
Submitted:
03 April 2023
Posted:
04 April 2023
You are already at the latest version
Abstract
The few studies dealing with leaf phenological responses to elevated nutrients in forest trees have given ambiguous results, i.e., while some reported delayed leaf-out and autumn leaf senescence, others reported advanced leaf phenology caused by increased nutrition. This study aimed to determine the effects of experimentally increased phosphorus (+P) on the leaf phenologies of two juvenile provenances of common beech and sessile oak. Other objectives were to determine whether there are interspecies differences as well as intraspecies variations. Saplings were excavated in two mixed beech-oak stands and transplanted into four wooden boxes filled with a commercial soil substrate. Phosphorus fertilizer was added to two of the boxes, while the remaining boxes served as controls. Both species responded to +P treatment with advanced autumn leaf senescence in the first year of the experiment. Leaf senescence in common beech began significantly earlier, while in both species, the process was accelerated compared to that in the control. In the second year, the leaf senescence response to +P treatment was even more pronounced in both species. The +P effect on leafing phenology was absent in both common beech provenances and in an oak provenance. However, the other oak provenance showed advanced leafing, indicating the existence of intraspecies differences.
Keywords:
Fagus sylvatica
; Quercus petraea
; flushing
; leaf senescence
; nutrition
; phosphorus effect
; phenology shift
1. Introduction
The leaf phenology of deciduous trees (i.e., multiphase development of leaves from spring bud burst to autumn senescence and leaf abscission) determines the length of a growth period and is one of the main drivers of ecological processes in temperate zone ecosystems [1]. It is known that the basic (typical, common) environmental factors that, in interaction with the genome, regulate leaf phenology are temperatures in different parts of the year (chilling, forcing and autumn temperature) and photoperiod [2,3,4,5]. However, there are additional (atypical, i.e., less-studied) environmental factors that influence phenological processes. Some of these factors are drought stress [6], soil nutrient availability [7,8,9], pathogen infections [10], increased CO2 [11], etc. A plant reacts to these atypical environmental factors with shifts in the timing of phenological stages as well as with changes in its dynamics (e.g., acceleration of leafing) [6,12,13]. Relatively little is known about how and to what extent the addition of nutrients affects the leaf phenology of forest trees, and the results of the few available studies are ambiguous. For example, a delay [14,15] as well as an advance of autumn phenology [16,17] have been reported as effects of higher availability of nutrients in different tree species.
Spring leaf development is linked with the translocation of carbohydrates and mineral nutrients from storage tissues to apical meristems and/or young leaves in their early stages of development, while autumn leaf senescence is linked with the translocation of mineral nutrients from leaves to storage tissues [18,19,20]. Both require a supply of energy and appropriate enzymes that regulate translocation within the plant [21]. Phosphorus (P) is one of the most important macronutrients for plant productivity and response to various stressors, although it is often the least available of all macronutrients in soil. Moreover, P is crucial for the capture, storage, and transmission of solar energy and therefore necessary for normal functioning of all metabolic and physiological processes. P is part of the energy transport molecule structure (ATP, CTP, GTP, UTP) as well as nucleic acids (DNA and RNA) necessary for the synthesis of enzymes mediating the remobilization of carbohydrates and mineral nutrients [22]. Thus, it may have a significant influence on spring leaf development and/or autumn leaf senescence dynamics.
The influence of elevated P concentrations (as well as concentrations of other biogenic elements) in soil and/or vegetative organs on shifts in the timing of spring and autumn leaf phenology of forest trees has rarely been investigated [23]. However, this knowledge could be important for the survival of forest ecosystems in the context of climate change consequences that make the natural regeneration of forest stands difficult [24,25]. Artificial reforestation practices in European forests are increasing, mostly due to various problems caused by climate change (extreme weather events, drought episodes, exotic pest outbreaks, etc.). Today, almost 30% of European forests have been artificially reforested [26]. For this reason, the need for the production of suitable forest reproductive material (FRM) is growing. The FRMs of common beech and sessile oak have significant shares in this process, especially in Central Europe [27]. Therefore, clonal seed orchards (CSOs) of these species are being established in an increasing number of EU countries (https://ec.europa.eu/forematis/). Effective production of quality FRM in CSOs and forest nurseries requires intensive management, i.e., implementation of the best agrotechnical practices, which includes fertilization. By applying fertilizers that most often contain essential biogenic elements in different proportions (including P), the concentrations of these elements in the soil increases, which ultimately results in an increased concentration of biogenic elements in plant organs [28,29,30]. This has a positive impact on the survival and later growth of the plants in natural habitats [31,32,33,34,35]. However, the influence of elevated concentrations of biogenic elements on shifts in forest tree spring leafing and/or autumn leaf senescence timing is still quite unknown. Leaf phenology shifts caused by increased concentrations of biogenic elements in vegetative organs together with the frequent occurrence of late spring and/or early autumn frosts could adversely affect seedling survival [4,5,36]. Most studies address the impacts of P deficiency on forest trees, emphasizing the negative consequences for growth and physiological processes [37]. Due to the results of such research, it is easy to conclude that ample nutrition should provide only benefits to trees. However, too much P in cells and tissues has a harmful effect on plants, which means that it is necessary to maintain a balance of nutrients [38].
Overall, further research is needed to determine the specific effects of elevated P on leaf phenology in different tree species and under different environmental conditions. Common beech and sessile oak are both deciduous broadleaf tree species, yet they differ significantly in their leaf phenology patterns. It is common for oaks to flush their leaves earlier in the spring than beeches. This is likely since oaks are more tolerant to cold temperatures, which allows them to begin their growing cycle earlier than beeches. Research considering comparison of leaf phenological responses to various environmental factors between these forest species are lacking, although they often form valuable mixed stands all over Europe [39,40]. Due to differences in their genetics and physiological processes, they may respond differently to elevated P, as well. It may be assumed that beech would respond faster than oak due to its shallower rooting system that allows faster P uptake upon fertilisation. Additionally, factors such as soil type, and climate can all influence how different provenances within species respond to elevated phosphorus levels, i.e., within-species variations may be expected. For example, soils in drier habitats may not provide the same amount of phosphorus as soils in wetter habitats. This could lead to different adaptation strategies of provenances and consequently their various responses to experimentally elevated P-levels.
Therefore, the main objectives of this research were (1) to determine the leaf phenology responses of juvenile common beech and sessile oak to increased phosphorus concentrations available in a growth substrate, (2) to determine whether there are differences in the responses between these two cohabiting but phenologicaly different species, and (3) to determine whether there are intraspecies variations i.e., are there variations between the provenances originating from different habitats.
2. Materials and Methods
2.1. Plant Material
Natural saplings of sessile oak and common beech were sampled at the beginning of March 2021 in two mixed stands dominated by these species. Saplings, 3-4 years old, were dug up from an area of approximately 1 ha, ensuring that they were evenly distributed throughout the stand (to avoid close relatedness and ensure proper representativeness of the stand) and that their roots were damaged as little as possible. Plants that were of uniform height were selected. 160 plants of each species were excavated in both stands (i.e., 160 saplings × 2 species × 2 stands = 640 saplings in total). The saplings were labelled and placed in moist sand (in PVC containers). On the same day, the saplings were transported to the location of the future experimental trial, where they were kept in shade until transplanting.
The selected stands differed from each other regarding prevailing habitat conditions (Table 1). The first is located within the city of Karlovac (Karlovac stand (KA); 45.4661986 N; 15.5219786 E; 170–185 m a.s.l.). It is a mixed stand of beech and oak, approximately 100 years old, with north‒northeast exposure, on deep soil. We considered this stand to be a representative of wetter and warmer habitats due to the higher average amount of precipitation, exposure and soil depth as well as the higher average air temperature at this location (Table 1). The second stand is located next to the village of Gornji Slatinik along a road that connects the towns of Našice and Slavonski Brod (Slavonski Brod stand (SB); 45.2733637 N; 17.9727061 E; 230–255 m a.s.l.). This is also a mixed common beech and sessile oak stand, approximately 105 years old, with south‒southeast exposure, on shallow soil. We considered this provenance to be representative of drier colder habitats due to the lower average precipitation, exposure and shallower soil and lower average air temperature at this location (Table 1).
