
Submitted:
15 April 2023
Posted:
17 April 2023
You are already at the latest version
Abstract
Introduction: Circulating tumor DNA (ctDNA) is newly diagnosed tumor DNA that can easily represent a tumor’s genetic and epigenetic change. Pheochromocytomas (PCCs) and Paragangliomas (PGLs) are rare tumors of adrenal gland tissue that have the potential to be detected by ctDNA. We aimed to study the potential of the methylation status of RDBP, SDHB, and SDHC genes in ctDNA of PCCs/PGLs patients as a diagnostic biomarker.
Materials and Methods: Clinical data, fresh frozen tissue, the blood of 12 PCCs/PGL patients, and the blood of 12 age/sex-matched normal patients were collected. The methylation status of RDBP, SDHB, and SDHC was compared between cases and controls by MS-HRM analysis.
Results: Amongst six promoter regions of RDBP, SDHB, and SDHC, promoter methylation quantification of SDHCa and RDBPb was significantly different between PCCs/PGLs and controls. SDHCa was methylated in 49.93% of PCCs/PGLs cases vs. 8.33 % of control samples, p-value: 0.026, area under curve AUC=0.757, and RDBPb in 74.9% of PCCs/PGLs cases vs. 25.0% of control samples, p-value: 0.032, AUC=0.750.
Conclusions: This study suggests the ctDNA potential for a less invasive source of tumor epigenetic modification in PCCs/PGLs malignancies. The SDHCa and RDBPb hypermethylation warrant further exploration as diagnostic tools for PCCs/PGLs.
Keywords:
ctDNA
; liquid biopsy
; Methylation
; SDHCa
; RDBPb
; Pheochromocytoma
; Paraganglioma
Introduction
Pheochromocytomas arise from chromaffin cells in the adrenal medulla, and PGLs arise from chromaffin cells in the ganglia of the autonomic nervous system. Paragangliomas originate from sympathetic or parasympathetic ganglia in the abdomen, thorax, and pelvis (1, 2). The annual incidence of PCCs/PGLs is about 0.8 per 100,000 people (3). Nevertheless, this is probably an underestimation because half of PCCs/PGLs might be diagnosed at autopsy (4). PCCs/PGLs mostly happen in the 3rd and 4th decades of age, equally in both genders (1). There is an extreme need to optimize biomarkers, mainly genetically and epigenetically, to discriminate the PCCs/PGLs and plan the exact treatment strategy at the precise time.
Recently, the new aspect of Liquid Biopsy (LB) has brought an exceptional tumor marker as the real-time representation of the tumor. Numerous studies focus on genetic and epigenetic circulating tumor DNA (ctDNA) as the tumor markers (5). However, the latest discoveries related to ctDNA genetic modification and circulating tumor cells (CTCs) in PCCs/PGLs are limited (6, 7).
More than genetic modifications, there are some epigenetic variations in DNA molecules that can change the gene expression of the tumoral cells with no change in the exact DNA sequences (8). Intergenic CpG sites are classically methylated in normal tissue. At the same time, they are often hypomethylated in the tumor, resulting in transcriptional changes by interfering with transcription factor binding and changing the euchromatin to heterochromatin (9, 10). DNA methylation patterns in some specific genes are reported to be associated with progression-free survival (PFS) and overall survival (OS) in several cancers (11). Three target genes were selected in this study, RDBP (Negative Elongation Factor Complex Member E) as the essential component of the NELF complex, a complex that negatively regulates the elongation of transcription by RNA polymerase II. SDHB and SDHC encoding subunits of succinate dehydrogenase are significant cause of hereditary paraganglioma and pheochromocytoma and demonstrate that these genes are classic tumor suppressors. Succinate dehydrogenase is a heterotetrameric protein complex and a component of the Krebs cycle and the mitochondrial respiratory chain (succinate: ubiquinone oxidoreductase or complex II). The ctDNAs can be considered a potential non-invasive source of methylation change in PCCs/PGLs. In this study, we examined the methylation quantification of PCCs/PGLs in ctDNA as the tumor transformation biomarker.
Materials and Methods
We ran this study on 12 consecutive PCCs/PGLs patients (cases) and 12 non-cancerous patients (controls). All patients signed the informed consent and all procedures were under National Institute for Medical Science Development Ethics Committee (IR.NIMAD.REC.1397.452). The blood samples were gathered in EDTA-containing vials before surgery for ctDNA analysis. Tumor tissues were captured during surgery in the Department of Surgery in Shariati hospital as the origin of the genomic DNA (gDNA) source. All surgically resected tissues were snap-frozen in liquid nitrogen for further analysis.
Tissue and ctDNA extraction:
Fresh frozen tissues of PCCs/PGLs were removed and kept in liquid nitrogen for a maximum of 1 month, then tissue DNA (gDNA) was extracted using DNeasy Blood & Tissue Kit (Qiagen, Netherlands, Cat No: 69504) (12). For ctDNA extraction, about 4-6 ml blood samples were collected from both PCCs/PGLs and control groups, and the ctDNA extraction was done within two hours. For ctDNA extraction, the plasma was first detached by centrifugation of blood at 2800 rpm for fifteen minutes using the Ficoll separation method. Then, plasma was moved to a sterilized tube, and ctDNA was extracted according to the Norgen Plasma/Serum Cell-Free Circulating DNA Purification Midi Kit protocol (Canada, Cat No: 55600). Lastly, the purity and concentration of ctDNA were determined by optical density at 260 and 280nm by Thermo Scientific™ NanoDrop™ spectrophotometers 2000c (Thermo Fisher Scientific Inc.). Both extracted ctDNA and gDNA were stored in the freezer at -80°C for further analysis.
