Submitted:
30 March 2023
Posted:
03 April 2023
You are already at the latest version
Abstract
The effect of biotic (e.g., growth characteristics) and a-biotic (e.g., crown closure) factors on cone production, fertility variation and gene diversity was evaluated in two consecutive years in 130 year old natural populations of Taurus cedar also called cedar of Lebanon (Cedrus libani A. Rich.). The a-biotic factors viz., altitude, temperature, aspect and rainfall varied; while some of them can be managed by cultural operations like management of crown and stand density. Impact of crown closure on cone production, fertility variation and related parameters were observed in Taurus cedar’s natural populations sampled from southern Turkey. Maximum cone production of 29 and 40/tree was recorded with full crown closure in two consecutive years by large differences among individuals within crown closure class. For instance, cone productions were between 10 and 67 in full crown closure of first year. The sibling coefficient, which is a measure of the fertility variation in a stand, was the highest (1.16) for full crown closure in the first (meaning 86% fertile trees) and the second year (1.55, 65%), while it was the lowest for medium crown closure (1.09, 92%) in both years. Gene diversity decreased from a degraded patch (0.987) to that with full crown closure (0.984). Results of variance analysis showed significant (p≤0.05) difference between crown closure classes for cone production within second year, and between years. Significant positive (p0.05) correlations were noted between years for cone production (r=0.22), and between cone production and crown closure in first and second years (r=0.29). However, growth characteristics had no effective on cone productions in individual crown closure classes, while there were significant (p0.05) correlations between diameter at breast height and cone production for both years (r= 0.15 and 0.17) in pooled populations.
Keywords:
crown cover
; effective size
; reproduction
; seed
; status number
; variation
1. Introduction
Taurus cedar also known cedar of Lebanon (Cedrus libani A. Rich.), has its main natural occurrence in the Taurus Mountains in southern Turkey, generally between 800 and 2100 m elevation. It can be found at lower (500-600 m) and higher (2400 m) elevations as small populations or small groups and individuals [1]. Its reported extent in southern Turkey is 405424 ha [2]. The species is classified as one of the economically and ecologically important tree species for Turkish forestry and the “National Tree Breeding and Seed Production Program” [3] because of valuable wood product and social-cultural importance of the species. The species is also used in reforestation and afforestation practices togetherwith landscape planning as monumental tree. Annual seed production is about 139 tones for different forestry practices of Taurus cedar based on forestry inventory [2]. 22 seed stands at 3437.8 ha were selected to supply seed demand of the species together with 9 seed orchards established at 60.55 ha [2]. Reported maxima, for different individual trees, are 1000 years age, 3 m stem diameter and 50 m height [1]. Taurus cedar generally bears cone by 30 years in natural stands. Male flowers of the dioecious species appear in July and elongate 3-5 cm in August, while female flowers can be seen in September. Pollination takes place in September or early October, depending on the elevation. The following year between April and June, conelets develop to mature cone sizes. Opening of the cone scales begins in October, about 25-26 months after flowering [4]. Good seed years have been reported as generally every 2-3 years [1] or 3-5 years [4] in the species. Knowledge of the fecundity and fertility (expressed as the individual's ability to fertilize - reproductive success), estimated from cones, flowers, pollen, fruit and seed production [5,6,7,8] is an important prerequisite of breeding and seed production program. Fertility variation estimated based on reproductive characters is easy, cheap, and light survey in plant science. While some studies reported reproductive data of the species [i.e., 4,9,10,11,12], the impact of crown closure on cone production and genetic parameters have not been studied. It is clear that many biological (i.e., number of individuals per area, age, species, growth characteristics, stand structure, population) [8,9,10,13,14] and environmental (i.e., edaphic, altitude, temperature, aspects and rainfall) [7,12,13,15,16] factors impact negatively or positively cone production and fertility variation. However, crown closure can be managed by silvicultural practices. While preparatory cutting practice for natural regeneration in shelter-wood method has many ecological and biological functions like improving the soil characteristics and seedbed, it also results increased strobili, cone and seed production in Taurus cedar. Boydak [1] reported that additional seeds are suggested for shelter-wood and clear-cut areas in natural regeneration practices of Taurus cedar. So, additional seeds are needed in natural regeneration practices of the species. One of the stages of the shelter-wood method is to be provoked of seed trees in natural regeneration practices by crown closure balancing to obtain additional cone/seed crops. Odabasi [4] reported that cone production could be related to crown closure. However, it has not been investigated detail, yet. Effect of crown closure on cone production, fertility variation and related parameters need to be observed to estimate the seed productivity of a population.
The purposes of this study are 1) to estimate the effect of crown closure classes on cone production, 2) to estimate fertility variation and its allied parameters such as effective number of parents and gene diversity in natural populations, 3) to estimate relations among the growth characteristics and cone productions, to improve natural regeneration, management of seed sources and other silvicultural practices of C. libani.
2. Materials and Methods
2.1. Study area
Populations of Taurus cedar were selected from natural stands available between 37°50' and 37°53' N and between 31°17' and 31°20' E at average 1630 m from southern Turkey. Averages of three heights (H), diameter at base (D0), diameter at breast height (d1.30), crown diameter (CD) and age, and altitude of the crown closure classes (CCCs) are given in Table 1.
Monthly maximum, minimum and average temperature (Figure 1) and average monthly precipitation and relative humidity (Figure 2) of the location were observed during the study period for two consecutive years (2020 - year 1 and 2021 - year 2).
Fifty genotypes each were selected from four crown closure classes [17]– crown closure classified using Spherical densitometer as: degraded forest (DF- up to 10%), low crown coverage (LC - 11% to 40%), medium crown coverage (MC - 41% to 70%) and full crown coverage (FC - 71% to 100%) from natural area of the species.
2.2. Data collection and analysis
The numbers of brown mature cones which had 8.5 cm length and 5 cm diameter (Odabasi, 1990) (Figure 3) were counted from the fifty phenotypically selected and marked trees during two consecutive years (2020-2021).
The following GLM of ANOVA was used to analyses the difference of cone productions among CCCs and years by the SAS software [18] in the present study.
Where Yijk is the observation from the kth tree of the jth crown closure in the ith year, μ is the overall mean of cone production, Fi is the effect of ith year, B(F)j(i) is the effect of the jth population in the ith year, and eijk is the random error.
Phenotypic Pearson’ correlation of cone production between years was estimated by Rohlf and Sokal [19].
Cone fertility variation (expressed as sibling coefficient -Ψ) was estimated as per Kang and Lindgren [20] and Bilir [7] as:
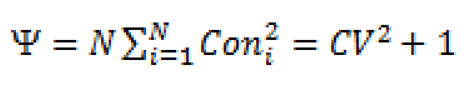
Where N is the census number of trees, Coni is the cone fertility of the ith individual, and CV is the coefficient of variation in total fertility. The coefficients of variations (CV) for cone fertility were calculated and applied to estimate the fertility variation in parents.