In the following text, the plants belonging to the sampled stands, i.e., provenances, will be labeled as follows:
1) Fs_KA - Fagus sylvatica provenance from the KA stand
2) Fs_SB - Fagus sylvatica provenance from the SB stand
3) Qp_KA - Quercus petraea provenance from the KA stand
4) Qp_SB - Quercus petraea provenance from the SB stand
2.2. The Experimental Trial
In mid-March 2021, the saplings were transplanted to 4 wooden boxes (W 155 x L 255 x D 80 cm (3.2 m3) located at the nursery of the Faculty of Forestry and Wood Technology in Zagreb, Croatia (45.820642 N; 16.022936 E). Each box was filled with commercial soil substrate Klasmann-Deilmann TS 3 (pH 6.0; 0.16 g/l P2O5). In each box, a total of 100 plants were planted in a random arrangement (spacing: 20 × 18 cm), with 25 plants per beech and oak provenance from the KA stand (Fs_KA and Qp_KA) and 25 plants per beech and oak provenance from the SB stand (Fs_SB and Qp_SB). Then, 1200 g of Triplex mineral fertilizer (45% P2O5) was added to the two boxes. Thus, the plants in two boxes (200 individuals in total) were exposed to an increased amount of phosphorus in the growth substrate (treatment +P), while in the remaining two boxes, the plants were exposed to the normal phosphorus content in the substrate (i.e., control - C).
2.3. Chemical Analyses
Soil samples from the original stands were extracted at the beginning of March 2021. The sampling methodology is described by Sever et al. [42]. There were no significant differences between the provenances regarding the chemical features of their soils. The total stocks of N, P and K at soil depths of 0-15 cm and 16-30 cm were similar, i.e., without significant differences between the provenances.
Sampling of the growth substrate and its chemical analysis were carried out during the establishment of the experimental trial but before adding phosphorus to the +P treatment boxes. The second series of chemical analyses of the substrate was performed in mid-September 2021, i.e., 6 months after the addition of mineral fertilizer. One composite (average) sample was collected from each box. Each composite sample was formed from nine individual samples collected in a diagonal (X) arrangement. Sampling of the substrate was carried out using a pedological probe at a depth of 0-30 cm. The samples were subjected to soluble component extraction by mixing with deionized water in a 1:2 ratio. The material was mixed for 1 hour using a rotating mixer. Then, the suspension was filtered through filter paper, and the clear filtrate was subjected to an analytical procedure aiming to determine the pH value, specific conductivity, concentrations of nitrate (NO3-), ammonia (NH4+), total nitrogen (N total), total phosphorus (P3-), potassium (K+), calcium (Ca2+), magnesium (Mg2+), chlorine (Cl-), and sodium (Na+). The applied methodology for chemical analyses of the substrate is described in more detail in [43].
Sampling of leaves in the experimental trial was performed twice, in mid-September 2021 and mid-May 2022. From the top of each plant, in all four boxes, three leaves were sampled. Three composite (average) samples were formed for each species provenance. Each sample consisted of a total of 25 randomly selected leaves from 25 plants per species provenance in a box.
The composite leaf samples were subjected to chemical analyses to determine their mineral nutrient contents. The samples were dried at 105 °C to a constant mass, minced and homogenized. The concentration of N was determined using the Kjeldahl method. Digestion was performed with concentrated nitric acid (HNO3) and perchloric acid (HClO4). The concentration of P was determined spectrophotometrically. The concentration of K was determined by a flame photometer. The concentrations of calcium (Ca) and magnesium (Mg) were determined by atomic absorption spectrophotometry. All analyses were carried out according to standardized international protocols [44].
2.4. Leaf Phenology Scoring
Spring leafing and autumn leaf senescence were scored in 2021 and 2022. Leaf phenological phases were scored once or twice a week (during the process) on all plants in the trial using a 1–7 ordinal scale: 1—bud scales completely closed, dormant buds; 2—buds swelling, scales slightly spaced; 3—bud burst, green leaf tips visible; 4—folded leaves visible. 5—leaves unfolding but not yet flattened, small. 6—leaves still relatively small but with flattened blades, blade edges bent downward, withered, lighter green or reddish; 7—leaves appear developed, larger but more tender than fully developed leaves and lighter green or reddish.
Autumn leaf phenological phases were scored (during the process) once a week in 2021 and twice a week in 2022 using a 0–5 ordinal scale: 0—leaves completely green with no visible discoloration; 1—up to 25% of plant leaves show discoloration; 2—up to 50% of plant leaves show discoloration; 3—more than 50% of plant leaves show discoloration; 4—more than 75% of plant leaves show discoloration; 5—leaves shed. All phenological scorings were done by one experienced person.
The day of the year when a plant reached leaf-out phase 3 was taken as its bud burst date, and the day of the year when a plant reached autumn leaf phenological phase 3 was taken as its autumn senescence date.
It should be noted that late spring frost affected the plants in the experimental trial from April 6th to 8th, 2021. At that time, many oak saplings (as well as few common beech saplings) had already started leafing out. Air temperature dropped to a minimum of -8 °C on the morning of April 6th, and then for three days repeatedly rose and fell to negative values. All the leaves on the plants that had started leafing out were destroyed, regardless of which phenophase they were in during that period. However, none of the plants died during this period, and they leafed out again approximately two weeks later.
2.5. Statistical Analyses
Assumptions of residual normality and variance homogeneity were tested by using the Shapiro–Wilk test and Levine’s test [45] with the GLM and UNIVARIATE procedures in SAS Studio software, a free version of SAS OnDemand for Academics by SAS Institute Inc., Cary, NC, USA. Residuals were plotted as a function of fitted values to test for variance homogeneity, and the distribution of residuals was also tested. Since data did not meet the criteria of normal distribution and homogeneity of variances (raw phenological data and mineral nutrition contents of the leaf samples) the nonparametric Kruskal‒Wallis and Mann–Whitney U tests were applied. These tests were used to assess the significance (p<0.05) of the +P treatment on the provenance leaf phenology phases on particular days of the year. Additionally, the U test was used to test the significance of leaf nutrition content differences between +P treatment and control plants. The nonparametric tests were performed using RStudio, version 1.2.5001 (© 2009-2019 RStudio, Inc).
3. Results
3.1. Substrate Nutrient Content Differences between the Treatments
The chemical traits of the substrate between the +P treatment and control differed significantly only regarding the concentration of total P. The concentrations of the other nutrients and pH value in both treatments were similar (Table 2).
3.2. Leaf Mineral Nutrition Variations
According to Mellert and Göttlein [46], the following value ranges in leaf dry matter indicate normal nutrition for common beech and sessile oak: P (1.2–1.9 mg g-1 and 1.4–2.1 mg g-1, respectively), N (18.7–23.2 mg g-1 and 19.8–26.8 mg g-1, respectively), K (6.1–9.7 mg g-1 and 7.2–11.4 mg g-1, respectively), Ca (6.7–14.0 mg g-1 and 5.3–10.2 mg g-1, respectively), and Mg (1.1–2.3 mg g-1 and 1.2–2.4 mg g-1, respectively).
Comparing the measured leaf dry matter nutrient contents from September 2021 (Table 3) with the abovementioned reference values, it is evident that the Fs_SB provenance had a P content in the upper part of the normal range in the +P treatment, while the Fs_KA provenance had a P surplus. The leaf P content of the control plants was within the lower normal limits. It should be emphasized that both common beech provenances had a significantly higher leaf P content in the +P treatment than in the control plants. The sessile oak provenances had a much lower leaf dry matter P content than the common beech provenances in the +P treatment, although it was still in the normal range. The control plants of both oak provenances showed latent P deficiency. Again, it should be emphasized that both oak provenances in the +P treatment had a significantly higher leaf P contents than the control plants.
