Submitted:
31 March 2023
Posted:
04 April 2023
You are already at the latest version
Abstract
The nematode Angiostrongylus cantonensis has been reported worldwide, however, some basic questions remain unanswered about the presence of A. cantonensis in Ecua-dor: 1) Was the invasion of A. cantonensis in Ecuador unique, or did it occur in different waves? 2) Was this invasion as recent as historical records suggest? 3) Did this inva-sion come from other regions of South America or elsewhere? To address these issues, we assessed the genetic diversity of MT-CO1 gene sequences from isolates obtained in eleven of Ecuador's 24 provinces. Our Bayesian inference phylogenetic tree recovered A. cantonensis as a well-supported monophyletic group. All eleven sequences from Ec-uador were identical and identified as AC17a. The haplotype AC17a, found in Ecuador and the USA, formed a cluster with AC17b (USA); AC13 (Thailand); and AC12a-b (Cambodia). It is notable that all the samples, obtained in different geographic and climatic regions of different Ecuadorian provinces, had no genetic difference between them. Despite the lack of genetic information on A. cantonensis in Latin America, ex-cept in Brazil, our finding differs from previous studies by its absence of genetic diver-sity in Ecuador. We concluded that the invasion of A. cantonensis in Ecuador may have occurred: 1) as a one-time event, 2) recently, and 3) from Asia via the USA. Further re-search should include samples from countries neighboring Ecuador to delve deeper in-to this.
Keywords:
Invasive species
; eosinophilic meningoencephalitis
; cytochrome c oxidase subunit I
1. Introduction
The rat lungworm Angiostrongylus cantonensis (Chen, 1935) was first described in the bronchi of the rodents Rattus rattus (Linnaeus, 1758) and Rattus norvegicus (Berkenhout, 1769) in Guangzhou (formerly Canton), China [1]. This nematode is the etiological agent of neuroangiostrongyliasis which is the main cause of eosinophilic meningitis (EM) or eosinophilic meningoencephalitis (EME) in humans, an infectious disease of the central nervous system [2]. This disease is characterized primarily by an increase in the proportion of eosinophils in peripheral blood and cerebrospinal fluid, among other symptoms such as fever and severe headache [3,4].
The first documented human case of neuroangiostrongyliasis was in Taiwan in 1944, although it took nearly two decades to establish a clear link between the parasite and the disease (i.e., A. cantonensis as a causative agent of EME) [5]. Since the first report, several outbreaks were reported globally as the parasite has spread from traditional endemic regions of Southern China and Southeast Asia to the Pacific islands, Japan, Australia, Africa, the Canary Islands, the Balearic Islands, and the Americas, including the USA, Caribbean islands, and Brazil [2,4,6,7]. By 2008, more than 2,800 cases of human angiostrongyliasis had already been recorded in 30 countries [8]. The spread of parasites in different regions poses a threat not only to people living in endemic areas but to a growing number of travelers visiting these regions [9].
In 2008, A. cantonensis was reported for the first time in Ecuador parasitizing the giant African land snail Achatina (Lissachatina) fulica Bowdich, 1822 and the rats R. rattus and R. norvegicus [10,11]. Since then, outbreaks and isolated cases have been reported to the Ministry of Public Health of Ecuador (MSP) [12], with most clinical-epidemiological suspicion and one necropsy-confirmed case [13]. The parasite is now considered endemic throughout most of the country [10,14]. The invasive pest A. fulica is one of the main intermediate hosts for A. cantonensis [15]. This mollusk lives in urban and rural areas and plays an important role in the spread of the parasite [16]. Humans may become infected by ingesting food contaminated with third-stage larvae or by eating infected raw snails [2]. Thus, A. fulica is an important transmitter of eosinophilic meningoencephalitis and ocular angiostrongyliasis [17].
Different molecular biology methods have been employed to detect A. cantonensis [18,19,20,21,22,23]. Furthermore, they have been applied to explore systematic and population genetic aspects of Angiostrongylus taxa, since there is great variability within populations [24,25,26,27,28,29,30,31]. The use of mitochondrial genes, such as cytochrome c oxidase subunit I (MT-CO1), as molecular markers for specific identification of the parasite has been efficient [32,33,34,35]. The MT-CO1 gene has been used in studies on phylogeny, phylogeography, and haplotype identification [36,37,38,39]. However, some basic questions remain unanswered about the presence of A. cantonensis in Ecuador: 1) Was the invasion of A. cantonensis in Ecuador a single event or did it occur in different waves? 2) Was this invasion as recent as historical records suggest? 3) Did this invasion come from other regions of South America or elsewhere?
To tackle these questions, we assessed the genetic diversity of MT-CO1 gene sequences from isolates obtained in eleven of Ecuador's 24 provinces. Thus, we were able to verify how many lineages could be found in different regions of Ecuador and whether there was enough time for the lineages to diversify. We also established the phylogenetic and phylogeographic relationships of these isolates, comparing them with other sequences from South America and the rest of the world. Consequently, we could retrace the possible origin of the lineages found in Ecuador.
2. Materials and Methods
Parasites and Experimental Infection
Third-stage larvae (L3) were obtained from A. fulica, collected in eleven provinces of Ecuador (Figure 1) using the catch-per-unit-effort method for 30 minutes in each locality [15]. The L3 were used to experimentally infect 12-week-old adult female Wistar strain R. norvegicus rats (200 ± 2 g body mass). The rats were supplied by the Instituto de Investigación en Salud Pública (INSPI) vivarium, with their corresponding health and genetic quality certificates. The cycle was maintained in the National Reference Center for Parasitology. An average of 150 L3 (counted in a Neubauer chamber) were orally administered to each rat using a pipette. Infected rats were separated into cages (two specimens per cage) and identified according to the locality (province) where the infected gastropods were collected. Rats were kept under controlled conditions of temperature (21-24 °C) and humidity (60%), alternating 12-hour light/dark cycle, and received food and water at pH 7.0. All procedures were performed following the guidelines for the maintenance and use of laboratory animals, following the specific legislation covering animals used for scientific purposes Directive 2010/63/EU as amended by European Union (EU) Regulation 2019/1010 [40].
Thirty-five days after administration of the larvae, rats were euthanized using CO2 and the thoracic cavity (heart, pulmonary arteries, and lungs) was examined for parasitic nematodes (juvenile or adult), according to protocols previously established at INSPI [10] Specific taxonomic characteristics as caudal bursa and the spicule length were used to identify the nematodes [41,42]. Approximately 30-40 adult A. cantonensis specimens, from two rats infected representing each province, were stored in a sterile labeled 50 mL Falcon tube with 70% ethanol in an ultra freezer at -80 °C.
Molecular Phylogenetic and Phylogeographic Analyses
To conduct phylogenetic and phylogeographic studies, we used DNA sequences obtained from adult parasites, as previously reported [38,43,44]. Genomic DNA samples were isolated from adult parasites recovered from the rats representing each province. Before DNA isolation, the nematodes were partitioned into tiny pieces with a scalpel and suspended in saline (0.9% NaCl). We used the QIAamp DNA Mini Kit (QIAGEN, Netherlands) for DNA isolation according to the manufacturer's protocol. Each isolated DNA sample was identified according to its origin and stored at -80 °C until further amplification by PCR technique. Genomic DNA concentration was measured directly in a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA).
