
Submitted:
06 April 2023
Posted:
07 April 2023
You are already at the latest version
Abstract
Atherogenesis and dyslipidemia increase the risk of cardiovascular disease, which is the leading cause of death in developed countries. While blood lipid levels have been studied as disease predictors, their accuracy in predicting cardiovascular risk is limited due to its high interindividual and interpopulation variability. The lipid ratios: atherogenic index of plasma (AIP=log TG/HDL-C) and the Castelli risk index 2 (CI2=LDL-C/HDL-C) have been proposed as better predictors of cardiovascular risk, but the genetic variability associated to these ratios has not been investigated. This study aimed to identify genetic associations with these indexes. The study population (n=426) included males (40%) and females (60%) aged 18-52 years (mean 39 years), the Infinium GSA array was used for genotyping. Regression models were developed using R and PLINK. AIP was associated with variation on APOC3, KCND3, CYBA, CCDC141/TTN, and ARRB1 (p-value < 2.1E-6) the three former previously associated to blood lipids, while CI2 was associated with variants on DIPK2B, LIPC, and 10q21.3 rs11251177 (p-value 1.1E-7) the latter previously linked to coronary atherosclerosis and hypertension. KCND3 rs6703437 was associated with both indexes. This study is the first to characterize the potential link between genetic variation and atherogenic indexes, AIP and CI2, highlighting the relation between genetic variation and dyslipidemia predictors. These results also contribute to consolidating the genetics of blood lipid and lipid indexes.
Keywords:
Dyslipidemia
; atherogenic indexes
; genetic associations
1. Introduction
Atherogenesis and dyslipidemia are key risk factors for coronary artery disease (CAD), the leading cause of mortality in the world [1]. Atherosclerosis is an inflammatory process that encompasses the formation of plaque in the artery walls contributing to cardiovascular disease (CVD), hypertension, stroke, and coronary artery disease [2]. Atherosclerosis is complex and multifactorial, involving genetics, the environment, lipid levels such as low-density lipoprotein cholesterol (LDL-C), total triglycerides (TG), and high-density lipoprotein cholesterol (HDL-C), the comorbidities, diabetes, dyslipidemia, hypercholesterolemia, and the accumulation of oxidized LDL [3]. Increasing age is highly correlated to atherosclerosis and current epidemiological studies suggest that early detection would lead to prevention and the deployment of interventions for treatment and control prior to cardiovascular disease onset. However, the characterization of early molecular phenotypes atherosclerotic key events remain to be fully elucidated [4]. Dyslipidemia, one of the main underlying factors in atherogenesis, is highly prevalent worldwide; for instance, it reaches 53% in the United States, 49% in China [5], and 80% in Turkey [6]. In Mexico City, the CARMELA study determined a prevalence of 50.5% for hypercholesterolemia and 32.5% for hypertriglyceridemia, and hypotheses are that nearly every two out of three city dwellers might have some type of dyslipidemia [7,8].
The early identification of atherosclerosis and dyslipidemia is urgently needed to pinpoint targeting therapies and preventive interventions. Lipids levels and lipid indexes have been of some help because of their association to cardiovascular risk. For example, LDL-C, TG, and HDL-C are strongly correlated to coronary heart disease (CHD) although their underlying genetics has not been fully characterized [9]. A genetic profile that may aid in the identification of dyslipidemia, and CVD or its predisposition could propel earlier diagnosis and interventions.
Up-today, the association between genetics and blood lipids has aided in the search for reliable markers of CVD, atherosclerosis, and dyslipidemia since in part, these have a heritable basis and the literature evidences their association with an extensive collection of genetic loci [10,11]. For example, single nucleotide variants (SNVs) on APOE, CETP, LPL, PCSK9, and GCKR have been significantly associated with lipid levels and dyslipidemia in different populations [10,11]. Despite the high prevalence of CVD and dyslipidemia in Mexico, only a few studies have investigated its relationship with genetic variation. One report found SNVs on APOA5, GCKR, LPL, and NPC1 associated with hypertriglyceridemia [12], while polymorphisms on ABCA1, CETP, LIPC, and LOC55908 have been associated with hypoalphalipoproteinemia, many of these variants are shared by different populations but some seem to be unique to certain geographical ancestries [10,13].
Recent studies have highlighted the accuracy and relevance of lipid ratios/indexes to better assess dyslipidemia and cardiovascular risk. The atherogenic index of plasma (AIP=log TG/HDL-C) can accurately predict hypertension, metabolic syndrome, and ischemic stroke even when HDL-C and TG levels seem normal or when isolated values of TG or HDLC-C cannot assess this risk [14,15]. The direct measurement of HDL-C and LDL-C has shown bias in assessing cardiovascular health (16) but their ratio i.e., the CI2 (LDL-C/HDL-C, CI2), although less cited, has been confirmed as a better predictor of cardiovascular risk [9,16,17]. Hence, there is an apparent, but not as frequently acknowledged, value of AIP and CI2 indexes to identify cardiovascular risk. It is possible that AIP and C2 ratios together with genetics could improve the clinical assessment of CVD risk and dyslipidemia. Nevertheless, little is known about the direct relationship between these indexes and genetic variation. Therefore, here we investigated the potential association between lipid indexes, AIP and CI2, and genetic variants in Mexican adults free of cardiovascular disease.
2. Materials and Methods
2.1. Population
Study participants were volunteers recruited between 2014 and 2016 for the longitudinal study Tlalpan 2020 (n=426) [18], all normotensive and with no previous diagnosis of cardiovascular disease. The study protocol followed the principles of the Declaration of Helsinki and was approved by the Institutional Bioethics Committee of the Instituto Nacional de Cardiologia numbers 13-802 and 16-983 and at INMEGEN, CEI2017/20. A blood sample was withdrawn after an overnight 12h fast in EDTA-Vacutainers. DNA was extracted using the PureBlood kit (Qiagen, Valencia CA, USA), nucleic acid quality control and concentration were assessed in a Nanodrop (ThermoFisher, Waltham, MA, USA) and aliquoted to 40 ng/μL, DNA samples were stored at -70o C until analysis. Anthropometric measurements included, height, weight, waist and hip circumference. Clinical determinations included, triglycerides (TG), HDL-C, LDL-C, uric acid, creatinine, and glucose. Blood pressure was reported as the average of three measurements with a calibrated sphygmomanometer after a resting period of 10 min. Circulating lipids were compared to reported reference values [19,20]. Total triglycerides, HDL-C, and LDL-C levels were used to calculate the Castelli risk index 2, CI2= LDL- C/HDL-C [18] and the atherogenic index of plasma, AIP = log (TG/HDL-C) [14,21,22].