Bisulfite modification
For methylation quantification analysis, ctDNA and its counterpart gDNA from each candidate PCCs/PGLs and ctDNA from controls were treated by the “EZ DNA Methylation-Gold™ Kit” (Zymoresearch, USA, Cat No: D5005) according to the manufacturer’s protocol. For the high methylation resolution melting analysis (HRM), six promoter regions of three target genes, RDBP, SDHB, and SDHC, were selected by specific primers (Table 1).
The MS-HRM program includes three main steps: holding step (94°C for 20 minutes), going to 40 cycles of 94°C for 10 seconds, several annealing temperatures depending on the primer (varying from 45°C to 60°C) for 35 seconds, and extension time of 72°C for 35 seconds leading to the final step of the melting curve. The melting curve step was made by heating the samples to 90°C for 15 seconds and 60°C for one minute leading to 65°C for 15 seconds, and afterward continuously heated up to 95°C with the acquisition of data during every 0.3°C rises in temperature.
All reaction mixture consisted of 10µl of master mix (Amplicon, Cat No: A325406), 20 pmol of each primer, and 2µl (almost 10ng) of bisulfite modified DNA template in the whole volume of 20 µl. Moreover 0%, 25%, 50%, 75%, and 100% methylated controls were run in each reaction. Standard curves with identified methylation ratios were comprised in all assays by assuming the methylation ratio of the unknown target. The ABI Step One Plus system did MS-HRM tests in triplicates.
Statistical analysis
Samples were considered hyper-methylated for curves between 25% and 100% of methylation compared to controls and non-methylated when the methylation amount was near 0% compared to the control. The comparison was made between two main groups: the case group comprising 12 PCCs/PGLs patients and the control group comprising 12 non-cancerous patients. The specificity and sensitivity of each target CpG site were assessed through a 2×2 table. The correlation of mutation and methylation of ctDNAs and their tissue counterparts were performed through spearman correlation analysis, and ctDNA’s methylation of PCCs/PGLs patient’s Receiver Operating Characteristics (ROC) curves were constructed. Statistical Package did all statical analyses for Science Software (SPSS) (version 17.0; SPSS Inc. Chicago, Illinois).
Results
Demographic characteristics of PCCs/PGLs patients and controls are presented in Table 2. The age of all patients ranged from 26 to 60. Among all 12 PCCs/PGLs, tumor size varied from 0.5-6 cm, and three patients (25%) were defined as homogeneous and hypoechoic solid lesions with well-defined borders. PCCs/PGLs patients showed significantly higher systolic blood pressure (SBP), diastolic blood pressure (DBP), and also considerably lower weight and body mass index (BMI).
For each run of MS-HRM, ten wells were allocated to the methylation controls 0%, 25%, 50%, 75%, and 100%, and test samples were run in triplicate. The melting curve was considered for additional analysis to determine the methylation status (Figure 1). The methylation level of each sample was estimated by comparing each sample’s normalized melt curve with the controls’ normalized melt curves.
The methylation status of all six target regions in the case and controls are presented in Table 3 and Figure 2. Among six promoter regions within three candidate genes, the SDHCa (the promoter region harboring initial ATG code) and RDBPb methylation statuses were hypermethylated (more than 25 % methylation) in PCCs/PGLs patients compared to the control group, and p-value 0.026 and 0.03, respectively (13). Bisulfite Pyrosequencing confirmed all results on tumor tissue DNA as an additional technique.
The maximum value for kappa observed was 0.919 between SDHBb and SDHCa and 0.805 between RDBPb and SDHCb (Table 4).
According to possible interpretation of Kappa, Poor agreement = Less than 0.20, Fair agreement = 0.20 to 0.40, Moderate agreement = 0.40 to 0.60, Good agreement = 0.60 to 0.80 and, Very good agreement = 0.80 to 1.00. *The highest kappa agreement score.
ROC curves were plotted based on the percentage of methylated reference (PMR) values in PCCs/PGLs and the control group to indicate if any of the targeted six promoter regions in circulating plasma ctDNA can be considered as PCCs/PGLs specific diagnostic markers (Figure 3).
The sensitivity and specificity of target CpG sites were defined based on correctly classifying subjects with methylated promoters into PCCs/PGLs. The area under the curve (AUC) ranged from 0.455 in RDBPa to 0.757 in SDHCa and then 0.750 in RDBPb (Table 5).
Discussion
Several genetic mutations are considered important molecular diagnostic biomarkers in PCCs/PGLs. Our pioneering study represents the role of specific genes DNA methylation as a PCCs/PGLs potential diagnostic tool. Herein, we report SDHC1 hypermethylation in 49.93% of PCCs/PGLs cases vs. 8.33 % of control samples, p-value: 0.026; and RDBP2 promoter region 74.9% hypermethylation in cases vs. 25.0% in controls, p-value: 0.03. These target regions have AUC 0.750 in RDBP2 and 0.757 in SDHC1 promoters.