DNA isolated from approximately 30 adult parasites was subjected to PCR to amplify the mitochondrially encoded cytochrome c oxidase I (MT-CO1) gene [37]. PCR reactions were performed in a 25 μL total volume containing 12.5 μL of GoTaq Colorless Master Mix (Promega, USA: 2x DNA polymerase, 400µM dATP, 400µM dGTP, 400µM dCTP, 400µM dTTP, and 3mM MgCl2, pH 8.5); 1.5 μL of 10 μM each MT-CO1 gene primer (Invitrogen, Thermo Fisher Scientific, USA); 5.5 μL of distilled water; and 4 μL of genomic DNA. We also used a positive control consisting of an adult parasite DNA obtained from a wild-type rat (R. rattus) and a negative control with ultrapure water. The primers used were:
co1-F (5′TAAAGAAAGAAAGAACATAATGAAAATG3′)
co1-R (3′TTTTTTTTTTGGCATTCCTGAGGAGGT5′)
Modifications have been made to the original thermal cycling protocol by Vitta et al. [37] to standardize the technique in the INSPI laboratory and obtain the desired amplicons of approximately 450 base pairs (bp) as follows: 94 °C for 5 minutes; followed by 30 cycles of 94 °C for 1 minute, 48 °C for 30 seconds, and 72 °C for 60 seconds; with a final extension at 72°C for 5 minutes. PCR was performed in a C1000 Touch thermal cycler (Bio-Rad Laboratories, Inc., USA).
We verified PCR products after 1.2% agarose (Promega, USA) gel electrophoresis in 0.04 M Tris-acetate running buffer, 1Mm ethylenediamine tetraacetic acid, pH 8.0 (Invitrogen, Thermo Fisher Scientific, USA). We added 10 µl of Syber® 1x (10000 x) dye (Invitrogen, Thermo Fisher Scientific, USA) to the agarose gel. Blue/Orange Loading Dye, 6x (Promega, USA) was used as loading buffer. TrackIt 100bp DNA Ladder (0.1 µg/µL), with 100 to 1,000 bp range (Invitrogen, Thermo Fisher Scientific, USA), was used as molecular weight marker. Electrophoresis was performed at 80 V for 55 min using a PowerPac HC power supply (Bio-Rad, USA). PCR products were visualized using the ChemiDoc XRS imaging system (Bio-Rad, USA).
Amplicons purification; cycle-sequencing of both strands via the Sanger method, using the abovementioned PCR primers; and product precipitation, formamide resuspension, and analysis using the 3130 DNA Analyzer (Applied Biosystems, USA) were performed at the biochemistry department of the Universidad de las Américas (Ecuador).
The resulting chromatograms were edited with the software platform Geneious R7.0 (Biomatters Australia) [45]. Sense and anti-sense sequences of each amplified and sequenced sample were assembled into contigs. The resulting consensus sequences corresponding to 11 Ecuadorian provinces were deposited in the GenBank (Table 1).
To construct our MT-CO1 dataset, we used A. cantonensis sequences found in GenBank that overlapped with ours (Table S1). As outgroups, we added one sequence of Angiostrongylus mackerrasae Bhaibulaya, 1968 (MN793157) and three sequences of Angiostrongylus malaysiensis Bhaibulaya and Cross, 1971 (KT947979, KU532150, KU532153), all from GenBank (Table S1). Sequences in the dataset were aligned by multiple alignment using MUSCLE [46] under default parameters, within the Geneious package. Final manual trimming of non-overlapping regions of the alignment was done using the Mesquite 3.70 software package [47].
Two different matrices were used in this study. In the first matrix, used for phylogenetic inferences, we excluded all duplicated sequences, keeping only one copy of each haplotype of A. cantonensis and the outgroup. In the second matrix, used for phylogeographic analyses, we included all A. cantonensis sequences and excluded the outgroup. To find the optimal partition clustering arrangements and corresponding log(ml) values in both matrices, we conducted Bayesian clustering of linked molecular data analyses using BAPS 6.0 [48,49].
Bayesian inference (BI) phylogenetic analyses were conducted using MrBayes 3.2.6 [50], on XSEDE within the CIPRES Science Gateway [51]. We used independent GTR+I+G models for each codon position, with unlinking of base frequencies and parameters. Sampling was performed by MCMC, for 10,000,000 generations, with four simultaneous chains, in two runs. Node supports were given by Bayesian posterior probabilities (BPP) of trees sampled every 100 generations, after removal of the first 25% ‘burn-in’ generations. We assessed sampling adequacy using the program Tracer 1.7.1 [52] to calculate the effective sample sizes (ESSs) of parameters. We considered robust values above 200 effectively independent samples.
An intraspecific phylogeographic network was inferred using the program PopART, version 1.7 [53] with the median-joining network method [54]. Using DnaSP 6.12.03 [55], we organized the sequences into groups according to their geographic localities (countries). We also calculated, using DnaSP, the genetic diversity by the numbers of haplotypes (H), polymorphic sites (S), haplotype diversity (Hd), and nucleotide diversity (π). We finally used DnaSP for neutrality tests Tajima’s D [56] and Fu’s Fs [57].
3. Results
Along with our 11 MT-CO1 gene sequences of A. cantonensis from Ecuador, we added 105 sequences of A. cantonensis from GenBank and four sequences of outgroups. The full dataset had 120 sequences of Angiostrongylus ranging from 255 to 1617 bp in length (Table S1). The haplotypes were named AC1-17, following the names for haplotypes previously adopted [36,39,58], adding letters to variants. All 11 sequences from Ecuador were identical and identified as AC17a. The sequences from Ecuador were also identical to five sequences from New Orleans, Louisiana, USA (USA-LA), retrieved from GenBank.
3.1. Molecular Phylogenetic Analyses
After multiple sequence alignment, trimming, and removal of all duplicates in the first matrix, for phylogenetic inferences, the matrix resulted in 29 taxa and 255 sites. Of these, 201 were constant characters and 41 were variable parsimony-informative characters. Angiostrongylus cantonensis was represented by 25 sequences while the outgroup by four. According to the population structure recovered using BAPS, Angiostrongylus specimens were distributed in five clusters in the 29 sequences matrix.
After 25% burn-in removal, the BI mean estimated marginal likelihood was -751.4969 and the median was -751.1709. The ESS values were well above 200 for all parameters. The BI phylogenetic tree (Figure 2) recovered A. cantonensis as a well-supported monophyletic group (BPP = 1.00). Within A. cantonensis, the sequence AC17a, from Ecuador and USA-LA was in a polytomy with AC17b (USA-LA); AC5a (Brazil, Japan, French Polynesia, and Hawaii, USA); AC5b (Japan); AC13 (Thailand); and a moderately supported clade (BPP = 0.70), formed by sequences AC8a (Brazil) and AC8b (Australia, Balearics, Canaries, Taiwan, and USA-LA). This polytomy was moderately supported (BPP = 0.78) and formed another polytomy with sequences AC12a and AC12b from Cambodia. This more inclusive polytomy was strongly supported (BPP = 0.98) and coincided with Cluster 3.
3.2. Phylogeographic Analyses
The second matrix, for phylogeographic analyses, included only sequences of A. cantonensis. This dataset included 11 sequences from Ecuador and 105 sequences from GenBank, excluding the outgroup, totaling 116 taxa and 255 sites, after multiple sequence alignment and trimming. The total number of sites excluding sites with gaps or missing data was 254. The number of haplotypes was H = 25, the number of polymorphic sites S = 36, the haplotype diversity Hd = 0.895, the nucleotide diversity π = 0.02546, Fu's Fs = -2.380, and Tajima’s D = -0.42728 (P > 0.10).
According to the population structure recovered using BAPS, A. cantonensis specimens were distributed in seven clusters in the 116 sequences matrix. We indicated the clusters in the intraspecific phylogeographic network (Figure 3). The haplotype AC17a, from Ecuador and USA-LA, formed a cluster with AC17b (USA-LA); AC13 (Thailand); and AC12a-b (Cambodia). This haplogroup was labeled Cluster 5 in the network.
