
Submitted:
05 April 2023
Posted:
06 April 2023
Read the latest preprint version here
Abstract
Drug-resistant tuberculosis (DR-TB) remains a major public health problem in South Africa. Gene mu-tations and the genotyping of DR-TB in a rural Eastern Cape Province have not previously been studied. Hence, we sought to detect prevalent mutations linked to DR-TB profiles, identify genetic diversity, and assign lineages. Sputum specimens were obtained from 1157 patients suspected with tuberculosis. They were assayed for rifampin-resistance using Xpert® MTB/RIF.and detection of mutations conferring re-sistance to anti-TB drugs was carried out using GenoType MTBDRplus VER 2.0. Thereafter, 441 isolates were spoligotyped. The most prevalent rifampin resistance-conferring mutations were in rpoB codon S531L. The INH resistant strains, 54.5% had mutations in katG gene at codon S315TB and 24.7% with mutations in inhA gene at codons C-15TB. Furthermore 69.9% strains displayed mutations involving both rpoB and katG genes, while 24.6% strains displayed mutations involving both rpoB and inhA genes. The overall prevalence of heteroresistance was 17.9%. Spoligotyping analysis generated 410 patterns of iso-lates which were found in the international database, with Beijing family being predominant. The diversity of mutations in the study provides information for investigating the evolutional lineages of M. tuberculosis isolates. The recognition frequency of rpoB, katG and inhA mutations in different study areas may help to guide decision‑making about standardization of treatment regimens or individualized treatment in areas where these mutations have been found. This calls for increased drug resistance surveillance and rein-forcement of TB control in this area.
Keywords:
Drug-resistant TB
; Heteroresistance
; Mutations
; Spoligotyping
1. Introduction
Global public health continues to be threatened by tuberculosis (TB), an infectious disease caused by Mycobacterium tuberculosis (M. tuberculosis) and ranked among the top 10 causes of mortality globally [1]. In 2020, the estimated number of incident cases of TB stands at 9.9 million out of which Africa had a 25% share. South Africa is one of the 30 countries with a high TB burden and has the eighth highest TB incidence globally having more than 500 cases per 100,000 population, which towers above the global average incidence of 127 cases per 100,000 population [2].
In South Africa, TB continues to be a disease of major importance and remained the leading cause of death for three consecutive years that is from 2016 to 2018 [3]. In 2019 alone, an estimated 360,000 South Africans became ill with TB and 58,000 people were estimated to have died from the disease [2]. The COVID-19 pandemic's effects have undone years of progress made in reducing the number of TB deaths worldwide, with the predicted number of deaths in 2020 returning to the level of 2017 [4]. The eradication of TB by 2035, a strategic goal of World Health Organization (WHO) cannot be actualized unless the emergence of resistance in TB is seriously addressed and controlled [5].
Drug-resistant TB (DR-TB) has emerged as a major risk to global TB control. Different mutations in genes such as rpoB, katG, inhA, pncA, embB, rpsL, gyrA, ethA and rrs have been identified conferring resistance to TB first-line drugs, second-line drugs or injectables and fluoroquinolones [6,7]. Mutations in codon 507 to 533 regions of the rpoB gene, called the rifampicin-resistance determining region (RRDR) are majorly responsible for rifampicin resistance while mutations in the katG and inhA genes are associated with INH resistance. The katG and inhA mutations give rise to high-level and low-level INH resistance respectively. Although mutations in both gyrA and gyrB genes are responsible for fluoroquinolone resistance, gyrA accounts for 60%-70% of all mutations [8,9,10]. Recent research has revealed that different mutations in M. tuberculosis can confer varying levels of phenotypic resistance to anti-TB medications [6,7,11]. Consequently, the aggregation of mutations at several positions has a comprehensive effect on drug resistance [12].
In rural areas of the Eastern Cape, drug resistance and gene mutations remain major barriers to effective control and management of TB. However, there was no report on the frequency of gene mutations associated with resistance to rifampicin (RIF) and isoniazid (INH). Hence, in this study, we report the prevalence of mutations in drug resistance genes (rpoB, katG and inhA) and identified the strains and lineages of DR TB strains.
2. Materials and Methods
2.1. Study site, patient population, and specimen collection
Sputum specimens were obtained from 1157 patients enrolled from different healthcare facilities and sent for testing and analysis to the National Health Laboratory Services (NHLS) at Mthatha over a 36-month duration (January 2018 to December 2020). These specimens were collected from patients showing clinical signs of TB. The patients were enrolled from one hundred and eighteen (118) healthcare facilities located in five districts including Oliver Reginald (OR) Tambo, Alfred Nzo, Amathole, Chris Hani, and Joe Gqabi.
2.2. Xpert® MTB/RIF assay
This test was carried out directly on TB sputum samples to detect mutations in the RRDR of M. tuberculosis. After decontamination of the sputum samples, a reagent buffer containing NaOH and isopropanol was added at a ratio of 2:1, followed by incubation at room temperature for 15 min [13]. Two milliliters of the final samples were then transferred into the Xpert MTB/RIF cartridge, then loaded into the GeneXpert instrument where the testing process is fully automated. At the end of the assay, the results were reported as M. tuberculosis negative or positive and RIF resistant or susceptible.
2.3. Phenotypic drug susceptibility testing (DST)
The test was carried out on sputum samples using the automated BACTEC Mycobacterial Growth Indicator Tube (MGIT) 960 (Becton Dickinson, USA) according to the manufacturer’s instructions. The following critical concentrations for 1st and 2nd line anti-TB drugs were used Isoniazid (0.1 µg/mL of medium), Rifampin (1.0 µg/mL of medium), Ofloxacin (2.0 µg/mL), Amikacin (1.0 µg/mL), Kanamycin (2.5 µg/mL) and Capreomycin (2.5 µg/mL) [14].
Briefly, reconstitution of PANTA powder with growth supplement (15mL) was done, followed by addition to MGIT tube and inoculation of each tube with 0.5ml of the processed specimen. The tubes were incubated at 37 0C in the BACTEC MGIT 960 instrument and monitored automatically every 60 min for increased fluorescence. Culture tubes were maintained until they became positive or for a maximum of 42 days to confirm if negative. Positive samples were removed from the instrument and recorded as positive along with the number of incubation days.
2.4. Genotypic DST
GenoType MTBDR plus version 2.0 (Hain Lifescience, Nehren, Germany) is aDeoxyribonucleic acid (DNA) strip-based method designed for simultaneous detection of the most important rpoB mutations, which confer RIF resistance and katG and inhA mutations, which confer high-level and low-level INH resistance, respectively [14] was used in this study. Three procedures namely DNA extraction, multiplex amplification with biotinylated primers, and reverse hybridization were performed following the manufacturer's instructions [15].
DNA extraction was performed using a Genolyse kit (Hain Lifescience, Nehren, Germany). Briefly, 1 ml of the decontaminated sputum was centrifuged for 15 min at 10000×g, and the supernatant was discarded. The sample was incubated at 950C for 5 minutes after the addition of 100 μl of lysis buffer in order to re-suspend the sediment. Subsequently, 100 μl of neutralization buffer was added, vortexed, and centrifuged for 5 minutes [16]. Multiplex amplification was carried out in different cycles at different temperatures as follows: 1 cycle of 15 min at 95°C, 20 cycles each of 30 s at 95°C and 2 min at 65°C, 30 cycles each of 25 s at 95°C, 40 s at 50°C, and 40 s at 70°C, and finally 1 cycle of 8 min at 70°C. Reverse hybridization was performed using an automated hybridization system, Auto-Lipa 48 system (Innogenetics) following the manufacturer’s instructions [15].
