
Submitted:
11 April 2023
Posted:
12 April 2023
You are already at the latest version
Abstract
This study examined the ventral process of C6 in extinct and extant Equus (sister taxa to Equus ferus caballus only) with the purpose of describing normal morphology and identifying anomalous variations relevant to recent studies describing a congenital malformation in E. ferus caballus. Overall, 83 specimens from 9 museums and 3 research/educational facilities were examined – 71 extinct specimens from 12 species and 12 extant specimens from 5 species. The lateral view revealed a large convexity existed in the ventral process between the cranial ventral tubercle (CrVT) and the caudal ventral tubercle (CVT) in the earliest ancestor Hyracotherium grangeri 55mya that receded throughout the millennia to a smaller convexity in E. ferus caballus and sister taxa. The CrVT was visibly shorter and narrower than the CVT with a constricted section directly ventral to the transverse process essentially demarcating the CrVT and CVT. No congenital malformations were evident. As the ventral process of C6 is an integral component for muscle attachment in supporting the head/neck during posture and locomotion, this would indicate the caudal module in the cervical column might be compromised when a partial or complete absence of the CVT is detected via radiographs in modern E. ferus caballus.
Keywords:
Caudal ventral tubercle (CVT)
; Cranial ventral tubercle (CrVT)
; Equus ferus caballus
; Longus colli
; Sixth cervical vertebra (C6)
; Ventral tubercle.
1. Introduction
In the mammalian neck evolutionary processes have produced a wide variety of morphological specialisations that include three complex cervical modules [1,2,3]. With few exceptions, the three modules comprise of seven cervical vertebrae (CV) - upper (C1, C2), middle (C3–C5) and lower (C6, C7). Each vertebrae in the upper and lower modules present highly specialised conserved traits specifically designed for a diverse range of daily functions [1,2,3]. When conserved traits are shared by a group of phylogenetically related animals, they are referred to as a ‘body plan’ [4]; as demonstrated in mammals by the specialised ventral process of C6 [1,2,3,5,6,7,8,9,10,11,12].
In Equus ferus caballus, veterinary text defines C6 as an ‘atypical’ vertebra inclusive of C1, C2 and C7; whereas C3–C5 are ‘typical’ [7,8,12,13,14,15]. The differentiation between CV that are typical and atypical are - a typical CV is relatively unspecialised; while an atypical CV present highly specialised structural features for soft tissue attachment/s critical for specific head and neck biomechanics [1,2,3,12,13,14,15,16,17,18,19]. In C6, its atypical feature is a bilateral ventral projection traversing the entire length of the vertebral body which is referred to in the literature by several names; ventral process, ventral lamina (lamina ventralis), ventral transverse process and or ventral tubercle [6,7,8,12,13,14,15,16,18,19,20,21,22,23,24,25,26,27,28,29]. For the purpose of this study the ventral projection will be referred to as the ventral process of C6.
The ventral process of C6 demonstrates two specifc osteological features; a cranial ventral tubercle (CrVT) and caudal ventral tubercle (CVT) and each provides attachments for the deep perivertebral muscle, longus colli [7,12,13,14,15,16]. Rhombach, Stubbs and Clayton [16] demonstrate the CrVT in E. ferus caballus is the attachment site for the cervical portion of the multi bundled longus colli muscle; while the CVT is the insertion point for the single bundled thoracal tendon of the longus colli muscle that extends caudally to either T5 or T6. This muscle aids in the fixation, stabilisation, rotation and flexion of the CV; and also acts as a site of force redirection during muscle contraction cranial to, and or caudal to C6 [12,13,14,15,18].
In domestic mammals, the literature primarily describes the ventral process of C6 relative to educational instruction or veterinary management [6,7,8,12,20,21,22,23,24,25,26,27,28,29]. Although, extensive literature on pathologies and peer reviewed journals describing anomalous presentations are well documented in some domestic mammals [15,19,24,27,28,30,31,32,33,34,35,36,37,38,39,40]; historically, congenital malformations in the fossil record are rare [41]. Therefore, when two domestic species presenting similar congenital malformations in the cervicothoracic junction involving the ventral process of C6 are reported, namely Bos taurus domesticus [30,31,32] and E. ferus caballus [34,35,36,37,38,39,40], further investigations into the fossil record are warranted.
Clinically, the condition in B. taurus domesticus, known as complex vertebral malformation (CVM) is fatal, with the animal either stillborn or spontanesouly aborted during gestation [31,32]. Hence, preservation in the fossil record is unlikely due to fragmentaion of the associative growth plates relative to the ventral process of C6 [7], plus poor preservation of the specimen, and or predation [41]. However, in modern E. ferus caballus, the condition is not known to be fatal, and the absent CVT on the ventral process of C6 is evident in mature adults, and therefore preservation is more likely [34,35,36,37,38,39,40,41]. Although clinically the absent CVT on C6 is yet to be defined, several authors have postulated, or documented via case studies symptoms conducive to the congenital malformation [34,38,40]. Therefore, the question is - is this congenital malformation of the absent CVT in C6 a recent event in modern E. ferus caballus or a normal variation in the population?
To examine this question further, this study will examine the fossil record in extinct and extant Equidae. It would be expected that such evidence, if evident, would be determinable as other anomalous variations of the CVT in C6 have been reported in the fossil record. For example; cervical rib facets on the CVT of C6 in Pleistocene Coelodonta antiquitatis [42] and Mammuthus promigenius [43], and a complete absence of the right CVT in C6 of Dendrohyrax arboreus [44]. Therefore, the purpose of this study is to determine if anomalous variations of the ventral process in C6 associated with extant modern E.ferus caballus is strictly a post-domestication abnormality with possible functional implications, or is a normal variation in the population.
2. Materials and Methods
2.1. Ethical Statement
No equids were euthanized for the purpose of this study and observational research was conducted from specimens in museum collections and educational facilities.
2.2. Terminology
2.3. Materials
Twenty-five applications to examine extinct and extant Equus collections were made to various museums and research/educational facilities. Collections only housing modern E. ferus caballus were excluded and for reasons of logistics, availability, and suitability of the collection, only 9 museums and three educational/research facilities were utilised for this study. Museums - American Museum of Natural History, New York, USA (AMNH), Monrepos Museum for Human Behavioural Evolution, Neuwied, Germany (MMHBE), Natural History Museum, Berlin, Germany (NHMB), Natural History Museum of Los Angeles County, USA (LACM), Oxford University Museum of Natural History, UK (OUMNH), Naturalis Biodiversity Center, Leiden, The Netherlands, (NBC), Smithsonian Institute, Washington, USA (USNM), The Page Museum Rancho La Brea, Los Angeles, USA (RLB), Yokohama Horse Museum, Yokohama, Japan (YHM). Educational facilities included the Australian College of Equine Podiotherapy, Yarck, Australia (ACEPT), Equine Studies, Asch, The Netherlands (ES), and the University of Florida, USA (UF).
Peer reviewed publications that nominated catalogued specimens presenting evidence of complete ventral processes of C6 were also examined.
To be eligible for the study, the specimens required minimal damage to at least one ventral process where a clear structural definition of the CrVT and CVT was determined. The purpose was to ascertain a normal and or anomalous presentation (Figure 1).
In those specimens where the ventral process is clearly divided into a CrVT and CVT, such as Pliohippus pernix (Figure 2), and where portions of either the CrVT or CVT on one side are lost to various pressures such as taphonomy or predation, then a revised observation was documented outlining the intact tubercles.
