
Submitted:
12 April 2023
Posted:
12 April 2023
You are already at the latest version
Abstract
Establishment of mixed-forests has gained increasing attention as a way to optimize forest production, to improve ecological benefits and as a safety net for impacts of future climate uncertainties. However, practical knowledge about which species and what proportion of them should be mixed is still lacking. Thus, this study was conducted with the aim of identify suitable species for mixture with Acacia cincinnata. The mixture tested in the present study was A. cincinnata + Eucalyptus robusta (6:4), A. cincinnata + Acacia mangium (3:1) and monospecifc plantation of A. cincinnata established in 2014. After 7 years of growth, we analyzed the effects of species mixture on growth of tree species, understory vegetation and soil physico-chemical properties as well as bacterial community structure and diversity. The results showed that species mixture had no significant effect on growth characteristics, such as diameter and singletree volume of A. cincinnata. However, mixed-species planting increased the total stocking volume compered to monospecific plantation of A. cincinnata. Furthermore, stand mixture significantly increased species diversity, biomass and nutrient stocks in the understory vegetation. The soil of mixed stand of A. cincinnata and A. mangium had the highest C and N contents, whereas the soil of pure A. cincinnata stand had the highest P content. The diversity of soil bacterial community were the highest in the mixed stand of A. cincinnata and E- robusta, followed by pure A. cincinnata stand and A. cincinnata + A. mangium stand. The relative abundance of Proteobacteria and Actinobacteria was higher in soils of mixed stands. Furthermore, the relative abundance of Firmicutes was high in the soil of A. cincinnata + A. mangium while the relative abundance of Verrucomicrobia was high in A. cincinnata + E. robusta stand. As a whole, the study demonstrated that establishing mixed-species plantation enhance the diversity and composition of understory vegetation, soil physico-chemical and soils bacterial community; thereby increasing biodiversity, nutrient cycling and carbon sequestration in the biomass and soil. From the viewpoints of forest productivity and ecological benefits, it is advisable to establish a mixed plantation of A. cincinnata and A- mangium in southern China. As a whole, our work revealed that the sustainability of mixed-species plantation relies on the interactions between soil attributes, vegetation, and bacterial community.
Keywords:
Acidobacterium
; Acacia mangium
; Eucalyptus robusta
; mixed-species forest
; soil biological property
; soil nutrients
1. Introduction
During rapid economic and social development, large-scale natural forest harvesting and conversion of natural forests into plantations commonly occur, resulting in forest degradation, soil erosion, and other forest and environmental problems [1]. To circumvent land degradation, the establishment of ecological welfare forestland was initiated in China, which is conducive to improving the ecological environment and the ecosystem services and functions of forests, which is of great significance to the restoration of fragile ecosystems and expansion of forest plantations [2,3]. Since China implemented classification-based forest management, a number of plantations with highly important ecological locations or extremely fragile ecological conditions has been established as ecological welfare forests, including water conservation forests, soil and water conservation forests, windbreak and sand dune fixation forests, and shore restoration forests [4]. These ecological welfare forests play an important role in territorial ecological security, biodiversity conservation, and sustainable economic and social development [5]. However, the quality of some ecological welfare forests in China is generally poor, and they have become low-yielding and inefficient with poor ecosystem stability and ecological functions, making it difficult to give full play to the ecological protection function of forests [6].
The practice of artificial afforestation shows that continuous growing of monoculture forests on the same land results in decline of soil fertility, reduction in forest stand productivity and the reduction of forest ecological functions [7]. Thus, mixed-species forest is opined to be a good model for afforestation and forest stand quality improvement in today's ecological welfare forest. Several studies have shown the advantages with mixed-species forests over monoculture forests. For instance, Maire et al. [8] analyzed the light energy utilization efficiency of pure and mixed forests of Eucalyptus grandis Hill ex Maiden and Acacia mangium Willd, and found that the mixed forest canopy was more favorable to the growth of plants in the stands that use sunlight to provide nutrients for growth. Forrester et al. [9] established 1:1 stand of E. robusta and Acacia mearnsii De Wilde and found that mixed stand improved stand water use efficiency, canopy photosynthetic capacity, and aboveground and underground carbon distribution. Liao et al. [10] found that a mixed forest of Eucalyptus grandis x urophylla and A. mangium was conducive to forest growth in a relatively barren red-soil mountainous area, and its ecological benefits, wind resistance, and disease resistance, were better than those of a pure forest was. Chen et al. [11] found that the mixture of E. urophylla and A. mangium can effectively increase the soil nutrient content and that the nutrient content of litter in mixed forests exceed that of pure forests. The study by Zang et al. [12] showed that the number and diversity of micro-organisms in the mixed forest of Acacia crassicarpa Benth and E. robusta were greater than those in the pure forest, and afforestation of mixed A. crassicarpa and E. robusta was beneficial to improve soil fertility and vegetation diversity.
Acacia cincinnata F. Muell is an evergreen tree of the legume family, native to the northeastern coastal region of Australia, introduced to China in the 1970s, and widely planted in Guangdong, Fujian, Hainan Provinces [13]. A. cincinnata has the advantages of fast growth, strong nitrogen fixing ability, resistance to drought and barrenness, wide timber use, and short rotation of 6 - 8 years. Therefore, it has been widely used for montane afforestation, soil and water conservation afforestation, and coastal conservation afforestation in southern Fujian, China [14]. Owing to its production of a large leaf litter, A. cincinnata has a good effect on improving soil fertility. It can also be used in crop rotation planting and suitable for establishment of mixed forest with E. robusta in southern Fujian to effectively improve the consumption of soil nutrients, fertilizer, and water on forestland due to the rapid growth of eucalyptus trees [15]. At the same time, A. cincinnata is also an excellent choice for establishment of mixed -forests with acacia species such as A. mangium, to effectively improve the stability and diversity of the forest stand structure [16]. Due to the long-term focus on the expansion of afforestation areas of A. cincinnata and insufficient attention to the improvement of stand management practices, especially neglecting stand-tending operation, many A. cincinnata forests in southern Fujian have become low-yielding, with poor ecosystem stability and ecological functions [17]. It is, therefore, difficult to truly achieve the multi-functional benefits of ecological welfare forests in maintaining water conservation, soil fertility, biodiversity conservation and recreational landscape effects [18].
Therefore, the establishment of A. cincinnata plantations mixed with E. robusta and A. mangium may be conducive to improving the soil fertility and water conservation capacity of barren land, which is of great significance for improving the ecological and economic benefits of forestland in southern China. However, the effects of these mixtures on growth of tree species, undergrowth vegetation diversity, soil physico-chemical and biological properties of A. cincinnata plantation are largely unexplored. Thus, the aims of this study were to examine whether mixture of A. cincinnata with A. mangium or E. robusta will have an impact on growth of tree species, if so which species, A. mangium or E. robusta has the largest impact; and to evaluate the effects of mixed species planting on understory vegetation, soil and bacterial community structure and diversity. The soil bacterial community was explored by sequencing of the 16S rRNA gene in soils from different forest stand types. Our research study endeavored to answer four main questions: (i) Does mixed-species planting improve growth of tree species? (ii) Does mixed-species planting increased the diversity and biomass of the understory vegetation as well as C, N and P concentrations in the soil, (iii) Is the bacterial community of pure and mixed-species stands different due to their higher nutrient inputs (litter fall and biomass) in the mixed stands? (iv) Is there any link between bacterial community and soil attributes (C, N, P, C:N, C:P and N:P ratios? The study will provide an insight into the scientific management of A. cincinnata mixed forest to meet a range of economic and ecological benefits in southern China.
2. Materials and Methods
2.1. Study site
The study was conducted in Daqiutian Wutai Mountain Forest Farm, Nan'an county, Quanzhou, Fujian, China (118°24′20″N, 25°16′45″E), which is located at an altitude of 80~1000 m. The area has a subtropical monsoon climate with an annual average temperature of 19.5 to 21.0 °C with a frost-free period of 350 d. The soil is red soil developed from sandstone shale with low soil fertility. The plantations were established in 2014, and the stand density was 1110 trees hm-2 of E. robusta (40%; 444 trees hm-2) and A. cincinnata (60%; 666 trees hm-2) forest, 1850 trees hm-2 of A. mangium (25%; 463 trees hm-2) and A. cincinnata (75%; 1387 trees hm-2) forest and 1230 trees hm-2 of A. cincinnata forest (100%), with a total area of about 6.13 hm2, 7.6 hm2, and 7.06 hm2, respectively.
2.2. Measurement of growth traits
In January 2021 (7 years after planting), three 20 × 20 m standard plots were set- up in each forest stand type. Thereafter, the diameter at breast height (DBH) and tree height of each tree were measured in the standard plot and the average volume and stand volume of each plot were calculated. The DBH of trees was divided into DBH class of 2 cm interval [19,20]. The number of tree in each diameter class was counted, and the average DBH of trees in each diameter class was calculated separately for each forest stand as follows:
Here, N is the total number of trees in the i-th diameter class, and di is the DBH of the i-th diameter class.
To improve the accuracy of the tree height measurements, the measured mean DBH was substituted into the growth model and then we fitted the tree height as follows:
where y and x is the fitted height and DBH, respectively
yA. Cincinnata = 3.2111x0.5254
yA. Mangium = 4.3692x0.4247
yE. Robusta = 2.598x0.6896
The volume per tree was calculated as follows:
For Acacia, V = 0.00005276×D1.882161×H1.009317
For E. robusta, V = 0.000109154×D(1.87892-0.00569186×(D+H))×H(0.652598+0.00784754×(D+H))
Where D is the average DBH, H is the fitted tree height.
To calculate the volume of each diameter class, the average volume per tree of each diameter class was multiplied by the number of trees in the diameter class. To calculate the stand volume, the volume of each diameter class was added up in the sample plot.
2.3. Biomass and nutrient concentrations of understory vegetation
The species richness of the understory vegetation was determined by conducting an inventory in three 1 m × 1 m plots in each forest stand type. All species were identified in situ using plant identification manual and its cover was determined. The aboveground biomass of understory vegetation (herbaceous plants and shrubs) was determined by harvesting all individuals in three 1m×1 m plots that were set-up in each forest stand. The underground (root) biomass was determined by digging each plant roots. The fresh biomass samples were weighed in the field and then the samples brought back to the laboratory where they were oven-dried at 105 °C to a constant mass for the determination of dry mass. After crushing the samples and passing through 0.15 mm sieve to remove impurities, the carbon (C) and nitrogen (N) concentrations were determined by Vario Max Elemental Carbon and Nitrogen Analyzer (Elementar, Germany), and the Molybdenum-antimony colorimetry method was used to determine the total phosphorus concentration in the biomass of understory vegetation.