2.5. Evaluation and interpretation of results
The developed strips were pasted on evaluation sheets provided with the kit in the designated fields by aligning the conjugate control and amplification control bands with the respective lines on the sheet. With the aid of GenoLyse package inserts, the mutation bands were interpreted; the absence of a wild-type band and the presence of a mutant band for a specific gene on the strip implied resistance.
2.6. Spoligotyping
Out of 1157 isolates, randomly selected 441 isolates were assayed by spoligotyping to determine genotypes of M. tuberculosis isolates. The samples were heat-killed and subjected to DNA extraction. Spoligotyping was performed using microbeads from TB-SPOL Kit (Beamedex®, Orsay, France) and the fluorescence intensity was measured using Luminex 200® (Austin, TX) following the manufacturer’s instructions. The hybridization patterns were translated into binary and octal formats. The generated binary codes of the isolates were entered into the SITVIT2 database of the Pasteur Institute of Guadeloupe and assigned specific spoligotype international type (SIT) [17]. Quality control was ensured by using M. tuberculosis H37Rv as a positive control.
3. Results
3.1. Profile of the isolates
A total of 1157 clinical isolates were analyzed over three years: 2018, 2019, and 2020 (Table 1), of which 950 (82.1%) were drug-resistant (DR-TB, multidrug-resistant TB (MDR-TB), extensively drug-resistant TB (XDR – TB)) and 207 (17.9%) were heteroresistant isolates. Table 1 displays the number of M. tuberculosis isolates over a 3-year period. The heteroresistance rate increased over time as displayed in Table 1.
3.2. Mutations in the rpoB, katG, and inhA genes
Among 1157 M. tuberculosis clinical isolates, rpoB531 and katG315 mutations were the most common mutations associated with RIF and INH resistance, respectively. Among RIF- resistant strains, known mutations in rpoB were found in 761 (65.8%), wild type was found in 381 strains while missing mutation accounted for the remaining 15 strains. The most prevalent RIF resistance-conferring mutations were in rpoB codons S531L (70.2%). Others included D516V (18.3%), H526D (3.8%), and H526Y (3.4%) (Figure 1). Out of 761 isolates, 532 (69.9%) isolates displayed mutations involving both rpoB and katG genes while 187 (24.6%) isolates displayed mutations involving both rpoB and inhA genes.
The INH-resistant strains totaled 916 isolates with mutations in the katG and inhA genes accounting for 630 and 286 isolates respectively. Mutations in the katG gene which are associated with a high level of INH resistance occurred in codon S315TB of the 630 (68.8%) (Figure 2); mutations in inhA which are associated with a low level of INH resistance were found in 286 (31.2%) isolates and were detected in codons C-15TB (88.5%), T8A (8.4%), T-8C (2.1%), A-16G (0.7%) with double mutations in codons A-16G and T8A (0.3%) (Figure 3).
Combined mutations involving either rpoB and katG genes or rpoB and inhA genes out of 761 RIF-resistant strains are shown in Table 2 below.
3.3. Heteroresistance mutations
Of 1157 DR-TB isolates, the overall prevalence of heteroresistance was 17.9% (n=207). This rate increased over time, ranging from 9.6% in 2018 to 32.2% in 2020. Heteroresistance to RIF was found in 58 (28%) cases whilst heteroresistance to INH was determined in 119 (57.5%) cases, of which 70 (58.8%) and 49 (41.2%) cases were katG and inhA-associated heteroresistance, respectively. In addition, combined heteroresistance to RIF and INH was found in 30 (14.5%) DR-TB isolates.
3.4. Spoligotyping
Of 441 isolates spoligotyped, 437 revealed distinct spoligotype patterns. Patterns of 410 (93.1%) isolates matched a pre-existing SIT in the SITVIT2 database, while 27 unique patterns (6.1%) were not in the database (Table 3).
4. Discussion
Spontaneous chromosomal mutations in particular locations of the bacterial genome is generally thought to be the cause of resistance to RIF and INH which several studies have proven [9]. Information associated with M. tuberculosis strain distribution, transmission levels (recent and previous) and transmission dynamics of DR strains is unknown in rural Eastern Cape and according to our knowledge, this is the first study detailing the distribution of drug-resistant genes, mutation sites and genotypes.
Mutations in M. tuberculosis mostly caused by rpoB, katG and inhA genes giving rise to MDR-TB is a cause of concern for clinicians [18]. RIF resistance is usually regarded as a marker for identifying MDR-TB. Hence, screening mutations in candidate genes constitute the foremost significant step to making a definite diagnosis in drug-resistant strains. The nucleotide sequences of the rpoB gene were analyzed for mutations in the 81 bp core region. The prevalence of mutations in the rpoB gene in this study was higher than in the other genes (katG and inhA genes). Analysis of the RIF-associated mutations revealed a prevalence of 65.8% of the rpoB gene. This is comparable to the study done by Otchere et al., with a prevalence of 52% [19]. Conversely, the studies of [9,20,21] reported a prevalent mutation of 93.5%, 94.9%, and 91.2% respectively in katG gene which is higher than what was obtained in this study. In this study, the most prevalent rifampin resistance-conferring mutations in rpoB occurred in S531L codon which is comparable with previous studies [9,20,22,23], and could be due to the spread of a prevalent clone. Mutation in this codon is identified as a hot spot for rpoB gene mutations in M. tuberculosis and has been reported in other Provinces of South Africa as well [24] showing that these mutations are prevalent in the country. This high frequency of occurrence may be due to the low fitness cost associated with rpoB S531L [24] and have been associated with major MDR-TB outbreaks. [23]. Low frequency of mutation was observed in codon 526 at 3.8% in this study but higher in Uganda at 12.5% [25]; China at 14.9% [20]; Brazil at 9.9%, [26] indicating that frequency varies with different geographic locations. The co-occurrence rate of codons 526D and 531 in this study (2.8%) was lower than the rate reported in Iran (23.9%) [27].