4. Discussion
The introduction of non-native mollusks, such as A. fulica, plays an important role in the transmission of A. cantonensis [59]. Since the mid-20th century, A. fulica has been introduced into the tropics and subtropics and has since been considered the most harmful snail pest in these regions [16]. In Brazil, these mollusks were possibly introduced more than once, on different occasions [60]. The first account is from the mid-1970s in the state of Minas Gerais [61]. The second, better documented, and probably the major introduction was in the late 1980s in the state of Paraná from specimens brought from Indonesia for commercial purposes (snail farming) that were unsuccessful [62]. The giant African snail is currently widespread in all 26 Brazilian states and the Federal District [63,64].
According to data from an Ecuadorian government organization, these snails were brought into the country in the mid-1990s. As in Brazil, this was for commercial purposes. Snail farms were built in some valleys of the Ecuadorian highlands, which offered an ideal temperature between 17 °C and 25 °C [65]. However, their breeding did not provide the expected economic returns, inevitably, most of the farms were abandoned and the snails were released into the environment. The result was a widespread infestation of urban and rural areas in almost all provinces of the country [11]. Achatina fulica was probably the vector that introduced A. cantonensis to the country, as was the case in Brazil [66] and China [67].
As for the definitive hosts, it is presumed that R. rattus arrived in Ecuador between the 16th and 17th centuries with the ships of the Spanish conquistadors [68]. Rattus norvegicus probably originated in China and spread to Europe, reaching North America through shipping during the second half of the 18th century. Both species are now widely distributed in urban areas around the planet [69].
In 2008, the snail A. fulica (intermediate host) and the rat R. rattus (definitive host) were found naturally infected by A. cantonensis in Ecuador [10,70]. The existence of intermediate and definitive hosts in almost the entire Ecuador has contributed to the endemic nature of angiostrongyliasis distribution, making the control of this disease even more complex [10]. Both intermediate (A. fulica) and definitive (R. rattus) hosts are non-native species to Ecuador and are considered among the 100 most important invasive species in the world, according to the World Conservation Union [71]. Invasive species in an ecosystem can affect biotic alter interactions, impacting the economy, the environment, or public and animal health [72,73]. Moreover, the practice of eating raw snails by elder Ecuadorians increases the risk of A. cantonensis infection [11].
Earlier studies using the MT-CO1 to distinguish A. cantonensis isolates have shown different geographical isolates in determinate regions [36,38,39,58]. Tokiwa et al. [39] reported seven different haplotypes (AC1 to AC7): five were found in Japan (AC1, AC2, AC3, AC5, and AC7), two in mainland China (AC2 and AC6) and only one in Taiwan (AC1). In Brazil, analyses from 15 geographic isolates determined the presence of three different MT-CO1 haplotypes (AC5, AC8, and AC9). Most of the sample sequences were AC5 or AC8, whereas AC9 was a new haplotype [36]. Rodpai et al. [58] identified different A. cantonensis haplotypes in Cambodia, Myanmar, Thailand, and Hawaii, USA. Two of them (AC2 and AC5) had been previously reported. The AC2 haplotype, previously reported in China and Japan, was found in Myanmar. The AC5 haplotype, previously reported in Brazil and Japan, was found in Hawaii. Additionally, four new haplotypes (AC10-AC13) were reported in Southeast Asia [58].
Such studies have shown that A. cantonensis in Asia has greater genetic diversity [38,39,58], indicating that this parasite has been circulating in these regions for a long time. Conversely, the sequence diversity of A. cantonensis is low in several regions outside Asia [74]. Otherwise, there is little or no genetic information on the parasite in other regions of the planet, such as the Americas, except in Brazil [36].
In the present study, all sequences of the isolates from Ecuador were identical, the haplotype AC17a. In our phylogenetic analyses, this haplotype was nested into a polytomy with other sequences from different parts of the world. Remarkably, all samples were obtained from provinces of Ecuador in different geographic and climatic regions, and yet they did not show any genetic divergence between them.
The findings reported here represent a novelty in the study of the genetic diversity of A. cantonensis isolates. Although there is a lack of information on the genetic diversity of this parasite in other Latin American countries, except for Brazil, our results are different from previous studies due to the complete absence of genetic diversity of A. cantonensis in Ecuador. Even admitting that the low number of nucleotide base pairs obtained could make the sequence homogeneous in the isolates from Ecuador, this same region of the MT-CO1 gene showed variations in the other haplotypes compared.
The fact that only one haplotype was found in 11 different Ecuadorian provinces, is revealing. It strongly advocates a single introduction event. Furthermore, this result suggests that A. cantonensis has been recently introduced in the country, as there was no time for new haplotypes to differentiate from the original. This may justify the non-existence of genetic diversity among different circulating isolates.
Interestingly, the sequences from Ecuador shared a recent common ancestor with two Brazilian haplotypes (AC5 and AC8) [75]. However, it is unlikely that this could indicate a historical connection between the strains from both countries. The AC5 haplotypes found in Brazil from Pirituba (state of São Paulo), Queimados, and Niterói (state of Rio de Janeiro) correspond to a haplotype found in Japan, Hawaii, and French Polynesia [58,74,76], suggesting that the arrival of the parasite in Rio de Janeiro or São Paulo may have occurred from the Asian continent [36]. This hypothesis is also considered for the AC8a haplotype, closely related to AC8b, found in Australia, the Balearics, the Canaries, Taiwan, and the United States of America (USA). This shows the possible spread of A. cantonensis, with the giant African land snail, as a vector, from the arrival localities in Brazil to the Southeast, Northeast, and North Brazilian regions [36].
The sequences obtained here also grouped with AC13 and AC17 haplotypes from Thailand and the USA, respectively. The haplotypes AC10, AC11, and AC13, from Thailand, and AC12, from Cambodia, were described by Rodpai et al. [58] in phylogenetic studies using different DNA regions of A. cantonensis and A. malaysiensis. The haplotypes AC17, from the USA, were reported in a study to identify A. cantonensis and determine the association between ecological characteristics and factors related to definitive hosts (R. rattus, R. norvegicus, Sigmodon hispidus, and Oryzomys palustris) associated with transmission risk of angiostrongyliasis in New Orleans [77]. The haplotypes AC12, AC13, and AC17 formed a cluster in the haplotype cluster analysis, suggesting that A. cantonensis may have arrived in Ecuador from Asia via the USA.
5. Conclusions
Our results suggest that the invasion of A. cantonensis in Ecuador occurred as a single event, since only one haplotype was present in all 11 provinces studied, encompassing different ecoregions of Ecuador. Moreover, this invasion took place very recently, as we found no variation from the initial haplotype. It is unlikely that A. cantonensis reached Ecuador from Brazil. It is conceivable that the lineage found in Ecuador came from Asia via the USA. Future studies should sample countries neighboring Ecuador to infer migratory routes into this country in more detail.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Table S1 - 120 documents from Angiostrongylus.txt.
Author Contributions
Conceptualization, L.S.A., L.R.R. and H.H.A.; methodology, H.H.A., L.R.R., L.S.A., R.V.V.; software, C.B.P., L.M.P., R.V.V.; validation, J.F.N., M.R.O.; formal analysis, C.B.P., J.F.N., L.M.P., R.V.V.; investigation, L.S.A., H.H.A.; resources, L.S.A., R.V.V.; data curation, C.B.P., L.S.A., R.V.V.; writing—original draft preparation, C.B.P., L.S.A.; writing—review and editing, C.B.P., L.S.A., R.V.V., R.O.S.; visualization, R.V.V., R.O.S.; supervision, L.S.A.; project administration, L.S.A.; funding acquisition, L.S.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All procedures were performed following the guidelines for the maintenance and use of laboratory animals, following the specific legislation covering animals used for scientific purposes Directive 2010/63/EU as amended by European Union (EU) Regulation 2019/1010.