Collectively, museums, educational/research facilities and peer reviewed literature yielded 68 extinct Equus specimens, 12 extant Equus specimens (non E. ferus caballus) and three peer reviewed pubications describing specimens - totalling 83 specimens. The age of each specimen at expiration was defined by the complete ossification of the caudal growth plate (epiphysis) on C6 and subsequently included. Adults were defined by a closed epiphysis with no delineation, while sub adults were determined by incomplete epiphyseal ossification and obvious delineation. When classification of a species was undifferentiated, the specimen was labelled Equus sp.
In museum specimens, 12 species of extinct Equus were examined totalling 68 specimens with geologic dates. Eighteen specimens were undifferentiated and categorised as Equus sp., according to the museum’s records (Table 1).
In extant Equus, five non E. ferus caballus species were examined with one undifferentiated specimen, totalling 12 specimens. In museums, specimens were recorded according to their catalogue number in the collection, except in one (NBC – Equus sp., mounted skeleton not E. ferus caballus). In educational facilities specimens were classified by the animal’s name pre-mortem (Table 2).
2.3. Methods
The ventral process of C6 was observed, described, and the left or right lateral profile/s were digitally recorded in extinct and extant species of Equus (not E. ferus caballus).
The assigned shape/s describing the lateral profile of the ventral process of C6 were derived from either botanical and or biological nomenclature, or from previous publications describing CV.
3. Results
3.1. General Anatomy
In all 83 specimens, the ventral process of C6 appeared on the lateral ventral border in an antero-posterior orientation separate and distal to the transverse process. In 82/83, the CrVT was anterior to the transverse process while the CVT remained posterior. Eurohippus messelensis was the exception and did not present a CrVT nor CVT according to the depictions and radiographs [48]. Excluding Eurohippus messelensis, in 82 specimens, the lateral profile of the ventral process depicts a separation between the CrVT and CVT that is noted by a convexity in the morphology directly distal to the transverse process. The extent of the convexity was not uniform between specimens and ranged from a lesser convexity between the CrVT and CVT to a distinct hourglass convexity (Figure 1a and Figure 2).
From the ventral view, the 82 specimens presented a tube-like morphology of the ventral process with a central constriction distal to the transverse process. Enthesis patterns were evident on both tubercles in relation to the attachment of the longus colli muscle – the cranial longus colli attachment to the CrVT and the thoracal tendon of the longus colli to the CVT. The CVT appeared more expanded than the CrVT, and when both cranial and caudal ventral tubercles were present, the overall outside width between left to right tubercles were greater across the CrVTs (Figure 4).
No congenital malformations or anomalous presentations as previously described were evident in the ventral process of C6 in the 83 specimens, nor in those specimens unsuitable for the study.
3.2. Lateral profile of the ventral process
In all 82/83 specimens, the CVT appeared longer than the CrVT. The size of the convexity in the ventral process presented three distinct morphological variations that were determined by size, and simply described as small, medium, and large (Figure 5).
In extinct Equus, the 71 specimens from museums and the literature presented a total of 38 small, 19 medium, and 13 large convexities with Eurohippus messelensis presenting a morphologically different bilateral ventral process with no convexities (Table 4).
Eurohippus messelensis presented bilateral ventral processes with no convexities that appeared as well-developed ventrolateral appendages from the vertebral body [48], similar to a trapezoid in profile. Gidley’s (1903) description of the ventral process describes the ventral surface of the 6th cervical as flat, turning downward laterally into the wing-like transverse processes, which are more strongly developed than in Equus [46]. Old literature describes these as cranial and caudal ventral transverse processes [49].
All 12 specimens in extant Equus presented small convexities in the ventral process, yet the demarcation between the CrVT and CVT was still clear (Table 5).
3.3. Variations of the Epiphyseal
During the examination of extinct and extant Equus variations of the CVT epiphyseal could be noted in sub adults as evident or absent. The absence was not anomalous variation, but a lost portion of the epiphyseal through destructive process such as degradation or human intervention through preparation of the vertebra (Figure 6).
In the extant specimen of Przewalskii’s horse from Equine Studies (Rideg), the caudal epiphyseal was intact upon dissection and maceration, its loss can be attributed to final preparations, such as degreasing.
4. Discussion
This study examined the morphology of the ventral process of C6 in extinct and extant Equus. The findings revealed the mammalian body plan for this specialised atypical structure remained a conserved trait in Equidae from its earliest ancestor – Hyracotherium grangeri [47]. The only noted variation in morphology was the size of the convexity between the CrVT and CVT receding from large to small over 55 million years, except in Eurohippus messelensis where a descent to modern E. ferus caballus could not be established [48].
The relationship between the convexity of the ventral process and its recedence might be functionally related to the increase in size of the head/neck, change in feeding regimes from predominantly browsing to predominantly grazing, and or environmental pressures such as predation [50]. The altered morphology of the ventral process led to an elongated CrVT and CVT with a reduced convexity, thus providing a longer attachment site for the Longus colli muscle to adequately support head/neck function. However, to verify this further morphometric studies would be necessary.
As for the variations seen in individual cervical vertebrae (C1–C7), they can be described in terms of intravertebral versus intervertebral modules, and defined as a component of a developing organism (e.g., an embryo) that are semi-autonomous relative to pattern formation and differentiation [51,52]. Arnold (2021) mapped the different possible modular schemes for cervical vertebrae based on developmental, morpho-functional and paleontological perspectives [1]. Similarly, Randau and Goswami (2017) demonstrated patterns of shape integration reflected modular organisation in felids [52], while in other carnivores, larger modules supported the integration between adjacent vertebrae to meet locomotory and functional demands [53].
In Equidae, as per most mammals, the cervical spine is held as vertically as possible to reduce the distance between the weight of the head and the sustaining cervicothoracic junction (C5–T2) [1,2]. This leads to the stereotypical vertical, s-shaped and self-stabilising resting posture of the cervical spine [54,55]. As head/neck movements start from this posture, orientation and gaze changes in the sagittal plane are restricted to the occiput to C2 module and C6–T1 module [1,2]. Arnold (2021) states that this functional modularity is supported by two prominent bony processes in mammals that provide major muscle attachment sites for head and neck motion: the enlarged spinous process on C2, and the ventral process of C6 [1]. These findings support Bainbridge’s (2018) comments that the ventral process of C6 acts as a site of force redirection during muscle contraction cranial to, and or caudal to C6 [15].
In modern E. ferus caballus, reports of the ventral process of C6 being unilaterally or bilaterally absent have been reported in multiple breeds from numerous countries [33,34,35,36,37,38,39,40]. Clinical symptoms are yet to be defined through longitudinal studies, yet the presentation across breeds and geographical regions suggests a familial connection [34]. With respect to the concepts of cervical modularity, this anomalous variation of the ventral process of C6, not seen in ancestral Equus, infers head and neck function could be compromised through the limitation, or lack thereof, for the attachment site of the thoracal longus colli muscle. May-Davis and Walker (2015) concurred through case studies based on gross examination of the thoracal Longus colli muscle, that it either and or, relocated with altered tendon morphology, hypertrophied on the affected side, and the authors also described potential symptomatic observations of afflicted horses bearing this anomalous variation [40]. Similar studies concurred with these findings and further described ataxic behaviours [34,38,56].