For the other nutrients, the beech provenances were in the normal range, mostly without significant differences between plants in the +P treatment and control plants. However, within the Fs_KA provenance, the control plants had significantly higher nitrogen and potassium contents than the plants in the +P treatment. On the other hand, the Fs_SB provenance in the +P treatment had a significantly higher magnesium contents than the control plants.
The average contents of the other nutrients in the oak provenances were in the normal range (except for nitrogen, of which there was a surplus), without significant differences between the plants in the +P treatment and control plants. However, the control plants of the Qp_KA provenance had significantly higher contents of calcium and magnesium than the plants of this provenance in the +P treatment.
A significant P increase in the oak provenances in the +P treatment was observed from leaf samples taken in May 2022, while the P contents in leaves of common beech provenances remained at approximately the same level as in the previous year (Table 3). The P contents in the leaves from the plants in the +P treatment were higher than those in the control plants in all the provenances, although the difference was not significant in the case of the Fs_KA provenance.
The values for the other nutrients in the beech provenances were in the normal range, mostly without significant differences between the plants in the +P treatment and control. The exception was the higher mean potassium content in the control plants of the Fs_KA provenance (Table 3). The average contents of the other nutrients were in the normal range for the oak provenances, with no significant differences between the plants in the +P treatment and control. It is interesting that nitrogen content fell compared to that in the previous year, while the calcium and magnesium values increased above the upper limit of the normal range (Table 3).
3.3. Variations Caused by the Elevated Phosphorus Treatment (+P)
As expected, the effect of increased phosphorus on spring phenology in 2021 was not recorded (Figure 1a). However, a statistically significant difference between the treatments was shown in autumn leaf senescence (Figure 1b). Namely, the plants in the +P treatment showed advanced leaf senescence dates, on average.
With more detailed analyses, it was possible to see that both species responded similarly, i.e., with significantly advanced autumn leaf phenology of plants in the +P treatment (Figure 2). Thus, the Fs_SB provenance in the +P treatment showed an average of more than 50% dead leaves on day 305; this threshold was reached three days later in the control (Figure 2a). The differences in average leaf senescence dynamics between the +P treatment and control were even more pronounced for the Fs_KA provenance (307 DoY and 311 DoY, respectively) (Figure 2b). For both beech provenances, the visible symptoms of leaf senescence started earlier and proceeded faster in the +P treatment. The dynamics of autumn leaf phenology were significantly different between the treatments (+P vs. control) for the oak provenances as well (Figure 2c,d), although not as intensely.
The effect of elevated phosphorus on leaf flushing phenology in the following year was not recorded for the common beech provenances, i.e., the leafing dynamics of plants in the +P treatment plants were not significantly different from those in the control (Figure 3a,b). However, advanced leaf flushing dynamics in the +P treatment were recorded for the oak provenance Qp_KA (Figure 3d), while there was no significant effect of the +P treatment for the provenance Qp_SB (Figure 3c).
The effect of increased phosphorus on autumn leaf phenology was even more pronounced in the fall of 2022 than in the year before (see Figure 2 and Figure 4). Significantly advanced leaf senescence in the +P treatment was recorded for both beech provenances as well as for the oak provenance Qp_SB. Plants of the Qp_KA provenance in the +P treatment also showed advanced leaf senescence compared to the control plants, although the difference was not statistically significant (Figure 4).
4. Discussion
Plants growing in phosphorus-enriched substrate showed earlier autumn leaf senescence in the first experimental year (Figure 1b). A closer analysis revealed that both beech provenances reacted to phosphorus addition with advanced autumn leaf phenology (Figure 2a,b). Both beech provenances started leafing out significantly earlier in the +P treatment, and the process itself was accelerated. The oak provenances also responded with advanced leaf senescence, although the difference was less pronounced than it was for the beech provenances (Figure 2c,d). Autumn leaf phenology between the +P treatment and the control differed even more in the second year in both species (Figure 4).
Autumn leaf phenology is triggered and its dynamics is shaped by photoperiod [47,48], early autumn temperatures [49,50], available soil moisture [51], and plant nutritional status [15,52]. Phosphorus deficiency induces leaf senescence, i.e., premature leaf discoloration is a typical symptom of P deficiency [53]. However, the influence of various nutrient contents on autumn leaf phenology varies significantly among tree species [15]. Such variations can also be observed in other plant species. For example, phosphorus addition had no effect on soybean (Glycine max (L.) Merr.) autumn leaf phenology [54]. On the other hand, a lack of phosphorus in soil delayed senescence of lower-stem leaves in corn (Zea mays L.), although the authors pointed out that this effect was not confirmed for upper-stem leaves [55]. Delayed autumn leaf phenology due to increased nutrition (a combination of nitrogen, phosphorus, and potassium) was reported by Fu et al. [15] for common beech. Our study, in contrast, showed advanced leaf senescence in response to increased phosphorus nutrition. The opposing autumn phenology alterations (i.e., advanced or delayed) observed in these two studies can be explained by the fact that the increased contents of several macronutrients (N, P and K) can have different effects on autumn phenology than the content of only one nutrient (P). Such differences were observed in larch (Larix gmelinii var. principis-rupprechtii (Mayr) Pilg.) between a combined nutrient increase treatment (N + P) and separate nutrient increase treatments (N or P) [14]. Opposing phenological shifts may also be explained by the different time periods in which fertilizer was applied. Namely, Fu et al. fertilized experimental plants at the end of May, while we added phosphorus immediately after planting (in March). However, since Zavišić and Polle [56] indicated that significant P uptake from the soil takes place at the end of summer and in autumn, this assumption seems less probable. It is more likely that the interaction effect of nitrogen and phosphorus reported by Fu et al. resulted in an autumn leaf phenological shift that was opposite to that observed in our study.
Advanced leaf senescence as a response to increased sunlight, and increased CO2 was reported by Zani et al. [16] for common beech, common birch (Betula pendula Roth), and Japanese spiraea (Spiraea japonica L.f.). The authors concluded that increased productivity, i.e., photosynthetic activity (due to higher sunlight or increased CO2), results in advanced autumn leaf senescence (i.e., the sink limitation hypothesis). Advanced oak autumn leaf phenology due to elevated CO2 was also reported [17]. Although Zani et al. did not specifically test the increased phosphorus effect on autumn phenology, in our case it can be assumed that plant productivity may have been increased due to greater P availability. Therefore, increased P nutrition resulted in the same effect in our research (advanced autumn leaf phenology in the +P treatment). This assumption is supported by the finding that beech photosynthetic activity increased after phosphorus fertilization, which was reported by Zavišić et al. [57]. Advanced autumn leaf phenology caused by elevated P might reduce plant susceptibility to early autumn frosts. For example, drought stress induces delayed autumn senescence in forest trees [6]. Since drought episodes during growth period have become more frequent, there is increasing probability for plants to be exposed to early autumn frosts. Therefore, it seems that improved plant P nutrition can be a part of solution to this problem, at least considering forest nurseries and/or seed orchards. It would be interesting to see if +P treatment may reduce adverse effects of drought. This question will be addressed in the extension of this study.
The observed differences between the species in their response intensities can be at least partially explained by significantly lower levels of absorbed phosphorus in oak than in beech in the +P treatment (Table 3). It is known that these two species differ significantly in terms of rooting depth. Beech roots are shallow, while oak roots are deep due to the intensive growth of their taproots. It was confirmed that oak saplings from studied provenances had significantly longer taproots with fewer fine roots than beech saplings [42]. Due to interspecies differences in root development patterns, it can be assumed that the beech saplings started to absorb nutrients (including P) from shallower parts of the substrate earlier and more intensively. This may be a reason that the oaks on average had lower phosphorus contents in their leaves (in September 2021) and therefore showed a less intense response in autumn phenology than the beech provenances.
