Sci. Transl. Med. 2021, 13, eaax8891. [CrossRef]
- Narendra, D.; Tanaka, A.; Suen, D.-F.; Youle, R.J. Parkin-induced mitophagy in the pathogenesis of Parkinson disease. Autophagy 2009, 5, 706–708. [CrossRef]
- Palacino, J.J.; Sagi, D.; Goldberg, M.S.; Krauss, S.; Motz, C.; Wacker, M.; Klose, J.; Shen, J. Mitochondrial dysfunction and oxidative damage in parkin-deficient mice. J. Biol. Chem. 2004, 279, 18614–18622. [CrossRef]
- Goldberg, M.S.; Pisani, A.; Haburcak, M.; Vortherms, T.A.; Kitada, T.; Costa, C.; Tong, Y.; Martella, G.; Tscherter, A.; Martins, A.; et al. Nigrostriatal dopaminergic deficits and hypokinesia caused by inactivation of the familial Parkinsonism-linked gene DJ-1. Neuron 2005, 45, 489–496. [CrossRef]
- Itier, J.-M.; Ibanez, P.; Mena, M.A.; Abbas, N.; Cohen-Salmon, C.; Bohme, G.A.; Laville, M.; Pratt, J.; Corti, O.; Pradier, L.; et al. Parkin gene inactivation alters behaviour and dopamine neurotransmission in the mouse. Hum. Mol. Genet. 2003, 12, 2277–2291. [CrossRef]
- Kitada, T.; Pisani, A.; Karouani, M.; Haburcak, M.; Martella, G.; Tscherter, A.; Platania, P.; Wu, B.; Pothos, E.N.; Shen, J. Impaired dopamine release and synaptic plasticity in the striatum of Parkin −/− mice. J. Neurochem. 2009, 110, 613–621. [CrossRef]
- Cortese, G.P.; Zhu, M.; Williams, D.; Heath, S.; Waites, C.L. Parkin Deficiency Reduces Hippocampal Glutamatergic Neurotransmission by Impairing AMPA Receptor Endocytosis. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 12243–12258. [CrossRef]
- Calì, T.; Ottolini, D.; Negro, A.; Brini, M. Enhanced parkin levels favor ER-mitochondria crosstalk and guarantee Ca(2+) transfer to sustain cell bioenergetics. Biochim. Biophys. Acta 2013, 1832, 495–508. [CrossRef]
- Bianchi, K.; Rimessi, A.; Prandini, A.; Szabadkai, G.; Rizzuto, R. Calcium and mitochondria: mechanisms and functions of a troubled relationship. Biochim. Biophys. Acta 2004, 1742, 119–131. [CrossRef]
- Jo, D.; Song, J. Irisin Acts via the PGC-1α and BDNF Pathway to Improve Depression-like Behavior. Clin. Nutr. Res. 2021, 10, 292–302. [CrossRef]
- Zhou, H.; Falkenburger, B.H.; Schulz, J.B.; Tieu, K.; Xu, Z.; Xia, X.G. Silencing of the Pink1 gene expression by conditional RNAi does not induce dopaminergic neuron death in mice. Int. J. Biol. Sci. 2007, 3, 242–250. [CrossRef]
- Madeo, G.; Schirinzi, T.; Martella, G.; Latagliata, E.C.; Puglisi, F.; Shen, J.; Valente, E.M.; Federici, M.; Mercuri, N.B.; Puglisi-Allegra, S.; et al. PINK1 heterozygous mutations induce subtle alterations in dopamine-dependent synaptic plasticity: Synaptic Plasticity in Pink1 +/− Mice. Mov. Disord. 2014, 29, 41–53. [CrossRef]
- Dave, K.D.; De Silva, S.; Sheth, N.P.; Ramboz, S.; Beck, M.J.; Quang, C.; Switzer, R.C.; Ahmad, S.O.; Sunkin, S.M.; Walker, D.; et al. Phenotypic characterization of recessive gene knockout rat models of Parkinson’s disease. Neurobiol. Dis. 2014, 70, 190–203. [CrossRef]
- Stauch, K.L.; Villeneuve, L.M.; Purnell, P.R.; Ottemann, B.M.; Emanuel, K.; Fox, H.S. Loss of Pink1 modulates synaptic mitochondrial bioenergetics in the rat striatum prior to motor symptoms: concomitant complex I respiratory defects and increased complex II-mediated respiration. Proteomics Clin. Appl. 2016, 10, 1205–1217. [CrossRef]
- Villeneuve, L.M.; Purnell, P.R.; Boska, M.D.; Fox, H.S. Early Expression of Parkinson’s Disease-Related Mitochondrial Abnormalities in PINK1 Knockout Rats. Mol. Neurobiol. 2016, 53, 171–186. [CrossRef]
- Creed, R.B.; Goldberg, M.S. Analysis of α-Synuclein Pathology in PINK1 Knockout Rat Brains. Front. Neurosci. 2018, 12, 1034. [CrossRef]
- Li, Z.; Jo, J.; Jia, J.-M.; Lo, S.-C.; Whitcomb, D.J.; Jiao, S.; Cho, K.; Sheng, M. Caspase-3 Activation via Mitochondria Is Required for Long-Term Depression and AMPA Receptor Internalization. Cell 2010, 141, 859–871. [CrossRef]
- Gautier, C.A.; Kitada, T.; Shen, J. Loss of PINK1 causes mitochondrial functional defects and increased sensitivity to oxidative stress. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 11364–11369. [CrossRef]
- Zhang, L.; Shimoji, M.; Thomas, B.; Moore, D.J.; Yu, S.-W.; Marupudi, N.I.; Torp, R.; Torgner, I.A.; Ottersen, O.P.; Dawson, T.M.; et al. Mitochondrial localization of the Parkinson’s disease related protein DJ-1: implications for pathogenesis. Hum. Mol. Genet. 2005, 14, 2063–2073. [CrossRef]