In this paper, the fertility of ith individual was estimated by the proportion of cone production in each crown closure. Therefore, the cone fertility represents the total contribution as zygotic parents.
The effective number of parents (Np), defined as the number of individuals in which an idealized population would produce the same number of offspring (sibs) as the real population, or defined as a function of fertility variation was estimated [20] as:
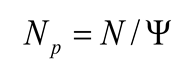
The gene diversity (GD) was estimated based on census number (N) and fertility variation (Ψ) [21] as:
3. Results
3.1. Cone production
The mean cone production varied between years and among crown closure classes within year (Figure 4), and among individuals within the crown closure class (Table 2).
Highest average cone production (29.5 and 40.3) was noticed in FC in both years (year 1 and 2 respectively- Table 2), while for both years pooled it ranged from 34.9 (FC), 27.2 (MC), 26.8 (DF), to 26.3 (LC) in the different CCCs (Figure 4).
The cone production was 20% higher (31.4/tree) in year 2 than year 1 (26.2). The most fecund five trees (10% of trees) produced about 17% and 24% of total cones in year 1 and 2 respectively in all the crown classes. Cone production varied among individual trees within crown class in both years ranging from 10 to 50 in LC (year 1), and 8 to 108 in FC (year 2) (Table 2). The cone production in FC was very different from the other crown classes in both years (Table 2). The coefficient of variation (CV) increased from 32.8% (DF) to 41.0% (FC) in year 1 and from 51.4% (DF) to 74.6% (FC) in year 2 (Table 2). It was also 37.4% in year 1, 67.1% in year 2, and 43.6% for pooled years across CCCs. The differences between years, and among CCCs in second year were significant (p≤0.05) for the cone productions, while CCCs x year interaction was not significant (p>0.05).
Correlation analysis showed positive and significant (p<0.05) correlations between years (r=0.22) for cone production, and between cone production and crown closure classes (r=0.29). Growth characteristics had no effective on cone productions in individual crown closure classes opposite to significant (p≤0.05) correlations between diameter at breast height and cone production (r= 0.15 and 0.17) for both years in pooled populations.
3.2. Fertility variation, effective number of parents and gene diversity
The fertility variation (sibling coefficient, Ψ) ranged from 1.11 (DF and LC) to 1.16 (FC) in year 1, increased with the crown closure from 1.26 (DF) to 1.55 (FC) in year 2. The fertility variations of CCCs in year 2 were decreased by combining the years, while the combined fertility variation (1.09) was lower than within each year (1.14 and 1.37) of MC (Table 3 and Figure 5). It was 1.14 (88% fertile trees), 1.45 (69%) and 1.19 (84%) at years 1 and 2, and across CCCs respectively.
The fertility variation was more or less similar between different crown closure classes in year 1 compared to year 2 - DF (1.26), LC (1.31), MC (1.37) and FC (1.55) (Table 3). Depending on the fertility variation the effective number of parents (Np) also showed the same trend ranging from 42.91 (86% fertile trees) in FC to 45.23 (91%) in DF in year 1, and from 32.35 (65%) in FC to 39.71 (79%) in DF in year 2 (Table 3). The effective population size, equivalent to the ideal populations, was 21% larger in year 1 than year 2, based on the fertility variation of individuals in the full crown closure, while it was 13% in low crown closure (Table 3).
Gene diversity was higher in first year than in second year in all CCCs (Table 3). Estimated loss of gene diversity from other crown closure classes to FC was 0.001 in year 1. Between the two years FC had highest loss of gene diversity of 0.004 (0.988 and 0.984 in two years). However, the gene diversity was higher (0.988 and 0.989) in pooled years of each CCCs than in year 2 (Table 3). The relative number of parents ranged from 65% to 91% in all classes of crowns (Table 3).
3.3. Cumulative contribution curve
The cumulative contribution of trees to the overall fertility estimates for years 1, 2 and pooled contribution of two years are shown in Figure 6, Figure 7 and Figure 8.
The cumulative contribution curve describes the relative proportion of trees to the accumulative gamete contribution. In an ideal situation, that is expecting an equal contribution from each of the individuals (a condition where Ψ=1), this cumulative curve attains a straight line. There was a deviation in cumulative contribution from expectation in all crown closure classes. It can be inferred that various crown closure classes (DF, LC, MC and FC) showed a similar trend in both years (Figure 6 and Figure 7). Only FC showed a skewed distribution in year 2. It could thus be concluded that the FC deviated from the ideal situation more compared to the other three crown closures only in year 2 (Figure 7) and fertility combined for two years (Figure 8). When seed crop of two years was mixed, the cumulative contribution increased (Figure 8) in all crown closure classes.
4. Discussion
4.1. Cone production
Average cone numbers of DF, LC, and MC (Table 2 and Figure 4) were slightly higher than two populations of the species reported earlier (20 and 22) [9] and a population (19) [11] of Taurus cedar, while it was dramatically higher in FC (40.3) in year 2. It could be due to more competition among trees in FC than the other crown closure classes. Rotation periods of Taurus cedar are 120-140 years on good sites and 160-180 years on poor sites [1]. The crown closure in populations which were at end of rotation age seemed stable for cone productions in the present study. It is generally suggested that taller trees allocate much of their energy to vegetative growth rather than to reproductive growth [21]. Initial research results and observation suggested that 15-20 kg/ha of additional seeds was sufficient for shelter-wood areas, but higher rates 20-30 kg/ha is necessary for strip clear-cut areas of the species [1]. The results of the study showed that additional cone/seed could be obtained by CCCs managements.
