
Submitted:
10 April 2023
Posted:
11 April 2023
You are already at the latest version
Abstract
The UNUSUAL FLORAL ORGANS (UFO) gene is an essential regulatory factor of class B genes and plays a vital role in the process of inflorescence primordial and flower primordial development. We investigated the role of UFO genes in soybean to better understand the development of floral organs through gene cloning, expression analysis, and gene knockout. In situ hybridization demonstrated predominant expression of the GmUFO1 gene in the flower primordium. Phenotypic observation of GmUFO1 knockout mutant lines (Gmufo1) showed obvious alteration in floral organ number and shape, and mosaic organ formation. In contrast, GmUFO2 knockout mutant lines (Gmufo2) showed no obvious difference in floral organs. However, the GmUFO1 and GmUFO2 double knockout lines (Gmufo1ufo2) showed a more severe mutant phenotype than the Gmufo1 lines. Gene expression analysis also showed differences in the expression of major ABC function genes in the knockout lines. Based on the phenotypic and expression analysis, our results suggest the major role of GmUFO1 in the regulation of flower organ formation in soybean and that GmUFO2 does not have any direct effect, but it might have an interaction role with GmUFO1 in the regulation of flower development. In conclusion, the present study identified UFO genes in soybean and improved our understanding of floral development that could be useful for flower design in hybrid soybean breeding.
Keywords:
GmUFOs
; knockout
; floral organ number
; floral organ shape
; soybean
1. Introduction
Cell division control is necessary for the proper arrangement of flower primordia and is regulated by the plant hormones auxin and cytokinin. The vast majority of angiosperm flowers have four-wheeled floral organs, which consist of sepals, petals, stamens, and the carpel, in that order, moving from the outside to the interior of the flower. The well-known "ABC model" is the functional model of floral organ attribute genes in Arabidopsis thaliana [1,2]. This model correctly describes the molecular process of flower organ primordium attributes, and it has been the subject of a great deal of research. The A functional genes AP1 (APETALA1) and AP2, B functional genes AP3 and PI (PISTILLATA), C functional gene AG (AGAMOUS), and E functional gene SEP (SEPALLATA) are the most typical genes studied in model plants [3]. This well-known "ABC model" is a scientific model of the process through which a pattern of gene expression is produced by flowering plants in their meristems, which in turn leads to the appearance of an organ oriented toward sexual reproduction [4]. This model correctly describes the molecular process of flower organ primordium attributes, and it has been the subject of a great deal of research.
The UFO gene is an F-box gene that encodes E3 ubiquitin ligase and was first discovered in Arabidopsis thaliana, where mutant flowers lack typical petals and stamens [1]. In Arabidopsis thaliana, the UFO gene co-activates the expression of the downstream AP3 gene with the AP1 gene as a co-activator of the LFY gene, thus preserving petal characteristics and petal development. LFY controls the temporal specificity of AP3 expression, while UFO and AP1 control the spatial pattern [5,6]. UFO has been reported to interact with the SCF E3 ubiquitin ligase subunits, ASK1 and CUL1, as well as subunits of the COP9 signalosome, which suggest the possible role of UFO in the ubiquitination of proteins that are involved in flower development [7,8]. The interaction of the UFO and ASK1 proteins may enhance the degradation of the downstream B functional gene AP3 and the PI inhibitor by the UFO gene, resulting in the activation and production of the B functional gene [5,9,10]. Strong loss-of-function mutations in the UFO gene cause flower abnormalities in all whorls. In sepals and petals, homeotic changes, organ number, and phyllotaxis are the most severe [8,11]. Sepals replace petals, and carpels replace stamens; however, mosaic organs are also prevalent in both whorls of the ufo mutant. UFO, similar to its Antirrhinum homolog FIM, is expressed in a complicated temporal and geographic pattern throughout floral development [12,13]. In floral meristems, transcription is inhibited in stage 1 and triggered in stage 2 in the central area. By stage 3, the core meristem loses expression but extends laterally in cone form. By stage 5, expression is confined to the petal primordia and remains there throughout floral organ development. AP3 and PI patterns are formed during stage 3 [14,15] before petal/stamen organ activation during stage 5, supporting UFO’s significance in B class function [16].
Soybean (Glycine max) is a globally important crop that is a rich source of edible protein and oil [17]. The reproductive period of soybean (starting from flower initiation till maturity) is essential for higher yield and quality [18]. Identifying and understanding the molecular mechanism of the genes involved in the regulation of the reproductive period and flower development in soybean are key targets for yield and quality improvement in soybean. In the present study, we reported that GmUFOs play a crucial role in regulating flower development, especially the quantity and characteristics of petals in connection with the development of the stamen. The GmUFOs mutation caused floral organ morphological defects and altered the expression of ABC flower development genes.
2. Results
2.1. Identification of UFO Genes
In the present study, we used the AtUFO gene (AT1G30950.1) of Arabidopsis thaliana as a query to search its orthologs in soybean. We identified two orthologues, Glyma.05G134000 and Glyma.08G088700, and named them GmUFO1 and GmUFO2, respectively. For phylogenetic analysis, we used GmUFO1 and GmUFO2, as well as another nine UFO genes from eight species, to construct the phylogenetic tree. The phylogenetic alignment showed that GmUFO1 and GmUFO2 were more closely related to the UFOs of leguminous plants (Figure 1A). Amino acid sequence alignment and protein 3D model analysis showed that the UFO protein domain was relatively conserved in these species (except cotton), and both GmUFO proteins in soybean had a conserved F-box (Figure 1B). Protein physical property analysis showed that the amino acid length of UFOs in these species was between 357 and 444, and the polypeptide isoelectric points were almost constant (Table S2).
2.2. Spatial and Temporal Expression Profiles of the GmUFO1 Gene in Soybean Flower Development
RNA in situ hybridization was used to detect GmUFO1 expression in the soybean flower primordium at different development stages (Figure 2A, B). In stage 1, the flower meristem formation stage, GmUFO1 was expressed in the central area of the whole flower meristem. In stage 2, which represents the sepal primordium formation stage, GmUFO1 was expressed between the sepal primordium and the carpel primordium, and the signal was also detected near the central dome of the sepal primordium. In stage 3, the petal primordium formation stage, UFO1 was mainly expressed in the petal primordium, but a few signals were detected in the sepal primordium. Stage 4 represents the stamen and carpal primordium formation stage, and in this stage, UFO1 was expressed in the petal primordia and almost absent in elongating sepal primordia. In stage 5, when the formation of all flower organs occurs, UFO1 was expressed at the boundary region of the petal between sepal and stamen.