INH is a first-line TB drug and a very important drug for the treatment of TB due to its early strong bactericidal activity. The detection of mutations in the katG or inhA promoter area is important and relevant to clinicians because it helps forecast the level of resistance to INH, and will determine the most appropriate treatment regimen used [28]. The catchment areas of the above-mentioned clinics need to be monitored for the changes occurring during the course of patient TB treatment as it shows a high level of resistance to INH drug. This study had both high-level resistant INH strains and the katG S315T gene mutation. The majority of mutations occur in codon 315 in 30%-90% of INH-resistant strains [16]. The findings of this study supported this assertion, demonstrating mutations at S315T linked with elevated levels of drug resistance to INH. A similar trend of mutation in codon S315T of katG gene have been reported in other countries including Zambia and Brazil [9,29]. Mutations in katG occurred only in codon agc/acc S315Tb in this study but the study of Jagielski et al. [30] found that mutations occurred in eleven other codons. The prevalence of katG S315T varied according to the geographic region: Sub-Saharan Africa (94.9%) [9], West Africa (64%) [31], Southeast Asia (29.3%) [32], and the United States (38%) [33] while the global frequency of katG S315 is estimated at 64.2% [34]. In addition to the katG gene, other genes associated with INH resistance of M. tuberculosis, include inhA, fabG1 and oxyR-ahpC. Mutations in the inhA promoter region have been found to occur in 20–42% of INH-resistant strains [33,34]. Previous studies have shown that mutations in the promoter region of the inhA gene confer low-level resistance to INH and is defined by the range of 8% to 43% [35]. In this study, low-level resistance ratio was 31.2%, close to the high limit of the reported range. Other studies have reported different proportions of inhA mutations in other locations of South Africa; Kwazulu Natal (27.5%) [36], and Free State (13.4%) [8]. On the contrary, a relatively low frequency (0.8% and 2.0%) of mutations in the inhA promoter region among INH-resistant strains was observed in Ethiopia and Zambia respectively [9,37]. Contrarily, Lempens et al. [38], reported a lower percentage of occurrence of C-15T gene mutation with a high level of INH resistance. This suggests that mutation of the inhA gene does not always indicate a low resistance level. Due to the common target of action, mutations in the inhA gene also confer cross-resistance to ethionamide (ETH), a second-line drug used in MDR therapy, and thus considered as a surrogate marker for early detection of ETH resistance [8,39,40]. Previously, the National Department of Health (NDoH) in South Africa included ETH in the treatment regimen for MDR-TB in South Africa. Due to this cross-resistance, using ETH to treat MDR-TB in the context of inhA mutations would not have been effective [8]. Consequently, in the clinical management of MDR-TB cases displaying inhA mutations, ETH must be excluded from the regimen The most common mutation in the inhA gene was found in C-15TB (88.5%) of our isolates corroborating the findings of Seifert et al., [41], who reported that the most common mutation in inhA gene was frequently observed in C-15TB.
The clinical and molecular characteristics of the M. tuberculosis strains vary in different areas. This was observed in this study in consonance with the findings of Liu et al. [12]. The diversity of mutations can also provide information for investigating the evolutional lineages of M. tuberculosis. The recognition frequency of rpoB, katG and inhA mutations in different areas of Mthatha may help to guide decision-making about standardization of treatment regimens or individualized treatment in each area where these mutations have been found. There is an indication of M. tuberculosis strains that are constantly mutating as we observed combined mutations (Table 2). This data can be used in the development of new anti-TB drugs.
Heteroresistance in the study area increased with time, by the third year of the study period, the heteroresistance rate was almost tripled that of the previous year (Table 1). The rpoB and katG combination had the highest number of heteroresistant isolates followed by rpoB and inhA combination. In rpoB and katG combination, the mutation regions rpoB S315L and katG 531ST had the highest number of isolates (Table 2). Under the selective pressure of inadequate anti-TB medication, separation into susceptible and resistant organisms most likely elucidates heteroresistance caused by infection with single strains. Several reports have documented the development of resistance as a result of insufficient treatment [42]. It has also been established that treatment outcomes can be negatively impacted by mixed-strain infections involving heteroresistant pathogens. Due to heteroresistance, the drug-resistant strain may go undetected, and treating such cases with first-line anti-tuberculosis medication may select for and amplify the drug-resistant strain in the host [43]. The rate of 17.9% of heteroresistance in this study is similar to the finding by Rinder et al. [44], who reported a rate of 17%. Other studies have reported significantly lower rates [45]. Heteroresistance was caused by a single strain of either the Beijing, LAM or X genotype, suggesting that splitting of a single strain into susceptible and resistant organisms is high likely the predominant underlying mechanism.
The M. tuberculosis population in this study area was genetically diverse. From the 441 clinical isolates, 23 spoligotypes were observed and classified into major M. tuberculosis lineages; lineages 1, 2, 3, and 4 as shown in Table 3. The population structure of Rifampicin resistant TB (RRTB) isolates in South Africa is dominated by Beijing and Euro-American (LAM, T, S, and X) strains which can be explained by the historical movement of strains as South Africa was located in a geographically central position in the historical trade route between East and West for hundreds of years [46].
One of the most widespread genotypes of M. tuberculosis found globally is the Beijing family, often known as lineage 2. It is usually linked to immune evasion and antibiotic resistance, which promotes rapid bacterial replication, spread, and transmission [47]. The Beijing family, which is more transmissible than other families [48] were prevalent in this study (42%). This lineage has been detected in studies reported from other parts of South Africa including Limpopo, Western Cape, and Mpumalanga [49,50]. According to Said et al. [49] Beijing family is predominant in the Eastern Cape followed by LAM. According to the study of Chihota et al. [51] based on the review of the repository and databases, South African M. tuberculosis strains revealed the greatest diversity and greatest abundance of Beijing families. Furthermore, the association between HLA-B27 and host-pathogen compatibility has accounted for the success of the Beijing lineage in South Africa [51]. Given the growing concern over the prevalence of Beijing strains and their success in evolving to fit into various human groups, suitable measures should be implemented for public health surveillance. The knowledge of the lineages circulating in the study area will help in understanding the drivers of drug resistance and their impact on treatment outcomes and management of TB transmission.
According to van Soolingen et al. [52] Beijing lineage was described for the first time in East Asian countries with a specific spoligotype pattern characterized by the presence of the last 10 spacers (spacers 34– 43). Historical evidence supported by genetic data established that the Beijing strains were historically recently brought to South Africa, not directly from its primary origin (China), but from the secondary one (Indonesia) [46]. Although varying theories have been proposed to support the introduction of Beijing family into Africa; Pokam et al. [53] opined that the dominance of the Beijing family is connected with the current influx of migration of the Asian population to the African region; as well as increased trade relations of Africans returning from travel to China for business, leading to the spread in the continent. It is important to actively carry out surveillance of the Beijing family to verify its heightened transmission and understand its importance in the management plan of TB in this area.
The LAM lineage (18.8%) was the subsequent most predominant in our study. This is not surprising considering the fact that this genotype has been discovered to be widespread in Eastern Cape and Free State Provinces [54,55]. This suggests there is a continuous TB transmission strain still ongoing throughout the province. On the other hand, the LAM genotype is prevalent in KwaZulu Natal, Northern Cape and Gauteng [49] and some other neighboring countries in Southern Africa, including Zambia and Zimbabwe [56,57]; but with least prevalence in Western Cape [49] The LAM strain is predominant in KwaZulu Natal, which is a neighboring Province to Eastern Cape; this might eventually bring it to par with the predominant Beijing strain thereby causing more havoc. Hence, continuous surveillance of the genetic diversity must be carried out to profile these strains. Concerning delineation of the LAM sub-lineages, five were found in our study namely LAM 3, 4, 5, 9 and LAM II-ZWE with LAM 3 being predominant at 74.7%. On the other hand, another study in South Africa reported that out of the 12 sub-lineages delineated globally, six sub-lineages were found of which LAM4 was the most predominant [54]. LAM11-ZWE has been reported to be a dominant subfamily in Zambia and Zimbabwe [56] whose origin has been traced to Portugal [51]. Two strains belonging to LAM 3 sub-lineage did not match any in the SITVIT2 database and were classified as orphans. LAM 3 in our study, corresponding to SIT33 and SIT 719 represented 8.2% and 3.9% respectively while the result obtained by Maguga-Phasha et al. [50] in Limpopo had LAM 3 (7.0%) corresponding to SIT 33 only. LAM 1, LAM 2 and LAM 6 were not reported in this study but reported elsewhere [29].