Cone production gives an estimate of seed yield in the different crown management practices. The large difference in cone productivity among individuals within each crown closure class (Table 2) shows the importance of individual selection of seed tree for regeneration and other silvicultural practices. Statistically significant (p≤0.05) difference in average cone production across years (26.2/tree and 31.4), and among CCCs within second year emphasized the need for considering the crown closure management practice in forestry for seed harvest, natural regeneration and other forestry practices. The results were well accordance with differences in cone numbers among aspects and years reported by Yazici and Bilir [12], and for strobili productions among years and populations by Bilir and Kang [9,10] in the species. Large differences among individuals within a population and among populations in fecundity and reproductive characteristics were also reported in natural stands of different forest tree species [eg., 14,15,16,22,23] and in Taurus cedar [eg., 9,10,12]. Odabasi [4] reported 144 seeds per cone in Taurus cedar which works out to 750 additional seeds from per tree in year 2 than in year 1. However, cone production which gives an estimate of seed yield could be affected by many biotic and environmental factors. Growth characteristics did not impact significantly (p>0.05) on cone productions in CCCs for both years opposite to significant (p≤0.05) correlations between diameter at breast height for both years (r= 0.15 and 0.17) in pooled populations. Correlation analysis showed significant (p≤0.05) positive correlation between cone production and crown closure during both years (r= 0.29). Similar observations were also made by Odabasi [4] in Cedrus libani and Eler [13] in Pinus brutia. Positive correlation in cone production (r=0.22) and crown closure across years observed in the study is in line with the findings on cone [12] and strobilus [10] production in the species, and other forest tree species [eg., 16,23]. The results highlight importance of considering fertility and crown closure in forest management. Climatic data recorded at the studied location (Figure 1 and Figure 2) are similar to climate of the natural distribution of the species reported by Boydak [1]. Eler [13] found that age, elevation and crown closure were important factors in seed yield in Pinus brutia. Differences in age and environmental variation, mainly in soil properties, may have influenced the observed variation in fruiting and seed set within each population in the natural forests [15]. Altitudinal differences were reported in strobili production in Pinus brutia [16], cone production among populations in P. nigra [8], and reproductive characteristics in other forest tree species [23]. In a results of recent study indicated a higher influence of the year and probably of the sensitivity to climate, and less variability between clones for cone production in a Silver fir (Abies alba Mill.) seed orchard [24]. They [24] also reported that repeatability of mean annual clone production suggests moderate continuity of cone crops in the production rank of individual clones, while heritability was under genetic control. However, crown closure is a silviculturally manageable.
Coefficient of variation in cone production ranged from 51.4% (DF) to 74.6% (FC) in second year, while it was varied between 32.8% (DF) and 41.0% (FC) in first year (Table 2). Lower cone production in year 1 than in year 2 is contradictory to an earlier estimation of strobilus production in the species [10]. Low coefficient of variation (CV) is desirable for higher gene diversity for forest managers. It is suggested as a rough generalized heuristic rule that CV should be lower than 140% (Ψ≤3) for natural populations [25], and the crown closure classes considered in the current study had lower CVs.
4.2. Fertility variation, effective number of parents and gene diversity
The fertility variation (Ψ) increased with crown closure that will reduce gene diversity in second year, while it was similar in different crown closure classes in the first year (Table 3 and Figure 5). It is suggested as a thumb rule that, Ψ should be lower than 3 in natural stands and less than 2 in managed populations like seed orchards [14]. The proportion of the numbers of cones and fertility variation estimated in the current study populations is acceptable especially with respect to the highest Ψ value of FC (1.56) in year 2 which is typical of natural populations [14,21] (Table 3). Fertility variation (cone production) of 1.55, 3.05, and 1.64 in pooled stands aspectual for three consecutive years [12], 1.43 in a natural population [11], 1.57 and 1.26 in two natural populations [9] were reported the species. Fertility with respect to strobili production varied between 1.22 and 1.91 in three natural populations of the species for three consecutive years [10]. The results have emphasized the importance of estimation of fertility and reproductive characteristics across years. Data collection on cone production was easier, cheaper and more accurate than that of other reproductive characteristics such as strobilus or pollen count as also suggested by Bilir and Kang [9]. The tree keeps the cone in longer period than strobili or pollen in a year.
Ψ could be also related by standard deviation (S) and average (x̄) of cone production based on CV given in equation 1 by Kang and Lindgren [26], and Bilir [7]. It showed the importance of balancing between amount of cone production and its variation among individual trees for lower fertility variation and higher gene diversity.
The effective number of parents (Np) decreased from DF to FC in both years. Effective number of parents (Np) was related by gene diversity (GD) as: GD=1-0.5/Np by Kang and Lindgren [26]. Estimated loss of gene diversity was 0.003 between crown closures in second year. The highest loss of gene diversity was in FC (0.004) (Table 3). However, it could be balanced to increase gene diversity by natural regeneration, forest tending or other forestry practices (i.e., mixed seed from different crown closure classes and years. For instance, gene diversity increased from seed crop of year 2 by mixing seed from both years (Table 3). Forestry practices such as seed cutting in natural regeneration, and seed harvesting were preferred generally in good seed year. However, poor seed year could be also used for seed collection if the right strategies are used to increase the genetic diversity using suitable management strategies. The results could be used in establishment of seed collection areas (e.g., spacing). It showed mixed cones should be harvested from different crown closure classes and years to increase gene diversity in natural regeneration or plantation forestry.
5. Conclusions
Differences in cone production and fertility among crown closure classes emphasized the importance of crown closure classes and which can be managed by forestry practices (i.e., natural regeneration or plantation forestry). Gene diversity could be enhanced by harvesting cones from different crown closure classes and years. Results of the study could be used in managing gene conservation and seed collection areas, and other forestry operations such as natural regeneration of the species. Impact of crown closure classes on cone production was investigated in the present study. New studies should be carried out to estimate impact of crown closure classes in quality and quantity of seed production.
Author Contributions
NY conducted data collection with the participation of NB and performed the bibliographic review. NB and NY analyzed data and organized the manuscript. NY prepared figures and tables. NB edited the final version of the manuscript. All authors participated in the discussion and approved the submitted version.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Authors thank the graduate students for their help in data collection, and the regional forest directorate for administrative support. We also thank the anonymous reviewers who made valuable comments which helped to improve the manuscript.
References
- Boydak, M. Regeneration of Lebanon cedar (Cedrus libani A. Rich.) on karstic lands in Turkey. For Ecol Manag. 2003, 178, 231–243. [Google Scholar] [CrossRef]
- Anonymous. Forest inventory of Turkey, Ankara, 2022. https://www.ogm.gov.tr.
- Koski, V.; Antola, J. National tree breeding and seed production programme for Turkey 1994-2003. The Research Directorate of Forest Tree Seeds and Tree Breeding, Ankara, 1993. https://ortohum.ogm.gov.tr.
- Odabasi, T. Research on cone and seed characters of Cedrus libani. General Directorate of Forestry. Ankara, Turkey, 1990. https://www.ogm.gov.tr.
- Griffin, A.R. (1982). Clonal variation in radiata pine seed orchards. I. Some flowering, cone and seed production traits. Aust. For. Res. 1982, 12, 295–302. http://jkv.50megs.com/afr.html.
- Savolainen, O.; Karkkainen, K.; Harju, A.; Nikkanen, T.; Rusanen, M. Fertility variation in Pinus sylvestris: a test of sexual allocation theory. American Journal of Botany 1993, 80, 1016–1020. [Google Scholar] [CrossRef]
- Bilir, N. Fertility variation in wild rose (Rosa canina) over habitat classes. International Journal of Agriculture and Biology 2011, 13, 110–114. [Google Scholar]
- Bilir, N.; Catal, Y.; Tekocak, S.; Cercioglu, M. Fertility variation in endemic populations of Ehrami black pine (Pinus nigra Arnold. subsp. Pallasiana var. pyramidata). J. For. Res. 2017, 28, 683–686. [Google Scholar] [CrossRef]
- Bilir, N.; Kang, K.S. Estimation of fertility variation by strobili and cone productions in Taurus cedar (Cedrus libani A. Rich.) populations. IUFRO Forest Tree Breeding Conference August 25-29, 2014, Prague, Czech Republic, p.42. http://www.iufrobreeding2014.org/.
