
Submitted:
15 April 2023
Posted:
17 April 2023
You are already at the latest version
Abstract
Cluster of differentiation 44 (CD44) promotes tumor progression through the recruitment of growth factors, and the acquisition of stemness, invasiveness, and drug resistance. CD44 has multiple isoforms including CD44 standard (CD44s) and CD44 variants (CD44v), which have common and unique functions in tumor development. Therefore, the elucidation of each CD44 isoform’s function in tumor is essential for the establishment of CD44-targeting tumor therapy. We have established various anti-CD44s and anti-CD44v monoclonal antibodies (mAbs) through immunization of CD44v3–10-overexpressed cells. In this study, we established C44Mab-6 (IgG1, kappa), which recognized the CD44 variant 3-encoded region (CD44v3) determined by enzyme-linked immunosorbent assay. C44Mab-6 reacted with CD44v3–10-overexpressed Chinese hamster ovary (CHO)-K1 cells (CHO/CD44v3–10) or some cancer cell lines (COLO205 and HSC-3) by flow cytometry. The apparent KD of C44Mab-6 for CHO/CD44v3–10, COLO205, and HSC-3 was 1.5 × 10−9 M, 6.3 × 10−9 M, and 1.9 × 10−9 M, respectively. C44Mab-6 could detect the CD44v3–10 in western blotting, and stained the formalin-fixed paraffin-embedded tumor sections in immunohistochemistry. These results indicate that C44Mab-6 is useful for detecting CD44v3 in various experiments, and expected for the application of tumor diagnosis and therapy.
Keywords:
CD44
; CD44 variant 3
; monoclonal antibody
; flow cytometry
; immunohistochemistry
1. Introduction
The cell surface glycoprotein, cluster of differentiation 44 (CD44) is broadly expressed by epithelial, mesenchymal, and hematopoietic cells, and is involved in the adhesion to extracellular matrix (ECM), lymphocytes homing, and their activation [1]. A growing body of evidences has been revealed critical roles of CD44 in tumor progression and metastasis [2,3]. The human CD44 gene consists of 19 exons, 10 of which are constant in all variants, and makes up the standard form of CD44 (CD44s) [4]. Furthermore, a large number of CD44 variants (CD44v) is generated due to the alternative splicing. The CD44v consists of the 10 constant exons in any combination with the remaining nine variant exons.
The translated CD44 usually receives post-translational modifications, such as N-/O-linked glycosylation or proteoglycans, including chondroitin sulfate, keratan sulfate, and heparan sulfate, which lead to the further diversity of CD44 function [5,6,7,8]. Therefore, the molecular weights of CD44s and CD44v show 75–95 kDa and 100~250 kDa, respectively [5]. These CD44 isoforms have both overlapping and unique functions. Both CD44s and CD44v (pan-CD44) possess hyaluronic acid (HA)-binding motifs that promote interaction with the microenvironment, which mediates cellular homing, migration, adhesion, and proliferation [9].
CD44v is overexpressed in tumors, which plays critical roles in the promotion of tumor invasion, metastasis, cancer-initiating properties [10], and resistance to therapies [2,11]. CD44v has ability to bind to heparin-binding growth factors, including fibroblast growth factors (FGFs) [7]. These growth factors bind to a heparan sulfate side chain attached to the SGSG sequence encoded by variant exon 3 [7,12]. Heparan sulfate proteoglycans play critical roles in cell proliferation and motility through presenting the growth factors to receptors. Therefore, the CD44 variant exon 3-containing isoform (CD44v3) can recruit heparin-binding growth factors to their receptor, and promote the growth factor receptor-mediated signal transduction [13,14,15]. Furthermore, the v6-encoded region makes the complex with hepatocyte growth factor and its receptor MET, which is essential for the activation [16]. The v8–10-encoded region interacts with a cystine–glutamate transporter (xCT) subunit, and mediates the oxidative stress resistance through regulation of the intracellular redox state [17].
Cancer stem cells (CSCs) exhibit the important properties, such as self-renewal, resistance to therapy, and promotion of tumor metastasis [18,19,20]. Several cell surface and intracellular proteins have been reported as CSC markers in hematopoietic malignancy and solid tumors [21,22]. Among them, CD44 was identified as a CSC marker in several solid tumors. In breast cancer, the CD44+CD24−/lowLineage− population was first shown to be 10- to 50-fold enriched for the ability to form tumors in immunodeficient mice relative to unfractionated tumor cells [23]. In head and neck squamous cell carcinoma (HNSCC), the CD44-high CSCs from HNSCC exhibited the elevated migration, invasiveness, and stemness [24,25] and could form metastatic foci in lung of immunodeficient mice. In contrast, the CD44-low populations failed to form the metastatic proliferation [26]. In above studies, anti-pan-CD44 monoclonal antibodies (mAbs) were used to isolate the CSCs from cultured cells and patient-derived tumor tissues. Furthermore, several CD44v specific mAbs were reported to separate CSCs from colorectal cancer using anti-CD44v6 [27] and anti-CD44v9 [28] mAbs. Therefore, specific mAbs against CD44s and CD44v are required for the isolation of CSCs and the analysis of their properties in detail.
Using the Cell-Based Immunization and Screening (CBIS) method, we established an anti-pan-CD44 mAb, C44Mab-5 (IgG1, kappa) [29]. We further established another anti-pan-CD44 mAb, C44Mab-46 (IgG1, kappa) by the immunization of CD44v3–10 ectodomain (CD44ec). [30]. Both C44Mab-5 and C44Mab-46 have been revealed to recognize the standard exons (1 to 5)-encoding sequences at the N-terminus [31,32,33]. Furthermore, both C44Mab-5 and C44Mab-46 are available for flow cytometry and immunohistochemical analyses in oral squamous cell carcinomas (OSCC) [29] and esophageal squamous cell carcinomas [30]. We further converted the mouse IgG1 subclass antibody (C44Mab-5) into an IgG2a subclass antibody (5-mG2a), and further produced a defucosylated version (5-mG2a-f) using fucosyltransferase 8-deficient ExpiCHO-S (BINDS-09) cells. The 5-mG2a-f exhibited in vitro antibody-dependent cellular cytotoxicity (ADCC) activity against OSCC cell lines (HSC-2 and SAS). Furthermore, the 5-mG2a-f suppressed the growth of HSC-2 and SAS xenograft [34].
Recently, we have established an anti-CD44v5 mAb [35] and an anti-CD44v6 mAb [36] by the CBIS method, an anti-CD44v7/8 mAb [37] by the immunization of CD44ec, and an anti-CD44v4 mAb by the peptide immunization [38]. In this study, we developed a novel anti-CD44v3 mAb, C44Mab-6 (IgG1, kappa) by CBIS method, and evaluated its applications, such as flow cytometry, Western blot, and immunohistochemical analyses.
2. Results
2.1. Development of C44Mab-6 as an anti-CD44v3 mAb
The CBIS method contains the immunization of antigen-overexpressed cells and the high throughput hybridoma screening using flow cytometry. We prepared a stable cell line, CHO/CD44v3–10, as an immunogen (Figure 1). The cells were immunized into mice, and hybridomas were plated into 96-well plates. We next performed the flow cytometry-based high throughput screening to select the supernatants, which are positive for CHO/CD44v3–10 cells, and negative for CHO-K1 cells. After the limiting dilution, anti-CD44 mAb-producing clones were finally established. Among them, C44Mab-6 (IgG1, kappa) was shown to recognize CD44p231–250 peptide (AGWEPNEENEDERDRHLSFS), which is corresponding to variant 3-encoded sequence (Figure 2 and Supplementary Table S1). In contrast, C44Mab-6 never recognized other extracellular region peptides of CD44v3-10. These results indicated that C44Mab-6 specifically recognizes CD44 variant 3-encoded sequence.
2.2. The reactivity of C44Mab-6 to CD44-Expressing Cells in Flow Cytometry
The reactivity of C44Mab-6 to CHO/CD44v3–10, CHO/CD44s, and CHO-K1 cells was investigated using flow cytometry. C44Mab-6 dose-dependently recognized CHO/CD44v3–10 cells (Figure 3A). In contrast, C44Mab-6 recognized neitherCHO/CD44s (Figure 3B) nor CHO-K1 (Figure 3C) cells. C44Mab-46, which is an anti-pan-CD44 mAb [30], recognized both CHO/CD44v3–10 and CHO/CD44s cells (Supplementary Figure S1). Moreover, C44Mab-6 could recognize a colorectal cancer cell line COLO205 (Figure 3D), and an oral squamous cell line HSC-3 (Figure 3E) in a dose-dependent manner.