The findings from this study indicate the anomalous variations reported in the ventral process of C6 in modern E. ferus caballus is a recent event and not indicative of a normal variation within the population of Equus. Nor was it present in pre-domestic Equus or ancestral Equus. However, the limitations in this study were identified as the small number of extant Equus (non E. ferus caballus), the limitation of complete specimens in the fossil record and access to further specimens. Even so, the evidence to date suggests this anomalous variation of the ventral process in C6, was not present in the fossil record (n=71). Therefore, in modern E. ferus caballus, when the CVT of C6 is absent, then functional ramifications could be of concern when the structural integrity of the caudal cervical module in head/neck function is compromised. These findings might benefit equine practitioners when reporting on diagnostic images of C6, and especially for potential buyers in pre-purchase cases.
5. Conclusions
The ventral process of C6 is a conserved trait in most mammals and its highly specialised atypical structure is present in extinct and extant Equus. Functionally, the muscles attaching to this process aid in flexion, stabilization, fixation, and force redirection of the cervical vertebrae, which are essential for head/neck posture and locomotion. When the CVT of the ventral process is absent, as reported in modern E. ferus caballus, there could be consequences relevant to normal function. This study provided evidence that absent CVTs are not present in extinct Equus, nor in a limited number extant species of Equus, only E. ferus caballus.
Therefore, as an integral part of the caudal cervical module, any anomalous variations in the ventral process of C6 might lead to dysfunctional ramifications of the cervicothoracic junction. This would require further studies, morphologically and biomechanically, to understand full implications.
Author Contributions
Conceptualization, SM-D.; methodology, SM-D and RW.; investigation, SM-D, RW, RH.; resources, SM-D, RW, RH.; data curation, SM-D, RW, RH.; writing—original draft preparation, SM-D.; writing—review and editing, SM-D, RW, RH.; funding acquisition, SM-D.
Funding
This research received no external funding.
Acknowledgments
I Dr. Aisling Farrell - Rancho La Brea Tarpits, Los Angeles, USA; Dr. Thomas Schossleitner – Berlin Natural History Museum, Germany; Dr. Elaine Turner - Monrepus Archaeological Research Centre and Museum for Human Behavioural Evolution, Neuwied Germany; Ms. Amanda Millhouse – Smithsonian Institute, Washington, USA; Mr. Samuel McLeod – Los Angeles County Museum, USA; Dr. Freitson Galis – Naturalis Biodiversity Centre, The Netherlands; Ms. Judith Galkin American Museum of Natural History, New York, USA; Ms. Pamela Blades Ekelberger – Equus Soma, USA; Mr. Andrew Bowe – Australian College of Equine Podiotherapy; Ms. Zefanja Vermeulen – Equine Studies, The Netherlands; Ms. Rachel Narducci – University of Florida.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arnold, P. Evolution of the mammalian neck and development, morpho-functional, and paleontological perspectives. J. Mamm. Evol. 2021, 28, 173–183. [Google Scholar] [CrossRef]
- Arnold, P.; Amson, E.; Fischer, M.S. Differential scaling patterns of vertebrae and the evolution of the neck length in mammals. Evo. 2017, 71, 587–599. [Google Scholar] [CrossRef]
- Buchholtz, E.A.; Bailin, H.G.; Laves, S.A.; Yang, J.T.; Chan, M-Y. ; Drozd, L.E. Fixed cervical count and the origin of the mammalian diaphragm. Evol. Dev. 2012, 5, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Willmore, K.E. The Body Plan Concept and Its Centrality in Evo-Devo. Evo Edu Outreach. 2012, 5, 219–230. [Google Scholar] [CrossRef]
- Flower, W.H. An Introduction to the Osteology of the Mammalia, 3rd ed.; Gadow, H., Ed.; Macmillion: London, United Kingdom, 1885; pp. 12–32. [Google Scholar]
- Chauveau, A. The Comparative Anatomy of the Domesticated Animals., 2nd ed.; Fleming, G. D. Appleton and Company: New York, USA, 1905; pp. 7–167. [Google Scholar]
- Getty, R. Equine Osteology. In The Anatomy of the Domestic Animals, 5th ed,; Sisson, S., Grossman, J. D., Eds.; Saunders: Philadelphia, USA, 1975; pp. 255–348. [Google Scholar]
- Frandson, R.D.; Wilke, W.L.; Fails, A.D. Anatomy and Physiology of Farm Animals. 7th ed,; Wiley-Blackwell: Ames, Iowa, USA, 2009; pp. 59–71. [Google Scholar]
- Janis, C.M. New ideas in ungulate phylogeny and evolution. Trends Ecol. Evol. 1988, 11, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Prothero, D.R. Evolutionary transitions in the fossil record of terrestrial hoofed mammals. Evo. Edu. Outreach. 2009, 289–302. [Google Scholar] [CrossRef]
- Narita, Y.; Kuratani, S. Evolution of the vertebral formulae in mammals: A perspective on developmental constraints. J Exp. Zool. 2005, 304, 91–106. [Google Scholar] [CrossRef]
- Sisson, S. Osteology. In A Textbook of Veterinary Anatomy. W.B. Saunders Company: Philadelphia, USA, 1910; pp. 31–35.
- Bradley, O. The Topical Anatomy of the Head and Neck of the Horse., 2nd ed.; Green and sons: Edinburgh, 1947. [Google Scholar]
- Denoix,J-M.; Pailloux, J-P. Anatomy and Basic Biomechanical Concepts. In Physical Therapy and Massage for the Horse, 2nd ed.; Trafalgar Square Publishing: North Pomfret, USA, 2001: pp. 19–69.
- Bainbridge, D. The Normal Anatomy of the Neck. In Equine Neck and Back Pathology, 2nd ed.; Henson., F.M.D., Ed.; John Wiley and Sons LTD: Hoboken, UK, 2018; pp. 1–9. [Google Scholar]
- Rhombach, N.; Stubbs, N.; Clayton, H. Gross anatomy of the deep perivertebral musculature in horses. J. Amer. Vet. Res. 2014, 75, 433–440. [Google Scholar] [CrossRef]
- Arnold, P.; Esteve-Altava, B.; Fischer, M.S. Musculoskeletal networks reveal topological disparity in mammalian neck evolution. BMC Evol Biol. 2017, 17, 251. [Google Scholar] [CrossRef]
- Davies, Z. Equine Science, 2nd ed,; John and Sons inc., Hoboken, New Jersey USA, 2018, pp 80–135.
- Haussler, K.K. Functional anatomy and clinical biomechanics of the equine cervical spine. Proceedings of the Association of American Equine Practitioners, 360˚ Pain in the Neck Conference. Functional Anatomy and Clinical Biomechanics of the Equine Cervical Soine. Retrieved from https://www.researchgate.net/publication/316053036_. 3160.
- Popesko, P. Atlas of Topographical Anatomy of the Domestic Animals, 4th ed.; W.B. Saunders: Philadelphia, USA, 1985; Volume 4, p. 185. [Google Scholar]
- Ballou, W.R. A Compendium of Equine Anatomy and Physiology. P. Blakiston’s Son and Co: Philadelphia USA, 1907; Replicated by Kessinger Legacy Reprints.