Changes in gene expression patterns that trigger the cell to malignancy can result from epigenetic modification (14, 15). Methylation-Sensitive High Resolution Melting (MS-HRM) is an in-tube; PCR-based method to detect methylation levels at specific loci of interest. A unique primer design facilitates a high sensitivity of the assays enabling detection of down to 0.1-1% methylated alleles in an un-methylated background. The only study so far in which the ctDNA was challenged as a PCCs/PGLs tumor genetic indicator was done by Wang et al. in 2018, and to the best of our knowledge, our study is the only one that seeks methylation of ctDNA (6). Nowadays, it has become common knowledge that ctDNA, as the liquid biopsy main component, has excellent potential to non-invasively show the tumor’s genetic status (16-19).
The global DNA methylation array indicates three distinct clusters: M1–3. M1 comprises tumors with SDHx mutations and hypermethylation, M2 VHL-mutated tumors, and M3 tumors with NF1 and RET mutations and hypomethylation (11, 20). Epithelial to Mesenchymal Transition (EMT) can be activated in metastatic PCCs/PGLs by SDHB gene mutations (21). It was shown by Astuti et al. that SDHB was hypermethylated in 21% of primary neuroblastomas and 32% of PCCs/PGLs (22). Our results indicate 25.0% SDHB promoter region hypermethylation and more than 75% of CpG sites harboring initial ATG.
However, based on posterior microsatellite instability and hypermethylation promoter studies, there is still doubt that SDHB methylation can play a role once it is unlikely to be related to either tumor initiation or progression in neuroblastoma (23). We have considered two upstream promoter regions of initial ATG with no difference in methylations between PCCs/PGLs and control patients. Contradictory, it was reported that EMT hypermethylation and in vitro acquisition of metastatic properties could be the consequence of SDHB loss of function (24). SDHB mutation might change the methylation status of methyltransferase enzymes like MGMT (25-28).
Considering gene silencing by promoter CpG methylation as an essential epigenetic mechanism in tumorigenesis, our results indicate that SDHC promoter hypermethylation could play a role in developing PCCs/PGLs. The altered expression of SDHC has been reported several times as the consequence of genetic and epigenetic changes (29-32). SDHC promoter methylation can cause SDHC inactivation, so it is essential to consider epigenetic changes and functional readouts in the genetic evaluation of patients (33). SDH-loss cells are selectively vulnerable to LDH genetic knock-down or chemical inhibition, suggesting that LDH inhibition may be an effective therapeutic strategy for SDH-loss and SDHC-loss transcriptional change correlate with baseline expression values in normal cells (34). Our results indicate that the CpG sites far from initial ATG are hypermethylated in PCCs/PGLs, and its methylation agrees with SDHB and RDBP. The RDBP gene is responsible for coding negative elongation factor E (NELF) as a complex that negatively regulates the elongation of transcription by RNA polymerase II (35). The first large-scale study of DNA methylation in metastatic PGL by de Cubas and colleagues supports that RDBP could be used for stratifying patients according to the risk of developing metastases (36).
Our study indicated RDBP promoter hypermethylation in PCCs/PGLs, in line with Backman et al., which suggested hypermethylated RDBP in metastasizing PGLs regardless of mutational status (11). RDBP hypermethylation should be further explored as malignancy and survival markers in patients with PCCs/PGLs (37-39). RDBP methylation can trigger the PCCs/PGLs to be malignant, and interestingly in 5 malignant patients in our study, the CpG sites in promoter regions harboring ATG were hypermethylated, similarly to Yong Joon Suh et al. findings (40). RDBP methylation status can predict outcome in PCCs/PGLs and the potential of targeted therapy (37, 41). Unlike SDHx which are methylated or mutated in several tumor types, the RDBP hypermethylation has been just reported in PCCs/PGLs and of the initial 86 candidate CpGs, from 47 genes, just RDBP was confirmed and could be used for stratifying patients according to the risk of developing metastases (36). However, this type of test still has significant limitations since it is only possible to be performed in centers with the technology available implicating in higher hospital structure and investment, which is not (yet) the reality of many centers worldwide. The primary study limitation was that a genetic analysis of ctDNA is very challenging because ctDNA is often highly fragmented and often limited in yield.
Conclusions
In line with current literature, our results support SDHCa and RDBPb hypermethylation in ctDNA of PCCs/PGLs as potential diagnostic and prognostic tools.
Author Contributions
S.M.K.A. and R.H. are principal investigators, L.O.R. edited the manuscript, M.A.P. and M.E. analyzed the data, S.H.N. and S.M.T. provided data and data curation, and F.K.H. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
Thanks to the urology research center, Tehran University of medical sciences, and the National Institute for Medical. Research Development (NIMAD) Grant number 977079.
Conflicts of Interest
All authors declare there is not any conflict of interest for this publication.
References
- Guerrero MA, Schreinemakers JM, Vriens MR, Suh I, Hwang J, Shen WT, et al. Clinical spectrum of pheochromocytoma. Journal of the American College of Surgeons. 2009;209(6):727-732. [CrossRef]
- Neumann HP, Young Jr WF, Eng C. Pheochromocytoma and paraganglioma. New England Journal of Medicine. 2019;381(6):552-565.
