
Submitted:
12 April 2023
Posted:
13 April 2023
You are already at the latest version
Abstract
Salmon aquaculture generates 80% of the total revenue of finfish aquaculture across Canada. Salmon farming is carried out in a multilevel process, and at least 60% of the total production is considered as by-products, including skin, head, viscera, trimming, frames, bones, and roes. These by-products are an excellent source of protein, which can be converted to protein hydrolysates through enzymatic hydrolysis and non-enzymatic processes such as chemical hydrolysis (acid and alkaline) and salt extraction. Several studies have reported that peptides from salmon protein hydrolysates possess bioactivities, including antihypertensive, antioxidant, anticancer, antimicrobial, antidiabetic, anti-allergic, and cholesterol-lowering effects. Incorporating in-silico computational methods is gaining more attention to identify potential peptides from source protein. The in-silico methods can be used to predict the properties of the peptides and thereby predetermine the processing, isolation, and purification steps that can be used for the peptides of interest. Therefore, it is essential to implement robust, standardized, and cost-effective processing techniques that can easily be transferrable and scale up for industrial applications. This contribution summarizes the latest research information on Atlantic salmon, production statistics, growth lifecycle, processing, protein production techniques, nutritional and functional properties, peptide production and purification processes, as well as potential health benefits as a nutraceutical product.
Keywords:
Atlantic salmon
; aquaculture
; protein hydrolysate
; bioactive peptide
; in-silico production
; health effect
1. Introduction
Fish protein is an important source of nutrition for millions of people around the world. Wild fisheries alone cannot sustain the global population growth hence aquaculture has provided a means to grow fish in a controlled environment. In both cases, the landed fish is processed and the fillets so produced are sold for human consumption. However, fish processing produces various by-products, including head, skin, trimmings, fins, frames, viscera, bones, and roe, accounting for 60% of the total processing output [1,2]. These processing by-products are often dumped in the ocean or landfills as waste in some parts of the world. In Canada, the USA, Norway, China, and many other countries, some of these by-products are sold at low cost to various industries for animal feed and fertilizer production. In addition, Canadian processors export fish heads to various markets as food delicacies [3,4].
These by-products are an excellent source of proteins, lipids, minerals, polysaccharides, and carotenoids. However, it is possible to increase the value of fish by-products by using a gentle, environmentally friendly process like enzymatic hydrolysis to produce high-quality products such as protein, oil, and minerals as primary products, which could be further processed and purified into omega-3 fatty acids, collagen, gelatin, bioactive peptides, enzymes, hydroxyapatite, and minerals [5,6]. Several proteolytic enzymes such as Alcalase, Flavourzyme, Pronase, Neutrase, Protamex, Sea-B-Zyme L200, Validase, bromelain, papain, pepsin, trypsin, chymotrypsin, pancreatin, and thermolysin can be used to hydrolyze fish by-products/ processing discards [2,7,8,9,10,11,12,13,14,15,16,17,18]. Protein hydrolysates are also produced along with oil, bones, and sludge upon enzymatic hydrolysis of fish by-products. During the hydrolysis process, proteins in fish by-products are solubilized, and the peptide bonds between amino acids are cleaved, generating smaller peptides. These peptides have a smaller molecular size with more ionizable amino and carboxyl groups [19,20]. The amino acid composition of fish proteins is well-balanced and provides high nutritional value for humans. In addition, the isolated protein hydrolysates have demonstrated various bioactivities, including antihypertensive, antioxidant, anticancer, antimicrobial, antidiabetic, anti-allergic, and cholesterol-lowering effects. Therefore, fish by-products play a vital role in developing various nutraceuticals and functional food products that can prevent or enhance the management and treatment of human diseases and maintain optimal human health [16,20]. To the best of our knowledge, this is the first literature review that attempts to thoroughly summarize the research work carried out on Atlantic salmon in order to produce bioactive peptides. It discusses the production status of Atlantic salmon in Canada, including Newfoundland, its production lifecycle, processing, and current utilization. It also reviews different methods of protein hydrolysate production and nutritional composition and functional properties of the resultant products. Finally, different bioactive production techniques, purification, separation, and characterization of peptides isolated from salmon processing discards are reviewed. In addition, the important health benefits and the commercial approval process of salmon peptides are provided.
2. Salmon Production
In Canada, salmon is produced by commercial fisheries (fresh and seawater), including wild capture and aquaculture. Canada is the fourth-largest producer of farmed salmon in the world. Three types of salmon, including Atlantic salmon (Salmo salar), Chinook salmon (Oncorhynchus tshawytscha), and Coho salmon (Oncorhynchus kisutch), are farmed in Canada. The United States of America is the largest export market for salmon farmed in Canada. Canada's aquaculture industry is a significant employer and economic driver in many coastal, rural, and Aboriginal communities. According to the Department of Fisheries and Oceans (DFO) Canada and Statistics Canada, the average annual finfish produced between 2008 and 2021 was 131,316 tonnes, with an average annual value of $947 million (Figure 1) [21]. The corresponding average shellfish production was 38,719 tonnes, with an average annual value of $85 million. Salmon, along with mussels, trout, scallops, oysters, clams, and other finfish species, is farmed in various Canadian regions (Figure 2). The production statistics between 2008 and 2021 indicate the dominancy of salmon among the majority of finfish aquaculture, with an average annual production of 111,752 tonnes valued at $781 million. Salmon aquaculture generates 80% of the total revenue of finfish aquaculture in Canada [21]. These statistics further reflect the importance of salmon aquaculture to the overall fisheries in Canada.
Newfoundland and Labrador's (NL) aquaculture has been steadily growing in the last decade, with a total finfish production of 19,635 tonnes and a value of $144 million in 2021 (Figure 3) [21]. Newfoundland is strategically located with 17,000 km of coastline with extensive open areas of clear cold water suited for aquaculture. Specific areas within the province are desirable aquaculture locations, including Notre Dame Bay, Green Bay, the Connaigre Peninsula, Bay d' Espoir, Fortune Bay, and Placentia Bay. Newfoundland's aquaculture industry is dominated by salmon and along with small quantities of steelhead trout and mussels. The NL salmon aquaculture produced 15,904 tonnes, valued at $138 million in 2021 and accounted for 82% of the total production and 96% of the total aquaculture revenue [22].
3. Atlantic Salmon Production Lifecycle
Salmon farming is carried out in a multilevel process as illustrated in Figure 4. In the first step of production, salmon eggs are collected from the broodstock (parent fish) selected from the best-performing fish on a sea farm and are moved into freshwater tanks or cages, usually during the autumn. The eggs are fertilized by mixing them with milt from mature male fish and nurtured at a hatchery in freshwater tanks for fifteen months. During the time inside the freshwater hatchery, the eggs hatch and young salmons are called “alevin”. The size of the alevin is about 2 cm in length. Then the young salmon enters the second stage called “fry”. In this stage, the salmon fry comes up from the gravel and starts feeding on microscopic life in the stream, and it grows to a length of 5-8 cm [23].
The fry matures and starts to feed (early spring), and they are transferred to small tanks in the hatchery. At 12 months (second spring), salmons undergo physiological transformation to reach a weight of 80-120 g, and the process is called smoltification. The smolts are placed in specialized grow-out farms where they are raised to marketable size in sea pens. The biomass development of Atlantic salmon depends on the season and hence smolts are usually released into the seawater during the warmer half of the year. Usually, under normal circumstances, high mortality rates of smolts are observed in the first two months after release to the seawater. The mortality is due to the smolt body's inability to absorb salt at the release time. The salmon are fed for 12-18 months prior to harvesting. Salmon are harvested once they reach a marketable weight of 4-8 kg [24,25,26]. During harvesting, fish are starved inside the sea farms for 2-3 days, and then transported to the slaughter plant alive. The fish is electrically stunned in the slaughter plant with a blow to the head, followed by bleeding and immersing in iced water. The fish is then gutted, washed, and chilled to maintain the flesh temperature at 3°C. Then depending upon the market, the salmon is sold as a whole or filleted, and head and frames are sold as raw materials for animal feed or pet food manufacturing. In another type of harvesting, salmons are pre-processed on board before being brought to the processing plant. In this case, harvested salmons are directly immersed in carbon dioxide-rich water for a short time and processed onboard by quickly rupturing their arteries and transferring them into ice-filled containers. The bled salmons are transported in ice to the processing plant and gutted, washed, chilled, graded by weight and packaged for the market. The harvesters use different types of proprietary fish feed based on various seasons during their growth. The salmon is harvested before it reaches maturity at 28 months after the first hatch. Salmon do not die after reaching sexual maturity. However, there is a degradation in quality due to spawning. Upon spawning, harvesting has to be delayed for one year [23,27,28].
4. Atlantic Salmon Processing and Utilization of By-Products
After harvesting, the salmon is processed in the processing plant, and the fish is sold as head-on-gutted, butterfly fillets, skin-on fillets, skinless fillets depending on the market requirement. In this case, the salmon skins are discarded as waste. Salmon heads and frames are mostly shipped to food and feed processing industries for low-end utilization, such as pet food, mink feed and fertilizer. Salmon guts are usually dumped as waste. However, these by-products can be utilized in a far better manner to produce various high-value nutraceuticals and other industrial products, including protein, fish oil, biodiesel, collagen, gelatin, bioactive peptides, enzymes, hydroxyapatite and minerals [5,29]. Thus, valorization of fisheries resources could be achieved by upgrading them into high-value products (Figure 5). The chain of events shown in Figure 5 starts from salmon by-products to fish meal to fish protein isolate to fish protein hydrolysates and finally to bioactive peptides with potential health effects. Fish meal and silage already exist as high-volume products with relatively low prices [10].
Protein isolates can be produced by employing at pH shift technique. In this technique, fish mince is solubilized with acid (below pH 3.5) to extract myofibrillar proteins. The lipids are skimmed, denser impurities are sedimented, and soluble protein is extracted by centrifugation. The supernatant is subjected to isoelectric precipitation at a pH of 5.2-5.5 (isoelectric point) to precipitate insoluble sarcoplasmic proteins. Finally, protein isolates are extracted by washing. Protein isolates provide more value addition when incorporated into seafood products [30,31]. The goal of moving towards the value chain in the form of salmon protein hydrolysates and bioactive peptides is often challenging in most parts of the world. The production of protein hydrolysates and bioactive peptides is relatively expensive compared to low-end products. These products are low-volume and high-value with documented health effects in the various scientific literature. The transformation of salmon processing by-products into high value products can revitalize many rural fish communities, fish producers and processors. However, production of these high-value products requires highly qualified personnel with technical expertise and skills in the field, which can be challenging at remote locations [2,10]. Besides, by-products variation in terms of amount and composition is common during salmon processing due to a diverse set of operations at various steps. The variable quality of the byproducts (raw materials), in some cases, makes them unfit to produce high-value products. Therefore, several steps need to be implemented to prevent raw material spoilage when the immediate production of protein hydrolysates is not possible. The spoilage of salmon by-products can be prevented by low-temperature storage (chilling), gutting, controlling water activity or by adding salt and sugar (to bind free water molecules to create an osmotic balance) and addition of food-grade antioxidants. Furthermore, to produce high-quality salmon protein and peptides, fresh raw materials without any oxidative changes, heating, pH change and freezing are recommended. Any chemical and physical changes in the raw material during storage can lead to variations in amino acids, digestibility and functional properties of the final product [10,32]
5. Chemical Composition of Atlantic Salmon
The proximate composition of salmon body parts is summarized in Table 1. The highest protein content (20.9%) is in the fillet portion of salmon. In contrast, the highest lipid (22.65%) and ash (1.94%) contents are present in salmon's frame/trimmings portion. Salmon fillets also contain a considerable amount of lipid (11.30%), with essential polyunsaturated fatty acids (PUFAs) and astaxanthin pigments responsible for the orange color of the fillets.
5.1. Lipid and fatty acid composition
The Atlantic salmon contains 17.30-22.12% lipid, which is mainly composed of both non-polar (neutral) and polar lipids. Neutral lipids, mainly triacylglycerols (TAGs), are dominant in salmon lipids. Meanwhile, salmon oil contains phospholipids (PL) as its intramuscular lipid [36]. The salmon body parts, including the head, frame and internal organs/ gut contribute to the composition of salmon oil. According to a recent study, salmon oil extracted from the head and frames and gut contained an average TAG content of 90.1 and 78.1%, respectively [37]. In another study, the crude oil extracted from farmed Atlantic salmon viscera contained 92.83% TAG [34]. Polvi and Ackman [38] reported 84-90% TAGs and 0.2-5.2% free fatty acids (FFA) in total lipids in cultured Atlantic salmon muscle, depending on the diet. Depending on the type of raw material, storage conditions, processing, oxidation parameters, and extraction technique, salmon oil has different levels of phospholipids and FFA. Liu et al. [37] reported an average phospholipid content of 2.1 and 3.8% in the oil extracted from salmon heads and frames, as well as guts, respectively. The study also indicated that a higher amount of FFA was present in the oil extracted from the gut (11.7%) compared to that from head and frames (1.5%). Dave et al. [34] reported 1.43% phospholipids and 1.23% FFA in the crude oil extracted from farmed Atlantic salmon viscera. On the other hand, Polvi and Ackman [38] reported 10-12% phospholipids and 0.2-5.2% FFA in total lipids in cultured Atlantic salmon muscle, depending on the diet.
The Atlantic salmon oil is mainly composed of monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs). Dave et al. [34] studied the fatty acid composition of oil extracted from by-products, including gut, head, and frame of farmed Atlantic salmon. The results indicated that the salmon gut, head, and frame contained saturated fatty acids (SFAs; 19.21-21.93 g/100 g), MUFAs (36.82-39.58 g/100 g), and PUFAs (38.89-39.83 g/100 g). The highest SFAs, MUFAs, and PUFAs present in the gut, head, and frame were palmitic acid, oleic acid, and linoleic acid. The omega-3 fatty acids including, eicosapentaenoic acid (EPA), docosapentaenoic acid (DPA), and docosahexaenoic acid (DHA) were present in all three parts of salmon. Similar results were reported elsewhere [39,40,41].
5.2. Protein and amino acid composition
The Atlantic salmon contains 19.1-20.4% protein, which is mainly composed of glutamic acid (5.96-6.46%), aspartic acid (4.63-4.92%), lysine (4.59-4.77%), leucine (3.78-4%), and valine (2.81-2.92%). Usually, farmed salmon has a lower total amino acid content than the wild salmon on a dry wet basis, though these are almost similar on a wet weight basis [42]. However, the content of amino acids is mainly dependent on the harvesting location, season, and processing techniques. For example, the protein contents of Atlantic salmon during different culinary treatments such as boiling, steaming, and oven-cooking were 16.69, 18.9, and 20.59%, respectively, while the raw fish had 14.73% protein [43]. They also reported that the major amino acids in cooked and raw salmon were aspartic acid and alanine. In particular, in cooked salmon, most essential amino acids, namely isoleucine, histidine, threonine, lysine, and valine, were significantly higher in raw salmon.
6. Production of Protein Hydrolysates
Traditionally, protein hydrolysates are prepared using several chemical extraction methods. However, techniques such as enzyme hydrolysis, microwave-assisted extraction, ultrasound-assisted extraction, high-pressure processing-assisted extraction, supercritical fluid extraction, pressurized solvent extraction, pulsed electric field-assisted extraction and fermentation are preferred [44,45]. Protein hydrolysates obtained from these processes are used in the food industry as milk replacers, protein supplements, stabilizers in beverages and flavor enhancers, among others.
6.1. Chemical hydrolysis
The chemical methods are carried out using acid and alkaline hydrolysis and salt extraction. One of the chemical methods still used in fisheries on a large scale is ensilaging. In the ensilaging process, spoiled fish, disease affected morts, underutilized species, by-products from marine fishing, commercial fish waste and industrial residues are mixed in large containers in the presence of various acids such as formic acid which activates endogenous enzymes and the pH is maintained at 3-4 for several days. During this process, the oil produced is collected and used for industrial purposes and/or as a feed ingredient if the silage is protected with antioxidants. Fish silage is an excellent protein product mixed with other ingredients and used for feed and fertilizer purposes [5].
6.2. Enzymatic hydrolysis
Enzymatic hydrolysis is the process of adding enzymes to fish material to break down tissues and produce protein hydrolysates with improved physiochemical, functional, and sensory properties of the native protein. Enzymatic hydrolysis process is carried out under mild extraction conditions and does not produce hydrolytic degradation products via racemization reactions common in acid and alkaline hydrolysis [46,47]. Enzymatic hydrolysis process is usually carried using different types of proteases. The proteases are categorized according to the specificity of their attack and mechanism against peptide bonds in the substrate. Based on the availability of structural and mechanistic information the proteases are classified into six distinct classes: aspartic, glutamic and metalloproteases, cysteine, serine and threonine proteases. Among these aspartic, glutamic and metalloproteases utilize activated water molecules as a nucleophile to attack the peptide bond of the substrate, whereas in cysteine, serine and threonine proteases the nucleophile is amino acid residue (C, S or T, respectively) located in the active site [48]. According to the catalytic mechanism, they are categorized as endopeptidases and exopeptidases. The endopeptidases hydrolyze the peptide bonds within protein molecules to produce relatively large peptides. The exopeptidases break the peptide bonds from either the N terminus (also called aminopeptidases) or the C terminus (also called carboxypeptidases). In some cases, both endopeptidases and exopeptidases are used for complete and effective hydrolysis of peptide bonds [47,49,50].
Enzymatic processing of salmon by-products helps producing a broad spectrum of food ingredients and industrial products for many applications. Several proteolytic enzymes are often used to hydrolyze fish protein hydrolysates. Some commercially produced proteases include Alcalase, Flavourzyme, Pronase, Neutrase, Protamex, protease N, protease A, Orientase, Sea-B-zyme, thermolysin, and Validase. In addition, there other various natural proteases such as chymotrypsin, pancreatin, papain, pepsin, trypsin, bromelain [2,46]. The muscle proteins present in salmon by-products are sensitive to various process parameters, including temperature, heating rate, pH, salt, ionic strength, oxidizing agent, and non-meat ingredients. Improper storage of salmon by-products and lack of controllable environment leads to poor protein quality [51,52].
The general hydrolysis procedure to extract protein from salmon by-products is shown in Figure 6. The first step in the enzymatic hydrolysis is the collection and processing of salmon by-products. During processing, salmon by-products are minced in a pilot-scale mincer producing particle size of less than 0.5 mm. During processing, salmon guts are always recommended to be processed separately due to relatively high amounts of endogenous enzymes, which could cause storage instability. To preserve the quality of raw materials and increase storage stability, food-grade antioxidants can be added before mincing. Minced salmon by-products are subjected to the hydrolysis process. During the hydrolysis process, various parameters are monitored and controlled according to optimum conditions. The hydrolysis parameters include reaction time, reaction temperature, type of enzyme, pH of the reaction system and enzyme concentration.
These parameters are optimized on a lab-scale, and the process can be scaled up to pilot level under optimum conditions. After hydrolysis, the reaction mixture is heated at 90°C for 10 min or the pH of the process is adjusted to deactivate the enzyme. The reaction mixture is subsequently subjected to separation techniques such as centrifugation in a lab-scale or using a decanter for a large scale. After separation, two main products, salmon oil and salmon protein hydrolysate, are obtained along with sludge and emulsion. The protein hydrolysate is then subjected to filtration to remove any suspended impurities and the oil. The filtered hydrolysate is then diafiltered to remove salt from the liquid. Some processes also include pasteurization of water-soluble protein to eliminate microorganisms. The diafiltered protein hydrolysate is then spray dried to obtain purified salmon protein powder. The crude salmon oil obtained after hydrolysis is flushed with nitrogen, and food-grade antioxidants are added before storage at -30°C. However, it is recommended to refine the oil immediately to preserve the quality of the oil. The traditional refining of salmon oil includes refining, bleaching and, deodorization, possibly followed by winterization, and addition of antioxidants [51,53,54,55,56]. Some of the work reported in the literature on the extraction of protein from salmon by-products are shown in Table 2.