2.3. The binding affinity of C44Mab-6 to CD44-Expressing Cells
The binding affinity of C44Mab-6 to CHO/CD44v3–10, COLO205, and HSC-3 was determined using flow cytometry. As shown in Figure 4, the KD of CHO/CD44v3–10 COLO205 and HSC-3 was determined as 1.5 × 10−9 M, 6.3 × 10−9 M, and 1.9 × 10−9 M, respectively. These results indicated that C44Mab-6 possesses high affinity for CD44v3–10 and endogenous CD44v3-expressing cells.
3.4. Western Blot Analysis
Western blot analysis was preformed to investigate the specificity of C44Mab-6. As shown in Figure 5A, C44Mab-6 detected CD44v3–10 as more than 180-kDa bands. However, C44Mab-6 did not detect any bands from lysates of CHO-K1 and CHO/CD44s cells at more than 48-kDa. An anti-pan-CD44 mAb, C44Mab-46, recognized the lysates from both CHO/CD44s (~75 kDa) and CHO/CD44v3–10 (> 180 kDa) (Figure 5B). These results indicated that C44Mab-6 specifically detects exogenous CD44v3–10, but not CD44s.
3.5. Immunohistochemical Analysis Using C44Mab-6 against Tumor Tissues
Immunohistochemical analysis against the formalin-fixed paraffin-embedded (FFPE) sections of OSCC was conducted to assess the availability of C44Mab-6. We used sequential sections of OSCC tissue microarray, and compared the staining patterns of C44Mab-6 and C44Mab-46. The clear membranous staining was observed by C44Mab-6 and C44Mab-46 in a well-differentiated OSCC section (Figure 6A,B). Figure 6C,D showed a OSCC section with the stromal invaded phenotype. C44Mab-6 strongly stained stromal invaded OSCC and could clearly distinguish tumor cells from surrounding stroma cells (Figure 6C). In contrast, C44Mab-46 stained both invaded tumor and stromal cells (Figure 6D). In Figure 6E,F, C44Mab-6 partially stained tumor cells, but not stromal cells (Figure 6E). In contrast, C44Mab-46 mainly stained stromal cells (Figure 6F). We summarized the data of immunohistochemical analysis of CD44 expression in tumor cells in Table 1; C44Mab-6 stained 44 out of 50 (88%) cases of OSCC. We also stained FFPE sections of colorectal cancer tissue microarray, and found that C44Mab-6 stained 7 out of 40 (18%) cases (Supplementary Table S2). However, the C44Mab-6 reactivity was faint and partly localized compared to that of C44Mab-46 (Supplementary Figure S2). These results indicated that C44Mab-6 is useful for immunohistochemical analysis of FFPE tumor sections.
3. Discussion
In this study, we developed C44Mab-6 using the CBIS method (Figure 1), and determined its epitope as variant 3-encoded region of CD44 (Figure 2 and supplemental Table S1). Then, we showed the usefulness of C44Mab-6 for multiple applications, including flow cytometry (Figure 3 and Figure 4), western blotting (Figure 5), and immunohistochemistry (Figure 6).
An anti-CD44v3 mAb (clone 3G5) was previously developed, and widely used for various applications [39]. The 3G5 was developed by the immunization of COS1-produced CD44v3-10-Fc protein. The specificity to exon was determined by indirect immunofluorescent staining of COS1 cells which expressed CD44v3–10, CD44v6–10, CD44v7–10, CD44v8–10, and CD44v10 [39]. Therefore, the 3G5 is thought to recognize the peptide or glycopeptide structure of CD44v3. However, the detailed binding epitope of 3G5 has not been determined. As shown in supplementary Table S1, C44Mab-6 recognized CD44p231–250 peptide (AGWEPNEENEDERDRHLSFS), but not CD44p241–260 peptides (DERDRHLSFSGSGIDDDEDF). The underlined SGSG sequence is heparan sulfate-modified sequence in variant 3-encoded region [7,12]. Therefore, the recognition of C44Mab-6 is probably not affected by the heparan sulfate modification.
Head and neck cancers are mainly derived from the oral cavity, larynx, pharynx, and nasal cavity [40]. The SCC is the common type. As shown in Figure 6, C44Mab-6 clearly stained the membrane of OSCC, and recognized a human OSCC cell line, HSC-3 (Figure 3 and Figure 4). The CD44v3 high and aldehyde dehydrogenase-1 (ALDH1)-high population of HSC-3 exhibited a potent tumorigenic potential in immunodeficient NOD/SCID mice [41]. The population showed the increased stemness-related transcriptional factors, including OCT4, SOX2, and NANOG [41]. In the future study, we will investigate the application of C44Mab-6 to isolate the cancer stem-like cell from cancer cell lines and/or OSCC tissue, and establish the strategy to deplete the cells for tumor therapy. We have investigated the antitumor activity using class-switched and defucosylated IgG2a mAbs [34,42,43,44,45,46,47,48]. The defucosylated IgG2a mAbs can be produced by fucosyltransferases 8-deficient CHO-K1 cells, and exhibited potent ADCC activity in vitro, and suppressed the xenograft growth [34,42,43,44,45,46,47,48]. Therefore, the production of defucosylated C44Mab-6 is one of the strategies to evaluate the antitumor activity in vivo.
The HNSCC treatments include surgery, chemotherapy, radiotherapy, molecular targeted therapy, immunotherapy, or a combination of these modalities [49]. Despite the progress of the therapies, drug resistance and metastasis are still the main causes of death [50]. In preclinical study, a pan-CD44 mAb applied to the novel modalities, including near-infrared photoimmunotherapy (NIR-PIT). The CD44 mAb–the photoactivatable dye IRDye700DX conjugate exhibited the significant antitumor effects after the NIR-light exposure against CD44-expressing OSCC [51]. However, a pan-CD44 mAb, C44Mab-46, recognized not only tumor cells but also stromal tissues (Figure 6D,F), and probably immune cells, which are important for antitumor immunity. Therefore, CD44v is a more rational tumor antigen for NIR-PIT, which could be a new modality for OSCC with locoregional recurrence.
Zen et al. established a unique mAb (clone C3H7), which recognized the basolateral membranes of epithelium and inhibited both the adhesion of epithelial cells to immobilized CD11b/CD18 and the transepithelial migration of leukocytes [52]. CD11b/CD18, also known as Macrophage-1 antigen, is a leukocyte integrin essential for firm adhesion to epithelial cells and transepithelial migration of leukocytes [53]. However, the receptor of CD11b/CD18 on epithelial cells has not been identified. They revealed that the antigen of C3H7 is CD44v3, which specifically binds to CD11b/CD18 through its heparan sulfate moieties [52]. The C3H7 antigen was increased by treatment with pro-inflammatory cytokine, including interferon-γ and tumor necrosis factor-α in epithelial monolayers [52], which support the previous finding that CD44v3 is increased in inflammatory diseases including ulcerative colitis [54]. C44Mab-6 also recognized the basolateral surface of colorectal cancer cells (Supplementary Figure S2A). Further investigations are required for the relationship between CD44v3 expression and the transepithelial migration of leukocytes. The study could provide the basis for the development of novel therapeutic applications of anti-CD44v3 mAbs, including C44Mab-6.
Chimeric antigen receptor T-cell (CAR-T) therapy have been developed in variety of hematopoietic malignances and solod tumors [55]. CAR-T cells have demonstrated remarkable success in treating CD19+ B cell leukemias [56]. However, CAR-T therapy for acute myeloid leukemia (AML) has been elusive, because of the target restriction and phenotypic heterogeneity [57]. Mutations of the FMS-like tyrosine kinase 3 (FLT3) and DNA methyltransferase 3A (DNMT3A) genes were identified as common driver mutations associated with poor prognosis of AML patients [58]. Tang et al. showed that AML cells expressed the high levels of CD44 mRNA, and the expression of AML-derived FLT3 and DNMT3A mutants promote the transcription of CD44 mRNA through suppression of CpG islands methylation in the CD44 promoter [59]. They also found that AML patients with FLT3 or DNMT3A mutations had higher expression of CD44v6 compared to normal specimens. Furthermore, they showed that CD44v6 CAR-T cells exhibited potent anti-leukemic effects [59]. Therefore, CD44v6 is thought to be a rational antigen of CAR-T therapy for AML with FLT3 or DNMT3A mutations.
Since CD44 mRNA is upregulated in AML, there is a possibility that other CD44 variants are also transcribed and expressed in AML. In humanized mouse model of chronic myeloid leukemia (CML) progression from chronic phase to blast crisis, a CD44 variant (CD44v8–10) was elevated, which is required for the maintenance of stemness [60]. Furthermore, we have examined the reactivity of C44Mab-6 against cell lines derived from hematopoietic malignancy, and found the increased reactivity in several cell lines (data not shown). Further studies are required for the selective expression of CD44v3 in leukemia cells, but not in hematopoietic stem cells to ensure its safety as a CAR-T antigen.