2.2. Lipid indexes and genetic analyses
Lipid values and indexes were assessed according to the above-mentioned equations, AIP and CI2 indexes showed a wide normal distribution, AIP: mean 0.419±0.281 and range (-0.40 – 1.610) and CI2: mean 2.66 ± 0.841 and range (0.430 – 5.36). DNA samples were genotyped using the GSA-Infinium array 24 v.10 (Illumina) for 670K variants. After bioinformatic quality control, we excluded redundant SNVs, variants with less than 95% genotype call rate, missing data per variant > 5%, missing data per individual > 2%, and minor allele frequency MAF < 1%, for statistical analyses we considered 330K variants
Independent linear regression models were developed with AIP and CI2 as dependent variables assuming additive effects on the allele dosage and selected covariates [23]. Descriptive genomic analyses included, Hardy-Weinberg p -value < 1.0 E-5, call rate > 0.95, and sex-check by heterozygosity [25]. Model covariates were selected according to their impact on the total variance for AIP or CI2 based on the value of the first component of PCA and included, uric acid, weight, waist circumference, and sex. Statistical significance was considered starting at p-value=1.0 E-5 given that our population was tested for 1.0 E5 SNVs (330K variants). Nevertheless, we identified genetic associations for AIP at p-value=1.0 E-6 and for CI2 between p-value=1.0 E-7 and 1.0 E-5, without correction for multiple testing since we defined this study as exploratory i.e., our observations would require future validation
To account for population stratification, we assessed genetic admixture using the Software Admixture 1.3 [26], and the 1000G project reference populations, Northern Europeans from Utah (CEU, Caucasians), Mexicans from Los Angeles (MXL), Yoruba in Ibadan from Nigeria (YRI), Chinese Han from Beijing (CHB), and Natives from Mexico. Genetic data from the GSA microarray were used to define 56,000 Ancestry Informative Markers (AIMs), setting an identity by descent value, IBD pi-hat Z0.5, and excluding markers in linkage disequilibrium or a physical distance < 500kb, ensuring that ancestry informative markers (AIMs) were uniformly distributed throughout the genome.
3. Results
3.1. Population and lipid characteristics
The study group consisted of 40% males and 60% females (n=170, 256), with a mean age of 39 years (17 – 53). Mean and median values were within reference laboratory ranges for uric acid, creatinine, glucose, HDL-C, LDL-C, and TG (Table 1). Males showed higher total TG (49%), LDL-C 32%, total cholesterol (50%), and HDL-C (45%) levels compared to females (p-value 1.9E-2 – 9.7E-4, Table 2). Dyslipidemia was identified in 70% of males and 72% of females, deviation from reference levels is depicted in Table 2. Also, females showed lower creatinine and uric acid levels compared to males and overall displayed a healthier lipid profile, in agreement with previous reports of lipid sex differences (16,27,28). Calculations for AIP and CI2 indexes showed that men presented 26% and 18% higher AIP and CI2 values compared to women (p-value ≤ 4E-6) reflecting an expected higher cardiovascular risk (Table 1 and Table 2).
3.2. Genetic analyses
The GSA array probes for 669,672 genetic variants, after quality control we considered a total of 309,635 SNVs for the development of the association models. Population admixture was assessed using publicly available genetic variation for European, Native Mexicans, Asian, and Sub-Saharan African ancestries. Our sample showed expected ancestral contributions of European 30% (0.1 – 0.80), Native Mexican 65% (0.16 – 0.99), CHB 1% (0 – 0.25), and YRI 4% (0 – 0.28) similar to previous results [11]. The admixture proportion was considered as a covariant for the association models and was graphically represented in Supplemental Figure S1.
3.3. Genotype-phenotype associations
3.3.1. Genotype-phenotype association for AIP
The AIP was associated with seven variants on, APOA1/APOC3, CYBA, ARRIB1, CCDC141, KCND3, and VLDR (p-value=1E-6, Table 3) and to other 26 variants with a p-value ~1E-5 (Supplemental Table S1 and Supplemental Figure S2). Top associated variants were, APOC3 rs5128, ARRB1 rs11236389, LIPC/ALDH1A2 rs261342, DIPK2B rs4294309, and KCND3 rs6703437, the latter was also associated with CI2 although with a lower significance (p-value=1.1E6 – 1.8E-5, Table 3). The visual impact of the most significant genotypes on AIP values were represented in Figure 1A. The variant allele of APOC3 rs5128 seems to have a detrimental effect on AIP values while the variant allele of ARRB1 rs11236389 is suggestive of a protective atherogenic effect showing an association with lower AIP values.
3.3.2. Genotype-phenotype association for CI2
We found a statistically significant association between CI2 and nine variants mostly in intergenic regions, four located on chromosome 12 in partial linkage disequilibrium (p-value <1E-7 – 1E-5, Table 3, and Supplemental Figure 2). Figure 1B depicts the impact of the most significant variants on CI2, where the minor allele of 10q21.3 rs11251177, and LINC0245 rs6582413 showed an association with a 25% increment in CI2 levels (p-value 5.5E-5). The current literature shows only a few reports on the clinical impact of the loci here identified (Table 3) or its relation to lipids and lipid indexes, hence we sought for their in-silico impact in The Regulome database (visited on December 2022), to infer its functional impact. For 10q21.3 rs11251177 and rs6582413 we found a neutral impact of these variants to clinical phenotypes, both in LD with low-ranking variants and within 500kb (28) (Table 3) leaving little room for interpretation and a void in information that ought to be addressed in future research.