6.3. Ultrasound-assisted enzymatic extraction (UAEE)
The ultrasound-assisted enzymatic extraction is the process of using ultrasound waves exceeding 20 kHz. The application can be divided into low power-high frequency (100 kHz-1 MHz) and high power-low frequency (0-100 kHz) ultrasound. Low power ultrasound is mainly used in medical diagnosis, whereas high power ultrasound is used to disrupt cell walls and membranes [57,58]. The ultrasound waves are mechanical waves that can be propagated by rarefactions and compression through solid, gas and liquid media. It acts by generating bubble cavitation in the biological matrix.
For example, Kangsanant et al. [68] studied the bioactivities of tilapia protein hydrolysates (TPI) extracted using ultrasound pretreatment and ultrasound-assisted enzymatic hydrolysis. The results from this study indicated that ultrasound-assisted enzymatic hydrolysis caused a reduction in the degree of hydrolysis ranging from 23 to 35% relative to that of the conventional process. The ultrasound-assisted enzymatic hydrolysis at 70 W produced the highest in-vitro antioxidant activity. The ultrasound-pretreated samples for 30 and 45 min produced the strongest nitric oxide inhibitory and antioxidative (macrophage cell lines) activities. In another study, Álvarez et al. [69] studied the acid/alkali isoelectric solubilization/ precipitation (ISP) of proteins from mackerel using ultrasound. The study indicated that traditional isoelectric solubilization/precipitation process generated a lower yield than conventional enzymatic hydrolysis. However, the application of ultrasound to alkaline extraction helped recover more than 95% of total protein from mackerel by-products. Moreover, Misir and Koral [70] studied the effect of ultrasound treatment on the structural, chemical, and functional properties of protein hydrolysate prepared from rainbow trout by-products and reported that the application of ultrasound for hydrolysis might cause unfolding of protein molecules, thus might result in e higher hydrophobic groups at the surface of molecules. This may in turn bring about interactions among these groups that might lead to larger aggregates, compared to smaller aggregates produced during conventional hydrolysis. The ultrasound hydrolysates also had significantly better foaming capacity, foaming stability, and oil-binding capacity as well as a higher antioxidant activity. On the other hand, Tian et al. [71] combined high-intensity ultrasound with an alkaline pH-shift process to develop TPI under various pH conditions. The ultrasound application resulted in a significant improvement in the consistency, protein solubility and sediment size during the pH shift process. The ultrasound improved the yield at lower pH (10.5) than the traditional alkaline pH process (pH 11.5).
6.4. High-pressure processing-enzymatic hydrolysis
High-pressure processing was first used in 1899 on a laboratory scale to destroy microbial contamination in milk and improve its shelf-life. During high-pressure processing, uniform pressure (100-1000 MPa) is applied instantaneously, independent of the size and geometry of food, on flexible packaging materials filled with liquid or solid food products. The pressure generation fluid is water, and the process can be carried out with or without heat [72,73,74]. During high-pressure processing, proteins are denatured where weak non-covalent chemical bonds such as hydrogen, hydrophobic, and ionic bonds are broken. This process is different from temperature or chemical denaturation, in which non-covalent bonds are broken, and the proteins are irreversibly unfolded and aggregated. Therefore, high-pressure processing modulates protein-protein and protein-solvent interactions. Proteins treated under high-pressure induce the formation of monomeric, oligomeric, and aggregated products without adding chemicals or high temperatures. Upon low-pressure treatments (< 400 MPa), the number of hydrogen bonds is increased, whereas, under high-pressure treatments (> 400 MPa), it is broken. The structural modifications caused by high-pressure treatment may be reversible or irreversible depending on the pressurization parameters and the protein properties [74,75]. Proteins treated under high-pressure have profound effects on both intramolecular and intermolecular interactions. The intramolecular interactions include ionic interactions, hydration, hydrophobic interactions, hydrogen bonds, van der Waals forces, and covalent bonds. The intermolecular interactions include protein-protein interactions, enzyme-substrate interactions, and protein-non protein interactions [76].
Hydration of proteins plays a significant role in the high-pressure denaturation process. During high-pressure processing, the water fills crevices between amino acids, screens out repulsive forces, and facilitates side chains and polypeptide backbones. Water penetration under pressure leads to conformational transitions resulting in protein unfolding. At this stage, pressure induces protein to adopt molten globule conformation, a compact, partially folded conformation without a specific tertiary structure. The modification of protein due to hydration under high-pressure can be due to two factors. First, the opening of cavities allows a solvent to occupy an internal volume. Second, the surface area in contact with solvent is larger for unfolded proteins than native proteins [74,75,76,77,78]. The hydrophobic interactions in protein induce non-polar chains to cluster inside proteins and significantly affect proper protein folding. Water molecules surrounding the non-polar groups have higher compressibility than both hydrophilic hydration and bulk water. Therefore, under high pressure, the hydrophobic core of globular protein is exposed to the solvent, resulting in decreased system volume. This effect results in the unfolding/denaturing process. The volume change reveals polar and non-polar groups, electrostriction, and elimination of cavities [76,79].
Under high-pressure, the hydrogen bonds present within protein shorten in length leading to the collapse of internal cavities. The collapse of cavities contributes to the compression of protein under pressure [76]. The thermostability of proteins is enhanced under high-pressure when the denatured state is less compressed than the native state. During high-pressure processing, hydrogen bond formation in the protein is promoted, and van der Waals forces are favored to maximize the packing density and reduce the protein volume. The phenomenon of protein stabilization under high-pressure is due to the opposing effects of pressure and temperature on hydrophobic interactions and hydrogen bond formation [80]. Furthermore, low-contact-order hydrogen bonds (helical turns or beta hairpins) hidden between amino acids and close together in the polypeptide sequence are moderately affected by temperature. However, high-contact-order hydrogen bonds have larger sequence separations and are less stable against pressure and temperature changes [81].
Alemán et al. [82] studied the enzymatic hydrolysis of catfish skin gelatin under atmospheric pressure and high pressure (100, 200, and 300 MPa for 15 and 30 min) using Alcalase at 50°C, as well as collagenase, trypsin, and pepsin at 37°C. This study's results indicated an enhanced degree of hydrolysis for the samples treated with enzyme under high-pressure. The antioxidant activities evaluated by ferric reducing power (FRAP) and ABTS radical scavenging ability were improved for all hydrolysates treated under high-pressure. Moreover, Hemker et al. [83] studied high-pressure assisted enzymatic hydrolysis on tilapia by-products to produce protein hydrolysates. The results from this study indicated that pressure and holding time influenced the soluble protein content which was increased with increasing pressure and holding time. The underlying mechanism is attributed to the activation of the enzyme and unfolding of protein substrate for the improved results under high-pressure. The UV-spectra of high-pressure treated protein hydrolysates exhibited significant changes in peak wavelengths and absorbance intensity. The study also showed that increasing pressure increased the release of soluble proteins and free amino acids, contributing to higher absorbance intensity. Under high-pressure, the protein unfolding occurs that exposes hydrophobic amino acid residues like tyrosine and tryptophan. The pressure-treated protein hydrolysate had improved solubility and emulsifying properties. The high-pressure treated protein hydrolysate had improved antioxidant capabilities compared to hydrolysates treated under atmospheric pressure.
7. Composition of Salmon Protein Hydrolysate
Several studies reported that the salmon protein hydrolysates produced from various parts of salmon by-products consisted of 69 to 89% protein, 0.06 to 16% lipids, 3.5 to 22% ash, and 0.9 to 5% moisture. The amount of protein and lipid are dependent on the type of salmon byproduct used in the hydrolysis. The high amounts of ash in the final protein hydrolysate are primarily due to added acid or base to adjust the pH. The final moisture content in the salmon protein hydrolysate is dependent on the type of sample and the temperatures employed during the drying process [2,8,16,59,60,66].
Salmon protein hydrolysates obtained after enzymatic hydrolysis are dominated by amino acids and short-chain peptides. The amino acid composition of salmon protein hydrolysates reported in various studies is shown in Table 3. There were significant differences in the amino acid composition of the protein hydrolysates obtained from the same species. The type of raw material used in the study plays an important role in the amino acid composition of the protein hydrolysate. Other factors which influence the amino acid composition include the type of enzyme and hydrolysis conditions. However, most of the salmon protein hydrolysates contained all the essential and non-essential amino acids and may serve as good sources of functional food ingredients [2,16,61].
7.1. Functional properties of salmon protein hydrolysate
The functional properties of salmon protein such as foaming, emulsification, water-holding capacities, solubility, and gelation are affected by the source of raw material, environmental, production, and processing factors. The production and processing factors include isolation, precipitation, drying or dehydration, concentration, enzymatic or chemical modification, and the environmental factors that include temperature, pH and ionic strength [10].
Enzymatic hydrolysis of salmon protein generates a mixture of amino acids, di-, tri-, and oligopeptides, increasing the number of polar groups and the hydrolysate's solubility. The choice of substrate and enzymes and degree of hydrolysis affect the resulting hydrolysate's physicochemical properties. The enzyme specificity strongly influences the molecular size and hydrophobicity of the hydrolysate. The control of the enzyme hydrolysis is essential; uncontrolled or prolonged hydrolysis may result in highly soluble peptides. The peptides produced in the uncontrolled reaction system totally lack the functional properties of the parent protein and may lead to undesirable bitter peptides. The physical and chemical properties that govern protein functionality include size, shape, amino acid composition and sequence, net charge and distribution of charges, hydrophobicity/hydrophilicity ratio, peptide structures, molecular flexibility, and ability to react with other components in a food system [47,86].
7.1.1. Solubility of proteins
The solubility of proteins is the most crucial functional property during developing and testing of new protein ingredients. Based on their solubility, fish muscle proteins, namelysarcoplasmic (most soluble), myofibrillar (soluble in dilute salt solutions), and stroma proteins (least soluble) may be affected. The solubility of proteins influences many other functional properties, such as emulsification, water-holding capacity, and foaming properties. Therefore, protein solubility is an excellent indicator of protein hydrolysate functionality and its potential applications. Protein solubility is influenced by amino acid composition and sequence, molecular weight and conformation, and the content of polar and non-polar groups in the amino acids. Protein solubility is also affected by environmental factors such as pH, temperature, ionic strength, type of solvent and processing conditions. In addition, hydrophobic and ionic interactions are significant factors that influence the solubility of proteins. The hydrophobic interactions are responsible for protein-protein interactions, resulting in decreased solubility, but ionic interactions are responsible for protein-water interactions and increased solubility [9,87,88].
The salmon protein is mainly composed of myofibrillar proteins with myosin and actin as its main components. The myofibrillar proteins are soluble in high ionic strength solutions [9]. Treatment of fish muscles with NaCl results in water uptake and lateral expansion of myofibrillar lattice. The amount of swelling is dependent on ionic strength. When the ionic strength is lowered or increased relative to the physiological value of 0.2 the swelling of fish muscle will decrease or increase, respectively due to the osmotic pressure [89]. Various researchers have worked on extracting fish protein using different salts such as sodium chloride and potassium chloride. For example, Munasinghe and Sakai [90] extracted proteins from adult yellowtail (Seriola quinqueradiata) using sodium chloride and potassium chloride. The protein extractability with sodium chloride and potassium chloride decreased when the pH changed from 6.0 to 8.0. The sodium chloride treated samples had the highest protein extractability at pH 6.0 and ionic strength of 0.8. Furthermore, Kim and Park [30] suggested that the presence of salt in the protein isolates adversely affected its gelation properties and promoted the salting-out effect.
Therefore, to increase the solubility of myofibrillar proteins in Atlantic salmon, enzymatic hydrolysis is recommended. Kristinsson and Rasco [9] studied the functional properties of Atlantic salmon muscle proteins hydrolyzed with various alkaline proteases. The results showed that alkaline proteases (Alcalase, Flavourzyme, Corolase PN-L, Corolase 7089, and endogenous extract) produced 92 to 99% soluble protein hydrolysates depending on the degree of hydrolysis. The results also indicated that the solubility of the hydrolysates was very high (above 90%) at pH 7.0 and 0.1 M NaCl. Electrophoresis results showed that the hydrolysates reached maximum solubility at 5% DH and remained unchanged at higher DH. The relationship between solubility and DH is due to the decrease in peptide size produced at higher DH. The solubility is also believed to be the delicate balance of hydrophilic and hydrophobic forces of the peptides. Gbogouri et al. [59] studied the influence of the degree of hydrolysis on the solubility of salmon byproducts hydrolysates extracted using Alcalase and reported higher solubility (above 90%) of hydrolysates at pH 6 to 7 while lower solubility at pH 3 to 4. The pH influences the charge on the weak acidic and basic side-chain groups. Therefore, protein hydrolysates display low solubility at their isoelectric point. In addition, Abdollahi and Undeland [51] suggested that the protein solubility of salmon by-products was highest at solubilization pH of 12.5. Usually, upon increasing the pH, the electrostatic charges of the proteins are improved, which increases their interaction capacity with the water (solubility), resulting in increased protein yield of the process. Similarly, a higher solubility (95%) of salmon head and backbone was observed at pH low and high pH, which could be relate to surface characteristics of their amino acids, conformational situation, and molecular weight [86].
7.1.2. Water holding capacity (WHC)
The water holding capacity of proteins is defined as the ability to maintain or imbibe water during the application of forces, pressing, centrifugation, or heating. Protein's water holding capacity is essential in the food industry to improve the texture. The hydration properties of proteins determine their applications in food systems. This functional property is dependent on the water-protein interaction which determines the functional properties of proteins such as water binding and retention, swelling, solubility, emulsifying properties, viscosity, gelation, and syneresis. The interaction of protein and water depends on the composition and conformation of the protein molecules. The interactions between water molecules and hydrophilic groups of protein side chains occur via hydrogen bonding. Proteins containing a high percentage of charged amino acids can bind large amounts of water. Therefore, the water holding capacity can be predicted from the amino acid composition of proteins [87,88,91].
Proteins with high water holding capacity can degrade other ingredients in the food system. Proteins with low water holding capacity are sensitive to storage humidity. Therefore, the choice of proteins with an appropriate water holding capacity is essential in food formulation. The recommended maximum water content of fish protein hydrolysate for storage is 0.075 g/g at less than 15% RH. Salmon protein hydrolysates are highly hygroscopic, and therefore proper packaging and low relative humidity of air during processing is an important consideration. Polar groups such as COOH and NH2 that increase during enzymatic hydrolysis substantially affect water and moisture absorption isotherm for fish protein [47].
Kristinsson and Rasco [9] studied the water holding capacity of salmon protein hydrolysates and reported that the degree of hydrolysis did not affect the property. However, the type of enzyme used for hydrolysis differentiated the water holding capacity of salmon protein. Alcalase hydrolyzed samples have the highest water holding effect compared to other alkaline proteases compared in the study. Samples hydrolyzed using Alcalase also produced the highest concentration of low-molecular-weight peptides than other enzymes. Corolase 7089 also produced similar concentrations of low-molecular-weight peptides as Alcalase; however, the water holding capacity was the lowest. This phenomenon is due to the type of peptides produced by different enzymes as per their specificities. Therefore, the selection of enzymes plays an important role in the production of functional protein hydrolysates. In addition, salmon proteins that recovered using the pH-shift method had a strong gel-forming capacity [51,86]. However, cod protein isolates showed a higher WHC than salmon protein and this could be due to a higher amount of lipids present in the salmon.
7.1.3. Emulsifying properties
The emulsifying properties of proteins are evaluated by their ability to take part in emulsion formation and stabilize the newly created emulsion. Proteins are major components in most food emulsions [87]. During homogenization at high speeds, protein adsorb to the surface of newly formed oil droplets and form a protective membrane that prevents droplets from coalescing [47]. Dried protein hydrolysates are surface-active molecules and possess good emulsifying properties due to the presence of both hydrophobic and hydrophilic amino acids. The surface-active nature of protein is an important property for its interfacial film-forming capacity [92]. Food emulsions are oil-in-water (O/W) and water-in-oil (W/O). When protein molecules are added to the oil in water emulsions, they migrate to the oil-water interface and realign to penetrate the oil droplets with their hydrophobic groups. In contrast, the hydrophilic groups are aligned with the aqueous phase. Following alignment, a robust viscoelastic film is formed around the oil droplets. The protein-rich interfacial layers protect the emulsion against strong destabilization processes by acting as an electrostatic, structural and mechanical energy barrier [9,59,93]. The emulsifying properties of proteins are described by emulsifying capacity (EC), emulsifying stability (ES) and emulsifying activity (EA). The emulsifying capacity (EC) is defined as the amount of oil (mL) emulsified by the protein hydrolysate (g) before the phase inversion occurs. The emulsification stability (ES) is determined by the percentage decrease in the emulsion's interfacial area, or the percentage volume of cream separated from the emulsion. The emulsifying activity (EA) is calculated as the interfacial area created per unit mass of protein [9,86,88,94]. The protein stabilized emulsions are affected by several factors such as pH, ionic strength, temperature, presence of low molecular-weight surfactants, sugars, oil phase volume, type of protein, and the melting point of the oil used during the process. Extrinsic factors such as type of equipment, energy input rate, and shear rate also the protein stabilized emulsions [94].
Gbogouri et al. [59] studied the emulsification capacity and stability of emulsions created using salmon protein hydrolysates. The study reported that the emulsification capacity varied between 0.02 and 0.2%. The results also showed that the emulsification capacity increased with increased protein concentration until reaching the maximum emulsification capacity and then decreased. The decrease of emulsification capacity was due to high protein concentration increasing the adsorption rate higher than the spreading rate. Moreover, Kristinsson and Rasco [9] studied the emulsification properties of salmon protein extracted using alkaline proteases such as Alcalase, Flavourzyme, Corolase PN-L, Corolase 7089, and endogenous extract. The study indicated that salmon protein hydrolysates produced using endogenous extract had the highest emulsification capacity at all degree of hydrolysis than other hydrolysates. The changes in the degree of hydrolysis affected the emulsification capacity of protein. They also reported a positive correlation between the peptide length and surface activity, and a minimum of 20 amino acid residues had good emulsifying and interfacial properties. The endogenous enzymes in producing salmon protein extracts retained large peptides (~2000 Da) at all degrees of hydrolysis, and it was responsible for the high emulsifying activity. The peptides released during Flavourzyme hydrolysis were similar in size to endogenous extract hydrolysis. However, Flavourzyme hydrolysates had poor emulsification capacities. Therefore, more than the peptide size, the physicochemical makeup of the peptides may play an important role in the difference in the emulsification capacity of protein. Large differences in emulsifications capacities were also observed between different hydrolysates at the same degree of hydrolysis. This property is due to the differences in enzyme specificity and hydrophobicity. Similarly, He et al. [95] studied the emulsifying capacity of salmon protein hydrolysates using Alcalase, Flavourzyme, and Neutrase and found that the emulsifying capacity decreased with longer processing time and a higher enzyme-to-substrate ratio. This trend is attributable to low molecular weight peptides produced at a higher enzyme-to-substrate ratio and increased reaction time. The low molecular weight peptides are less efficient in decreasing the water-oil interface tension because they cannot unfold and reorient on the water-oil surface to stabilize the emulsion system. Flavourzyme hydrolyzed salmon produced the highest emulsifying capacity (51 ± 1.8 m2/ g) at a 0.5% enzyme-to-substrate ratio and 30 min reaction time.
7.1.4. Fat absorption capacity
The capacity of protein hydrolysates to absorb fat is an important attribute that influences the taste and texture of food. Fat absorption of proteins is affected by protein source, processing conditions, presence and composition of additives, particle size, and temperature. Fat absorption is a form of physical entrapment of oil by proteins. Low-density protein powder with small particle size entraps more oil than high-density protein powders. Insoluble and hydrophobic proteins have high oil absorption capacity. Oxidized lipids accelerate lipid-protein interactions. The proteins act as a trap for lipid peroxides and secondary lipid oxidation products. The breakdown products of oxidations can interact with terminal functional groups of amino acids in proteins and enzymes. These interactions influence the functional and nutritional properties of proteins and flavor. Four types of bonds involved in the protein-lipid interactions are hydrophobic, electrostatic, hydrogen, and non-covalent bonds [19,87,88].