- Guzman, J.N.; Sanchez-Padilla, J.; Wokosin, D.; Kondapalli, J.; Ilijic, E.; Schumacker, P.T.; Surmeier, D.J. Oxidant stress evoked by pacemaking in dopaminergic neurons is attenuated by DJ-1. Nature 2010, 468, 696–700. [CrossRef]
- Goldberg, J.A.; Guzman, J.N.; Estep, C.M.; Ilijic, E.; Kondapalli, J.; Sanchez-Padilla, J.; Surmeier, D.J. Calcium entry induces mitochondrial oxidant stress in vagal neurons at risk in Parkinson’s disease. Nat. Neurosci. 2012, 15, 1414–1421. [CrossRef]
- Heo, J.Y.; Park, J.H.; Kim, S.J.; Seo, K.S.; Han, J.S.; Lee, S.H.; Kim, J.M.; Park, J.I.; Park, S.K.; Lim, K.; et al. DJ-1 Null Dopaminergic Neuronal Cells Exhibit Defects in Mitochondrial Function and Structure: Involvement of Mitochondrial Complex I Assembly. PLoS ONE 2012, 7, e32629. [CrossRef]
- Andres-Mateos, E.; Perier, C.; Zhang, L.; Blanchard-Fillion, B.; Greco, T.M.; Thomas, B.; Ko, H.S.; Sasaki, M.; Ischiropoulos, H.; Przedborski, S.; et al. DJ-1 gene deletion reveals that DJ-1 is an atypical peroxiredoxin-like peroxidase. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 14807–14812. [CrossRef]
- Lopert, P.; Patel, M. Brain mitochondria from DJ-1 knockout mice show increased respiration-dependent hydrogen peroxide consumption. Redox Biol. 2014, 2, 667–672. [CrossRef]
- Chen, W.; Liu, H.; Liu, S.; Kang, Y.; Nie, Z.; Lei, H. Altered prefrontal neurochemistry in the DJ-1 knockout mouse model of Parkinson’s disease: complementary semi-quantitative analyses with in vivo magnetic resonance spectroscopy and MALDI-MSI. Anal. Bioanal. Chem. 2022, 414, 7977–7987. [CrossRef]
- Dringen, R.; Hirrlinger, J. Glutathione pathways in the brain. Biol. Chem. 2003, 384, 505–516. [CrossRef]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [CrossRef]
- Dringen, R.; Gutterer, J.M.; Hirrlinger, J. Glutathione metabolism in brain metabolic interaction between astrocytes and neurons in the defense against reactive oxygen species. Eur. J. Biochem. 2000, 267, 4912–4916. [CrossRef]
- Junn, E.; Jang, W.H.; Zhao, X.; Jeong, B.S.; Mouradian, M.M. Mitochondrial localization of DJ-1 leads to enhanced neuroprotection. J. Neurosci. Res. 2009, 87, 123–129. [CrossRef]
- Irrcher, I.; Aleyasin, H.; Seifert, E.L.; Hewitt, S.J.; Chhabra, S.; Phillips, M.; Lutz, A.K.; Rousseaux, M.W.C.; Bevilacqua, L.; Jahani-Asl, A.; et al. Loss of the Parkinson’s disease-linked gene DJ-1 perturbs mitochondrial dynamics. Hum. Mol. Genet. 2010, 19, 3734–3746. [CrossRef]
- Im, J.-Y.; Lee, K.-W.; Woo, J.-M.; Junn, E.; Mouradian, M.M. DJ-1 induces thioredoxin 1 expression through the Nrf2 pathway. Hum. Mol. Genet. 2012, 21, 3013–3024. [CrossRef]
- Dolgacheva, L.P.; Berezhnov, A.V.; Fedotova, E.I.; Zinchenko, V.P.; Abramov, A.Y. Role of DJ-1 in the mechanism of pathogenesis of Parkinson’s disease. J. Bioenerg. Biomembr. 2019, 51, 175–188. [CrossRef]
- Kitada, T.; Tong, Y.; Gautier, C.A.; Shen, J. Absence of nigral degeneration in aged parkin/DJ-1/PINK1 triple knockout mice. J. Neurochem. 2009, 111, 696–702. [CrossRef]
- Farshim, P.P.; Bates, G.P. Mouse Models of Huntington’s Disease. Methods Mol. Biol. Clifton NJ 2018, 1780, 97–120. [CrossRef]
- Pouladi, M.A.; Morton, A.J.; Hayden, M.R. Choosing an animal model for the study of Huntington’s disease. Nat. Rev. Neurosci. 2013, 14, 708–721. [CrossRef]
- Yu, Z.-X.; Li, S.-H.; Evans, J.; Pillarisetti, A.; Li, H.; Li, X.-J. Mutant huntingtin causes context-dependent neurodegeneration in mice with Huntington’s disease. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 2193–2202. [CrossRef]
- Choo, Y.S.; Johnson, G.V.W.; MacDonald, M.; Detloff, P.J.; Lesort, M. Mutant huntingtin directly increases susceptibility of mitochondria to the calcium-induced permeability transition and cytochrome c release. Hum. Mol. Genet. 2004, 13, 1407–1420. [CrossRef]
- Panov, A.V.; Gutekunst, C.-A.; Leavitt, B.R.; Hayden, M.R.; Burke, J.R.; Strittmatter, W.J.; Greenamyre, J.T. Early mitochondrial calcium defects in Huntington’s disease are a direct effect of polyglutamines. Nat. Neurosci. 2002, 5, 731–736. [CrossRef]
- Mangiarini, L.; Sathasivam, K.; Seller, M.; Cozens, B.; Harper, A.; Hetherington, C.; Lawton, M.; Trottier, Y.; Lehrach, H.; Davies, S.W.; et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell 1996, 87, 493–506. [CrossRef]
- Menalled, L.; El-Khodor, B.F.; Patry, M.; Suárez-Fariñas, M.; Orenstein, S.J.; Zahasky, B.; Leahy, C.; Wheeler, V.; Yang, X.W.; MacDonald, M.; et al. Systematic behavioral evaluation of Huntington’s disease transgenic and knock-in mouse models. Neurobiol. Dis. 2009, 35, 319–336. [CrossRef]