- Bilir, N.; Kang, K.S. Fertility variation, seed collection and gene diversity in natural stands of Taurus cedar (Cedrus libani). European Journal of Forest Research 2021, 40, 199–208. [Google Scholar] [CrossRef]
- Bilir, N.; Ozel, H.B. Fertility variation in a natural stand of Taurus cedar (Cedrus libani A. Rich.). International Forestry and Environment Symposium (IFES), 7-10 November, 2017, Trabzon. http://www.ktu.edu.tr/ifes2017.
- Yazici, N.; Bilir, N. Aspectual fertility variation and its effect on gene diversity of seeds in natural stands of Taurus cedar (Cedrus libani A. Rich.). IJ. Genomics 2017, 2960624. [CrossRef]
- Eler, U. Seed yield in Calabrian cluster pine (Pinus brutia Ten.) by age. Forest Research Institute, Technical Bulletin No. 225, Antalya, 1990, 78 pp. https://centralanatolia.ogm.gov.tr.
- Kang, K.S.; Bila, A.D.; Harju, A.M.; Lindgren, D. Fertility variation in forest tree populations. Forestry 2003, 76, 329–344. [Google Scholar] [CrossRef]
- Bila, A.D.; Lindgren, D. Fertility variation in Milletias thuhlmannii, Brachystegia spiciformis, Brachystegia bohemii and Leucaena leucocephala and its effects on relatedness in seeds. Forest Genetics 1998, 5, 119–129. [Google Scholar]
- Bilir, N.; Kang, K.S.; Lindgren, D. Fertility variation in six populations of Brutian pine (Pinus brutia Ten.) over altitudinal ranges. Euphytica, 2005, 141, 163–168. [CrossRef]
- Saatcioglu, F. Silviculture-I. Forestry Faculty of Istanbul University press, Istanbul, 1976. https://cerrahpasa.iuc.edu.tr/.
- SAS. Statistical Analysis System, SAS Institute, Inc. Cary., N.C., USA, 2004. https://www.sas.com.
- Rohlf, F.J.; Sokal, R.R. Statistical Tables. Macmillan. 1995. https://www.scirp.org.
- Kang, K.S.; Lindgren, D. Fertility variation among clones of Korean pine (Pinus koraiensis S. et Z.) and its implications on seed orchard management. Forest Genetics 1999, 6, 191–200. [Google Scholar]
- Kang, K.S. Genetic gain and gene diversity of seed orchard crops. Ph.D Thesis. Swedish University of Agricultural Science, Acta Universitatis Agriculturae Sueciae, Silvestria 187, Umeå, Sweden, 2001, pp.75. https://www.upsc.se.
- Shea, K. L. Effects of population structure and cone production on out crossing rates in Engelmann spruce and Subalpine fir. Evolution 41, 1987,124-136. [CrossRef]
- Kamalakannan, R.; Varghese, M.; Park, J.M.; Kwon, S.H.; Song, J.H.; Kang, K.S. Fertility variation and its impact on effective population size in seed stands of Tamarindus indica and Azadirachta indica. Silvae Genetica 2015, 64, 91–99. [Google Scholar] [CrossRef]
- Teodosiu, M.; Botezatu, A.; Ciocîrlan, E.; Mihai, G. Variation of cones production in a Silver fir (Abies alba Mill. ) clonal seed orchard. Forests 2023, 14, 17. [Google Scholar] [CrossRef]
- Kang, K.S.; Bilir, N. Seed orchards (Establishment, Management and Genetics). OGEM-VAK Press, Ankara, 2021. https://www.ogemvak.org.tr/.
- Kang, K.S.; Lindgren, D. Fertility variation and its effect on the relatedness of seeds in Pinus densiflora, Pinus thunbergii and Pinus koraiensis clonal seed orchards. Silvae Genetica 1998, 47, 196–201. [Google Scholar]
Figure 1.
Monthly maximum, minimum and average temperature of the studied location during study period.
Figure 1.
Monthly maximum, minimum and average temperature of the studied location during study period.
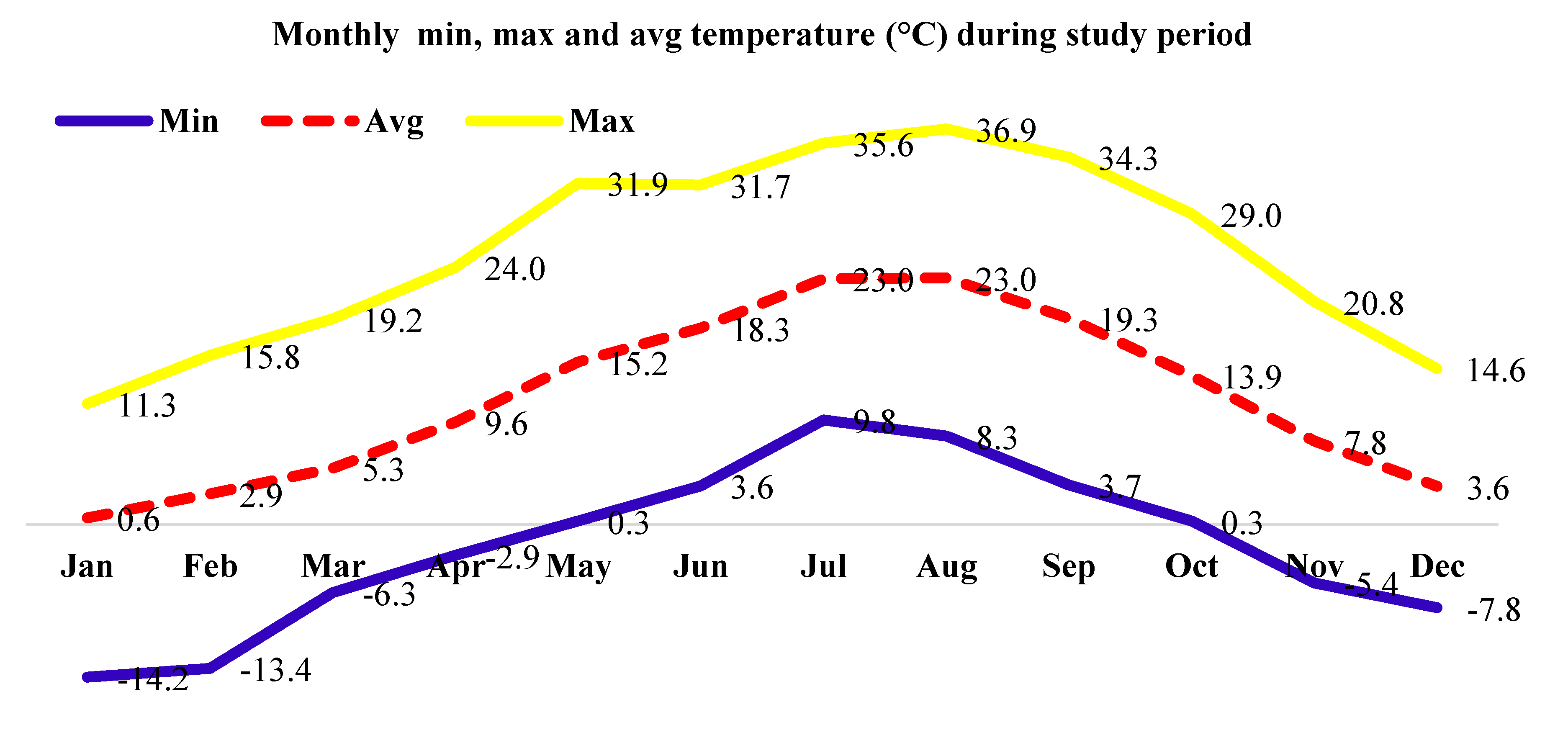
Figure 2.