Gbogouri et al. [59] studied the fat absorption capacity of protein hydrolysates prepared using Alcalase and reported that the best corn oil absorption was achieved at a degree of hydrolysis (DH) of 11.5%. The fat absorption capacity decreased with an increase in the DH, possibly due to the physical entrapment of oil and, thus, the higher bulk density of protein. On the other hand, Kristinsson and Rasco [9] studied the fat absorption properties of salmon protein extracted using alkaline proteases such as Alcalase, Flavourzyme, Corolase PN-L, Corolase 7089, and endogenous extract. This study also reported higher fat absorption at a 5% degree of hydrolysis for all extracts, but only limited research has been conducted on the fat absorption of fish protein hydrolysates. The enzyme-substrate specificity also plays a major role in the fat-binding capacity of hydrolysates.
7.2. Sensory properties of salmon protein hydrolysate
The sensory properties, both taste and odor, of salmon protein hydrolysates should be acceptable to the consumers alongside their quality and functional properties. Significant problems associated with protein hydrolysates are the bitterness and fishy odor of the final product. The protein hydrolysate's bitterness is related to various factors such as hydrophobic amino acids, degree of hydrolysis, molecular weight, enzyme type, and peptide sequences. The amino acids responsible for the bitter taste are valine, isoleucine, phenylalanine, tryptophan, leucine, and tyrosine. The hydrophobicity (Q) values of more than 1400 Cal/mole and molecular masses of less than 6 kDa exhibit bitterness [10,66,96,97,98,99]. Several methods have been tried to mask the bitterness of protein hydrolysates, including the addition of glutamic acid, glutamyl-rich peptides, polyphosphates, gelatin, or glycine in the products [99]. Recently, Xu et al. [100] proposed a mechanism in which adding sodium chloride decreased the surface hydrophobicity. The screening of Na+ reduced hydrophobic interactions between the protein hydrolysates and bitter taste receptors. The addition of sodium chloride creates a salting-in effect, causing the self-folding of the peptide and Cl- bind to the hydrophobic cavities and decrease the surface hydrophobicity. The study also indicated that the addition of sodium chloride decreased the particle size and turbidity of the protein hydrolysates. Specifically, plastein reaction is carried out as a strategy to reduce the bitterness of peptides and this is often carried out by the addition of glutamic acid, as it was explained earlier. Other measures might also be considered in this regard [101].
Aspevik et al. [102] studied the sensory properties of protein hydrolysates prepared from Atlantic salmon using Alcalase, Promod, and Protex enzymes at 17, 25 and 30% enzyme concentrations, respectively. The formation of bitter taste was attributed to both the degree of hydrolysis and enzyme specificity. The results indicated that protein hydrolysates obtained using Alcalase were more bitter and astringent than Promod and Protex hydrolysates. The subtilisin-activity of Alcalase releases more bitter-peptides from the salmon substrate than Protex and Promod enzymes. The study concluded that the degree of hydrolysis plays a significant role in several sensory attributes, such as bitter, umami, sea, fish, and pungent tastes and odors. The maximum intensity and threshold level of bitter taste were between medium to high based on the degree of hydrolysis. Similarly, Idowu et al. [66] suggested that the hydrolysate obtained from salmon frames using Alcalase yielded with higher bitterness than papain, which could be linked to hydrophobic groups toward their C-terminal. Furthermore, sensory attributes of protein isolate prepared from salmon (Salmo salar), herring (Clupea harengus), and cod (Gadus morhua) by-products using the pH-shift method was studied [86]. Herring and salmon proteins scored high compared to cod protein for sensorial characteristics related to off-flavor. This could be due to the lipid oxidation during protein isolation through the pH-shift process due to heme-protiens. Thus, extra precautions should be taken to prevent lipid oxidation for fatty fish-like salmon since oxidation products could play a part in the development of bitter taste [67]. In contrast, Aspevik et al. [67] suggested that salmon hydrolysates were linked with larger peptides (>2 kDa), low flavor intensity, and pleasant flavors, including umami taste and sea flavor.
8. Bioactive Peptides
Proteins available in foods are sources of energy and essential amino acids that contribute to the body's growth and maintenance. Amino acids present in proteins are necessary to synthesize a wide variety of proteins with essential functions, including carriers of oxygen, vitamins, CO2, enzymes, and structural proteins. Besides the nutritional aspect of proteins, they are also responsible for various physiochemical and sensory properties of foods and may act as functional and health-promoting ingredients [2,103,104]. Most of the proteins' physiological and functional properties are attributed to the biologically active peptides encrypted in the protein molecules. Bioactive peptides are released from the parent protein source by the digestive enzymes (gastrointestinal digestion), food processing (ripening, fermentation and cooking), storage or by in-vitro enzyme hydrolysis [104,105].
8.1. Empirical production of bioactive peptides and their functions
The bioactive peptides can be produced from protein sources using different methods, including enzymatic hydrolysis with digestive enzymes, enzymatic hydrolysis using proteases, fermentation, microwave-assisted hydrolysis and other techniques, as mentioned earlier. Enzymatic hydrolysis is the most popular one for producing protein hydrolysates from marine sources. Crude protein hydrolysates are subjected to various assays to screen for bioactivities. After detecting bioactivities, crude protein hydrolysates are fractionated based on peptide size via ultrafiltration. The fraction showing the highest bioactivity is further purified using various chromatographic techniques. The isolated peptides are then sequenced using tandem mass spectrometry and protein sequencing. Finally, based on the peptide sequence, peptides are synthesized, and the assays are repeated to confirm the bioactivities [106,107]. The production of bioactive peptides is shown in Figure 7. Some of the peptides extracted from Atlantic salmon reported in the literature are given in Table 4.
Several studies have been conducted on the bioactive peptides of Atlantic salmon and their beneficial action in health promotion, including angiotensin-converting enzyme (ACE) inhibitory activity. ACE inhibitors could help to relax the arteries and veins, resulting in lower blood pressure. For example, Neves et al. [18] extracted bioactive peptides from salmon trimmings with ACE inhibitory and dipeptidyl peptidase IV (DPP-IV) inhibitory activities as well as, oxygen radical absorbance capacity (ORAC). Salmon protein procured from trimmings was isolated using the pH shift method and hydrolyzed with Alcalase, Flavourzyme, Promod, and Corolase PP. The IC50 values for ACE inhibitory activities of hydrolysed salmon trimming protein ranged between 0.74 and 1.69 mg/mL. Alcalase treated hydrolysates generated a more potent ACE inhibitory peptide with an IC50 value of 0.74 mg/mL. Promod hydrolysates produced the least potent ACE inhibitory peptide with an IC50 value of 1.69 mg/mL. The IC50 values for DPP-IV inhibitory activities extracted from salmon trimming protein ranged between 0.30 and 2.37 mg/mL. Corolase PP-treated hydrolysates generated a more potent DPP-IV inhibitory peptide with an IC50 value of 0.30 mg/mL. Promod hydrolysates produced the least potent DPP-IV inhibitory peptide with an IC50 value of 2.37 mg/mL. Similarly, Neves et al. [17] extracted gelatin from salmon trimming protein and hydrolyzed it with Alcalase, Alcalase+Flavourzyme, Corolase PP, Promod, and Brewer’s Clarex. There were distinct differences in the peptide profiles of hydrolysates generated with different enzymes used. The increase in reaction time generated more low molecular weight peptides using all enzymes. More potent ACE inhibitory hydrolysates were generated using Corolase PP and Alcalase+Flavourzyme with IC50 values of 0.13 and 0.28 mg/mL, respectively. More potent DPP-IV inhibitory hydrolysates were generated using Corolase PP and Alcalase+Flavourzyme with IC50 values of 0.08 and 0.10 mg/mL, respectively. The ORAC values of antioxidant peptides extracted from salmon gelatin hydrolysates ranged between 103 and 540.94 µmol Trolox/g. The hydrolysates extracted using Corolase PP and Promod generated the most antioxidant peptides based on ORAC values. The salmon gelatin hydrolysate extracted using Corolase PP was used for further analysis. The purified Corolase salmon gelatin hydrolysate was administered to spontaneously hypertensive rats (SHR). The in vivo study indicated improved mean arterial blood pressure (MAP), systolic (SBP) and diastolic blood pressure (DBP), and decreased the heart rate (HR) of SHR compared to synthetic drug CaptoprilTM. In addition, the ORAC values of antioxidant peptides extracted using Promod ranged from 587.41 to 882.58 µmol Trolox/g. Peptides generated by Alcalase, Alcalase+Flavourzyme, and Corolase PP had ORAC values higher than 601.47 µmol Trolox equivalents/g. In another study, peptides derived from salmon demonstrate strong ACE inhibition due to their high affinity to ACE active sites. In particular, peptides with molecular weight (>1500 Da), C terminal peptide sequence, short chain length, hydrophobic amino acids, and presence of lysine or arginine at C end exhibited higher ACE inhibitory activity [108].
Ahn et al. [109] identified ACE inhibitory peptides from salmon protein by-product via enzymatic hydrolysis using Alcalase, Flavourzyme, Neutrase, pepsin, Protamex, and trypsin. The results indicated that Alcalase hydrolysates possessed the highest ACE inhibitory activity. The chromatographically purified ACE inhibitory peptides were VWDPPKFD (P1), FEDYVPLSCF (P2) and FNVPLYE (P4) and their IC50 values were 9.10, 10.77 and 7.72 µM, respectively. The study also indicated that the VWDPPKFD peptide was a non-competitive inhibitor, and FEDYVPLSCF and FNVPLYE had mixed inhibition modes. The presence of phenylalanine, leucine and tyrosine at the C-terminal appears to play an important role in their inhibition activity. In another study, low-molecular-weight ACE inhibitory peptides were isolated from Atlantic salmon skin by Gu et al. [110]. The peptides had a high quantity (90.79%) of oligopeptides below 1 kDa, with most of them in the range of 132-576 Da (69%). The filtered salmon peptides had an IC50 of 1.165 mg/mL. To better understand and identify the peptides, the samples were subjected to RP-HPLC, and 11 different fractions were obtained. Two fractions out of 11 had high ACE inhibitory activities. A total of eleven peptides were identified from the two high ACE fractions, including three dipeptides, one tripeptide, five tetrapeptides, and two pentapeptides. Two dipeptides, AP and VR, exhibited an IC50 of 0.06 and 0.332 mg/mL ACE inhibition, respectively, which is 20 and 40-folds more potent than filtered salmon peptides (1.165 mg/mL).
Slizyte et al. [62] screened defatted salmon backbone protein hydrolysates using Corolase PP, Corolase 7089, papain, bromelain, Protex, Seabzyme L200, and trypsin for bioactivities. The highest DPPH radical scavenging activity was obtained from Protamex hydrolysates without oil separation before hydrolysis, followed by Corolase hydrolysates. The iron-chelating activities ranged between 54 and 87% for all hydrolysates. Bromelain+papain hydrolysates after 20 min had the best iron-chelating ability indicating that larger peptides have better ability to chelate iron, and this property weakens when the peptide size is reduced. Trypsin hydrolysates had the highest ACE inhibitory activity (IC50 = 0.9 mg/mL) after 120 min hydrolysis. Bromelain+papain hydrolysates (8 mg/mL) had the most significant glucose transport inhibiting activity (39% reduction) than other enzymes. The 250-300 Da dipeptides are reported to have a role in the regulation of glucose uptake. On the other hand, Picot et al. [116] studied the antiproliferative activity of fish protein hydrolysates against human breast cancer cell lines. Seven different types of fishes, including blue whiting, Atlantic cod, Atlantic salmon, Atlantic emporer, pollack, plaice, and Portuguese dogfish was used to produce protein hydrolysates. The protein hydrolysates from blue whiting (35 and 28%), cod (40 and 20%), plaice (35 and 30%) and salmon (25 and 18%) exhibited significant growth inhibition relative to control against two human breast carcinoma cell lines, MCF-7/6 and MDA-MB-231 cells grown in vitro. Moreover, antioxidant and anti-inflammatory properties of peptide fractions extracted from salmon by-products were studied by Ahn et al. [117]. Salmon byproduct protein from pectoral fin was used for enzymatic hydrolysis. All salmon protein hydrolysates exhibited potent DPPH activities in a dose-dependent manner. The purified pepsin-hydrolysate showed high DPPH scavenging activity (73% at 4 mg/mL) and hydrogen peroxide scavenging activity (87% at 4 mg/mL). The salmon hydrolysate was tested for inhibiting intracellular reactive oxygen species (ROS) and lipid peroxidation and its effect on glutathione (GSH) level in Chang liver cells. The pepsin hydrolysate decreased intracellular ROS generation by 2.14-fold compared to the control group. It enhanced the GSH level in Chang liver cells in a time and dose-dependent manner at a concentration of 1 mg/mL. The pepsin hydrolysate also inhibited LPS-induced nitric oxide (NO) production (3.61 times) in a concentration-dependent manner with the highest reduction at 400 µg/mL. The pepsin hydrolysate also inhibited proinflammatory cytokine production, including TNF-α (200 µg/mL), IL-6 (200 µg/mL), and IL-1β (200 µg/mL) in RAW264.7 macrophage cells in a concentration-dependent manner. In a follow-up study, Ahn et al. [111] purified and studied the anti-inflammatory action of a tripeptide from salmon pectoral fin by-product protein hydrolysate. The purified anti-inflammatory peptide was identified as PAY, and it significantly inhibited the production of nitric oxide (NO) by 63% and prostaglandin E2 (PGE2) by 45.33% in RAW264.7 cells at 0.75 mM concentration. The PAY tripeptide also inhibited LPS-stimulated TNF-α, IL-6, and IL-1β production in RAW264.7 macrophage cells at 0.75 mM concentration.
Li-Chan [112] studied the dipeptidyl-peptidase IV (DPP-IV)-inhibitory activity of peptides derived from Atlantic salmon skin gelatin hydrolyzed by Alcalase, Flavourzyme, and bromelain The Flavourzyme hydrolysate extracted at 6% enzyme/substrate concentration exhibited the DPP-IV highest inhibition rate of 45.2%, followed by Alcalase (30%) and Bromelain (23.1%). The peptides obtained within the <1 kDa ultrafiltrate fraction exhibited the highest DDP-IV inhibition rate of 61.2% whereas, the >2.5 and 1-2.5 kDa fractions had 29.6 and 43.2% inhibition rates, respectively. The IC50 value of <1 kDa fraction was 1.35 mg/mL. The <1 kDa fraction was purified, and DPP-IV inhibitory activities were studied at a concentration of 100 µg solid/mL. The purified fraction had an IC50 value of 57.3 µg/mL. The peptides identified in the purified fraction were GPAE (372.4 Da) and GPGA (300.4 Da). The IC50 values of the two synthetic peptides, GPAE and GPGA, were 49.6 and 41.9 μM, respectively. Moreover, Jin et al. [113] identified dipeptidyl-peptidase IV (DPP-IV)-inhibitory peptides from salmon skin collagen hydrolysate. The ultrafiltered sample analysis indicated that < 3kDa fraction had the highest DPP-IV inhibitory activity with an IC50 value of 1.54 mg/mL. The study reported three DPP-IV inhibitory peptides out of 24 identified peptides, including YYGYTGAFR, LDKVFR, and VLATSGPG, with IC50 values of 1.21, 0.10, and 0.18 mg/mL, respectively. The study reported a novel hexapeptide (LDKVFR), and it also had the highest DPP-IV inhibitory activity. The molecular docking studies also revealed that six hydrogen bonds and eight hydrophobic interactions between LDKVFR and DPP-IV contributed to DPP-IV inhibition.
Atlantic salmon peptides have been reported to exhibit strong antioxidant potential. For instance, Wang et al. [114] studied the purification and characterization of antioxidant peptides from salmon protamine hydrolysate. Protamine is derived from fish milt, and it is usually discarded as an industrial by-product in fish plants The salmon protamine hydrolysate was fractionated using size exclusion chromatography, and various fractions were analyzed for scavenging activity on hydroxyl radical, DPPH radical, and superoxide radical. The highest hydroxyl radical scavenging activity peptide (PR) had an IC50 value of 91.3 µg/mL. Likewise, Girgih et al. [118] studied the antioxidant properties of peptides extracted from salmon frame protein hydrolysates that were hydrolyzed sequentially with pepsin and trypsin+chymotrypsin, and the resulting hydrolysate was ultrafiltered through < 1 kDa membrane. Later, ultrafiltered protein hydrolysate was separated using reverse-phase HPLC into four peptide fractions (1-4). The results from this study indicated that fractions 2-4 exhibited higher ORAC values between 1315 and 1541 µM Trolox equivalent (TE)/g compared to non-fractionated protein hydrolysate (819.3 µM TE/g). A similar trend was seen for DPPH and superoxide radical scavenging activities. However, the non-fractionated salmon protein hydrolysates had higher metal chelating activity than the peptide fractions. The peptide fractions also strongly inhibited linoleic acid oxidation. In addition, the pressurized liquid extraction (PLE) technique was used to prepare antioxidant peptides from salmon muscle remains, skin, heads, viscera, and tailfins and found that both ORAC and Trolox equivalent antioxidant capacity (TEAC) assays exhibited strong antioxidant activity, mainly those were prepared from viscera [119]. Hydrophobic amino acids, including alanine, proline, leucine, and valine, in salmon by-products could play an important role in showing antioxidant activity. Moreover, Hanachi et al. [120] isolated salmon head peptides using membrane filtration, which exhibited strong ABTS and DPPH radical scavenging activities, ORAC, and metal chelation activity (MCA), as well as ACE inhibitory activity, with an IC50 value of 413.43 ± 13.12 µg/mL. The MCA could be linked to the presence of histidine and glutamic acid in salmon head, providing more carboxylic groups and imidazole rings to bioactive peptides and increasing the electrostatic interaction with Fe2+ ions. Likewise, Rajendran et al. [121] developed protein hydrolysates from Atlantic salmon processing waste (viscera) using lactic acid fermentation with the formic acid treatment and Flavourzyme and found that they showed higher metal chelation and ferric reducing capacities. On the other hand, anti-allergic peptides were isolated from Atlantic salmon byproducts using sephadex G-15 gel permeation chromatography, HPLC, and mass spectrometry [122]. Result suggested that the isolated peptide (TPEVHIAVDKF) exerted excellent anti-allergic activity by inhibiting the release of β-hexosaminidase in immunoglobulin E (IgE)-mediated RBL-2H3 cell degranulation at IC50 value of 1.39 mg/mL.
8.2. In-silico production of bioactive peptides
The in-silico analysis involves computational methods applied to manage, curate, and interpret various biological systems. Protein databases such as UniProtKB, NCBI, and BIOPEP contain different protein sequences that can be utilized to analyze amino acid profiles of precursor proteins. Online tools such as BIOPEP and ExPASY-PeptideCutter can be used to select enzymes and proteins and predict theoretical bioactive peptide profiles. The in-silico hydrolysis results can be compared with the bioactive peptides reported in the literature and databases [123,124,125,126]. The in-silico approach for identifying and processing bioactive peptides is shown in Figure 8. The different databases and tools are given in Table 5.
Darewicz et al. [115] studied the ACE inhibitory peptides from salmon protein hydrolysates using three different methods such as in-silico, ex-vivo and in-vitro. In the in-silico analysis, the 52 amino acid sequences of salmon proteins were selected from the UniProt database. The proteolysis simulation using pepsin, trypsin and chymotrypsin was conducted using the procedure built into the BIOPEP database. The ex-vivo digestion was carried out using human gastric juice (HGJ) and human duodenal juice (HDJ). The in-vitro digestion was carried out using pepsin and Corolase PP. The in-silico digestion identified 11 ACE peptides from two types of salmon proteins (myofibrillar and sarcoplasmic). The two-step ex-vivo and in-vitro digestion were conducted to mimic the human digestion process. The results indicated that porcine enzymes more easily degraded salmon proteins than gastrointestinal enzymes. The sarcoplasmic proteins were broken down more easily than myofibrillar proteins. This study identified 9 and 7 peptides from ex-vivo and in-vitro hydrolysates, respectively, compared to 11 peptides identified in the in-silico analysis (Table 4). The study indicated that results generated by in-silico simulation of hydrolysis were not confirmed in the in-vitro studies due to the oversimplification assumed for the availability of all bonds susceptible to the enzyme in the polypeptide chain of the protein. Sometimes, incomplete data on the specificity of the enzyme can also lead to discrepancies [124].