- Bogdanov, M.B.; Andreassen, O.A.; Dedeoglu, A.; Ferrante, R.J.; Beal, M.F. Increased oxidative damage to DNA in a transgenic mouse model of Huntington’s disease. J. Neurochem. 2001, 79, 1246–1249. [CrossRef]
- Tabrizi, S.J.; Workman, J.; Hart, P.E.; Mangiarini, L.; Mahal, A.; Bates, G.; Cooper, J.M.; Schapira, A.H. Mitochondrial dysfunction and free radical damage in the Huntington R6/2 transgenic mouse. Ann. Neurol. 2000, 47, 80–86. [CrossRef]
- Petersen, M.H.; Willert, C.W.; Andersen, J.V.; Madsen, M.; Waagepetersen, H.S.; Skotte, N.H.; Nørremølle, A. Progressive Mitochondrial Dysfunction of Striatal Synapses in R6/2 Mouse Model of Huntington’s Disease. J. Huntingt. Dis. 2022, 11, 121–140. [CrossRef]
- Milnerwood, A.J.; Raymond, L.A. Corticostriatal synaptic function in mouse models of Huntington’s disease: early effects of huntingtin repeat length and protein load. J. Physiol. 2007, 585, 817–831. [CrossRef]
- Giralt, A.; Saavedra, A.; Alberch, J.; Pérez-Navarro, E. Cognitive Dysfunction in Huntington’s Disease: Humans, Mouse Models and Molecular Mechanisms. J. Huntingt. Dis. 2012, 1, 155–173. [CrossRef]
- Ghiglieri, V.; Bagetta, V.; Calabresi, P.; Picconi, B. Functional interactions within striatal microcircuit in animal models of Huntington’s disease. Neuroscience 2012, 211, 165–184. [CrossRef]
- Nithianantharajah, J.; Hannan, A.J. Dysregulation of synaptic proteins, dendritic spine abnormalities and pathological plasticity of synapses as experience-dependent mediators of cognitive and psychiatric symptoms in Huntington’s disease. Neuroscience 2013, 251, 66–74. [CrossRef]
- Ghiglieri, V.; Campanelli, F.; Marino, G.; Natale, G.; Picconi, B.; Calabresi, P. Corticostriatal synaptic plasticity alterations in the R6/1 transgenic mouse model of Huntington’s disease. J. Neurosci. Res. 2019, 97, 1655–1664. [CrossRef]
- Rosenstock, T.R.; Bertoncini, C.R.A.; Teles, A.V.; Hirata, H.; Fernandes, M.J.S.; Smaili, S.S. Glutamate-induced alterations in Ca2+ signaling are modulated by mitochondrial Ca2+ handling capacity in brain slices of R6/1 transgenic mice. Eur. J. Neurosci. 2010, 32, 60–70. [CrossRef]
- Clark, J.B. N-acetyl aspartate: a marker for neuronal loss or mitochondrial dysfunction. Dev. Neurosci. 1998, 20, 271–276. [CrossRef]
- Jenkins, B.G.; Klivenyi, P.; Kustermann, E.; Andreassen, O.A.; Ferrante, R.J.; Rosen, B.R.; Beal, M.F. Nonlinear decrease over time in N-acetyl aspartate levels in the absence of neuronal loss and increases in glutamine and glucose in transgenic Huntington’s disease mice. J. Neurochem. 2000, 74, 2108–2119. [CrossRef]
- Andreassen, O.A.; Dedeoglu, A.; Ferrante, R.J.; Jenkins, B.G.; Ferrante, K.L.; Thomas, M.; Friedlich, A.; Browne, S.E.; Schilling, G.; Borchelt, D.R.; et al. Creatine increase survival and delays motor symptoms in a transgenic animal model of Huntington’s disease. Neurobiol. Dis. 2001, 8, 479–491. [CrossRef]
- Jenkins, B.G.; Andreassen, O.A.; Dedeoglu, A.; Leavitt, B.; Hayden, M.; Borchelt, D.; Ross, C.A.; Ferrante, R.J.; Beal, M.F. Effects of CAG repeat length, HTT protein length and protein context on cerebral metabolism measured using magnetic resonance spectroscopy in transgenic mouse models of Huntington’s disease. J. Neurochem. 2005, 95, 553–562. [CrossRef]
- Tkac, I.; Dubinsky, J.M.; Keene, C.D.; Gruetter, R.; Low, W.C. Neurochemical changes in Huntington R6/2 mouse striatum detected by in vivo 1H NMR spectroscopy. J. Neurochem. 2007, 100, 1397–1406. [CrossRef]
- Browne, S.E. Mitochondria and Huntington’s disease pathogenesis: insight from genetic and chemical models. Ann. N. Y. Acad. Sci. 2008, 1147, 358–382. [CrossRef]
- Lopes, C.; Ferreira, I.L.; Maranga, C.; Beatriz, M.; Mota, S.I.; Sereno, J.; Castelhano, J.; Abrunhosa, A.; Oliveira, F.; De Rosa, M.; et al. Mitochondrial and redox modifications in early stages of Huntington’s disease. Redox Biol. 2022, 56, 102424. [CrossRef]
- Wright, D.J.; Renoir, T.; Smith, Z.M.; Frazier, A.E.; Francis, P.S.; Thorburn, D.R.; McGee, S.L.; Hannan, A.J.; Gray, L.J. N-Acetylcysteine improves mitochondrial function and ameliorates behavioral deficits in the R6/1 mouse model of Huntington’s disease. Transl. Psychiatry 2015, 5, e492. [CrossRef]
- Herbst, E. a. F.; Holloway, G.P. Exercise training normalizes mitochondrial respiratory capacity within the striatum of the R6/1 model of Huntington’s disease. Neuroscience 2015, 303, 515–523. [CrossRef]