2.3. CRISPR/Cas9 System Generated Target Mutations for GmUFO Genes
To obtain the GmUFO mutants, CRISPR/Cas9-mediated genome editing of the GmUFO genes was performed, and the target site for both GmUFO1 and GmUFO2 was located ahead of the F-box domain (Figure 3A, B). We obtained four T0 generation transgenic lines and 40 T1 generation transgenic lines. In the T1 generation, the GmUFOs were sequenced, and based on the GmUFO target site sequencing data, we identified 10 Gmufo1 single mutants, six Gmufo2 single mutants, and three Gmufo1ufo2 double mutants (Figure 3C). These mutants were used for further investigation in our study.
2.4. Knockout of the GmUFO1 Gene Caused Abnormal Flower Organs
All 10 mutant plants obtained in Gmufo1 of the T1 knockout lines showed heterozygosity at the GmUFO1 target site, whereas no mutation was found at the GmUFO2 gene, and three of the mutants were selected for further analysis (Figure 4). The Gmufo1 seedlings showed no obvious differences compared with the wild type (Figure 3D), whereas the floral organ was different from the wild type (Figure 4A). Compared with the wild type, Gmufo1 showed floral organ number and shape alterations in sepals, petals, and stamens, and also mosaic flower organ formation with green sepal-like petals (Figure 4A, B).
Furthermore, five ABC function genes, namely GmAP1a, GmAP2, GmAP3, GmPI04G, and GmAG, were selected, and their expression was detected using qPCR in the three Gmufo1 lines and the wild type. The expression levels of GmAP1a, GmAP3, GmPI04G, and GmAG decreased in all three lines, while the expression levels of GmAP2 increased (Figure 4C). We also examined the expression of GmUFO1 and GmUFO2 gene transcripts in Gmufo1 and found that they did not differ from the wild type (Figure 4D).
2.5. Knockout of the GmUFO2 Gene Had no Effect on Flower Organs
Six positive plants obtained in Gmufo2 of the T1 knockout lines showed heterozygosity at the GmUFO2 target site, whereas no mutants possessed at the GmUFO1 gene, and three of them were selected for further analysis (Figure 5). Gmufo2 seedlings had no obvious differences compared with the wild type (Figure 3E) and did not show differences in floral organ shape or number (Figure 5A). Further statistics also showed that the number of Gmufo2 organs to the wild type had not changed (Figure 5A, B). The five ABC functional genes (i.e., GmAP1a, GmAP2, GmAP3, GmPI04G, and GmAG) in the three Gmufo2 lines were detected using qPCR. The GmPI04G expression decreased, while GmAP2 expression increased in all three Gmufo2 lines (Fig. 5C). We examined the expression of GmUFO1 and GmUFO2 gene transcripts in Gmufo2 and found that they did not differ from the wild type (Figure 5D).
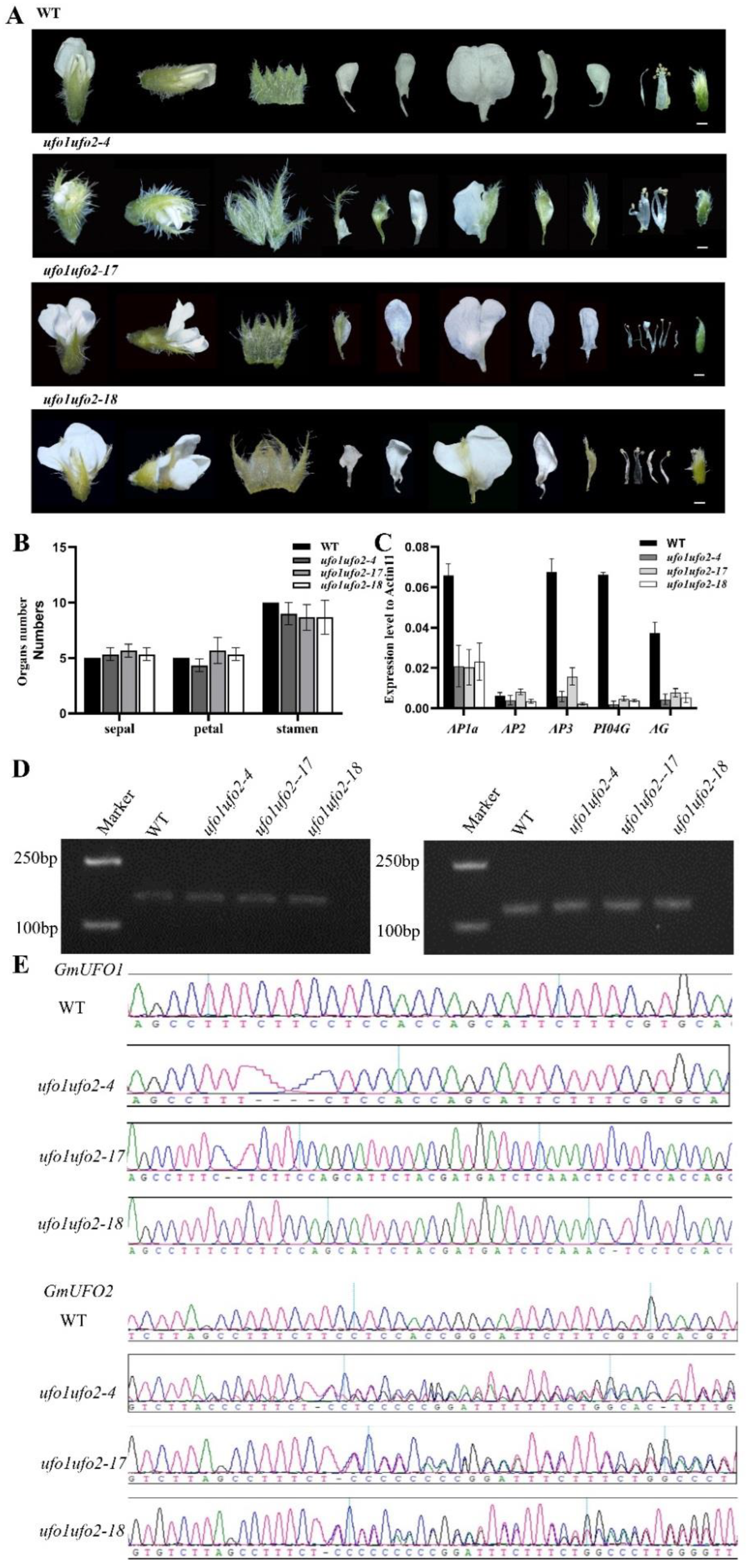
2.6. Double Knockout of GmUFO1 and GmUFO2 Had More Serious Defects in Organ Number and Shape
Among the T1 knockout lines, three lines were double mutated and were selected for further analysis. In the case of the GmUFO1 gene, line Gmufo1ufo2-4 had a 4-bp deletion at the target site, Gmufo1ufo2-17 had a 1-bp deletion at the target site, and Gmufo1ufo2-18 showed a 1-bp deletion after the target site. However, in the case of the GmUFO2 gene, all three selected mutant plants were heterozygotes at the target site (Figure 6). The vegetative appearance showed no obvious difference in the seedlings of double mutants compared with the wild type (Figure 3F), whereas the floral organ showed even more serious defects than Gmufo1 single mutants compared with the wild type (Figure 4A and Figure 6A). We observed more mosaic organs formed with sepal-like petals and petal-like stamens, and the alteration of organ number and shape in the double mutant relative to the Gmufo1 single mutant (Figure 6A).