Statistical nonsignificance of the differences between the Qp_KA in the +P treatment and the control was surprising (though phenology was advanced in the +P treatment). We hypothesize that excess and deficient phosphorus could have similar effects on autumn leaf phenology, i.e., earlier senescence occurs in both cases. Based on such an assumption, it seems likely that the Qp_KA provenance could have shown a convergence of the autumn phenology curves due to the occurrence of the largest difference in P content between the +P treatment (2.15 mg g-1) and the control (1.25 mg g-1). Of course, this assumption should be proven experimentally.
As expected, exposure to increased phosphorus contents in the substrate (+P treatment) had no effect on spring leaf phenology in the first year of the study (Figure 1a). Fertilization of the substrate with phosphorus was carried out approximately 15 days before plant leaf-out in the trial. It is evident that the saplings in the +P treatment could not absorb enough phosphorus during that short period to influence their leafing. The probable reason for the inability to absorb the P was insufficient time for the fertilizer to dissolve and become available to the plants. On the other hand, research on uptake dynamics and P allocation in common beech [56] indicates that P uptake from soil is extremely low in spring (during the leafing phase). The relatively high phosphorus content in leaves is the result of stored phosphorus allocation toward the new photosynthetic apparatuses. This means that plants mainly rely on stored phosphorus reserves during leafing and not on the current source in the soil. Nutrient uptake intensifies after the development of leaf mass. According to the same study [56], the main P uptake from soil occurs in late summer and fall.
The increased phosphorus effect on spring phenology in 2022 (the second experimental year) was almost completely absent, i.e., neither beech provenances nor an oak provenance showed any differences in flushing phenology between the +P treatment and the control. However, one oak provenance showed advanced spring phenology in the +P treatment (Figure 3d). Advanced flushing of holm oak (Quercus ilex L.) seedlings in response to NPK fertilization was reported [58]. Additionally, common beech seedlings originating from a phosphorus-rich habitat showed advanced leafing [23]. The latter report is based on a comparison of two provenances that differed significantly in soil phosphorus content (rich vs. poor P content). Although plants in our study were exposed to significantly different P levels, the soils were not P-rich or P-poor but rather in the upper normal vs. lower normal P content range (or lower normal vs. latent deficiency, in the case of oak - Table 3). In this context, it can be assumed that excess and deficient phosphorus have opposite effects on spring phenology (i.e., phosphorus surplus induces advance, while deficiency delays spring phenology). However, advanced flushing is shown when comparing groups of plants that strongly differ in phosphorus content, i.e., when comparing a phosphorus-deficit group with a group containing a P surplus. This assumption is supported by the fact that in this study, significantly advanced spring phenology was shown only by one oak provenance (Qp_Ka, see Figure 3), with the largest difference in phosphorus content between the +P treatment and control (Table 3, year 2022). On the other hand, it is possible that plants should be exposed to elevated phosphorus for more than a growing season to show a more intense response in spring leaf phenology. It should be recalled that at the beginning of leafing, young trees (of common beech) rely on phosphorus reserves stored mainly in coarse roots [56]. In that case, the accelerated flushing of the Qp_KA provenance could have indicated its greater efficiency in storing phosphorus, a phenomenon for which we currently have no direct evidence.
5. Conclusions
In general, the results showed that both species similarly responded to elevated phosphorus content in substrate i.e., with advanced autumn leaf phenology, although common beech responded more intensely in the first experimental year. A more pronounced response (i.e., more advanced autumn leaf phenology in the +P treatment than in the control) to the phosphorus treatment was shown in both species in the second experimental year. Advanced leaf senescence as response to elevated phosphorus may be due to increased plant productivity which corresponds with sink-limitation hypothesis.
There was no spring leaf phenology response to elevated phosphorus in common beech, and in one sessile oak provenance. However, the other oak provenance responded with advanced leafing indicating within-species variations. Therefore, the effect of elevated phosphorus was more clearly manifested in autumn than in spring leaf phenology, at least in the first growth period after fertilization, especially for common beech.
Author Contributions
Conceptualization, K.S. and S.B.; methodology, M.B., I.K.B. and S.B.; validation, M.B. and S.B.; formal analysis M.B. and S.B.; investigation, M.B., I.K.B. and S.B.; resources, K.S., S.B.; data curation, M.B. and S.B.; writing—original draft preparation, M.B. and S.B.; writing—review and editing, K.S. and I.K.B.; visualization M.B., K.S. and S.B.; supervision, S.B.; project administration, K.S.; funding acquisition, K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Croatian Science Foundation, grant number IP-2020-02-5204; Project title ”Phenotypic response of common beech and sessile oak provenances to long-lasting drought in interaction with different phosphorus concentrations in the soil”.
Data Availability Statement
The data in this study are available from the authors upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Polgar, C.A.; Primack, R.B. Leaf-out phenology of temperate woody plants: from trees to ecosystems. New Phytol. 2011, 191, 926–941. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Montgomery, R.A. Photoperiod constraints on tree phenology, performance and migration in a warming world. Plant, Cell Environ. 2015, 38, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Kuster, T.M.; Arend, M.; Günthardt-Goerg, M.S.; Schulin, R. Root growth of different oak provenances in two soils under drought stress and air warming conditions. Plant Soil 2012, 369, 61–71. [Google Scholar] [CrossRef]
- Vitasse, Y.; Lenz, A.; Kã¶Rner, C. The interaction between freezing tolerance and phenology in temperate deciduous trees. Front. Plant Sci. 2014, 5, 541–541. [Google Scholar] [CrossRef] [PubMed]
- Lenz, A.; Hoch, G.; Körner, C.; Vitasse, Y. Convergence of leaf-out towards minimum risk of freezing damage in temperate trees. Funct. Ecol. 2016, 30, 1480–1490. [Google Scholar] [CrossRef]
- ehulić, I.; Sever, K.; Katičić Bogdan, I.; Jazbec, A.; Škvorc, Ž.; Bogdan, S. Drought Impact on Leaf Phenology and Spring Frost Susceptibility in a Quercus Robur L. Provenance Trial. Forests 2019, 10, 50. [Google Scholar] [CrossRef]
- Murray, M.B.; Smith, R.I.; Leith, I.D.; Fowler, D.; Lee, H.S.J.; Friend, A.D.; Jarvis, P.G. Effects of elevated CO2, nutrition and climatic warming on bud phenology in Sitka spruce (Picea sitchensis) and their impact on the risk of frost damage. Tree Physiol. 1994, 14, 691–706. [Google Scholar] [CrossRef]
- Fløistad, I.S.; Kohmann, K. Influence of nutrient supply on spring frost hardiness and time of bud break in Norway spruce (Picea abies (L.) Karst.) seedlings. New For. 2004, 27, 1–11. [Google Scholar] [CrossRef]