5. Conclusions
The identification of areas where DR-TB is concentrated in rural areas where TB is a burden could assist policy makers to implement targeted interventions aimed at the prevention and management of TB transmission. This is particularly important in resource-limited settings and in high DR-TB burden areas like rural areas of Mthatha and environs. Targeted interventions to rural communities are vital as these areas find it impossible to provide DR-TB services across the communities as the diagnosis and treatment of DR-TB is challenged by many factors such as co-infection with HIV and poor adherence to treatment.
Little is known about the effects of distribution on antibiotic-resistance conferring mutations on the bacterial fitness and spoligotypes of M. tuberculosis, therefore, knowledge about the distribution of gene mutations and spoligotypes among treating physicians and the value of reporting these mutations by the laboratories is of high importance. Genetic mutations are responsible for the high rate of resistance observed in the treatment of tuberculosis, hence continuous surveillance is recommended in this study area. The prevention and control of the spread of these drug-resistant strains depend in large part on the early detection of drug-resistant tuberculosis.
Author Contributions
Conceptualization, L.M.F.; methodology, L.M.F.; formal analysis, L.M.F, AD, SO and RW.; investigation, L.M.F.; resources, L.M.F. and RW; data curation, A.D. and S.O.; writing—original draft preparation, L.M.F, NS and M.C.H.; writing—review and editing, L.M.F, N.S and M.C.H.; visualization, L.M.F. and M.C.H.; supervision, R.M.W., S.V., and T.A.; project administration, L.M.F.; funding acquisition, L.M.F. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for this study was obtained from the South African Medical Research Council (SAMRC) Research development grant (Pilot grant).
Institutional Review Board Statement
This study was conducted in accordance with the Decla-ration of Helsinki and approved by the Research Ethics and Biosafety Committee of the Faculty of Health Sciences of Walter Sisulu University (Ref. No. 026/2019) and Eastern Cape Department of Health (Ref No EC_201904_011).
Informed Consent Statement
Not applicable. This study used routine samples received in National Health Laboratory Services (NHLS) TB laboratory.
Data Availability Statement
Data will be made available upon request from the corresponding author.
Acknowledgments
The authors are grateful to the NHLS TB laboratory staff and participating clinics for their support during sample analysis and data collection.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bwalya, P.; Yamaguchi, T.; Solo, E.S.; Chizimu, J.Y.; Mbulo, G.; Nakajima, C.; Suzuki, Y. Characterization of Mutations Associated with Streptomycin Resistance in Multidrug-Resistant Mycobacterium tuberculosis in Zambia. Antibiotics 2021, 10, 1169. [CrossRef]
- World Health Organization. Global tuberculosis report, 2020 World Health Organization. Geneva, Switzerland, available at https://apps.who.int/ iris/handle/10665/336069 [accessed August 6, 2022].
- Statistics South Africa. Mortality and causes of death in South Africa. Findings from death notification. Pretoria, South Africa. Stats SA’s June 2021 report [accessed 21 January 2022].
- World Health Organization. Global tuberculosis report 2021: supplementary material [accessed August 8, 2022].
- Kabir, S.; Junaid, K.; Rehman, A. Variations in rifampicin and isoniazid resistance associated genetic mutations among drug naïve and recurrence cases of pulmonary tuberculosis. Int. J. Infect. Dis. 2021, 103, 56-61. [CrossRef]
- Libiseller-Egger, J.; Phelan, J.; Campino, S.; Mohareb, F.; Clark, T.G. Robust detection of point mutations involved in multidrug-resistant Mycobacterium tuberculosis in the presence of co-occurrent resistance markers. PLoS Comput. Biol. 2020, 16(12), e1008518. [CrossRef]
- Wan, L.; Liu, H.; Li, M.; Jiang, Y.; Zhao, X.; Liu, Z.; Wan, K.; Li, G.; Guan, C-x. Genomic Analysis Identifies Mutations Concerning Drug-Resistance and Beijing Genotype in Multidrug-Resistant Mycobacterium tuberculosis Isolated From China. Front. Microbiol. 2020, 11, 1444. [CrossRef]
- Pitso, L.; Potgieter, S.; Van der Spoel van Dijk, A. Prevalence of isoniazid resistance-conferring mutations associated with multidrug-resistant tuberculosis in Free State Province, South Africa. S Afr. Med. J. 2019, 109(9), 659-664. [CrossRef]
- Solo, E.S.; Nakajima, C.; Kaile, T.; Bwalya, P.; Mbulo, G.; Fukushima, Y.; Chila, S.; Kapata, N.; Shah, Y.; Suzuki, Y. Mutations in rpoB and katG genes and the inhA operon in multidrug-resistant Mycobacterium tuberculosis isolates from Zambia. J. Glob. Antimicrob. Res. 2020, 22, 302-307. [CrossRef]
- Diriba, G.; Kebede, A.; Tola, H.H.; Alemu, A.; Yenew, B.; Moga, S.; Addise, D.; Mohammed, Z.; Getahun, M.; Fantahun, M.; Tadesse, M. Utility of line probe assay in detecting drug resistance and the associated mutations in patients with extra pulmonary tuberculosis in Addis Ababa, Ethiopia. SAGE Open Med. 2022, 10, 20503121221098241.
- Valafar, S.J. Systematic review of mutations associated with isoniazid resistance points to continuing evolution and subsequent evasion of molecular detection, and potential for emergence of multidrug resistance in clinical strains of Mycobacterium tuberculosis. Antimicrob. Agents. Chem. 2021, 65(3), e02091-20. [CrossRef]
- Liu, L.; Zhao, X.; Wu, X.; Li, S.; Liu, B.; Rajaofera, M.J.; Zeng, Y.; Dong, S.; Bei, Z.; Pei, H.; Xia, Q. Prevalence and molecular characteristics of drug-resistant Mycobacterium tuberculosis in Hainan, China: from 2014 to 2019. BMC. Microbiol. 2021, 21, 185. [CrossRef]
- Bodmer, T.; Ströhle, A. Diagnosing pulmonary tuberculosis with the Xpert MTB/RIF test. J.Vis. Exp. 2012, 62, e3547.
- World Health Organization. The impact of the roll-out of rapid molecular diagnostic testing for tuberculosis on empirical treatment in Cape Town, South Africa, Bulletin WHO, 2017, 95, 545-608.
- Hain Lifescience. Company history and product releases. 2016. https://www.hain-lifescience.de/en/company/history.html. (Accessed 11 February 2021).