2.4. Soil analysis
Soil samples were collected from tree plots in each stand at different soil depth: 0-10 cm, 10-20 cm and 20-40 cm. The soil physical properties, including soils water content, bulk density, water holding capacity and non-capillary and capillary porosity, were determined using the ring knife method, keeping the soil structure intact. The chemical properties were measured using air-dried soil (sieved at 0.15 mm). Carbon and nitrogen contents were measured with an elemental analyzer (Vario MAX CNS, Elementar, Hanau, Germany) while P was determined after digestion by HF, HClO4, and HNO3 using the molybdenum-blue method.
2.5. Extraction and Amplification of Soil Bacterial DNA for Sequencing
The DNA in soil samples was extracted using the CTAB method, and the quality of the extracted DNA was checked using electrophoresis in agarose gels (1% w/v in TAE buffer). After electrophoresed for 30 min, the samples were thawed on ice, centrifuged and thoroughly mixed. The quality of the samples was detected by Nanodrop, and 30 ng sample were taken for PCR amplification. PCR products were purified using the Agencourt AMPure XP Nucleic Acid Purification Kit. The V3–V4 hypervariable regions of the 16S rDNA of the bacteria were amplified. The primer sequences were ACTCCTACGGGAGGCAGCAG and GGACTACHVGGGTWTCTAAT. The amplification was conducted under the following reaction conditions: 5 min of initial denaturation at 94 °C, 30 s of denaturation at 94 °C, 30 s of annealing at 55 °C, and 28 cycles of 1 min of elongation at 72 °C. The amplified PCR products were sequenced on an Illumina MiSeq (PE300) sequencing platform. The original sequencing data were deposited in the NCBI SRA database under the accession number PRJNA954280. The high-quality Clean Tags sequences were obtained after quality control filtering, and the Clean Tags sequences were clustered to produce operational taxonomic units (OTUs) based on 97% sequence similarity. The UNITE taxonomic database and the RDP Classifier database were used for species annotation of OUT representative sequences and analysis of bacterial community composition at each taxonomic level to obtain information on bacterial species in soils of different forest stand types.
2.6. Statistical analysis
The obtained OUT clustering results and species information were analyzed using Mothur software, and the abundance index of Chao 1 and Observed species, phylogenetic diversity (PD_whole_tree) and Shannon’s diversity index were calculated. One-way analysis of variance (ANOVA) was conducted to examine significant differences in growth traits of A. cincinnata, biomass of understory vegetation, and bacterial diversity among the different forest stand types while two-way ANOVA was conducted to determine significant differences in soil physico-chemical properties among different soil depths and stand types. Means that exhibited significant differences were compared using Tukey’s honestly significant test using SPSS software (version 22, IBM, New York, USA). The relationship between soil C, N and P content and the relative abundance of the dominant bacterial phyla was analyzed by redundancy analysis using Canoco software (version 5). All data were expressed as the mean ± standard error (SE).
3. Results
3.1. Growth of A. cincinnata in mixed and pure stands
Growth characters of tree species showed significant differences among forest stand types (Table 1). Diameter, height and individual tree volume of A. cincinnata did not differ between mixed-species planting and monospecific planting. Diameter and volume of individual A. mangium and E. robusta trees were higher than A. cincinnata in mixed-species plantings. Whereas height of E. robusta was higher than A. cincinnata and A. mangium Compared with the pure stand of Acacia cincinnata, mixed management was conducive to the increase of stand volume as evidenced in significantly (p < 0.05) higher stand volume of mixed stand of A. cincinnata and A. mangium than that of pure stand of A. cincinnata. The total volume of the standing stock was higher in A. cincinnata + A. mangium stand than the monospecific stand of A. cincinnata. Even though it was not significant, the total volume of A. cincinnata + E. robusta stand was relatively higher than that of monospecific stand.
3.2. Biomass and nutrient concentrations of understory vegetation
Diversity of understory vegetation differed among forest stand types; particularly species richness was higher in A. cincinnata + E. robusta stand than the other stands (Appendix A). Biomass of understory vegetation varied significantly (p < 0.05) among stand types, where both above- and belowground herbaceous biomass was higher in A. cincinnata and mixed stand of A. cincinnata and A. mangium than mixed stand of A. cincinnata and E. robusta (Table 2). Similarly, biomass of roots, branches and leaves of shrub species in the understory varied significantly (p < 0.05) among different forest stand types (Table 2). Mixed stand of A. cincinnata and A. mangium resulted in higher biomass of roots and branches and leaves of shrub species than mixed stand of A. cincinnata and E. robusta but similar with A. cincinnata stand.
Nutrient concentration of different parts of the understory vegetation also varied among stand types (Table 3). N concentration in the aboveground parts of herbaceous species was higher in the pure stand of A. cincinnata than mixed stands while it was the opposite in the underground parts. C and P concentrations in the aboveground parts of herbaceous species did not vary significantly among stand types. The C concentration of underground parts was higher in mixed stands than A. cincinnata stand while the P concentration of the underground part was higher in pure A. cincinnata stand than mixed stands. Among understory shrub species, the concentrations of N and P in the roots were the highest in the mixed stand of the two acacias. The concentration of P in the branch was slightly higher in the pure than mixed stands. The concentrations of N and C in the leaf was the highest in mixed than pure stands.
3.3. Soil physical properties and C:N:P stoichiometry
Soil physical properties differed significantly (p < 0.05) among soil layers and the different stand types (Figure 1). In mixed stand of A. cincinnata and E. robusta, the soil water content was high in 20-40 cm soil layer while it was significantly low in 10-20 cm soil layer in mixed stand of the two acacias compared to monospecific planting of A. cincinnata and mixed stand of A. cincinnata a and E. robusta. The bulk density in 0-10 cm soil layer was significantly higher in a mixed stand of A. cincinnata + A. mangium than the pure stand of A. cincinnata. Of the three stand types, the maximum water holding capacity in 0 - 10 cm soil layer was the highest in the mixed stand of A. cincinnata and A. mangium. In the 0-10 cm soil layer, the minimum water-holding capacity of the three forest stands was in the order pure stand of A. cincinnata < mixed stand of A. cincinnata + E. robusta < mixed stand of A. cincinnata + A. mangium. The maximum water holding capacity, capillary water holding capacity, minimum water holding capacity, capillary porosity, and total porosity of 20-40 cm soil layer were all improved in mixed stands compared to pure stand of A. cincinnata. As a whole, mixture of the two acacias (A. cincinnata + A. mangium) resulted in improved soil physical properties more than mixed stand of Acacia cincinnata and E. robusta.
Analysis of soil stoichiometry revealed significant differences (p < 0.05) in soil C, N and P concentrations as well as C:N, C:P and N:P ratios in different soil layer and among different forest stands (Figure 2). The concentration of C decreased along the soil depth in all stand types; especially in mixed stands whereas it remained the same among stands within each soil layer. In mixed stands, the concentration of N was higher in the 0-10 cm soil layer than the other soil layers while in pure stand of A. cincinnata the N concentration was higher in 10-20 cm soil layer than the other soil layers. The P concentration was higher in monospecific than mixed-species stands across the soil depth. The C:N ratio was significantly higher in mixed stand of A. cincinnata and A. mangium than in mixed stand of A. cincinnata and E. robusta and monospecific stand. The C:P ratio was higher in the 10-20 cm soil layer than the other soil layers in mixed stands while it remained statistically similar in the pure stand of A. cincinnata. Similarly, the N:P ratio was higher in the upper than in the lower soil layer in mixed stands while it was the same across the soil depth in the pure stand of A. cincinnata.
3.4. Soil bacterial diversity
With 97% similarity, the number of OTU was obtained for each stand, and the total number of OTU for A. Cincinnata + A mangium, A. cincinnata + E. robusta and A. Cincinnata stands were 1493, 1830, and 1737, respectively. There were 1,149 OTUs in all acacia stands, of which 158, 89, and 70 OTU were unique to A. cincinnata + A. mangium , A. cincinnata + E. robusta, and A. cincinnata stands (Figure 3). The richness of soil bacteria community, as measured by Chao1 and Observed_ species, showed significant differences in response to species mixture. Both Chao1 and Observed_ species indices were higher in mixed stand of A. cincinnata and E. robusta and pure stand of A. cincinnata than in mixed stand of A. cincinnata and A. mangium (Table 4). The phylogenetic diversity (PD_whole_tree) showed similar pattern as Observed- species index, whereas species diversity (Shannon index) remained similar among the different stand types.
3.5. Composition of Soil Bacteria Community
The composition of bacterial community differed among the different stand types. At the phylum level, Acidobacterium, Chloroflexi, Proteobacteria and Actinobacteria were the most abundant bacterial among the top 11 phyla (Figure 4). The relative abundance of Acidobacterium was relatively high in the soil, followed by the relative abundance of Chloroflexi and the relative abundance of Proteobacteria; together accounting for about 80% of the phylum composition in pure A. cincinnata stand and about 78% in mixed- species stand. The relative abundance of Proteobacteria and Actinobacteria was higher in soils of mixed stands than in pure A. cincinnata stand. While the relative abundance of Firmicutes in the soil of A. cincinnata + A. mangium stand was higher than that of the other two forest types, and the relative abundance of Verrucomicrobia was higher in A. cincinnata + E. robusta stand.
3.6. Relationship between soil bacteria and C, N and P stoichiometry
To further explore what shapes bacterial community composition, redundancy analysis was performed using soil stoichiometry as explanatory variables with the first axis explained 78% and the second axis explained 22% of the variation (Figure 5). Actinomycetes and Plactomycetes were positively correlated with Total N and C:P but negatively correlated with C:N. Acidobacteria was positively related with N:P but negatively related with Total C and Total P. Chloroflexi and Gemmatimonadetes were positively correlated with Total C and Total P but negatively correlated with Total N, C:P and N:P. Verrucomicrobia and Proteobacteria were positively correlated with C:N but negatively correlated with Total N.