- Gardian, G.; Browne, S.E.; Choi, D.-K.; Klivenyi, P.; Gregorio, J.; Kubilus, J.K.; Ryu, H.; Langley, B.; Ratan, R.R.; Ferrante, R.J.; et al. Neuroprotective effects of phenylbutyrate in the N171-82Q transgenic mouse model of Huntington’s disease. J. Biol. Chem. 2005, 280, 556–563. [CrossRef]
- Hermel, E.; Gafni, J.; Propp, S.S.; Leavitt, B.R.; Wellington, C.L.; Young, J.E.; Hackam, A.S.; Logvinova, A.V.; Peel, A.L.; Chen, S.F.; et al. Specific caspase interactions and amplification are involved in selective neuronal vulnerability in Huntington’s disease. Cell Death Differ. 2004, 11, 424–438. [CrossRef]
- Wellington, C.L.; Ellerby, L.M.; Hackam, A.S.; Margolis, R.L.; Trifiro, M.A.; Singaraja, R.; McCutcheon, K.; Salvesen, G.S.; Propp, S.S.; Bromm, M.; et al. Caspase cleavage of gene products associated with triplet expansion disorders generates truncated fragments containing the polyglutamine tract. J. Biol. Chem. 1998, 273, 9158–9167. [CrossRef]
- Ona, V.O.; Li, M.; Vonsattel, J.P.; Andrews, L.J.; Khan, S.Q.; Chung, W.M.; Frey, A.S.; Menon, A.S.; Li, X.J.; Stieg, P.E.; et al. Inhibition of caspase-1 slows disease progression in a mouse model of Huntington’s disease. Nature 1999, 399, 263–267. [CrossRef]
- Carroll, J.B.; Southwell, A.L.; Graham, R.K.; Lerch, J.P.; Ehrnhoefer, D.E.; Cao, L.-P.; Zhang, W.-N.; Deng, Y.; Bissada, N.; Henkelman, R.M.; et al. Mice lacking caspase-2 are protected from behavioral changes, but not pathology, in the YAC128 model of Huntington disease. Mol. Neurodegener. 2011, 6, 59. [CrossRef]
- Avenali, M.; Cerri, S.; Ongari, G.; Ghezzi, C.; Pacchetti, C.; Tassorelli, C.; Valente, E.M.; Blandini, F. Profiling the Biochemical Signature of GBA-Related Parkinson’s Disease in Peripheral Blood Mononuclear Cells. Mov. Disord. 2021, 36, 1267–1272. [CrossRef]
- Petrillo, S.; Schirinzi, T.; Di Lazzaro, G.; D’Amico, J.; Colona, V.L.; Bertini, E.; Pierantozzi, M.; Mari, L.; Mercuri, N.B.; Piemonte, F.; et al. Systemic Activation of Nrf2 Pathway in Parkinson’s Disease. Mov. Disord. 2020, 35, 180–184. [CrossRef]
- Schirinzi, T.; Salvatori, I.; Zenuni, H.; Grillo, P.; Valle, C.; Martella, G.; Mercuri, N.B.; Ferri, A. Pattern of Mitochondrial Respiration in Peripheral Blood Cells of Patients with Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 10863. [CrossRef]
- Annesley, S.J.; Lay, S.T.; De Piazza, S.W.; Sanislav, O.; Hammersley, E.; Allan, C.Y.; Francione, L.M.; Bui, M.Q.; Chen, Z.-P.; Ngoei, K.R.W.; et al. Immortalized Parkinson’s disease lymphocytes have enhanced mitochondrial respiratory activity. Dis. Model. Mech. 2016, 9, 1295–1305. [CrossRef]
- Haylett, W.; Swart, C.; van der Westhuizen, F.; van Dyk, H.; van der Merwe, L.; van der Merwe, C.; Loos, B.; Carr, J.; Kinnear, C.; Bardien, S. Altered Mitochondrial Respiration and Other Features of Mitochondrial Function in Parkin-Mutant Fibroblasts from Parkinson’s Disease Patients. Park. Dis. 2016, 2016, 1819209. [CrossRef]
- Antony, P.M.A.; Kondratyeva, O.; Mommaerts, K.; Ostaszewski, M.; Sokolowska, K.; Baumuratov, A.S.; Longhino, L.; Poulain, J.F.; Grossmann, D.; Balling, R.; et al. Fibroblast mitochondria in idiopathic Parkinson’s disease display morphological changes and enhanced resistance to depolarization. Sci. Rep. 2020, 10, 1569. [CrossRef]
- Fais, M.; Dore, A.; Galioto, M.; Galleri, G.; Crosio, C.; Iaccarino, C. Parkinson’s Disease-Related Genes and Lipid Alteration. Int. J. Mol. Sci. 2021, 22, 7630. [CrossRef]
- Smith, A.M.; Depp, C.; Ryan, B.J.; Johnston, G.I.; Alegre-Abarrategui, J.; Evetts, S.; Rolinski, M.; Baig, F.; Ruffmann, C.; Simon, A.K.; et al. Mitochondrial dysfunction and increased glycolysis in prodromal and early Parkinson’s blood cells. Mov. Disord. Off. J. Mov. Disord. Soc. 2018, 33, 1580–1590. [CrossRef]
- Havelund, J.F.; Heegaard, N.H.H.; Færgeman, N.J.K.; Gramsbergen, J.B. Biomarker Research in Parkinson’s Disease Using Metabolite Profiling. Metabolites 2017, 7, 42. [CrossRef]
- Willkommen, D.; Lucio, M.; Moritz, F.; Forcisi, S.; Kanawati, B.; Smirnov, K.S.; Schroeter, M.; Sigaroudi, A.; Schmitt-Kopplin, P.; Michalke, B. Metabolomic investigations in cerebrospinal fluid of Parkinson’s disease. PloS One 2018, 13, e0208752. [CrossRef]