- Ogari, C.O.; Nyamache, A.K.; Nonoh, J.; Amukoye, E. Prevalence and detection of drug resistant mutations in Mycobacterium tuberculosis among drug naïve patients in Nairobi, Kenya. BMC Infect. Dis. 2019, 19, 279. [CrossRef]
- Couvin, D.; David, A.; Zozio, T.; Rastogi, N. Macro-geographical specificities of the prevailing tuberculosis epidemic as seen through SITVIT2, an updated version of the Mycobacterium tuberculosis genotyping database. Infect. Genet.Evol. 2019, 72, 31-43. [CrossRef]
- Bhembe, N.L.; Green, E. Characterization of mutations in the rpoB gene conferring rifampicin resistance in Mycobacterium tuberculosis complex isolated from lymph nodes of slaughtered cattle from South Africa. Braz. J. Microbiol. 2020, 51(4), 1919-1927. [CrossRef]
- Otchere, I.D.; Asante-Poku, A.; Osei-Wusu, S.; Baddoo, A.; Sarpong, E.; Ganiyu, A.H.; Aboagye, S.Y.; Forson, A.; Bonsu, F.; Yahayah, A.I.; Koram, K. Detection and characterization of drug-resistant conferring genes in Mycobacterium tuberculosis complex strains: a prospective study in two distant regions of Ghana. Tuberculosis (Edinb). 2016, 99, 147-154. [CrossRef]
- Jia, H.; Xu, Y.; Sun, Z. Analysis on Drug-Resistance-Associated Mutations among Multidrug-Resistant Mycobacterium tuberculosis Isolates in China. Antibiotics. 2021, 10, 1367. [CrossRef]
- Isakova, J.; Sovkhozova, N.; Vinnikov, D.; Goncharova, Z.; Talaibekova, E.; Aldasheva, N.; Aldashev, A. Mutations of rpoB, katG, inhA and ahp genes in rifampicin and isoniazid-resistant Mycobacterium tuberculosis in Kyrgyz Republic. BMC Microbiol. 2018, 18, 22. [CrossRef]
- Uddin, M.K.; Rahman, A.; Ather, M.F.; Ahmed, T.; Rahman, S.M.; Ahmed, S.; Banu, S. Distribution and frequency of rpoB mutations detected by Xpert MTB/RIF assay among Beijing and non-Beijing rifampicin resistant Mycobacterium tuberculosis isolates in Bangladesh. Infect. Drug Resist. 2020, 13, 789. [CrossRef]
- Meftahi, N.; Namouchi, A.; Mhenni, B.; Brandis, G.; Hughes, D.; Mardassi, H. Evidence for the critical role of a secondary site rpoB mutation in the compensatory evolution and successful transmission of an MDR tuberculosis outbreak strain. J. Antimicrob. Chemother. 2016, 71, 324-332.
- Evans, J.; Stead, M.C.; Nicol, M.P.; Segal, H. Rapid genotypic assays to identify drug-resistant Mycobacterium tuberculosis in South Africa. J. Antimicrob. Chemother. 2009, 63, 11-16. [CrossRef]
- Ssengooba, W.; Meehan, C.J.; Lukoye, D.; Kasule, G.W.; Musisi, K.; Joloba, M.L.; Cobelens, F.G.; de Jong, B.C. Whole genome sequencing to complement tuberculosis drug resistance surveys in Uganda. Infect. Genet. Evol. 2016, 40, 8-16. [CrossRef]
- Salvato, R.S.; Schiefelbein, S.; Barcellos, R.B.; Praetzel, B.M.; Anusca, I.S.; Esteves, L.S.; Halon, M.L.; Unis, G.; Dias, C.F.; Miranda, S.S.; de Almeida, I.N.. Molecular characterisation of multidrug-resistant Mycobacterium tuberculosis isolates from a high-burden tuberculosis state in Brazil. Epidemiol. Infec. 2019, 147.
- Tajbakhsh, A.; Ghasemi, F.; Mirbagheri, S.Z.; Heravi, M.M.; Rezaee, M.; Meshkat, Z. Investigation of the rpoB mutations causing rifampin resistance by rapid screening in Mycobacterium tuberculosis in North-East of Iran. Iran. J. Pathol. 2018, 13(4), 429.
- Bollela, V.R., Namburete, E.I.; Feliciano, C.S.; Macheque, D.; Harrison, L.H.; Caminero, J.A. Detection of katG and inhA mutations to guide isoniazid and ethionamide use for drug-resistant tuberculosis. Int. J. Tuberc. Lung Dis. 2016, 20(8), 1099-1104. [CrossRef]
- Verza, M.; Scheffer, M.C.; Salvato, R.S.; Schorner, M.A.; Barazzetti, F.H.; Machado, H.D.; Medeiros, T.F.; Rovaris, D.B.; Portugal, I.; Viveiros, M.; Perdigão, J. Genomic epidemiology of Mycobacterium tuberculosis in Santa Catarina, Southern Brazil. Sci. Rep. 2020, 10, 12891.
- Jagielski, T.; Grzeszczuk, M.; Kamiński, M.; Roeske, K.; Napiórkowska, A.; Stachowiak, R.; Augustynowicz-Kopeć, E.; Zwolska, Z.; Bielecki, J. Identification and analysis of mutations in the katG gene in multidrug-resistant Mycobacterium tuberculosis clinical isolates. Adv. Resp. Med. 2013, 81(4), 298-307. [CrossRef]
- Abanda, N.N.; Djieugoué, J.Y.; Lim, E.; Pefura-Yone, E.W.; Mbacham, W.; Vernet, G.; Penlap, V.M.; Eyangoh, S.I.; Taylor, D.W.; Leke, R.G. Diagnostic accuracy and usefulness of the Genotype MTBDRplus assay in diagnosing multidrug-resistant tuberculosis in Cameroon: A cross-sectional study. BMC infect. Dis. 2017, 17, 379. [CrossRef]
- Tseng, S.T.; Tai, C.H.; Li, C.R.; Lin, C.F.; Shi, Z.Y. The mutations of katG and inhA genes of isoniazid-resistant Mycobacterium tuberculosis isolates in Taiwan. J. Microbiol. Immunol. Infect. 2015, 48(3), 249-255. [CrossRef]
- Charoenpak, R.; Santimaleeworagun, W.; Suwanpimolkul, G.; Manosuthi, W.; Kongsanan, P.; Petsong, S.; Puttilerpong, C. Association between the phenotype and genotype of isoniazid resistance among Mycobacterium tuberculosis isolates in Thailand. Infect. Drug Resist. 2020, 13, 627.
- Norouzi, F.; Moghim, S.; Farzaneh, S.; Fazeli, H.; Salehi, M.; Esfahani B.N. Significance of the coexistence of non-codon 315 katG, inhA, and oxyR-ahpC intergenic gene mutations among isoniazid-resistant and multidrug-resistant isolates of Mycobacterium tuberculosis: a report of novel mutations. Pathog. Glob. Health. 2022, 116(1), 22-29.
- Tessema, B.; Beer, J.; Emmrich, F.; Sack, U.; Rodloff, A.C. Analysis of gene mutations associated with isoniazid, rifampicin and ethambutol resistance among Mycobacterium tuberculosis isolates from Ethiopia. BMC Infect. Dis. 2012, 12, 37.