4. Discussion
4.1. Effects of species mixture on the growth of Acacia cincinnata
Mixed-species planting did not improve the growth of individual A. cincinnata trees in relation to its monoculture while improved the growth of accompanying species, A. mangium and E. robusta. However, the introduction of E. robusta or A. mangium in the A. cincinnata plantation significantly improved stand volume of A. cincinnata trees. Mixed-species management is currently an important afforestation method to improve productivity and the ecological benefits of plantations, as well as to improve biodiversity [21). Reasonable mixing of species can promote stand growth due to improvement in physico-chemical properties of the forest soil, such that the soil is looser with good aeration and improved nutrient availability that promotes the extension of tree roots in the soil layer [22]. In mixed-species planting management, tree growth depends on the characteristics of the species itself and interspecific belowground interaction. Interspecific belowground interactions can be competition or facilitation, where the latter represents that one species positively modifies the environment of the other species; thereby benefiting its growth [23]. For instance, N2-fixing species, such as A. cincinnata and A. mangium, can meet its nitrogen requirement through fixing atmospheric N in their roots [24], while supplying N for accompanying species in mixed planting management [25]. Short-term belowground N transfer from A. mangium, N2 fixing tree species, to Eucalyptus trees has been observed in a mixed-species plantation [26]. Likewise, it was found that non-N2-fixing trees utilize atmospheric N fixed by the N2-fixing trees, when grown together, presumably through constricted root connections or organic forms of N from the litter layer. In addition, studies have shown that competition between tree species, when grown in mixed-species model, can lead to differentiation in horizontal or vertical root distribution [27,28]. Thus, mixing tree species with contrasting rooting traits can result in a more effective exploration of the soil by the roots, while spatial and temporal niche separation in the soil can result in increased root biomass [29]. One example could be that A. cincinnata and E. robusta, when grown together, could increase the tree height and diameter growth as well as stand volume (note that the stem density is half of that of monospecific plantation of A. cincinnata) due to increased complementarity by spatial niche partitioning. This is because E. robusta has deep root system, which can avoid competition in the shallow root zone with A. cincinnata while the latter species could provide N to the former species. This has been observed in significantly higher total stock of mixed forest of E. robusta + A. cincinnata (84.6 m3/hm2) than that of pure A. cincinnata (61.1 m3/hm2) stand as well as in a mixed-species plantation of E. globulus and A. decurrens in Ethiopia [Error! Reference source not found.]. Similarly, in a mixed plantation of Gmelina arborea and Cordia alliodora, it was observed that G. arborea accounted for 40% of the total stand volume compared to 21% by C. alliodora [30].
Mixed-species did not improve the diameter growth of individual A. cincinnata tree in relation to its monospecific planting and accompanying tree species. This result is similar with previous studies that demonstrated lack of significant differences in diameter growth of E. urograndis and A. mearnsii in different arrangements of monospecifc and mixed-species planting at the age of 18 months [31]. In a study made to examine the growth of E. camaldulensis and Leucaena leucocephala in monospecifc and mixed-species planting systems, the growth in DBH did not show significant difference between the different planting systems, which indicates that there is a lack of positive or negative interspecifc interactions that favored the growth in DBH of both species in different planting arrangements [32]. Similarly, the volume growth of individual A. cincinnata tree in monoculture and mixed-species plantations did not display significant differences between the different plantation systems, but significantly higher volume of accompanying species was observed. Similar results have been observed in mixed-species plantations of E. globulus and A. mearnsii [33] and in monoculture and mixed-species plantation of E. saligna and A. mearnsii [34].
4.2. Effects of species mixture on composition and diversity of understory vegetation
Mixed-species planting; particularly A. cincinnata + E. robusta, resulted in higher diversity (species richness) while mixed species planting of A. cincinnata and A. mangium resulted in higher biomass of understory vegetation. Several studies showed that tree species mixture increases species richness and biomass of the understory vegetation [35,36]. Each tree species used in mixed-species forests has an explicit impact on the understory environment, which in turn is associated with its influences on resource availability (light, water and soil nutrients) and other environmental factors, such as microclimate, precipitation throughfall and competition [37,38,39]. It has often been opined that the composition and diversity of understory vegetation in mixed forests lies in between species composition and diversity of individual monoculture communities [40]. In other words, each tree species occurring in mixed-species forest contributes all or some of the species associated with it. In our study, five species occurring in the understory of monospecific plantation of A. cincinnata occurred in mixed planting of A. cincinnata and E. robusta. The effect of species admixture is usually large when the tree species involved in mixed-species plantation change to a greater extent the understory environment, such as light availability, soil nutrient content, litter and root turnover [41]. Furthermore, the stand density regulates species richness of the understory vegetation; i.e. increasing stand density will lead to low species richness [42]. This is further evidenced in our study where low stand density of A. cincinnata + E. robusta resulted in higher species richness in the understory vegetation than the other forest stand types. However, the biomass of understory vegetation and the N content of the roots of herbaceous species as well as shrubs were higher in mixed stand of A, cincinnata and A. mangium. This could be related with increased N status in the soils of this stand through atmospheric N fixation by both Acacia species. Mixed-species planting, especially A. cincinnata and A, mangium increased the C content of the underground part of herbaceous species and the leaves of understory shrubs; suggesting better growth of the understory vegetation under mixed-species stand, as a mixed forest plantation contains more varied microhabitats than the respective monocultures do [43].
4.3. Effects of species mixture on soil physical properties and C:N:P stoichiometry
As an indispensable resource affecting the growth and development of plant communities in terrestrial ecosystems, soil provides nutrients and water for plants, and its physicochemical properties directly affect the growth of plants [Error! Reference source not found.]. The soil physicochemical properties are the main drivers of changes in the structure and spatial distribution of soil microbial communities, which play a key role in ensuring the normal growth and development of plants [Error! Reference source not found.] as well as in regulating soil nutrients and ecosystem processes. Owing to the different biological characteristics of various tree species, different effects of species mixture on soil physico-chemical properties are observed [44]. In this study, soil physical properties exhibited significant differences along the soil layer among different forest types. The average soil water content was high in 20-40 cm soil layer in mixed stand of A. cincinnata and E. robusta, while it was significantly low in 10-20 cm soil layer in mixed stand of the two acacias compared to monospecific planting of A. cincinnata. Generally soil water content is dependent on the interactions between precipitation, soil evapotranspiration and surface cover [45] With increasing soil depth, evaporation decreases whereas water infiltration increases, thus resulting in a higher soil water content at a lower depth [46]. In addition, Eucalyptus trees, with their deep root system, can draw more water in the deeper soil layer than the upper soil layer. Mixed forests significantly reduced the bulk density of soil due to the interspersion of plant roots that make the soil looser and improves soil aeration [47]. Mixed-species plantations significantly improved soil aeration and water holding capacity as well as the soil porosity (i.e., non-capillary and capillary porosity, and total porosity) and water holding capacity, capillary water holding capacity, maximum water holding capacity, and minimum water holding capacity of topsoil and subsoil compared to monoculture Chinese fir plantations [44] – a result similar to our study. Similarly, a decrease in soil bulk density, but an increase in water holding capacity was observed in mixed-species forest during restoration of sodic lands [48]. This can be attributed to the development of root network and increased leaf litter production in mixed-species forests [48].
The present study also demonstrated that the C and N concentrations tended to be higher in mixed-species than in monospecific plantations, while the opposite pattern was observed in the P concentration. Mixed-species increases the quality and quantity of litter in the understory, enhances the biological activity of microorganisms, and increases the concentration of organic matter [49]. The current soil C content is the result of the balance between C input and output processes, which are chiefly derived from litter fall, soil microorganisms, and soil-animal and plant-root turnover [50,51]. Both A. mangium and E. robusta had substantial litter fall input, resulting in a higher C input into the soil of mixed stands compared with pure stand of A. cincinnata. Studies have shown that litter decomposition is dependent on micro-environmental conditions of the forest understory and abundance and diversity of earthworms [52], which might be more conducive in the mixed stand than monospecific stand in our case. Soil C:N ratio reflects nitrogen mineralization capacity, and a lower value improves increased N available and soil fertility [53]. In this study, mixed stand of A. cincinnata and E. robusta and monospecific planting of A. cincinnata significantly decreased soil C:N ratio in both top and sub soil compared with mixed stand of A. cincinnata and A. mangium, indicating that the effect of mixed-forests on soil nutrient status is specifies-specific. When both N2-fixing species, A. cincinnata and A. mangium, are grown together, the N concentration in the soil will be higher than when grown together with non-N2-fixing species, such as E. robusta, resulting in lower C:N ratio. Similar result was reported where soils of coniferous mixed forests have lower C.N ratio than broad-leaved mixed forest [54].
With regard to soil N and P content, mixed stands presented higher N content in the topsoil while the P concentration was higher in monospecific than mixed-species stands across the soil depth. Soil N is mainly derived from the decomposition of organic matter (mainly litter turnover) as well as atmospheric N fixation; as a result, higher total N concentrations were observed in mixed stand of both acacia species, where larger biomass of herbaceous species and N content in leaves of understory shrub species were also recorded. It should be noted that turnover of herbaceous flora is the major source of organic matter and the composition and diversity of soil bacterial and fungal communities are controlled by the contents of nitrogen, cellulose and lignin in leaf litter of standing vegetation [48]. On the other hand, the P content in the soil is strongly influenced by the mineralization of soil microbiome, which allows more P to be stored in the soil [53]. The fact that higher total P was found under monospecific plantation across all soil depths in the present study suggest that mixed-species plantations are not conducive for P accumulation in the soil. The C:P ratio reflects the efficiency of the plant's use of P [55], and N:P ratio represents the plant's nutrient limitation status [56]. We found higher C:P ratio in the subsoil of mixed stands while higher N:P ratio in the topsoil of mixed stands. This can be attributed to generally low availability of P in the soils of the study area itself [57]; and hence the P content is small compared to carbon and nitrogen contents.
4.4. Effect of species mixture on abundance and diversity of soil bacterial community
Mixed forests improve soil bacterial structure and increase bacterial diversity compared to pure forests due to differences in the degree of soil decomposition of understory litter, the soil water content, C, N, P contents, and other soil physico-chemical properties [58,59]. In the present study, mixed-species planting, especially A. cincinnata + E. robusta, increased the richness (Chao1 and Observed number of species) and phylogenetic diversity compared with mixed stand of A. cincinnata and A. mangium. Despite the highest N content in A, cincinnata + A. mangium plantation, the soil bacterial richness and diversity were the lowest compared with A. cincinnata + E. robusta. This suggest that tree species have a significant effect on community structure than the soil environment [60]. A similar result has reported from mixed-species plating of A. mangium and Eucalyptus in Congo [61]. Addition of N in a subtropical deciduous oak mixed forest in China resulted in a significant decline in bacterial richness and diversity [62,63]. It was suggested that the loss of one or more bacterial species does not dramatically affect the ecosystem functions, probably due to the high functional redundancy of soil microbes [61]. The fact that A, cincinnata + E. robusta had the largest number of soil bacteria, and the observed species index, and PD_whole_tree index could be linked to its high soil moisture content, which improved the soil environment and enriched the soil bacterial community structure [64]. The metabolic processes of different plant roots, as well as the decomposition of soil litter, will affect soil permeability, porosity, compactness, and other soil attributes, such as C, N, and P concentrations in the soil, which will then affect soil bacterial community structure [65]. Our result is consistent with the study by Zang et al. [12] who demonstrated increased number and diversity of microorganisms in the mixed forest of A. crassicarpa and E. robusta compared with the pure forest.