- Ashdown, R.R.; Done, S.H. Color atlas of veterinary anatomy., 2nd ed.; Mosby-Wolfe: London UK, 2000. [Google Scholar]
- Budras, K-D. ; Sack, W.O.; Röck, S. Anatomy of the Horse., 5th ed.; Schlütersche Verlagsgesellschaft & Co: Hannover Germany, 2009. [Google Scholar]
- 19, de Lahunta, A. Veterinary neuroanatomy and clinical neurology., 4th ed.; Elsevier: St. Louis USA, 2015. [Google Scholar]
- Dyce, K.M.; Sack, W.O.; Wensing, C. J. Textbook of veterinary anatomy., 5th ed.; Elsevier: Pennsylvania USA, 2018. [Google Scholar]
- Fails, A.D. Functional anatomy of the equine musculoskeletal system. In Adams and Stashak’s Lameness in Horses., 7th ed.; Ed. Gary Baxter. Blackwell Publishing: Oxford UK, 2020; pp. 1–65. [Google Scholar]
- Hodgins, J.E.; Haskett, T.H. The anatomy and diseases and treatment of domestic animals. The Veterinary Science Company: Detroit USA, 1905.
- Percivall, W. Anatomy, physiology, and pathology of the horse. 1826; retrieved from https://archive.org/.
- Steel, J.H. STEEL, J. H. (1876). Outlines of Equine Anatomy: A Manual for the use of Veterinary Students in the Dissection Room. William Wood and Co: New York USA, 1876. Replicated by Hardpress Publishing.
- Agerholm, J.S.; Bendixen, C.; Andersen, O.; Arnbjerg, J. Complex vertebral malformation in Holstein calves. J. Vet. Diagn. Invest. 2001, 13, 283–289. [Google Scholar] [CrossRef]
- Agerholm, J.S. Complex vertebral malformation Syndrome in Holstein cattle: The Story so Far. Acta. Vet. Scan. 2007, 49, S5. [Google Scholar] [CrossRef]
- Nielsen US, Aamand GP, Andersen O, Bendixen C, Nielsen VH, Agerholm JS: Effects of complex vertebral malformation on fertility traits in Holstein cattle. Livestock Prod. Sci. 2003, 79: 233-238. 10.1016/S0301-6226(02)00170-7.
- May-Davis, S. The occurrence of a congenital malformation in the sixth and seventh cervical vertebrae predominantly observed in thoroughbred horses. J. Equine. Vet. Sci. 2014, 18, 1313–1317. [Google Scholar] [CrossRef]
- DeRouen, A.; Spriet, M.; Aleman, M. Prevalence of anatomical variation of the sixth cervical vertebra and association with vertebral canal stenosis and articular process osteoarthritis in the horse. Vet. Radiol. Ultrasound. 2016, 57, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Santinelli, I.; Beccati, F.; Arcelli, R. Anatomical variation of the spinous and transverse processes in the caudal cervical vertebrae and the first thoracic vertebra in horses. Equine Vet. J. 2016, 48, 45–49. [Google Scholar] [CrossRef]
- Veraa, S.; Bergmann, W.; van den Belt, A-J. ; Wijnberg, I,; Back, W. Ex vivo computed tomographic evaluation of morphology variations in equine cervical vertebrae. Vet. Rad. Ultra. 2016, 0, 1–7. [Google Scholar]
- Veraa, S.; de Graaf, K.; Wijnberg, I.D.; Back, W.; Vernooij, H. ; Nielen, M; Belt, A,J, Caudal Cervical Vertebral Morphological Variation is not Associated with Clinical Signs in Warmblood Horses. Equine Vet. J. 2019, 52, 210–224. [Google Scholar]
- Beccati, F.; Pepe, M.; Santinelli, I.; Gialletti, R.; Di Meo, A.; Romero, J.M. Radiographic findings and anatomical variations of the caudal cervical area in horses with neck pain and ataxia: case–control study on 116 horses. Vet. Rec. 2020, 187, e79. [Google Scholar] [CrossRef]
- May-Davis, S. Congenital Malformations of the 1st Sternal Rib. J. Equine Vet. Sci. 2014, 49, 92–100. [Google Scholar] [CrossRef]
- May-Davis, S.; Walker, C. Variations and Implications of the Gross Morphology in the Longus colli Muscle in Thoroughbred and Thoroughbred Derivative Horses Presenting with a Congenital Malformation of the Sixth and Seventh Cervical Vertebrae. J. Equine Vet. Sci, 2015, 35, 560–568. [Google Scholar] [CrossRef]
- Witzmann, F.; Haridy, Y.; Hilger, A.; Manke, I.; Asbach, P. Rarity of congenital malformation and deformity in the fossil record of vertebrates – A non-human perspective. Int J Paeopatho. 2021, 33, 30–42. [Google Scholar] [CrossRef]
- van der Geer, A.A.E.; Galis, F. High incidence of cervical ribs indicates vulnerable condition in Late Pleistocene woolly rhinoceroses. PeerJ. 2017, e3684. [Google Scholar] [CrossRef] [PubMed]
- Reumer, J.W.F.; ten Broek, C.M.A.; Galis, F. Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths. PeerJ. 2014, 2:318. [CrossRef]
- Varela-Lasheras, I.; Bakker, A.J.; van der Mije, S.D.; Metz, J.A.J.; van Alphen, J.; Galis, F. Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations. EvoDevo, 2011, 2:11.
- Gasse, H., W. Van Den Broeck. and P. Simeons. Nomina Anatomica Veterinaria. 6th ed. Published by the Editorial Committee for the World Association of Veterinary Anatomists. Hanover, Ghent, Columbia MO and Rio de Janeiro. 2017; pp. 17.
- Gidley, J.W. A new three-toed horse. J. Morhol. 1903, 29, 465–476. [Google Scholar]
- Wood, A.R.; Bebej, R.M.; Manz, C.L; Begun, D.L; Gingerich, P.D. Postcranial functional morphology of Hyracotherium (Equidae, Perissodactyla) and locomotion in the earliest horse. J. Mammal. Evol. 2011, 18, 1–32. [Google Scholar] [CrossRef]
- Franzen, J.F.; Habersetzer, J. Complete skeleton of Eurohippus messelensis (Mammalia, Perissodactyla, Equoidea) from the early middle Eocene of Grube Messel (Germany). Palaeobio. Palaeoenv. 2017, 97, 807–832. [Google Scholar] [CrossRef]
- Chauveau, M.D. (1908). The Comparative Anatomy of the Domesticated Animals. 2nd ed. Translation George Fleming. Appleton and Company: New York, USA, 1908; pp. 53–80.
- MacFadden, B.J. Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae. Cambridge University Press: Cambridge UK, 1992.
- Randau, M.; Goswani, A. Morphological modularity in the vertebral column of Felidae (Mammalia, Carnivora). BMC Evol. Biol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, G.; Wagner, G.P. Modularity in Development and Evolution. University of Chicago Press: Chicago, USA, 2004.
- Figueirido, B.; Pérez-Ramos, A.; Martín-Serra, A. Intravertebral vs. intervertebral integration and modularity in the vertebral column of mammalian carnivorans. J. Anat. 2022, 4, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Vidal, P.P.; Graf, W.; Berthoz, A. The orientation of the cervical vertebral column in unrestrained awake animals. Exp Brain Res, 1986, 61, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Graf, W.; de Waele, C.; Vidal, P.; Wang, D.; Evinger, C. The orientation of the cervical vertebral column in unrestrained awake animals. II. Movement strategies. Brain Behav. Evol. 1995, 45, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, C.M.; Wright, L.; Kristoffersen, M.; Puchalski, S.M. Computed tomography and myelography of the equine cervical spine: 180 cases (2013–2018). EVJ. 2020, 9, 475–483. [Google Scholar] [CrossRef]
Figure 1.
