Submitted:
06 April 2023
Posted:
07 April 2023
Read the latest preprint version here
Abstract
Recently, two pig-to-human kidney transplants and a pig-to-human heart transplant were completed. The kidney trials involved a patient who was deceased and a patient who was brain dead. They seemed to indicate that pig kidneys can be at least somewhat functional in humans. However, patients still have to be under severe immunosuppression - and the first patient to receive a porcine heart passed away after two months. It is difficult to know exactly which proteins we need to overexpress or underexpress/knockout in a porcine organ to negate the human recipient’s immunological response to it. And testing different porcine organ genetic modifications in baboons can cost around $500,000 per transplant. But there might be a way to decrease immunogenicity where we don’t have to worry so much about modifying the animal’s organs genetically. First, however, we would have to prevent complement factor-mediated lysis of the porcine vascular endothelial cells, which we have made much progress on with triple knockout animals. Then, we could modify the porcine organ so that the cells of said organ secrete a small molecule or peptide that acts as a chemorepellent for the host immune cells. The host immune cells can be modified via bone marrow transplant or vector delivery to express the chemorepulsion receptor.
Keywords:
Xenotransplantation
; chemorepulsion
; adaptive immunity
; innate immunity
; dysregulated coagulation
; complement cascade
Introduction
8,000 people die each year in the U.S. alone from organ (especially kidney) shortages [1]. Currently, porcine or baboon organ xenotransplantation into human hosts is limited by issues of immunogenicity [2,3]. Patients who receive transplants from porcine organs or non-donor matches have to be on immunosuppression indefinitely and as a result suffer from many side effects such as much more frequent infections.
While hyper-acute rejection has been successfully reduced via a triple-knockout strategy in porcine organs, vascular issues and long-term rejection are still problematic [4]. This relates to the interaction between the host immune system and the vascular endothelial cells (VECs) in porcine organs, as well as infiltration by adaptive and innate immune cells into the organ over time [5].
The Hypothesis
I hypothesize that a large portion of the host immune inflammatory response to a xenograft could be curtailed by keeping the host immune cells out of the transplanted organ. You obviously cannot keep the host immune cells out of circulation, and so they are bound to come in contact with VECs of the transplanted organ, but you could express proteins like human endomucin [6] on the surface of these VECs to prevent the attachment of host immune cells and therefore ideally prevent much inflammation. Perhaps if such proteins are employed, synthetic versions should be designed that only relate to their attachment properties if possible - i.e., with no other activities that could be detrimental [7].
To keep the host immune cells out of the transplanted organ parenchyma, one could reprogram a patient’s hematopoietic stem cells (HSCs), either with the delivery system I mentioned here [8] targeted to VECs in the “zip codes” of HSC niches or autologous bone marrow transplant [9] with engineered HSCs (perhaps with longer telomeres to allow for repopulation of leukocytes). One would install a chemorepulsion module in the patient’s immune cells that is sensitive to an orthogonal small molecule or peptide produced by the porcine organ. The chemorepulsion effect would be akin to a reversal of the type of system cited here [10]. Of course, you could potentially simply program the porcine tissue to secrete natural chemorepellants, but the immune system is very complex, and some immune cell types may actually be attracted by endogenous chemorepellants of other immune cell types.
Evaluation of the Hypothesis
Binding of human antibodies to porcine cells has been markedly reduced through the knockout of three antigens: αGal, Neu5GC, and SDa. Further improvements in avoiding immunogenicity can be achieved by expressing certain human proteins in the porcine cells, like CD47 [11]. However, antibodies to VEC macromolecules in the lumen of the porcine blood vessels could be a problem over time. Perhaps there is a way to develop B-cell tolerance to all of the porcine VEC markers. One possible approach to negate complement cascade and antibody binding issues is to replace the porcine endothelial cells with human endothelial cells - either embryonically [12] or post-removal of the organ [13]. Moreover, the porcine VECs could be bioengineered in such a way that endocytosis is upregulated - and that the vascular endothelial glycocalyx is turned over more frequently. One, more difficult intervention might be to bioengineer autologous leukocytes to bind and roll along the porcine VECs, internalizing but not reacting to antibodies on the surface. Another option is to engineer tolerance against at least the major porcine antigens using BAR Tregs [14]. Circulating antibodies may negatively affect this approach [13]. If so, the engineered TCRs could be overexpressed and contain degrons in one or both of the ζ-chains. Co-culturing recipient Tregs with porcine-antigen-loaded tolerogenic dendritic cells could also help [15].