Informed Consent Statement
The protocol was approved by the Human Research Ethics Commission (CEISH) of the Kennedy Clinic of the City of Guayaquil, approved by the Ministry of Health of Ecuador, unique approval code DIS-CEISH-HCK-09-014. Doctors were completely free to decide to participate in the investigation. At the end of the survey, they signed and entered the number of their identification document in it.
Data Availability Statement
The data presented in this study are openly available in GenBank, accession numbers MW390965 to MW390974 and MW391020
Acknowledgments
The authors thank members of the Parasitology Laboratory, Centro de Referencia Nacional de Parasitología y Micología, Instituto de Investigación en Salud Pública, INSPI, and the staff of the Microbiology laboratory, Facultad de Medicina, Universidad de Guayaquil. We are also grateful to Dr. Alessandra L. Morassutti and Dr. Arnaldo Maldonado Júnior, who put us in contact to carry out this work.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. The researchers declare that they have no conflict of interest.
References
- Chen, H.-T. Un Nouveau Nématode Pulmonaire, Pulmonema Cantonensis, n. g., n. Sp. Annales de Parasitologie Humaine et Comparée 1935, 13, 312–317. [Google Scholar] [CrossRef]
- Eamsobhana, P. Eosinophilic Meningitis Caused by Angiostrongylus Cantonensis--a Neglected Disease with Escalating Importance. Tropical biomedicine 2014, 31, 569–578. [Google Scholar] [PubMed]
- Martins, Y.C.; Tanowitz, H.B.; Kazacos, K.R. Central Nervous System Manifestations of Angiostrongylus Cantonensis Infection. Acta Tropica 2015, 141, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Wu, Z.D.; Wei, J.; Owen, R.L.; Lun, Z.R. Human Angiostrongylus Cantonensis: An Update. European Journal of Clinical Microbiology & Infectious Diseases 2011 31:4 2012, 31, 389–395. [Google Scholar] [CrossRef]
- Alicata, J.E. Biology and Distribution of the Rat Lungworm, Angiostrongylus Cantonensis, and Its Relationship to Eosinophilic Meningoencephalitis and Other Neurological Disorders of Man and Animals**Paper Presented at the First International Congress of Parasitology, Rome, Italy. In Advances in Parasitology; Dawes, B., Ed.; Academic Press, 19–26 September 1965; pp. 223–248. [Google Scholar]
- Maldonado-Júnior, A.; Simões, R.O.; Oliveira, A.P.M.; Motta, E.M.; Fernandez, M.A.; Pereira, Z.M.; Monteiro, S.S.; Torres, E.J.L.; Thiengo, S.C. First Report of Angiostrongylus Cantonensis (Nematoda: Metastrongylidae) in Achatina Fulica (Mollusca: Gastropoda) from Southeast and South Brazil. Memórias do Instituto Oswaldo Cruz 2010, 105, 938–941. [Google Scholar] [CrossRef]
- Cowie, R.H.; Ansdell, V.; Panosian Dunavan, C.; Rollins, R.L. Neuroangiostrongyliasis: Global Spread of an Emerging Tropical Disease. The American Journal of Tropical Medicine and Hygiene 2022, tpmd220360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Lai, D.H.; Zhu, X.Q.; Chen, X.G.; Lun, Z.R. Human Angiostrongyliasis. Lancet Infect Dis 2008, 8, 621–630. [Google Scholar] [CrossRef]
- Ansdell, V.; Wattanagoon, Y. Angiostrongylus Cantonensis in Travelers: Clinical Manifestations, Diagnosis, and Treatment. Current Opinion in Infectious Diseases 2018, 31, 399–408. [Google Scholar] [CrossRef]
- Solorzano, L.F.; Martini Robles, L.; Hernandez, H.; Sarracent, J.; Muzzio, J.; Rojas, L. Angiostrongylus cantonensis: un parásito emergente en Ecuador. Revista Cubana Medicina Tropical 2014, 66. [Google Scholar]
- Martini-Robles, L.; Gómez-Landires, E.; Muzzio-Aroca, J.; Solórzano-Álava, L. Descripcion Del Primer Foco de Transmision Natural de Angiostrongylus Cantonensis En Ecuador. In Angiostrongylus cantonensis - Emergencia en América; Martini-Robles, L., Dorta-Contreras, A.J., Eds.; Editorial Academia La Habana: La Habana, 2016; ISBN 978-959-270-368-1. [Google Scholar]
- Pincay, T.; García, L.; Narváez, E.; Decker, O.; Martini, L.; Moreira, J.M. Angiostrongiliasis Por Parastrongylus (Angiostrongylus) Cantonensis En Ecuador. Primer Informe En Sudamérica. Tropical Medical Int Health 2009, 14, S37. [Google Scholar]
- Guerrero, M.; Vargas, F.M.; Rosero, A.R.; Nuques, M. de L.; Bolaños, E.S.; Briones, M.T.; Martínez, W.Z.; Gómez, A.O. Meningitis eosinofílica por angiostrongylus cantonensis. Reporte de caso de autopsia. Medicina 2008, 13, 312–318. [Google Scholar]
- Solórzano, L.; Sánchez-Amador, F.; Valverde, T. Angiostrongylus (Parastrongylus) cantonensis en huéspedes intermediarios y definitivos en Ecuador, 2014-2017. Biomédica 2019, 39, 370–384. [Google Scholar] [CrossRef] [PubMed]
- Muzzio, J. Hospederos intermediarios de Angiostrongylus cantonensis en Ecuador; Editorial Academica Española: Saarbrücken, 2014. [Google Scholar]
- Thiengo, S.C.; Faraco, F.A.; Salgado, N.C.; Cowie, R.H.; Fernandez, M.A. Rapid Spread of an Invasive Snail in South America: The Giant African Snail, Achatina Fulica, in Brasil. Biological Invasions 2007, 9, 693–702. [Google Scholar] [CrossRef]
- Moreira, V.L.C.; Giese, E.G.; Melo, F.T.V.; Simões, R. de O.; Thiengo, S.C.; Maldonado-Júnior, A.; Santos, J.N. Endemic Angiostrongyliasis in the Brazilian Amazon: Natural Parasitism of Angiostrongylus Cantonensis in Rattus Rattus and R. Norvegicus, and Sympatric Giant African Land Snails, Achatina Fulica. Acta tropica 2013, 125, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Sullivan, J.J.; Bishop, H.S.; Hollingsworth, R.; da Silva, A.J. PCR-Based Detection of Angiostrongylus Cantonensis in Tissue and Mucus Secretions from Molluscan Hosts. Applied and Environmental Microbiology 2007, 73, 1415–1419. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; da Silva, A.J.C.A.; Teem, J.L.; Hollingsworth, R.; Bishop, H.S.; Graeff-Teixeira, C.; Aramburu Da Silva, A.C. Improved Molecular Detection of Angiostrongylus Cantonensis in Mollusks and Other Environmental Samples with a Species-Specific Internal Transcribed Spacer Based TaqMan Assay. Applied and Environmental Microbiology 2010, 76, 5287–5289. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; Xayavong, M.; da Silva, A.C.A.; Park, S.Y.; Whelen, A.C.; Calimlim, P.S.; Sciulli, R.H.; Honda, S.A.A.; Higa, K.; Kitsutani, P.; et al. Real-Time Polymerase Chain Reaction Detection of Angiostrongylus Cantonensis DNA in Cerebrospinal Fluid from Patients with Eosinophilic Meningitis. The American Journal of Tropical Medicine and Hygiene 2016, 94, 176–181. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; Bishop, H.S.; da Silva, A.J. Detection of Rat Lungworm in Intermediate, Definitive, and Paratenic Hosts Obtained from Environmental Sources. Hawai’i Journal of Medicine & Public Health: A Journal of Asia Pacific Medicine & Public Health 2013, 72, 63–69. [Google Scholar]