- Beard C, Sheps S, Kurland L, Carney J, Lie J, editors. Occurrence of pheochromocytoma in Rochester, Minnesota, 1950 through 1979. Mayo Clinic Proceedings; 1983.
- Sutton M, Sheps S, Lie J, editors. Prevalence of clinically unsuspected pheochromocytoma. Review of a 50-year autopsy series. Mayo Clinic Proceedings; 1981. [CrossRef]
- Aghamir SMK, Heshmat R, Ebrahimi M, Khatami F. Liquid Biopsy: The Unique Test for Chasing the Genetics of Solid Tumors. Epigenetics Insights. 2020;13:2516865720904052. [CrossRef]
- Wang L, Li Y, Guan X, Zhao J, Shen L, Liu J. Exosomal double-stranded DNA as a biomarker for the diagnosis and preoperative assessment of pheochromocytoma and paraganglioma. Molecular cancer. 2018;17(1):1-6. [CrossRef]
- Khatami F, Tavangar SM. Current diagnostic status of pheochromocytomaand future perspective: A mini review. Iranian journal of pathology. 2017;12(3):313. [CrossRef]
- Kulis M, Esteller M. DNA methylation and cancer. Advances in genetics. 70: Elsevier; 2010. p. 27-56.
- Khatami F, Mohammadamoli M, Tavangar SM. Genetic and epigenetic differences of benign and malignant pheochromocytomas and paragangliomas (PPGLs). Endocrine regulations. 2018;52(1):41-54. [CrossRef]
- Nazar E, Khatami F, Saffar H, Tavangar SM. The Emerging Role of Succinate Dehyrogenase Genes (SDHx) in Tumorigenesis. International Journal of Hematology-Oncology and Stem Cell Research. 2019;13(2):72. [CrossRef]
- Backman S, Maharjan R, Falk-Delgado A, Crona J, Cupisti K, Stålberg P, et al. Global DNA Methylation Analysis Identifies Two Discrete clusters of Pheochromocytoma with Distinct Genomic and Genetic Alterations. Sci Rep. 2017;7:44943-44943. [CrossRef]
- Fatemeh Khatami BL, Ramin Heshmat, Shirzad Nasiri, Hiva Saffar, Gita Shafiee, Azam Mossafa, Seyed Mohammad Tavangar. Promoter Methylation of Four Tumor Suppressor Genes in Human Papillary Thyroid Carcinoma. Iranian Journal of Pathology. 2018;In Press. [CrossRef]
- Brigliadori G, Foca F, Dall’Agata M, Rengucci C, Melegari E, Cerasoli S, et al. Defining the cutoff value of MGMT gene promoter methylation and its predictive capacity in glioblastoma. Journal of Neuro-Oncology. 2016;128(2):333-339. [CrossRef]
- Baylin SB, Jones PA. Epigenetic determinants of cancer. Cold Spring Harbor perspectives in biology. 2016;8(9):a019505. [CrossRef]
- Chatterjee A, Rodger EJ, Eccles MR, editors. Epigenetic drivers of tumourigenesis and cancer metastasis. Seminars in cancer biology; 2018: Elsevier. [CrossRef]
- Ma M, Zhu H, Zhang C, Sun X, Gao X, Chen G. “Liquid biopsy”—ctDNA detection with great potential and challenges. Annals of translational medicine. 2015;3(16). [CrossRef]
- Khatami F, Larijani B, Tavangar SM. The presence of tumor extrachomosomal circular DNA (ecDNA) as a component of liquid biopsy in blood. Medical hypotheses. 2018;114:5-7. [CrossRef]
- Khatami F, Aghaii M, Aghamir SMK. Prime editing: The state-of-the-art of genome editing. Meta Gene. 2020;24:100661. [CrossRef]
- Tamehri Zadeh SS, Taheri D, Shivarani S, Khatami F, Kazemi R. Liquid Biopsy in Prostate Cancer Diagnosis and Prognosis: A Narrative Review. Translational Research In Urology. 2020;2(4):139-146. [CrossRef]
- Letouzé E, Martinelli C, Loriot C, Burnichon N, Abermil N, Ottolenghi C, et al. SDH mutations establish a hypermethylator phenotype in paraganglioma. Cancer cell. 2013;23(6):739-752. [CrossRef]
- Loriot C, Burnichon N, Gadessaud N, Vescovo L, Amar L, Libé R, et al. Epithelial to mesenchymal transition is activated in metastatic pheochromocytomas and paragangliomas caused by SDHB gene mutations. The Journal of Clinical Endocrinology & Metabolism. 2012;97(6):E954-E962. [CrossRef]
- Astuti D, Morris M, Krona C, Abel F, Gentle D, Martinsson T, et al. Investigation of the role of SDHB inactivation in sporadic phaeochromocytoma and neuroblastoma. British journal of cancer. 2004;91(10):1835-1841. [CrossRef]
- Grau E, Oltra S, Orellana C, Hernández-Martí M, Castel V. There is no evidence that the SDHB gene is involved in neuroblastoma development. Oncology Research Featuring Preclinical and Clinical Cancer Therapeutics. 2005;15(7-8):393-398. [CrossRef]