4. Materials and Methods
4.1. Cell lines
COLO205 (a human colorectal cancer cell line) was obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer at Tohoku University (Miyagi, Japan). HSC-3 (a human OSCC cell line) and LN229 (a human glioblastoma cell line) were obtained from the Japanese Collection of Research Bioresources (Osaka, Japan). P3X63Ag8U.1 (P3U1; a mouse multiple myeloma) and Chinese hamster ovary (CHO)-K1 cell lines were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). HSC-3 and LN229 were cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/mL of penicillin, 100 μg/mL streptomycin, 0.25 μg/mL amphotericin B. CHO-K1, COLO205, and P3U1 were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (Nacalai Tesque, Inc.) supplemented with 10% FBS and antibiotics, as indicated above. All the cells were grown in a humidified incubator at 37°C with 5% CO2.
4.2. Construction of expression plasmids and stable transfectants
Using LN229 cDNA as a template, CD44s cDNA was amplified using HotStar HiFidelity Polymerase Kit (Qiagen Inc., Hilden, Germany). We obtained CD44v3–10 cDNA from the RIKEN BRC through the National Bio-Resource Project of the MEXT, Japan. The CD44s and CD44v3–10 cDNAs were cloned into pCAG-Ble-ssPA16 vector with signal sequence and N-terminal PA16 tag (GLEGGVAMPGAEDDVV) [29,61,62,63,64], which is detected by NZ-1, originally developed as an anti-human podoplanin mAb [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80]. The pCAG-Ble/PA16-CD44s and pCAG-Ble/PA16-CD44v3–10 vectors were transfected into CHO-K1 cells using a Neon transfection system (Thermo Fisher Scientific, Inc.), and CHO/CD44s and CHO/CD44v3–10 were finally established, as described previously [36].
4.3. Production of hybridomas
All animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2019NiA-001). The female BALB/c mice (CLEA Japan., Tokyo, Japan) were intraperitoneally immunized with CHO/CD44v3–10 (1 × 108 cells) with Imject Alum (Thermo Fisher Scientific Inc.) as an adjuvant. The three additional immunizations per week, and a booster injection were performed two days before harvest of the spleen cells. The hybridomas were produced by the fusion of splenocytes and P3U1 cells using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN, USA). The supernatants, which are positive for CHO/CD44v3–10 cells and negative for CHO-K1 cells, were selected using flow cytometry, SA3800 Cell Analyzers (Sony Corp. Tokyo, Japan).
4.4. ELISA
We obtained fifty-eight peptides, which cover the extracellular domain of CD44v3–10 [31], from Sigma-Aldrich Corp. (St. Louis, MO, USA). We immobilized them on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific, Inc) at 1 µg/mL for 30 min at 37°C. The immunoplates washing was performed using HydroSpeed Microplate Washer (Tecan, Zürich, Switzerland) with phosphate-buffered saline (PBS) containing 0.05% (v/v) Tween 20 (PBST; Nacalai Tesque, Inc.). After the blocking with 1% (w/v) bovine serum albumin (BSA) in PBST for 30 min at 37°C, C44Mab-6 (10 µg/mL) was added to each well. Then, the wells were further incubated with anti-mouse immunoglobulins peroxidase-conjugate (1:2000 diluted; Agilent Technologies Inc., Santa Clara, CA, USA) for 30 min at 37°C. One-Step Ultra TMB (Thermo Fisher Scientific Inc.) was used for enzymatic reactions. An iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA, USA) was used to mesure the optical density at 655 nm.
4.5. Flow cytometry
CHO-K1, CHO/CD44s, CHO/CD44v3–10, COLO205, and HSC-3 cells were obtained using 0.25% trypsin and 1 mM ethylenediamine tetraacetic acid (EDTA; Nacalai Tesque, Inc.). The cells were treated with C44Mab-6, C44Mab-46, or blocking buffer (control) (0.1% BSA in PBS) for 30 min at 4˚C. Then, the cells (1 × 105 cells/sample) were treated with anti-mouse IgG conjugated with Alexa Fluor 488 (1:2,000; Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 min at 4˚C. The data were analyzed using the SA3800 Cell Analyzer and SA3800 software ver. 2.05 (Sony Corp.).
4.6. Determination of dissociation constant (KD) via flow cytometry
In CHO/CD44v3–10 cells, we prepared from 130 to 0.008 nM (diluted by 1/2) of C44Mab-6. In COLO201 and HSC-3 cells, we prepared from 1300 to 0.08 nM (diluted by 1/2) of C44Mab-6. The serially diluted C44Mab-6 was suspended with 2 × 105 cells. Then, the cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488 (1:200). BD FACSLyric and BD FACSuite software version 1.3 (BD Biosciences, Franklin Lakes, NJ, USA) were used for the fluorescence data analyses. The KD was determined by the fitting binding isotherms to built-in one-site binding models of GraphPad Prism 8 (GraphPad Software, Inc., L a Jolla, CA, USA).
4.7. Western blot analysis
The 10 μg of cell lysates were subjected to SDS-polyacrylamide gel for electrophoresis using polyacrylamide gels (5–20%; FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The separated proteins were transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA, Darmstadt, Germany). The blocking was performed using 4% skim milk (Nacalai Tesque, Inc.) in PBST. The membranes were incubated with 10 μg/mL of C44Mab-6, 10 μg/mL of C44Mab-46, or 1 μg/mL of an anti-β-actin mAb (clone AC-15; Sigma-Aldrich Corp.), and then incubated with peroxidase-conjugated anti-mouse immunoglobulins (diluted 1:1,000; Agilent Technologies, Inc.). Finally, the signals were enhanced using a chemiluminescence reagent, ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation), and were detected by a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
4.8. Immunohistochemical analysis
FFPE sections of OSCC tissue array (OR601c) and colorectal carcinoma tissue array (CO483a) were purchased from US Biomax Inc. (Rockville, MD, USA). The tissue arrays were autoclaved in EnVision FLEX Target Retrieval Solution High pH (Agilent Technologies, Inc.) for 20 min. After blocking with SuperBlock T20 (Thermo Fisher Scientific, Inc.), the sections were incubated with C44Mab-6 (1 μg/mL) and C44Mab-46 (1 μg/mL) for 1 h at room temperature. The sections further were incubated with the EnVision+ Kit for mouse (Agilent Technologies Inc.) for 30 min. Then, the chromogenic reaction using 3,3′-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies Inc.) was conducted. Hematoxylin (FUJIFILM Wako Pure Chemical Corporation) was used for the counterstaining. To examine the sections and obtain images, we used Leica DMD108 (Leica Microsystems GmbH, Wetzlar, Germany).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1, The determination of the binding epitope of C44Mab-6 by ELISA. Supplementary Table S2, Immunohistochemical analysis using C44Mab-6 and C44Mab-46 against colorectal cancer tissues. Supplementary Figure S1, Recognition of CHO/CD44s and CHO/CD44v3–10 by C44Mab-46 using flow cytometry. Supplementary Figure S2, Immunohistochemical analysis using C44Mab-6 and C44Mab-46 against colorectal carcinoma tissues.