4. Discussion
The identification of a quantitative relationship between genetics and CVD surrogates such as AIP and CI2 is of health transcendence due to the high mortality associated with cardiovascular disease underlied by atherogenesis. Several lipid levels and their indexes have attempted to predict cardiovascular risk and support prevention strategies. There are a couple of studies that associate lipids and lipoproteins measurements with genetic loci in different populations, but high interindividual or population variability has clouded their interpretation and potential application [30,31]. The AIP and CI2 indexes have emerged as surrogate markers of cardiovascular health as they have been reliably correlated with cardiovascular risk, lipoprotein size [17,29], or plasma atherogenicity [30]. They have demonstrated to be better CVD predictors compared to TG/HDL-C alone [31,32]. M. Dobiasova and J. Frohlich showed that the AIP index closely correlates to lipoprotein particle size and fractional esterification rate of HDL-C which in turn is a predictor of coronary artery disease risk [33,34], cerebrovascular accident [35], the thickness of the carotid intima-media, statin response, and ischemic stroke [36]. Current reports have provided valuable molecular insights into lipid metabolic pathways and dyslipidemia, but no study has identified the relationship between the ratios, AIP and CI2 with genetics [10].
Here, we report statistically significant associations between gene variants and lipid indexes AIP and CI2 previously reported as relevant for lipid levels and CVD including, APOC3/APOA1, 10q.21.3 rs1125117, KCND3, and VLDR. Evidence from other fields suggests that adding genetic information to clinical CVD prevention may fine-tune the utility of lipid indexes for disease prediction [20], likely facilitating the development of specific laboratory tests and algorithms. Below, we discuss the relevance of our findings in the scope of novel and previous genetic associations.
The identification of variants on APOC3 and VLDR associated to atherogenic indexes confirmed previous inferences, since these genes are well known to impact lipid levels. APOC3 has been repeatedly associated with dyslipidemia [37] and blood lipids. Several studies confirm a variety of loci, not always in LD, mapping on the APOC1, APOC3, and APOA5 clusters and its relation to blood lipids [38,39,40]. Variant APOC3 rs147210663 has been reported over 40 times associated to dyslipidemia, cholesterol, and BMI, it is in LD with APOC3 rs5128 here identified, and whose association with triglyceride levels in Pima Amerindians has been reported as a founder mutation [41]. Also, a recent multi-ancestry analysis on 170,000 exomes including 16,440 individuals of “Hispanic” origin reported that APOC3 is a relevant gene for HDL-C and the TG/HDL-C ratio [11]. To further delve into the relevance of the APOC3 and chromosome 11 loci, Jurado-Camacho et al. described the APOA1/C3/A5-ZPR1-BUD13 cluster and its impact on several lipid traits including, HDL-C and TG [41]. These observations agree with our results of the intron variant APOC3 rs5128 as significantly associated to the AIP index in Mexican adults highlighting that the connection of APOC3 and blood triglycerides is likely population independent [42]. Also associated with AIP were intronic variants, ARRB1 rs11236389 and CYBA rs12709102 the former codes for the cytosolic protein, arrestin beta 1, with immune functions but no clinical reports were found. The second one is part of the microbicidal oxidase system of phagocytes that has also been related to CAD, the thickness of the carotid intima media, and as a direct indicator of atherogenicity and obesity validating in part our observations [43]. Although the link between lipid metabolism and variants rs11236389 and rs12709102 here identified have not been previously reported, it might not necessarily be an unexpected observation since these genes, or their paralogs seem to bear variants in relation to cardiovascular risk [1,43].
TTN/CCDC141 rs10497525 is an intron variant of the large sarcomeric protein, titin, variations on this gene cause muscle disorders and cardiomyopathies [44]. TTN/CCDC141 is highly expressed in the heart [45] suggesting its potential role in biochemical pathways and cardiovascular health, but not yet discussed under the scope of dyslipidemia and atherogenic indexes. Our results may give rise to the biochemical connection between heart health, blood lipid levels and genetics identified in adults under 53 years.
The last variant associated with the AIP index was KCND3 rs6703437, this gene codes for a potassium channel responsible for smooth muscle contraction and it is associated with the Brugada syndrome and cardiac conduction [46]. KCND3 rs6703437 is 0.6 Kb apart and in partial LD with variant rs672757, this latter directly associated with obesity in patients with asthma [47], hinting towards a potential role of heart disease under a lipid imbalance.
Overall and according to the recent literature, the variants here associated with the AIP index may be indicative of cardiovascular health, heart function, lipid transport, and metabolism. Our observations confirm previous correlations between lipid levels and genes, APOC3, TTN/CCDC141, KCND3, CYBA, and ARRB1 and attest for the first time to a genetic relationship with AIP.
For the CI2, we identified six variants on intergenic and non-coding loci, four of them on chromosome 12 with few or no reports of their clinical relevance. We identified DIPK2B rs4294309 an intron variant located on chromosome Xp11.3. DIPK2B codes for a protein kinase domain 2B and known variants influence autism and intestinal carcinoma, but its relation to lipid metabolism or cardiovascular health has not been previously reported. However, DIPK2B maybe indirectly related to lipid and lipid indexes since its association with autism has been linked to alterations of cholesterol levels, decreased HDL-C, apolipoprotein A1 (ApoA1), and apolipoprotein B (ApoB) (48,49), suggesting a potential lipid-gene-autism relationship that may possibly pinpoint to a genetic marker. On chromosome 12 we identified four variants associated with CI2, rs6582413, rs12817366, rs34115639, and rs10880344 the two former on the Long Intergenic Non-Protein Coding RNA 2451, LINC02451, and the two latter in intergenic regions. Genome-wide linkage and meta-analyses of chromosome 12 have confirmed the presence of variants relevant to premature myocardial infarction and atherogenic plaque of the carotid intimal media, but these reported loci do not appear to be in close LD with the variants here listed. It is possible that several regions on chromosome 12 point towards genetic regulation or coding genes correlated to the CI2 and cardiovascular health that together may be considered as a polygenic cluster on chromosome 12.