Mixed hematopoietic chimerism [16] and thymic grafting [17] are two ways of instilling tolerance. However, xenogeneic HSCs are eliminated rather quickly by the new host organism, even when intra-bone injections are administered and the donor HSCs express the host CD47 protein [18] - although perhaps an initial injection of as well as periodic injections of autologous, porcine-specific recipient Tregs would help protect them, at least until central tolerance is established [15,19].
For both mixed hematopoietic chimerism and thymic grafting, a patient’s T and B cells should be ablated. Donor HSCs could repel all immune cells except immature T cells, immature B cells, and perhaps naïve B cells in general - or at least those that matured via interaction with the xenogeneic HSCs (perhaps marked via SynNotch [20] circuitry). The thymic graft that would then be transplanted could repel all immune cells except immature T cells, thymocytes, and subsequent naïve T cells. These tactics could allow the HSCs or thymic graft to be safe and effectively instill tolerance.
For the thymic grafting technique, at least for younger patients who still have a very functional thymus, thymectomy seems to be required [21] - although perhaps not with the chemorepulsion tactic detailed here. However, repelling naïve T cells may interfere with maturation inside the graft. So, perhaps the thymic graft TEC cells could have a ligand on their surface that activates a SynNotch pathway in immature T cells and thymocytes - making them immune to subsequent chemorepulsion throughout and after the maturation process. Or, perhaps a peptide could be secreted by the thymic graft that in high concentrations starts a clock in immature T cells or thymocytes that prevents chemorepulsion until the clock runs down, even when they subsequently become naïve T cells - allowing them the time to mature inside the graft before being repelled.
If thymectomy is required despite these and perhaps other interventions, mixed hematopoietic chimerism would certainly be better - at the very least for younger patients. At the very least, the thymectomy approach, if necessary, could give younger patients more time to receive a human organ - and their thymi could potentially be regrown by a minimal scaffold seeded with autologous fibroblasts - along with FOXN1 overexpression [22].
Notably, if HSC-based chimerism is employed, those cells could also potentially reseed the vascular endothelium of the transplanted organ when cells are naturally lost or if some form of damage occurs. (I believe it is still unclear to what extent the vascular endothelium replenishes itself vs. being seeded by HSCs.)
Finally, an even more complicated strategy to avoid excessive antibody production against the xenotransplanted VECs or VEC glyocalyx would be to magnetically draw carrier macrophages [23] into the thymus - and deliver miniaturized porcine VECs to the thymic cells that replicate up until a certain copy number, at which point they stop via quorum sensing [24]. Every so often, through competence circuitry [25], a small number would lyse, releasing antigens to be sampled by the cells of thymus that instill T-cell tolerance [26]. The miniature porcine VECs may need to replicate in a vacuole for this strategy to work. It is well known that activated CD4+ helper T cells are generally required for B-cell activation [27]. This could work for hematopoietic stem cells as well - either through magnetism, my proposed delivery system, or during a bone marrow transplant - in a sort of take on mixed hematopoietic chimerism.
Figure 1.
Immune cells are chemorepelled from a transplanted porcine heart, for example. Neutrophils that would otherwise infiltrate the foreign organ and release neutrophil extracellular traps (NETs) and damage the tissue would be unable to do so [28]. Along these lines, when macrophages encounter NETs, they release cytokines and inflammatory markers; NETs appear to them to be damage-associated molecular patterns. This process would also not occur. Moreover, T-cells would not get close enough to bind SLA1 or SLA2 on the porcine cells - which would otherwise trigger cytokine release and target cell death.
Figure 1.
Immune cells are chemorepelled from a transplanted porcine heart, for example. Neutrophils that would otherwise infiltrate the foreign organ and release neutrophil extracellular traps (NETs) and damage the tissue would be unable to do so [28]. Along these lines, when macrophages encounter NETs, they release cytokines and inflammatory markers; NETs appear to them to be damage-associated molecular patterns. This process would also not occur. Moreover, T-cells would not get close enough to bind SLA1 or SLA2 on the porcine cells - which would otherwise trigger cytokine release and target cell death.