- Loriot C, Domingues M, Berger A, Menara M, Ruel M, Morin A, et al. Deciphering the molecular basis of invasiveness in Sdhb-deficient cells. Oncotarget. 2015;6(32):32955-32965. [CrossRef]
- Ricci R, Martini M, Ravegnini G, Cenci T, Milione M, Lanza P, et al. Preferential MGMT methylation could predispose a subset of KIT/PDGFRA-WT GISTs, including SDH-deficient ones, to respond to alkylating agents. Clinical Epigenetics. 2019;11(1):2. [CrossRef]
- Liu Y, Pang Y, Caisova V, Ding J, Yu D, Zhou Y, et al. Targeting NRF2-governed glutathione synthesis for SDHB-mutated pheochromocytoma and paraganglioma. Cancers. 2020;12(2):280. [CrossRef]
- Lamy C, Hadoux J, Durand S, Alghuzlan A, Riviere J, Lefevre D, et al. Preclinical evaluation of new therapeutic strategies on SDHB invalidated clones from human pheochromocytoma cells. AACR; 2019. [CrossRef]
- Rashedi S. Landscape of Circular Ribonucleic Acids in Urological Cancers. Translational Research Urology. 2021;3(2):45-47. [CrossRef]
- Gill AJ, Benn DE, Chou A, Clarkson A, Muljono A, Meyer-Rochow GY, et al. Immunohistochemistry for SDHB triages genetic testing of SDHB, SDHC, and SDHD in paraganglioma-pheochromocytoma syndromes. Human pathology. 2010;41(6):805-814. [CrossRef]
- Remacha L, Comino-Méndez I, Richter S, Contreras L, Currás-Freixes M, Pita G, et al. Targeted exome sequencing of Krebs cycle genes reveals candidate cancer–predisposing mutations in pheochromocytomas and paragangliomas. Clinical Cancer Research. 2017;23(20):6315-6324. [CrossRef]
- Shi C, Zeng Z, Zhao D, Hanzhong L, Miao Q, Zhu W, et al. Application of SDHB and SDHC immunohistochemistry in the differentiation of malignant and benign pheochromocytoma and paraganglioma. Chinese Journal of Endocrinology and Metabolism. 2018;34(6):472-478.
- Karimaei S, Oliveira Reis L. Cytotoxicity and Apoptotic Effect of Nisin as an Effective Bacteriocin on the Cancer Cells. Translational Research Urology. 2020;2(2):45-47. [CrossRef]
- Richter S, Klink B, Nacke B, de Cubas AA, Mangelis A, Rapizzi E, et al. Epigenetic mutation of the succinate dehydrogenase C promoter in a patient with two paragangliomas. The Journal of Clinical Endocrinology & Metabolism. 2016;101(2):359-363. [CrossRef]
- Smestad J, Hamidi O, Wang L, Holte MN, Al Khazal F, Erber L, et al. Characterization and metabolic synthetic lethal testing in a new model of SDH-loss familial pheochromocytoma and paraganglioma. Oncotarget. 2018;9(5):6109. [CrossRef]
- Narita T, Yamaguchi Y, Yano K, Sugimoto S, Chanarat S, Wada T, et al. Human transcription elongation factor NELF: identification of novel subunits and reconstitution of the functionally active complex. Molecular and cellular biology. 2003;23(6):1863-1873. [CrossRef]
- De Cubas AA, Korpershoek E, Inglada-Pérez L, Letouzé E, Currás-Freixes M, Fernández AF, et al. DNA methylation profiling in pheochromocytoma and paraganglioma reveals diagnostic and prognostic markers. Clinical Cancer Research. 2015;21(13):3020-3030. [CrossRef]
- Goncalves J, Lussey-Lepoutre C, Favier J, Gimenez-Roqueplo A-P, Castro-Vega LJ, editors. Emerging molecular markers of metastatic pheochromocytomas and paragangliomas. Annales d'endocrinologie; 2019: Elsevier. [CrossRef]
- Björklund P, Backman S. Epigenetics of pheochromocytoma and paraganglioma. Molecular and Cellular Endocrinology. 2018;469:92-97. [CrossRef]
- Oishi T, Iino K, Okawa Y, Kakizawa K, Matsunari S, Yamashita M, et al. DNA methylation analysis in malignant pheochromocytoma and paraganglioma. Journal of clinical & translational endocrinology. 2017;7:12-20. [CrossRef]
- Job S, Georges A, Burnichon N, Buffet A, Amar L, Bertherat J, et al. Transcriptome analysis of lncRNAs in pheochromocytomas and paragangliomas. The Journal of Clinical Endocrinology & Metabolism. 2020;105(3):898-907. [CrossRef]
- Nicolas M, Dahia P. Predictors of outcome in phaeochromocytomas and paragangliomas. F1000Research. 2017;6. [CrossRef]
Figure 1.
The MS-HRM for studying the methylation status of six CpG sites of three target genes.