- Saiki, S.; Hatano, T.; Fujimaki, M.; Ishikawa, K.-I.; Mori, A.; Oji, Y.; Okuzumi, A.; Fukuhara, T.; Koinuma, T.; Imamichi, Y.; et al. Decreased long-chain acylcarnitines from insufficient β-oxidation as potential early diagnostic markers for Parkinson’s disease. Sci. Rep. 2017, 7, 7328. [CrossRef]
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [CrossRef]
- Schirinzi, T.; Di Lazzaro, G.; Sancesario, G.M.; Summa, S.; Petrucci, S.; Colona, V.L.; Bernardini, S.; Pierantozzi, M.; Stefani, A.; Mercuri, N.B.; et al. Young-onset and late-onset Parkinson’s disease exhibit a different profile of fluid biomarkers and clinical features. Neurobiol. Aging 2020, 90, 119–124. [CrossRef]
- Schirinzi, T.; Vasco, G.; Zanni, G.; Petrillo, S.; Piemonte, F.; Castelli, E.; Bertini, E.S. Serum uric acid in Friedreich Ataxia. Clin. Biochem. 2018, 54, 139–141. [CrossRef]
- Schirinzi, T.; Di Lazzaro, G.; Colona, V.L.; Imbriani, P.; Alwardat, M.; Sancesario, G.M.; Martorana, A.; Pisani, A. Assessment of serum uric acid as risk factor for tauopathies. J. Neural Transm. Vienna Austria 1996 2017, 124, 1105–1108. [CrossRef]
- Wei, Z.; Li, X.; Li, X.; Liu, Q.; Cheng, Y. Oxidative Stress in Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Mol. Neurosci. 2018, 11, 236. [CrossRef]
- Ueno, S.-I.; Hatano, T.; Okuzumi, A.; Saiki, S.; Oji, Y.; Mori, A.; Koinuma, T.; Fujimaki, M.; Takeshige-Amano, H.; Kondo, A.; et al. Nonmercaptalbumin as an oxidative stress marker in Parkinson’s and PARK2 disease. Ann. Clin. Transl. Neurol. 2020, 7, 307–317. [CrossRef]
- Yamagishi, Y.; Saigoh, K.; Saito, Y.; Ogawa, I.; Mitsui, Y.; Hamada, Y.; Samukawa, M.; Suzuki, H.; Kuwahara, M.; Hirano, M.; et al. Diagnosis of Parkinson’s disease and the level of oxidized DJ-1 protein. Neurosci. Res. 2018, 128, 58–62. [CrossRef]
- Sancesario, G.M.; Di Lazzaro, G.; Grillo, P.; Biticchi, B.; Giannella, E.; Alwardat, M.; Pieri, M.; Bernardini, S.; Mercuri, N.B.; Pisani, A.; et al. Biofluids profile of α-Klotho in patients with Parkinson’s disease. Parkinsonism Relat. Disord. 2021, 90, 62–64. [CrossRef]
- Fernández-Espejo, E.; Rodriguez de Fonseca, F.; Suárez, J.; Martín de Pablos, Á. Cerebrospinal fluid lactoperoxidase level is enhanced in idiopathic Parkinson’s disease, and correlates with levodopa equivalent daily dose. Brain Res. 2021, 1761, 147411. [CrossRef]
- Sun, W.; Zheng, J.; Ma, J.; Wang, Z.; Shi, X.; Li, M.; Huang, S.; Hu, S.; Zhao, Z.; Li, D. Increased Plasma Heme Oxygenase-1 Levels in Patients With Early-Stage Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 621508. [CrossRef]
- Shamir, R.; Klein, C.; Amar, D.; Vollstedt, E.-J.; Bonin, M.; Usenovic, M.; Wong, Y.C.; Maver, A.; Poths, S.; Safer, H.; et al. Analysis of blood-based gene expression in idiopathic Parkinson disease. Neurology 2017, 89, 1676–1683. [CrossRef]
- Picca, A.; Guerra, F.; Calvani, R.; Marini, F.; Biancolillo, A.; Landi, G.; Beli, R.; Landi, F.; Bernabei, R.; Bentivoglio, A.R.; et al. Mitochondrial Signatures in Circulating Extracellular Vesicles of Older Adults with Parkinson’s Disease: Results from the EXosomes in PArkiNson’s Disease (EXPAND) Study. J. Clin. Med. 2020, 9, 504. [CrossRef]
- Paul, B.D.; Snyder, S.H. Impaired Redox Signaling in Huntington’s Disease: Therapeutic Implications. Front. Mol. Neurosci. 2019, 12, 68. [CrossRef]
- Jędrak, P.; Mozolewski, P.; Węgrzyn, G.; Więckowski, M.R. Mitochondrial alterations accompanied by oxidative stress conditions in skin fibroblasts of Huntington’s disease patients. Metab. Brain Dis. 2018, 33, 2005–2017. [CrossRef]
- Vanisova, M.; Stufkova, H.; Kohoutova, M.; Rakosnikova, T.; Krizova, J.; Klempir, J.; Rysankova, I.; Roth, J.; Zeman, J.; Hansikova, H. Mitochondrial organization and structure are compromised in fibroblasts from patients with Huntington’s disease. Ultrastruct. Pathol. 2022, 46, 462–475. [CrossRef]
- Neueder, A.; Orth, M. Mitochondrial biology and the identification of biomarkers of Huntington’s disease. Neurodegener. Dis. Manag. 2020, 10, 243–255. [CrossRef]
- Chen, C.-M.; Wu, Y.-R.; Cheng, M.-L.; Liu, J.-L.; Lee, Y.-M.; Lee, P.-W.; Soong, B.-W.; Chiu, D.T.-Y. Increased oxidative damage and mitochondrial abnormalities in the peripheral blood of Huntington’s disease patients. Biochem. Biophys. Res. Commun. 2007, 359, 335–340. [CrossRef]
- Spinelli, J.B.; Zaganjor, E. Mitochondrial efficiency directs cell fate. Nat. Cell Biol. 2022, 24, 125–126. [CrossRef]
- Ahmad, M.; Wolberg, A.; Kahwaji, C.I. Biochemistry, Electron Transport Chain. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2022.