The three selected double mutant lines showed obvious variations in the number of sepals, petals, and stamens (Figure 6A, B). The expression of the five ABC functional genes (GmAP1a, GmAP2, GmAP3, GmPI04G, and GmAG) in the three lines was detected using qPCR. The expression levels of all of the ABC genes, GmAP1a, GmAP2, GmAP3, GmPI04G, and GmAG, decreased in all three Gmufo1ufo2 plants, and the decrease was greater than that in Gmufo1 (Figure 6C). We also detected the expression of GmUFO1 and GmUFO2 gene transcripts in Gmufo1ufo2 and found that they did not differ from the wild type (Figure 6D).
3. Discussion
In previous studies, the ufo mutant of different plant species has been observed to show the abnormal development of flower organs. However, there was a difference among the ufo mutants of various plants in regard to abnormal flower organ development, which might be due to some differences in the underlying molecular mechanism originating from plant evolution. For example, the single mutant of UFO in Arabidopsis thaliana (Atufo) showed a slight reduction in inflorescences at co-florescence, but the meristem of all primary and co-flowering buds in Atufo stopped growing after the appearance of structures fused with sepal-carpus organs. The attribute changes in Atufo flower organs were mainly concentrated in the second and third round organs, and the quantitative changes were reflected in all round organs. There are always deletions in the second round of early primordium development [20]. In the model legume Lotus japonicus, an orthologue of the Arabidopsis UFO gene mutation resulted in the pfo mutant [21]. In the pfo mutant, the flower primordium was unable to form normal flower organs, resulting in some sepal-like structures with filamentous structures, and petals failed to develop [21]. In our study, the changes in flower organs in soybean ufo mutants were mainly concentrated in the second whorl organ, and the increase in sepals and the decrease in stamens were also found in the first and third whorl organs. Even the most severe mutant phenotype of Gmufo did not show complete replacement of the second whorl organ with sepals, and sepal-like petals mostly appeared on the ventral part. Previous studies have documented that by controlling the interaction between UFO and LFY, the UFO gene can regulate the downstream targets of AP3 and PI, thereby affecting the flower phenotype [8]. In this context, the interaction among the UFO, ABC, LFY, WUS, and CAL genes has been explored previously in different studies at the petal formation stage. For example, LFY interacts with UFO, CAL, and AP1, and UFO is expressed downstream of AP1 [22,23,24]. The interaction between LFY and UFO, as well as between LFY and AP1, is mutual and activates each other [23,24]. LFY directly activates AG, and AG is also activated by WUS, but AG acts as a WUS antagonist [25,26,27]. In addition, AG directly activates AP3 and PI. Moreover, AP2 is an AG antagonist, and AP2 activates AP3 and PI directly [28]. UFO also directly activates AP3 and PI [29]. Based on the above interactions of UFO with ABC and other genes, it is evident that the loss-of-function mutation of the UFO gene affects the expression of ABC genes, both directly and indirectly. For example, it indirectly reduces the expression of downstream AP3 and PI by affecting LFY expression, which in turn affects AG and AP1 expression [23,30] and directly decreases the expression of downstream AP3 and PI [29]. Our qPCR analysis is consistent with previous findings. We also demonstrated that knockout of GmUFO in Gmufo1 and Gmufo1ufo2 soybean mutants reduced the expression of ABC genes, GmAP1a, GmAP2, GmAP3, GmPI04G, and GmAG (Figure 4C, Figure 5C, and Figure 6C). Hence, GmUFO can be regarded as a key regulator of ABC function genes in soybean. The ABC model of flower organ development works as follows: A alone controls sepal development, A and B together regulate petal development, B and C together control stamen identity, and C regulates carpel development alone. In our study, we observed that the Gmufo1ufo2 and Gmufo1 mutants showed abnormal development of the sepals, petals, and stamens (increased sepals, decreased stamens, and petal looks like sepals), but carpel development was normal. Therefore, in agreement with the ABC model, our results also showed that knockout of GmUFO genes reduced the expression of ABC genes, viz., A (GmAP1a, GmAP2), B (GmAP3, GmPI04G), and C (GmAG) in the ufo1ufo2 and ufo1 mutants, which in turn led to the abnormal development of sepals, petals, and stamens. Although GmAG expression was reduced in the ufo1ufo2 and ufo1 mutants, carpel development was still normal in the mutant plants. This suggests that there might be some other interaction factors working together with GmAG in the regulation of carpel development in soybean, which needs to be further explored. The ufo2 mutant did not show any difference in flower development compared with the wild type. We also observed only the expression levels of GmAP2 and GmPI04G, which decreased in the ufo2 mutant, and the expression of the remaining ABC genes did not differ relative to the wild type. In addition, our study documented that the effect of the ufo1ufo2 mutant on flower development was more severe than that of ufo1. These findings suggest the major role of GmUFO1 in the regulation of flower organ formation in soybean. GmUFO2 did not have any direct effect on flower development, but it might have an interaction role with GmUFO1 in the regulation of flower development.
In conclusion, our work demonstrated that GmUFO genes regulated the development of flower organs, viz., soybean petals, stamens, and sepals, in soybean by controlling the expression of ABC genes. These findings will be the basis for further investigation of the in-depth mechanism involved in the GmUFO-mediated regulation of flower development in soybean. This, in turn, will have a significant effect on soybean hybrid breeding, leading to the development of high-yielding cultivars.
4. Materials and Methods
4.1. GmUFO Identification and Cloning
The Arabidopsis thaliana UFO gene (AT1G30950.1) sequence was downloaded from The Arabidopsis Information Resource (TAIR) database (https://www.arabidopsis.org/), and this gene was used as a query to identify putative orthologs of UFO genes in the Glycine max using BLASTp. The GmUFO1 gene fragment was amplified from soybean cultivar ‘Williams 82’, and the primers were designed using Primer5 software (https://primer.software.informer.com/5.2/) (Table S1). PCR amplifications were performed using 2 × Phanta Max Master Mix (Dye Plus) DNA (Vazyme Biotech Co., Nanjing, Jiangsu, China) polymerase in a 50 μL mix containing 2 μmol L−1 of each primer, 25 units of 2 × Phanta Max Master Mix (Dye Plus), and 1 μL of cDNA. The PCR experiments were performed as follows: 95°C for 30 s, 32 cycles of 95°C for 15 s, 56°C for 15 s, 72°C for 2 min, and 72°C for 5 min. The PCR products were cloned using the TA cloning vector pGEM-Teasy (Promega Biotech Co., Beijing, China) and sequenced. After verifying the sequences, these products were used to construct the in situ hybridization and subcellular localization vector.