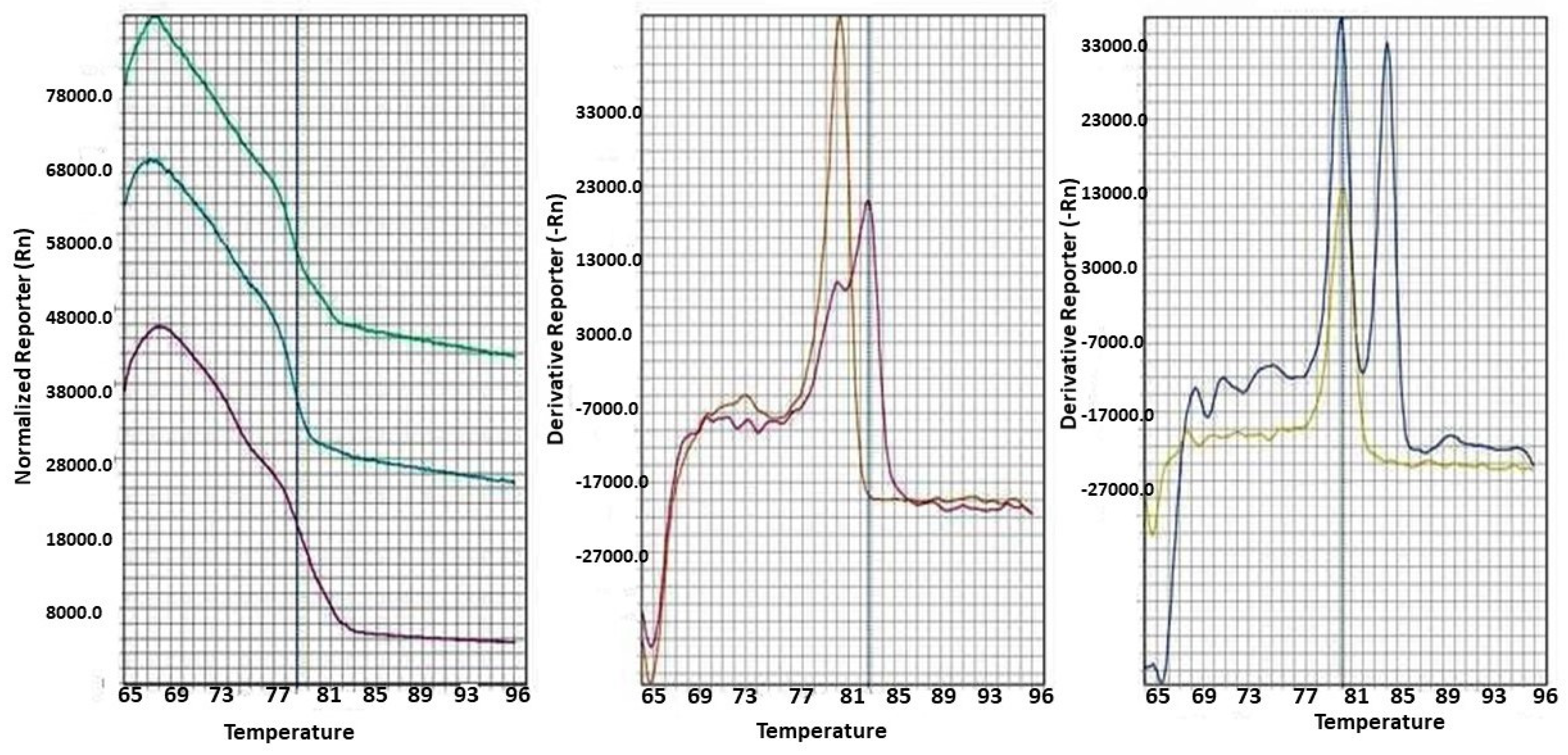
Figure 2.
Comparison of methylation quantification in six target promoter regions of SDHB, SDHC, and RDBP genes between PCCs/PGLs cases and non-cancerous patients. PCC is an abbreviation of PCCs/PGLs.
Figure 2.
Comparison of methylation quantification in six target promoter regions of SDHB, SDHC, and RDBP genes between PCCs/PGLs cases and non-cancerous patients. PCC is an abbreviation of PCCs/PGLs.