Genetic variation on 10q21.3 here, rs7762658, rs11251177, have been associated with coronary artery disease in the GENOA study and in a pedigree of familial hypercholesterolemia [50]. Loci 10q.21.3 has been suggested to harbor genes with a role in subclinical coronary atherosclerosis [1], we identified this same locus as 10q21.3 rs1125117 with the highest statistical significance and size effect associated to CI2 (p-value 1.07e-7). Lange L. et al. mentioned that this variant is enriched in families with hypertension which is one of the future goals of the present cohort, i.e., the identification of markers predictive of hypertension and cardiovascular health supporting the relevance of cluster 10q21.3 in particular variant rs1125117.
Other variants associated with CI2 were observed on KCND3, DIPK2B, and LIPC/ALDH1A2 which have already been identified in lipoprotein and dyslipidemia studies. Here, KCND3 rs6703437 was associated to both, AIP and CI2 indexes (p-value 2.06e-6 and 1.7e-5) suggesting a concomitant association of this variant with HDL-C and triglycerides. Current reports on KCND3 indicate a link between genetic variation and cognitive impairment [51,52] suggesting a shared relationship between neurological disorders and lipids and hence the importance of monitoring genetic variation associated with lipid indexes as potential predictors of several physiological systems.
Associated to CI2 was also the gene LIPC/ALDH1A2 with the dual function of triglyceride hydrolase and ligand/bridging factor for receptor-mediated lipoprotein uptake. LIPC/ALDH1A2 rs261342 is located on the 5’ promoter region of LIPC and has been strongly associated with HDL-C in women as part of a haplotype [53] and used to assess CVD risk and its relation to lipids and apolipoproteins [54]. We found this variant associated with CI2 confirming its relationship to HDL-C levels, in males and females, i.e., in a sex independent manner opposed to the sexual dimorphism listed in previous results [53]. For CI2, we corroborate previous associations between this atherogenic index and 10q21.3 rs11251177, and to loci clustered on chromosomes 12 and 6. Also, we confirmed that gene variation on LIPC and KCND3 impact CI2 interindividual variation and the latter may have a stronger health impact as we found it associated with both, AIP and CI2.
5. Conclusions
Our observations introduce variants not previously reported and provide to the collection of loci associated to the lipid indexes AIP and CI2. The clinical utility of these SNVs as indicators of cardiovascular disease risk remains to be investigated, but its association to genetic variation contributes to the not fully accounted genetic impact on lipids [48]. Here, we attest the relationship between genetics and the atherogenic indexes AIP and CI2 that to the best of our knowledge has not been previously reported. We showed that the genetic variants listed here are associated to these lipid indexes overlapping previous reports of genetic associations to specific blood lipids. Future studies should aim to integrate and validate a list of genetic markers consolidating knowledge on genetics and lipids that could be directed to cardiovascular prevention.
6. Patents
No patents are filed or intended from the work reported in this manuscript.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Admixture of the study population; Table S1: All genetic variants associated to Atherogenic Plasma Index and Castelli 2 Index (p-value < 0.51.E-2). Supplemental Table S1: All genetic variants and its association to Atherogenic Plasma Index.
Author Contributions
SR-M and EC-R contributed to the cohort conceptualization, volunteer recruitment, sample management, clinical processing, clinical analyses, and genetic conceptualization. T.T contributed to sample management, processing, genetic analyses, data bioinformatic and statistical analysis and writing the manuscript. RC-R contributed to the cohort conceptualization, volunteer recruitment, and clinical data acquisition. KR-C, MR-D wrote sections of the manuscript and contributed to population statistical analyses. DK defined patient grouping, revised the manuscript and bioethics documentation. VG-C defined the genetics study, analyzed and interpreted results, wrote the paper, and contributed to sample processing, and data acquisition. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This research was funded by CONACyT- FOSSIS 2016-1-272994 and 272795. Tomas-Texis is being awarded with a scholarship from Conacyt under the Ciencias Bioquimicas Graduate Program at UNAM, Mexico.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of Instituto Nacional de Cardiología Ignacio Chávez (protocol code 13-802 and date of approval 16/05/2017) and at INMEGEN CEI 2017/20.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patient(s) to publish this paper”
Data Availability Statement
Data available on request from the corresponding author due to privacy restrictions and multiple institutions managing the future purpose of the cohort as a longitudinal study.