4.2. Phylogenetic Analysis and Physical Properties of UFO Genes
Phylogenetic analysis was performed using MEGA11 software [31] following the neighbor-joining method to analyze the evolutionary relationships among the 11 UFO genes from nine plants, including both monocots and dicots, namely Arabidopsis, soybean, Medicago, cowpea, kidney bean, cotton, rice, maize, and barley.
ProtParam (https://web.expasy.org/protparam/database) was used to determine the physical properties of the UFO proteins, such as the molecular weight, the length of each protein, and the protein isoelectric point (PI). Protein structure prediction was performed using an online website (https://robetta.bakerlab.org/domains/robetta/).
4.3. In Situ Hybridization
Tissue fixation for in situ hybridization was performed following the protocol of Feng et al [32]. The tissues were embedded in paraffin (Paraplast Plus, Sigma-Aldrich, Saint Louis, MO, USA) and sliced into 8 μm sections with a microtome (Leica, Wetzlar, Germany). The 3’-region of GmUFO1 cDNA was subcloned and used as a template to generate sense and antisense RNA probes (Table S1). Digoxigenin-labeled RNA probes were prepared with a DIG RNA Labeling Kit (T7/SP6) (Cat. No. 11175025910, Roche) according to the manufacturer’s instructions. The slides were observed under a bright field using a Leica microscope (DMI8, Leica, Wetzlar, Germany).
4.4. Subcellular Localization
For the GmUFO protein subcellular experiment, the full-length CDS (removal terminator) sequence of GmUFO1 was amplified and linked with a GFP fluorescent protein sequence and inserted into the pUC19 vector using NdeΙ and SalΙ for double digestion. The subcellular expression vector pUC19-GFP-GmUFO1 was constructed containing the 35S promotor controlling expression of the GFP-GmUFO1 fusion construct. A modified polyethylene glycol (PEG) method was used for the transfection of protoplasts [33,34,35]. The pUC19-GFP-GmUFO1 and pUC19-AHL22-mCherry plasmids were extracted by TIANprep Mini Plasmid Kit (TIANGEN Biotech Co., Beijing, China), and the two plasmids were transiently transformed into Arabidopsis protoplasts. The transformed protoplasts were subsequently cultured at room temperature in the dark for 16 h, and fluorescence observation and photography were performed using a Leica microscope (DMI8, Leica, Wetzlar, Germany).
4.5. Cytological Analysis
The floral primordium of wild-type ‘Williams 82’ during five developmental stages (stages 1–5) was observed by scanning electron microscopy. After removing the bracts, they were fixed in a formaldehyde alcohol acetic acid (FAA) fixation solution for at least 12 h. All materials were dehydrated with ethanol (30, 50, 70, 90, 95, 100%) and then dehydrated with ethanol-tert-butanol to 100% tert-butanol. The samples were dissected using a positive microscope and held in small Petri dishes with conductive tape. The floral primordium was scanned using InTouchScope Scanning Electron (JSM-IT500, Japan Electronics, Tokyo, Japan). The anatomical observation of flower organs was performed using an orthomicroscope (SZ2-ILST, Olympus Co., Tokyo, Japan) and photographed with cellSens Standard imaging software (Olympus cellSens Standard, Version 1.16, Olympus Co., Tokyo, Japan).
4.6. Gene Knockout and Material Identification
A Cas9/sgRNA plasmid construction kit (Viewsolid Biotech Co., Beijing, China) was used to construct the CRISPR/Cas9 plasmids. These plasmids contained a dicotyledon codon-optimized dpCas9 (under the GmUbi3 promoter) and a gRNA scaffold (under the GmU6-2 promoter). Construction of the CRISPR/Cas9 vector was performed following the manufacturer’s instructions (Viewsolid Biotech Co., Beijing, China). The constructed CRISPR/Cas9-gDNA plasmid was transformed into Agrobacterium EHA105, and soybean genetic transformation was performed following Yamada et al [36]. Genomic DNA was extracted from the seedlings of wild-type and all positive transgenic plants, and DNA fragments spanning the target sites were amplified by PCR. The target site for GmUFO1 was amplified with the primers GmUFO-F (5′-ATGGAAGGTTTTCACCCTTCTATGT-3′) and GmUFO1-R (5′-AGCATTGAAACTCTGGAAGGGC-3′). The target site for GmUFO2 was amplified with the primers GmUFO-F (5′-ATGGAAGGTTTTCACCCTTCTATGT-3′) and GmUFO2-R (5′-TTGGGCACGTTCAGCTTGCTCTTCTCA-3′).
4.7. Total RNA Extraction and qPCR
Total RNA was extracted using the Trizol method [37], and the cDNA was synthesized by the process of reverse transcription using the TransScript One-Step genomic DNA (gDNA) removal and cDNA synthesis Super-Mix kit (TransGen Biotech Co., Beijing, China), with gDNA removal in one step. The cDNA was diluted in sterile water to 100 ng/μl, with 2 μl diluent as a template for qPCR. qPCR was performed using 2 × RealStar Green Fast mix (GenStar Biosolutions Co., Beijing, China) on a Stratagene Mx3005P sequence detection system (Stratagene, La Jolla, CA, USA) following the manufacturer’s instructions. Three biological replicates were used to quantify the expression levels. Differences between groups were calculated using the 2−ΔΔCt method. The soybean GmActin 11 (Glyma.18G290800) gene was used as an internal control, and the relative expression levels were calculated. Primers used for RT-qPCR analysis are shown in Supplemental Table S1.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Primers used in this research; Table S2: Protein physical information of 11 UFO genes from nine species.
Author Contributions
H.Y performed the experiment and wrote manuscripts. X.Y, Z.Z and T.W helped to guide the experiment. J.F and Y.J helped to complete experiments. Y.Z and X.F discussed the results and wrote the manuscript. Y.Z, M.H.K, J.A.B and F.W revised the manuscript. X.F and Y.W designed the project and provided resources. All authors read and approved the final manuscript.
Funding
This research was funded by National Natural Science Foundation of China (U21A20215), Jilin Province Science and Technology Development Plan Project (20210302005NC), Zhejiang Lab (2021PE0AC04) and Hainan Yazhou Bay Seed Laboratory (B21HJ0101).