Soil microbes regulate major biogeochemical cycles and can significantly affect ecosystem functions [66]; hence, the soil environment contains a diversity of microorganisms, dominantly Acidobacteria, Proteobacteria, Actinobacteria, Verrucomicrobia, Bacteroidetes, and Planctomycetes [67]. In the present study, Acidobacterium, Chloroflexi, Proteobacteria and Actinobacteria dominated the soil bacterial community in pure and mixed-species plantations. The dominance of Proteobacteria, Acidobacteria, and Actinobacteria have been often associated with the acidic nature of forest soils [44,68]. The relative abundance of Acidobacterium, Chloroflexi and Proteobacteria accounted for about 80% of the phylum composition in pure A. cincinnata stand and for about 78% in mixed-species planting of A. cincinnata and E. robusta as well as A, cincinnata and A. mangium. Acidobacterium was the dominant bacterial phylum of the soil from pure A, cincinnata plantation, but it had a relative abundance of only 25-35%, which was much lower than that of areas with acidic soil. For instance, Acidobacterium accounted for 53.3%, 67.8% and 60% of the soil samples of coniferous forest, mixed forest and broadleaf forest, respectively in the soil of Dinghu forest with pH 3.00-4.50 [69]. Soils of Fujian province, where the study was carried out, are acidic, but other soil attributes might differ between the two sites. However, the relative abundance of Proteobacteria and Actinobacteria was higher in soils of mixed stands than in pure A. cincinnata stand; whereas the relative abundance of Firmicutes in the soil of A. cincinnata + A. mangium stand was higher than that of the other two forest types, and the relative abundance of Verrucomicrobia was higher in A. cincinnata + E. robusta stand. This suggests a shift in the bacterial community as results of different planting systems, involving N2-fixing trees, as reforestation, N input and availability of mineral N are known to increase microbial diversity [70,71]. The relatively higher abundance of Firmicutes in A. cincinnata + A. mangium stand is probably linked to enhanced soil N cycling in this stand and to increased N content in the soil through atmospheric N fixation compared to A. cincinnata + E. robusta stand. This has been observed in the introduction of A. mangium into Eucalyptus plantation in the Congolese Coastal Plains, where N inputs change bacterial community structure and microbial biomass [61]. A study made on the distribution of soil microorganisms in different vegetation types in the new wetland of the Yellow River Delta also concluded that the different types and quantities of rhizosphere exudates of different plants directly affected the distribution of microbial populations [72].
N and P content is the main indicator of soil nutritional status in soil organic matter, are the most important nutrient elements in the process of bacterial metabolism, and have the most significant influence on the reproduction rate and quantity of bacteria [72]. Correlations between soil C, N, P and C:N:P ratios and relative abundance of dominant phyla of bacterial community has been detected, with specific phylum being associated with different soil attributes. According to the redundancy analysis of soil microbial communities, it was found that soil C:P and total P had the most significant effects on soil microbial community. Our results are consistent with previous studies that demonstrate a positive significant link between soil attributes and bacterial communities in Acacia and Eucalyptus mixed planting in the Congolese coastal plains [61] and in a broad-leaved forest ecosystem in central Germany [60], which are attributed to the effects of litterfall and root exudates. Significant differences in the distribution of soil bacterial community between Robinia pseudoacacia L. + Ailanthus altissima mixed forest and pure forest in the Yellow River Delta were observed [73,74] as well as in Larix gmelinii (Rupr.) Kuzen. and Pinus sylvestris var.Mongholica Litv, and Larix gmelinii + Pinus sylvestris mixed forest [75]. As a whole, several factors create heterogeneous environment in mixed-species stand; e.g. a shift in forest floor composition and quality and quantity of litter fall, which in turn, results in a different composition of the soil bacterial community.
5. Conclusions
Our study demonstrated the benefit of mixed-species planting in terms of increasing forest productivity and enhancing ecological values. Mixed-species planting of A. cincinnata and A. mangium provided the largest stocking volume compared to monoculture of A. cincinnata. Mixed-species planting improved the soil C, N, P contents due to increased litter fall and higher biomass of understory vegetation, whose turnover increases the physico-chemical properties of the soil. Soil bacterial diversity and community structure is shaped by species mixture. Thus, mixed-species planting of A. cincinnata and A. mangium is recommended to augment the value of ecological welfare forests by introducing economical valuable timber species. The increased biomass of understory vegetation could play an important in climate change mitigation through increased sequestration of carbon in the biomass. In addition, the turnover of the herbaceous species not only improves the nutrient status of the forest floor but also serves as bio-diverse energy sources for the soil bacterial to flourish. In conclusion, our work revealed that the sustainability of mixed-species plantation relies on the interactions between soil attributes, vegetation, and bacterial community.
Author Contributions
Conceptualization, D.W. and M.L.; Formal analysis, T.L.; Y.S. and L.W.; Funding acquisition, T.L. and D.W.; Investigation, R.X.; L.W. and M.L.; Methodology, Y.S.; R.X.; L.W. and M.L.; Supervision, M.L.; Validation, M.T.; Visualization, T.L.; Y.S. and L.W.; Writing—original draft, M.L.; Writing—review & editing, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Guizhou Provincial Science and Technology Projects ([2018]1160, [2017]1189) and ‘liang shan’ research project in Li shui University (FGLS202202, FGLS202204).
Data Availability Statement
The original sequencing data were deposited in the NCBI SRA database under the accession number PRJNA954280.
Acknowledgments
We thank staff members of the institute of forest and environment, college of Forestry, Fujian Agriculture and Forestry University for valuable discussion and assistance in fieldwork.
Conflicts of Interest
“The authors declare no conflict of interest.”
Appendix A
List of species recorded in the understory of different A. cincinnata stands, together with their height and coverage.
| A. cincinnata stand | ||
| Species | Height (cm) | Coverage(%) |
| Rubus corchorifolius | 35 | 20 |
| Pteris dispar | 25 | 7 |
| Digitaria sanguinalis | 20 | 20 |
| Dryopteris chinensis | 10 | 10 |
| Dicranopteris dichotoma | 30 | 30 |
| Glochidion puberum | 50 | 25 |
| Adiantum flabellulatum | 5 | 0.5 |
| Smilax glabra | 10 | 0.4 |
| Lygodium japonicum | 5 | 4 |
| Clerodendrum cyrtophyllum | 40 | 5 |
| Mean ± SE | 23 ± 5 | 12.2 ± 3.4 |
| A. cincinnata+ E. robusta Stand | ||
| Dicranopteris dichotoma | 40 | 70 |
| Digitaria sanguinalis | 15 | 2 |
| Toxicodendron vernicifluum | 27 | 1 |
| Hedyotis hedyotidea | 6 | 1 |
| Urena lobate | 15 | 0.4 |
| Sapium discolor | 40 | 2 |
| Adiantum flabellulatum | 10 | 0.4 |
| Rubus corchorifolius | 8 | 0.4 |
| Smilaz china | 5 | 0.5 |
| Paederia cruddasiana | 60 | 16 |
| Embelia laeta | 37 | 50 |
| Gahnia tristis | 24 | 5 |
| Litsea rotundifolia var. Oblongifolia | 40 | 6 |
| Mean ± SE | 25.2 ± 4.8 | 11.9± 6.1 |
| A. cincinnata+ A. mangium stand | ||
| Adiantum flabellulatum | 15 | 4 |
| Pericampylus glaucus | 10 | 5 |
| Embelia ribes | 40 | 20 |
| Gahnia tristis | 90 | 15 |
| Pteris dispar | 10 | 6 |
| Dicranopteris dichotoma | 100 | 20 |
| Sapium discolor | 60 | 5 |
| Ficus hirta Vahl | 60 | 5 |
| Melicope pteleifolia | 110 | 10 |
| Mean ± SE | 55 ± 13 | 10 ± 2.2 |
References
- Liu, S.R.; Yang, Y.J.; Wang, H. Development strategy and countermeasures of Chinese industrial and forestry management: from single-objective management pursuing wood yield to multi-objective management to improving the quality and efficiency of ecosystem services. Acta Ecologica Sinica 2018, 38, 1–10. [Google Scholar]
- Xu, C.; Lin, F.; Zhu, C. Li, C.; Cheng, B. Does Classification-Based Forest Management Promote Forest Restoration? Evidence from China’s Ecological Welfare Forestland Certification Program. Forests 2022, 13, 573. [Google Scholar] [CrossRef]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.-M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J.; Drössler, L.; Fahlvik, N.; Gong, P.; Holmström, E.; Keskitalo, E.C.H.; Klapwijk, M.J.; Laudon, H.; Lundmark, T.; Niklasson, M.; Nordin, A.; Pettersson, M.; Stenlid, J.; Sténs, A.; Wallertz, K. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 2016, 45 (Suppl 2), 124–139. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.J.; Zhang, J.X. Reflections on sustainable management of plantations. Science 2016, 68, 37–40. [Google Scholar]
- Sheng, L.L. Research on sustainable management of ecological public welfare forests in China. Northeast Forestry University 2011. [Google Scholar]
- Zou, J.M.; Li, J.Z.; Yu, H.Q.; Qiao, J.D.; Liao, W.M. Research on the value assessment of ecosystem services of ecological public welfare forest: A case study of Jiangxi Province. China Forestry Economy 2022, 8–14. [Google Scholar]
- Lv, C.Q. Preliminary study on afforestation of Acacia curly pod in southern Fujian. Anhui Agricultural Science Bulletin 2021, 27, 70–71. [Google Scholar]
- Maire, G.L. Tree and stand light use efficiencies over a full rotation of single- and mixed-species Eucalyptus grandis and Acacia mangium plantations. Forest Ecology and Management 2013, 288, 31–42. [Google Scholar] [CrossRef]
- Forrester, D.I. Soil Organic Carbon is Increased in Mixed-Species Plantations of Eucalyptus and Nitrogen-Fixing Acacia. Ecosystems 2013, 16, 123–132. [Google Scholar] [CrossRef]
- Liao, J.L. Experimental study on afforestation of pure forest and mixed forest of Eucalyptus grandis x urophylla and Acacia mangium. Journal of South China Normal University (Natural Science Edition) 2000, 70–73.