- Luoranen, J.; Rikala, R. Nutrient loading of Norway spruce seedlings hastens bud burst and enhances root growth after outplanting. Silva Fenn. 2011, 45. [Google Scholar] [CrossRef]
- Mutz, J.; McClory, R.; van Dijk, L.J.A.; Ehrlén, J.; Tack, A.J.M. Pathogen infection influences the relationship between spring and autumn phenology at the seedling and leaf level. Oecologia 2021, 197, 447–457. [Google Scholar] [CrossRef]
- Maurer, S.; Egli, P.; Spinnler, D.; Körner, C. Carbon and water fluxes in Beech-Spruce model ecosystems in response to long-term exposure to atmospheric CO2 enrichment and increased nitrogen deposition. Funct. Ecol. 1999, 13, 748–755. [Google Scholar] [CrossRef]
- Nord, E.A.; Lynch, J.P. Plant phenology: a critical controller of soil resource acquisition. J. Exp. Bot. 2009, 60, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Sigurdsson, B.D. Elevated [CO2] and nutrient status modified leaf phenology and growth rhythm of young Populus trichocarpa trees in a 3-year field study. Trees 2001, 15, 403–413. [Google Scholar] [CrossRef]
- Wang, P.; Fu, C.; Wang, L.; Yan, T. Delayed autumnal leaf senescence following nutrient fertilization results in altered nitrogen resorption. Tree Physiol. 2022, 42, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hänninen, H.; Geng, X.; Peñuelas, J.; Zhang, X.; A Janssens, I.; Campioli, M. Nutrient availability alters the correlation between spring leaf-out and autumn leaf senescence dates. Tree Physiol. 2019, 39, 1277–1284. [Google Scholar] [CrossRef]
- Zani, D.; Crowther, T.W.; Mo, L.; Renner, S.S.; Zohner, C.M. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees. Science 2020, 370, 1066–1071. [Google Scholar] [CrossRef]
- Asshoff, R.; Zotz, G.; Körner, C. Growth and phenology of mature temperate forest trees in elevated CO2. Glob. Chang. Biol. 2006, 12, 848–861. [Google Scholar] [CrossRef]
- Ueda, M.U.; Mizumachi, E.; Tokuchi, N. Allocation of nitrogen within the crown during leaf expansion in Quercus serrata saplings. Tree Physiol. 2009, 29, 913–919. [Google Scholar] [CrossRef]
- Netzer, F.; Mueller, C.W.; Scheerer, U.; Grüner, J.; Kögel-Knabner, I.; Herschbach, C.; Rennenberg, H. Phosphorus Nutrition of Populus$\times$ Canescens Reflects Adaptation to High P-Availability in the Soil. Tree Physiol. 2018, 38, 6–24. [Google Scholar] [CrossRef]
- Netzer, F.; Herschbach, C.; Oikawa, A.; Okazaki, Y.; Dubbert, D.; Saito, K.; Rennenberg, H. Seasonal Alterations in Organic Phosphorus Metabolism Drive the Phosphorus Economy of Annual Growth in F. sylvatica Trees on P-Impoverished Soil. Front. Plant Sci. 2018, 9, 723. [Google Scholar] [CrossRef]
- Poirier, Y.; Bucher, M. Phosphate Transport and Homeostasis in Arabidopsis. Arab. Book 2002, 1, e0024–e0024. [Google Scholar] [CrossRef]
- Malhotra, H.; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore. 2018; pp. 171–190. [CrossRef]
- Yang, N.; Zavišić, A.; Pena, R.; Polle, A. Phenology, Photosynthesis, and Phosphorus in European Beech (Fagus Sylvatica L. ) in Two Forest Soils with Contrasting P Contents. J. Plant Nutr. Soil Sci. 2016, 179, 151–158. [Google Scholar]
- Kolström, M.; Lindner, M.; Vilén, T.; Maroschek, M.; Seidl, R.; Lexer, M.J.; Netherer, S.; Kremer, A.; Delzon, S.; Barbati, A.; et al. Reviewing the Science and Implementation of Climate Change Adaptation Measures in European Forestry. Forests 2011, 2, 961–982. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Ina, A.B.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. For. Int. J. For. Res. 2014, 87, 492–503. [Google Scholar] [CrossRef]
- Forest Europe 2020. State of Europe’s Forests 2020. Available online: https://foresteurope.org/state-of-europes-forests/ (accessed on 3 January 2023).
- Jansen, S.; Konrad, H.; Geburek, T. Crossing borders – European forest reproductive material moving in trade. J. Environ. Manag. 2019, 233, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Sever, K.; Bogdan, S.; Škvorc. ; Sever, M.Z.O.; Franjić, J. Estimation of leaf nitrogen concentrations in Quercus robur L. using the CCM-200 portable chlorophyll meter for different patterns of vegetative growth and acorn production. New For. 2016, 47, 513–527. [Google Scholar] [CrossRef]
- Sever, K.; Bogdan, S.; Škvorc. Response of photosynthesis, growth, and acorn mass of pedunculate oak to different levels of nitrogen in wet and dry growing seasons. J. For. Res. 2022, 34, 167–176. [Google Scholar] [CrossRef]
- Schmal, J.L.; Jacobs, D.F.; O’reilly, C. Nitrogen budgeting and quality of exponentially fertilized Quercus robur seedlings in Ireland. Eur. J. For. Res. 2011, 130, 557–567. [Google Scholar] [CrossRef]
- Walters, M.B.; Reich, P.B. Seed Size, Nitrogen Supply, and Growth Rate Affect Tree Seedling Survival in Deep Shade. Ecology 2000, 81, 1887–1901. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puértolas, J.; Peñuelas, J.L.; Planelles, R. Effect of nitrogen fertilization in the nursery on the drought and frost resistance of Mediterranean forest species. 2005, 14, 408. [CrossRef]
- Villar-Salvador, P.; Heredia, N.; Millard, P. Remobilization of acorn nitrogen for seedling growth in holm oak (Quercus ilex), cultivated with contrasting nutrient availability. Tree Physiol. 2010, 30, 257–263. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Penuelas, J.L.; Nicolás-Peragón, J.L.; Benito, L.F.; Domínguez-Lerena, S. Is Nitrogen Fertilization in the Nursery a Suitable Tool for Enhancing the Performance of Mediterranean Oak Plantations? New For. 2013, 44, 733–751. [Google Scholar] [CrossRef]
- Shi, W.; Villar-Salvador, P.; Li, G.; Jiang, X. Acorn size is more important than nursery fertilization for outplanting performance of Quercus variabilis container seedlings. Ann. For. Sci. 2019, 76, 22. [Google Scholar] [CrossRef]
- Vitra, A.; Lenz, A.; Vitasse, Y. Frost hardening and dehardening potential in temperate trees from winter to budburst. New Phytol. 2017, 216, 113–123. [Google Scholar] [CrossRef]
- Geilfus, C.-M.; Carpentier, S.C.; Zavišić, A.; Polle, A. Changes in the Fine Root Proteome of Fagus Sylvatica L. Trees Associated with P-Deficiency and Amelioration of P-Deficiency. J. Proteomics 2017, 169, 33–40. [Google Scholar]
- Pan, Y.; Song, Y.; Zhao, L.; Chen, P.; Bu, C.; Liu, P.; Zhang, D. The Genetic Basis of Phosphorus Utilization Efficiency in Plants Provide New Insight into Woody Perennial Plants Improvement. Int. J. Mol. Sci. 2022, 23, 2353. [Google Scholar] [CrossRef] [PubMed]
- Bolte, A.; Czajkowski, T.; Kompa, T. The north-eastern distribution range of European beech a review. For. Int. J. For. Res. 2007, 80, 413–429. [Google Scholar] [CrossRef]
- Maleki, K.; Zeller, L.; Pretzsch, H. Oak often needs to be promoted in mixed beech-oak stands - the structural processes behind competition and silvicultural management in mixed stands of European beech and sessile oak. iForest - Biogeosciences For. 2020, 13, 80–88. [Google Scholar] [CrossRef]
- Marchi, M.; Castellanos-Acuña, D.; Hamann, A.; Wang, T.; Ray, D.; Menzel, A. ClimateEU, scale-free climate normals, historical time series, and future projections for Europe. Sci. Data 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Sever, K.; Vukmirović, A.; Hodak, L.; Bogdan, S.; Katičić Bogdan, I.; Krstonošić, D.; Karažija, T.; Franjić, J.; Škvorc, Ž. Funkcionalna Prilagodba Prirodnog Pomlatka Hrasta Kitnjaka i Obične Bukve Na Različite Stanišne Prilike. Šumar. List 2022, 146, 293–307 (In Croatian with English summary), (In Croatian with English summary). [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. Methods of Soil Analysis, Part 3: Chemical Methods; John Wiley & Sons, 2020; Vol. 14.