- Gliddon, H.D.; Frampton, D.; Munsamy, V.; Heaney, J.; Pataillot-Meakin, T.; Nastouli, E.; Pym, A.S.; Steyn, A.J.; Pillay, D.; McKendry, R.A. A Rapid Drug Resistance Genotyping Workflow for Mycobacterium tuberculosis, Using Targeted Isothermal Amplification and Nanopore Sequencing. Microbiol. Spectr. 2021, 9(3), e00610. [CrossRef]
- Abate, D.; Tedla, Y.; Meressa, D.; Ameni, G. Isoniazid and rifampicin resistance mutations and their effect on second-line anti-tuberculosis treatment. Int. J. Tuberc. Lung Dis. 2014, 18(8), 946-951. [CrossRef]
- Lempens, P.; Meehan, P.J.; Vandelannoote, K.; Fissette, K.; de Rijk, P.; Van Deun, A.; Rigouts, L.; de Jong, B.C. Isoniazid resistance levels of Mycobacterium tuberculosis can largely be predicted by high confidence resistance-conferring mutations. Sci. Rep. 2018, 8:3246. [CrossRef]
- Sarin, R.; Bhalla, M.; Kumar, G.; Singh, A.; Myneedu, V.P.; Singhal, R. Correlation of inhA mutations and ethionamide susceptibility: Experience from national reference center for tuberculosis. Lung India. 2021, 38(6), 520-523. [CrossRef]
- Maitre, T.; Morel, F.; Brossier, F.; Sougakoff, W.; Jaffre, J.; Cheng, S.; Veziris, N.; Aubry, A.; NRC-MyRMA. How a PCR Sequencing Strategy Can Bring New Data to Improve the Diagnosis of Ethionamide Resistance. Microorganisms 2022, 10(7), 1436. [CrossRef]
- Seifert, M.; Catanzaro, D.; Catanzaro, A.; Rodwell, T.C. Genetic Mutations Associated with Isoniazid Resistance in Mycobacterium tuberculosis: A Systematic Review. PLoS ONE, 2015, 10(3), e0119628. [CrossRef]
- Hofmann-Thiel, S.; van Ingen, J.; Feldmann, K.; Turaev, L.; Uzakova, G.T.; Murmusaeva, G.; van Soolingen, D.; Hoffmann, H. Mechanisms of heteroresistance to isoniazid and rifampin of Mycobacterium tuberculosis in Tashkent, Uzbekistan. Eur. Respir. J. 2009, 33, 368–374. [CrossRef]
- Shin, S.S.; Modongo, C.; Baik, Y.; Allender, C.; Lemmer, D.; Colman, R.E.; Engelthaler, D.M.; Warren, R.M.; Zetola, N.M. Mixed Mycobacterium tuberculosis–strain infections are associated with poor treatment outcomes among patients with newly diagnosed tuberculosis, independent of pretreatment heteroresistance. J. Infect. Dis. 2018, 218(12), 1974-1982.
- Rinder, H.; Mieskes, K.T.; Löscher, T. Heteroresistance in Mycobacterium tuberculosis. Int. J. Tuberc. Lung Dis. 2001, 5(4), 339-345.
- de Assis Figueredo, L.J.; de Almeida, I.N.; Augusto, C.J.; Soares, V.M.; Suffys, P.N.; da Silva Carvalho, W.; de Miranda, S.S. Characterization of Mycobacterium tuberculosis heteroresistance by genotyping. Int. J. Mycobacteriol. 2020, 9(4), 368-372. [CrossRef]
- Said, H.; Ratabane, J.; Erasmus, L.; Gardee, Y.; Omar, S.; Dreyer, A.; Ismail, F.; Bhyat, Z.; Lebaka, T.; van der Meulen, M.; Gwala, T.; Adelekan, A.; Diallo, K.; Ismail, N. Distribution and Clonality of drug-resistant tuberculosis in South Africa. BMC Microbiol. 2021, 21, 157. [CrossRef]
- Mokrousov, I.; Ly, H.M.; Otten, T.; Lan, N.N.; Vyshnevskyi, B.; Hoffner, S.; Narvskaya, O. Origin and primary dispersal of the Mycobacterium tuberculosis Beijing genotype: clues from human phylogeography. Genome Res. 2005, 15(10), 1357-1364. [CrossRef]
- María Irene, C.C.; Juan Germán, R.C.; Gamaliel, L.L.; Dulce Adriana, M.E.; Estela Isabel, B.; Brenda Nohemí, M.C.; Payan Jorge, B.; Zyanya Lucía, Z.B.; Myriam, B.D.; Adrian, O.L.; Martha Isabel, M. Profiling the immune response to Mycobacterium tuberculosis Beijing family infection: a perspective from the transcriptome. Virulence 2021, 12(1), 1689-1704. [CrossRef]
- Karmakar, M.; Trauer, J.M.; Ascher, D.B.; Denholm, J.T.; Hyper transmission of Beijing lineage Mycobacterium tuberculosis: systematic review and meta-analysis. J. Infect. 2019, 79(6), 572-581. [CrossRef]
- Maguga-Phasha, N.T.; Munyai, N.S.; Mashinya, F.; Makgatho, M.E.; Mbajiorgu, E.F. Genetic diversity and distribution of Mycobacterium tuberculosis genotypes in Limpopo, South Africa. BMC Infect. Dis. 2017, 17, 764. [CrossRef]
- Chihota, V.N.; Niehaus, A.; Streicher, E.M.; Wang, X.; Sampson, S.L.; Mason, P.; Källenius, G.; Mfinanga, S.G.; Pillay, M.; Klopper, M.; Kasongo, W. Geospatial distribution of Mycobacterium tuberculosis genotypes in Africa. PLoS ONE 2018, 13(8), e0200632. [CrossRef]
- van Soolingen, D.; Qian, L.; De Haas, P.E.; Douglas, J.T.; Traore, H.; Portaels, F.; Qing, H.Z.; Enkhsaikan, D.; Nymadawa, P.; van Embden, J.D. Predominance of a single genotype of Mycobacterium tuberculosis in countries of east Asia. J. Clin. Microbiol. 1995, 33(12), 3234-3238. [CrossRef]
- Pokam, B.D.; Yeboah-Manu, D.; Amiteye, D.; Asare, P.; Guemdjom, P.W.; Yhiler, N.Y.; Morton, S.N.; Ofori-Yirenkyi, S.; Laryea, R.; Tagoe, R.; Asuquo, A.E. Molecular epidemiology and multidrug resistance of Mycobacterium tuberculosis complex from pulmonary tuberculosis patients in the Eastern region of Ghana. Heliyon 2021, 7(10), e08152. [CrossRef]
- Bhembe, N.L.; Nwodo, U.U.; Okoh, A.I.; Obi, C.L.; Mabinya, L.V.; Green, E. Clonality and genetic profiles of drug-resistant Mycobacterium tuberculosis in the Eastern Cape Province, South Africa. Microbiology Open 2019, 8(3), e00449.
- Van der Spoel van Dijk, A.; Makhoahle, P.M.; Rigouts, L.; Baba, K. Diverse molecular genotypes of Mycobacterium tuberculosis complex isolates circulating in the Free State, South Africa. Int. J. Microbiol. 2016, 2016. [CrossRef]
- Solo, E.S.; Suzuki, Y.; Kaile, T.; Bwalya, P.; Lungu, P.; Chizimu, J.Y.; Shah, Y.; Nakajima, C. Characterization of Mycobacterium tuberculosis genotypes and their correlation to multidrug resistance in Lusaka, Zambia. Int. J. Infect. Dis. 2021, 102, 489-496. [CrossRef]
- Sagonda, T.; Mupfumi, L.; Manzou, R.; Makamure, B.; Tshabalala, M.; Gwanzura, L.; Mason, P.; Mutetwa, R. Prevalence of extensively drug resistant tuberculosis among archived multidrug resistant tuberculosis isolates in Zimbabwe. Tuberc. Res. Treat. 2014, 349141. [CrossRef]
Figure 1.
Distribution of mutations in rpoB gene.