Data Availability Statement
The datasets used in the study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Levin, J.Z.; Meyerowitz, E.M. UFO: an Arabidopsis gene involved in both floral meristem and floral organ development. Plant Cell. 1995, 7, 529–548. [Google Scholar] [PubMed]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls - genetic interactions controlling flower development. Nature. 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Thomson, B.; Wellmer, F. Molecular regulation of flower development. Curr. Top. Dev. Biol. 2019, 131, 185–210. [Google Scholar] [PubMed]
- Sun, L.Y.; Nie, T.J.; Chen, Y.; Li, J.; Yang, A.X.; Yin, Z.F. Gene identification and tissue expression analysis inform the floral organization and color in the basal angiosperm Magnolia polytepala (Magnoliaceae). Planta. 2023, 257, 4. [Google Scholar] [CrossRef] [PubMed]
- Laufs, P.; Coen, E.; Kronenberger, J.; Traas, J.; Doonan, J. Separable roles of UFO during floral development revealed by conditional restoration of gene function. Development. 2003, 130, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.X.; Li, K.P.; Zhang, Z.R.; Li, J.G.; Wang, B.; Zhang, Z.M.; Zhu, Y.Y.; Pan, C.C.; Sun, K.; He, C.Y. Transcriptomic comparison sheds new light on regulatory networks for dimorphic flower development in response to photoperiod in Viola prionantha. BMC Plant Biol. 2022, 22, 336. [Google Scholar] [CrossRef]
- Wang, X.P.; Feng, S.H.; Nakayama, N.; Crosby, W.L.; Irish, V.; Deng, X.W.; Wei, N. The COP9 signalosome interacts with SCFUFO and participates in Arabidopsis flower development. Plant Cell. 2003, 15, 1071–1082. [Google Scholar] [CrossRef]
- Rieu, P.; Turchi, L.; Thévenon, E.; Zarkadas, E.; Nanao, M.; Chahtane, H.; Tichtinsky, G.; Lucas, J.; Blanc-Mathieu, R.; Zubieta, C. The F-box protein UFO controls flower development by redirecting the master transcription factor LEAFY to new cis-elements. Nat. Plants. 2023, 9, 315–329. [Google Scholar] [CrossRef]
- Pelosi, J.A.; Sessa, E.B. From genomes to populations: a meta-analysis and review of fern population genetics. Int. J. Mol. Sci. 2021, 182, 325–343. [Google Scholar] [CrossRef]
- Zhao, D.; Yang, M.; Solava, J.; Ma, H. The ASK1 gene regulates development and interacts with the UFO gene to control floral organ identity in Arabidopsis. Dev. Genet. 1999, 25, 209–223. [Google Scholar] [CrossRef]
- Levin, J.Z.; Meyerowitz, E.M. UFO: an Arabidopsis gene involved in both floral meristem and floral organ development. Plant Cell. 1995, 7, 529–548. [Google Scholar] [PubMed]
- Ingram, G.C.; Goodrich, J.; Wilkinson, M.D.; Simon, R.; Haughn, G.W.; Coen, E.S. Parallels between UNUSUAL FLORAL ORGANS and FIMBRIATA, genes controlling flower development in Arabidopsis and Antirrhinum. Plant Cell. 1995, 7, 1501–1510. [Google Scholar] [PubMed]
- Peréz-Mesa, P.; Suárez-Baron, H.; Ambrose, B.A.; González, F.; Pabón-Mora, N.J.E. Floral MADS-box protein interactions in the early diverging angiosperm Aristolochia fimbriata Cham.(Aristolochiaceae: Piperales). Evol. Dev. 2019, 21, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.J.; Ye, L.X.; Ai, X.Y.; Hu, C.G.; Cheng, Z.P.; Zhang, J.Z. Functional analysis of a PISTILLATA-like gene CcMADS20 involved in floral organs specification in citrus. Plant Sci. 2022, 6, 319. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Meng, Q.; Tan, X.M.; Yang, L.; Zhang, K.L.; Xu, Z.Q. Functional identification of the different regions in B-class floral homeotic MADS-box proteins IiAP3 and IiPI from Isatis indigotica. Physiol. Plantarum. 2022, 174, e13713. [Google Scholar] [CrossRef]
- Zhang, A.J.; He, H.B.; Li, Y.; Wang, L.X.; Liu, Y.X.; Luan, X.C.; Wang, J.X.; Liu, H.J.; Liu, S.Y.; Zhang, J.; et al. MADS-Box subfamily gene GmAP3 from Glycine max regulates early flowering and flower development. Int. J. Mol. Sci. 2023, 24, 2751. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Kim, W.S.; Oehrle, N.W.; Smith, J.R.; Gillman, J.D. Effect of heat stress on seed protein composition and ultrastructure of protein storage vacuoles in the Cotyledonary Parenchyma cells of soybean genotypes that are either tolerant or sensitive to elevated temperatures. Int. J. Mol. Sci. 2020, 21, 4775. [Google Scholar] [CrossRef]
- Lyu, J.; Cai, Z.D.; Li, Y.H.; Suo, H.C.; Yi, R.; Zhang, S.; Nian, H. The floral repressor GmFLC-like is involved in regulating flowering time mediated by low temperature in soybean. Int. J. Mol. Sci. 2020, 21, 1322. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Haughn, G.W. UNUSUAL FLORAL ORGANS controls meristem identity and organ primordia fate in Arabidopsis. Plant Cell. 1995, 7, 1485–1499. [Google Scholar] [CrossRef]
- Zhang, S.; Sandal, N.; Polowick, P.L.; Stiller, J.; Stougaard, J.; Fobert, P.R. Proliferating Floral Organs (Pfo), a Lotus japonicus gene required for specifying floral meristem determinacy and organ identity, encodes an F-box protein. Plant J. 2003, 33, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Jofuku, K.D.; den Boer. B.G.; Van Montagu.M.; Okamuro J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell. 1994, 6, 1211–1225. [Google Scholar]