- McAdam, E.; Brem, R.; Karran, P. Oxidative Stress-Induced Protein Damage Inhibits DNA Repair and Determines Mutation Risk and Therapeutic Efficacy. Mol. Cancer Res. MCR 2016, 14, 612–622. [CrossRef]
- Korovila, I.; Hugo, M.; Castro, J.P.; Weber, D.; Höhn, A.; Grune, T.; Jung, T. Proteostasis, oxidative stress and aging. Redox Biol. 2017, 13, 550–567. [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [CrossRef]
- Scarpulla, R.C. Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol. Rev. 2008, 88, 611–638. [CrossRef]
- Scarpulla, R.C. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim. Biophys. Acta 2011, 1813, 1269–1278. [CrossRef]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: a potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc. Natl. Acad. Sci. U. S. A. 1994, 91, 1309–1313. [CrossRef]
- Saxton, W.M.; Hollenbeck, P.J. The axonal transport of mitochondria. J. Cell Sci. 2012, 125, 2095–2104. [CrossRef]
- Schwarz, T.L. Mitochondrial trafficking in neurons. Cold Spring Harb. Perspect. Biol. 2013, 5, a011304. [CrossRef]
- Misgeld, T.; Schwarz, T.L. Mitostasis in Neurons: Maintaining Mitochondria in an Extended Cellular Architecture. Neuron 2017, 96, 651–666. [CrossRef]
- Franco-Iborra, S.; Vila, M.; Perier, C. Mitochondrial Quality Control in Neurodegenerative Diseases: Focus on Parkinson’s Disease and Huntington’s Disease. Front. Neurosci. 2018, 12. [CrossRef]
- Keating, D.J. Mitochondrial dysfunction, oxidative stress, regulation of exocytosis and their relevance to neurodegenerative diseases. J. Neurochem. 2008, 104, 298–305. [CrossRef]
- Schirinzi, T.; Madeo, G.; Martella, G.; Maltese, M.; Picconi, B.; Calabresi, P.; Pisani, A. Early synaptic dysfunction in Parkinson’s disease: Insights from animal models: Early Synaptic Dysfunction in PD. Mov. Disord. 2016, 31, 802–813. [CrossRef]
- Gerencser, A.A.; Doczi, J.; Töröcsik, B.; Bossy-Wetzel, E.; Adam-Vizi, V. Mitochondrial swelling measurement in situ by optimized spatial filtering: astrocyte-neuron differences. Biophys. J. 2008, 95, 2583–2598. [CrossRef]
- O’Sullivan, J.D.B.; Bullen, A.; Mann, Z.F. Mitochondrial form and function in hair cells. Hear. Res. 2023, 428, 108660. [CrossRef]
- Woo, J.; Cho, H.; Seol, Y.; Kim, S.H.; Park, C.; Yousefian-Jazi, A.; Hyeon, S.J.; Lee, J.; Ryu, H. Power Failure of Mitochondria and Oxidative Stress in Neurodegeneration and Its Computational Models. Antioxid. Basel Switz. 2021, 10, 229. [CrossRef]
- Polyzos, A.A.; McMurray, C.T. The chicken or the egg: mitochondrial dysfunction as a cause or consequence of toxicity in Huntington’s disease. Mech. Ageing Dev. 2017, 161, 181–197. [CrossRef]
- Lang, A.E.; Espay, A.J. Disease Modification in Parkinson’s Disease: Current Approaches, Challenges, and Future Considerations: Disease Modification in PD. Mov. Disord. 2018, 33, 660–677. [CrossRef]
- Lee, L.-H.N.; Huang, C.-S.; Chuang, H.-H.; Lai, H.-J.; Yang, C.-K.; Yang, Y.-C.; Kuo, C.-C. An electrophysiological perspective on Parkinson’s disease: symptomatic pathogenesis and therapeutic approaches. J. Biomed. Sci. 2021, 28, 85. [CrossRef]
- Jamwal, S.; Kumar, P. Insight Into the Emerging Role of Striatal Neurotransmitters in the Pathophysiology of Parkinson’s Disease and Huntington’s Disease: A Review. Curr. Neuropharmacol. 2019, 17, 165–175. [CrossRef]
- Terreros-Roncal, J.; Moreno-Jiménez, E.P.; Flor-García, M.; Rodríguez-Moreno, C.B.; Trinchero, M.F.; Cafini, F.; Rábano, A.; Llorens-Martín, M. Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science 2021, 374, 1106–1113. [CrossRef]
- Levine, M.S.; Cepeda, C.; Hickey, M.A.; Fleming, S.M.; Chesselet, M.-F. Genetic mouse models of Huntington’s and Parkinson’s diseases: illuminating but imperfect. Trends Neurosci. 2004, 27, 691–697. [CrossRef]
- Carmo, C.; Naia, L.; Lopes, C.; Rego, A.C. Mitochondrial Dysfunction in Huntington’s Disease. Adv. Exp. Med. Biol. 2018, 1049, 59–83. [CrossRef]
Figure 1.