Acknowledgments
We gratefully acknowledge the technical support of Cintia K. Guzman Cruz during DNA extraction and sample management.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lange LA, Lange EM, Bielak LF, Langefeld CD, Kardia SL, Royston P, Turner ST, Sheedy PF, Boerwinkle E, Peyser PA. Autosomal genome-wide scan for coronary artery calcification loci in sibships at high risk for hypertension. Arterioscler Thromb Vasc Biol (2002) 22:418– 423. [CrossRef]
- Herrington W, Lacey B, Sherliker P, Armitage J, Lewington S. Epidemiology of Atherosclerosis and the Potential to Reduce the Global Burden of Atherothrombotic Disease. Circ Res (2016) 118:535–546. [CrossRef]
- Araújo JA, Lusis AJ. “Atherosclerosis.,” Encyclopedia of Endocrine Diseases. Elsevier (2004). p. 282–288. [CrossRef]
- Scipione CA, Cybulsky MI. Early atherogenesis: new insights from new approaches. Curr Opin Lipidol (2022) 2022:2–12. [CrossRef]
- Huang Y, Gao L, Xie X, Tan SC. Epidemiology of dyslipidemia in Chinese adults: meta- analysis of prevalence, awareness, treatment, and control. Popul Health Metr (2014) 12:28. [CrossRef]
- Bayram F, Kocer D, Gundogan K, Kaya A, Demir O, Coskun R, Sabuncu T, Karaman A, Cesur M, Rizzo M, et al. Prevalence of dyslipidemia and associated risk factors in Turkish adults. J Clin Lipidol (2014) 8:206–216. [CrossRef]
- Urina-Jassir M, Pacheco-Paez T, Paez-Canro C, Urina-Triana M. Statin associated adverse reactions in Latin America: A scoping review. BMJ Open (2021) 11:. [CrossRef]
- Pu J, Romanelli R, Zhao B, Azar KMJ, Hastings KG, Nimbal V, Fortmann SP, Palaniappan LP. Dyslipidemia in Special Ethnic Populations. Endocrinol Metab Clin North Am (3AD) 45:205–216. [CrossRef]
- Gao P, Wen X, Ou Q, Zhang J. Which one of LDL-C /HDL-C ratio and non-HDL-C can better predict the severity of coronary artery disease in STEMI patients. BMC Cardiovasc Disord (2022) 22:318. [CrossRef]
- Teslovich TM, Musunuru K, Smith A V, Edmondson AC, Stylianou IM, Koseki M, Pirruccello JP, Ripatti S, Chasman DI, Willer CJ, et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature (2010) 466:707–713. [CrossRef]
- Hindy G, Dornbos P, Chaffin MD, Liu DJ, Wang M, Selvaraj MS, Zhang D, Park J, Aguilar- Salinas CA, Antonacci-Fulton L, et al. Rare coding variants in 35 genes associate with circulating lipid levels—A multi-ancestry analysis of 170,000 exomes. Am J Hum Genet (2022) 109:81–96. [CrossRef]
- Weissglas-Volkov D, Aguilar-Salinas CA, Nikkola E, Deere KA, Cruz-Bautista I, Arellano- Campos O, Muñoz-Hernandez LL, Gomez-Munguia L, Ordoñez-Sánchez ML, Linga Reddy PMV, et al. Genomic study in Mexicans identifies a new locus for triglycerides and refines European lipid loci. J Med Genet (2013) 50:298. [CrossRef]
- Weissglas-Volkov D, Aguilar-Salinas CA, Sinsheimer JS, Riba L, Huertas-Vazquez A, Ordoñez-Sánchez ML, Rodriguez-Guillen R, Cantor RM, Tusie-Luna T, Pajukanta P. Investigation of Variants Identified in Caucasian Genome-Wide Association Studies for Plasma High-Density Lipoprotein Cholesterol and Triglycerides Levels in Mexican Dyslipidemic Study Samples. Circ Cardiovasc Genet (2010) 3:31–38. [CrossRef]
- Olamoyegun M, Oluyombo R, Asaolu S. Evaluation of dyslipidemia, lipid ratios, and atherogenic index as cardiovascular risk factors among semi-urban dwellers in Nigeria. Ann Afr Med (2016) 15:194–199. [CrossRef]
- Liu H, Liu K, Pei L, Li S, Zhao J, Zhang K, Zong C, Zhao L, Fang H, Wu J, et al. Atherogenic Index of Plasma Predicts Outcomes in Acute Ischemic Stroke. Front Neurol (2021) 12:. [CrossRef]
- Dobiášová M, Frohlich J. The plasma parameter log (TG/HDL-C) as an atherogenic index: correlation with lipoprotein particle size and esterification rate in apoB-lipoprotein-depleted plasma (FER(HDL)). Clin Biochem (2001) 34:583–588. [CrossRef]
- Rivera-Mancía S, Colín-Ramírez E, Cartas-Rosado R, Infante O, Vargas-Barrón J, Vallejo M. Indicators of accumulated fat are stronger associated with prehypertension compared with indicators of circulating fat: A cross-sectional study. Medicine (Baltimore) (2018) 97:. [CrossRef]
- Colín-Ramírez E, Rivera-Manciá S, Infante-Vázquez O, Cartas-Rosado R, Vargas-Barrón J, Madero M, Vallejo M. Protocol for a prospective longitudinal study of risk factors for hypertension incidence in a Mexico City population: The Tlalpan 2020 cohort. BMJ Open (2017) 7:. [CrossRef]
- Rivera-Mancía S, Colín-Ramírez E, Cartas-Rosado R, Infante O, Vargas-Barrón J, Vallejo M. Indicators of accumulated fat are stronger associated with prehypertension compared with indicators of circulating fat: A cross-sectional study. Medicine (Baltimore) (2018) 97:. [CrossRef]
- Liu T, Liu J, Wu Z, Lv Y, Li W. Predictive value of the atherogenic index of plasma for chronic total occlusion before coronary angiography. Clin Cardiol (2021) 44:518–525. [CrossRef]
- Fernández-Macías JC, Ochoa-Martínez AC, Varela-Silva JA, Pérez-Maldonado IN. Atherogenic Index of Plasma: Novel Predictive Biomarker for Cardiovascular Illnesses. Arch Med Res (2019) 50:285–294. [CrossRef]
- Koca TT, Tugan CB, Seyithanoglu M, Kocyigit BF. The Clinical Importance of the Plasma Atherogenic Index, Other Lipid Indexes, and Urinary Sodium and Potassium Excretion in Patients with Stroke. Eurasian J Med (2019) 51:171–175. [CrossRef]
- Team RC. A Language and Environment for Statistical Computing. https://www.R-project.org. (2014).