- Bradley, D.; Ratcliffe, O.; Vincent, C.; Carpenter, R.; Coen, E. Inflorescence commitment and architecture in Arabidopsis. Science. 1997, 275, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Yanofsky, M.F. Activation of the Arabidopsis B class homeotic genes by APETALA1. Plant Cell. 2001, 13, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.A.; Bomblies, K.; Weigel, D. Activation of a floral homeotic gene in Arabidopsis. Science. 1999, 285, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Mena, C.; de Folter, S.; Costa, M.M.R.; Angenent, G.C.; Sablowski, R. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development. 2005, 132, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.G.; Kim, Y.J.; Muller, R.; Yumul, R.E.; Liu, C.Y. Pan Y.Y.; Cao X.F.; Goodrich J.; Chena X.M. AGAMOUS terminates floral stem cell maintenance in Arabidopsis by directly repressing WUSCHEL through recruitment of polycomb group proteins. Plant Cell. 2011, 23, 3654–3670. [Google Scholar] [CrossRef]
- Drews, N.; Bowman, J.L.; Meyerowitz, E.M. Negative regulation of the Arabidopsis homeotic gene AGAMOUS by the APETALA2 product. Cell. 1991, 65, 991–1002. [Google Scholar] [CrossRef]
- Chae, E.; Tan, Q.K.; Hill, T.A.; Irish, V.F. An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development. Development. 2008, 135, 1235–1245. [Google Scholar] [CrossRef]
- Ng, M.; Yanofsky, M.F. Function and evolution of the plant MADS-box gene family. Nat. Rev. Genet. 2001, 2, 186–195. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.Z.; Zhao, Z.; Tian, Z.X.; Xu, S.L.; Luo, Y.H.; Cai, Z.G.; Wang, Y.M.; Yang, J.; Wang, Z.; Weng, L.; et al. Control of petal shape and floral zygomorphy in Lotus japonicus. Proc. Natl. Acad. Sci. USA. 2006, 103, 4970–4975. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tiwari, S.B.; Hagen, G.; Guilfoyle, T.J. AUXIN RESPONSE FACTOR7 restores the expression of auxin-responsive genes in mutant Arabidopsis leaf mesophyll protoplasts. Plant Cell. 2005, 17, 1979–1993. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.S.; Jiang, L.W. Transient expression of fluorescent fusion proteins in protoplasts of suspension cultured cells. Nat. Protoc. 2007, 2, 2348–2353. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shen, W.J.; Chen, H.F.; Chu, L.T.; Xu, Y.C.; Zhou, X.C.; Liu, C.L.; Chen, C.M.; Zeng, J.H.; Liu, J.; et al. Characterization and subcellular localization of histone deacetylases and their roles in response to abiotic stresses in soybean. BMC Plant Biol. 2018, 18, 226. [Google Scholar] [CrossRef]
- Tetsuya, Y.; Satoshi, W.; Maiko, A.; Kyuya, H.; Keisuke, K. Cotyledonary node pre-wounding with a micro-brush increased frequency of Agrobacterium-mediated transformation in soybean. Plant Biotechnol. 2010, 27, 217–220. [Google Scholar]
- Rio, D.C.; Ares, M.J.; Hannon, G.J.; Nilsen, T.W. Preparation of cytoplasmic and nuclear RNA from tissue culture cells. Cold. Spring. Harb. Protoc. 2010, 6, 5441. [Google Scholar] [CrossRef]
Figure 1.
(a) Phylogenetic analysis of the selected 11 UFO proteins from nine species, namely Arabidopsis thaliana (AT1G30950), Glycine max (Glyma.05G134000, Glyma.08G088700), Medicago truncatula (Medtr4g094748), Vigna unguiculate (Vigun03g153100), Phaseolus vulgaris (Phvul.002G188300), Gossypium raimondii (Gorai.002G139700), Oryza sativa (Os06g45460), Zea mays (GRMZM2G109966, GRMZM2G360081), and Hordeum vulgare (Hr1G108970). The neighbor-joining tree was constructed using MEGA11, and the bootstrap value was set at 1000 replicates. (b) Amino acid sequence alignment of UFOs in soybean and other species. The orange line represents the F-box domain.
Figure 1.
(a) Phylogenetic analysis of the selected 11 UFO proteins from nine species, namely Arabidopsis thaliana (AT1G30950), Glycine max (Glyma.05G134000, Glyma.08G088700), Medicago truncatula (Medtr4g094748), Vigna unguiculate (Vigun03g153100), Phaseolus vulgaris (Phvul.002G188300), Gossypium raimondii (Gorai.002G139700), Oryza sativa (Os06g45460), Zea mays (GRMZM2G109966, GRMZM2G360081), and Hordeum vulgare (Hr1G108970). The neighbor-joining tree was constructed using MEGA11, and the bootstrap value was set at 1000 replicates. (b) Amino acid sequence alignment of UFOs in soybean and other species. The orange line represents the F-box domain.

Figure 2.
(a) In situ hybridization of the UFO1 gene in the soybean flower primordium. The periods from left to right are stage 1: flower primordium initiation, stage 2: sepal primordium formation stage, stage 3: petal primordium formation stage, stage 4: stamen and carpal primordium formation, and stage 5: formation of all flower organs. Bar = 100 μm. (b) Scanning electron microscopy (SEM) observations of in situ hybridization and stereoscopic models of GmUFO1 gene expression at five developmental stages. Bar = 100 μm. (c) Expression location of GmUFO1-GFP in Arabidopsis thaliana. Bar = 10 μm.
Figure 2.
(a) In situ hybridization of the UFO1 gene in the soybean flower primordium. The periods from left to right are stage 1: flower primordium initiation, stage 2: sepal primordium formation stage, stage 3: petal primordium formation stage, stage 4: stamen and carpal primordium formation, and stage 5: formation of all flower organs. Bar = 100 μm. (b) Scanning electron microscopy (SEM) observations of in situ hybridization and stereoscopic models of GmUFO1 gene expression at five developmental stages. Bar = 100 μm. (c) Expression location of GmUFO1-GFP in Arabidopsis thaliana. Bar = 10 μm.