Monthly maximum, minimum and average temperature of the studied location during study period.
Figure 2.
Monthly maximum, minimum and average temperature of the studied location during study period.
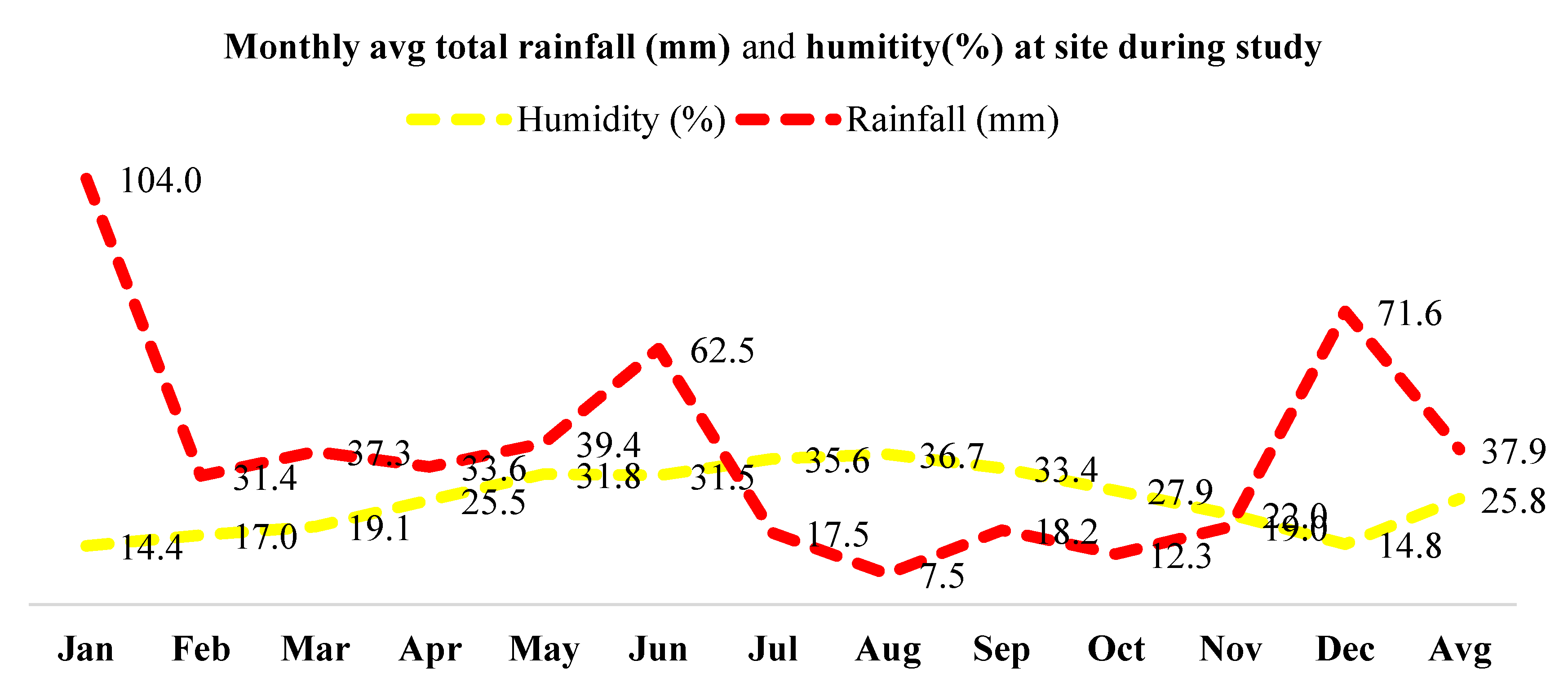
Figure 3.
Mature cones of Taurus cedar in studied population.

Figure 4.
Averages of cone production.