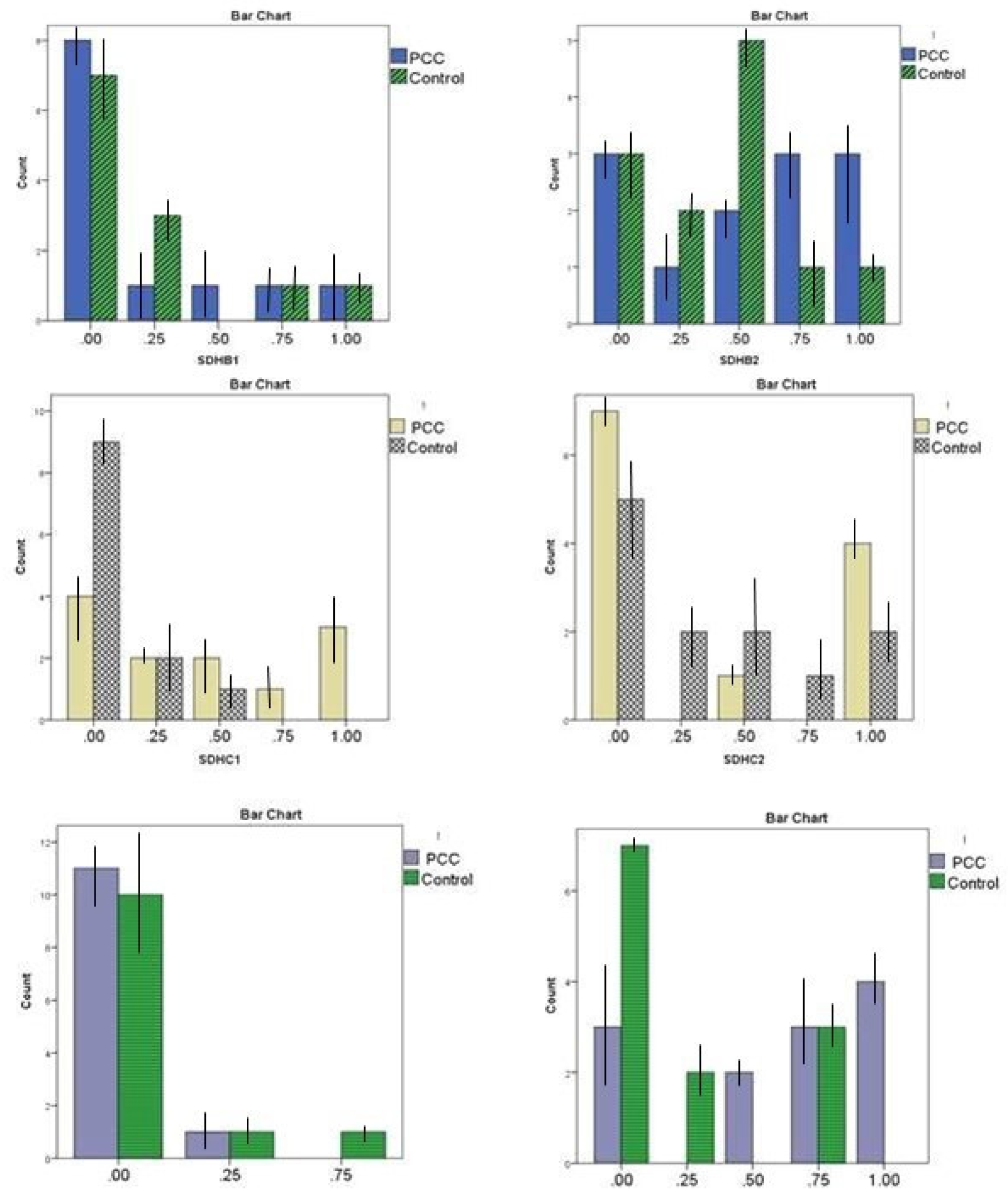
Figure 3.
Receiver Operating Characteristic (ROC) curves for the DNA methylation markers (as ranked by p-value), using the current collection of circulating plasma ctDNA.
Figure 3.
Receiver Operating Characteristic (ROC) curves for the DNA methylation markers (as ranked by p-value), using the current collection of circulating plasma ctDNA.


Table 1.
Specific primer sequences for methylation quantification through MS-HRM analysis of six selected promoter regions of RDBP, SDHB, and SDHC.
Table 1.
Specific primer sequences for methylation quantification through MS-HRM analysis of six selected promoter regions of RDBP, SDHB, and SDHC.
| Gene | Part | Forward Primer | Reverse Primer | Tann | Number of CpG sites in the amplicon |
|---|---|---|---|---|---|
|
RDBP |
a | 5' GGTAAGTTTTTTGTTTTTTAT 3' | 5' TTTAAATACATATAATTCA 3' | 56°C | 15 |
| b | 5' GGATATAGTTTGGTTTAAG 3' | 5’ ACATCTTTCTCCACTATTAC 3’ | 52°C | 9 | |
| SDHB | a |
5' GTTAGTGTTTTAGTGGATGT 3' |
5' AAACTCACCTACAAACAAAC 3' | 57°C | 17 |
| b | 5' GGGAAGTTAAATGGGT 3' | 5' TCCACTAAAACCCACT 3' | 55°C | 14 | |
| SDHC | a |
5' GTAATTAGTTAGGTAGAG 3' |
5' ACTAAATCACCTCAACA 3' | 50°C | 14 |
| b |
5'TAGATGTAGATTTTGAGTTA 3' |
5'ACTCTACTAACTAATTTAC 3' | 49°C | 6 |
Table 2.
Demographics of PCCs/PGLs (cases) and non-cancer patients (controls).
| Variables | PCCs/PGLs cases (n= 12) | Controls (n=12) | P-value |
|---|---|---|---|
| Age (years) | 41.25 (±10.532) | 42.42 (±11.828) | - |
|
Gender: Female Male |
8 (66.7%) 4 (33.3%) |
8 (66.7%) 4 (33.3%) |
- |
| Weight (kg) | 61.58 (±5.299) | 74.00 (±11.201) | 0. 002 |
| Height (cm) | 165.50 (±9.200) | 165.32 (±9.55) | 0.370 |
| BMI | 23.68 (±3.884) | 27.05 (±3.833) | 0.043 |
| SBP (mm/Hg) | 13.72(±1.190) | 11.50 (±1.167) | <.001 |
| DBP (mm/Hg) | 10.36(±1.68) | 7.91 (±1.37) | 0.001 |
| Educated (After High School) | 7 (58.3%) | 10 (83.3%) | 0.146 |
| Malignant | 5(41.6%) | - | - |
| Tumor size | 0.8-12 cm | - | - |
| Norepinephrine level | 4.2 ± 1.5% | - | - |
| Epinephrine | 58.02(±5.12) | - | - |
| Norepinephrine | 61.3 (±3.459) | - | - |
| Dopamine | 220 (±2.023) | - | - |
BMI: body mass index, SBP: systolic blood pressure, DBP: diastolic blood pressure.