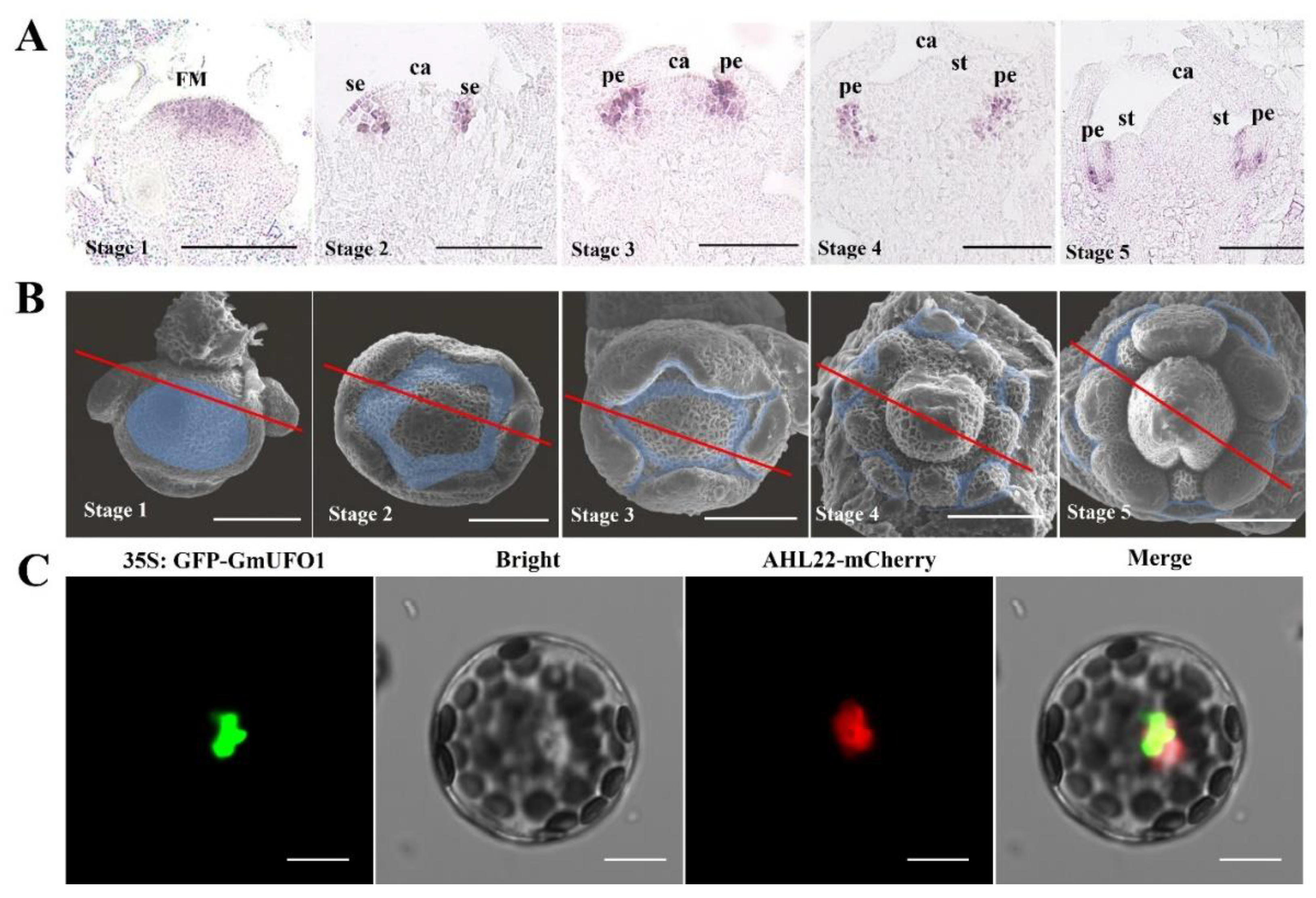
Figure 3.
Summary of CRISPR/Cas9-mediated genome editing of GmUFO genes. (a) Structural diagram of the CRISPR/Cas9 vector. (b) Target sites designed for GmUFO1 and GmUFO2 in CRISPR/Cas9. (c) Summary of mutation frequency in the T1 generation. (d-f) Seedling phenotype of Gmufo1 single mutant lines (d), Gmufo2 single mutant lines (e), and Gmufo1ufo2 double mutant lines (f) compared with the wild type. Scale bar = 10 cm.
Figure 3.
Summary of CRISPR/Cas9-mediated genome editing of GmUFO genes. (a) Structural diagram of the CRISPR/Cas9 vector. (b) Target sites designed for GmUFO1 and GmUFO2 in CRISPR/Cas9. (c) Summary of mutation frequency in the T1 generation. (d-f) Seedling phenotype of Gmufo1 single mutant lines (d), Gmufo2 single mutant lines (e), and Gmufo1ufo2 double mutant lines (f) compared with the wild type. Scale bar = 10 cm.

Figure 4.
(a) Floral organ phenotype of the three Gmufo1 mutant lines compared with the wild type. Scale bar = 1 mm. (b) Statistics of the number of sepals, stamens, and petals in Gmufo1 mutant lines. (c) ABC function gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo1-1, Gmufo1-15, and Gmufo1-16. All data presented are mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
Figure 4.
(a) Floral organ phenotype of the three Gmufo1 mutant lines compared with the wild type. Scale bar = 1 mm. (b) Statistics of the number of sepals, stamens, and petals in Gmufo1 mutant lines. (c) ABC function gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo1-1, Gmufo1-15, and Gmufo1-16. All data presented are mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
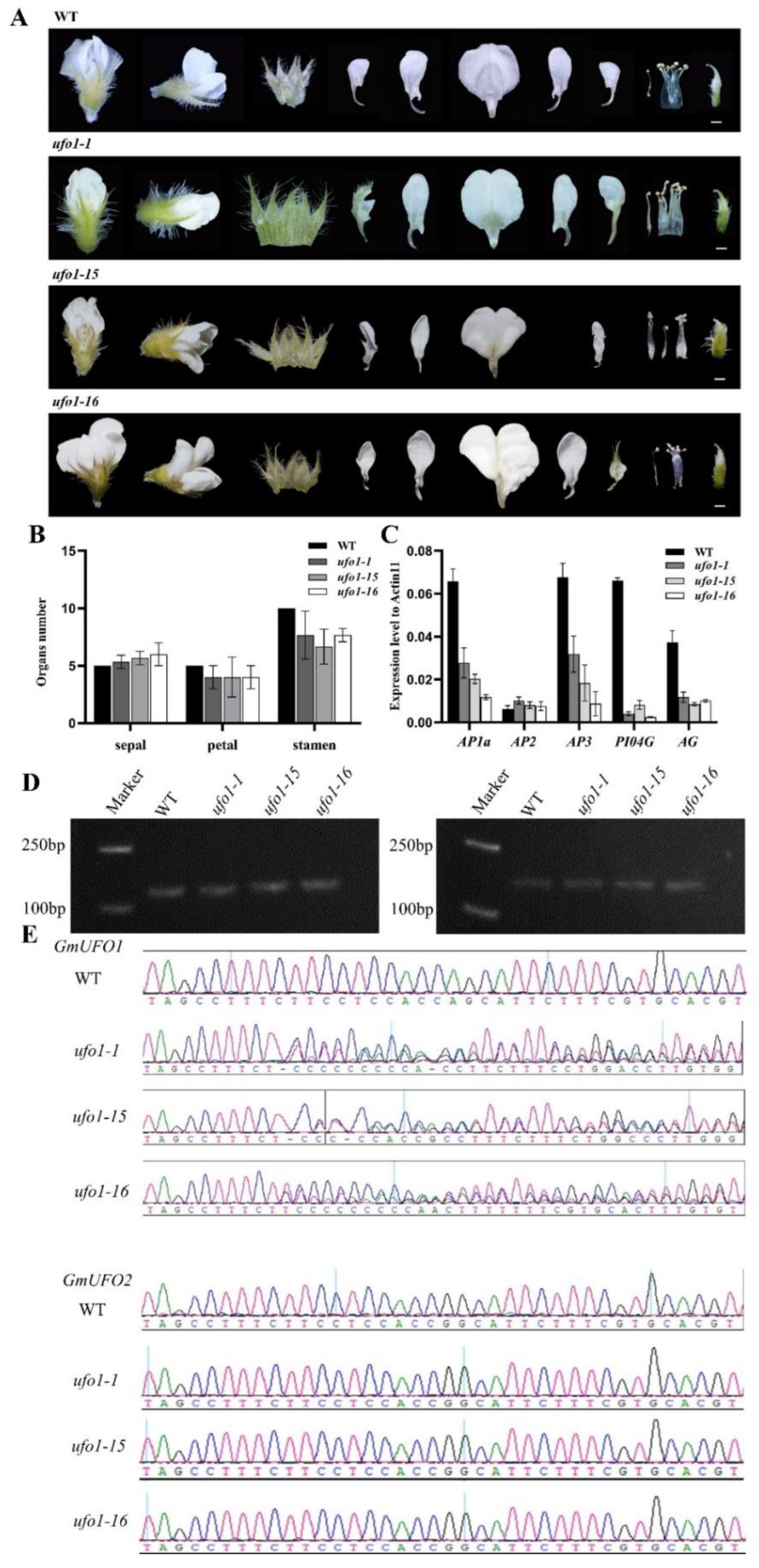
Figure 5.