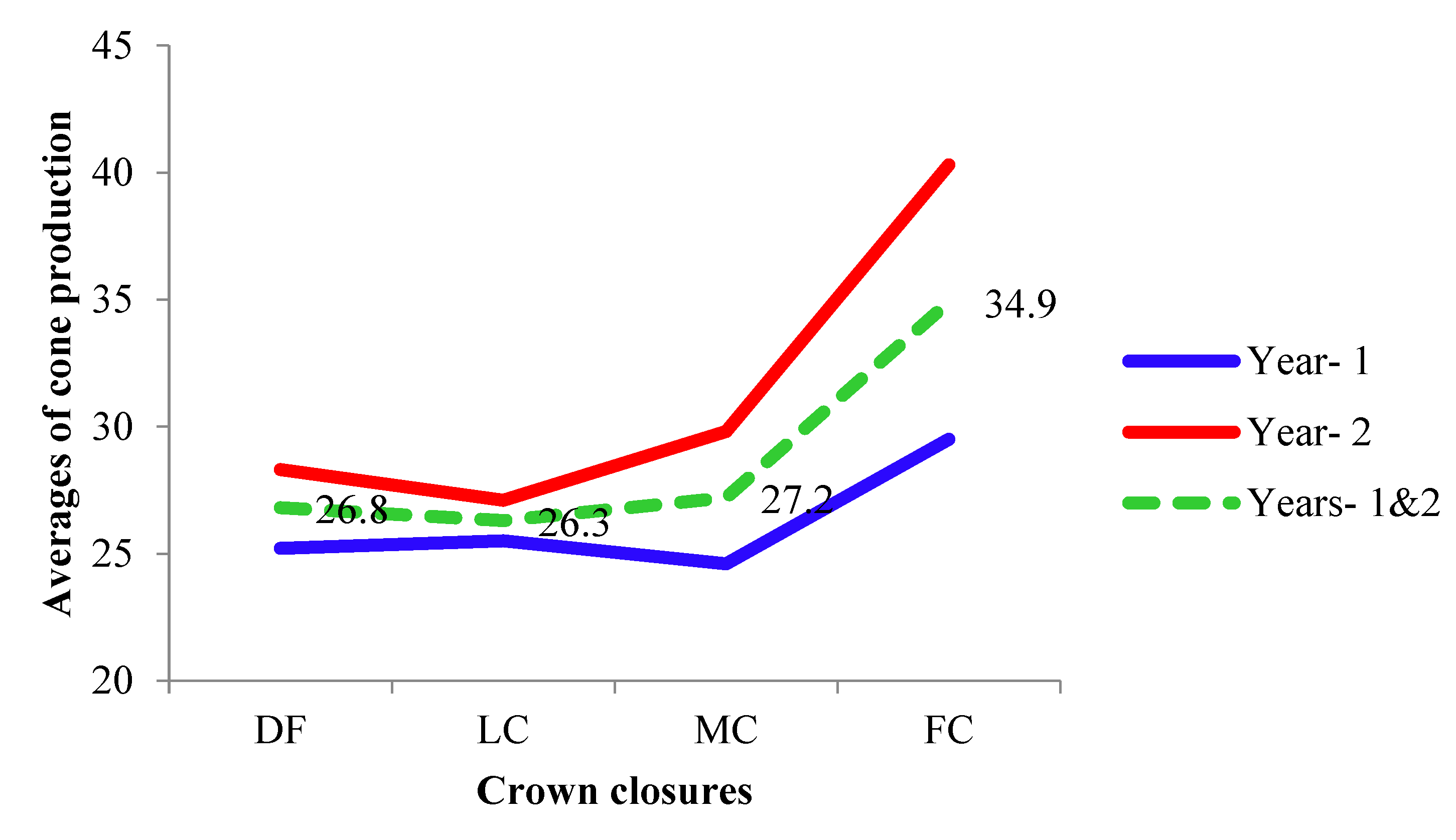
Figure 5.
Fertility variation for crown closures and years.
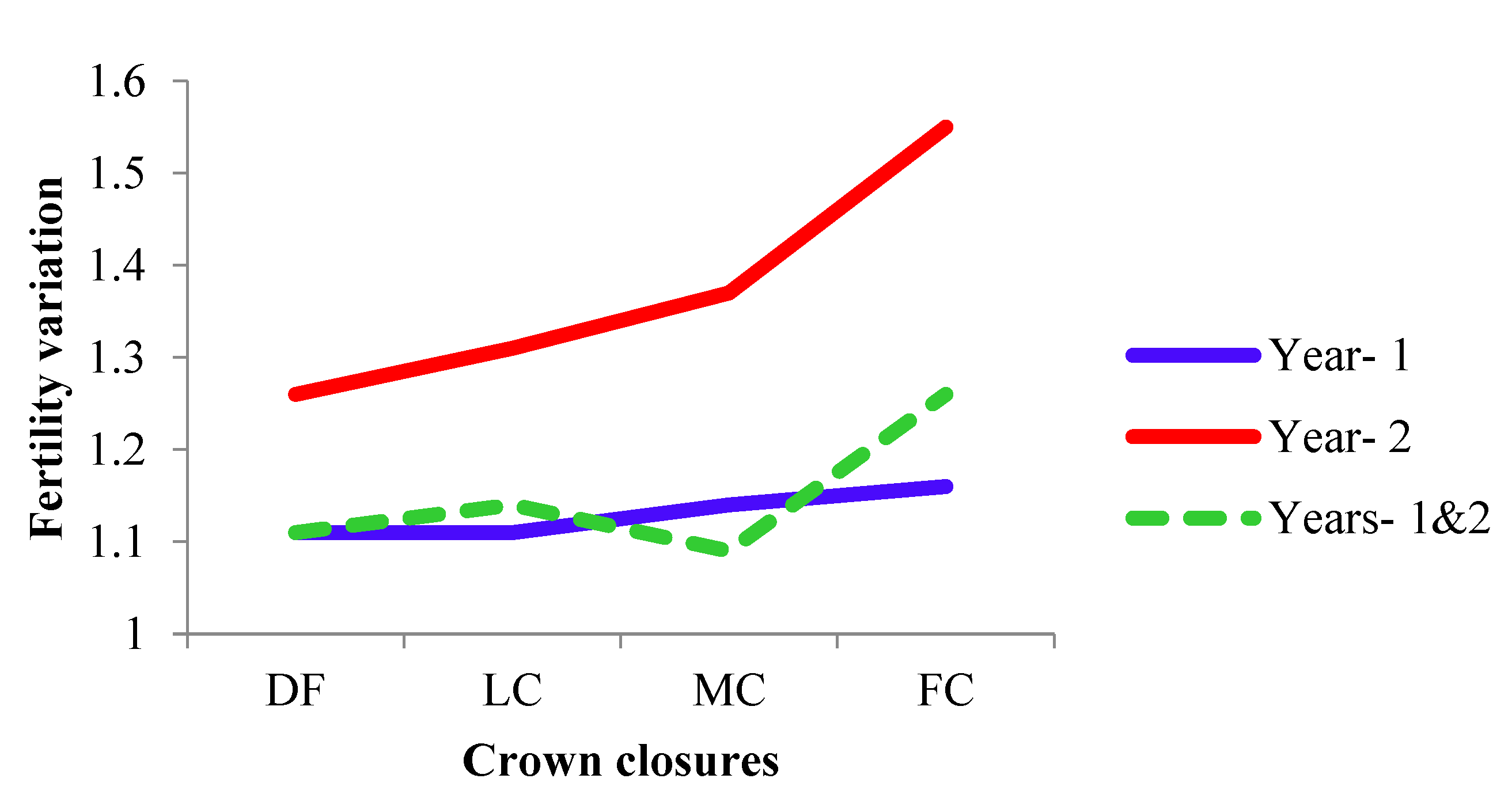
Figure 6.
Cumulative contribution of Parental-balance curves in different crown closure classes in year 1.
Figure 6.
Cumulative contribution of Parental-balance curves in different crown closure classes in year 1.