Wang et al. [127] studied the potential of in-silico approach for predicting DPP-IV inhibitory activity, in-vitro, of protein hydrolysates. In this study, 294 edible protein sequences (80 animal and 214 plant sources) and five commercial proteases were analyzed in-silico. The authors hypothesized that protein hydrolysates with high contents of peptides having XP and XA might potentially be DPP-IV inhibitors. This study identified the frequency of specific amino acids by calculating the ratio of the number of peptides with proline, alanine, or proline+alanine as the second N-terminal residues to the total peptide fragments released by proteases. The in-silico analysis was carried out on all combinations of protein sources and proteases. Based on these results, fifteen protein and protease combinations were selected for in-vitro analysis. The parvalbumin-beta 2, Atlantic salmon (Salmo salar; BIOPEP ID 1739), have the most alanine content (19.4 alanine/100 amino acid residues) and was suggested to have great potential as the precursor of DPP-IV inhibitory peptides. Unfortunately, the salmon proteins were not chosen for further in-vitro studies. However, the in-vitro studies from other proteins indicated that the DPP-IV inhibition rate increased with the degree of hydrolysis and hydrolysis time. The correlation studies between DPP-IV inhibition rates vs frequency of proline, alanine or proline+alanine (A%) showed that the selection of proline+alanine during in-silico analysis showed a strong correlation with in-vitro DPP-IV inhibition rates. However, the in-silico selection of proline and alanine had a moderate and weak correlation with the in-vitro analysis. Similarly, Lacroix and Li-Chan [128] evaluated the potential of various dietary proteins as precursors of DPP-IV inhibitors by an in-silico approach. The study used three proteins from Atlantic salmon (actin-cytoplasmic 1, myosin regulatory light chain 2, and slow myosin heavy chain) and one protein from Chum salmon (Type 1 collagen alpha 2 chain) reported in the UniProt KB database. The in-silico study identified 499 fragments matching sequences known to present an inhibitory activity against the DPP-IV enzyme. The GP and PG were the most frequently occurring sequences and were mainly found in the collagen proteins due to the high contents of proline and glycine. Collagen from Atlantic salmon and Chum salmon had a DPP-IV peptides occurrence frequency value of 0.110 and 0.305, respectively. Unlike other protein sources investigated in this study, salmon collagens contained relatively high numbers of DPP-IV inhibitory tripeptides APG and GPA. The study concluded that salmon proteins were one of the best potential precursors of DPP-IV inhibitors. In contrast, the plant commodity oat was the least promising potential source of DPP-IV inhibitors. In addition, de la Fuente et al. [119] identified 67 peptides from salmon viscera and predicted their antioxidant activity using the BIOPEP-UMP database. Bioinformatics analysis exhibited few antioxidant small peptides encrypted in amino acid sequences, mainly glycine-alanine-alanine and glycine-alanine-alanine. Most of these predicted antioxidant peptides were di- and tri-peptides in nature.
8.3. Computational characterization of peptides
The peptides obtained from in-silico methods are subjected to computational models such as quantitative structure-activity relationships (QSAR) and quantitative structure-property relationships (QSPR). The QSAR methodology is used to describe the biological activity (φ) as a function of chemical structures (C) differentiated by molecular or physicochemical variables. In 1868, Crum-Brown and Fraser published the first equation for QSAR, as shown below [129,130].
φ = f(C)
The QSAR modelling starts with data collection and pre-processing. During this step, the bioactive peptide library is built by collating target peptide sequences for QSAR models. The data collected is transformed using smoothing, normalization and aggregation. The data are further divided into two data sets: training and test. The training data set is used to formulate the QSAR model, while the test data is used to validate its predictability and accuracy. The molecular descriptors and features are selected. In this process, the peptides are described using selected physicochemical or amino acid descriptors. When a large set of descriptors are obtained, the features are filtered using filter and wrapper methods to reduce the dataset horizontally. This technique removes collinearity between the descriptor pairs. The heuristic methods used to select features are based on multiple linear regression, and it discards constant values and removes descriptors for all structures containing no value. The QSAR model is constructed using various statistical methods. The statistical techniques are classified into regression-based, classification-based and machine learning techniques. Once the model is built, it is validated using the test data. The parameters used in the model should be interpretable. The mechanistic interpretation is used to understand the influence of descriptors in the predicted activity. The applicability domain analysis is used to know whether the built models can be used for any set of compounds. In some instances, confirmatory studies allow for external validation of the QSAR model by synthetically manufacturing peptides for robustness and bioactivity [131,132,133]. The schematic representation of QSAR modelling is shown in Figure 9.
Over the years, amino acid descriptors have played a vital role in the peptide's structural variation. Amino acid descriptors help the modelling and prediction of biological activity as a function of molecular structure. The properties of the entire peptide can be forecasted with amino acid descriptors. The two-dimensional (2D) amino acid descriptors include hydropathic scale, amino acid z-scales for hydrophilicity/hydrophobicity (z1), molecular size/bulkiness (z2) and electronic properties/charge (z3), molecular electronegativity edge vector (MEEV) scales, divided physiochemical property score (DPPS) descriptor, the vectors of hydrophobic, steric and electronic properties (VHSE) scales. The three-dimensional (3D) amino acid descriptors use isotropic surface area and electronic charge index (ISA-ECI) and molecular surface-weighted holistic invariant molecular (MS-WHIM) scales [123]. In QPSR studies, the physiochemical properties of chemical compounds are determined based on the molecular structure information. Physiochemical properties such as melting point, boiling point, stability, dielectric constant, diffusion coefficient, thermodynamic properties and hydrophobicity are tested for determining QSPR [133].
9. Purification and Separation of Peptides
The bioactive peptides are receiving increased interest to be produced commercially. The isolation and purification of bioactive peptides are essential for studying their physicochemical properties and evaluating the bioactive properties by in-vitro and in-vivo assays. The three main factors that affect the purification process include purity, cost of production and process time. Conventional purification of biological material involves various steps, including removing of the insolubles, isolation and concentration of products, purification, and polishing [134]. It is estimated that about 70% of production cost is for separation and purification processes alone. Therefore, the purification steps should be straightforward, simple, and involve minimum processing steps. The most important is to know about the target peptide's structure, function, and properties to set up a correct purification strategy. Traditional isolation and purification methods for peptides include selective precipitation, membrane filtration and chromatographic techniques. The most common technologies used in the separation and purification of peptides are shown in Figure 10 [10,123,134,135,136].
9.1. Selective precipitation
The selective precipitation processes such as ammonium sulfate precipitation, isoionic precipitation, salting out, and solvent extraction can be used before the separation process to remove protein, enzymes, and other crude extract components [103,134]. These processes can be used at all stages of purification, from bulk recovery to selective isolation and are easy to scale up. The precipitation process is a cheap, simple, high throughput process that can be utilized in continuous operations. However, some precipitation processes are not effective for some proteins and peptides. Hydrophobic peptides with low solubility in aqueous and organic solvents are difficult to separate by this process. The precipitation process also requires a subsequent cleaning step to remove the precipitation agent from the peptide of interest [123].
9.2. Membrane filtration
Membrane filtration uses selective barriers to transmit specific components while retaining other parts under pressure. The most common membrane filtration technologies include microfiltration (MF), ultrafiltration (UF), nanofiltration (NF) and reverse osmosis (RO). The microfiltration membranes filter particles and globules with pore sizes between 100 and 10,000 nm. The ultrafiltration membranes filter protein and peptides with pore sizes between 2 and100 nm. The nanofiltration membrane filters salt, solutes and amino acids with pore sizes between 0.5 and 2 nm. The reverse osmosis process is mainly used to filtrate monovalent ions (Na+, K+, Cl-) with pore sizes between 0.1 and 1 nm [134,137,138]. Membrane filtration processes like ultrafiltration and nanofiltration are the most common for bioactive peptide filtration and concentration. These processes have several advantages including, (a) high throughput and scalable for continuous operation, (b) maintains protein and peptide integrity, (c) improved enzyme yield and process productivity levels, (d) uses insoluble separating agent eliminating the use of any additives, (e) allows selective transport and sound separation and (f) can be performed at isothermic conditions and a fixed pH [123,137]. Some of the disadvantages of using membrane filtration process include (a) poor selectively while separating similar size peptides, (b) membrane fouling, (c) increased viscosity of the retentate and (d) low mass transfer rates for highly concentrated raw material [107,123]. In recent years, applying the external electrical field as an additional driving force to the pressure gradient is being investigated to improve the separation efficiency of charged bioactive molecules. Some of these combination membrane processes include electromembrane filtration (EMF) and electrodialysis with UF membrane (EMUF) [139].
Electromembrane filtration combines membrane filtration and electrophoresis mechanisms, making it more selective than membrane filtration and less costly than chromatograph. In addition, operating parameters such as type of membrane, electrical field strength, salination, and hydrolysate concentration can be manipulated to improve the product transfer and rate of separation [135]. Suwal et al. [140] extracted antioxidant peptides in a two-step process from rainbow trout protein hydrolysate using EMF. The use of EMF in this study led to the highest peptide migration rate and a significant improvement in peptide antioxidant activities.
Electrodialysis with ultrafiltration membranes (EUDF) combines the charge selectivity feature of electrodialysis and size separation capacity (molecular weight cut-off) of ultrafiltration. This process also allows selective and simultaneous separation of anionic and cationic peptides. A study reported that the EUDF process allowed a selective and simultaneous separation of anionic and cationic peptides from the snow crab by-products protein hydrolysates. Two anti-cancer peptides were obtained in this study using the EUDF process [141]. Roblet et al. [142] studied the separation of salmon frame bioactive peptides with antidiabetic effects using the EUDF process. In this study, two UF membranes and two ion-exchange membranes were used, and the researchers were able to obtain both anionic and cationic peptides. The results indicated that cationic peptides obtained using EUDF significantly enhanced the glucose uptake in L6 muscle cells grown in tissue culture.
9.3. Chromatographic techniques
Chromatographic techniques are considered the most powerful technique to isolate and purify bioactive peptides. The most used chromatographic techniques are reversed-phase, ion exchange, size exclusion and affinity chromatography. Some of the common advantages of using chromatographic techniques include (a) highly selective and offers high resolution in a short time, (b) an insoluble separating agent and (c) concentrate and stabilize target peptides. The disadvantages of using chromatography include (a) costly, especially during scale-up operations, (b) difficulty in treating viscous materials, (c) slow binding, (d) bead deformation during high pressure drops in scale-up operations, (e) solvent waste resulting environmental concern, (f) mass transfer limitation and steric hindrance can cause reduced capacity [123].
9.3.1. Ion-exchange chromatography (IEC)
The ion-exchange chromatography is based on the principle of the attraction of oppositely charged molecules. Each protein molecule carries a surface charge depending on pI and pH. Each ion exchange membrane is made up of porous beads, which assist in the adsorption of the target molecule. Generally, protein above their pI is negatively charged and can bind to oppositely charged anion exchange beds. Whereas protein below their pI is positively charged and can bind to oppositely charged cation exchange beds [107,134]. ACE inhibitory peptides were purified from salmon by-products using Alcalase hydrolysates and ion-exchange chromatography (DEAE FF ion-exchange column). The IEC fractions were monitored at 280 nm and investigated for the active ACE fraction [109]. After SEC purification of salmon protamine hydrolysate, the highest antioxidant activity fraction was purified for the second time using Macro-Prep High Q Support ion-exchange column. The IEC fractions were studied for antioxidant activity and were subjected to further purification steps [114].
9.3.2. Size Exclusion chromatography (SEC)
Size exclusion chromatography (SEC), also known as gel filtration chromatography (GFC), has been employed for the separation, desalting and molecular weight estimation of peptides and protein. The molecules fractionated using SEC do not bind to the chromatography medium, so the buffer composition does not directly affect the resolution [134,143]. The SEC is used primarily in the early stages of the purification of peptides. The SEC's resolution is influenced by particle size, particle uniformity, bed height, column packing quality, flow rate, sample concentration, and volume. Also, separating the peptide of interest with other closely related peptides is impossible [107].
ACE inhibitory peptides were purified from salmon by-product peptic hydrolysate using size exclusion chromatography (BioBasic SEC 60 column). The SEC fractions were monitored at 280 nm and investigated for active ACE fractions [117]. The size exclusion chromatography was used as a second purification step for purifying salmon ACE fraction, which was previously purified using IEC. The SEC purification (Sephadex G-25 gel filtration column) was monitored at 280 nm and pooled, and the ACE inhibitory activity was determined [109]. Wang et al. [114] studied the antioxidant capabilities of salmon protamine hydrolysate. In this study, protamine hydrolysate was fractionated using size exclusion chromatography on the Sephadex G-35 column. These fractions were studied for antioxidant activity and were subjected to a further purification process. Picot et al. [116] determined the molecular weight distributions of salmon protein hydrolysates using SEC in FPLC mode on a Superdex Peptides HR 10/30 column. Similarly, Slizyte et al. [62] used SEC to separate salmon peptides based on molecular weight using Superdex Peptides HR 10/300 column.
9.3.3. Reversed-phase high-performance liquid chromatography (RP-HPLC)
The reversed-phase high-performance liquid chromatography (RP-HPLC) is a widely used technique for fractionating peptides based on their hydrophobic properties. The RP-HPLC contains a stationary phase of lower polarity and a mobile phase of higher polarity. The main advantages of using RP-HPLC include ease of use, high resolution and sensitivity, shorter run time than SEC and IEC [107,134,143]. RP-HPLC was used as the third and final purification and fractionation of active salmon ACE inhibitory peptides after two prior purifications with IEC and SEC. The column used was ODS C18 (Hypersil Gold, 4.6 mm x 250 mm). The RP-HPLC purified salmon ACE inhibitory peptides were used for peptide sequencing and inhibition mode studies [109]. RP-HPLC was used as the third and final purification and fractionation of protamine peptides using the YMC-Pack Protein-RP column. The highest antioxidant activity RP-HPLC fraction was collected and used in further characterization studies [114].
10. Identification and Characterization of Peptides
HPLC as a separation technique together with identification tools such as tandem mass spectrometric detection (HPLC-MS/MS) is the standard method for characterizing peptide sequences. The tandem MS/MS contains two or three quadrupoles and a TOF analyzer. The analysis of peptides in MS/MS happens in two stages: (a) predetermined set of mass-to-charge (m/z) ions are separated from the rest of the ions from the ion source and fragmented via chemical reaction, and (b) mass spectra are produced for the fragments [107,134,143]. All peptides reported from Atlantic salmon shown in Table 4 were identified using tandem mass spectrometric detection [17,18,109,110,111,114,144].
11. Health Effects and Commercial Approval
In Canada, bioactive peptides, especially collagen peptides, are available in the market as natural health products (NHPs) for promoting skin health and other effects. The collagen peptides are sold under the Health Canada monograph as hydrolyzed collagen from bovine, porcine, chicken, fish, and shark (NPN: 80108792). ACE inhibitory peptides extracted from shrimp protein hydrolysate are also approved by Health Canada to be sold as a natural health product (NPN: 80080580). As per Health Canada approved claim, ACE inhibitory peptides help to maintain healthy blood pressure and support cardiovascular health. The beneficial effects of bioactive peptides as functional foods can be represented with disease risk reduction claims or physiological function claims [13,145].
The food health claims must be supported by substantial evidence from human studies. Health Canada assesses the substantiation of food health claims through ten guiding principles. The guiding principles include systematic approach, transparency, comprehensiveness, human evidence, high level of certainty, demonstration of causality, biological relevance of the claimed effect, feasibility of consumption of effective dose, health claim wording and substantiation of one food-health relationship in a submission. In-vitro studies from animal studies can be used to support the results of human studies [146,147]. Other essential regulatory requirements for food health claims include a summary of the physical, chemical, and microbiological properties of peptides, manufacturing steps of the bioactive peptides, stability data of the peptides under suggested storage conditions and the results of batch variability [13].
The necessary testing and research work for regulatory approval is very complex, time-consuming, and expensive. Most fish producers and processors operate in remote areas and are small-scale operations. It is challenging for small-scale producers and processors to take up such a complex process on their own. Some large-scale producers and processors are solely focused on producing and selling high-quality fish. Most of their revenue is from the edible portion of fish. At present, most of them are comfortable selling their by-products for cheaper returns as fish feed and fertilizer. However, with the proper foundational research work and testing, high quality bioactive peptides can be produced with considerable economic returns than feed or fertilizers.
12. Conclusion
Atlantic salmon and its by-products have proven to be an excellent repository of potent bioactive molecules such as protein, omega-3 oil, collagen, gelatin, peptides, enzymes, hydroxyapatite, and minerals. Biomolecules such as bioactive peptides display potent antioxidant, antihypertension, antimicrobial, anticoagulant and antidiabetic activities. However, these by-products are currently used mainly to produce low-value products such as fertilizer, silage, pet food and fish meal. There are many reasons, including the cost of production, human resources with high technical knowledge and skills to work in remote places of the country, the quality control process for by-products handling during processing and infrastructure such as additional cold storage facilities, especially for by-products can impede many processors and producers to be involved in the production of high-value products. However, these biomolecules present in Atlantic salmon are low-volume and high-value products. The transformation of the salmon processing by-products into high-value products can revitalize many rural fish communities, fish producers and processors. Therefore, it is essential to look at different processing techniques such as ultrasound and high-pressure to assist the production processes to save time and money with no compromise to quality. Processes like ultrasound and high-pressure processing are already available on an industrial scale. They have been proven to effectively reduce cost and increase the performance and quality of the final product. Furthermore, a critical impeding factor in marketing these biomolecules such as bioactive peptides to human health and nutrition requires in-vivo human studies. Most of the studies reported in the literature about bioactive peptides remain at the in-vitro cell culture stage and thus, extensive in-vivo animal studies are required in order to use them as functional ingredients.