- Sears, W.J.; Qvarnstrom, Y.; Nutman, T.B. RPAcan3990: An Ultrasensitive Recombinase Polymerase Assay To Detect Angiostrongylus Cantonensis DNA. J Clin Microbiol 2021, 59, e0118521. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Song, H.-Q.; Zhang, R.-L.; Chen, M.-X.; Xu, M.-J.; Ai, L.; Chen, X.-G.; Zhan, X.-M.; Liang, S.-H.; Yuan, Z.-G.; et al. Specific Detection of Angiostrongylus Cantonensis in the Snail Achatina Fulica Using a Loop-Mediated Isothermal Amplification (LAMP) Assay. Molecular and Cellular Probes 2011, 25, 164–167. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Steinmann, P.; Utzinger, J.; Zhou, X.-N. The Genetic Variation of Angiostrongylus Cantonensis in the People’s Republic of China. Infectious Diseases of Poverty 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Zhang, R.L.; Chen, M.X.; li, J.; Ai, L.; Wu, C.Y.; Zhu, X.-Q.; Lin, Q. Characterisation of Angiostrongylus Cantonensis Isolates from China by Sequences of Internal Transcribed Spacers of Nuclear Ribosomal DNA. Journal of Animal and Veterinary Advances 2011, 10, 593–596. [Google Scholar] [CrossRef]
- Peng, J.; He, Z.; Zhang, S.; Lun, Z.; Wu, Z.; Fan, C.-K.; Brown, C.L.; Cheng, P.; Peng, S.; Yang, T. Phylogeography of Angiostrongylus Cantonensis (Nematoda: Angiostrongylidae) in Southern China and Some Surrounding Areas. PLOS Neglected Tropical Diseases 2017, 11, e0005776. [Google Scholar] [CrossRef] [PubMed]
- Fontanilla, I.K.C.; Wade, C.M. The Small Subunit (SSU) Ribosomal (r) RNA Gene as a Genetic Marker for Identifying Infective 3rd Juvenile Stage Angiostrongylus Cantonensis. Acta Tropica 2008, 105, 181–186. [Google Scholar] [CrossRef]
- Eamsobhana, P.; Lim, P.E.; Yong, H.S. Phylogenetics and Systematics of Angiostrongylus Lungworms and Related Taxa (Nematoda: Metastrongyloidea) Inferred from the Nuclear Small Subunit (SSU) Ribosomal DNA Sequences. Journal of Helminthology 2015, 89, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Nabholz, B.; Glémin, S.; Hurst, G.D.D. Mitochondrial DNA as a Marker of Molecular Diversity: A Reappraisal. Molecular Ecology 2009, 18, 4541–4550. [Google Scholar] [CrossRef] [PubMed]
- Blouin, M.S. Molecular Prospecting for Cryptic Species of Nematodes: Mitochondrial DNA versus Internal Transcribed Spacer. International Journal for Parasitology 2002, 32, 527–531. [Google Scholar] [CrossRef]
- Yong, H.S.; Eamsobhana, P.; Song, S.-L.; Prasartvit, A. Molecular Phylogeography of Angiostrongylus Cantonensis (Nematoda: Angiostrongylidae) and Genetic Relationships with Congeners Using Cytochrome b Gene Marker. Acta Tropica 2015, 148, 66–71. [Google Scholar] [CrossRef]
- Almeida, L.R. de; Souza, J.G.R. de; Santos, H.A.; Torres, E.J.L.; Vilela, R. do V.; Cruz, O.M.S.; Rodrigues, L.; Pereira, C.A. de J.; Maldonado-Júnior, A.; Lima, W. dos S. Angiostrongylus Minasensis n. Sp.: New Species Found Parasitizing Coatis (Nasua Nasua) in an Urban Protected Area in Brazil. Revista Brasileira de Parasitologia Veterinária 2020, 29, e018119. [Google Scholar] [CrossRef]
- Eamsobhana, P.; Lim, P.-E.; Solano, G.; Zhang, H.; Gan, X.; Yong, H.S. Molecular Differentiation of Angiostrongylus Taxa (Nematoda: Angiostrongylidae) by Cytochrome c Oxidase Subunit I (COI) Gene Sequences. Acta tropica 2010, 116, 152–156. [Google Scholar] [CrossRef]
- Caldeira, R.L.; Carvalho, O. dos S.; Mendonça, C.L.G.F.; Graeff-Teixeira, C.; Silva, M.C.; Ben, R.; Maurer, R.; Lima, W. dos S.; Lenzi, H.L. Molecular Differentiation of Angiostrongylus Costaricensis, A. Cantonensis, and A. Vasorum by Polymerase Chain Reaction-Restriction Fragment Length Polymorphism. Memórias do Instituto Oswaldo Cruz 2003, 98, 1039–1043. [Google Scholar] [PubMed]
- Valentyne, H.; Spratt, D.M.; Aghazadeh, M.; Jones, M.K.; Šlapeta, J. The Mitochondrial Genome of Angiostrongylus Mackerrasae Is Distinct from A. Cantonensis and A. Malaysiensis. Parasitology 2020, 147, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Monte, T.C.C.; Simões, R. de O.; Oliveira, A.P.M.; Novaes, C.F.; Thiengo, S.C.; Silva, A.J.; Cordeiro-Estrela, P.; Maldonado-Júnior, A. Phylogenetic Relationship of the Brazilian Isolates of the Rat Lungworm Angiostrongylus Cantonensis (Nematoda: Metastrongylidae) Employing Mitochondrial COI Gene Sequence Data. Parasites & vectors 2012, 5, 248. [Google Scholar] [CrossRef]
- Vitta, A.; Srisongcram, N.; Thiproaj, J.; Wongma, A.; Polsut, W.; Fukruksa, C.; Yimthin, T.; Mangkit, B.; Thanwisai, A.; Dekumyoy, P. Phylogeny of Angiostrongylus Cantonensis in Thailand Based on Cytochrome c Oxidase Subunit I Gene Sequence. The Southeast Asian journal of tropical medicine and public health 2016, 47, 377–386. [Google Scholar]
- Eamsobhana, P.; Song, S.L.; Yong, H.S.; Prasartvit, A.; Boonyong, S.; Tungtrongchitr, A. Cytochrome c Oxidase Subunit I Haplotype Diversity of Angiostrongylus Cantonensis (Nematoda: Angiostrongylidae). Acta Tropica 2017, 171, 141–145. [Google Scholar] [CrossRef]
- Tokiwa, T.; Harunari, T.; Tanikawa, T.; Komatsu, N.; Koizumi, N.; Tung, K.-C.; Suzuki, J.; Kadosaka, T.; Takada, N.; Kumagai, T.; et al. Phylogenetic Relationships of Rat Lungworm, Angiostrongylus Cantonensis, Isolated from Different Geographical Regions Revealed Widespread Multiple Lineages. Parasitology International 2012, 61, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Legislation for the Protection of Animals Used for Scientific Purposes - Environment - European Commission Available online:. Available online: https://ec.europa.eu/environment/chemicals/lab_animals/legislation_en.htm (accessed on 29 October 2020).