- Chen, J.B.; Liang, J.P.; Li, G.M.; Huang, D.Y.; Huang, R.L.; Zou, J.Y.; Hu, Y.G.; Zeng, J.J. Study on the effect of mixed forest on soil nutrients in. Guangxi Forestry Science 2004, 1-5+14. [Google Scholar]
- Zang, M.H.; Huang, B.L.; Cheng, L.; Lv, C.Q.; Li, Z.X.; Wei. L.X. Study on soil microbial characteristics of mixed forest land of Eucalyptus robusta - Acacia. Guangxi Agricultural Sciences 2009, 40, 681–685. [Google Scholar]
- Chen, W.l. Cultivation technology of timber forest in short rotation period of Acacia curly pod in southern Fujian mountain. Forestry Investigation and Design 2015, 13–17. [Google Scholar]
- Zhan, N.; Huang, L.J. ; Review of research progress of acacia tree species in China. Tropical Forestry 2015, 43, 41–45. [Google Scholar]
- Yang, H.G. Experimental study on artificial mixed management technology of Eucalyptus robusta - Acacia cincinnata. Forestry Investigation and Planning 2015, 40, 91–95. [Google Scholar]
- Zhao, W.D.; Li, K.; Wang, J.; He, Z.M. Current stock and water-holding characteristics of five kinds of artificial Acacia forest litter. Journal of Sichuan Agricultural University 2020, 38, 677–684. [Google Scholar]
- Hartemink A, E. Soil fertility decline in the tropics: with case studies on plantations. Cabi 2003. [Google Scholar]
- Chen, X.Z. Analysis of ecological forest supply problems and countermeasures in Yanping District, Fujian. China forestry economics 2017, 33–35. [Google Scholar]
- Guo, D.Q.; Lu, L.F.; Deng, Z.Y.; Lan, C.Z.; Mo, K.Z.; Chen, J.B. Research progress of eucalyptus mixed forest in China. Eucalyptus. Science and Technology 2018, 35, 27–32. [Google Scholar]
- Nigussie, M.B.; Habeteyohannes, L.; Teshome, G. The Effect of Mixed Plantation on the Stand Yield and Soil Attributes of Eucalyptus globulus and Acacia decurrens in North Shewa Zone, Ethiopia. J Forest Res 2021, 10, 282. [Google Scholar]
- Zagatto, M.R.G. Interactions between mesofauna, microbiological and chemical soil attributes in pure and intercropped Eucalyptus grandis and Acacia mangium plantations. Forest Ecology and Management 2019, 433, 240–247. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, M.; Qi, L.; Zhao, C.; Zhang, W.; Zhang, Y.; Wen, W.; Yuan, J. Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration. Forests 2022, 13, 648. [Google Scholar] [CrossRef]
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology, Individuals, Populations and Communities, 3rd ed.; Blackwell Science: London, UK, 1996. [Google Scholar]
- Paula, R.R.; Bouillet, J-P.; de M. Gonçalves, J.L.; Trivelin, P. C. O.; Balieiro, F. de C.; Nouvellon, Y.; Oliveira, J. de C.; Júnior, J.C. de D.; Bordron, B.; Laclau, J-P. Nitrogen fixation rate of Acacia mangium Wild at mid rotation in Brazil is higher in mixed plantations with Eucalyptus grandis Hill ex Maiden than in monocultures. Annals of Forest Science 2018, 75, 14.
- Hoogmoed, M.; Cunningham, S.C.; Baker, P.; Beringer, J.; Cavagnaro, T.R. N-fixing trees in restoration plantings: Effects on nitrogen supply and soil microbial communities. Soil Biology & Biochemistry 2014, 77, 203–212. [Google Scholar]
- Oliveira, I.R.; Bordron, B.; Laclau, J-P.; Paula, R.R.; Ferraz, A.V.; Gonçalves, J.L.M.; le Maire, G.; Bouillet, J.P. Nutrient deficiency enhances the rate of short-term belowground transfer of nitrogen from Acacia mangium to Eucalyptus trees in mixed-species plantations. Forest Ecology and Management 2021, 491, 119192.
- Edberg, S.; Tigabu, M.; Odén, P.C. Commercial Eucalyptus Plantations with Taungya System: Analysis of Tree Root Biomass. Forests 2022, 13, 1395. [Google Scholar] [CrossRef]
- Leuschner, C.; Hertel, D.; Coners, H.; Büttner, V. Root competition between beech and oak: a hypothesis. Oecologia 2001, 126, 276–284. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Laganière, J.; Reich, P.B.; Bergeron, Y.; Paré, D.; Yuan, Z.; Chen, H. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 2013, 210–219. [Google Scholar] [CrossRef]
- Ruiz-Blandon, B.A.; Salcedo-Pérez, E.; Rodríguez-Macías, R.; Hernández-Álvarez, E.; Campo, J.; Merino, A. Growth, biomass, carbon and nutrient pools in Gmelina arborea established in pure and mixed forest stand production systems in Mexico. New Forests 2022, 53, 735–757. [Google Scholar] [CrossRef]
- Viera M, Schumacher MV, Liberalasso E (2011) Crescimento e produtividade de povoamentos monoespecíficos e mistos de eucalipto e acácia-negra. Pesqui Agropecu Trop 41:415–421. [CrossRef]
- Tang, G.; Li, K.; Zhang, C.; Gao, C.; Li, B. Accelerated nutrient cycling via leaf litter, and not root interaction, increases growth of Eucalyptus in mixed-species plantations with Leucaena. For Ecol Manag 2013, 310, 45–53. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For Ecol Manag 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Vezzani, F.M.; Tedesco, M.J.; Barros, N.F. Alterações dos nutrientes no solo e nas plantas em consórcio de eucalipto e acácia negra. Rev Bras Ciênc Solo 2001, 25, 225–231. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved – a critical review for temperate and boreal forests. For. Ecol. Manage. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Messier, C.; Parent, S.; Bergeron, Y. Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests. J. Veg. Sci. 1998, 9, 511–520. [Google Scholar] [CrossRef]
- Augusto, L.; Dupoey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Lindroos, A.-J.; Derome, J.; Derome, K.; Smolander, A. The effect of Scots pine, Norway spruce and silver birch on the chemical composition of stand throughfall and upper soil percolation water in northern Finland. Boreal Env. Res. 2011, 16, 240–250. [Google Scholar]
- Macdonald, S.E.; Fenniak, T.E. Understory plant communities of boreal mixed wood forests in western Canada: Natural patterns and response to variable-retention harvesting. For. Ecol. Manage. 2007, 242, 34–48. [Google Scholar] [CrossRef]
- Jonsson, M.; Bengtsson, J.; Gamfeldt, L.; Moen, J.; Snäll, T. Levels of forest ecosystem services depend on specific mixtures of commercial tree species. Nat. Plants 2019, 5, 141–147. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Holmström, E.; Lindbladh, M.; Felton, A. Concealed by darkness: How stand density can override the biodiversity benefits of mixed forests. Ecosphere 2019, 10, e02835. [Google Scholar] [CrossRef]
- Ampoorter, E.; Selvi, F.; Auge, H.; Baeten, L.; Berger, S.; Carrari, E.; Coppi, A.; Fotelli, M.; Radoglou, K.; Setiawan, N.N.; Vanhellemont, M.; Verheyen, K. Driving mechanisms of overstorey – understorey diversity relationships in European forests. Perspect. Plant Ecol. 2016, 19, 21–29. [Google Scholar]
- Guo, J.; Feng, H.; McNie, P.; Liu, Q.; Xu, X.; Pan, C.; Yan, K.; Feng, L.; Goitom, E.A.; Yu, Y. Species mixing improves soil properties and enzymatic activities in Chinese fir plantations: A meta-analysis. Catena 2023, 220, 106723. [Google Scholar] [CrossRef]
- López-Vicente, M.; Álvarez, S. Stability and patterns of topsoil water content in rainfed vineyards, olive groves, and cereal fields under different soil and tillage conditions. Agric. Water Manag. 2018, 201, 167–176. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, L.; Gong, W.; Yang, X.; Kang, Y. Evaluation of the Water Conservation Function of Different Forest Types in Northeastern China. Sustainability 2019, 11, 4075. [Google Scholar] [CrossRef]
- Wen, X.F.; Li, Y.; Yang, W.F. Study on growth and soil physicochemical properties of mixed forest of Pinus koraiensis and Alnus sibirica. Liaoning Forestry Science and Technology 2012, 19+34. [Google Scholar]
- Singh, K.; Singh, B.; Singh, R.R. Changes in physico-chemical, microbial and enzymatic activities during restoration of degraded sodic land: Ecological suitability of mixed forest over monoculture plantation. Catena 2012, 96, 57–67. [Google Scholar] [CrossRef]
- Huang, W.Y. Analysis of the effect of Acacia cincinnata in the second generation budding of Eucalyptus grandis x urophylla. Collection 2019, 17. [Google Scholar]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8. [Google Scholar] [CrossRef]
- Awad, Y.M.; Blagodatskaya, E.; Yong, S.O.; Kuzyakov, Y. Effects of polyacrylamide, biopolymer, and biochar on decomposition of soil organic matter and plant residues as determined by C and enzyme activities. Eur. J. Soil Biol. 2012, 48, 1–10. [Google Scholar] [CrossRef]
- Berger, T.W.; Berger, P. Greater accumulation of litter in spruce (Picea abies) compared to beech (Fagus sylvatica) stands is not a consequence of the inherent recalcitrance of needles. Plant Soil 2012, 358, 349e369. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Wen, D.; Yu, K. . Soil potential labile but not occluded phosphorus forms increase with forest succession. Biol. Fertil. Soils 2016, 52, 41e51. [Google Scholar] [CrossRef]
- Xie, H.; Tang, Y.; Yu, M.; Wang, G.G. The effects of afforestation tree species mixing on soil organic carbon stock, nutrients accumulation, and understory vegetation diversity on reclaimed coastal lands in Eastern China. Global Ecology and Conservation 2021, 26, e01478. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Pienaar, L.V.; Shiver, B.D. Early results from an old-field loblolly pine spacing study in the Georgia Piedmont with competition control. South J. Appl. For. 1993, 17, 193–196. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, Z. Research advance on adaptation mechanism of forest tree to low-phosphorus stress and genetics of phosphorus efficiency. Forest Research 2002, 15, 734–740. [Google Scholar]
- Zhang, W.; Li, J.J.; Xiang, M.Z.; Huang, H.M.; Li, C.H.; Yan, J.L.; Gao, G.N.; Su, X.Y.; You, Y.M.; Huang, X.M. Effects of nitrogen-fixing tree Acacia mangium on particle size distribution and stability of soil aggregates in Eucalyptus grandis plantations. Flora of Guangxi 1–18.