- AOAC Official Methods of Analysis of AOAC.; International 17th edition.; Association of Analytical Communities Gaithersburg, MD, USA, 2000.
- Sabin, T.E.; Stafford, S.G. Assessing the need for transformation of response variables. For. Res. Lab. Or. State Univ. Corvallis Spec. Publ. 1990, 31 p.
- Mellert, K.H.; Göttlein, A. Comparison of new foliar nutrient thresholds derived from van den Burg’s literature compilation with established central European references. Eur. J. For. Res. 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- Perry, T.O. Dormancy of Trees in Winter. Science 1971, 171, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lagercrantz, U.L.F. At the End of the Day: A Common Molecular Mechanism for Photoperiod Responses in Plants? J. Exp. Bot. 2009, 60, 2501–2515. [Google Scholar] [CrossRef]
- Olsson, C.; Jönsson, A.M. A model framework for tree leaf colouring in Europe. Ecol. Model. 2015, 316, 41–51. [Google Scholar] [CrossRef]
- ufar, K.; De Luis, M.; Saz, M.A.; Črepinšek, Z.; Kajfež-Bogataj, L. Temporal Shifts in Leaf Phenology of Beech (Fagus Sylvatica) Depend on Elevation. Trees 2012, 26, 1091–1100. [Google Scholar]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and boreal forest tree phenology: from organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef]
- Weih, M. Genetic and environmental variation in spring and autumn phenology of biomass willows (Salix spp.): effects on shoot growth and nitrogen economy. Tree Physiol. 2009, 29, 1479–1490. [Google Scholar] [CrossRef]
- Smart, C.M. Gene expression during leaf senescence. New Phytol. 1994, 126, 419–448. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J. Phosphorus Nutrition Influence on Leaf Senescence in Soybean. Plant Physiol. 1992, 98, 1128–1132. [Google Scholar] [CrossRef]
- Colomb, B.; Kiniry, J.R.; Debaeke, P. Effect of Soil Phosphorus on Leaf Development and Senescence Dynamics of Field-Grown Maize. Agron. J. 2000, 92, 428–435. [Google Scholar] [CrossRef]
- Zavišić, A.; Polle, A. Dynamics of phosphorus nutrition, allocation and growth of young beech (Fagus sylvatica L.) trees in P-rich and P-poor forest soil. Tree Physiol. 2017, 38, 37–51. [Google Scholar] [CrossRef]
- Zavišić, A.; Yang, N.; Marhan, S.; Kandeler, E.; Polle, A. Forest Soil Phosphorus Resources and Fertilization Affect Ectomycorrhizal Community Composition, Beech P Uptake Efficiency, and Photosynthesis. Front. Plant Sci. 2018, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Oliet, J.A.; Salazar, J.M.; Villar, R.; Robredo, E.; Valladares, F. Fall fertilization of Holm oak affects N and P dynamics, root growth potential, and post-planting phenology and growth. Ann. For. Sci. 2011, 68, 647–656. [Google Scholar] [CrossRef]
Figure 1.
a) Bud burst date distribution by treatment in the first experimental year (2021). b) Leaf senescence date distribution by treatment in the first experimental year. +P treatment – elevated phosphorus; C – control with no phosphorus added. Different letters denote a statistically significant difference between the two treatments (at p<0.05).
Figure 1.
a) Bud burst date distribution by treatment in the first experimental year (2021). b) Leaf senescence date distribution by treatment in the first experimental year. +P treatment – elevated phosphorus; C – control with no phosphorus added. Different letters denote a statistically significant difference between the two treatments (at p<0.05).
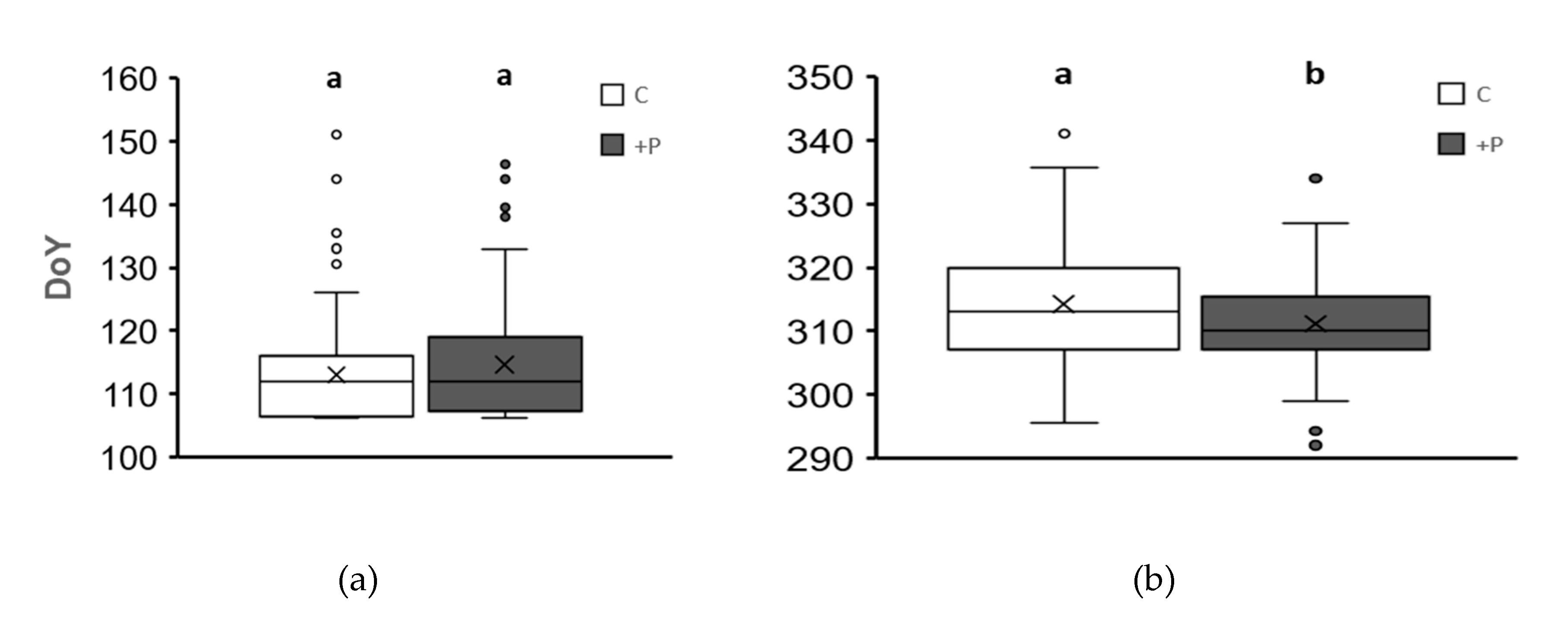
Figure 2.
Autumn leaf phenologies of beech (Fs) and oak (Qp) saplings in 2021, by provenance and treatment. (a) beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; C – control (no phosphorus added). The markers represent the means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).
Figure 2.
Autumn leaf phenologies of beech (Fs) and oak (Qp) saplings in 2021, by provenance and treatment. (a) beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; C – control (no phosphorus added). The markers represent the means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).

Figure 3.
Spring leaf phenology of the beech (Fs) and oak (Qp) saplings in 2022 by provenance and treatment. (a) Beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; Control – no phosphorus was added. The markers represent means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).
Figure 3.
Spring leaf phenology of the beech (Fs) and oak (Qp) saplings in 2022 by provenance and treatment. (a) Beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; Control – no phosphorus was added. The markers represent means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).