Author Contributions
H.S., K.K, N.G., K.I, T.O, and T.T. performed the experiments. M.K.K. and Y.K designed the experiments. H.S. and Y.K analyzed the data. H.S. and Y.K. wrote the manuscript. All authors have read and agreed to the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP22ama121008 (to Y.K.), JP22am0401013 (to Y.K.), JP22bm1004001 (to Y.K.), JP22ck0106730 (to Y.K.), and JP21am0101078 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 21K20789 (to T.T.), 22K06995 (to H.S.), 21K07168 (to M.K.K.), and 22K07224 (to Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2019NiA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article and supplementary material.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: from adhesion molecules to signalling regulators. Nat Rev Mol Cell Biol 2003, 4, 33-45. [CrossRef]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11. [CrossRef]
- Zöller, M. CD44: can a cancer-initiating cell profit from an abundantly expressed molecule? Nat Rev Cancer 2011, 11, 254-267. [CrossRef]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl Med 2015, 4, 1033-1043. [CrossRef]
- Mereiter, S.; Martins Á, M.; Gomes, C.; Balmaña, M.; Macedo, J.A.; Polom, K.; Roviello, F.; Magalhães, A.; Reis, C.A. O-glycan truncation enhances cancer-related functions of CD44 in gastric cancer. FEBS Lett 2019, 593, 1675-1689. [CrossRef]
- Takahashi, K.; Stamenkovic, I.; Cutler, M.; Dasgupta, A.; Tanabe, K.K. Keratan sulfate modification of CD44 modulates adhesion to hyaluronate. J Biol Chem 1996, 271, 9490-9496. [CrossRef]
- Bennett, K.L.; Jackson, D.G.; Simon, J.C.; Tanczos, E.; Peach, R.; Modrell, B.; Stamenkovic, I.; Plowman, G.; Aruffo, A. CD44 isoforms containing exon V3 are responsible for the presentation of heparin-binding growth factor. J Cell Biol 1995, 128, 687-698. [CrossRef]
- Brown, T.A.; Bouchard, T.; St John, T.; Wayner, E.; Carter, W.G. Human keratinocytes express a new CD44 core protein (CD44E) as a heparan-sulfate intrinsic membrane proteoglycan with additional exons. J Cell Biol 1991, 113, 207-221. [CrossRef]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angiogenesis in vascular disease: uncovering RHAMM and CD44 receptor signaling pathways. Matrix Biol 2007, 26, 58-68. [CrossRef]
- Guo, Q.; Yang, C.; Gao, F. The state of CD44 activation in cancer progression and therapeutic targeting. Febs j 2021. [CrossRef]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int J Biochem Cell Biol 2016, 81, 166-173. [CrossRef]
- Jackson, D.G.; Bell, J.I.; Dickinson, R.; Timans, J.; Shields, J.; Whittle, N. Proteoglycan forms of the lymphocyte homing receptor CD44 are alternatively spliced variants containing the v3 exon. J Cell Biol 1995, 128, 673-685. [CrossRef]
- DiGabriele, A.D.; Lax, I.; Chen, D.I.; Svahn, C.M.; Jaye, M.; Schlessinger, J.; Hendrickson, W.A. Structure of a heparin-linked biologically active dimer of fibroblast growth factor. Nature 1998, 393, 812-817. [CrossRef]
- Spivak-Kroizman, T.; Lemmon, M.A.; Dikic, I.; Ladbury, J.E.; Pinchasi, D.; Huang, J.; Jaye, M.; Crumley, G.; Schlessinger, J.; Lax, I. Heparin-induced oligomerization of FGF molecules is responsible for FGF receptor dimerization, activation, and cell proliferation. Cell 1994, 79, 1015-1024. [CrossRef]
- Yayon, A.; Klagsbrun, M.; Esko, J.D.; Leder, P.; Ornitz, D.M. Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell 1991, 64, 841-848. [CrossRef]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev 2002, 16, 3074-3086. [CrossRef]
- Ishimoto, T.; Nagano, O.; Yae, T.; Tamada, M.; Motohara, T.; Oshima, H.; Oshima, M.; Ikeda, T.; Asaba, R.; Yagi, H.; et al. CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc(-) and thereby promotes tumor growth. Cancer Cell 2011, 19, 387-400. [CrossRef]
- Maitland, N.J.; Collins, A.T. Cancer stem cells - A therapeutic target? Curr Opin Mol Ther 2010, 12, 662-673.
- Prince, M.E.; Ailles, L.E. Cancer stem cells in head and neck squamous cell cancer. J Clin Oncol 2008, 26, 2871-2875. [CrossRef]
- Ailles, L.E.; Weissman, I.L. Cancer stem cells in solid tumors. Curr Opin Biotechnol 2007, 18, 460-466. [CrossRef]
- Yu, S.S.; Cirillo, N. The molecular markers of cancer stem cells in head and neck tumors. J Cell Physiol 2020, 235, 65-73. [CrossRef]
- Tahmasebi, E.; Alikhani, M.; Yazdanian, A.; Yazdanian, M.; Tebyanian, H.; Seifalian, A. The current markers of cancer stem cell in oral cancers. Life Sci 2020, 249, 117483. [CrossRef]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A 2003, 100, 3983-3988. [CrossRef]
- Lee, Y.; Shin, J.H.; Longmire, M.; Wang, H.; Kohrt, H.E.; Chang, H.Y.; Sunwoo, J.B. CD44+ Cells in Head and Neck Squamous Cell Carcinoma Suppress T-Cell-Mediated Immunity by Selective Constitutive and Inducible Expression of PD-L1. Clin Cancer Res 2016, 22, 3571-3581. [CrossRef]
- Prince, M.E.; Sivanandan, R.; Kaczorowski, A.; Wolf, G.T.; Kaplan, M.J.; Dalerba, P.; Weissman, I.L.; Clarke, M.F.; Ailles, L.E. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proc Natl Acad Sci U S A 2007, 104, 973-978. [CrossRef]
- Davis, S.J.; Divi, V.; Owen, J.H.; Bradford, C.R.; Carey, T.E.; Papagerakis, S.; Prince, M.E. Metastatic potential of cancer stem cells in head and neck squamous cell carcinoma. Arch Otolaryngol Head Neck Surg 2010, 136, 1260-1266. [CrossRef]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell 2014, 14, 342-356. [CrossRef]
- Kimura, Y.; Goi, T.; Nakazawa, T.; Hirono, Y.; Katayama, K.; Urano, T.; Yamaguchi, A. CD44variant exon 9 plays an important role in colon cancer initiating cells. Oncotarget 2013, 4, 785-791. [CrossRef]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C(44)Mab-5. Biochem Biophys Rep 2018, 14, 64-68. [CrossRef]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int J Mol Sci 2022, 23. [CrossRef]
- Takei, J.; Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using Alanine-Scanning Mutagenesis and Surface Plasmon Resonance. Monoclon Antib Immunodiagn Immunother 2021, 40, 219-226. [CrossRef]
- Asano, T.; Kaneko, M.K.; Takei, J.; Tateyama, N.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 156-161. [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 162-167. [CrossRef]
- Takei, J.; Kaneko, M.K.; Ohishi, T.; Hosono, H.; Nakamura, T.; Yanaka, M.; Sano, M.; Asano, T.; Sayama, Y.; Kawada, M.; et al. A defucosylated antiCD44 monoclonal antibody 5mG2af exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Oncol Rep 2020, 44, 1949-1960. [CrossRef]
- Kudo, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 variant 5 Monoclonal An-tibody C44Mab-3 for Multiple Applications against Pancreatic Carcinomas. Antibodies 2023, in press.
- Ejima, R.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 6 Monoclonal Antibody C(44)Mab-9 for Multiple Applications against Colorectal Carcinomas. Int J Mol Sci 2023, 24. [CrossRef]
- Suzuki, H.; Ozawa, K.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 7/8 Monoclonal Antibody, C44Mab-34, for Multiple Applications against Oral Carcinomas. Biomedicines 2023, 11, 1099. [CrossRef]
- Suzuki, H.; Tanaka, T.; Goto, N.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry. Curr Issues Mol Biol 2023, 45, 1875-1888. [CrossRef]
- Fox, S.B.; Fawcett, J.; Jackson, D.G.; Collins, I.; Gatter, K.C.; Harris, A.L.; Gearing, A.; Simmons, D.L. Normal human tissues, in addition to some tumors, express multiple different CD44 isoforms. Cancer Res 1994, 54, 4539-4546.