Author Contributions
Conceptualization, V.V.R., A.H., D.D., and F.S.; software, V.V.R.; investigation, V.V.R., A.H., D.D., and F.S.; resources, D.D., and F.S.; data curation, V.V.R. and A.H.; writing—original draft preparation, V.V.R.; writing—review and editing, V.V.R., A.H., D.D., and F.S.; visualization, V.V.R., A.H.; supervision, D.D. and F.S.; project administration, D.D., and F.S.; funding acquisition, D.D. All authors have read and agreed to the published version of the manuscript
Funding
This research was funded by the Ocean Frontier Institute (OFI), through an award from the Canada First Research Excellence Fund.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dekkers, E.; Raghavan, S.; Kristinsson, H.G.; Marshall, M.R. Oxidative stability of mahi mahi red muscle dipped in tilapia protein hydrolysates. Food Chem. 2011, 124, 640–645. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Dinesh, K.B.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.R.; Newton, R.W.; Tlusty, M.; Little, D.C. The rise of aquaculture by-products: Increasing food production, value, and sustainability through strategic utilisation. Mar. Policy 2018, 90, 115–124. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish waste: From problem to valuable resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef]
- Ghaly, A.E.; Ramakrishnan, V.V.; Brooks, M.S.; Budge, S.M.; Dave, D. Fish processing wastes as a potential source of proteins, amino acids and oils: A critical review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar]
- Routray, W.; Dave, D.; Ramakrishnan, V.V.; Murphy, W. Production of high quality fish oil by enzymatic protein hydrolysis from cultured atlantic salmon by-products: Investigation on effect of various extraction parameters using central composite rotatable design. Waste Biomass Valorization 2018, 9, 2003–2014. [Google Scholar] [CrossRef]
- Amarowicz, R.; Shahidi, F. Antioxidant activity of peptide fractions of capelin protein hydrolysates. Food Chem. 1997, 58, 355–359. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; and Rasco, B.A. Kinetics of the hydrolysis of Atlantic salmon (Salmo salar) muscle proteins by alkaline proteases and a visceral serine protease mixture. Process Biochem. 2000, 36, 131–139. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Hydrolysis of salmon muscle proteins by an enzyme mixture extracted from Atlantic salmon (Salmo salar) pyloric caeca. J. Food Biochem. 2000, 24, 177–187. [Google Scholar] [CrossRef]
- Thorkelsson, G.; Kristinsson, H. Bioactive peptides from marine sources. State of art. Report to the NORA fund. Peptides 2009, 1–19. [Google Scholar]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Keskin, U.S.; Hong, H.; Wu, J. Regulatory requirements of bioactive peptides (protein hydrolysates) from food proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G. Lipid oxidation and fishy odour in protein hydrolysate derived from Nile tilapia (Oreochromis niloticus) protein isolate as influenced by haemoglobin. J. Sci. Food Agric. 2014, 94, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Kishimura, H. Antioxidant and sensory properties of protein hydrolysate derived from Nile tilapia (Oreochromis niloticus) by one- and two-step hydrolysis. J. Food Sci. Technol. 2015, 52, 3336–3349. [Google Scholar] [CrossRef] [PubMed]
- Opheim, M.; Šližyte, R.; Sterten, H.; Provan, F.; Larssen, E.; Kjos, N.P. Hydrolysis of Atlantic salmon (Salmo salar) rest raw materials - Effect of raw material and processing on composition, nutritional value, and potential bioactive peptides in the hydrolysates. Process Biochem. 2015, 50, 1247–1257. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide identification in a salmon gelatin hydrolysate with antihypertensive, dipeptidyl peptidase IV inhibitory and antioxidant activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [CrossRef]
- Ishak, N.H.; Sarbon, N.M. A review of protein hydrolysates and bioactive peptides deriving from wastes generated by fish processing. Food Bioprocess Technol. 2018, 11, 2–16. [Google Scholar] [CrossRef]
- DFO. Aquaculture production quantities and value. In: Fish Ocean Canada, 2021. https://www.dfo-mpo.gc.ca/stats/aqua/aqua-prod-eng.htm. Accessed 2 Jul 2021.
- GovNL. Aquaculture - Newfoundland & Labrador Canada. In: Invest Aquac. 2021. https://www.findnewfoundlandlabrador.com/invest/aquaculture/. Accessed 2 Jul 2021.
- Seafish. Fishmeal and fish oil figures. 2011. Seafish 30.
- DFO. State of salmon aquaculture technologies. 2019. 2019.
- MOWI. Salmon Farming Industry Handbook 2019. 2019.
- ACCFA. Growing our fish. In: Atl. Fish Farmers. 2021. https://www.atlanticfishfarmers.com/growing-our-fish%0Ahttps://www.atlanticfishfarmers.com/growing-our-fish#:~:text=Stocking,between 15%2C000 and 30%2C000 smolts. Accessed 2 Jul 2021.
- Borderías, A.J; a Sánchez-alonso, I. First processing steps and the quality of wild and farmed fish. J. Food Sci. 2011, 76, R1–R5. [Google Scholar] [CrossRef] [PubMed]
- Frida, N.-U.; Nesse, A.G. Smoother harvest of farmed salmon, value-adding or costly? Norwegian University of Science and Technology 2014. [Google Scholar]
- Dave, D.; Routray, W. Current scenario of Canadian fishery and corresponding underutilized species and fishery byproducts: A potential source of omega-3 fatty acids. J. Clean. Prod. 2018, 180, 617–641. [Google Scholar] [CrossRef]
- Kim, Y.S.; Park, J.W. Negative roles of salt in gelation properties of fish protein isolate. J Food Sci. 2008, 73, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Nolsøe, H.; Undeland, I. The acid and alkaline solubilization process for the isolation of muscle proteins: State of the art. Food Bioproc. Tech. 2009, 2, 1–27. [Google Scholar] [CrossRef]
- Ghaly, A.E.; Dave, D.; Brooks, M.S.; Budge, S. Production of biodiesel by enzymatic transesterification: Review. Am. J. Biochem. Biotechnol. 2010, 6, 54–76. [Google Scholar] [CrossRef]
- Aas, T.S.; Åsgård, T.; Ytrestøyl, T. Chemical composition of whole body and fillet of slaughter sized Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss) farmed in Norway in 2020. Aquac. Rep. 2022, 25, 101252. [Google Scholar] [CrossRef]
- Dave, D.; Ramakrishnan, V.V.; Trenholm, S.; Manuel, H.; Pohling, J.; Murphy, W. Marine oils as potential feedstock for biodiesel production: Physicochemical characterization. J Bioprocess Biotech. 2014, 4, 1–12. [Google Scholar] [CrossRef]
- Malcorps, W.; Newton, W.R.; Sprague, M.; Glencross, B.D.; Little, C.D. Nutritional characterisation of European aquaculture processing by-products to facilitate strategic utilisation. Front Sustain Food Syst. 2021, 5, 720595. [Google Scholar] [CrossRef]
- Kralovec, J.A.; Zhang, S.; Zhang, W.; Barrow, C.J. A review of the progress in enzymatic concentration and microencapsulation of omega-3 rich oil from fish and microbial sources. Food Chem. 2012, 131, 639–644. [Google Scholar] [CrossRef]
- Liu, Y.; Ramakrishnan, V.V.; Dave, D. Lipid class and fatty acid composition of oil extracted from Atlantic salmon by-products under different optimization parameters of enzymatic hydrolysis. Biocatal Agric Biotechnol. 2020, 30, 101866. [Google Scholar] [CrossRef]
- Polvi, S.M.; Ackman, R.G. Atlantic salmon (Salmo salar) muscle lipids and their response to alternative dietary fatty acid sources. J. Agric. Food Chem. 1992, 40, 1001–1007. [Google Scholar] [CrossRef]
- Głowacz-Różyńska, A.; Tynek, M.; Malinowska-Pańczyk, E.; Martysiak-Żurowska, D.; Pawłowicz, R.; Kołodziejska, I. Comparison of oil yield and quality obtained by different extraction procedures from salmon (Salmo salar) processing byproducts. Eur. J. Lipid Sci. Technol. 2016, 118, 1759–1767. [Google Scholar] [CrossRef]
- Liaset, B.; Julshamn, K.; Espe, M. Chemical composition and theoretical nutritional evaluation of the produced fractions from enzymic hydrolysis of salmon frames with ProtamexTM. Process Biochem. 2003, 38, 1747–1759. [Google Scholar] [CrossRef]
- Haq, M.; Ahmed, R.; Cho, Y.J.; Chun, B.S. Quality Properties and bio-potentiality of edible oils from Atlantic salmon by-products extracted by supercritial carbon dioxide and conventional methods. Waste Biomass Valorization 2017, 8, 1953–1967. [Google Scholar] [CrossRef]
- Colombo, S.M.; Mazal, X. Investigation of the nutritional composition of different types of salmon available to Canadian consumers. J. Agric. Food Res. 2020, 2, 100056. [Google Scholar] [CrossRef]
- Fomena-Temgoua, N.S.; Sun, Z.; Okoye, C.O.; Pan, H. Fatty acid profile, physicochemical composition, and sensory properties of Atlantic salmon fish (Salmo salar) during Different culinary treatments. J. Food Qual. 2022, 7425142. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. Functional and physiochemical properties of protein isolates from different body parts of North Atlantic sea cucumber (Cucumaria frondosa). Food Biosci 2023, 52, 102511. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Antioxidant potential of sea cucumbers and their beneficial effects on human health. Mar. Drugs 2002, 20, 521. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional enzymes in life and disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, T.R.L.; Dave, D.; Shahidi, F. Antioxidant potential and physicochemical properties of protein hydrolysates from body parts of North Atlantic sea cucumber (Cucumaria frondosa). Food Prod. Process. Nutr. 2021, 3, 1–22. [Google Scholar] [CrossRef]
- Shen, H.B.; Chou, K.C. Identification of proteases and their types. Anal. Biochem. 2009, 385, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Undeland, I. Physicochemical and gel-forming properties of protein isolated from salmon, cod and herring by-products using the pH-shift method. LWT - Food Sci. Technol 2019, 101, 678–684. [Google Scholar] [CrossRef]
- Xiong, Y.L. Muscle proteins. In Muscle proteins. In: Proteins in Food Processing; Elsevier Ltd., 2004; pp. 100–122. [Google Scholar]
- Ramakrishnan, V.V.; Ghaly, A.E.; Brooks, M.S.; Budge, S.M. Extraction of proteins from mackerel fish processing waste using Alcalase enzyme. J. Bioprocess. Biotech. 2013, 3, 1–9. [Google Scholar]
- Dave, D.; Vegneshwaran, V.R.; Julia, P.; Sukhinder, K.C.; Sheila, T.; Heather, M.; Wade, M. Investigation on oil extraction methods and its influence on omega-3 content from cultured salmon. J. Food Process. Technol. 2014, 5, 1000401. [Google Scholar]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Petrova, I.; Tolstorebrov, I.; Eikevik, T.M. Production of fish protein hydrolysates step by step: Technological aspects, equipment used, major energy costs and methods of their minimizing. Int. Aquat. Res. 2018, 10, 223–241. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Álvarez, C.; O’Donnell, C.P. Ultrasound applications for the extraction, identification and delivery of food proteins and bioactive peptides. Trends Food Sci. Technol. 2015, 46, 60–67. [Google Scholar] [CrossRef]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [PubMed]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproducts hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- See, S.F.; Hoo, L.L.; Babji, A.S. Optimization of enzymatic hydrolysis of salmon (Salmo salar) skin by Alcalase. Int. Food Res. J. 2011, 18, 1359–1365. [Google Scholar]
- Zhang, X.; Dai, Z.; Zhang, Y.; Dong, Y.; Hu, X. Structural characteristics and stability of salmon skin protein hydrolysates obtained with different proteases. LWT - Food Sci. Technol 2022, 153, 112460. [Google Scholar] [CrossRef]
- Slizyte, R.; Rommi, K.; Mozuraityte, R.; Eck, P.; Five, K.; Rustad, T. Bioactivities of fish protein hydrolysates from defatted salmon backbones. Biotechnol. Rep. 2016, 11, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Aspevik, T.; Egede-Nissen, H.; Oterhals, Å. A systematic approach to the comparison of cost efficiency of endopeptidases for the hydrolysis of Atlantic salmon (Salmo salar) by-products. Food Technol. Biotechnol 2016, 54, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Liaset, B.; Nortvedt, R.; Lied, E.; Espe, M. Studies on the nitrogen recovery in enzymic hydrolysis of Atlantic salmon (Salmo salar, L.) frames by Protamex™ protease. Process Biochem 2002, 37, 1263–1269. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Sotelo, C.G.; Sanz, N.; Pérez-Martín, R.I.; Amado, I.R.; Valcarcel, J. Valorization of aquaculture by-products of salmonids to produce enzymatic hydrolysates: Process optimization, chemical characterization and evaluation of bioactives. Mar. Drugs 2019, 17, 676. [Google Scholar] [CrossRef]
- Idowu, A.T.; Benjakul, S.; Sinthusamran, S.; Sookchoo, P.; Kishimura, H. Protein hydrolysate from salmon frames: Production, characteristics and antioxidative activity. J. Food Biochem. 2019, 43, e12734. [Google Scholar] [CrossRef]
- Aspevik, T.; Steinsholm, S.; Vang, B.; Carlehög, M.; Arnesen, J.A.; Kousoulaki, K. Nutritional and sensory properties of protein hydrolysates based on salmon (Salmo salar), mackerel (Scomber scombrus), and herring (Clupea harengus) heads and backbones. Front. Nutr. 2021, 8, 695151. [Google Scholar] [CrossRef]
- Kangsanant, S.; Murkovic, M.; Thongraung, C. Antioxidant and nitric oxide inhibitory activities of tilapia (Oreochromis niloticus) protein hydrolysate: Effect of ultrasonic pretreatment and ultrasonic-assisted enzymatic hydrolysis. Int. J. Food Sci. Technol. 2014, 49, 1932–1938. [Google Scholar] [CrossRef]
- Álvarez, C.; Lélu, P.; Lynch, S.A.; Tiwari, B.K. Optimised protein recovery from mackerel whole fish by using sequential acid/alkaline isoelectric solubilization precipitation (ISP) extraction assisted by ultrasound. LWT - Food Sci. Technol 2018, 88, 210–216. [Google Scholar] [CrossRef]
- Misir, G.B.; Koral, S. Effects of ultrasound treatment on structural, chemical and functional properties of protein hydrolysate of rainbow trout (Oncorhynchus mykiss) by-products. Ital J Food Sci. 2019, 31, 205–223. [Google Scholar]
- Tian, J.; Wang, Y.; Zhu, Z.; Zeng, Q.; Xin, M. Recovery of tilapia (Oreochromis niloticus) protein isolate by high-intensity ultrasound-aided alkaline isoelectric solubilization/precipitation process. Food Bioproc. Tech. 2015, 8, 758–769. [Google Scholar] [CrossRef]
- Elamin, W.M.; Endan, J.B.; Yosuf, Y.A.; Shamsudin, R.; Ahmedov, A. High pressure processing technology and equipment evolution: A review. J. Eng. Sci. Technol. Rev. 2015, 8, 75–83. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Effect of high-pressure processing (hpp) on phenolics of North Atlantic sea cucumber (Cucumaria frondosa). J. Agric. Food Chem. 2022, 70, 3489–3501. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, A.; Suwal, S.; Naderi, N.; Pouliot, Y.; Doyen, A. Enhancing enzymatic hydrolysis of food proteins and production of bioactive peptides using high hydrostatic pressure technology. Trends Food Sci. Technol. 2018, 80, 187–198. [Google Scholar] [CrossRef]
- Rivalain, N.; Roquain, J.; Demazeau, G. Development of high hydrostatic pressure in biosciences: Pressure effect on biological structures and potential applications in Biotechnologies. Biotechnol. Adv 2010, 28, 659–672. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, B.B.; Park, C.B.; Clark, D.S. Pressure effects on intra- and intermolecular interactions within proteins. Biochim. Biophys. Acta - Protein Struc. Mol. Enzymol 2002, 1595, 235–249. [Google Scholar] [CrossRef]
- Royer, C.A. Insights into the role of hydration in protein structure and stability obtained through hydrostatic pressure studies. Braz. J. Med. Biol. Res. 2005, 38, 1167–1173. [Google Scholar] [CrossRef]
- Silva, J.L.; Foguel, D. Hydration, cavities and volume in protein folding, aggregation and amyloid assembly. Phys. Biol. 2009, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Grigera, J.R.; McCarthy, A.N. The behavior of the hydrophobic effect under pressure and protein denaturation. Biophysical J. 2010, 98, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Knorr, D.; Heinz, V.; Buckow, R. High pressure application for food biopolymers. Biochim. Biophys. Acta - Proteins Proteom 2006, 1764, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, G.; Schwalbe, H. Protein NMR spectroscopy: Hydrogen bonds under pressure. Nat. Chem. 2012, 4, 693–695. [Google Scholar] [CrossRef] [PubMed]
- Alemán, A.; Giménez, B.; Gómez-Guillén, M.C. ’ Montero, P. Enzymatic hydrolysis of fish gelatin under high pressure treatment. Int. J. Food Sci. Technol. 2011, 46, 1129–1136. [Google Scholar] [CrossRef]
- Hemker, A.K.; Nguyen, L.T.; Karwe, M.; Salvi, D. Effects of pressure-assisted enzymatic hydrolysis on functional and bioactive properties of tilapia (Oreochromis niloticus) by-product protein hydrolysates. LWT - Food Sci. Technol. 2020, 122, 109003. [Google Scholar] [CrossRef]
- Nakajima, K.; Yoshie-Stark, Y.; Ogushi, M. Comparison of ACE inhibitory and DPPH radical scavenging activities of fish muscle hydrolysates. Food Chem. 2009, 114, 844–851. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Atlantic salmon (Salmo salar) co-product-derived protein hydrolysates: A source of antidiabetic peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar] [CrossRef]
- Abdollahi, M.; Undeland, I. Structural, functional, and sensorial properties of protein isolate produced from salmon, cod, and herring by-products. Food Bioproc. Tech. 2018, 11, 1733–1749. [Google Scholar] [CrossRef]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and bioactive properties of peptides derived from marine side streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef]
- Zayas, J.F. Functionality of proteins in food; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Jones, S.M. Quality and Grading of Carcasses of Meat Animals. CRC Press Jungbauer, A. and Hahn, R. 2009, Chapter 22 Ion-Exchange Chromatography. In: Methods in Enzymology. Academic Press Inc., pp 349–371.
- Munasinghe, D.M.S.; Sakai, T. Sodium chloride (0.8 M) as a better protein extractant for fish meat quality assessments. J. Food Sci 2003, 68, 1059–1062. [Google Scholar] [CrossRef]
- Chan, S.K.W.; Roth, B.; Jessen, F.; Jakobsen, A.N.; Lerfall, J. Water holding properties of Atlantic salmon. Compr. Rev. Food Sci. Food Saf. 2021, 21, 477–498. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.H.; Nickerson, M.T. Food proteins: A review on their emulsifying properties using a structure-function approach. Food Chem. 2013, 141, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.A.; Timilsena, Y.P.; Adhikari, B. Food proteins, structure, and function. Ref. Modul. Food Sci. 2016. [Google Scholar]
- Damodaran, S.; Parkin, K.L.; Fennema, O.R. Fennema’s food chemistry, 4th Eds., 2007, CRC Press.
- He, S.; Franco, C.; Zhang, W. Process optimisation and physicochemical characterisation of enzymatic hydrolysates of proteins from co-products of Atlantic salmon (Salmo salar) and yellowtail kingfish (Seriola lalandi). Int. J. Food Sci. Technol. 2012, 47, 2397–2404. [Google Scholar] [CrossRef]
- Saha, B.C.; Hayashi, K. Debittering of protein hydrolyzates. Biotechnol. Adv. 2001, 19, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.J.; Unklesbay, N.; Hsieh, F.H.; Clarke, A.D. Hydrophobicity of bitter peptides from soy protein hydrolysates. J. Agric. Food Chem 2004, 52, 5895–5901. [Google Scholar] [CrossRef]
- Sun, X.D. Enzymatic hydrolysis of soy proteins and the hydrolysates utilisation. Int. J. Food Sci. Technol. 2011, 46, 2447–2459. [Google Scholar] [CrossRef]
- Benjakul, S.; Yarnpakdee, S.; Senphan, T.; Halldorsdottir, S.M.; Kristinsson, H.G. Fish protein hydrolysates: Production, bioactivities, and applications. In: Antioxidants and Functional Components in Aquatic Foods. 2014, John Wiley & Sons, Ltd, Chichester, UK, pp 237–281.