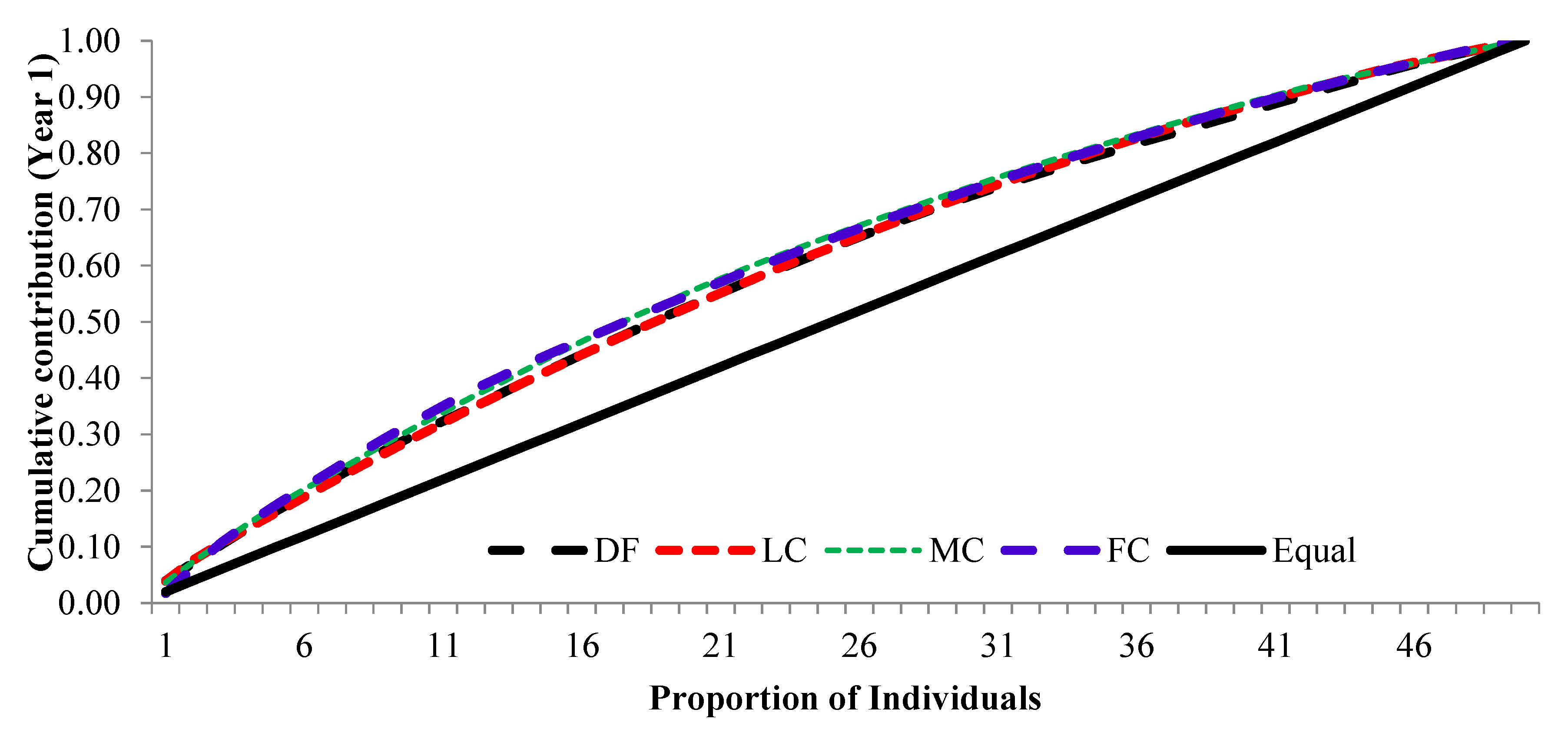
Figure 7.
Cumulative contribution of Parental-balance curves in different crown closure classes in year 2.
Figure 7.
Cumulative contribution of Parental-balance curves in different crown closure classes in year 2.
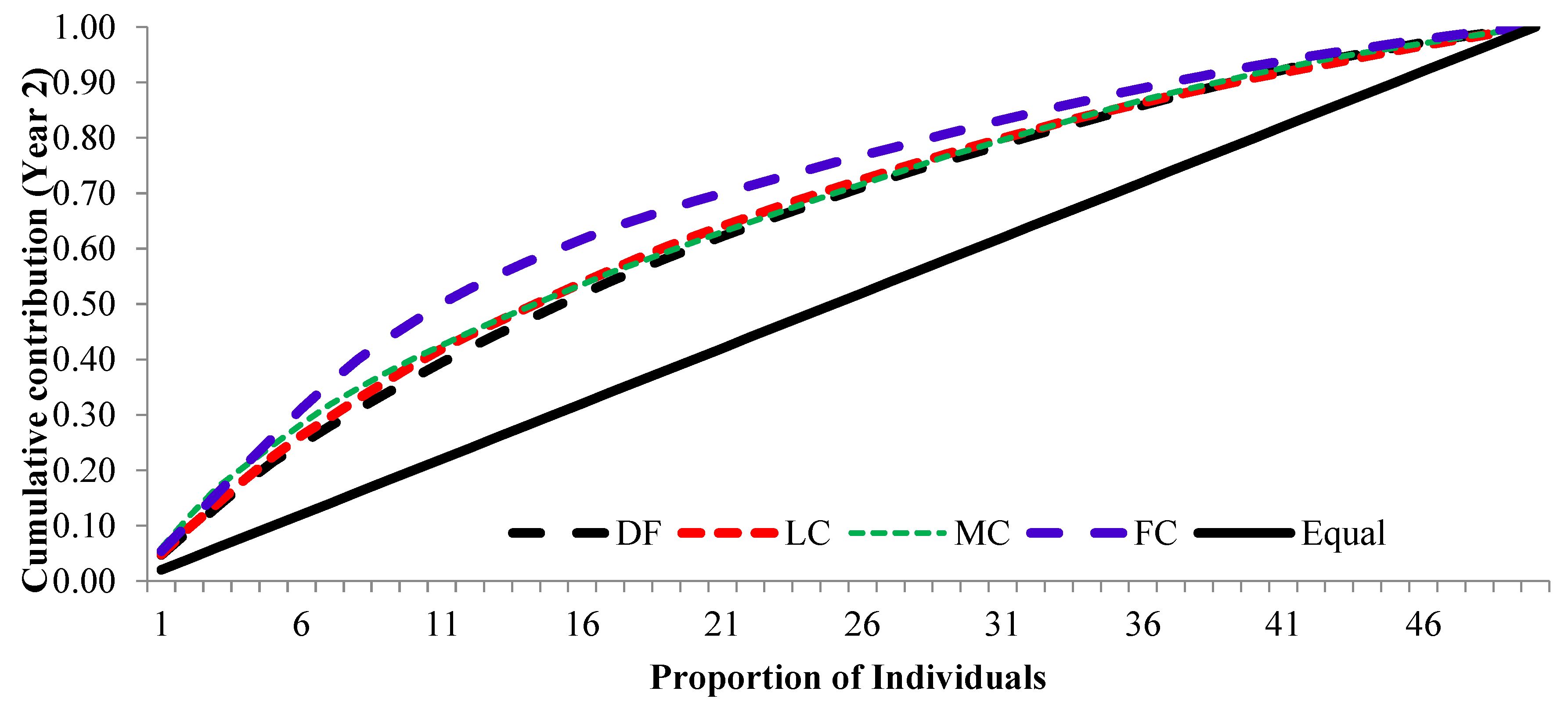
Figure 8.