- Mody, M.D.; Rocco, J.W.; Yom, S.S.; Haddad, R.I.; Saba, N.F. Head and neck cancer. Lancet 2021, 398, 2289-2299. [CrossRef]
- Bourguignon, L.Y.; Wong, G.; Earle, C.; Chen, L. Hyaluronan-CD44v3 interaction with Oct4-Sox2-Nanog promotes miR-302 expression leading to self-renewal, clonal formation, and cisplatin resistance in cancer stem cells from head and neck squamous cell carcinoma. J Biol Chem 2012, 287, 32800-32824. [CrossRef]
- Li, G.; Suzuki, H.; Ohishi, T.; Asano, T.; Tanaka, T.; Yanaka, M.; Nakamura, T.; Yoshikawa, T.; Kawada, M.; Kaneko, M.K.; et al. Antitumor activities of a defucosylated anti-EpCAM monoclonal antibody in colorectal carcinoma xenograft models. Int J Mol Med 2023, 51. [CrossRef]
- Nanamiya, R.; Takei, J.; Ohishi, T.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Handa, S.; Tateyama, N.; et al. Defucosylated Anti-Epidermal Growth Factor Receptor Monoclonal Antibody (134-mG(2a)-f) Exerts Antitumor Activities in Mouse Xenograft Models of Canine Osteosarcoma. Monoclon Antib Immunodiagn Immunother 2022, 41, 1-7. [CrossRef]
- Kawabata, H.; Suzuki, H.; Ohishi, T.; Kawada, M.; Kaneko, M.K.; Kato, Y. A Defucosylated Mouse Anti-CD10 Monoclonal Antibody (31-mG(2a)-f) Exerts Antitumor Activity in a Mouse Xenograft Model of CD10-Overexpressed Tumors. Monoclon Antib Immunodiagn Immunother 2022, 41, 59-66. [CrossRef]
- Kawabata, H.; Ohishi, T.; Suzuki, H.; Asano, T.; Kawada, M.; Suzuki, H.; Kaneko, M.K.; Kato, Y. A Defucosylated Mouse Anti-CD10 Monoclonal Antibody (31-mG(2a)-f) Exerts Antitumor Activity in a Mouse Xenograft Model of Renal Cell Cancers. Monoclon Antib Immunodiagn Immunother 2022. [CrossRef]
- Asano, T.; Tanaka, T.; Suzuki, H.; Li, G.; Ohishi, T.; Kawada, M.; Yoshikawa, T.; Kaneko, M.K.; Kato, Y. A Defucosylated Anti-EpCAM Monoclonal Antibody (EpMab-37-mG(2a)-f) Exerts Antitumor Activity in Xenograft Model. Antibodies (Basel) 2022, 11. [CrossRef]
- Tateyama, N.; Nanamiya, R.; Ohishi, T.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Saito, M.; Asano, T.; Tanaka, T.; et al. Defucosylated Anti-Epidermal Growth Factor Receptor Monoclonal Antibody 134-mG(2a)-f Exerts Antitumor Activities in Mouse Xenograft Models of Dog Epidermal Growth Factor Receptor-Overexpressed Cells. Monoclon Antib Immunodiagn Immunother 2021, 40, 177-183. [CrossRef]
- Takei, J.; Ohishi, T.; Kaneko, M.K.; Harada, H.; Kawada, M.; Kato, Y. A defucosylated anti-PD-L1 monoclonal antibody 13-mG(2a)-f exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Biochem Biophys Rep 2020, 24, 100801. [CrossRef]
- Xing, D.T.; Khor, R.; Gan, H.; Wada, M.; Ermongkonchai, T.; Ng, S.P. Recent Research on Combination of Radiotherapy with Targeted Therapy or Immunotherapy in Head and Neck Squamous Cell Carcinoma: A Review for Radiation Oncologists. Cancers (Basel) 2021, 13. [CrossRef]
- Muzaffar, J.; Bari, S.; Kirtane, K.; Chung, C.H. Recent Advances and Future Directions in Clinical Management of Head and Neck Squamous Cell Carcinoma. Cancers (Basel) 2021, 13. [CrossRef]
- Nagaya, T.; Nakamura, Y.; Okuyama, S.; Ogata, F.; Maruoka, Y.; Choyke, P.L.; Allen, C.; Kobayashi, H. Syngeneic Mouse Models of Oral Cancer Are Effectively Targeted by Anti-CD44-Based NIR-PIT. Mol Cancer Res 2017, 15, 1667-1677. [CrossRef]
- Zen, K.; Liu, D.Q.; Li, L.M.; Chen, C.X.; Guo, Y.L.; Ha, B.; Chen, X.; Zhang, C.Y.; Liu, Y. The heparan sulfate proteoglycan form of epithelial CD44v3 serves as a CD11b/CD18 counter-receptor during polymorphonuclear leukocyte transepithelial migration. J Biol Chem 2009, 284, 3768-3776. [CrossRef]
- Colgan, S.P.; Comerford, K.M.; Lawrence, D.W. Epithelial cell-neutrophil interactions in the alimentary tract: a complex dialog in mucosal surveillance and inflammation. ScientificWorldJournal 2002, 2, 76-88. [CrossRef]
- Rosenberg, W.M.; Prince, C.; Kaklamanis, L.; Fox, S.B.; Jackson, D.G.; Simmons, D.L.; Chapman, R.W.; Trowell, J.M.; Jewell, D.P.; Bell, J.I. Increased expression of CD44v6 and CD44v3 in ulcerative colitis but not colonic Crohn's disease. Lancet 1995, 345, 1205-1209. [CrossRef]
- Oliveira, G.; Wu, C.J. Dynamics and specificities of T cells in cancer immunotherapy. Nat Rev Cancer 2023. [CrossRef]
- Haslauer, T.; Greil, R.; Zaborsky, N.; Geisberger, R. CAR T-Cell Therapy in Hematological Malignancies. Int J Mol Sci 2021, 22. [CrossRef]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424-447. [CrossRef]
- Patel, J.P.; Gönen, M.; Figueroa, M.E.; Fernandez, H.; Sun, Z.; Racevskis, J.; Van Vlierberghe, P.; Dolgalev, I.; Thomas, S.; Aminova, O.; et al. Prognostic relevance of integrated genetic profiling in acute myeloid leukemia. N Engl J Med 2012, 366, 1079-1089. [CrossRef]
- Tang, L.; Huang, H.; Tang, Y.; Li, Q.; Wang, J.; Li, D.; Zhong, Z.; Zou, P.; You, Y.; Cao, Y.; et al. CD44v6 chimeric antigen receptor T cell specificity towards AML with FLT3 or DNMT3A mutations. Clin Transl Med 2022, 12, e1043. [CrossRef]
- Holm, F.; Hellqvist, E.; Mason, C.N.; Ali, S.A.; Delos-Santos, N.; Barrett, C.L.; Chun, H.J.; Minden, M.D.; Moore, R.A.; Marra, M.A.; et al. Reversion to an embryonic alternative splicing program enhances leukemia stem cell self-renewal. Proc Natl Acad Sci U S A 2015, 112, 15444-15449. [CrossRef]
- Kato, Y.; Yamada, S.; Furusawa, Y.; Itai, S.; Nakamura, T.; Yanaka, M.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K. PMab-213: A Monoclonal Antibody for Immunohistochemical Analysis Against Pig Podoplanin. Monoclon Antib Immunodiagn Immunother 2019, 38, 18-24. [CrossRef]
- Furusawa, Y.; Yamada, S.; Itai, S.; Sano, M.; Nakamura, T.; Yanaka, M.; Fukui, M.; Harada, H.; Mizuno, T.; Sakai, Y.; et al. PMab-210: A Monoclonal Antibody Against Pig Podoplanin. Monoclon Antib Immunodiagn Immunother 2019, 38, 30-36. [CrossRef]
- Furusawa, Y.; Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K.; Kato, Y. PMab-219: A monoclonal antibody for the immunohistochemical analysis of horse podoplanin. Biochem Biophys Rep 2019, 18, 100616. [CrossRef]
- Furusawa, Y.; Yamada, S.; Itai, S.; Nakamura, T.; Takei, J.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K.; Kato, Y. Establishment of a monoclonal antibody PMab-233 for immunohistochemical analysis against Tasmanian devil podoplanin. Biochem Biophys Rep 2019, 18, 100631. [CrossRef]
- Kato, Y.; Kaneko, M.K.; Kuno, A.; Uchiyama, N.; Amano, K.; Chiba, Y.; Hasegawa, Y.; Hirabayashi, J.; Narimatsu, H.; Mishima, K.; et al. Inhibition of tumor cell-induced platelet aggregation using a novel anti-podoplanin antibody reacting with its platelet-aggregation-stimulating domain. Biochem Biophys Res Commun 2006, 349, 1301-1307. [CrossRef]
- Chalise, L.; Kato, A.; Ohno, M.; Maeda, S.; Yamamichi, A.; Kuramitsu, S.; Shiina, S.; Takahashi, H.; Ozone, S.; Yamaguchi, J.; et al. Efficacy of cancer-specific anti-podoplanin CAR-T cells and oncolytic herpes virus G47Delta combination therapy against glioblastoma. Mol Ther Oncolytics 2022, 26, 265-274. [CrossRef]