The normal and anomalous presentation of the 6th cervical vertebra in modern Equus ferus caballus. (a) and (c) normal presentation; (b) and (d) anomalous presentation of an absent left caudal ventral tubercle. (Not to scale) (Photographs curtesy of Equus Soma, Aiken SC) .
Figure 1.
The normal and anomalous presentation of the 6th cervical vertebra in modern Equus ferus caballus. (a) and (c) normal presentation; (b) and (d) anomalous presentation of an absent left caudal ventral tubercle. (Not to scale) (Photographs curtesy of Equus Soma, Aiken SC) .
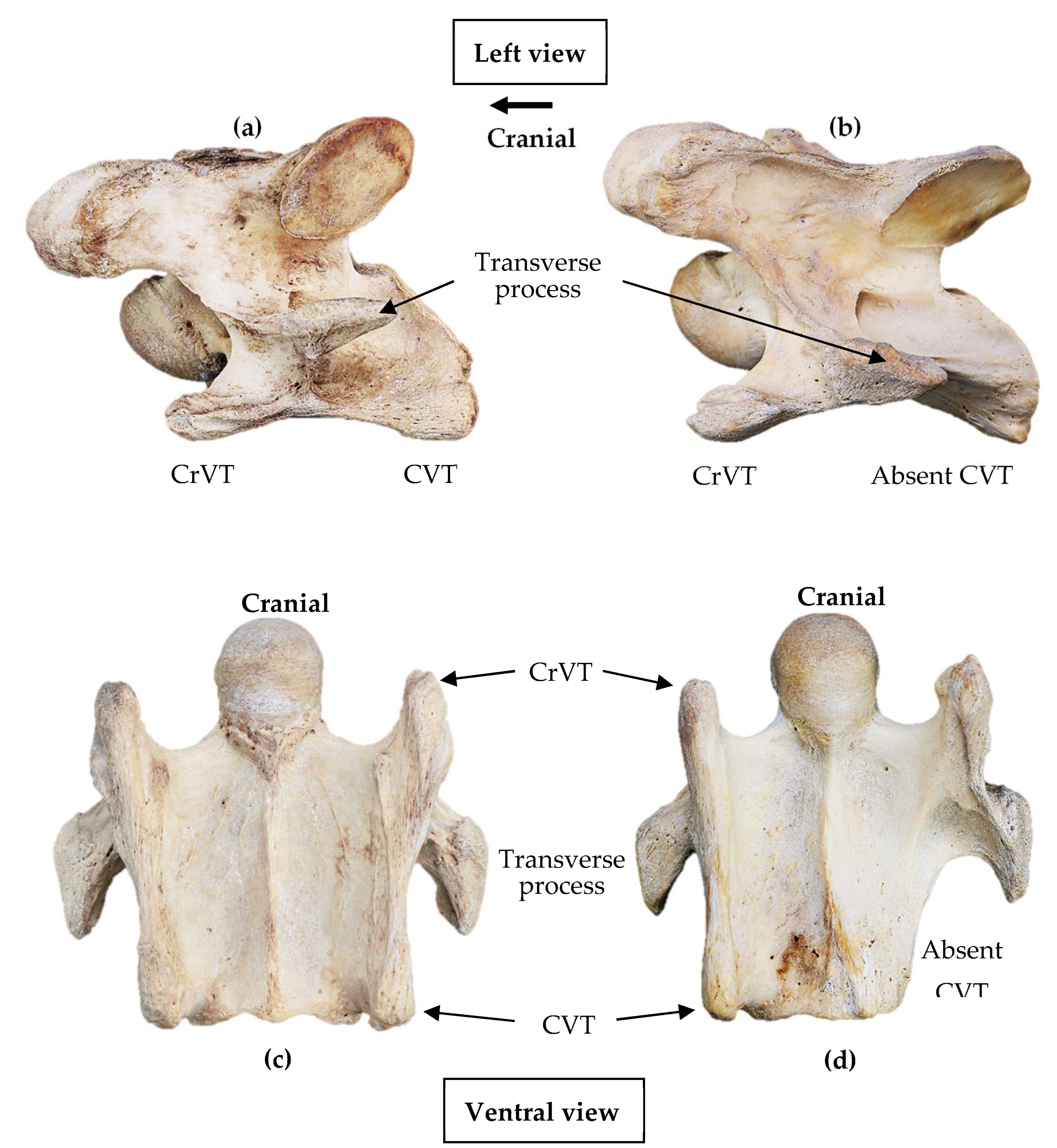
Figure 2.
The left ventral process of C6 in Pliohippus pernix - AMNH catalogue no. 60810 presenting a clearly divided cranial and caudal ventral tubercle. (Not to scale).
Figure 2.
The left ventral process of C6 in Pliohippus pernix - AMNH catalogue no. 60810 presenting a clearly divided cranial and caudal ventral tubercle. (Not to scale).
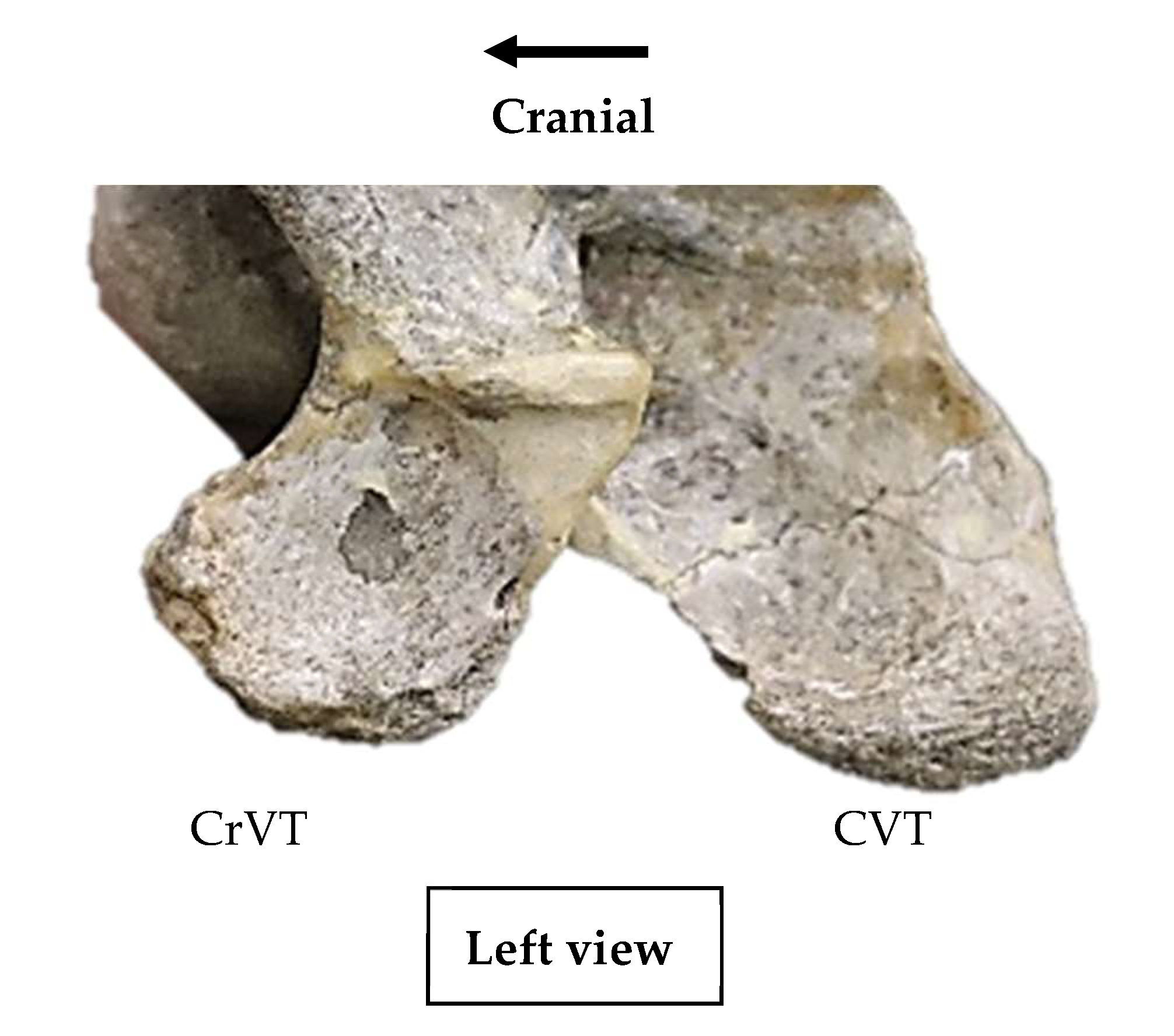
Figure 4.