- Alexander DH, Lange K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinformatics (2011) 12:246. [CrossRef]
- Turner S, Armstrong LL, Bradford Y, Carlson CS, Crawford DC, Crenshaw AT, de Andrade M, Doheny KF, Haines JL, Hayes G, et al. Quality Control Procedures for Genome-Wide Association Studies. Curr Protoc Hum Genet (2011) 68:1.19.1-1.19.18. [CrossRef]
- Alexander DH, Lange K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinformatics (2011) 12:1–6. [CrossRef]
- Gonzalez-Covarrubias V. Lipidomics in longevity and healthy aging. Biogerontology (2013) 14:663–672. [CrossRef]
- Boyle AP, Hong EL, Hariharan M, Cheng Y, Schaub MA, Kasowski M, Karczewski KJ, Park J, Hitz BC, Weng S, et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res (2012) 22:1790–1797. [CrossRef]
- Afsin A, Kaya H, Suner A, Uzel KE, Bursa N, Hosoglu Y, Yavuz F, Asoglu R. Plasma atherogenic indices are independent predictors of slow coronary flow. BMC Cardiovasc Disord (2021) 21:608. [CrossRef]
- Millán J, Pintó X, Muñoz A, Zúñiga M, Rubiés-Prat J, Pallardo LF, Masana L, Mangas A, Hernández-Mijares A, González-Santos P, et al. Lipoprotein ratios: Physiological significance and clinical usefulness in cardiovascular prevention. Vasc Health Risk Manag (2009) 5:757– 765. [CrossRef]
- Kammar-García A, López-Moreno P, Hernández-Hernández ME, Ortíz-Bueno AM, Martínez- Montaño M de LC. Atherogenic index of plasma as a marker of cardiovascular risk factors in Mexicans aged 18 to 22 years. Proc (Bayl Univ Med Cent) (2020) 34:22–27. [CrossRef]
- Ahn SS, Lee LE, Pyo JY, Song JJ, Park YB, Lee SW. Atherogenic index of plasma predicts cerebrovascular accident occurrence in antineutrophil cytoplasmic antibody-associated vasculitis. Lipids Health Dis (2020) 19:1–11. [CrossRef]
- Du R, Li M, Wang X, Wang S, Li S, Tian H, Wu Y, Zhang C. LDL-C/HDL-C ratio associated with carotid intima-media thickness and carotid plaques in male but not female patients with type 2 diabetes. Clin Chim Acta (2020) 511:215–220. [CrossRef]
- Talmud PJ, Hawe E, Miller GJ, Humphries SE. Nonfasting Apolipoprotein B and Triglyceride Levels as a Useful Predictor of Coronary Heart Disease Risk in Middle-Aged UK Men. Arter Thromb Vasc Biol (2002) 22:1918–1923. [CrossRef]
- Ko A, Cantor RM, Weissglas-Volkov D, Nikkola E, Reddy PMVL, Sinsheimer JS, Pasaniuc B, Brown R, Alvarez M, Rodriguez A, et al. Amerindian-specific regions under positive selection harbour new lipid variants inLatinos. Nat Commun (2014) 5:. [CrossRef]
- Luo H, Zhang X, Shuai P, Miao Y, Ye Z, Lin Y. Genetic variants influencing lipid levels and risk of dyslipidemia in Chinese population. J Genet (2017) 96:985–992. [CrossRef]
- Song Y, Zhu L, Richa M, Li P, Yang Y, Li S. Associations of the APOC3 rs5128 polymorphism with plasma APOC3 and lipid levels: a meta-analysis. (2015). [CrossRef]
- Wu J, Zhou Q, Wei Z, Wei J, Cui M. Atherogenic Index of Plasma and Coronary Artery Disease in the Adult Population: A Meta-Analysis. Front Cardiovasc Med (2021) 0:1927. [CrossRef]
- Rocco CA, Mecikovsky D, Aulicino P, Bologna R, Sen L. Hypercholesterolemia Is Associated with the Apolipoprotein C-III (APOC3) Genotype in Children Receiving HAART: An Eight- Year Retrospective Study. PLoS One (2012) 7:39678. [CrossRef]
- Hsueh W-C, Nair AK, Kobes S, Chen P, Göring HHH, Pollin TI, Malhotra A, Knowler WC, Baier LJ, Hanson RL. Identity-by-Descent Mapping Identifies Major Locus for Serum Triglycerides in Amerindians Largely Explained by an APOC3 Founder Mutation. Circ Cardiovasc Genet (2017) 10:. [CrossRef]
- Jurado-Camacho PA, Cid-Soto MA, Barajas-Olmos F, García-Ortíz H, Baca-Peynado P, Martínez-Hernández A, Centeno-Cruz F, Contreras-Cubas C, González-Villalpando ME, Saldaña-Álvarez Y, et al. Exome Sequencing Data Analysis and a Case-Control Study in Mexican Population Reveals Lipid Trait Associations of New and Known Genetic Variants in Dyslipidemia-Associated Loci. Front Genet (2022) 13:807381. [CrossRef]
- Davis JP, Vadlamudi S, Roman TS, Zeynalzadeh M, Iyengar AK, Mohlke KL. Enhancer deletion and allelic effects define a regulatory molecular mechanism at the VLDLR cholesterol GWAS locus. Hum Mol Genet (2019) 28:888. [CrossRef]