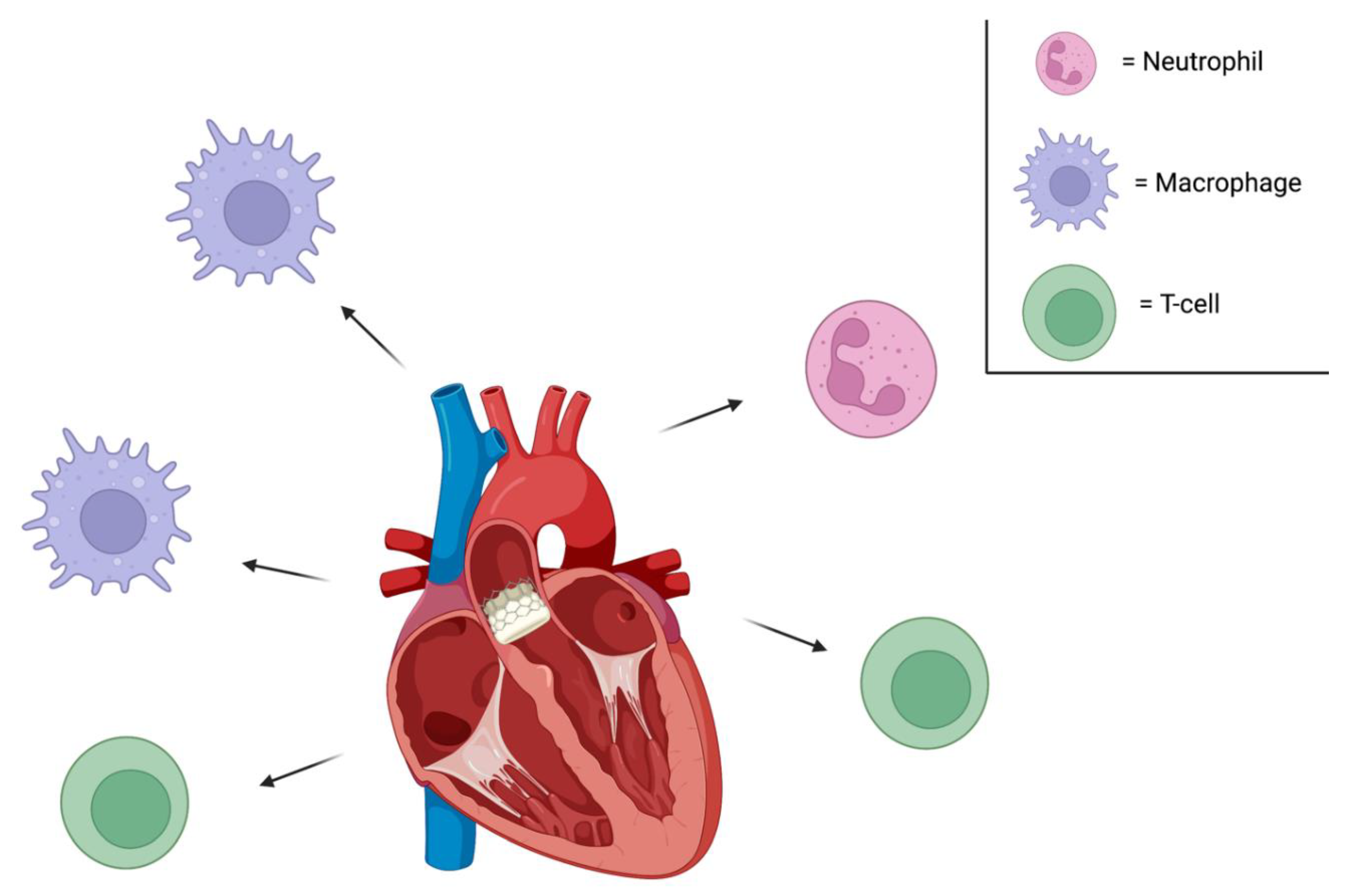
If all of a patient’s immune cells are chemorepelled from the porcine organ, CD8+ T-cells will be unable to bind foreign cells and lyse them. Macrophages and neutrophils will be unable to phagocytose them and secrete inflammatory cytokines. I believe the benefits of such an approach are fairly clear.
In order to avoid pathogen infections wherein they have evolved to secrete the small molecule in question, it would almost certainly be wise to utilize multiple small molecules that only achieve chemorepulsion when all of them are detected.