Ventral view of the 6th cervical vertebra in Equus sp. Leisey 1A B-Y UF 82283 depicting the tube-like formation of the tubercles (orange lineation), widths of cranial ventral tubercle, caudal ventral tubercle, and central constriction (red arrows), and the overall outside width between the left and right cranial ventral tubercules and caudal ventral tubercles (black arrows). (Not to scale).
Figure 4.
Ventral view of the 6th cervical vertebra in Equus sp. Leisey 1A B-Y UF 82283 depicting the tube-like formation of the tubercles (orange lineation), widths of cranial ventral tubercle, caudal ventral tubercle, and central constriction (red arrows), and the overall outside width between the left and right cranial ventral tubercules and caudal ventral tubercles (black arrows). (Not to scale).
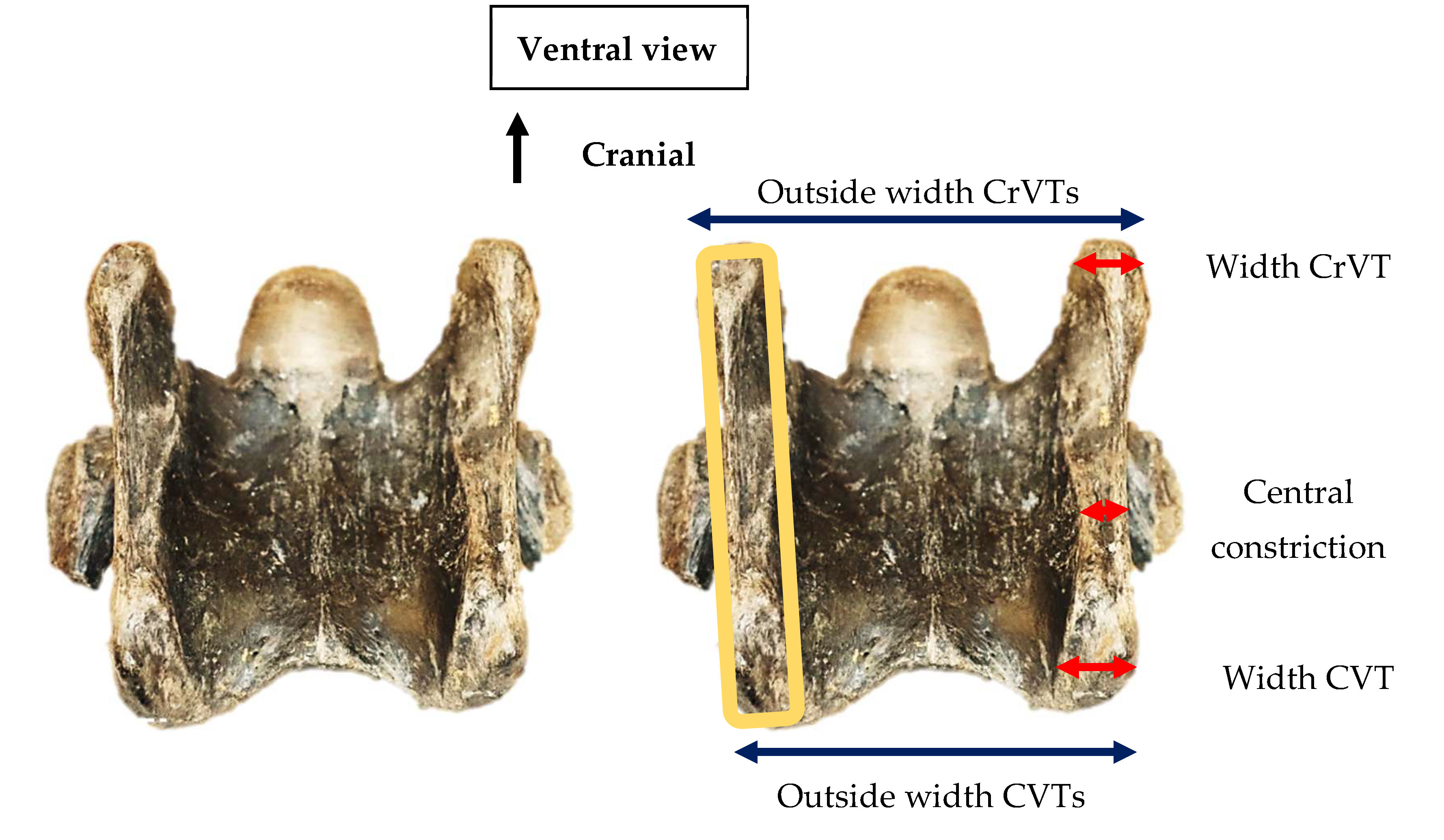
Figure 5.
Left lateral profile of the ventral process of the 6th cervical vertebra in extinct Equus noting the convexity in the morphology between the cranial ventral tubercle and the caudal ventral tubercle (*). (a) Small sized convexity – Equus simplicidens USNM PAL 785556. (b) Medium sized convexity – Leisey 1A UF 151092. (c) Large sized convexity – Pliohippus pernix 60803. (Not to scale). .
Figure 5.
Left lateral profile of the ventral process of the 6th cervical vertebra in extinct Equus noting the convexity in the morphology between the cranial ventral tubercle and the caudal ventral tubercle (*). (a) Small sized convexity – Equus simplicidens USNM PAL 785556. (b) Medium sized convexity – Leisey 1A UF 151092. (c) Large sized convexity – Pliohippus pernix 60803. (Not to scale). .