Schematic representation of the mitochondrial functions.From left: Under physiological conditions, mitochondria supply ATP through OXPHOS. Krebs cycle enzymes use acetyl-coenzyme A to reduce NADH and FADH2, which are used for energy transfer to the electron transport chain (ETC) embedded in the inner mitochondrial membrane. OXPHOS is also an important source of ROS, whose basal levels are maintained by the radical scavenging network. Mitochondria also play a crucial role in calcium homeostasis. The voltage-dipendent anion channel (VDAC) and the mitochondrial Ca2+ uniporter complex (MCU) finely controls Ca2+ passage across the mitochondrial membranes while the mitochondrial Na+/Ca2+ exchanger (NCLX) is one of the central units involved in Ca2+ extrusion. Under normal conditions, through ROS generation and redox signaling, mitochondria can control cellular metabolism, physiology, inflammatory response, and immune function and act as important signaling molecules in the cell by activating various protein kinases. In contrast, the overproduction of ROS and dysregulation of the redox signaling system result in oxidative stress that can lead to mitochondrial damage. Malfunctioning mitochondria can be selectively removed through mitophagy, or, as all other defense mechanisms fail, the neuron can orchestrate its self-destruction by activating the intrinsic suicide program of apoptosis.
Figure 1.
Schematic representation of the mitochondrial functions.From left: Under physiological conditions, mitochondria supply ATP through OXPHOS. Krebs cycle enzymes use acetyl-coenzyme A to reduce NADH and FADH2, which are used for energy transfer to the electron transport chain (ETC) embedded in the inner mitochondrial membrane. OXPHOS is also an important source of ROS, whose basal levels are maintained by the radical scavenging network. Mitochondria also play a crucial role in calcium homeostasis. The voltage-dipendent anion channel (VDAC) and the mitochondrial Ca2+ uniporter complex (MCU) finely controls Ca2+ passage across the mitochondrial membranes while the mitochondrial Na+/Ca2+ exchanger (NCLX) is one of the central units involved in Ca2+ extrusion. Under normal conditions, through ROS generation and redox signaling, mitochondria can control cellular metabolism, physiology, inflammatory response, and immune function and act as important signaling molecules in the cell by activating various protein kinases. In contrast, the overproduction of ROS and dysregulation of the redox signaling system result in oxidative stress that can lead to mitochondrial damage. Malfunctioning mitochondria can be selectively removed through mitophagy, or, as all other defense mechanisms fail, the neuron can orchestrate its self-destruction by activating the intrinsic suicide program of apoptosis.
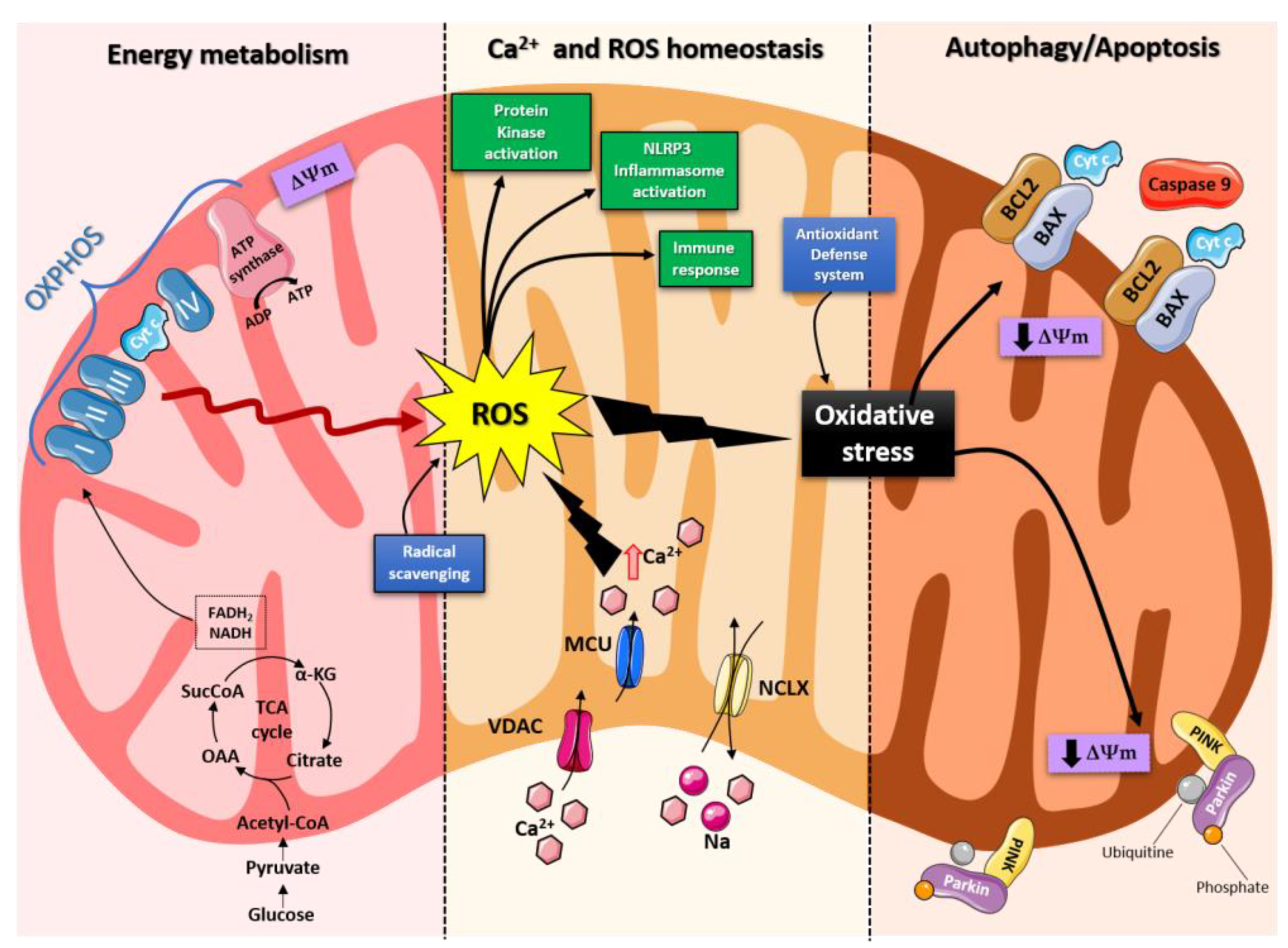
Figure 3.