Table 3.
Methylation pattern of six target promoter regions in PCCs/PGLs and normal cases.
| Promoter Region | Methylation | PCCs/PGLs patients Number (percentage) | Control (percentage) | P-value |
|
SDHBa |
0-12.5% methylated (non-methylated) | 8 (66.6%) | 7 (58.3%) | 0.886 |
| 12.5 ≤, <25% methylated | 1 (8.33%) | 3 (25.0%) | ||
| 25≤, <50% methylated | 1 (8.33%) | 0 (0.0%) | ||
| 50≤, <75 % methylated | 1 (8.33%) | 1 (8.33%) | ||
| 75- 100% methylated | 1 (8.33%) | 1 (8.33%) | ||
|
SDHBb |
0-12.5% methylated (non-methylated) | 3 (25.0%) | 3 (25.0%) | 0.507 |
| 12.5 <, ≥25% methylated | 1 (8.33%) | 2 (16.6%) | ||
| 25≤, <50% methylated | 2 (16.6%) | 5 (41.6%) | ||
| 50≤, <75 % methylated | 3 (25.0%) | 1 (8.33%) | ||
| 75- 100% methylated | 3 (25.0%) | 1 (8.33%) | ||
|
SDHCa |
0-12.5% methylated (non-methylated) | 4 (33.3%) | 9 (75.0%) | 0.026* |
| 12.5 ≤, <25% methylated | 2 (16.6%) | 2 (16.6%) | ||
| 25≤, <50% methylated | 2 (16.6%) | 1 (8.33%) | ||
| 50≤, <75 % methylated | 1 (8.33%) | 0 (0.0%) | ||
| 75- 100% methylated | 3 (25.0%) | 0 (0.0%) | ||
|
SDHCb |
0-12.5% methylated (non-methylated) | 7 (58.3%) | 5 (41.6%) | 0.750 |
| 12.5 ≤, <25% methylated | 0 (0.0%) | 2 (16.6%) | ||
| 25≤, <50% methylated | 1 (8.33%) | 2 (16.6%) | ||
| 50≤, <75 % methylated | 0 (0.0%) | 1 (8.33%) | ||
| 75- 100% methylated | 4 (33.3%) | 2 (16.6%) | ||
| RDBPa | 0-12.5% methylated (non-methylated) | 11 (891.6%) | 10 (83.3%) | 0.987 |
| 12.5 ≤, <25% methylated | 1 (8.33%) | 1 (8.33%) | ||
| 25≤, <50% methylated | 0 (0.0%) | 1 (8.33%) | ||
| 50≤, <75 % methylated | 0 (0.0%) | 0 (0.0%) | ||
| 75- 100% methylated | 0 (0.0%) | 0 (0.0%) | ||
| RDBPb | 0-12.5% methylated (non-methylated) | 3 (25.0%) | 7 (58.3%) | 0.032* |
| 12.5 ≤, <25% methylated | 0 (0.0%) | 2 (16.6%) | ||
| 25≤, <50% methylated | 2 (16.6%) | 0 (0.0%) | ||
| 50≤, <75 % methylated | 3 (25.0%) | 3 (25.0%) | ||
| 75- 100% methylated | 4 (33.3%) | 0 (0.0%) |
*If the p-value is less than 0.05, we reject the null hypothesis that there’s no difference between the two groups and conclude that a significant difference does exist. *If at least one targeted promoter region was methylated more than 25%, the final methylation status of the target gene was hypermethylated.
Table 4.
Observed agreements between six CpG sites of SDHB, SDHC, and RDBP2 in plasma ctDNA.
| RDBP1 | RDBP2 | SDHB1 | SDHB2 | |
|---|---|---|---|---|
| SDHCa | .567 | .668 | .715 | .919 * |
| SDHCb | .624 | .824 * | .805 | .649 |
Table 5.
AUC, Sensitivity & Specificity Analysis for six targeted promoter regions of PCCs/PGLs from patients’ ctDNA.
Table 5.
AUC, Sensitivity & Specificity Analysis for six targeted promoter regions of PCCs/PGLs from patients’ ctDNA.
| The ctDNA promoter Locus | AUC (95 % CI) | Sensitivity | Specificity |
|---|---|---|---|
| SDHBa | 0.476 (0.240-0.712) | 25% | 83.3% |
| SDHBb | 0.622 (0.388 - 0.855) | 66.6% | 41.6% |
| SDHCa | 0.757 (0.558 - 0.956) | 50% | 91.6% |
| SDHCb | 0.483 (0.242–0.723) | 41.6% | 58.3% |
| RDBPa | 0.455 (0.220 – 0.690) | 21% | 91.6% |
| RDBPb | 0.750 (0.549 - 0.951) | 75% | 75% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