- Ishikawa, A.; Waseda, M.; Ishii, T.; Kaneko, M.K.; Kato, Y.; Kaneko, S. Improved anti-solid tumor response by humanized anti-podoplanin chimeric antigen receptor transduced human cytotoxic T cells in an animal model. Genes Cells 2022, 27, 549-558. [CrossRef]
- Tamura-Sakaguchi, R.; Aruga, R.; Hirose, M.; Ekimoto, T.; Miyake, T.; Hizukuri, Y.; Oi, R.; Kaneko, M.K.; Kato, Y.; Akiyama, Y.; et al. Moving toward generalizable NZ-1 labeling for 3D structure determination with optimized epitope-tag insertion. Acta Crystallogr D Struct Biol 2021, 77, 645-662. [CrossRef]
- Kaneko, M.K.; Ohishi, T.; Nakamura, T.; Inoue, H.; Takei, J.; Sano, M.; Asano, T.; Sayama, Y.; Hosono, H.; Suzuki, H.; et al. Development of Core-Fucose-Deficient Humanized and Chimeric Anti-Human Podoplanin Antibodies. Monoclon Antib Immunodiagn Immunother 2020, 39, 167-174. [CrossRef]
- Fujii, Y.; Matsunaga, Y.; Arimori, T.; Kitago, Y.; Ogasawara, S.; Kaneko, M.K.; Kato, Y.; Takagi, J. Tailored placement of a turn-forming PA tag into the structured domain of a protein to probe its conformational state. J Cell Sci 2016, 129, 1512-1522. [CrossRef]
- Abe, S.; Kaneko, M.K.; Tsuchihashi, Y.; Izumi, T.; Ogasawara, S.; Okada, N.; Sato, C.; Tobiume, M.; Otsuka, K.; Miyamoto, L.; et al. Antitumor effect of novel anti-podoplanin antibody NZ-12 against malignant pleural mesothelioma in an orthotopic xenograft model. Cancer Sci 2016, 107, 1198-1205. [CrossRef]
- Kaneko, M.K.; Abe, S.; Ogasawara, S.; Fujii, Y.; Yamada, S.; Murata, T.; Uchida, H.; Tahara, H.; Nishioka, Y.; Kato, Y. Chimeric Anti-Human Podoplanin Antibody NZ-12 of Lambda Light Chain Exerts Higher Antibody-Dependent Cellular Cytotoxicity and Complement-Dependent Cytotoxicity Compared with NZ-8 of Kappa Light Chain. Monoclon Antib Immunodiagn Immunother 2017, 36, 25-29. [CrossRef]
- Ito, A.; Ohta, M.; Kato, Y.; Inada, S.; Kato, T.; Nakata, S.; Yatabe, Y.; Goto, M.; Kaneda, N.; Kurita, K.; et al. A Real-Time Near-Infrared Fluorescence Imaging Method for the Detection of Oral Cancers in Mice Using an Indocyanine Green-Labeled Podoplanin Antibody. Technol Cancer Res Treat 2018, 17, 1533033818767936. [CrossRef]
- Tamura, R.; Oi, R.; Akashi, S.; Kaneko, M.K.; Kato, Y.; Nogi, T. Application of the NZ-1 Fab as a crystallization chaperone for PA tag-inserted target proteins. Protein Sci 2019, 28, 823-836. [CrossRef]
- Shiina, S.; Ohno, M.; Ohka, F.; Kuramitsu, S.; Yamamichi, A.; Kato, A.; Motomura, K.; Tanahashi, K.; Yamamoto, T.; Watanabe, R.; et al. CAR T Cells Targeting Podoplanin Reduce Orthotopic Glioblastomas in Mouse Brains. Cancer Immunol Res 2016, 4, 259-268. [CrossRef]
- Kuwata, T.; Yoneda, K.; Mori, M.; Kanayama, M.; Kuroda, K.; Kaneko, M.K.; Kato, Y.; Tanaka, F. Detection of Circulating Tumor Cells (CTCs) in Malignant Pleural Mesothelioma (MPM) with the "Universal" CTC-Chip and An Anti-Podoplanin Antibody NZ-1.2. Cells 2020, 9. [CrossRef]
- Nishinaga, Y.; Sato, K.; Yasui, H.; Taki, S.; Takahashi, K.; Shimizu, M.; Endo, R.; Koike, C.; Kuramoto, N.; Nakamura, S.; et al. Targeted Phototherapy for Malignant Pleural Mesothelioma: Near-Infrared Photoimmunotherapy Targeting Podoplanin. Cells 2020, 9. [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; Nogi, T.; Kato, Y.; Takagi, J. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014, 95, 240-247. [CrossRef]
- Kato, Y.; Kaneko, M.K.; Kunita, A.; Ito, H.; Kameyama, A.; Ogasawara, S.; Matsuura, N.; Hasegawa, Y.; Suzuki-Inoue, K.; Inoue, O.; et al. Molecular analysis of the pathophysiological binding of the platelet aggregation-inducing factor podoplanin to the C-type lectin-like receptor CLEC-2. Cancer Sci 2008, 99, 54-61. [CrossRef]
- Kato, Y.; Vaidyanathan, G.; Kaneko, M.K.; Mishima, K.; Srivastava, N.; Chandramohan, V.; Pegram, C.; Keir, S.T.; Kuan, C.T.; Bigner, D.D.; et al. Evaluation of anti-podoplanin rat monoclonal antibody NZ-1 for targeting malignant gliomas. Nucl Med Biol 2010, 37, 785-794. [CrossRef]
Figure 1.
Anti-human CD44 mAbs production. (A) The CD44 structure. CD44s mRNA is assembled by the first five (1 to 5) and the last five (16 to 20) exons, and translates CD44s. CD44v mRNAs are generated by the alternative splicing of variant exons, and translate multiple CD44v isoforms, such as CD44v3-10, CD44v4-10, CD44v6-10, and CD44v8-10. (B) BALB/c mice were intraperitoneally immunized with CHO/CD44v3–10 cells. (C) The hybridomas were produced by fusion of the splenocytes and P3U1 cells. (D) The flow cytometry-mediated screening was conducted using parental CHO-K1 and CHO/CD44v3–10 cells. (E) After cloning and additional screening, a clone C44Mab-6 (IgG1, kappa) was established. Finally, the binding epitopes were determined by enzyme-linked immunosorbent assay (ELISA) using peptides, which cover the extracellular domain of CD44v3–10.
Figure 1.
Anti-human CD44 mAbs production. (A) The CD44 structure. CD44s mRNA is assembled by the first five (1 to 5) and the last five (16 to 20) exons, and translates CD44s. CD44v mRNAs are generated by the alternative splicing of variant exons, and translate multiple CD44v isoforms, such as CD44v3-10, CD44v4-10, CD44v6-10, and CD44v8-10. (B) BALB/c mice were intraperitoneally immunized with CHO/CD44v3–10 cells. (C) The hybridomas were produced by fusion of the splenocytes and P3U1 cells. (D) The flow cytometry-mediated screening was conducted using parental CHO-K1 and CHO/CD44v3–10 cells. (E) After cloning and additional screening, a clone C44Mab-6 (IgG1, kappa) was established. Finally, the binding epitopes were determined by enzyme-linked immunosorbent assay (ELISA) using peptides, which cover the extracellular domain of CD44v3–10.

Figure 2.
Determination of C44Mab-6 epitope by ELISA. The synthesized peptides, which cover the CD44v3–10 extracellular domain, were immobilized on immunoplates. The plates were incubated with C44Mab-6, followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins. Optical density was measured at 655 nm. The CD44p231–250 sequence (AGWEPNEENEDERDRHLSFS) is corresponding to variant 3-encoded sequence. ELISA, enzyme-linked immunosorbent assay. NC, negative control (solvent; DMSO in PBS).
Figure 2.
Determination of C44Mab-6 epitope by ELISA. The synthesized peptides, which cover the CD44v3–10 extracellular domain, were immobilized on immunoplates. The plates were incubated with C44Mab-6, followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins. Optical density was measured at 655 nm. The CD44p231–250 sequence (AGWEPNEENEDERDRHLSFS) is corresponding to variant 3-encoded sequence. ELISA, enzyme-linked immunosorbent assay. NC, negative control (solvent; DMSO in PBS).