- Xu, Q.; Hong, H.; Yu, W.; Jiang, X.; Yan, X.; Wu, J. Sodium chloride suppresses the bitterness of protein hydrolysates by decreasing hydrophobic interactions. J. Food Sci. 2019, 84, 86–91. [Google Scholar] [CrossRef]
- Gong, M.; Mohan, A.; Gibson, A.; Udenigwe, C.C. Mechanisms of plastein formation, and prospective food and nutraceutical applications of the peptide aggregates. Biotechnol. Rep. 2015, 5, 63–69. [Google Scholar] [CrossRef]
- Aspevik, T.; Totland, C.; Lea, P.; Oterhals, Å. Sensory and surface-active properties of protein hydrolysates based on Atlantic salmon (Salmo salar) by-products. Process Biochem. 2016, 51, 1006–1014. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Bioactive peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Sefora, R.G.; Taneyo, S.D.L.; Gianotti, A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Daliri, E.B.M.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, H.; Xing, R.; Li, P. Characterization, preparation, and purification of marine bioactive peptides. Biomed. Res. Int. 2017, 2017, 9746720. [Google Scholar] [CrossRef] [PubMed]
- Phadke, G.G.; Rathod, N.B.; Ozogul, F.; Elavarasan, K.; Karthikeyan, M.; Shin, K.-H.; Kim, S.-K. Exploiting of secondary raw materials from fish processing industry as a source of bioactive peptide-rich protein hydrolysates. Mar. Drugs 2021, 19, 480. [Google Scholar] [CrossRef]
- Ahn, C.B.; Jeon, Y.J.; Kim, Y.T.; Je, J.Y. AngiotensinI converting enzyme (ACE) inhibitory peptides from salmon byproduct protein hydrolysate by Alcalase hydrolysis. Process Biochem. 2012, 47, 2240–2245. [Google Scholar] [CrossRef]
- Gu, R.Z.; Li, C.Y.; Liu, W.Y.; Yi, W.X.; Cai, M.Y. Angiotensin I-converting enzyme inhibitory activity of low-molecular-weight peptides from Atlantic salmon (Salmo salar L.) skin. Food Res. Int 2011, 44, 1536–1540. [Google Scholar] [CrossRef]
- Ahn, C.B.; Cho, Y.S.; Je, J.Y. Purification and anti-inflammatory action of tripeptide from salmon pectoral fin byproduct protein hydrolysate. Food Chem. 2015, 168, 151–156. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y.; Hunag, S.L.; Jao, C.L.; Ho, K.P.; Hsu, K.C. Peptides derived from Atlantic salmon skin gelatin as dipeptidyl-peptidase IV inhibitors. J. Agric. Food Chem. 2012, 60, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP–IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, F.; Han, F.; Wang, H. Purification and characterization of antioxidative peptides from salmon protamine hydrolysate. J. Food Biochem. 2008, 32, 654–671. [Google Scholar] [CrossRef]
- Darewicz, M.; Borawska, J.; Vegarud, G.E.; Minkiewicz, P.; Iwaniak, A. Angiotensin I-converting enzyme (ACE) inhibitory activity and ace inhibitory peptides of salmon (Salmo salar) protein hydrolysates obtained by human and porcine gastrointestinal enzymes. Int. J. Mol. Sci. 2014, 15, 14077–14101. [Google Scholar] [CrossRef] [PubMed]
- Picot, L.; Bordenave, S.; Didelot, S. Antiproliferative activity of fish protein hydrolysates on human breast cancer cell lines. Process Biochem. 2006, 41, 1217–1222. [Google Scholar] [CrossRef]
- Ahn, C.B.; Je, J.Y.; Cho, Y.S. Antioxidant and anti-inflammatory peptide fraction from salmon byproduct protein hydrolysates by peptic hydrolysis. Food Res. Int. 2012, 49, 92–98. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Hasan, F.M.; Gill, T.A.; Aluko, R.E. Antioxidant properties of Salmon (Salmo salar) protein hydrolysate and peptide fractions isolated by reverse-phase HPLC. Food Res. Int. 2013, 52, 315–322. [Google Scholar] [CrossRef]
- De La Fuente, B.; Pallarés, N.; Berrada, H.; Barba, F.J. Salmon (Salmo salar) side streams as a bioresource to obtain potential antioxidant peptides after applying pressurized liquid extraction (PLE). Mar. Drugs 2021, 19, 323. [Google Scholar] [CrossRef]
- Hanachi, A.; Bianchi, A.; Kahn, C.J.F.; Velot, E.; Arab-Tehrany, E.; Cakir-Kiefer, C.; Linder, M. Encapsulation of salmon peptides in marine liposomes: Physico-chemical properties, antiradical activities and biocompatibility assays. Mar. Drugs 2022, 20, 249. [Google Scholar] [CrossRef]
- Rajendran, S.R.C.K.; Mohan, A.; Khiari, Z.; Udenigwe, C.C.; Mason, B. Yield, physicochemical, and antioxidant properties of Atlantic salmon visceral hydrolysate: Comparison of lactic acid bacterial fermentation with Flavourzyme proteolysis and formic acid treatment. J. Food Process. Preserv. 2018, 42, e13620. [Google Scholar] [CrossRef]
- Wang, K.; Siddanakoppalu, P.N.; Ahmed, I.; Pavase, T.R.; Lin, H.; Li, Z. Purification and identification of anti-allergic peptide from Atlantic salmon (Salmo salar) byproduct enzymatic hydrolysates. J. Funct. Foods 2020, 72, 104084. [Google Scholar] [CrossRef]
- Agyei, D.; Ongkudon, C.M.; Wei, C.Y.; Chan, A.S.; Danquah, M.K. Bioprocess challenges to the isolation and purification of bioactive peptides. Food Bioprod. Process. 2016, 98, 244–256. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. In Silico analysis of bioactive peptides produced from underutilized sea cucumber by-products—A bioinformatics approach. Mar. Drugs 2022, 20, 610. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. TRAC - Trends Analyt. Chem 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-Y.; Hsieh, C.-H.; Hung, C.-C.; Jao, C.-L.; Lin, P.-Y.; Hsieh, Y.-L.; Hsu, K.-C. A study to evaluate the potential of an in silico approach for predicting dipeptidyl peptidase-IV inhibitory activity in vitro of protein hydrolysates. Food Chem. 2017, 234, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Tichý, M.; Hanzlíková, I.; Rucki, M.; Pokorná, A.; Uzlová, R.; Tumová, J. Acute toxicity of binary mixtures: Alternative methods, QSAR and mechanisms. Interdiscip. Toxicol. 2008, 1, 15–17. [Google Scholar] [CrossRef]
- Gad, S.C. Encyclopedia of Toxicology, 2014, Third Edition. Elsevier, pp 1–9.
- Nongonierma, A.B.; Fitzgerald, R.J. Learnings from quantitative structure-activity relationship (QSAR) studies with respect to food protein-derived bioactive peptides: A review. RSC Adv. 2016, 6, 75400–75413. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef]
- Peter, S.C.; Dhanjal, J.K.; Malik, V.; Radhakrishnan, N.; Jayakanthan, M.; Sundar, D.; Sundar, D. Quantitative structure-activity relationship (QSAR): Modeling approaches to biological applications. In: Encyclopedia of Bioinformatics and Computational Biology: ABC of Bioinformatics. 2018, Elsevier, pp 661–676.
- Lemes, A.C.; Sala, L.; Ores, J.D.C.; Braga, A.R.C.; Egea, M.B.; Fernandes, K.F. A review of the latest advances in encrypted bioactive peptides from protein-richwaste. Int. J. Mol. Sci. 2016, 17, 950. [Google Scholar] [CrossRef] [PubMed]
- Agyei, D.; Danquah, M.K. Industrial-scale manufacturing of pharmaceutical-grade bioactive peptides. Biotechnol Adv. 2011, 29, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, L.; Firdaous, L. Separation of bioactive peptides by membrane processes: Technologies and devices. Recent Pat. Biotechnol. 2013, 7, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Hakami, M.W.; Alkhudhiri, A.; Al-Batty, S.; Zacharof, M.P.; Maddy, J.; Hilal, N. Ceramic microfiltration membranes in wastewater treatment: Filtration behavior, fouling and prevention. Membranes 2020, 10, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Muro, C.; Riera, F.; Fernandez, A. (2013) Advancements in the fractionation of milk biopeptides by means of membrane processes. In: Bioactive Food Peptides in Health and Disease. 2013, InTech.
- Suwal, S.; Ketnawa, S.; Liceaga, A.M.; Huang, J.Y. Electro-membrane fractionation of antioxidant peptides from protein hydrolysates of rainbow trout (Oncorhynchus mykiss) byproducts. Innov. Food Sci. Emerg. Technol. 2018, 45, 122–131. [Google Scholar] [CrossRef]
- Doyen, A.; Beaulieu, L.; Saucier, L.; Pouliot, Y.; Bazinet, L. Demonstration of in vitro anticancer properties of peptide fractions from a snow crab by-products hydrolysate after separation by electrodialysis with ultrafiltration membranes. Sep. Purif. Technol. 2011, 78, 321–329. [Google Scholar] [CrossRef]
- Roblet, C.; Akhtar, M.J.; Mikhaylin, S.; Pilon, G.; Gill, T.; Marette, A.; Bazinet, L. Enhancement of glucose uptake in muscular cell by peptide fractions separated by electrodialysis with filtration membrane from salmon frame protein hydrolysate. J. Funct. Foods 2016, 22, 337–346. [Google Scholar] [CrossRef]
- Perez Espitia, P.J.; de Fátima Ferreira Soares, N.; dos Reis Coimbra, J.S.; de Andrade, N.J.; Souza Cruz, R.; Alves Medeiros, E.A. Bioactive peptides: Synthesis, properties, and applications in the packaging and preservation of food. Compr. Rev. Food Sci. Food Saf. 2012, 11, 187–204. [Google Scholar] [CrossRef]
- Ahn, C.B.; Kim, J.G.; Je, J.Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef]
- Health Canada. Licensed natural health products database (LNHPD). In: Online query Phoslax. 2021, https://health-products.canada.ca/lnhpd-bdpsnh/info.do?licence=80000689#fn1-rf. Accessed 20 Apr 2021.
- Health Canada Questions and Answers about Preparing Submissions for Food Health Claims. In: Heal. Canada. 2015, https://www.canada.ca/en/health-canada/services/food-nutrition/legislation-guidelines/guidance-documents/questions-answers-about-preparing-submissions-food-health-claims-2015.html. Accessed 20 Apr 2021.
- Bureau of Nutritional Sciences. Guidance Document for Preparing a Submission for Food Health Claims; Health Canada, 2009; pp. 1–45. [Google Scholar]
Figure 1.
Canadian finfish and shellfish production and values (2008-2021).
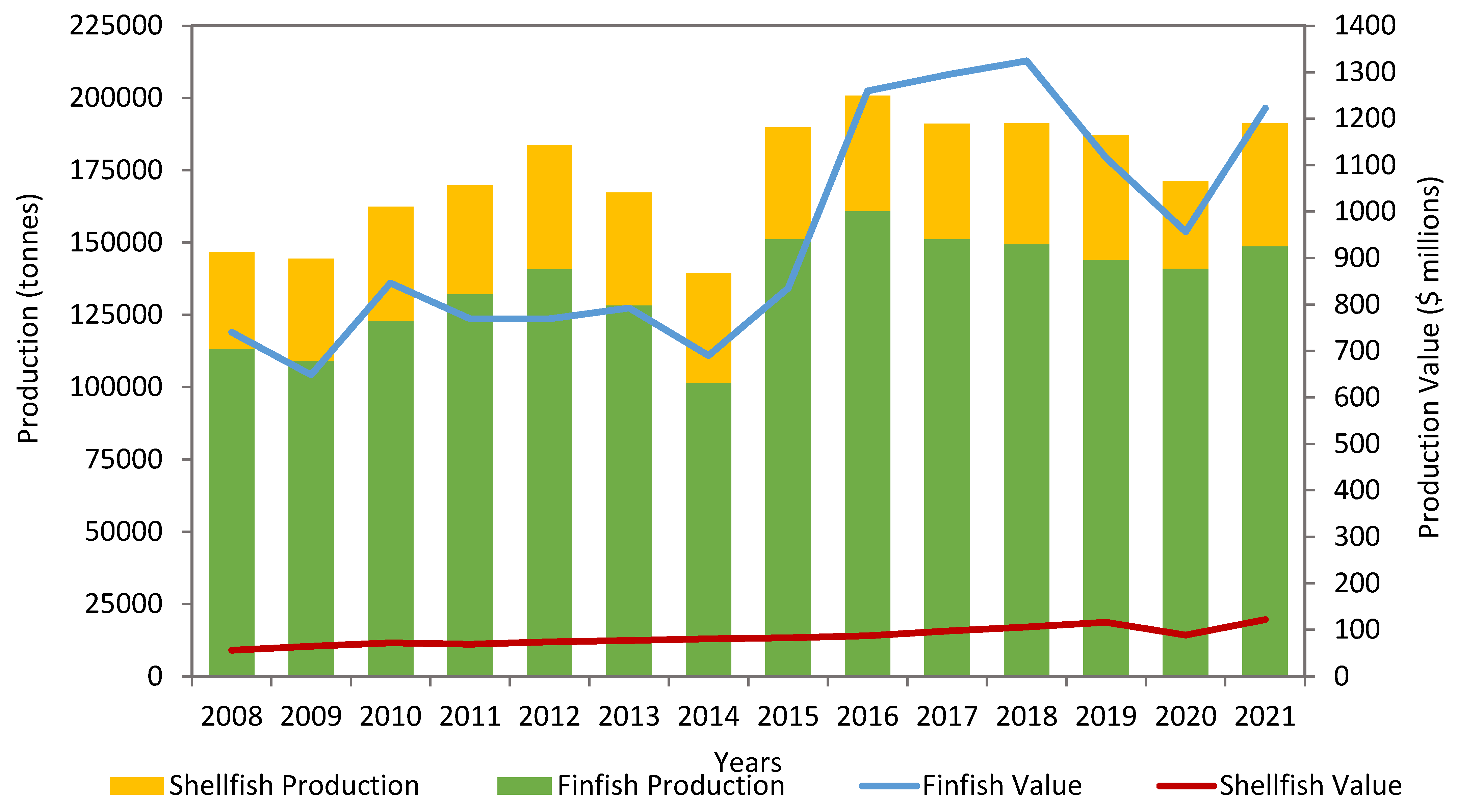
Figure 2.
Various types of Canadian finfish production and values (2008-2021).
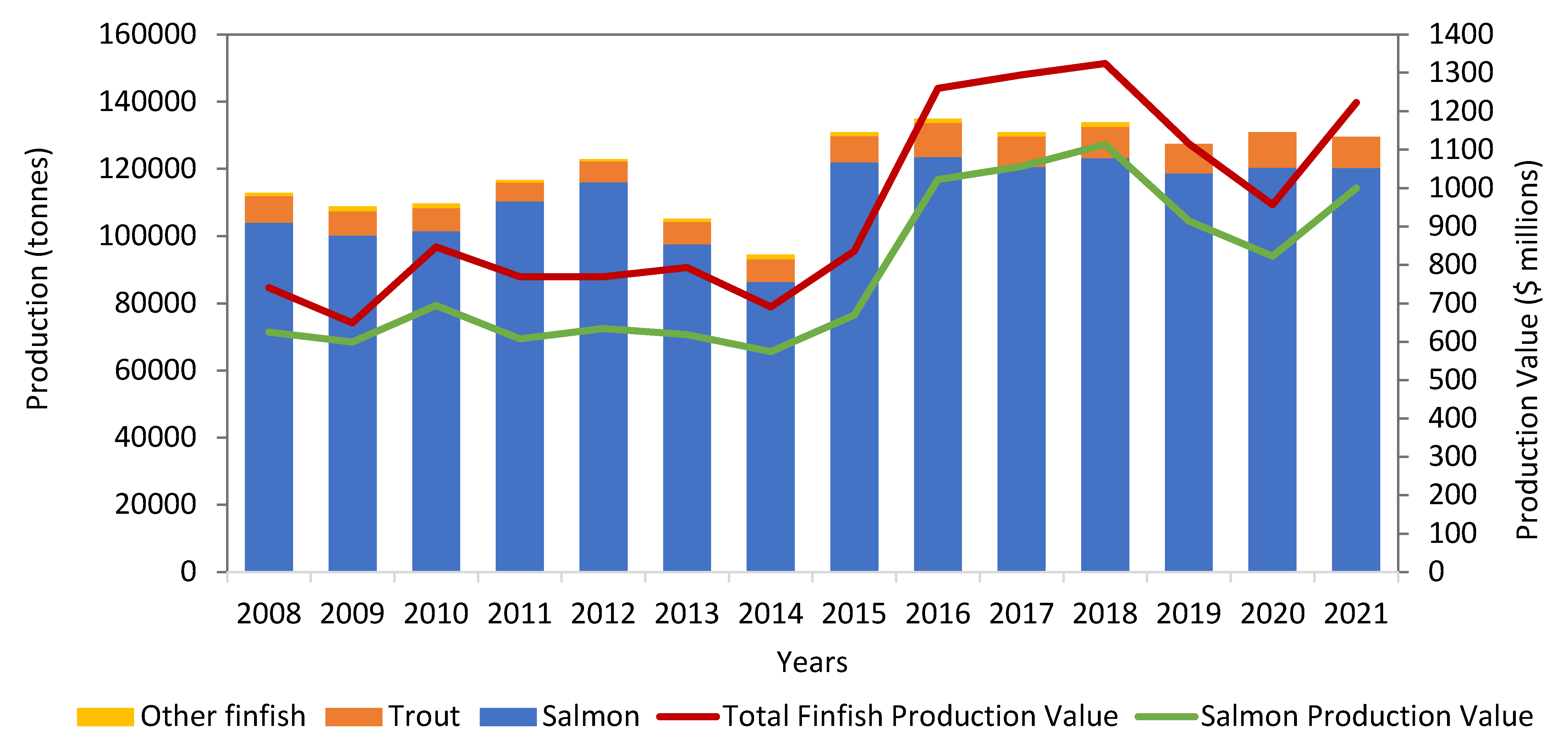
Figure 3.
Newfoundland and Labrador finfish, salmon and shellfish production and values (2008-2021).
Figure 3.
Newfoundland and Labrador finfish, salmon and shellfish production and values (2008-2021).
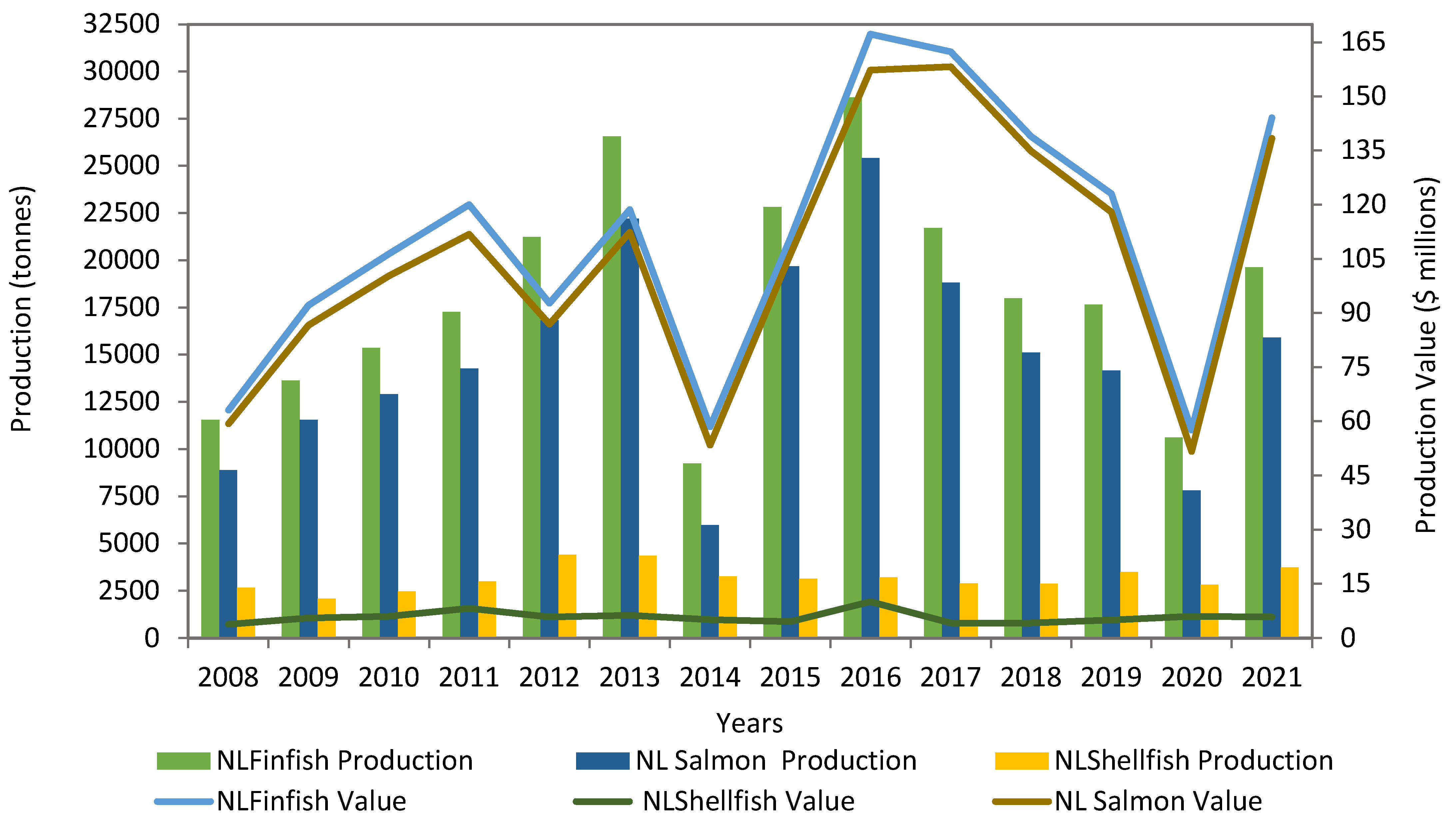
Figure 4.