- Van Den Berg ME, Warren HR, Cabrera CP, Verweij N, Mifsud B, Haessler J, Bihlmeyer NA, Fu YP, Weiss S, Lin HJ, et al. Discovery of novel heart rate-associated loci using the Exome Chip. Hum Mol Genet (2017) 26:2346. [CrossRef]
- Savarese M, Maggi L, Vihola A, Jonson PH, Tasca G, Ruggiero L, Bello L, Magri F, Giugliano T, Torella A, et al. Interpreting Genetic Variants in Titin in Patients With Muscle Disorders. JAMA Neurol (2018) 75:557–565. [CrossRef]
- Tharp CA, Haywood ME, Sbaizero O, Taylor MRG, Mestroni L. The Giant Protein Titin’s Role in Cardiomyopathy: Genetic, Transcriptional, and Post-translational Modifications of TTN and Their Contribution to Cardiac Disease. Front Physiol (2019) 10:1436. [CrossRef]
- Ye D, Zhou W, Hamrick SK, Tester DJ, Kim CSJ, Barajas-Martinez H, Hu D, Giudicessi JR, Antzelevitch C, Ackerman MJ. Acacetin, a Potent Transient Outward Current Blocker, May Be a Novel Therapeutic for KCND3 -Encoded Kv4.3 Gain-of-Function-Associated J-Wave Syndromes. Circ Genomic Precis Med (2022) 15:419–428. [CrossRef]
- Zhu Z, Guo Y, Shi H, Liu CL, Panganiban RA, Chung W, O’Connor LJ, Himes BE, Gazal S, Hasegawa K, et al. Shared genetic and experimental links between obesity-related traits and asthma subtypes in UK Biobank. J Allergy Clin Immunol (2020) 145:537–549. [CrossRef]
- El-Ansary A, Chirumbolo S, Bhat RS, Dadar M, Ibrahim EM, Bjørklund G. The Role of Lipidomics in Autism Spectrum Disorder. Mol Diagn Ther (2020) 24:31–48. [CrossRef]
- Tierney E, Remaley AT, Thurm A, Jager LR, Wassif CA, Kratz LE, Bailey-Wilson JE, Bukelis I, Sarphare G, Jung ES, et al. Sterol and lipid analyses identifies hypolipidemia and apolipoprotein disorders in autism associated with adaptive functioning deficits. Transl Psychiatry (2021) 11:. [CrossRef]
- Yano Y, Butler KR, Hall ME, Schwartz GL, Knopman DS, Lirette ST, Jones DW, Wilson JG, Hall JE, Correa A, et al. Associations of Nocturnal Blood Pressure With Cognition by Self- Identified Race in Middle-Aged and Older Adults: The GENOA (Genetic Epidemiology Network of Arteriopathy) Study. J Am Heart Assoc (2017) 6:. [CrossRef]
- Zanni G, Hsiao C-T, Fu S-J, Tang C-Y, Capuano A, Bosco L, Graziola F, Bellacchio E, Servidei S, Primiano G, et al. Novel KCND3 Variant Underlying Nonprogressive Congenital Ataxia or SCA19/22 Disrupt K V 4.3 Protein Expression and K+ Currents with Variable Effects on Channel Properties. Int J Mol Sci Artic Int J Mol Sci (2021) 22:4986. [CrossRef]
- Pollini L, Galosi S, Tolve M, Caputi C, Carducci C, Angeloni A, Leuzzi V. Molecular Sciences KCND3-Related Neurological Disorders: From Old to Emerging Clinical Phenotypes. [CrossRef]
- Feitosa MF, Myers RH, Pankow JS, Province MA, Borecki IB. LIPC variants in the promoter and intron 1 modify HDL-C levels in a sex-specific fashion. Atherosclerosis (2009) 204:171– 177. [CrossRef]
- Talmud PJ, Drenos F, Shah S, Shah T, Palmen J, Verzilli C, Gaunt TR, Pallas J, Lovering R, Li K, et al. Gene-centric association signals for lipids and apolipoproteins identified via the HumanCVD BeadChip. Am J Hum Genet (2009) 85:628–642. [CrossRef]
Figure 1.
Impact of APOC, ARRB1, LINC02451, and 10q21.3 variants on AIP and Castelli 2 indexes. A. association between AIP and APOC3 rs5128 (left) and ARRB1 rs11236389 (right), horizontal middle line depicts lipid indexes mean value. B. association between CI2 and 10q21.3 rs11251177 (left) and LINC02451 rs65822413 (right), horizontal line reflect the mean values of each lipid index, statistical significance of mean differences is displayed at the top.
Figure 1.
Impact of APOC, ARRB1, LINC02451, and 10q21.3 variants on AIP and Castelli 2 indexes. A. association between AIP and APOC3 rs5128 (left) and ARRB1 rs11236389 (right), horizontal middle line depicts lipid indexes mean value. B. association between CI2 and 10q21.3 rs11251177 (left) and LINC02451 rs65822413 (right), horizontal line reflect the mean values of each lipid index, statistical significance of mean differences is displayed at the top.
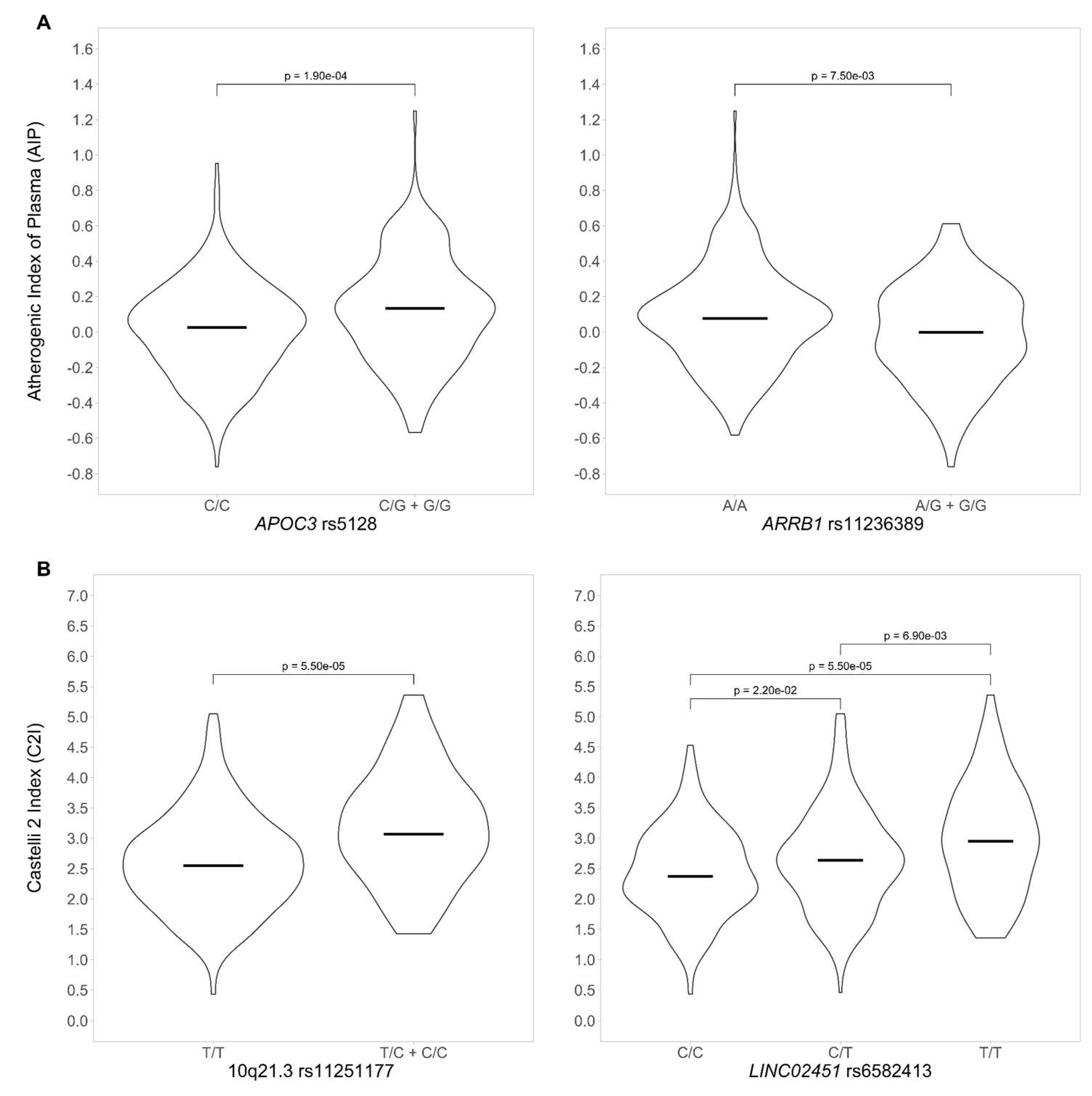

Table 1.