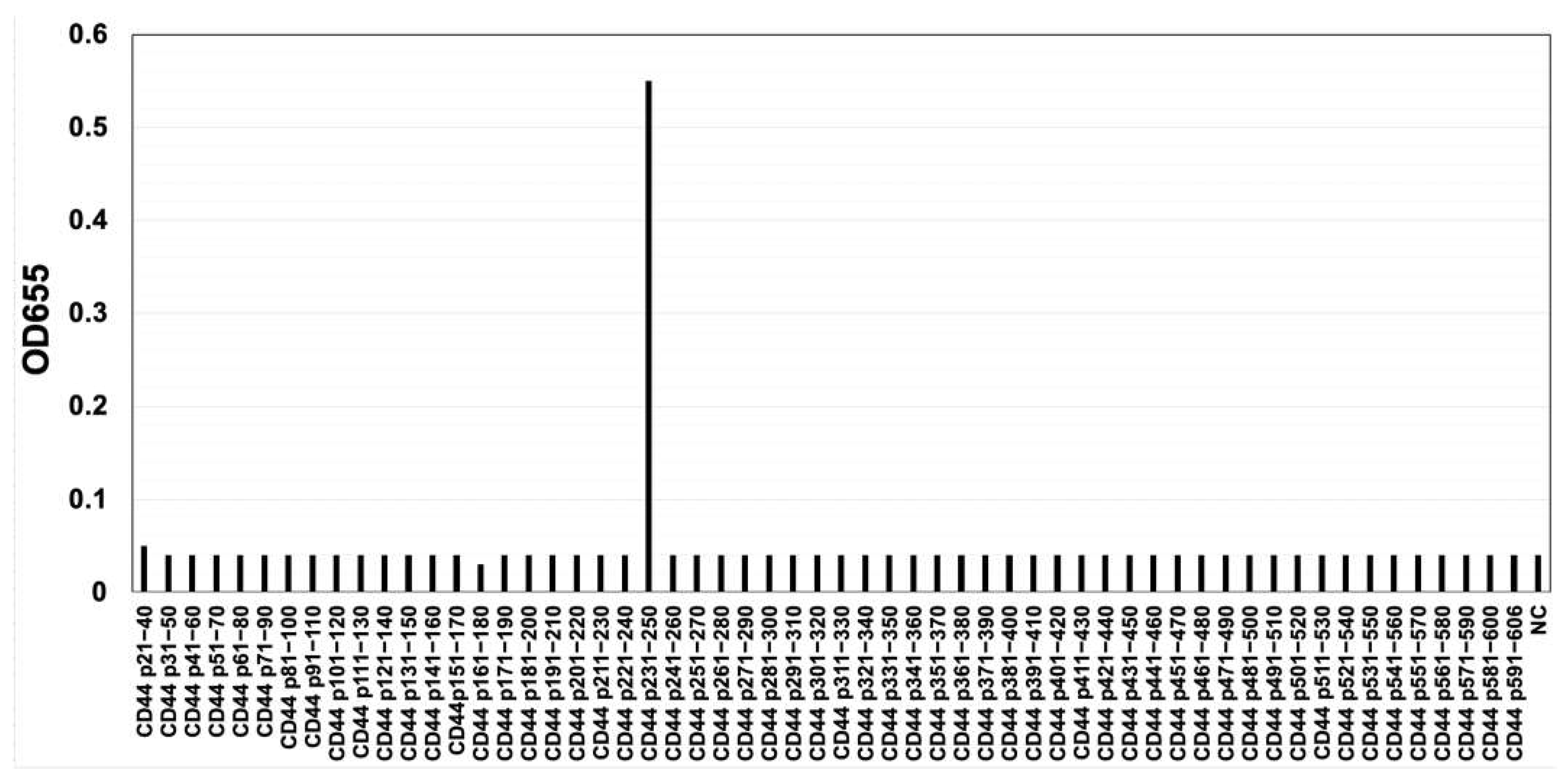
Figure 3.
The reactivity of C44Mab-6 to CD44-Expressing Cells in Flow Cytometry. CHO/CD44v3–10 (A), CHO/CD44s (B), CHO-K1 (C), COLO205 (D), and HSC-3 (E) cells were treated with C44Mab-6 at 0.01-10 µg/mL, followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488 (Red line). Black line, negative control (blocking buffer).
Figure 3.
The reactivity of C44Mab-6 to CD44-Expressing Cells in Flow Cytometry. CHO/CD44v3–10 (A), CHO/CD44s (B), CHO-K1 (C), COLO205 (D), and HSC-3 (E) cells were treated with C44Mab-6 at 0.01-10 µg/mL, followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488 (Red line). Black line, negative control (blocking buffer).
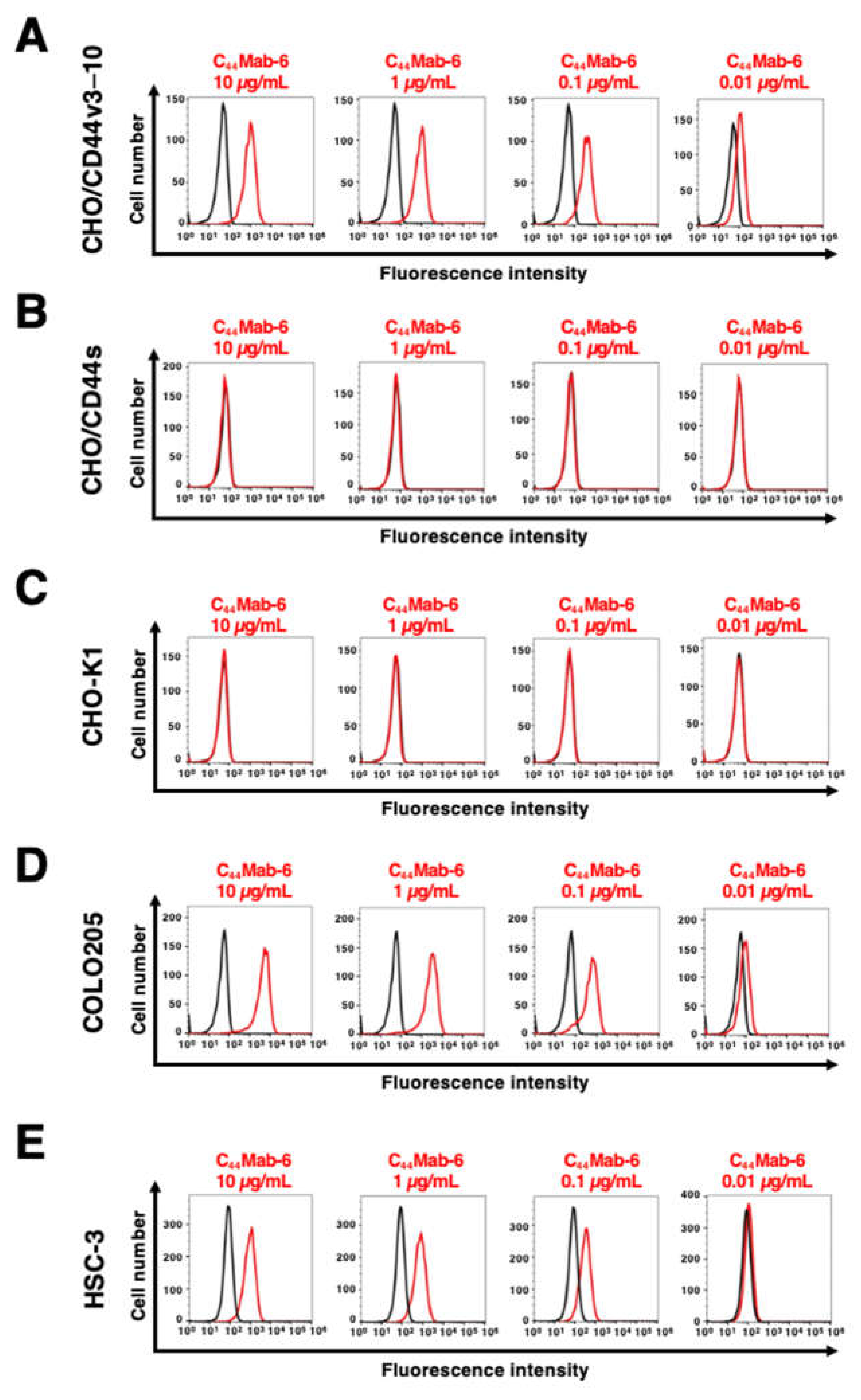
Figure 4.
The binding affinity of C44Mab-6 to CD44-expressing cells. CHO/CD44v3–10 (A), COLO205 (B), and HSC-3 (C) cells were suspended in C44Mab-6 at the indicated concentrations. Cells were incubated with Alexa Fluor 488-conjugated secondary antibody. Fluorescence data were collected, and the apparent dissociation constant (KD) was calculated of by GraphPad PRISM 8.
Figure 4.
The binding affinity of C44Mab-6 to CD44-expressing cells. CHO/CD44v3–10 (A), COLO205 (B), and HSC-3 (C) cells were suspended in C44Mab-6 at the indicated concentrations. Cells were incubated with Alexa Fluor 488-conjugated secondary antibody. Fluorescence data were collected, and the apparent dissociation constant (KD) was calculated of by GraphPad PRISM 8.