- Aguiar, P.H.; Morera, P.; Pascual, J. First Record of Angiostrongylus Cantonensis in Cuba. The American Journal of Tropical Medicine and Hygiene 1981, 30, 963–965. [Google Scholar] [CrossRef]
- Ubelaker, J.E. Systematics of Species Referred to the Genus Angiostrongylus. The Journal of Parasitology 1986, 72, 237–244. [Google Scholar] [CrossRef]
- Chan, A.H.E.; Chaisiri, K.; Dusitsittipon, S.; Jakkul, W.; Charoennitiwat, V.; Komalamisra, C.; Thaenkham, U. Mitochondrial Ribosomal Genes as Novel Genetic Markers for Discrimination of Closely Related Species in the Angiostrongylus Cantonensis Lineage. Acta Tropica 2020, 211, 105645. [Google Scholar] [CrossRef]
- Eamsobhana, P.; Yong, H.-S.; Song, S.-L.; Gan, X.-X.; Prasartvit, A.; Tungtrongchitr, A. Molecular Phylogeography and Genetic Diversity of Angiostrongylus Cantonensis and A. Malaysiensis (Nematoda: Angiostrongylidae) Based on 66-KDa Protein Gene. Parasitology International 2019, 68, 24–30. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Research 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis 2021.
- Corander, J.; Marttinen, P.; Sirén, J.; Tang, J. Enhanced Bayesian Modelling in BAPS Software for Learning Genetic Structures of Populations. BMC Bioinformatics 2008, 9, 539. [Google Scholar] [CrossRef] [PubMed]
- Corander, J.; Sirén, J.; Arjas, E. Bayesian Spatial Modeling of Genetic Population Structure. Computational Statistics 2008, 23, 111–129. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Systematic Biology 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE); IEEE, November 2010; pp. 1–8. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Systematic Biology 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-Feature Software for Haplotype Network Construction. Methods in Ecology and Evolution 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Molecular Biology and Evolution 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Molecular Biology and Evolution 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.-X. Statistical Tests of Neutrality of Mutations Against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Rodpai, R.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Sadaow, L.; Laymanivong, S.; Aung, W.P.; Phosuk, I.; Laummaunwai, P.; Maleewong, W. Angiostrongylus Cantonensis and A. Malaysiensis Broadly Overlap in Thailand, Lao PDR, Cambodia and Myanmar: A Molecular Survey of Larvae in Land Snails. PLOS ONE 2016, 11, e0161128. [Google Scholar] [CrossRef] [PubMed]
- de Wit, L.A.; Ricketts, T.H. Trade and Deforestation Predict Rat Lungworm Disease, an Invasive-Driven Zoonosis, at Global and Regional Scales. Frontiers in Public Health 2021, 9, 680986. [Google Scholar] [CrossRef]
- Zanol, J.; Fernandez, M.A.; de Oliveira, A.P.M.; Russo, C.A. de M.; Thiengo, S.C. O Caramujo Exótico Invasor Achatina Fulica (Stylommatophora, Mollusca) No Estado Do Rio de Janeiro (Brasil): Situação Atual. Biota Neotropica 2010, 10, 447–451. [Google Scholar] [CrossRef]
- Barçante, J.M. de P.; Barçante, T.A.; Dias, S.R.C.; Lima, W. dos S. Ocorrência de Achatina Fulica Bowdich, 1822 (Mollusca: Gastropoda: Achatinoidea) No Estado de Minas Gerais, Brasil. Boletim do Museu de Biologia Mello Leitão 2005, 18, 65–70. [Google Scholar]
- Teles, H.M.S.; Fontes, L.R. Implicações Da Introdução e Dispersão de Achatina Fulica Bowdich, 1822 No Brasil. Boletim do Instituto Adolfo Lutz 2002, 12, 3–5. [Google Scholar]
- Thiengo, S.C.; Simões, R. de O.; Fernandez, M.A.; Maldonado-Júnior, A. Angiostrongylus Cantonensis and Rat Lungworm Disease in Brazil. Hawai’i journal of medicine & public health : a journal of Asia Pacific Medicine & Public Health 2013, 72, 18–22. [Google Scholar]
- Arruda, J.O.; Santos, L. First Record of Achatina Fulica Bowdich, 1822 (Mollusca, Achatinidae), for the State of Rio Grande Do Sul, Brazil. Biotemas 2022, 35, 1–6. [Google Scholar] [CrossRef]
- Borrero, F.J.; Breure, A.S.H.; Christensen, C.C.; Correoso, M.; Avila, V.M. Into the Andes: Three New Introductions of Lissachatina Fulica (Gastropoda, Achatinidae) and Its Potential Distribution in South America. Tentacle 2009, 6–8. [Google Scholar]
- Caldeira, R.L.; Mendonça, C.L.G.F.; Goveia, C.O.; Lenzi, H.L.; Graeff-Teixeira, C.; Lima, W.S.; Mota, E.M.; Pecora, I.L.; De Medeiros, A.M.Z.; Carvalho, O.D.S. First Record of Molluscs Naturally Infected with Angiostrongylus Cantonensis (Chen, 1935) (Nematoda: Metastrongylidae) in Brazil. Memórias do Instituto Oswaldo Cruz 2007, 102, 887–889. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Liu, H.-X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.-Y.; Utzinger, J.; Zhou, X.-N. Invasive Snails and an Emerging Infectious Disease: Results from the First National Survey on Angiostrongylus Cantonensis in China. PLoS Neglected Tropical Diseases 2009, 3, e368. [Google Scholar] [CrossRef]
- Tirira S., D. Guía de Campo de Los Mamíferos Del Ecuador; Murciélago Blanco: Quito, 2007; ISBN 9978-44-651-6. [Google Scholar]
- Kosoy, M.; Khlyap, L.; Cosson, J.-F.; Morand, S. Aboriginal and Invasive Rats of Genus Rattus as Hosts of Infectious Agents. Vector-Borne and Zoonotic Diseases 2015, 15, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Martini L Muzzio J, Solorzano L., G.E. Descripcion Del Primer Foco de Transmision Natural de Angiostrongylus Cantonensis En Ecuador. In Angiostrongylus cantonensis; Emergencia en América, 2016; pp. 209–220. ISBN 978-959-270-368-1.
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection From The Global Invasive Species Database. Aliens 2000, 12, 1–12. [Google Scholar] [CrossRef]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The Spread of Invasive Species and Infectious Disease as Drivers of Ecosystem Change. Frontiers in Ecology and the Environment 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Chinchio, E.; Crotta, M.; Romeo, C.; Drewe, J.A.; Guitian, J.; Ferrari, N. Invasive Alien Species and Disease Risk: An Open Challenge in Public and Animal Health. PLOS Pathogens 2020, 16, e1008922. [Google Scholar] [CrossRef]
- Červená, B.; Modrý, D.; Fecková, B.; Hrazdilová, K.; Foronda, P.; Alonso, A.M.; Lee, R.; Walker, J.; Niebuhr, C.N.; Malik, R.; et al. Low Diversity of Angiostrongylus Cantonensis Complete Mitochondrial DNA Sequences from Australia, Hawaii, French Polynesia and the Canary Islands Revealed Using Whole Genome next-Generation Sequencing. Parasites & Vectors 2019, 12, 241. [Google Scholar] [CrossRef]
- Monte, T.C.C.; Gentile, R.; Garcia, J.S.; Mota, E.; dos Santos, J.N.; Maldonado-Júnior, A. Brazilian Angiostrongylus Cantonensis Haplotypes, Ac8 and Ac9, Have Two Different Biological and Morphological Profiles. Memorias do Instituto Oswaldo Cruz 2014, 109, 1057–1063. [Google Scholar] [CrossRef]
- Tokiwa, T.; Hashimoto, T.; Yabe, T.; Komatsu, N.; Akao, N.; Ohta, N. First Report of Angiostrongylus Cantonensis (Nematoda: Angiostrongylidae) Infections in Invasive Rodents from Five Islands of the Ogasawara Archipelago, Japan. PLoS ONE 2013, 8, 1–5. [Google Scholar] [CrossRef]
- Rael, R.C.; Peterson, A.C.; Ghersi-Chavez, B.; Riegel, C.; Lesen, A.E.; Blum, M.J. Rat Lungworm Infection in Rodents across Post-Katrina New Orleans, Louisiana, USA. Emerging Infectious Diseases 2018, 24, 2176–2183. [Google Scholar] [CrossRef]
Figure 1.