(a) Floral organ phenotype of the three Gmufo2 mutant lines compared with the wild type. Scale bar =1 mm. (b) Statistics of the number of sepals, stamens, and petals in Gmufo2 mutant lines. (c) ABC model gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo2-1, Gmufo2-2, and Gmufo2-3. All data presented are mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
Figure 5.
(a) Floral organ phenotype of the three Gmufo2 mutant lines compared with the wild type. Scale bar =1 mm. (b) Statistics of the number of sepals, stamens, and petals in Gmufo2 mutant lines. (c) ABC model gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo2-1, Gmufo2-2, and Gmufo2-3. All data presented are mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
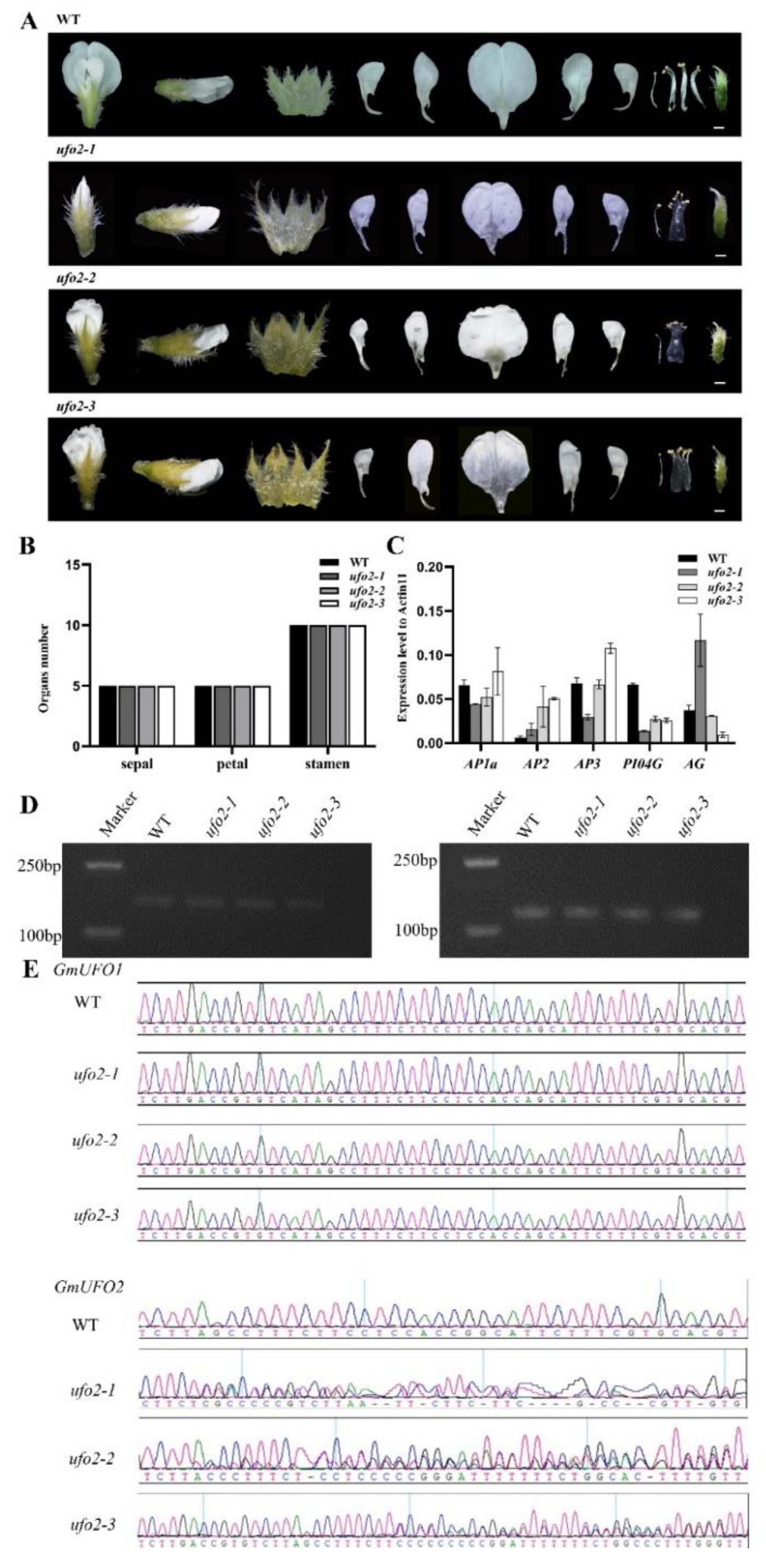
Figure 6.
(a) Floral organ phenotype of the three Gmufo1ufo2 mutant lines compared with the wild type. Scale bar = 1 mm. (b) Statistics of the number of sepals, stamens, and petals in the Gmufo1ufo2 mutant lines. (c) ABC model gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo1ufo2-4, Gmufo1ufo2-17, and Gmufo1ufo2-18. All data presented are the mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
Figure 6.
(a) Floral organ phenotype of the three Gmufo1ufo2 mutant lines compared with the wild type. Scale bar = 1 mm. (b) Statistics of the number of sepals, stamens, and petals in the Gmufo1ufo2 mutant lines. (c) ABC model gene expression was analyzed in the flowers of ‘Williams 82’, Gmufo1ufo2-4, Gmufo1ufo2-17, and Gmufo1ufo2-18. All data presented are the mean ± SD for three biological replicates. (d) GmUFO1 and GmUFO2 expression level in three mutant lines and the wild type. (e) CRISPR/Cas9 target site sequencing of GmUFO1 and GmUFO2 in the three selected lines.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