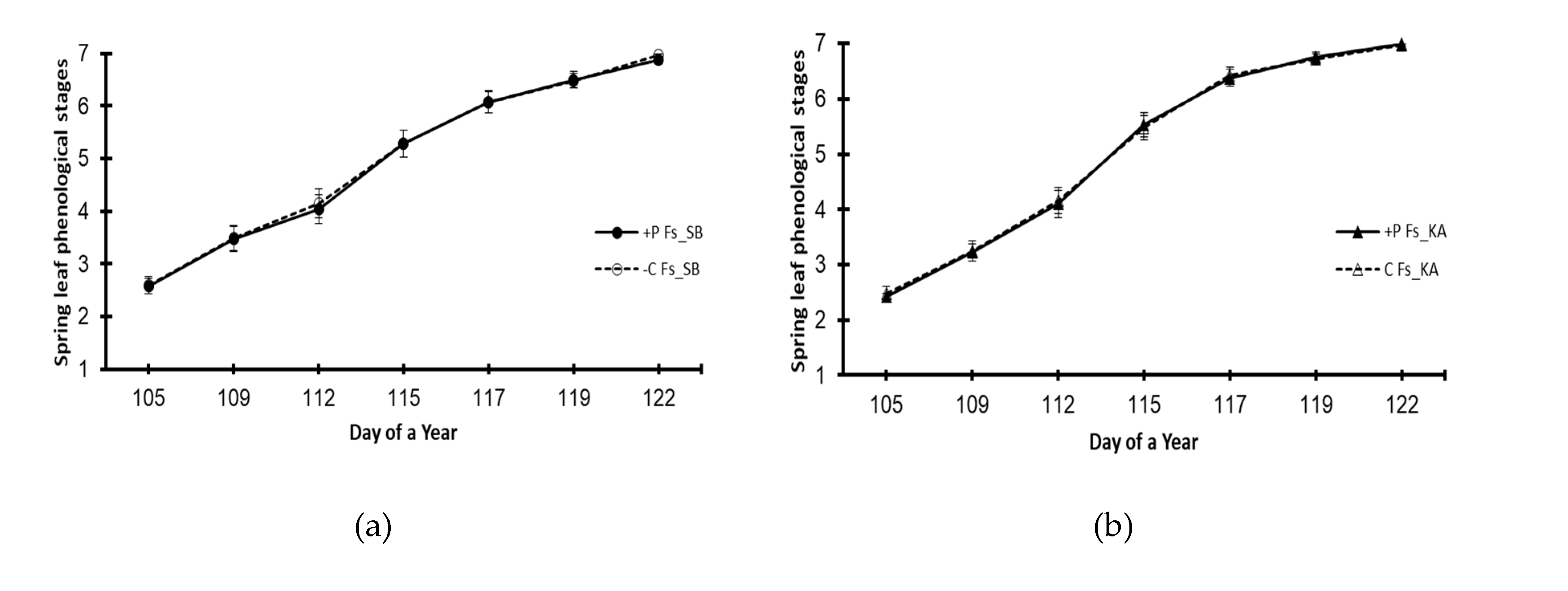
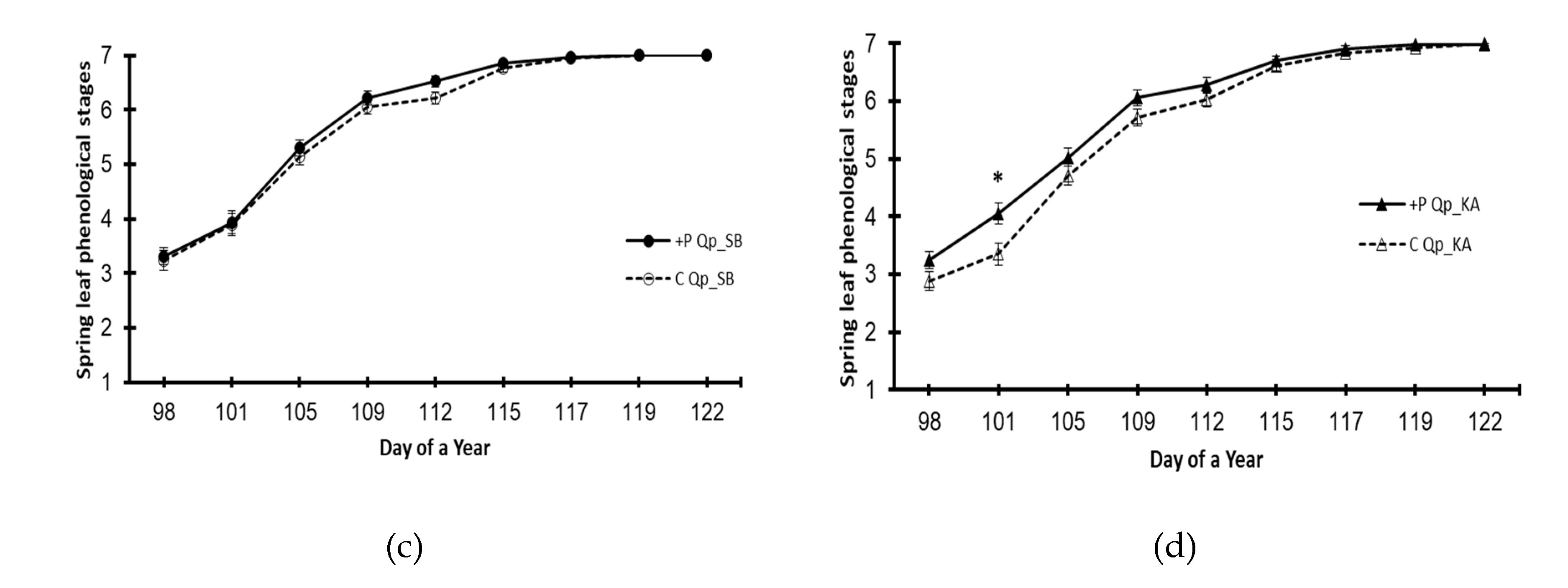
Figure 4.
Autumn leaf phenology of beech (Fs) and oak (Qp) saplings in 2022 by provenance and treatment. (a) Beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; C – control (no phosphorus added). The markers represent the means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).
Figure 4.
Autumn leaf phenology of beech (Fs) and oak (Qp) saplings in 2022 by provenance and treatment. (a) Beech provenance SB; (b) beech provenance KA; (c) oak provenance SB; (d) oak provenance KA. +P treatment – elevated phosphorus; C – control (no phosphorus added). The markers represent the means ± S.E.s (n=50). A star denotes a statistically significant difference between the treatments on a given day of the year (p<0.05).