- Huang, P.; Wang, N.; Zhou, Z.Y.; Wang, T.; Yuan, Z.L.; Chang, J.T.; Ye, Y.Z. Correlation analysis of soil bacterial community structure and environmental factors in deciduous broad-leaved forest in Baiyun Mountain. Journal of Henan Agricultural University 2020, 54, 415–421. [Google Scholar]
- Dukunde, A.; Schneider, D.; Schmidt, M.; Veldkamp, E.; Daniel, R. Tree Species Shape Soil Bacterial Community Structure and Function in Temperate Deciduous Forests. Front. Microbiol. 2019, 10, 1519. [Google Scholar] [CrossRef] [PubMed]
- Koutika, L-S.; Fiore, A.; Tabacchioni, S.; Aprea, G.; de Araujo Pereira, A. P.; Bevivino, A. Influence of Acacia mangium on Soil Fertility and Bacterial Community in Eucalyptus Plantations in the Congolese Coastal Plains. Sustainability 2020, 12, 8763. [CrossRef]
- Li, J.G.; Shen, M.C.; Hou, J.F.; Li, L.; Wu, J.X.; Dong, Y.H. Effect of Different Levels of Nitrogen on Rhizosphere Bacterial Community Structure in Intensive Monoculture of Greenhouse Lettuce. Sci. Rep. 2016, 6, 25305. [Google Scholar] [CrossRef]
- Zhu, H.; He, X.; Wang, K. Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem. European Journal of Soil Biology 2012, 51, 1–7. [Google Scholar] [CrossRef]
- Zhang, Z.F. Effects of mixed breeding of precious native tree species and Eucalyptus on biomass, carbon storage and soil physicochemical properties. Guangxi University 2020. [Google Scholar]
- Alves, L.F.; Vieira, S.A.; Scaranello, M.A. Forest structure and live aboveground biomass variation along an elevational gradient of tropical Atlantic moist forest (Brazil). Forest ecology and management 2010, 260, 679–691. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Abrantes, I.; Barra Caracciolo, A.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proença, D.N. Belowground Microbiota and the Health of Tree Crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-Based Assessment of Soil PH as a Predictor of Soil Bacterial Community Structure at the Continental Scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Liu, C.L.; Zuo, W.Y.; Zhao, Z.Y.; Qiu, L.H. Diversity of forest soil bacteria at different succession stages in Dinghu Mountain. Acta Microbiol 2012, 52, 1489–1496. [Google Scholar]
- Liu, J.; Yang, Z.; Dang, P.; Zhu, H.; Gao, Y.; Ha, V.N.; Zhao, Z. Response of Soil Microbial Community Dynamics to Robinia Pseudoacacia, L. Afforestation in the Loess Plateau: A Chronosequence Approach. Plant Soil 2018, 423, 327–338. [Google Scholar] [CrossRef]
- Li, Y.; Tian, D.; Wang, J.; Niu, S.; Tian, J.; Ha, D.; Qu, Y.; Jing, G.; Kang, X.; Song, B. Differential Mechanisms Underlying Responses of Soil Bacterial and Fungal Communities to Nitrogen and Phosphorus Inputs in a Subtropical Forest. PeerJ 2019, 7, e7631. [Google Scholar] [CrossRef]
- Chen, W.F.; Shi, Y.X. Microbial distribution characteristics of soils of different vegetation types in the new wetlands of the Yellow River Delta. Journal of Grassland 2010, 18, 859–864. [Google Scholar]
- Ding, X.J.; Huang, Y.L.; Jing, R.Y.; Ma, F.Y.; An, R.; Tian, Q.; Chen, B.J. Study on soil bacterial structure and diversity of four plantations in the Yellow River Delta based on high-throughput sequencing. Acta Ecologica Sinica 2018, 38, 5857–5864. [Google Scholar]
- Ding, X.J.; Xie, G.L.; Jing, R.Y.; Ma, F.Y.; Liu, F.C.; Ma, H.L. Decomposition characteristics of litter in different artificial Robinia pseudoacacia mixed forests in the Yellow River Delta. Journal of Soil and Water Conservation 2016, 30, 249–253+307. [Google Scholar]
- Zou, L.; Tang, Q.M.; Wang, Y. Community distribution characteristics of soil microorganisms in pure and mixed forests of Larix gmelinii and Pinus sylvestris. Journal of Northeast Forestry University 2010, 38, 63–64+79. [Google Scholar]
Figure 1.
Soil physical properties of Acacia cincinnata stands with different species mixture (Mean ± SE). Means followed by different lowercase letters varied significantly (P < 0.05) among soil layers within the same forest stand type and uppercase letter denotes significant differences (P < 0.05) among forest stand types within the same soil layer.
Figure 1.
Soil physical properties of Acacia cincinnata stands with different species mixture (Mean ± SE). Means followed by different lowercase letters varied significantly (P < 0.05) among soil layers within the same forest stand type and uppercase letter denotes significant differences (P < 0.05) among forest stand types within the same soil layer.
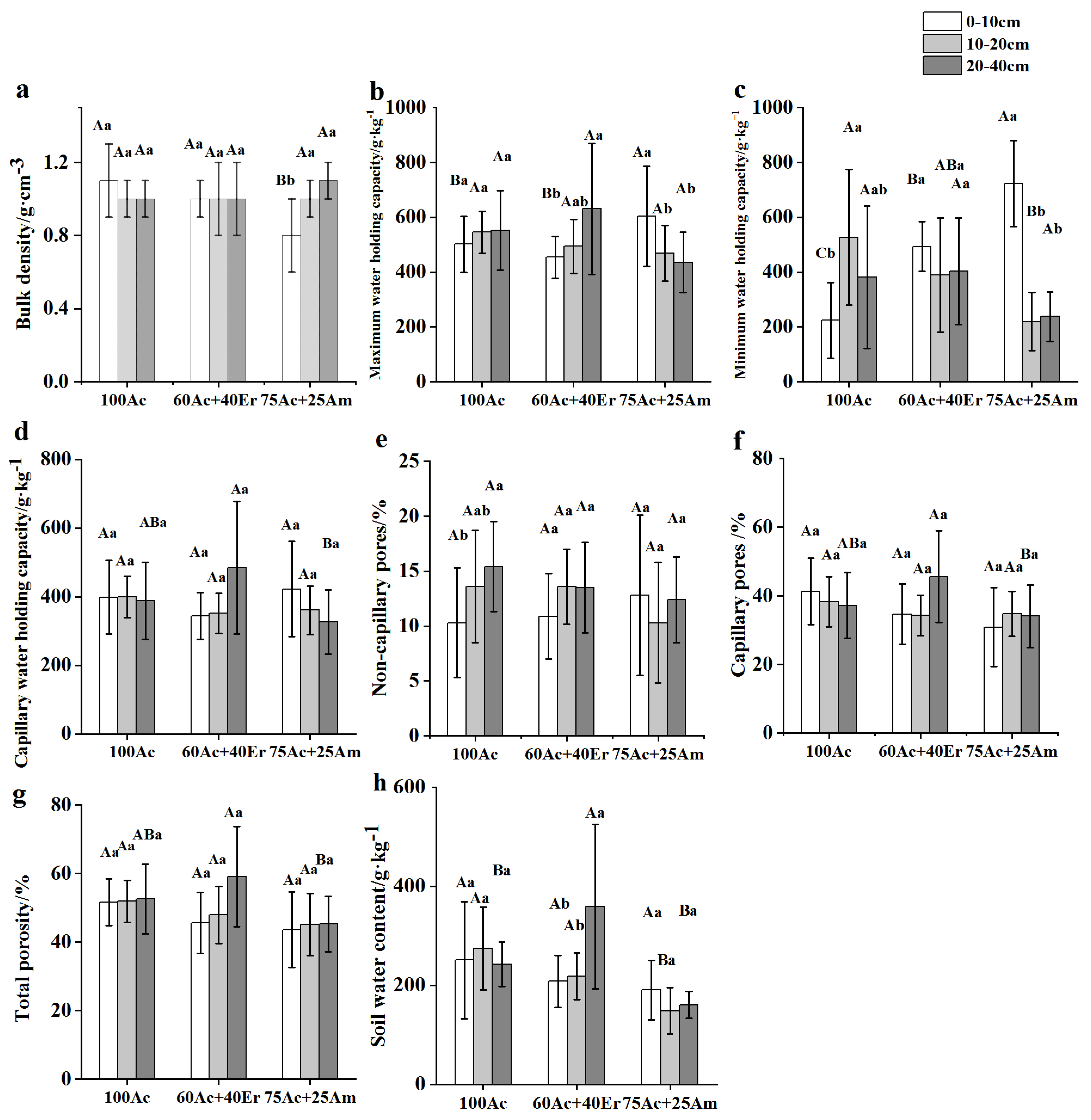
Figure 2.
Soil Carbon, Nitrogen and Phosphorus contents and soil stoichiometry of Acacia cincinnata under different stand mixture (Mean ± SE). For the stand mixture; 75Ac + 25Am represents 75% A. cincinnata and 25% A. mangium; 60Ac + 40Er represents 60% A. cincinnata and 40% E. robusta; and 100Ac represents 100% A. cincinnata in the stand. Means followed by different lowercase letters varied significantly (P < 0.05) among soil layers within the same forest stand type and uppercase letter denotes significant differences (P < 0.05) among forest stand types within the same soil layer.
Figure 2.
Soil Carbon, Nitrogen and Phosphorus contents and soil stoichiometry of Acacia cincinnata under different stand mixture (Mean ± SE). For the stand mixture; 75Ac + 25Am represents 75% A. cincinnata and 25% A. mangium; 60Ac + 40Er represents 60% A. cincinnata and 40% E. robusta; and 100Ac represents 100% A. cincinnata in the stand. Means followed by different lowercase letters varied significantly (P < 0.05) among soil layers within the same forest stand type and uppercase letter denotes significant differences (P < 0.05) among forest stand types within the same soil layer.