Also, while they are using CRISPR to remove porcine retroviruses, they could also potentially just genetically install a porcine version of DRACO [29] in the porcine organs that are to be transplanted.
Conclusion
This approach with xenotransplantation also applies to non-donor matched allogeneic transplantation if my proposed delivery system can be developed [8]. However, lentivirus-mediated suppression of HLA antigens in donor organs [30,31] achieved during normothermic ex vivo perfusion - could work as well. In fact, perhaps only the donor VECs would need to be affected [32]. Moreover, the donor VECs could perhaps be stripped, and recipient VECs cultured ex vivo could be used to reseed them. DRACO could also be installed in the donor cells simultaneously to help manage infections given HLA antigen knockout or knockdown.
Funding
Funding not received for the study.
Acknowledgements
Thank you to my family and friends for very helpful conversations. The figure in this piece was created with BioRender.com.
Competing Interests
The author declares no competing interests.
References
- Anonymous. Facts and Benefits of Organ Donation | UF Health, University of Florida Health. n.d. Available from: https://ufhealth.org/blog/facts-and-benefits-organ-donation [Last accessed: 11/11/2022].
- Montgomery RA, Stern JM, Lonze BE, et al. Results of Two Cases of Pig-to-Human Kidney Xenotransplantation. N Engl J Med 2022;386(20):1889–1898. [CrossRef]
- Wang W, He W, Ruan Y, et al. First pig-to-human heart transplantation. Innovation (Camb) 2022;3(2):100223. [CrossRef]
- Fischer K, Kraner-Scheiber S, Petersen B, et al. Efficient production of multi-modified pigs for xenotransplantation by ‘combineering’, gene stacking and gene editing. Scientific Reports 2016;6:29081. [CrossRef]
- Fox A, Mountford J, Braakhuis A, et al. Innate and Adaptive Immune Responses to Nonvascular Xenografts: Evidence That Macrophages Are Direct Effectors of Xenograft Rejection. The Journal of Immunology 2001;166(3):2133–2140. [CrossRef]
- Zahr A, Alcaide P, Yang J, et al. Endomucin Prevents Leukocyte–Endothelial Cell Adhesion and Has a Critical Role under Resting and Inflammatory Conditions. Nat Commun 2016;7(1):10363. [CrossRef]
- Park-Windhol C, Ng YS, Yang J, et al. Endomucin Inhibits VEGF-Induced Endothelial Cell Migration, Growth, and Morphogenesis by Modulating VEGFR2 Signaling. Sci Rep 2017;7(1):17138. [CrossRef]
- Renteln M. Lipofuscin as the main driving force of current age-related disease: justification and strategies for removal. Current Aging Science (Submitted: February, 2022).
- Persaud SP, Ritchey JK, Kim S, et al. Antibody-drug conjugates plus Janus kinase inhibitors enable MHC-mismatched allogeneic hematopoietic stem cell transplantation. J Clin Invest n.d.;131(24):e145501. [CrossRef]
- Park JS, Rhau B, Hermann A, et al. Synthetic Control of Mammalian-Cell Motility by Engineering Chemotaxis to an Orthogonal Bioinert Chemical Signal. Proc Natl Acad Sci USA 2014;111(16):5896–5901. [CrossRef]
- Ma D, Hirose T, Lassiter G, et al. Kidney transplantation from triple-knockout pigs expressing multiple human proteins in cynomolgus macaques. Am J Transplant 2022;22(1):46–57. [CrossRef]
- Das S, Koyano-Nakagawa N, Gafni O, et al. Generation of human endothelium in pig embryos deficient in ETV2. Nat Biotechnol 2020;38(3):297–302. [CrossRef]
- Cohen S, Partouche S, Gurevich M, et al. Generation of vascular chimerism within donor organs. Sci Rep 2021;11(1):13437. [CrossRef]
- Scott DW. Genetic Engineering of T Cells for Immune Tolerance. Molecular Therapy - Methods & Clinical Development 2020;16:103–107. [CrossRef]