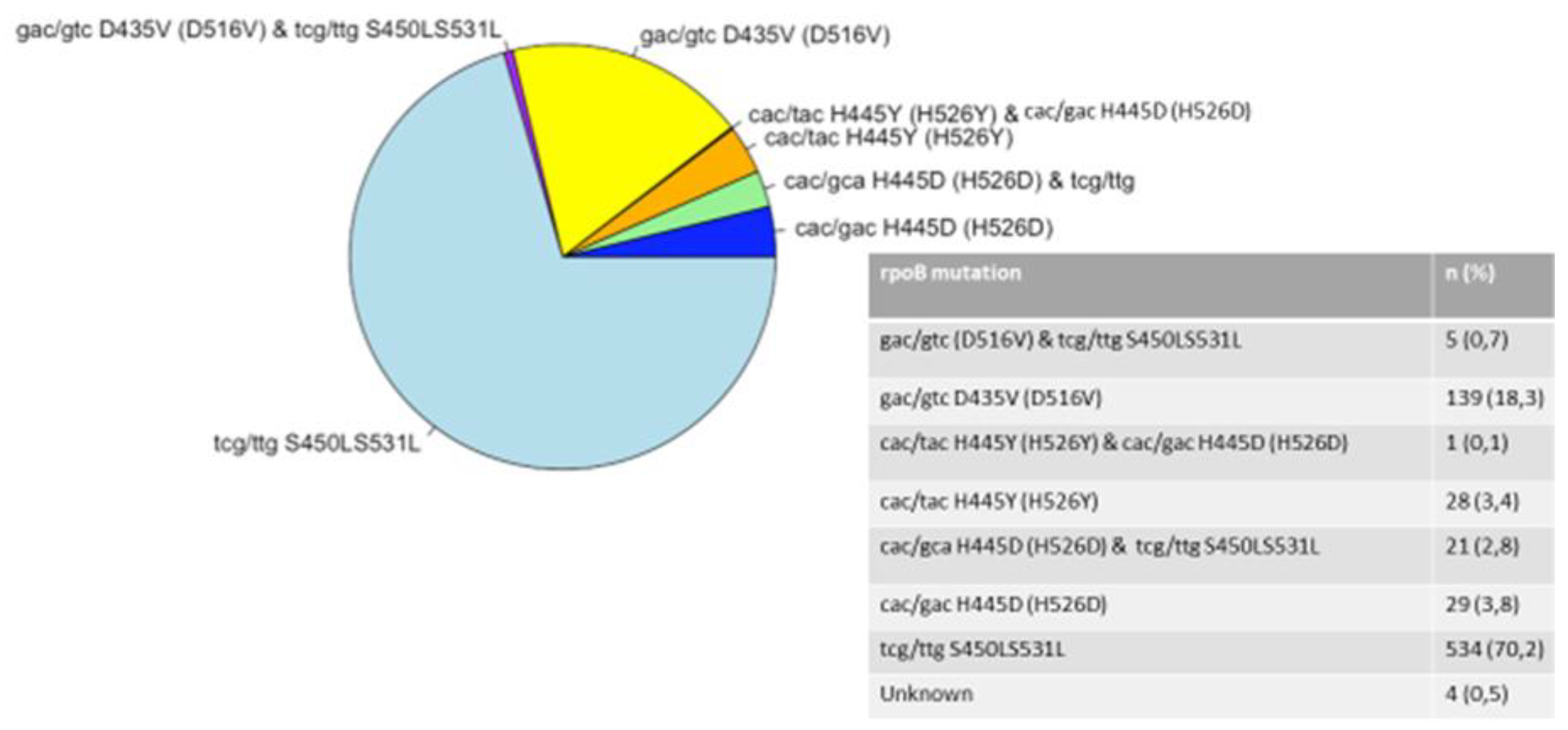
Figure 2.
Distribution of mutations in katG gene.
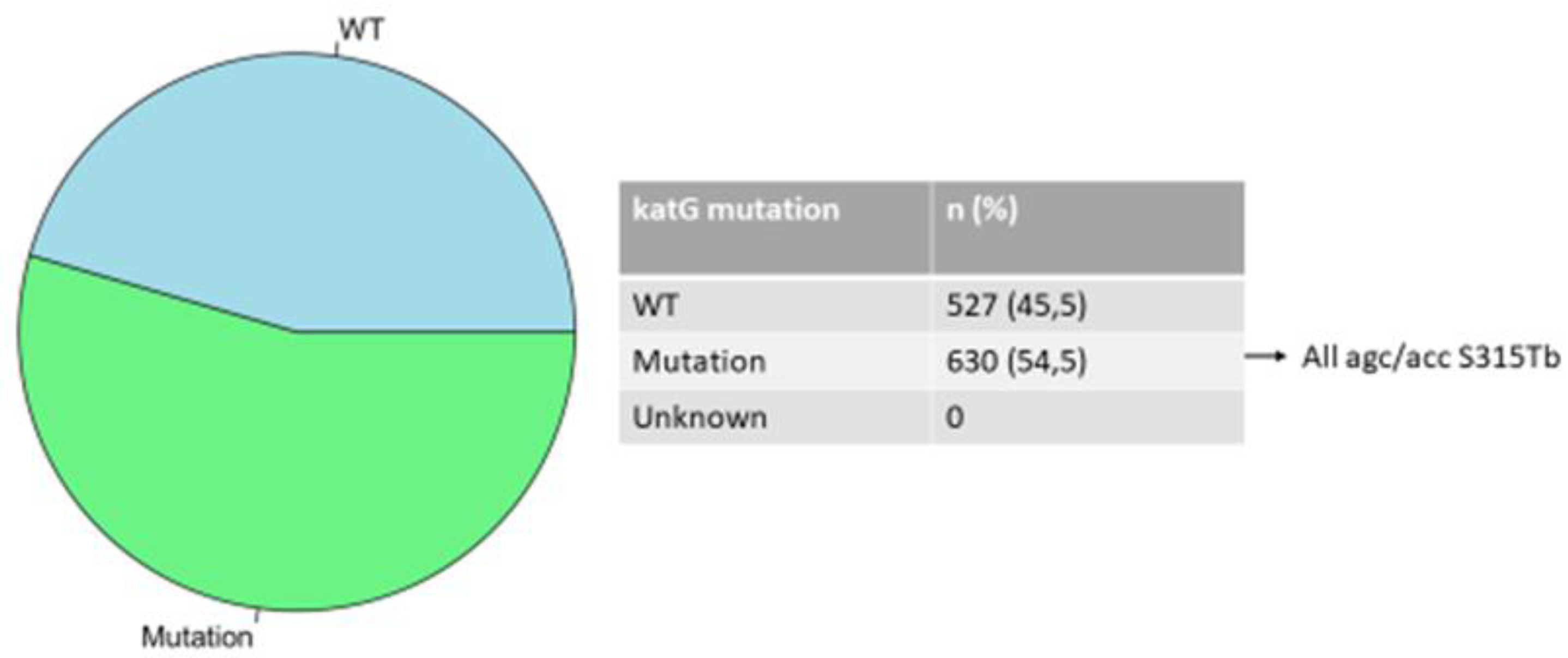
Figure 3.
Distribution of mutations in inhA gene.