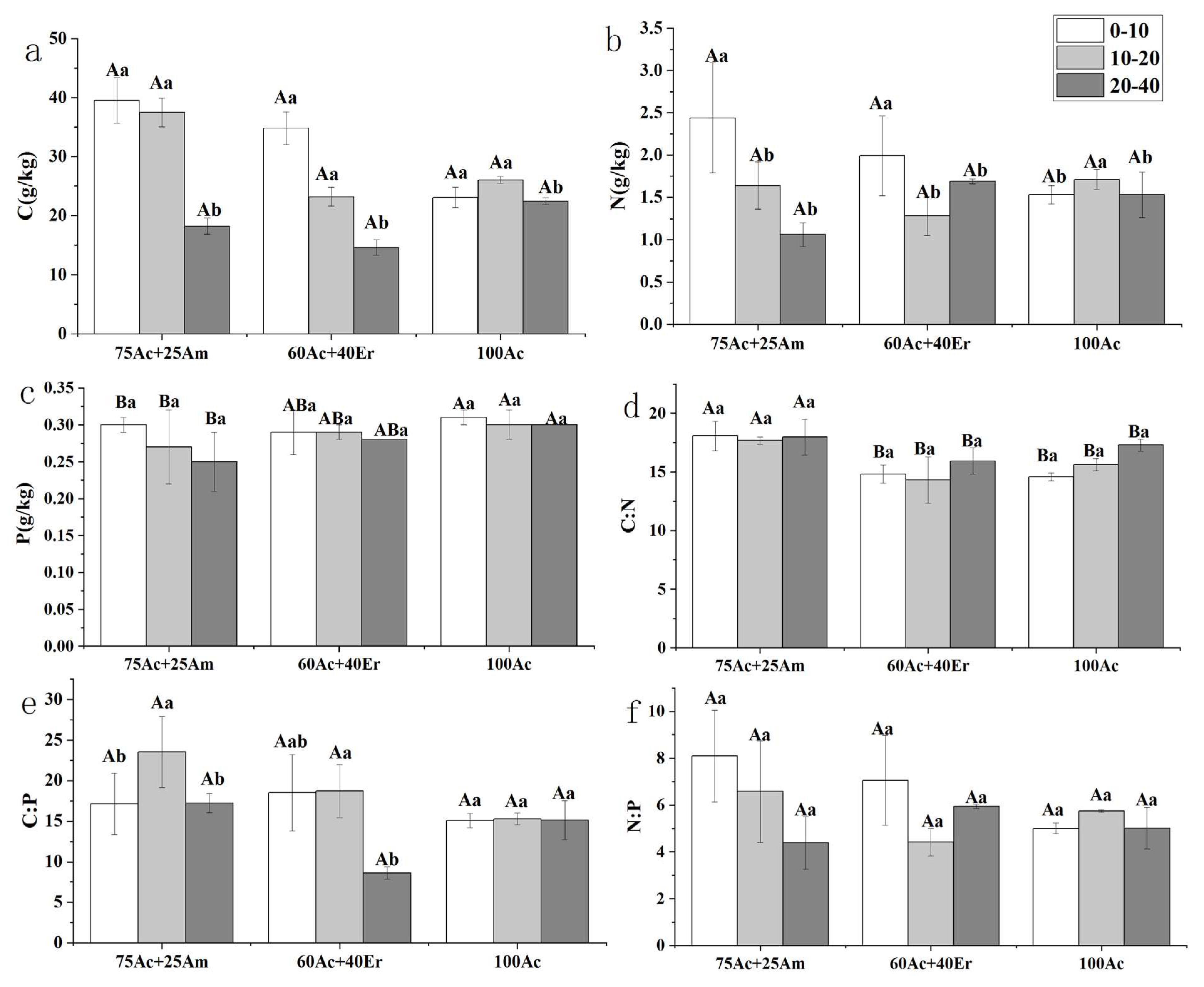
Figure 3.
Venn diagram of the distribution of soil bacterial operational taxonomic units in different acacia stand types.
Figure 3.
Venn diagram of the distribution of soil bacterial operational taxonomic units in different acacia stand types.
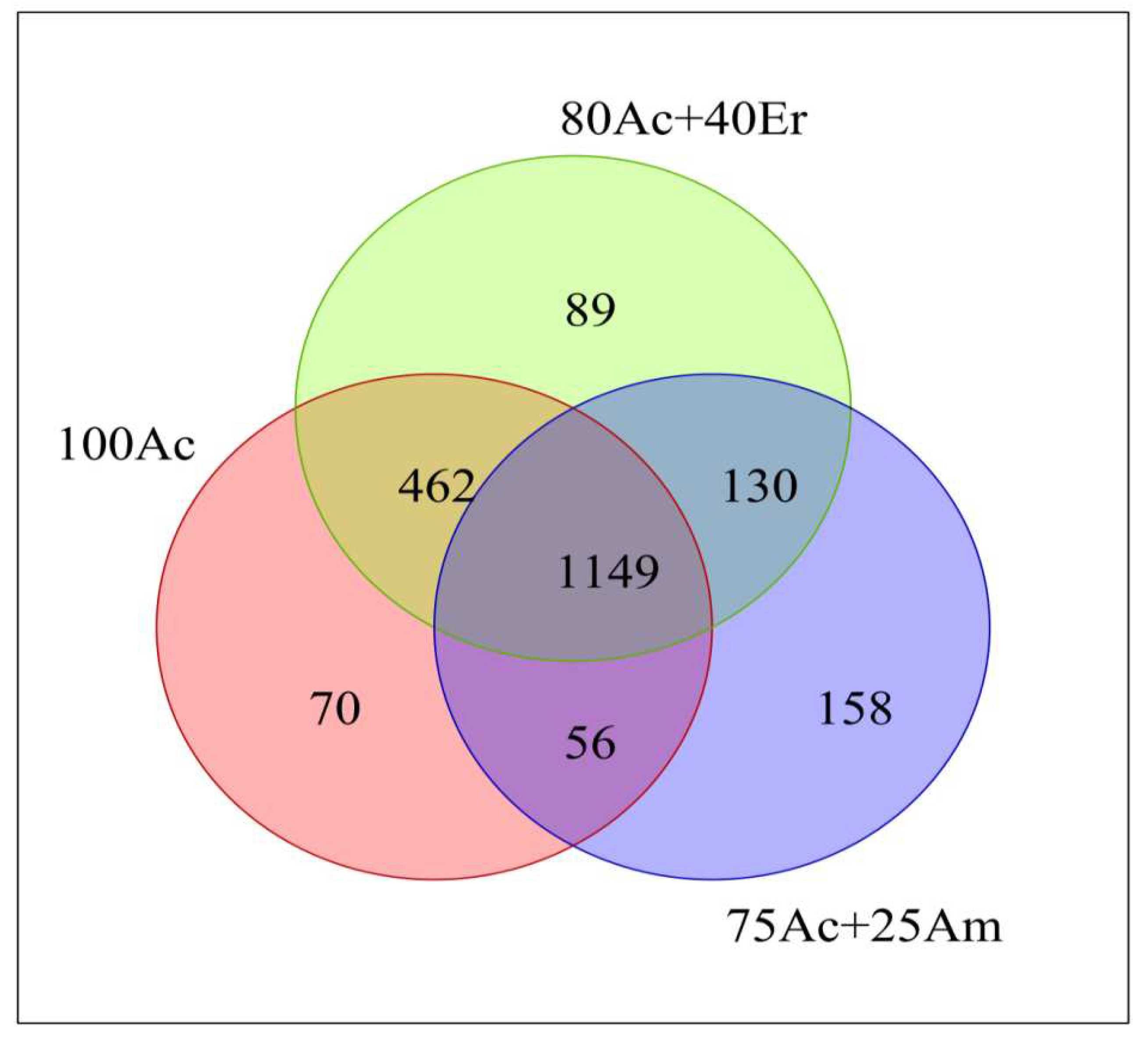
Figure 4.
Relative abundance of bacterial community composition at the phylum level.
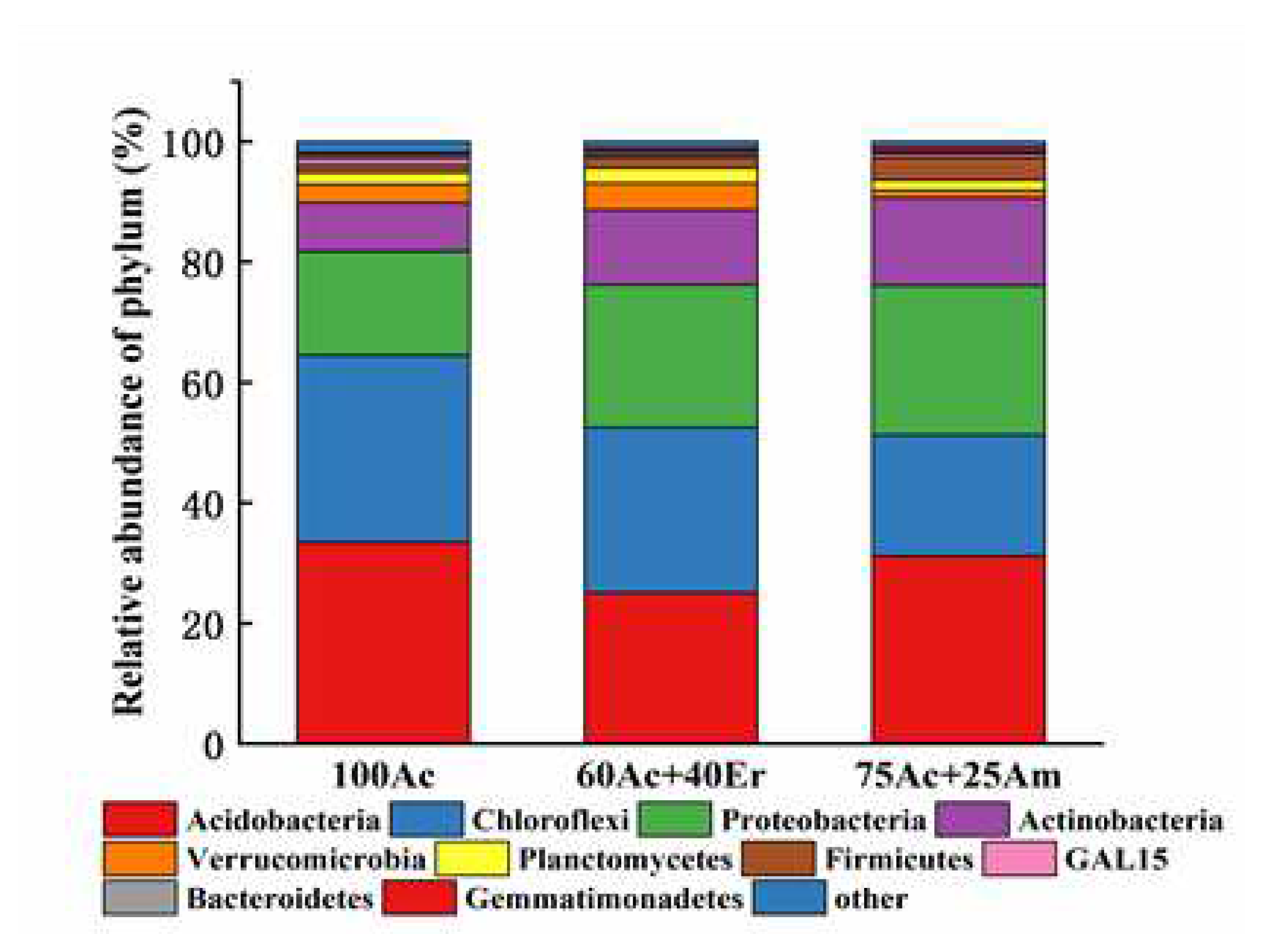
Figure 5.