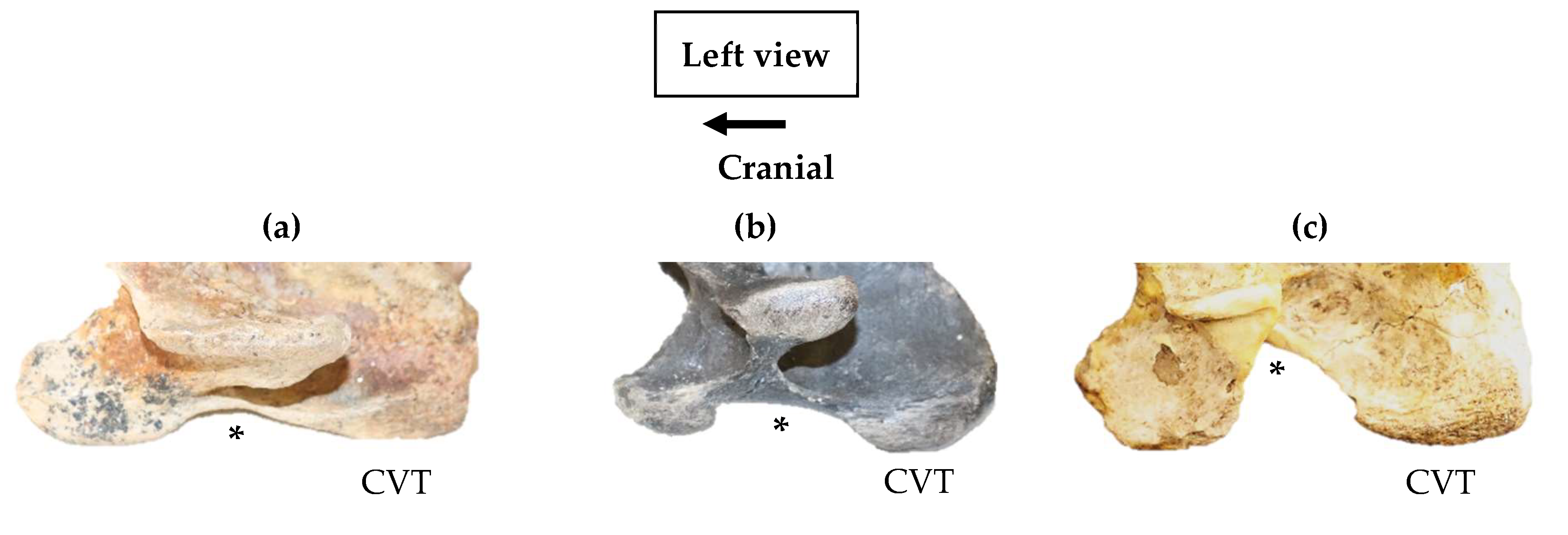
Figure 6.
Degradation and partial loss (black arrow) of the left epiphyseal in the caudal ventral tubercle of the 6th cervical vertebra in extinct and extant Equus. (a) Equus sp. LACM (CIT) 138 120. (b) Equus sp. LACM (CIT) 138 120 outline of the caudal epiphyseal (red dotted line). (c) Partial loss of the epiphyseal in Equine Studies ‘Rideg’ - 3 yr old male Przewalskii’s horse. (Not to scale). .
Figure 6.
Degradation and partial loss (black arrow) of the left epiphyseal in the caudal ventral tubercle of the 6th cervical vertebra in extinct and extant Equus. (a) Equus sp. LACM (CIT) 138 120. (b) Equus sp. LACM (CIT) 138 120 outline of the caudal epiphyseal (red dotted line). (c) Partial loss of the epiphyseal in Equine Studies ‘Rideg’ - 3 yr old male Przewalskii’s horse. (Not to scale). .


Table 1.
Extinct Equus specimens from eight museums and one educational facility.
| Species (n) and Geologic | Institute (n=8) |
Collection/ Catalogue No. | Adult or Sub adult |
|---|---|---|---|
| Equus Sp (4) | NHMB | MB. Ma. 45354 | Adult |
| MB. Ma. 24107 | Sub adult | ||
| MB. Ma. 24410 | Adult | ||
| MB. Ma. 24463 | Adult | ||
|
Equus mosbachensis (4) 80–600kya |
MMHBE | GO 374 3' 8P 197 | Adult |
| Scho 13 II 4 95 690 22 14 | |||
| Scho 13 II 4 95 690 18 45 | |||
| Scho 13 II 4 96 692 27 | |||
| Equus sp 10–40kya (1) | Ko 100-108 | Sub adult | |
| Equus occidentalis (20) 10–40kya | TPMRLB | 2-T22,8'9 | Adult |
| E-2,9 | Sub adult | ||
| 3 E2,16 | Sub adult (left side only) | ||
| 3,E-1,17.5 | Adult | ||
| 3,E-2, 12 | |||
| 3,F4,12 | Sub adult | ||
| 3,F-4,12 | Adult |
||
| 4 B2 M-19 | |||
| 4 B5,20 | |||
| 4 C-2,17-19 | |||
| 4 C-4 13-15 | |||
| 4 D2-4,8,17 | |||
| 4 D2-99 | |||
| 4, C5, 17 246 | Sub adult | ||
| 60 D-12,14-16 | Adult | ||
| 4, G + H-2+3,8 | Sub adult | ||
| 67 H-9,15-18.5 | |||
| 77 E-9,11 | Adult | ||
| Circle 61 | Sub adult | ||
| GIZ, 14.5-16.5 | Adult (right side only) | ||
| Pliohippus (1) | YHM | Mounted replica | Adult |
| Hyracotherium (1) | |||
| Mesohippus (1) | |||
| Hyracotherium (1) 55mya | AMNH | 15428 | Adult (right side only) |
| Merychippus isonesus (1) 15–5mya | ESP. 5263638 | ||
| Mesohippus bairdii (1) 37–32mya | 1477 | Adult | |
| Miohippus doliquideus (1) 32–25mya | 39115 | Adult (left side only) | |
| Pliohippus mirabilis (1) 12–6mya | 60810 | ||
| Pliohippus pernix (1) 12–6mya | 60803 | Adult | |
| Mesohippus (cast) (1) | OUMNH | Mounted Replica | Adult |
| Pliohippus (1) 12–6mya | LACM | (CIT) 210 | Adult |
| Equus Sp (6) | LACM | (CIT) 138 120 | Sub adult |
| (CIT) 138 138 | Adult (left side only) | ||
| (CIT) 138 699 | Adult | ||
| 192 L8 | Adult (right side only) | ||
| LACM San Josecito |
(CIT) 192 | Adult (right side only) | |
| LACM Row XIX |
(CIT) 192 (duplicate) | Adult (right side only) | |
|
Equus simplicidens (15) 5.3–1.8mya |
USNM | USNM 13791 | Adult |
| USNM 12573 | |||
| USNM PAL 785552 | Sub adult | ||
| USNM V 12575 | Adult | ||
| USNM PAL 785561 | |||
| USNM 785560 | |||
| USNM PAL 785553 | |||
| USNM 78559 | |||
| USNM 12155 | |||
| USNM 12580 | |||
| USNM PAL 785557 | |||
| USNM PAL 785556 | |||
| USNM PAL 785559 | |||
| USNM PAL 785555 | |||
| USNM PAL 785554 | |||
| Equus sp. (7) | UF | Leisey 1A UF 82283 | Adult |
| Leisey 1A UF 151092 | |||
| Leisey 1A UF 151020 | Adult (right side only) | ||
| UF 242520 | |||
| UF 242521 | Adult | ||
| Leisey 1A UF 151094 | Adult (Left side only) | ||
| UF 242522 | Adult (right side only) |
Table 2.