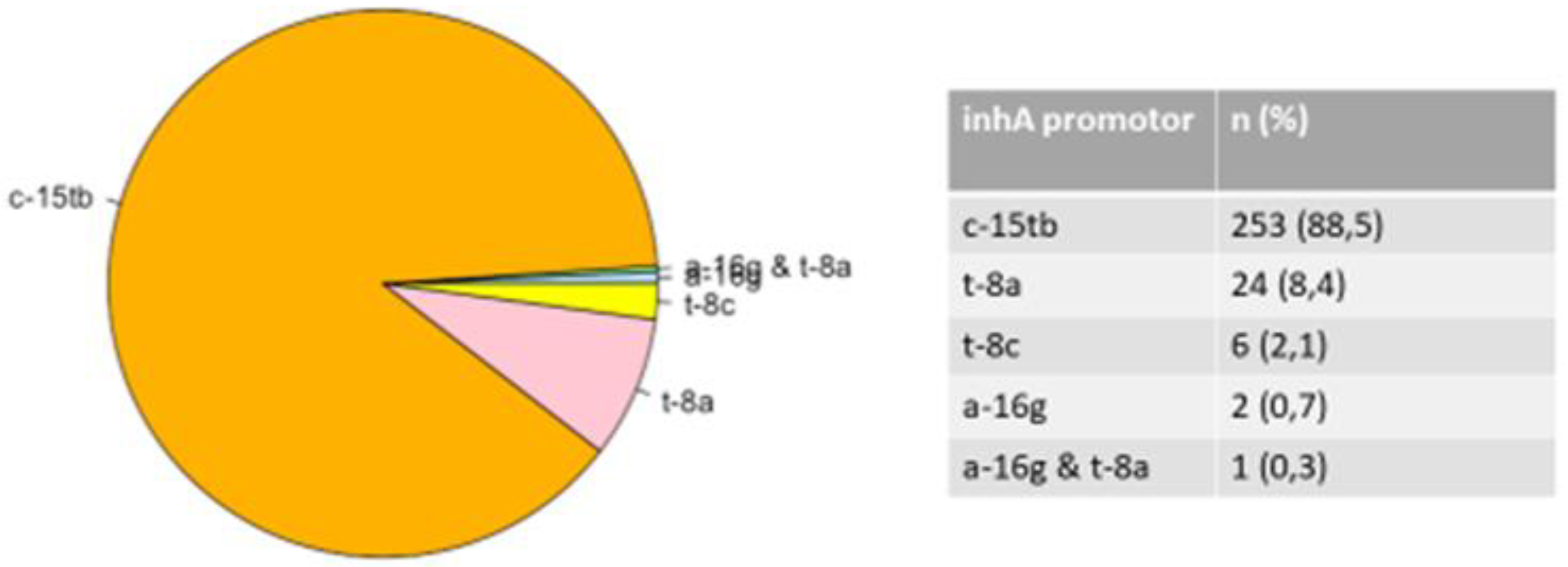

Table 1.
Number of DR-TB isolates over a 3-year period.
| Year of diagnosis | DR-TB case n (%) |
Annual heteroresistance rate n (%) |
|---|---|---|
| 2018 | 385 (33.3) | 37 (9.6) |
| 2019 | 376 (32.5) | 43 (11.4) |
| 2020 | 396 (34.2) | 127 (32.2) |
Table 2.
Isolates with predominant gene mutations and mutation regions.
| Combined mutation | Number of isolates (%) | Mutation regions | Number of isolates (%) |
|---|---|---|---|
| rpoB and katG | 532 (69.9) | rpoB S315L and katG 531ST | 366 (68,8%) |
| rpoB and inhA | 187 (24.6) | rpoB S315L and inhA c-15tb | 171 (91,4%) |
| rpoB S315L and inhA a-16g | 1 (0,5%) | ||
| rpoB S315L and inhA t-8c | 2 (1,1%) | ||
| rpoB S315L and inhA t-8a | 13 (6,9%) |
Table 3.
Lineages and sub-lineages from spoligotyped isolates.
| Lineage |
Number of Isolates |
Sub-lineage | Number of Isolates | SIT | Number of Isolates |
|---|---|---|---|---|---|
| Beijing | 185 | 1 | 185 | ||
| LAM | 83 | LAM3 | 62 | 33 | 36 |
| 719 | 17 | ||||
| ORPHAN | 2 | ||||
| 4 | 1 | ||||
| 130 | 1 | ||||
| 376 | 1 | ||||
| 2014 | 1 | ||||
| 2284 | 1 | ||||
| 2302 | 1 | ||||
| not in SITVIT | 1 | ||||
| LAM4 | 15 | 60 | 15 | ||
| LAM5 | 2 | 93 | 1 | ||
| 136 | 1 | ||||
| LAM9 | 2 | 42 | 2 | ||
| LAM11-ZWE | 2 | 811 | 1 | ||
| 1873 | 1 | ||||
| X | 48 | X1 | 18 | 2022 | 10 |
| 2226 | 4 | ||||
| 119 | 3 | ||||
| 336 | 1 | ||||
| X2 | 3 | 18 | 1 | ||
| 137 | 1 | ||||
| 2016 | 1 | ||||
| X3 | 27 | 92 | 21 | ||
| 2286 | 4 | ||||
| 2020 | 2 | ||||
| T | 34 | T1 | 29 | 53 | 13 |
| 926 | 4 | ||||
| 334 | 2 | ||||
| 501 | 2 | ||||
| 156 | 1 | ||||
| 245 | 1 | ||||
| 373 | 1 | ||||
| 519 | 1 | ||||
| 732 | 1 | ||||
| 1122 | 1 | ||||
| 1144 | 1 | ||||
| ORPHAN | 1 | ||||
| T2 | 1 | 52 | 1 | ||
| T2/3 | 2 | 73 | 2 | ||
| T3 | 1 | ORPHAN | 1 | ||
| T5-RUS1 | 1 | 254 | 1 | ||
| T- TUSCANY | 2 | 1737 | 2 | ||
| S | 31 | 34 | 18 | ||
| 789 | 4 | ||||
| 71 | 3 | ||||
| Not in SITVIT | 2 | ||||
| 790 | 1 | ||||
| 1211 | 1 | ||||
| Not in SITVIT | 1 | ||||
| Not in SITVIT | 1 | ||||
| EAI | 16 | EAI1-SOM | 10 | 806 | 6 |
| 48 | 2 | ||||
| 1649 | 2 | ||||
| EAI5 | 5 | 625 | 3 | ||
| ORPHAN | 2 | ||||
| EAI | 1 | Not in SITVIT | 1 | ||
| H | 6 | H1 | 5 | 62 | 2 |
| 2375 | 2 | ||||
| 47 | 1 | ||||
| H3 | 1 | 50 | 1 | ||
| CAS | 5 | CAS1-Kili | 2 | 21 | 2 |
| CAS1-Delhi | 1 | 1092 | 1 | ||
| CAS | 2 | Not in SITVIT | 2 | ||
| unknown | 4 | Unknown | 1 | 2018 | 1 |
| Not in SITVIT | 3 | Not in SITVIT | 1 | ||
| Not in SITVIT | 1 | ||||
| Not in SITVIT | 1 | ||||
| no result | 27 | ||||
| TOTAL | 441 |
SIT= Shared International Type; SITVIT = international spoligotyping database; SITIV 2 = is a genotyping molecular markers database focusing on Mycobacterium tuberculosis complex; CAS = Central Asian; Orphan = isolates showed unknown patterns that were not as-signed to any known major lineages in the SITVIT2 database; LAM: Latin American; EAI: East-African Indian; Delhi/CAS: Delhi/Central Asian, H: Haarlem); no = number.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