The lifecycle of Atlantic salmon in aquaculture.
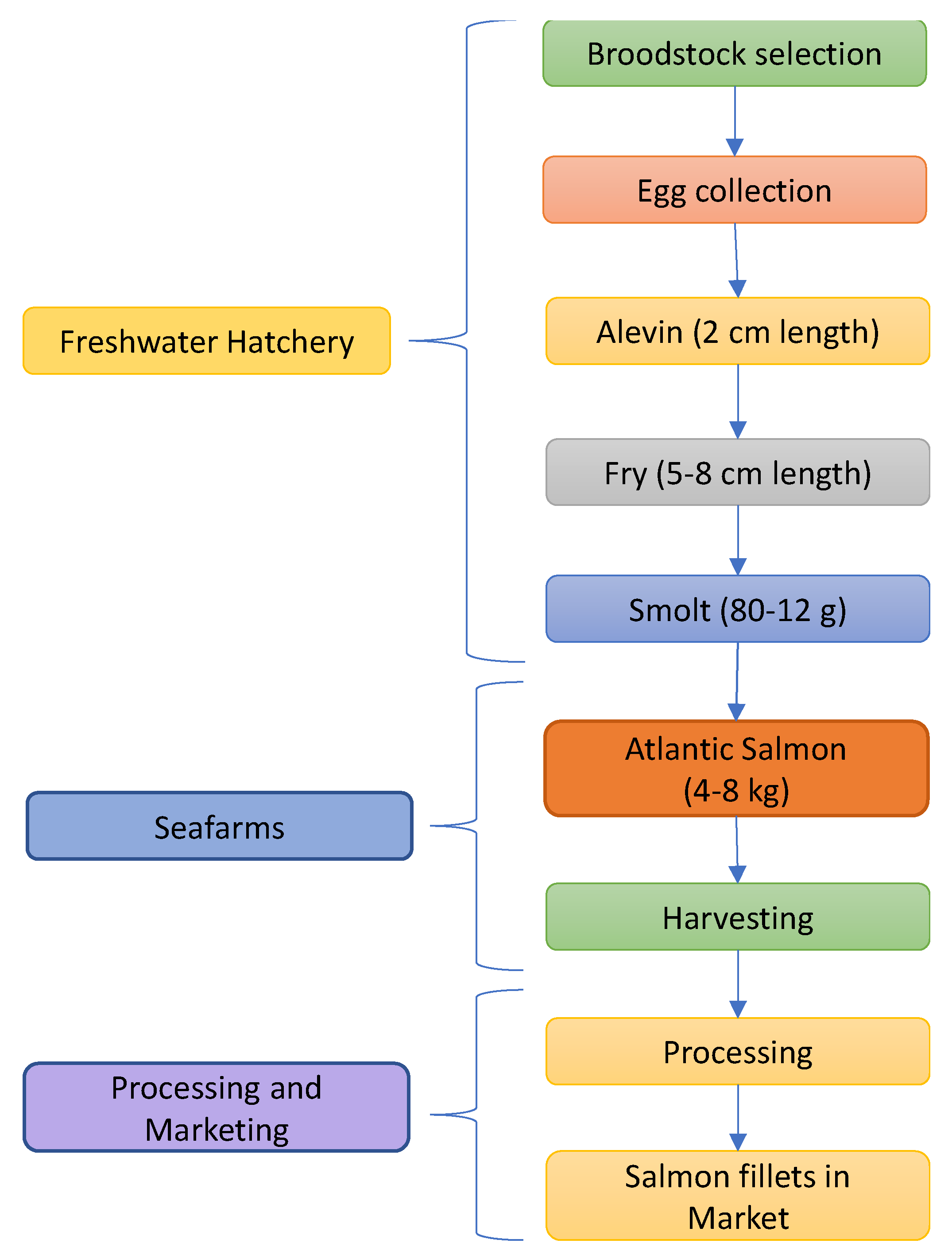
Figure 5.
Value-added utilization of salmon processing by-products.
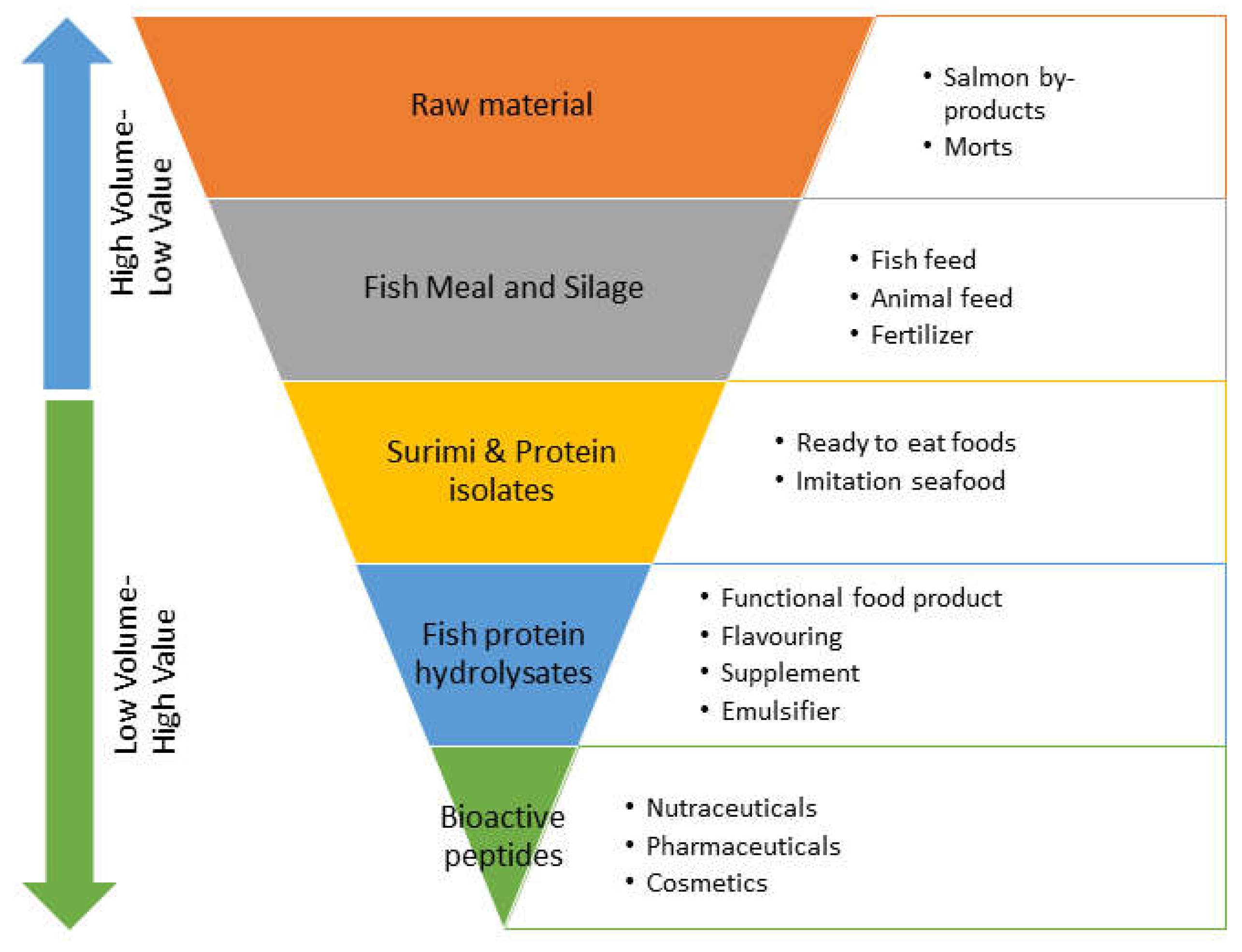
Figure 6.
Production of salmon protein via enzymatic hydrolysis process .
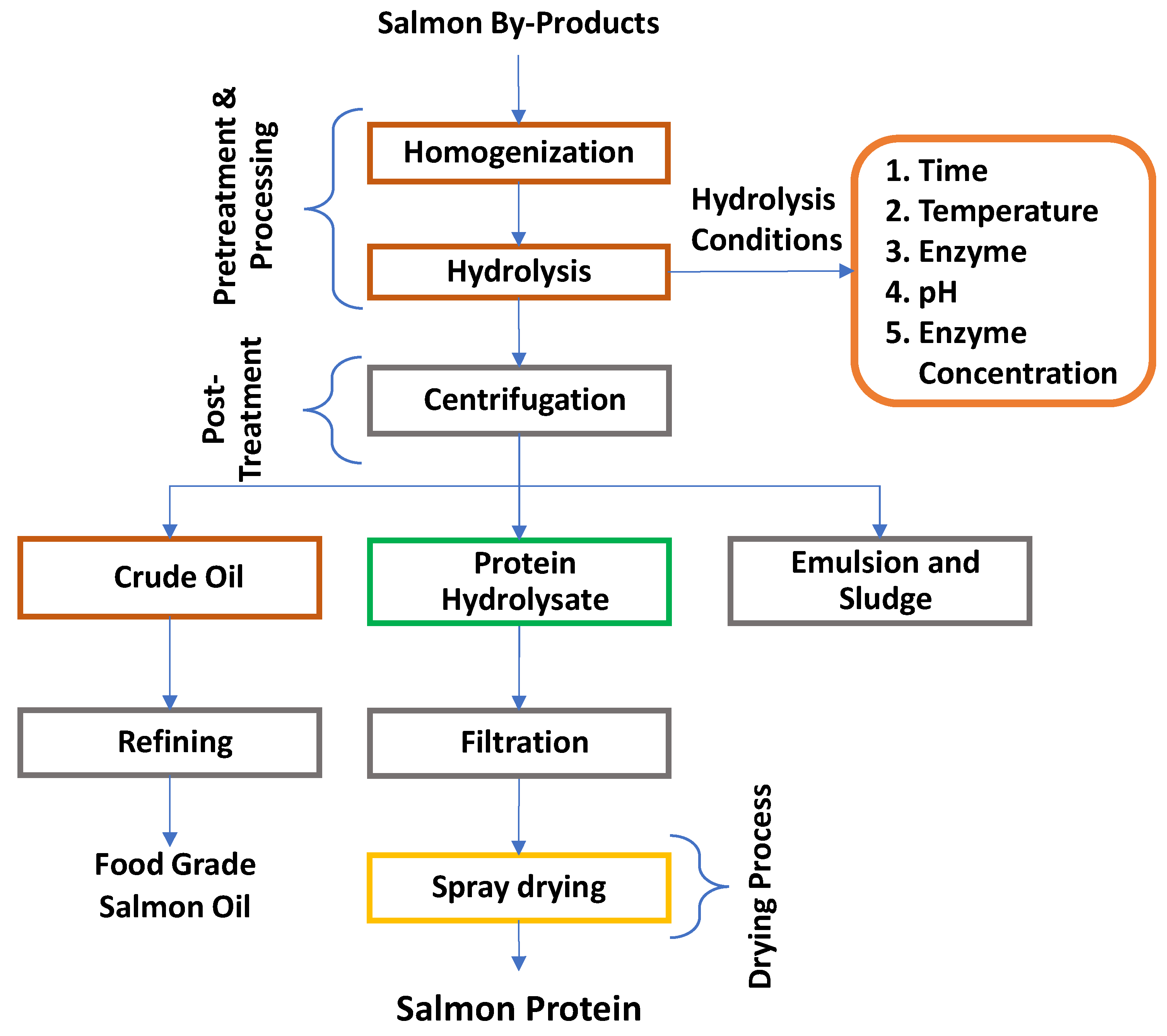
Figure 7.
Empirical production of bioactive peptides. .

Figure 8.
In-silico production of bioactive peptides.
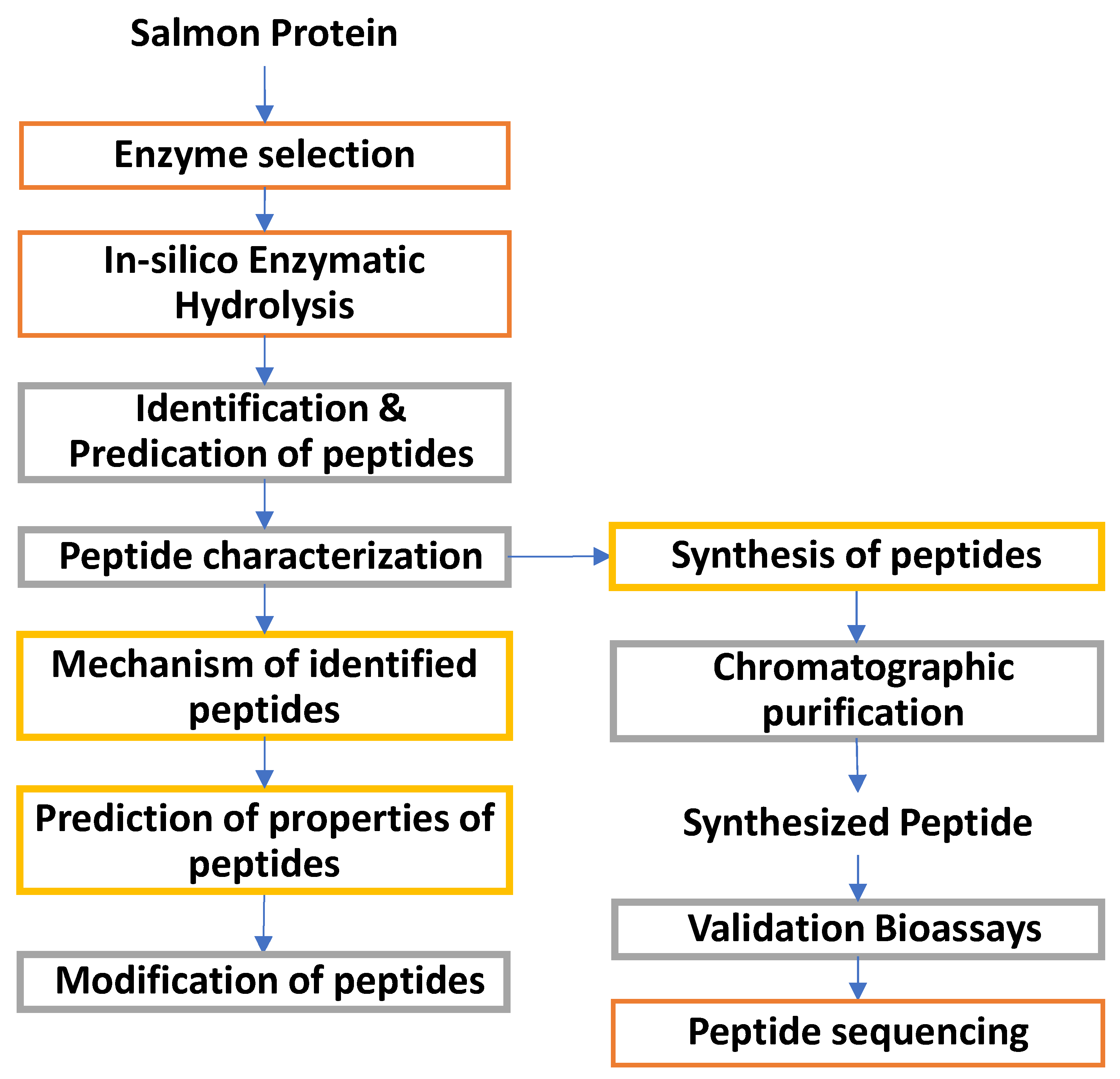
Figure 9.
Schematic representation of QSAR modelling.
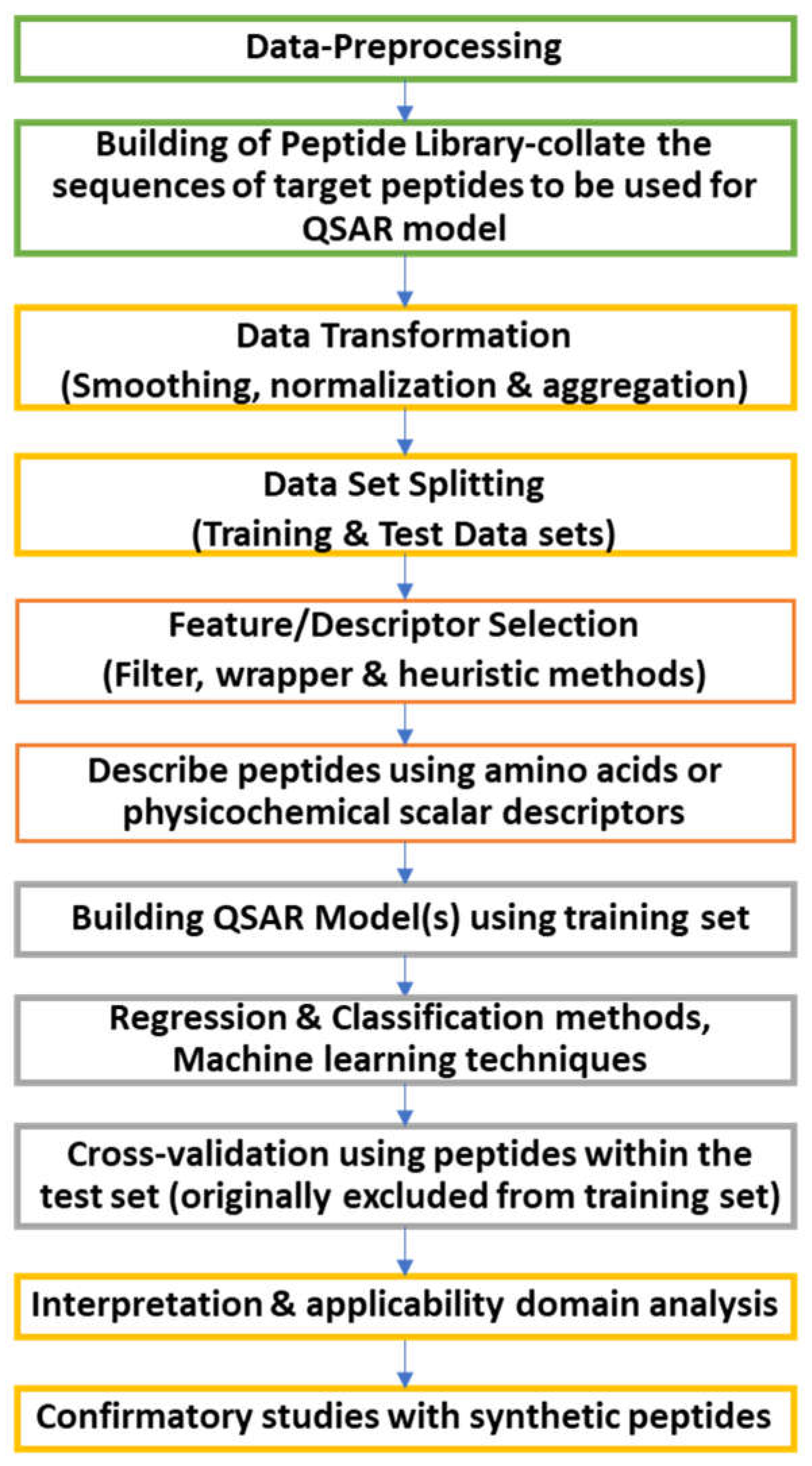
Figure 10.