Map of Ecuador showing the study area highlighting the sampled provinces.
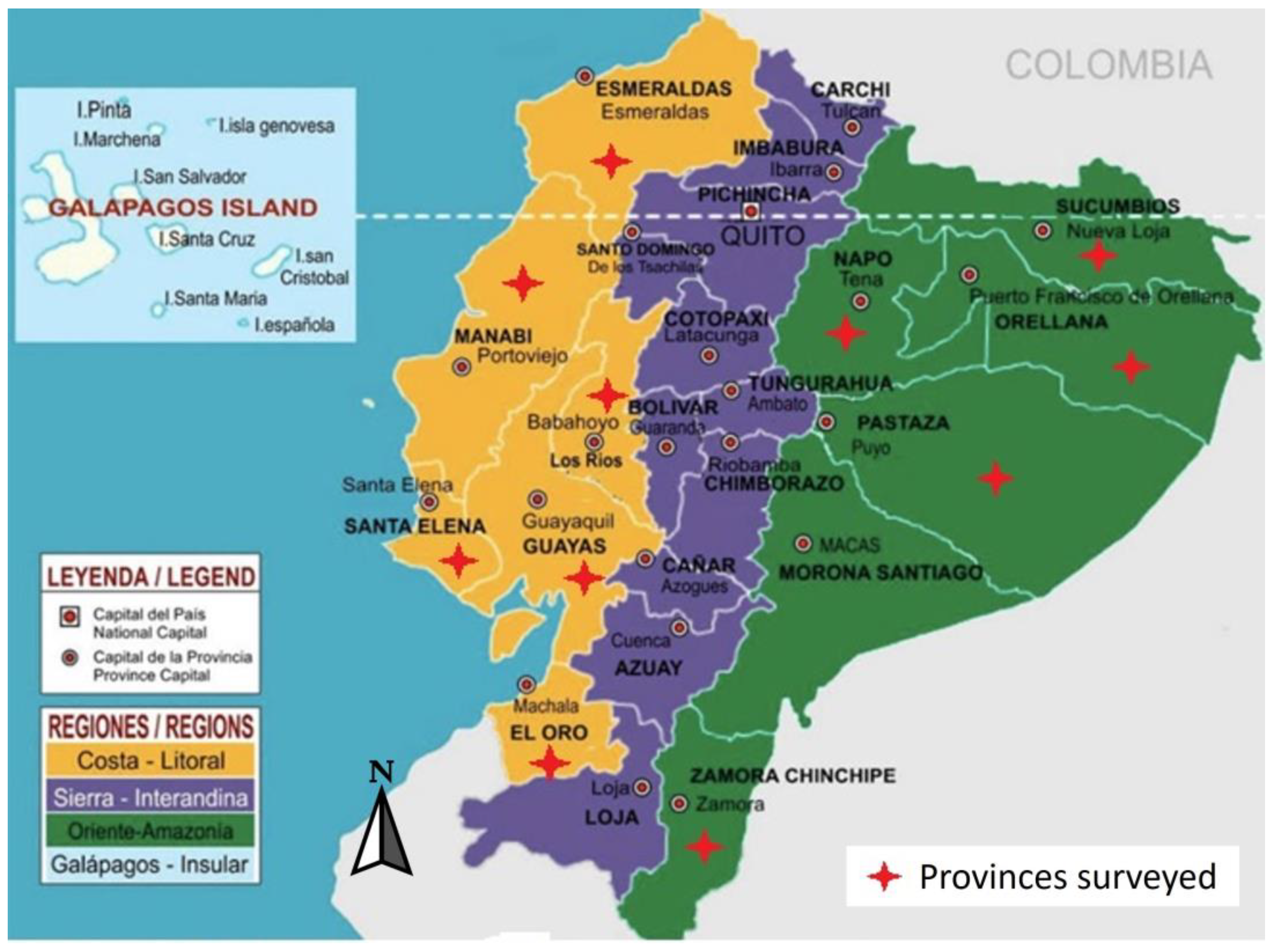
Figure 2.
Bayesian inference (BI) phylogenetic relationships of A. cantonensis specimens and outgroups unique MT-CO1 gene sequences (255 bp). The values at the nodes are BPPs (> 0.50). The scale bar is the number of substitutions per site. Sequence names are colored based on the clusters recovered in the BAPS cluster analysis (bottom right). Sequences are named AC1-17, following the names for haplotypes previously adopted [36,39,58], adding letters to variants, followed by the localities (countries) where they are found. Clusters 1-5 were recovered in the BAPS cluster analysis for the 29 sequences matrix.
Figure 2.
Bayesian inference (BI) phylogenetic relationships of A. cantonensis specimens and outgroups unique MT-CO1 gene sequences (255 bp). The values at the nodes are BPPs (> 0.50). The scale bar is the number of substitutions per site. Sequence names are colored based on the clusters recovered in the BAPS cluster analysis (bottom right). Sequences are named AC1-17, following the names for haplotypes previously adopted [36,39,58], adding letters to variants, followed by the localities (countries) where they are found. Clusters 1-5 were recovered in the BAPS cluster analysis for the 29 sequences matrix.