- Li M, Eckl J, Geiger C, et al. A novel and effective method to generate human porcine-specific regulatory T cells with high expression of IL-10, TGF-β1 and IL-35. Sci Rep 2017;7(1):3974. [CrossRef]
- Sachs DH. Transplantation Tolerance Through Mixed Chimerism: From Allo to Xeno. Xenotransplantation 2018;25(3):e12420. [CrossRef]
- Yamada K, Ariyoshi Y, Pomposelli T, et al. Co-transplantation of Vascularized Thymic Graft with Kidney in Pig-to-Nonhuman Primates for the Induction of Tolerance Across Xenogeneic Barriers. Methods Mol Biol 2020;2110:151–171. [CrossRef]
- Watanabe H, Ariyoshi Y, Pomposelli T, et al. Intra-bone bone marrow transplantation from hCD47 transgenic pigs to baboons prolongs chimerism to >60 days and promotes increased porcine lung transplant survival. Xenotransplantation 2020;27(1):e12552. [CrossRef]
- Pilat N, Granofszky N, Wekerle T. Combining Adoptive Treg Transfer with Bone Marrow Transplantation for Transplantation Tolerance. Curr Transplant Rep 2017;4(4):253–261. [CrossRef]
- Morsut L, Roybal KT, Xiong X, et al. Engineering Customized Cell Sensing and Response Behaviors Using Synthetic Notch Receptors. Cell 2016;164(4):780–791. [CrossRef]
- Kamano C, Vagefi PA, Kumagai N, et al. Vascularized thymic lobe transplantation in miniature swine: Thymopoiesis and tolerance induction across fully MHC-mismatched barriers. Proc Natl Acad Sci U S A 2004;101(11):3827–3832. [CrossRef]
- Bredenkamp N, Ulyanchenko S, O’Neill KE, et al. An organized and functional thymus generated from FOXN1-reprogrammed fibroblasts. Nat Cell Biol 2014;16(9):902–908. [CrossRef]
- Muthana M, Kennerley AJ, Hughes R, et al. Directing cell therapy to anatomic target sites in vivo with magnetic resonance targeting. Nature Communications 2015;6(1):8009. [CrossRef]
- Miller ST, Xavier KB, Campagna SR, et al. Salmonella typhimurium Recognizes a Chemically Distinct Form of the Bacterial Quorum-Sensing Signal AI-2. Molecular Cell 2004;15(5):677–687. [CrossRef]
- Mirouze N, Dubnau D. Chance and Necessity in Bacillus subtilis Development. Microbiology Spectrum 2013;1(1):1.1.02. [CrossRef]
- Nunes-Alves C, Nobrega C, Behar SM, et al. Tolerance has its limits: how the thymus copes with infection. Trends Immunol 2013;34(10). [CrossRef]
- Hu M, Hawthorne WJ, Yi S, et al. Cellular Immune Responses in Islet Xenograft Rejection. Frontiers in Immunology 2022;13.
- Arabi TZ, Sabbah BN, Lerman A, et al. Xenotransplantation: Current Challenges and Emerging Solutions. Cell Transplant 2023;32:09636897221148771. [CrossRef]
- Rider TH, Zook CE, Boettcher TL, et al. Broad-Spectrum Antiviral Therapeutics. PLOS ONE 2011;6(7):e22572. [CrossRef]
- Yuzefovych Y, Valdivia E, Rong S, et al. Genetic Engineering of the Kidney to Permanently Silence MHC Transcripts During ex vivo Organ Perfusion. Frontiers in Immunology 2020;11.
- Berkhout B. A Fourth Generation Lentiviral Vector: Simplifying Genomic Gymnastics. Molecular Therapy 2017;25(8):1741–1743. [CrossRef]
- Figueiredo C, Carvalho Oliveira M, Chen-Wacker C, et al. Immunoengineering of the Vascular Endothelium to Silence MHC Expression During Normothermic Ex Vivo Lung Perfusion. Hum Gene Ther 2019;30(4):485–496. [CrossRef]
- Su LT, Gopal K, Wang Z, et al. Uniform Scale-Independent Gene Transfer to Striated Muscle After Transvenular Extravasation of Vector. Circulation 2005;112(12):1780–1788. [CrossRef]
- Greelish JP, Su LT, Lankford EB, et al. Stable restoration of the sarcoglycan complex in dystrophic muscle perfused with histamine and a recombinant adeno-associated viral vector. Nat Med 1999;5(4):439–443. [CrossRef]
- Gregorevic P, Blankinship MJ, Allen JM, et al. Systemic delivery of genes to striated muscles using adeno-associated viral vectors. Nat Med 2004;10(8):828–834. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