Peptide purification technologies.
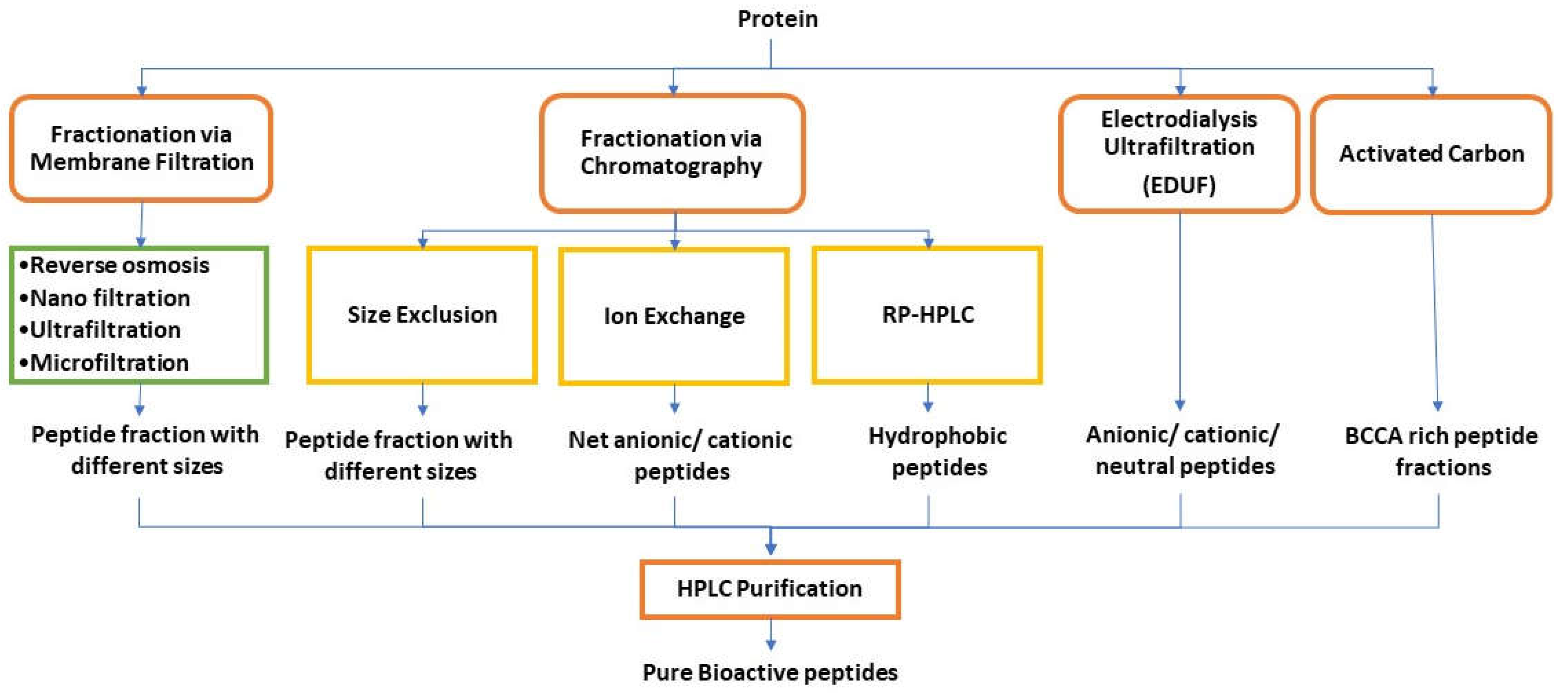

Table 1.
Proximate composition (%) of salmon body parts.
| Sample | Moisture | Protein | Lipid | Ash | References |
|---|---|---|---|---|---|
| Gut | 50-60.45 | 10.38-13.8 | 22.12-30 | 1.8-1.94 | [33,34,35] |
| Head | 62-63.35 | 11.31-14 | 17-21.86 | 3.51-4.4 | |
| Frame/ trimmings | 55-57.18 | 16.36-19 | 22.65-27 | 3.64-6 | |
| Fillet | 68.80-75 | 18.7-20.90 | 11.30-21.3 | 1.80-2.2 | |
| Whole fish | 64.50-75 | 16.7-17.90 | 17.30-22.12 | 1.88-2.2 |
Table 2.
Enzymatic hydrolysis of salmon protein.
| Enzymes | Salmon Part | pH | Time (min) | Temperature (°C) | Enzyme Concentration | Protein Yield (%) | Reference |
|---|---|---|---|---|---|---|---|
| Alcalase | Muscle | 7.5 | 180 | 40 | 7.5% | 86.92-88.39 | [9] |
| Flavourzyme | 78.95-84.26 | ||||||
| Corolase PN-L | 72.88-79.32 | ||||||
| Corolase 7089 | 82.41-86.48 | ||||||
| Endogenous | 71.67-79.12 | ||||||
| Alcalase | Head | 8 | 120 | 55 | 5.5% | 71.00 | [59] |
| Alcalase | Skin | 8.39 | 120 | 55 | 2.5% | 77.03 | [60] |
| Neutrase Alcalase Flavourzyme and Protamex | Skin | 6.7-8.0 | 240 | 53 | 10000 U/g | 45.11 | [61] |
| Endogenous | Viscera | N/A | 60 and 120 | 52 | 0.1% | 77.40 | [16] |
| Protamex +Endogenous | Viscera, head and frame | 82.80 | |||||
| Papain + Bromelain + Endogenous | Viscera, head and frame | 0.05+0.05% | 86.0 | ||||
| Protamex + Endogenous | Viscera | 0.1% | 75.50 | ||||
| Papain plus Bromelain + Endogenous | Viscera | 76.60 | |||||
| Papain plus Bromelain | Viscera | 69.90 | |||||
| Corolase PP | Backbone | N/A | 120 | 50 | 0.1% | 11.50 | [62] |
| Corolase 7089 | 0.1% | 9.30 | |||||
| Protamex | 0.1% | 9.40 | |||||
| Bromelain 400 GDU/g + Papain 100TU/mg | 0.05+0.05% | 11.60 | |||||
| Protex 6L | 0.1% | 8.50 | |||||
| Seabzyme L200 | 0.1% | 9.90 | |||||
| Trypsin | 0.1% | 12.10 | |||||
| Alcalase | Head and backbone | 6.5 | 120 | 50 | 5.5 - 88 U/g | 64.70 | [63] |
| Promod 671L | 66.00 | ||||||
| Protex 7L | 62.20 | ||||||
| Protamex | Frame | 6.5 | 60 | 50-56 | 10-90 AU/kg | 43.00-61.00 | [64] |
| Alcalase | Head, trimming, and frame | 8.98 | 180 | 64.2 | 0.2% |
84 |
[65] |
| Alcalase and papain | Frame | 8.0 | 240 | 85 | 1-3% | 79.2-82.01 | [66] |
| Bromelain BR1200 and FoodPro PNL | Backbone and head | N/A | 60 | 50 | 10 U/g |
89.3-92.1 |
[67] |
Table 3.
Amino acid composition of Atlantic salmon protein hydrolysates.
| Amino acids | Hydrolysates (mg/100 g) | Skin (g/100 g) |
Trimmings (g/100 g) | Viscera+head+frame (mg/g) | Viscera (mg/g) |
||
| Pepsin | Pepsin + pancreatin | Thermolysin | |||||
| Aspartic acid | 0.4 ± 0.2 | 11.6 ± 1.1 | 12.4 ± 0.5 | 5.72 | 7.31 | 77.8 ± 0.8 | 77.0 ± 0.1 |
| Threonine | 2.9 ± 0.2 | 6.0 ± 0.3 | 17.8 ± 0.8 | 2.03 | 3.23 | 39.3 ± 0.1 | 42.4 ± 0.3 |
| Serine | 4.8 ± 0.2 | 6.4 ± 0.3 | 21.8 ± 0.6 | 3.92 | 3.19 | 39.2 ± 0.2 | 41.2 ± 0.9 |
| Glutamic acid | 3.1 ± 0.1 | 29.0 ± 1.2 | 54.2 ± 5.4 | 9.05 | 11.07 | 113.1 ± 1.3 | 109.6 ± 1.9 |
| Glycine | 13.7 ± 0.8 | 15.2 ± 1.0 | 33.0 ± 2.7 | 22.6 | 7.07 | 85.8 ± 1.4 | 66.5 ± 0.0 |
| Alanine | 24.7 ± 1.5 | 35.5 ± 0.9 | 95.0 ± 5.3 | 7.81 | 5.22 | 57.0 ± 0.0 | 53.4 ± 2.0 |
| Valine | 2.5 ± 0.1 | 15.2 ± 0.3 | 18.4 ± 1.7 | 1.71 | 3.83 | 42.6 ± 0.2 | 45.6 ± 0.1 |
| Methionine | 1.3 ± 0.1 | 11.8 ± 0.4 | 31.9 ± 1.6 | 2.46 | 2.66 | 24.3 ± 0.1 | 24.4 ± 0.8 |
| Isoleucine | 1.2 ± 0.1 | 21.9 ± 2.2 | – | 1.05 | 2.98 | 36.0 ± 0.2 | 37.4 ± 0.0 |
| Leucine | 2.4 ± 0.1 | 84.3 ± 1.4 | 69.8 ± 2.3 | 1.75 | 4.97 | 57.9 ± 0.4 | 62.6 ± 0.3 |
| Tyrosine | 1.9 ± 0.1 | 76.9 ± 4.3 | 11.4 ± 0.5 | 0.35 | 1.76 | 18.9 ± 0.5 | 22.2 ± 0.2 |
| Phenylalanine | 3.1 ± 0.3 | 76.3 ± 5.3 | 137.0 ± 4.0 | 1.71 | 2.53 | 31.2 ± 0.2 | 33.9 ± 0.3 |
| Histidine | 16.4 ± 0.9 | 21.0 ± 0.8 | 62.4 ± 2.7 | 1.33 | 3.88 | 26.3 ± 0.2 | 23.2 ± 0.7 |
| Arginine | 1.3 ± 0.6 | 196 ± 14 | – | 7.8 | 5.04 | 56.2 ± 0.3 | 50.8 ± 0.7 |
| Tryptophan | – | 18.3 ± 0.5 | – | 8.2 ± 0.0 | 10.4 ± 0.0 | ||
| Lysine | – | – | – | 2.93 | 6.4 | 71.2 ± 0.6 | 69.4 ± 0.4 |
| Proline | – | – | – | 10.63 | 3.98 | 49.2 ± 0.5 | 47.0 ± 0.3 |
| Cysteine | 0.49 | 0.94 | 9.4 ± 0.0 | 12.4 ± 0.0 | |||
| Reference | [84] | [85] | [16] | ||||
Table 4.
Atlantic Salmon derived bioactive peptides.
| Peptide/AA | Activities | Reference | ||||
|---|---|---|---|---|---|---|
| ACE IC50 | DPP-IV IC50 | ORAC (µmol TE/µmol peptide) |
AIA (mM) | HRSA (µg/mL) | ||
| GPAV | 415.91±2.65 (µM) | 245.58 ± 7.15 (µM) | 9.51 ± 1.40 | [17] | ||
| VP | 1215.15 ± 83.14 (µM) | 758.15 ± 12.45 (µM) | 19.45 ± 2.15 | |||
| VC | 134.45 ± 5.15 (µM) | 5413.45 ± 62.15 (µM) | 3.45 ± 0.78 | |||
| YP | 5.21 ± 0.94 (µM) | 7564.02 ± 42.45 (µM) | 17.18 ± 1.54 | |||
| FF | 59.15 ± 0.54 (µM) | 546.84 ± 8.15 (µM) | 8.47 ± 1.05 | |||
| PP | 1912.46 ± 63.15 (µM) | 4343.48 ± 29.78 (µM) | 12.48 ± 1.47 | |||
| W | 135.48 ± 5.48 (µM) | 438.15 ± 10.97 (µM) | 4.31 ± 0.19 | |||
| F | 125.15 ± 2.64 (µM) | 295.15 ± 6.45 (µM) | 4.59 ± 0.89 | |||
| Y | 182.84 ± 0.52 (µM) | 75.15 ± 0.84 (µM) | 2.74 ± 0.65 | |||
| GF | 178.14 ± 24.51 (µM) | 1547.15 ± 34.15 (µM) | 19.74 ± 1.01 | [18] | ||
| GPVA | 445.61 ± 6.94 (µM) | 264.74 ± 1.59 (µM) | 9.48 ± 0.94 | |||
| GGPAGPAV | 673.16 ± 15.03 (µM) | 8139.11 ± 134.68 (µM) | 5.47 ± 0.94 | |||
| R | 98.04 ± 0.15 (µM) | 110.44 ± 0.47 (µM) | 4.71 ± 0.09 | |||
| PAY | 0.75 | [111] | ||||
| VWDPPKFD | 9.12 ± 1.07 (µM) | [109] | ||||
| FEDYVPLSCF | 13.72 ± 2.54 (µM) | |||||
| FNVPLYE | 6.79 ± 0.78 (µM) | |||||
| AP | 0.060 ± 0.001 mg/mL | [110] | ||||
| VR | 0.332 ± 0.005 mg/mL | |||||
| GAAGR | ||||||
| AGPS | ||||||
| VDGK | ||||||
| RER | ||||||
| LN | ||||||
| VTGK | ||||||
| GHAGE | ||||||
| VGGK | ||||||
| GHGR | ||||||
| GAPE | 49.6 (µM) | [112] | ||||
| GPGA | 41.9 (µM) | |||||
| YYGYTGAFR | 1.21 mg/mL | [113] | ||||
| LDKVFR | 0.10 mg/mL | |||||
| VLATSGPG | 0.18 mg/mL | |||||
| PA | 91.3 | [114] | ||||
| IVY | 0.48 (µM) | [115] | ||||
| VW | 1.4 (µM) | |||||
| IY | 2.1 (µM) | |||||
| IW | 4.7 (µM) | |||||
| VY | 7.1 (µM) | |||||
| TVY | 15 (µM) | |||||
| VFPS | 0.46 (µM) | |||||
| VTVNPYKWLP | 5.5 (µM) | |||||
| IWHHT | 5.8 (µM) | |||||
| YALPHA | 9.8 (µM) | |||||
| ALPHA | 10 (µM) | |||||
| ACE: Angiotensin-I converting enzyme inhibition; DPP-IV: Dipeptidyl-peptidase IV (DPP-IV)-inhibitory activity; ORAC: Oxygen radical absorbance capacity; TE: Trolox equivalent; AIA: Anti-Inflammatory Activity; HRSA: Hydrogen Radical Scavenging Activity | ||||||
Table 5.
Common in-silico databases reported in the literature.
| Databases | Name | Webpage | Purpose | Reference |
|---|---|---|---|---|
| Protein/peptide sequence databases | BIOPEP | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Protein and peptide database | [119,123,124,125,126,] |
| PepBank | http://pepbank.mgh.harvard.edu/ | Peptide database | ||
| BioPD | http://biopd.bjmu.edu.cn/ | Peptide database | ||
| SwePep | http://www.swepep.org/ | Sweden peptide database | ||
| EROP-Moscow | http://erop.inbi.ras.ru/ | Peptide database | ||
| MilkAMP | http://milkampdb.org/home.php | Dairy antimicrobial peptide database | ||
| UniProtKB | http://www.uniprot.org/ | Primary sequence and structural information for proteins | ||
| NCBI Protein database | http://www.ncbi.nlm.nih.gov | Basic sequence information for proteins | ||
| PeptideDB | http://www.peptides.be/ | Peptide database | ||
| AVPdb | http://crdd.osdd.net/servers/avpdb/index.php | Database of anti-viral peptides | ||
| CAMPR3 | http://www.camp.bicnirrh.res.in/ | Collection of anti-microbial peptides | ||
| HIPdb | http://crdd.osdd.net/servers/hipdb/index.php | Database of HIV inhibiting peptides | ||
| BACTIBASE | http://bactibase.hammamilab.org/main.php | Database of bacteriocins | ||
| APD | https://wangapd3.com/main.php | Antimicrobial peptide database | ||
| http://aps.unmc.edu/AP/ | ||||
| BitterDB | http://bitterdb.agri.huji.ac.il/dbbitter.php | Database for bitter peptides | ||
| In silico digestion Tools | BIOPEP | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Tools for peptide predication | |
| PeptideCutter | http://web.expasy.org/peptidecutter/ | Predict peptide cleavage sites | ||
| POPS | http://pops.csse.monash.edu.au/pops-cgi/index.php | Tool for modelling and predicting protease specificity | ||
| http://www.mybiosoftware.com/pops-2-1-1-prediction-protease-specificity.html | ||||
| NeuroPred | http://stagbeetle.animal.uiuc.edu/cgi-bin/neuropred.py | Prediction of cleavage sites and mass from neuropeptide precursors | ||
| CAMPSign | http://www.campsign.bicnirrh.res.in/ | Identification of AMPs using AMP family signatures | ||
| Enzyme Predictor | http://bioware.ucd.ie/~enzpred/Enzpred.php | Evaluation tool to determine the cleavage ability of enzymes | ||
| Potential bioactivity prediction | PeptideRanker | http://bioware.ucd.ie/ ∼compass/biowareweb/ | Pediction of bioactive peptides | |
| BIOPEP | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Pediction of bioactive peptides | ||
| AntiBP2 | http://www.imtech.res.in/raghava/antibp2/ | Antibacterial peptide prediction tool | ||
| AVPpred | http://crdd.osdd.net/servers/avppred/submit.php | Collection and prediction of antiviral peptides | ||
| AVP-IC50Pred | http://crdd.osdd.net/servers/ic50avp/index.html | Prediction of peptide antiviral activity in terms of IC50 | ||
| Toxicity/allergenicity prediction | ||||
| ToxinPred | http://crdd.osdd.net/raghava/toxinpred/ | Predicting the toxicity of peptides | ||
| BIOPEP | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Allergenic protein database | ||
| AlgPred | http://crdd.osdd.net/raghava/algpred/ | Predicting allergenic proteins and peptides | ||
| Allerdictor | https://openebench.bsc.es/tool/allerdictor | Allergen prediction tool | ||
| EPIMHC | http://bio.med.ucm.es/epimhc/ | Database of Naturally Processed MHC-restricted Peptide Ligands and Epitopes for Customized Computational Vaccinology | ||
| Physicochemical characteristics prediction | Expasy-Compute pI/Mw | http://web.expasy.org/compute_pi/ | A tool to compute the theoretical pI (isoelectric point) and MW (mol weight) | |
| ProtParam | http://web.expasy.org/protparam/ | A tool to compute the grand average of hydropathicity (GRAVY) and Instability index | ||
| Peptide Structure Prediction Server | PepDraw | http://www.tulane.edu/~biochem/WW/PepDraw/ | A tool to compute net charge and hydrophobicity | |
| Pep-Fold | https://bioserv.rpbs.univ-paris-diderot.fr/services/PEP-FOLD/ | Predicting peptide structures from amino acid sequences. | ||
| PEPstrMOD | http://osddlinux.osdd.net/raghava/pepstrmod/ | Predicts the tertiary structure of small peptides with sequence length varying between 7 to 25 residues | ||
| I-TASSER | https://zhanglab.ccmb.med.umich.edu/I-TASSER/ | Protein structure prediction and structure-based function annotation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