Cumulative contribution of Parental-balance curves in different crown closure classes in years 1&2.
Figure 8.
Cumulative contribution of Parental-balance curves in different crown closure classes in years 1&2.
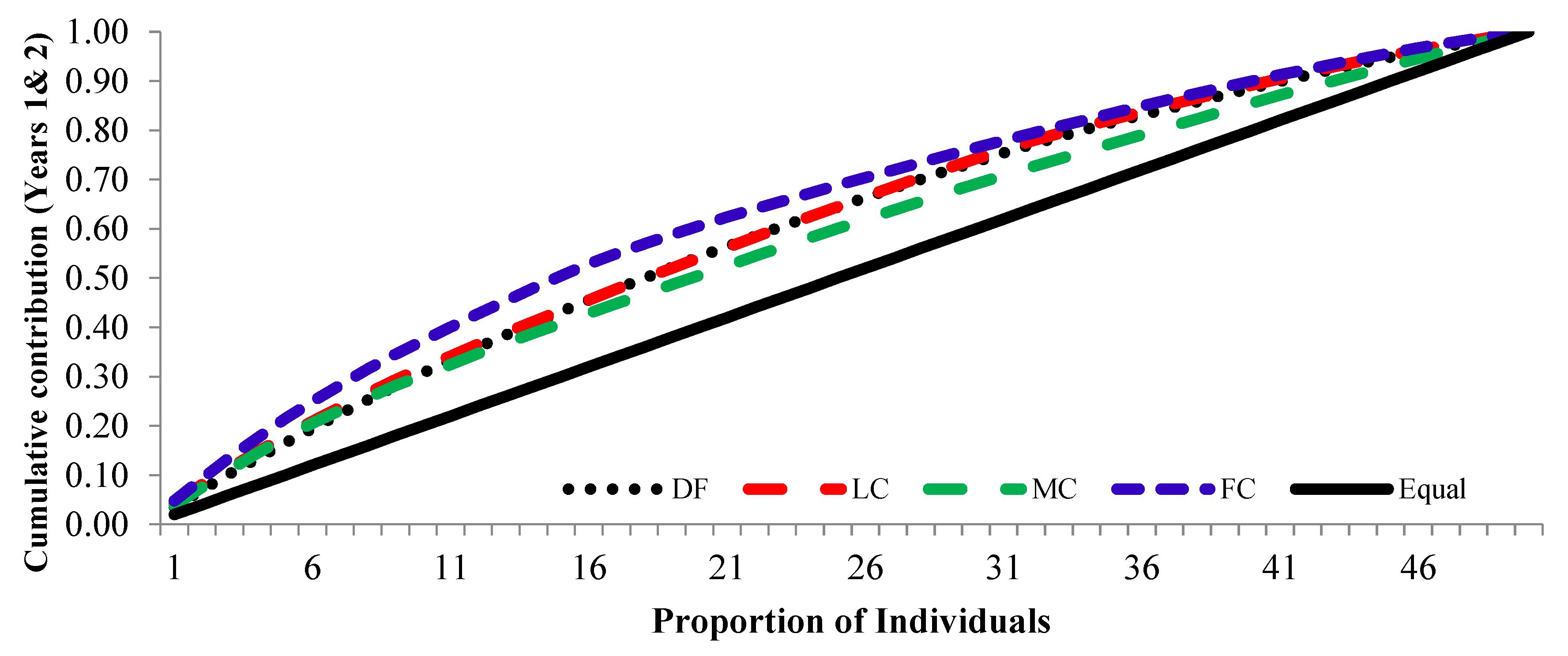

Table 1.
Growth details and altitude of the crown closure classes.
| CCCs | ||||
|---|---|---|---|---|
| Characteristics | DF | LC | MC | FC |
| H (m) | 20.3 | 21.2 | 20.0 | 21.6 |
| D0 (cm) | 64.1 | 74.9 | 72.1 | 89.4 |
| d1.30 (cm) | 53.9 | 63.4 | 62.4 | 78.2 |
| CD (m) | 7.3 | 8.9 | 8.4 | 10.0 |
| Age (year) | 121 | 131 | 127 | 139 |
| Altitude (m) | 1632 | 1630 | 1628 | 1628 |
Table 2.
Average (x̄), range and coefficient of variation (CV %) in numbers cone production for the crown closures and years.
Table 2.
Average (x̄), range and coefficient of variation (CV %) in numbers cone production for the crown closures and years.
| CCCs | DF | LC | MC | FC | ||||
|---|---|---|---|---|---|---|---|---|
| Years | Year 1* | Year 2 | Year 1 | Year 2 | Year 1 | Year 2 | Year 1 | Year 2 |
| x̄ | 25.2a | 28.3a | 25.5a | 27.1a | 24.6a | 29.8a | 29.5b | 40.3b |
| Range | 11-50 | 9-66 | 10-50 | 10-65 | 10-45 | 10-88 | 10-67 | 8-108 |
| CV | 32.8 | 51.4 | 33.5 | 56.0 | 37.5 | 61.1 | 41.0 | 74.6 |
*; The same letters showed not significantly different at p > 0.05 within year
Table 3.
Fertility variation (Ψ), effective number of parents (Np), relative effective number of parent (Nr) and gene diversity (GD) in the crown closure classes and years.
Table 3.
Fertility variation (Ψ), effective number of parents (Np), relative effective number of parent (Nr) and gene diversity (GD) in the crown closure classes and years.
| CCCs | DF | LC | MC | FC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Years | Year 1 | Year 2 | 1&2 | Year 1 | Year 2 | 1&2 | Year 1 | Year 2 | 1&2 | Year 1 | Year 2 | 1&2 |
| Ψ | 1.11 | 1.26 | 1.12 | 1.11 | 1.31 | 1.14 | 1.14 | 1.37 | 1.09 | 1.16 | 1.55 | 1.26 |
| Np | 45.23 | 39.71 | 44.60 | 45.04 | 38.25 | 43.73 | 43.95 | 36.62 | 45.79 | 42.92 | 32.35 | 39.60 |
| Nr* | 0.91 | 0.79 | 0.89 | 0.90 | 0.77 | 0.87 | 0.88 | 0.73 | 0.92 | 0.86 | 0.65 | 0.79 |
| GD | 0.989 | 0.987 | 0.989 | 0.989 | 0.987 | 0.989 | 0.989 | 0.986 | 0.989 | 0.988 | 0.984 | 0.988 |
*; Nr=Np/N.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