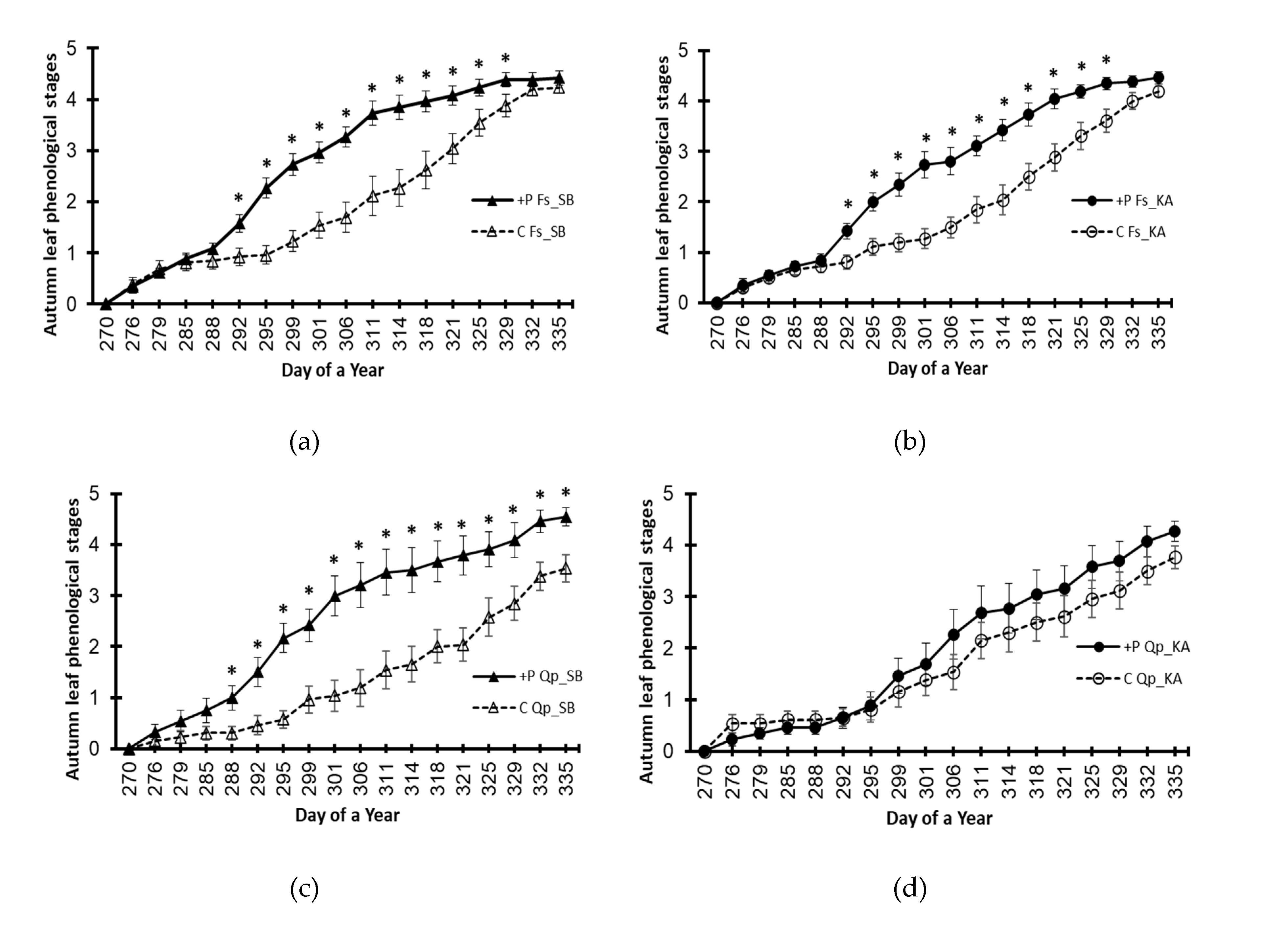

Table 1.
Basic climate data for the sampled stands. Generated for the 1991–2020 period by ClimateEU software (https://sites.ualberta.ca/~ahamann/data/climateeu.html; [41].
Table 1.
Basic climate data for the sampled stands. Generated for the 1991–2020 period by ClimateEU software (https://sites.ualberta.ca/~ahamann/data/climateeu.html; [41].
| Stand/provenance | Mean annual precipitation (mm) | Mean summer precipitation (mm) | Mean annual temperature (°C) | Mean coldest month temperature (°C) | Mean warmest month temperature (°C) |
| KA | 1099 | 508 | 12.3 | 2.0 | 22.4 |
| SB | 848 | 414 | 11.0 | 0.4 | 21.1 |
KA – the Karlovac provenance (stand); SB – the Slavonski Brod provenance.
Table 2.
Mean nutrient contents of the growth substrates in the treatment boxes. The +P treatment had elevated phosphorus content. The control had no added P.
Table 2.
Mean nutrient contents of the growth substrates in the treatment boxes. The +P treatment had elevated phosphorus content. The control had no added P.
| Parameter | +P treatment | Control | ||
| Value | Description1 | Value | Description1 | |
| pH (H2O) | 6.8 | Neutral | 7.1 | Neutral |
| P3- (mg/L) | 8.4 | High | 2.7 | Moderate |
| NH4+ (mg/L) | 13.2 | Optimal | 13.8 | Optimal |
| NO3- (mg/L.) | 59.5 | Optimal | 60.0 | Optimal |
| N total (mg/L) | 46.9 | Medium - normal | 49.7 | Medium - normal |
| K+ (mg/L) | 63.0 | Medium - normal | 60.9 | Medium - normal |
| Mg2+ (mg/L) | 61.4 | Moderate | 54.5 | Moderate |
| Ca2+ (mg/L) | 204 | Low | 234 | Low |
| Cl- (mg/L) | 54.8 | Medium -normal | 52.4 | Medium -normal |
| Na+ (mg/L) | 34.79 | Moderate | 36.6 | Moderate |
| E.C. (mS/cm) | 1.185 | Medium - normal | 1.156 | Medium - normal |
| Salt (%) | 0.151 | Medium - normal | 0.147 | Medium - normal |
1 Description according to [46].
Table 3.
Mean nutrient contents ± standard errors (mg g-1) in dried leaf samples of the beech and oak provenances by treatment. Samples were taken in September 2021 and May 2022.
Table 3.
Mean nutrient contents ± standard errors (mg g-1) in dried leaf samples of the beech and oak provenances by treatment. Samples were taken in September 2021 and May 2022.
| Nutrient | Year | Fs_KA | Fs_SB | Qp_KA | Qp_SB | ||||
| +P | C | +P | C | +P | C | +P | C | ||
| P | 2021 | 1.96*±0.04 | 1.31±0.03 | 1.72*±0.09 | 1.24±0.02 | 1.36*±0.08 | 1.08±0.04 | 1.45*±0.02 | 1.06±0.08 |
| 2022 | 1.82±0.16 | 1.64±0.07 | 1.95*±0.10 | 1.28±0.08 | 2.15*±0.35 | 1.22±0.10 | 1.76*±0.15 | 1.31±0.05 | |
| N | 2021 | 21.15±0.45 | 23.00*±0.32 | 21.75±0.19 | 22.50±0.77 | 28.67±0.43 | 27.30±0.60 | 28.08±0.45 | 29.00±0.54 |
| 2022 | 22.34±2.26 | 24.56±1.16 | 22.12±0.98 | 22.25±0.85 | 21.51±0.84 | 22.34±2.26 | 22.34±2.26 | 22.34±2.26 | |
| K | 2021 | 6.38±0.13 | 7.19*±0.30 | 6.57±0.15 | 7.54±0.13 | 7.73±0.09 | 7.73±0.17 | 7.69±0.10 | 7.83±0.11 |
| 2022 | 6.82±0.27 | 8.06*±0.41 | 6.57±0.34 | 7.31±0.34 | 9.67±0.46 | 9.13±0.40 | 9.59±0.28 | 9.15±0.44 | |
| Ca | 2021 | 8.06±0.27 | 8.89±0.43 | 9.18±0.10 | 9.41±0.08 | 6.34±0.27 | 7.72*±0.22 | 6.86±0.16 | 6.59±0.57 |
| 2022 | 7.47±0.62 | 6.71±0.28 | 9.18±0.53 | 8.68±0.63 | 10.41±0.88 | 10.91±0.71 | 10.32±0.76 | 10.98±1.23 | |
| Mg | 2021 | 1.53±0.12 | 1.35±0.06 | 1.58*±0.05 | 1.25±0.08 | 1.87±0.09 | 2.23*±0.04 | 2.00±0.05 | 1.97±0.09 |
| 2022 | 2.19±0.10 | 2.37±0.09 | 2.52±0.11 | 2.31±0.09 | 2.86±0.14 | 3.09±0.21 | 2.89±0.13 | 2.76±0.18 | |
Fs_KA – Fagus sylvatica provenance in the KA stand; Fs_SB – Fagus sylvatica provenance in the SB stand; Qp_KA – Quercus petraea provenance in the KA stand; Qp_SB – Quercus petraea provenance in the SB stand. +P – elevated phosphorus treatment; C – control; Asterisk indicates significantly larger nutrient contents in the treatment (within a provenance). P – phosphorus; N – nitrogen; K – potassium; Ca – calcium; Mg – magnesium.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