Population characteristics.
| Males N = 170 |
Females N = 256 |
All N = 426 |
|
|---|---|---|---|
| Age, y | 38 (18-53) | 40 (17-52) | 39 (17-53) |
| Weight, kg | 78.1 (51.2 -125) | 63.1 (41.7 - 119) | 70 (41.7 - 125) |
| Height, m | 1.70 (1.50 - 1.99) | 1.57 (1.36 - 1.72) | 1.61 (1.36 - 1.99) |
| BMI, kg/m2 | 26.8 (16.9 - 40.3) | 26.2 (16.9 - 47.1) | 26.4 (16.8 - 47.1) |
| Waist circumference, cm | 94.0 (63.0 – 130) | 85.0 (54.0 – 126) | 89.0 (54.0 – 130) |
| Glucose, mg/dL | 94.0 (72.0 – 166) | 90.0 (74.0 – 241) | 92.0 (72.0 – 241) |
| Uric acid, mg/dL | 6.34 (1.82 – 10.0) | 4.62 (2.30 – 7.58) | 5.31 (1.82 – 10.0) |
| Creatinine, mg/dL | 0.95 (0.62 – 1.40) | 0.69 (0.44 – 1.19) | 0.77 (0.44 – 1.40) |
| Cholesterol, mmol/dL | 4.62 (2.96 – 8.30) | 4.39 (2.16 – 7.06) | 4.49 (2.16 – 8.30) |
| HDL-C, mmol/dL | 1.10 (0.60 – 2.12) | 1.22 (0.73 – 2.27) | 1.16 (0.60 – 2.27) |
| LDL-C, mmol/dL | 3.09 (0.98 – 6.87) | 2.93 (0.54 – 5.38) | 3.00 (0.54 – 6.87) |
| Triglycerides (TG), mmol/dL | 1.48 (0.47 – 15.4) | 1.22 (0.22 – 5.86) | 1.32 (0.22 – 15.34) |
| Dyslipidemia, n (%) | 119 (70%) | 85 (72%) | 304 (71%) |
| Castelli risk index 2 (CI2)1 | 2.91 (1.25 – 5.36) | 2.45 (0.43 – 4.92) | 2.60 (0.43 – 5.36) |
| Atherogenic index of plasma (AIP)2 | 0.48 (-0.18 – 1.61) | 0.38 (-0.40 – 1.17) | 0.42 (-0.40 – 1.61) |
Values indicate the mean and range. 1 CI2 = LDL-C/HDL-C; 2AIP = log (TG/HDL-C).
Table 2.
Proportion of lipid levels outside reference values.
| High TG > 1.9 mmol/L | High cholesterol > 5 mmol/L | High LDL-C > 3.9 mmol/L | Low HDL-C < 1.04 mmol/L | |
|---|---|---|---|---|
| Males %, n1 | 30%, 51 | 35.3%, 60 | 16.5%, 28 | 38%, 65 |
| Females %, n1 | 16%, 41 | 22.3%, 59 | 10.9%, 28 | 26%, 67 |
| p-value2 | 8.74E-06 | 2.32E-03 | 1.80E-03 | 2.59E-05 |
1 n, sample size for each sex. 2p-value for sex differences using an ANOVA test.
Table 3.
Variants associated with atherogenic indexes.
| Gene | Chr | rs identifier | Coefficient | p-value |
|---|---|---|---|---|
| Variants associated to AIP | ||||
| APOA1/APOC3 | 11 | rs5128 | 0.094 | 2.61E-06 |
| CYBA | 16 | rs12709102 | 0.078 | 3.91E-06 |
| ARRIB1 | 11 | rs11236389 | -0.102 | 6.63E-06 |
| TTN/CCDC141 | 2 | rs10497528 | 0.089 | 8.29E-06 |
| APOA1/APOC3 | 11 | rs5072 | 0.091 | 8.94E-06 |
| Variants associated to CI2 | ||||
| Intergenic | 10q21.3 | rs11251177 | 0.606 | 1.07E-07 |
| LINC02451 | 12 | rs6582413 | 0.259 | 5.19E-07 |
| LINC02451 | 12 | rs12817366 | 0.254 | 1.88E-06 |
| Intergenic | 12 | rs34115639 | 0.244 | 7.06E-06 |
| Intergenic | 12 | rs10880344 | -0.233 | 7.10E-06 |
| Intergenic | 6 | rs7762658 | -0.247 | 2.03E-06 |
| LIPC/ALDH1A2 | 15 | rs261342 | 0.227 | 1.10E-06 |
| DIPK2B | 23 | rs4294309 | 0.306 | 1.18E-05 |
| KCND3 | 1 | rs6703437 | -0.234 | 1.76E-05 |
Lead SNVs generated by GLM considering uric acid, weight, waist circumference, sex, and genetic ancestry. The raw output of the GLM analysis is in ST2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