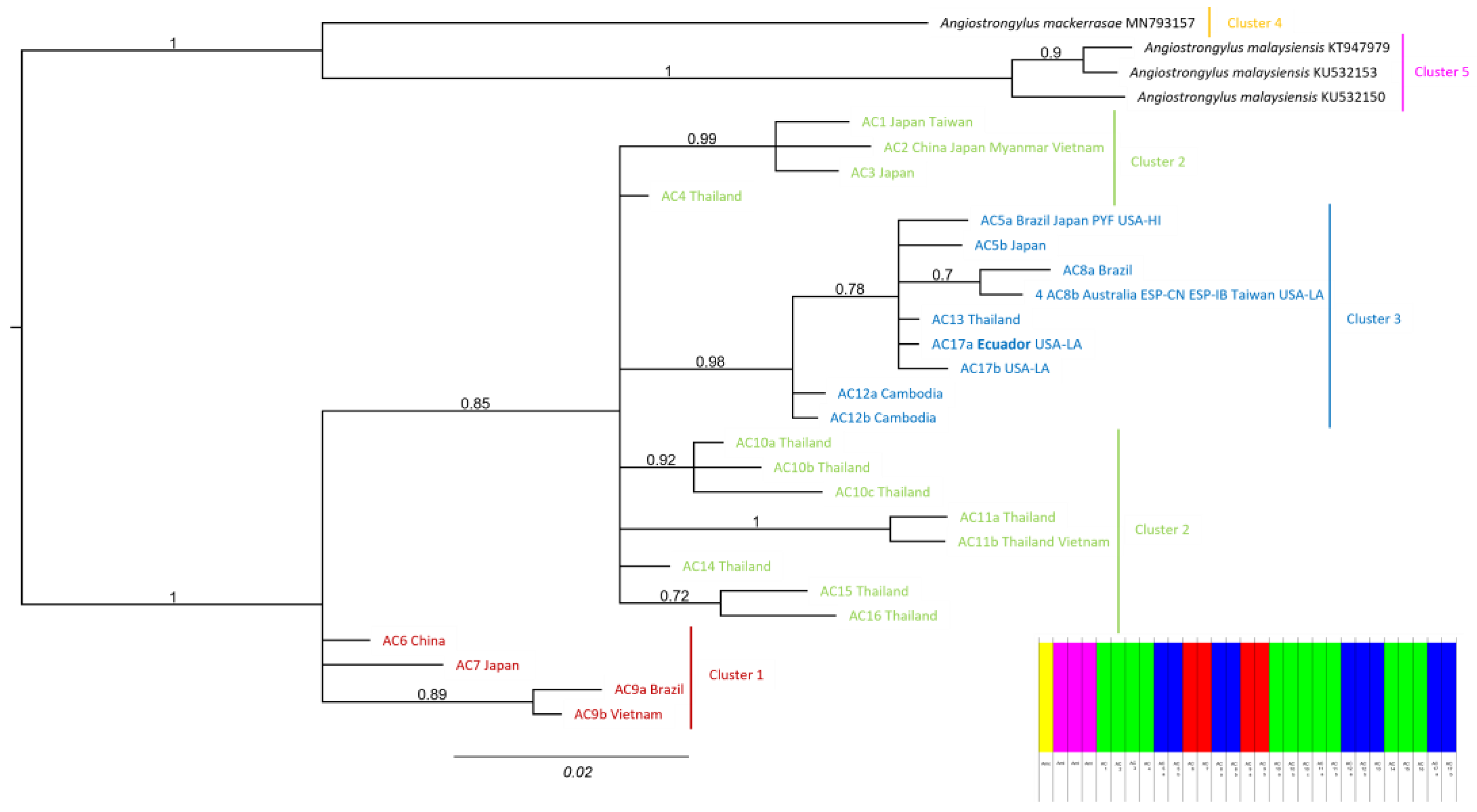
Figure 3.
Median-joining haplotype network of A. cantonensis (25 haplotypes) based on 116 partial MT-CO1 gene sequences (255 bp). The size of the circles represents the frequency of haplotypes. The colors of the circles represent the localities (countries) of occurrence of each haplotype. Black circles are median vectors. Sequences are named AC1-17, following the names for haplotypes previously adopted [36,39,58], adding letters to variants. Clusters 1-7 were recovered in the BAPS cluster analysis for the 116 sequences matrix.
Figure 3.
Median-joining haplotype network of A. cantonensis (25 haplotypes) based on 116 partial MT-CO1 gene sequences (255 bp). The size of the circles represents the frequency of haplotypes. The colors of the circles represent the localities (countries) of occurrence of each haplotype. Black circles are median vectors. Sequences are named AC1-17, following the names for haplotypes previously adopted [36,39,58], adding letters to variants. Clusters 1-7 were recovered in the BAPS cluster analysis for the 116 sequences matrix.
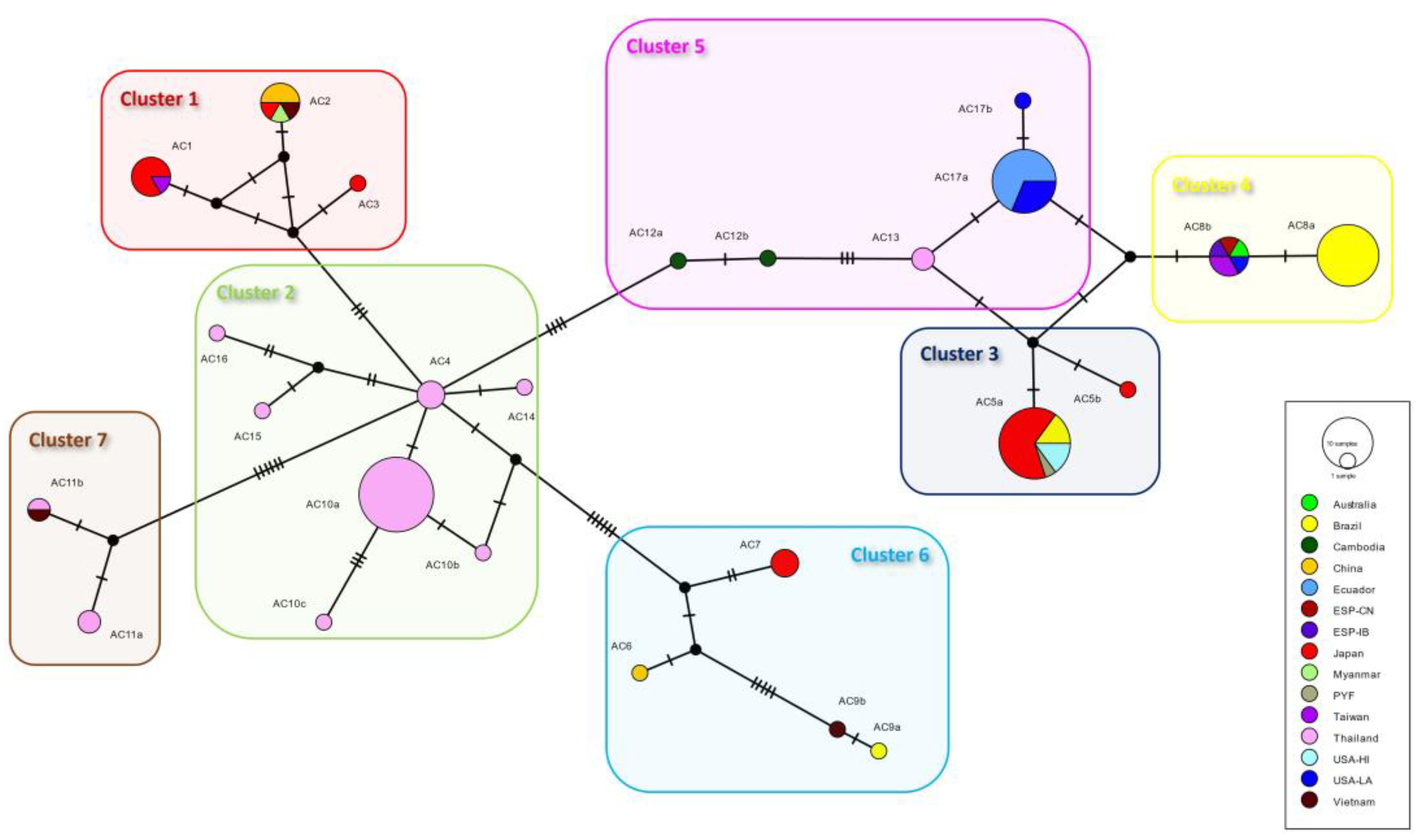

Table 1.
Identification and GenBank accession numbers of sequences obtained in this study, followed by their respective sampling localities.
Table 1.
Identification and GenBank accession numbers of sequences obtained in this study, followed by their respective sampling localities.
| Identification | GenBank accession number | Province |
| LSA-01 | MW391020 | Esmeraldas |
| LSA-02 | MW390970 | Santa Elena |
| LSA-03 | MW390971 | El Oro_ |
| LSA-04 | MW390972 | Guayas |
| LSA-05 | MW390967 | Zamora |
| LSA-06 | MW390974 | Pastaza_ |
| LSA-07 | MW390969 | Orellana |
| LSA-08 | MW390973 | Manabi |
| LSA-09 | MW390968 | Napo |
| LSA-10 | MW390966 | Los Rios |
| LSA-11 | MW390965 | Sucumbios |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