PubMed evidences of mitochondria involvement on neuro-degenerative/degenerative disease. Records were obtained from research articles published until January 2023, 2020. Article publications were obtained from following searches on PubMed: “Neurodegenerative disease and mitochondria; degenerative disease and mitochondria; HD and mitochondria; PD and mitochondria”. To realize the pies on sub-items, we have selected “ PD, genes and mitochondria; PD, PINK1 and mitochondria; PD, PINK1, mitophagy and mitochondria; HD, mHHT and mitochondria; HD, mHHT, mithophagy and mitochondria.
Figure 3.
PubMed evidences of mitochondria involvement on neuro-degenerative/degenerative disease. Records were obtained from research articles published until January 2023, 2020. Article publications were obtained from following searches on PubMed: “Neurodegenerative disease and mitochondria; degenerative disease and mitochondria; HD and mitochondria; PD and mitochondria”. To realize the pies on sub-items, we have selected “ PD, genes and mitochondria; PD, PINK1 and mitochondria; PD, PINK1, mitophagy and mitochondria; HD, mHHT and mitochondria; HD, mHHT, mithophagy and mitochondria.
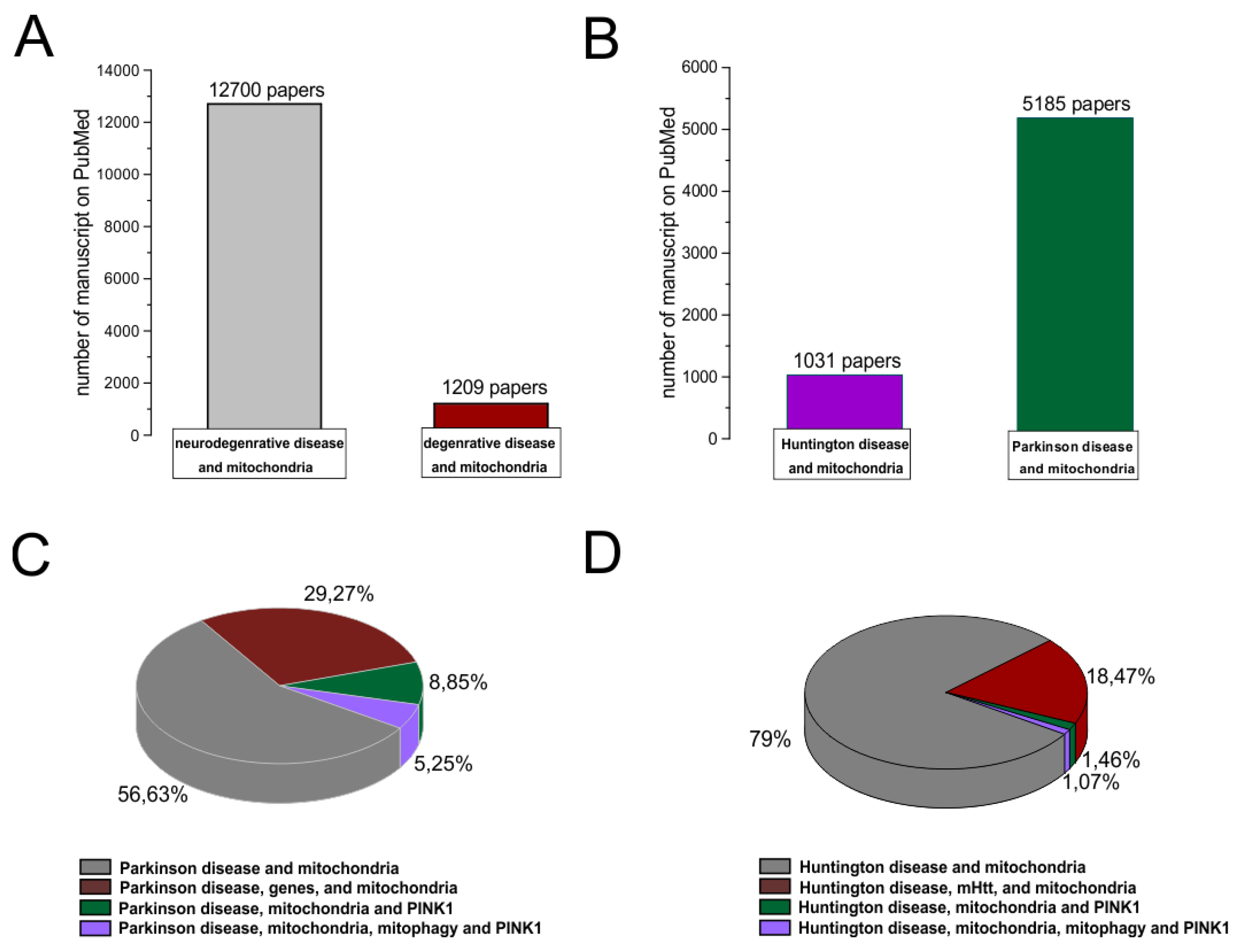
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