Extant Equus specimens from two museums and two educational facilities.
| Species (n) | Institute | Collection/Catalogue No. | Adult or Sub adult |
|---|---|---|---|
| Equus africanus (1) | NBC | N.A.M. 28.-11-1972 |
Adult |
| Equus hermionus (1) | No. 509 | ||
| Equus sp. (1) | Equus sp mounted skeleton | ||
| Equus asinus (3) | ACEPT | Donald | Adult |
| Sebastian | |||
| Kam | |||
| Equus przewalskii (5) | ES | Heteni | |
| Rideg | Sub adult | ||
| YHM | YHM – Live Pony Park | Adult | |
| NHMB | MB. Ma. 16464 | ||
| MB. Ma. 45373 | |||
| Equus quagga boehmi (1) | ES | Kimberley – Zoo bred |
Table 3.
Extinct Equus specimens from three peer reviewed publications.
| Species (n) | Institute | Collection/Catalogue No. | Adult or Sub adult |
|---|---|---|---|
| Neohipparion whitneyi (1) 13.6–4.9mya [46] | AMNH | 9815 | Adult |
| Hyracotherium grangeri (1) 55mya [47] | UM | 115547 | |
| Eurohippus messelensis (1) 56–33.9mya [48] | SMF | SMF ME 11034 |
Key: UM – University of Michigan; SMF – Senckenberg Museum und Forschungsinstitut.
Table 4.
The size of the convexity presented in the lateral profile of the ventral process in the sixth cervical process in extinct Equus (museum specimens and literature) in chronological order, except Equus sp.
Table 4.
The size of the convexity presented in the lateral profile of the ventral process in the sixth cervical process in extinct Equus (museum specimens and literature) in chronological order, except Equus sp.
| Species (n) and Geologic | Institute (n=8) |
Collection Catalogue No. | Size of Convexity |
|---|---|---|---|
| Hyracotherium (1) | YHM | Mounted replica | Large |
| Hyracotherium (1) 55mya | AMNH | 15428 | Large |
| Hyracotherium grangeri (1) 55mya [47] | UM | 115547 | Large |
| Eurohippus messelensis (1) 56–33.9mya [48] | SMF | SMF ME 11034 | * |
| Mesohippus (cast) (1) | OUMNH | Mounted Replica | Large |
| Mesohippus bairdii (1) 37–32mya | AMNH | 1477 | Large |
| Mesohippus (1) | YHM | Mounted replica | Large |
| Miohippus doliquideus (1) 32–25mya | AMNH | 39115 | Large |
| Merychippus isonesus (1) 15–5mya | AMNH | ESP. 5263638 | Large |
| Neohipparion whitneyi (1) 13.6–4.9mya [46] | AMNH | 9815 | Large^ |
| Pliohippus mirabilis (1) 12–6mya | AMNH | 60810 | Large |
| Pliohippus pernix (1) 12–6mya | AMNH | 60803 | Large |
| Pliohippus (1) | YHM | Mounted replica | Large |
| Pliohippus (1) 12–6mya | LACM | (CIT) 210 | Large |
|
Equus simplicidens (15) 5.3–1.8mya |
USNM | USNM 13791 | Small |
| USNM 12573 | Small | ||
| USNM PAL 785552 | Small | ||
| USNM V 12575 | Small | ||
| USNM PAL 785561 | Small | ||
| USNM 785560 | Small | ||
| USNM PAL 785553 | Medium | ||
| USNM 785559 | Medium | ||
| USNM 12155 | Medium | ||
| USNM 12580 | Small | ||
| USNM PAL 785557 | Medium | ||
| USNM PAL 785556 | Small | ||
| USNM PAL 785559 | Small | ||
| USNM PAL 785555 | Medium | ||
| USNM PAL 785554 | Small | ||
| Equus occidentalis (20) 10–40kya | TPMRLB | 2-T22,8'9 | Small |
| E-2,9 | Small | ||
| 3 E2,16 | Small | ||
| 3,E-1,17.5 | Medium | ||
| 3,E-2, 12 | Medium | ||
| 3,F4,12 | Medium | ||
| 3,F-4,12 | Small | ||
| 4 B2 M-19 | Small | ||
| 4 B5,20 | Small | ||
| 4 C-2,17-19 | Small | ||
| 4 C-4 13-15 | Small | ||
| 4 D2-4,8,17 | Small | ||
| 4 D2-99 | Small | ||
| 4, C5, 17 246 | Small | ||
| 60 D-12,14-16 | Small | ||
| 4, G + H-2+3,8 | Small | ||
| 67 H-9,15-18.5 | Small | ||
| 77 E-9,11 | Medium | ||
| Circle 61 | Medium | ||
| GIZ, 14.5-16.5 | Small | ||
|
Equus mosbachensis (4) 80–600kya |
MMHBE | GO 374 3' 8P 197 | Small |
| Scho 13 II 4 95 690 22 14 | Small | ||
| Scho 13 II 4 95 690 18 45 | Small | ||
| Scho 13 II 4 96 692 27 | Small | ||
| Equus sp 10–40kya (1) | Ko 100-108 | Small | |
| Equus Sp (4) | NHMB | MB. Ma. 24107 | Small |
| MB. Ma. 24410 | Medium | ||
| MB. Ma. 24463 | Medium | ||
| MB. Ma. 45354 | Medium | ||
| Equus Sp (6) | LACM | (CIT) 138 120 | Small |
| (CIT) 138 138 | Small | ||
| (CIT) 138 699 | Medium | ||
| 192 L8 | Medium | ||
| LACM San Josecito |
(CIT) 192 | Small | |
| LACM Row XIX |
(CIT) 192 (duplicate) | Small | |
| Equus sp. (7) | UF | Leisey 1A UF 82283 | Small |
| Leisey 1A UF 151092 | Medium | ||
| Leisey 1A 151120 | Medium | ||
| UF 242520 | Small | ||
| UF 242521 | Small | ||
| Leisey 1A UF 151094 | Medium | ||
| UF 242522 | Medium |
Key: * - no convexity; ^ - Gidley’s (1903) description.
Table 5.
The size of the convexity presented in the lateral profile of the ventral process in the sixth cervical process in extant Equus (museum specimens and educational facilities), except E. ferus caballus.
Table 5.
The size of the convexity presented in the lateral profile of the ventral process in the sixth cervical process in extant Equus (museum specimens and educational facilities), except E. ferus caballus.
| Species (n) | Institute | Collection/Catalogue No. | Adult or Sub adult |
|---|---|---|---|
| Equus africanus (1) | NBC | N.A.M. 28.-11-1972 |
Small |
| Equus hermionus (1) | No. 509 | ||
| Equus sp. (1) | Equus sp mounted skeleton | ||
| Equus asinus (3) | ACEPT | Donald | Small |
| Sebastian | |||
| Kam | |||
| Equus przewalskii (7) | ES | Heteni | |
| Rideg | Small | ||
| YHM | YHM – Live Pony Park | Small | |
| NHMB | MB. Ma. 16464 | ||
| MB. Ma. 45373 | |||
| Equus quagga boehmi (1) | ES | Kimberley |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