Redundancy analysis of rhizosphere soil physical and chemical properties and bacterial community (phylum level) of Acacia cincinnata plantations.
Figure 5.
Redundancy analysis of rhizosphere soil physical and chemical properties and bacterial community (phylum level) of Acacia cincinnata plantations.
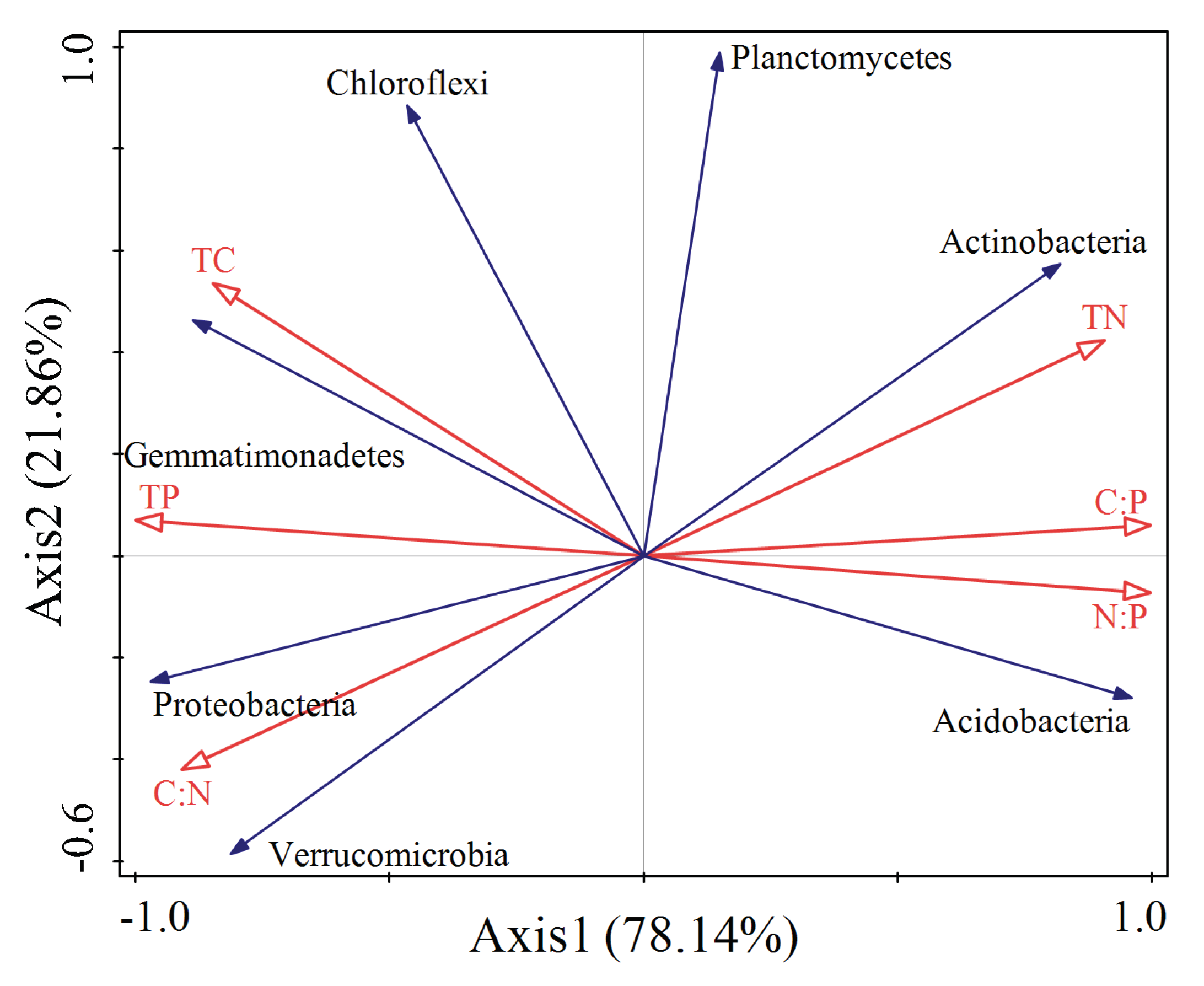

Table 1.
Growth of tree species under different species mixture modes. Means across the column that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
Table 1.
Growth of tree species under different species mixture modes. Means across the column that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
| Stand type* | Species | Diameter (cm) | Height (m) | Volume per tree (m3) | Stand volume (m3/hm2) |
Total stock (m3/hm2) |
|---|---|---|---|---|---|---|
| 75Ac+ 25Am | Acacia cincinnata | 9.6 ± 0.7a | 10.1 ± 0.2a | 1.06 ± 0.2a | 18.34 ± 3.03a | 109.6 ± 14.9b |
| A. mangium | 11.3 ± 0.1b | 11.9 ± 0.1bc | 1.62 ± 0.04b | 91.23 ± 15.27c | ||
| 60Ac + 40Er | Acacia cincinnata | 9.9 ± 0.4a | 10.5 ± 0.1a | 1.12 ± 0.1a | 28.37 ± 12.56a | 84.6 ± 23.9ab |
| E. robusta | 13.4 ± 2.3b | 14.9 ± 2.16c | 2.80 ± 1.01b | 56.20 ± 18.75b | ||
| 100Ac | Acacia cincinnata | 10.4 ± 0.1a | 10.8 ± 0.01a | 1.24 ± 0.04a | 61.12 ± 12.95b | 61.1 ± 9.8a |
For stand type, 75Ac + 25Am represents 75% A. cincinnata and 25% A. mangium; 60Ac + 40Er represents 60% A. cincinnata and 40% E. robusta; and 100Ac represents 100% A. cincinnata in the stand.
Table 2.
Biomass of herbaceous and shrub species in the understory of different Acacia cincinnata stands (Mean ± SE). Means across the column that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
Table 2.
Biomass of herbaceous and shrub species in the understory of different Acacia cincinnata stands (Mean ± SE). Means across the column that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
| Forest stand type* | Herbal biomass(Kg/ha) | Shrub biomass (Kg/ha) | |||||
|---|---|---|---|---|---|---|---|
| Above ground | Under ground | Total biomass | root | branch | leaf | Total biomass | |
| 75Ac + 25Am | 820.1±200.4a | 429.9 ± 71.2a | 1250.0 ± 251.0a | 285.3 ± 186.5a | 378.7 ± 171.7a | 151.9 ± 40.7a | 815.8 ± 392.4a |
| 60Ac + 40Er | 254.4±103.7b | 258.1 ± 12.2b | 512.5 ± 106.9b | 54.6 ± 12.4b | 141.4 ± 52.3b | 88.0 ± 30.0ab | 284.1 ± 79.1b |
| 100Ac | 752.8±123.4a | 386.6 ± 87.7a | 1139.4 ± 184.1a | 109.8 ± 31.5a | 112.6 ± 37b | 76.9 ± 26.5b | 299.2 ± 81.7b |
Table 3.
Nutrient concentrations of understory vegetation in different Acacia cincinnata forest stand types (Mean ± SE). Means across the rows that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
Table 3.
Nutrient concentrations of understory vegetation in different Acacia cincinnata forest stand types (Mean ± SE). Means across the rows that were followed by different lowercase letters varied significantly (P < 0.05) among forest stand types.
| Vegetation | Plant Parts | Nutrient | Forest stand type* | ||
|---|---|---|---|---|---|
| 75Ac + 25Am | 60Ac + 40Er | 100Ac | |||
| Herbal biomass | Above ground | N | 9.1 ± 1.1a | 11.6 ± 1.1ab | 13.7 ± 1.3b |
| C | 363.7 ± 61.6a | 402.3 ± 28.8a | 420.8 ± 14.6a | ||
| P | 1.0 ± 0.1a | 0.8 ± 0.3a | 1.2 ± 0.3a | ||
| Under ground | N | 8.1 ± 0.7b | 7.1 ± 1.2b | 5.7 ± 0.3a | |
| C | 244.8 ± 26.4b | 282.4 ± 40.9b | 185.8 ± 3.2a | ||
| P | 0.8 ± 0.1a | 0.7 ± 0.3a | 1.5 ± 0.0b | ||
| Shrub biomass | root | N | 15.5 ± 1.5c | 10.3 ± 0.1b | 7.6 ± 0.0a |
| C | 404.5 ± 18.4a | 420.2 ± 5.6a | 420.5 ± 0a | ||
| P | 0.6 ± 0.1b | 0.3 ± 0.1a | 0.5 ± 0.0b | ||
| branch | N | 11.4 ± 1.3a | 13.1 ± 4.01a | 12.7 ± 0.0a | |
| C | 453.6 ± 3.9a | 445.0 ± 9.2a | 450.5 ± 0.0a | ||
| P | 0.6 ± 0.1a | 0.5 ± 0.2a | 0.9 ± 0.0b | ||
| leaf | N | 22.5 ± 0.6c | 17.3 ± 1.9b | 14.3 ± 0.0a | |
| C | 466.0 ± 1.3c | 431.7 ± 5.0a | 449.6 ± 0.0b | ||
| P | 0.5 ± 0.0a | 0.5 ± 0.3a | 0.7 ± 0.0a | ||
* For forest stand type, 75A + 25A represents 75% A. cincinnata and 25% A. mangium; 60A + 40E represents 60% A. cincinnata and 40% E. robusta; and 100A represents 100% A. cincinnata in the stand.
Table 4.
Soil bacterial diversity indices in three different Acacia cincinnata stands (Mean ± S). Means followed by different letters ae significantly different (P < 0.05) between forest stand types.
Table 4.
Soil bacterial diversity indices in three different Acacia cincinnata stands (Mean ± S). Means followed by different letters ae significantly different (P < 0.05) between forest stand types.
| Forest stand* | |||
|---|---|---|---|
| Bacterial diversity index | 100Ac | 60Ac + 40Er | 75Ac + 25Am |
| Chao1 | 1605.5 ± 49.9a | 1546.4 ± 45.3a | 1297.4 ± 86.0b |
| Observed_ species | 1327.8 ± 47.3a | 1315.9 ± 76.26a | 1109.3 ± 55.1b |
| PD_whole_tree | 85.8 ± 2.9a | 84.8 ± 4.6a | 71.3 ± 3.1b |
| Shannon | 8.1 ± 0.2a | 8.3 ± 0.1a | 7.8 ± 0.1a |
* 100Ac, 60Ac + 40Er and 75Ac + 25Am represents A. cincinnata forest stand, A. cincinnata + E. robusta mixed forest stand and Acacia cincinnata + Acacia mangium forest stand, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