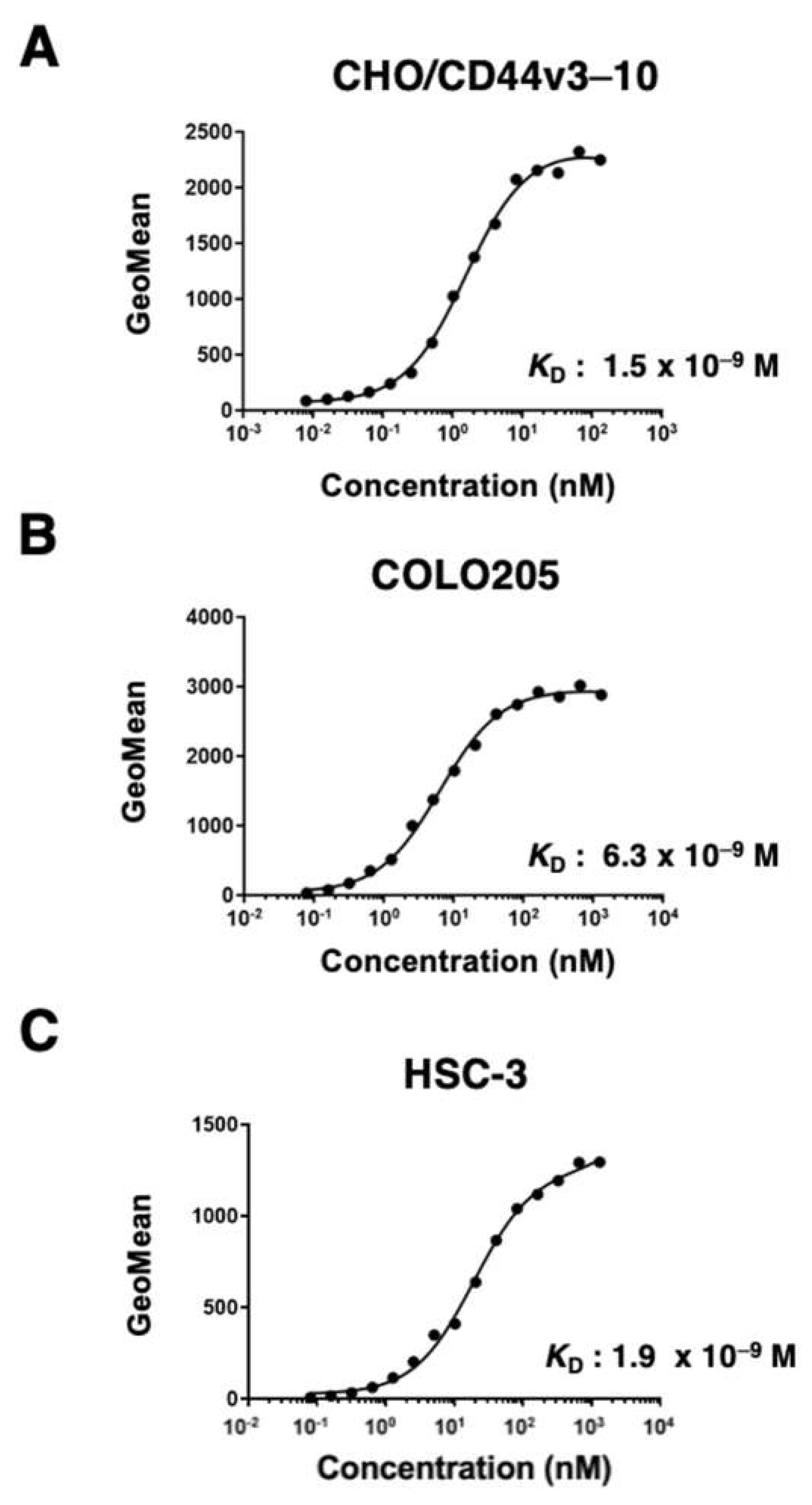
Figure 5.
Western blot analysis using C44Mab-6. The cell lysates of CHO-K1, CHO/CD44s, and CHO/CD44v3–10 (10 µg) were electrophoresed, and transferred onto polyvinylidene fluoride membranes. The membranes were incubated with 10 µg/mL of C44Mab-6 (A), 10 µg/mL of C44Mab-46 (B), and 1 µg/mL of an anti-β-actin mAb (C). Then, the membranes were incubated with anti-mouse immunoglobulins conjugated with peroxidase. The black arrow indicates the CD44s (~75 kDa). The red arrows indicate the CD44v3–10 (>180 kDa).
Figure 5.
Western blot analysis using C44Mab-6. The cell lysates of CHO-K1, CHO/CD44s, and CHO/CD44v3–10 (10 µg) were electrophoresed, and transferred onto polyvinylidene fluoride membranes. The membranes were incubated with 10 µg/mL of C44Mab-6 (A), 10 µg/mL of C44Mab-46 (B), and 1 µg/mL of an anti-β-actin mAb (C). Then, the membranes were incubated with anti-mouse immunoglobulins conjugated with peroxidase. The black arrow indicates the CD44s (~75 kDa). The red arrows indicate the CD44v3–10 (>180 kDa).

Figure 6.
Immunohistochemical analysis using C44Mab-6 and C44Mab-46 against OSCC tissues. After antigen retrieval, serial sections of OSCC tissue array (Catalog number: OR601c) were incubated with 10 μg/mL of C44Mab-6 (A, C, E) or 1 μg/mL of C44Mab-46 (B, D, F), followed by treatment with the Envision+ kit. The chromogenic reaction was conducted using 3,3′-diaminobenzidine tetrahydrochloride (DAB). The counterstaining was performed using hematoxylin. Scale bar = 100 μm.
Figure 6.
Immunohistochemical analysis using C44Mab-6 and C44Mab-46 against OSCC tissues. After antigen retrieval, serial sections of OSCC tissue array (Catalog number: OR601c) were incubated with 10 μg/mL of C44Mab-6 (A, C, E) or 1 μg/mL of C44Mab-46 (B, D, F), followed by treatment with the Envision+ kit. The chromogenic reaction was conducted using 3,3′-diaminobenzidine tetrahydrochloride (DAB). The counterstaining was performed using hematoxylin. Scale bar = 100 μm.


Table 1.
Immunohistochemical analysis using C44Mab-6 and C44Mab-46 against OSCC tissue arrays.
| No. | Age | Sex | Anatomic Site | Pathology diagnosis | TNM | Grade | Type | C44Mab-6 | C44Mab-46 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 78 | M | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | ++ | + |
| 2 | 40 | M | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | ++ | ++ |
| 3 | 75 | F | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | + | + |
| 4 | 35 | F | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | ++ | ++ |
| 5 | 61 | M | Tongue | SCC a of tongue | T2N0M0 | 1 | Malignant | +++ | +++ |
| 6 | 41 | F | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | + | + |
| 7 | 64 | M | Tongue | SCC of right tongue | T2N2M0 | 1 | Malignant | ++ | ++ |
| 8 | 76 | M | Tongue | SCC of tongue | T1N0M0 | 1 | Malignant | ++ | ++ |
| 9 | 50 | F | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | ++ | ++ |
| 10 | 44 | M | Tongue | SCC of tongue | T2N1M0 | 1 | Malignant | +++ | +++ |
| 11 | 53 | F | Tongue | SCC of tongue | T1N0M0 | 1 | Malignant | ++ | ++ |
| 12 | 46 | F | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | + | + |
| 13 | 50 | M | Tongue | SCC of root of tongue | T3N1M0 | 1 | Malignant | +++ | + |
| 14 | 36 | F | Tongue | SCC of tongue | T1N0M0 | 1 | Malignant | +++ | +++ |
| 15 | 63 | F | Tongue | SCC of tongue | T1N0M0 | 1 | Malignant | ++ | + |
| 16 | 46 | M | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | +++ | - |
| 17 | 58 | M | Tongue | SCC of tongue | T2N0M0 | 1 | Malignant | + | + |
| 18 | 64 | M | Lip | SCC of lower lip | T1N0M0 | 1 | Malignant | +++ | +++ |
| 19 | 57 | M | Lip | SCC of lower lip | T2N0M0 | 1 | Malignant | +++ | +++ |
| 20 | 61 | M | Lip | SCC of lower lip | T1N0M0 | 1 | Malignant | ++ | ++ |
| 21 | 60 | M | Gum | SCC of gum | T3N0M0 | 1 | Malignant | + | + |
| 22 | 60 | M | Gum | SCC of gum | T1N0M0 | 1 | Malignant | +++ | +++ |
| 23 | 69 | M | Gum | SCC of upper gum | T3N0M0 | 1 | Malignant | ++ | ++ |
| 24 | 53 | M | Bucca cavioris | SCC of bucca cavioris | T2N0M0 | 1 | Malignant | + | + |
| 25 | 55 | M | Bucca cavioris | SCC of bucca cavioris | T1N0M0 | 1 | Malignant | ++ | + |
| 26 | 58 | M | Tongue | SCC of base of tongue | T1N0M0 | 1 | Malignant | ++ | ++ |
| 27 | 63 | M | Oral cavity | SCC | T1N0M0 | 1 | Malignant | ++ | ++ |
| 28 | 48 | F | Tongue | SCC of tongue | T1N0M0 | 1--2 | Malignant | ++ | + |
| 29 | 80 | M | Lip | SCC of lower lip | T1N0M0 | 1--2 | Malignant | +++ | +++ |
| 30 | 77 | M | Tongue | SCC of base of tongue | T2N0M0 | 1--2 | Malignant | +++ | ++ |
| 31 | 59 | M | Tongue | SCC of tongue | T2N0M0 | 2 | Malignant | + | - |
| 32 | 77 | F | Tongue | SCC of tongue | T1N0M0 | 2 | Malignant | ++ | ++ |
| 33 | 56 | M | Tongue | SCC of root of tongue | T2N1M0 | 2 | Malignant | + | + |
| 34 | 60 | M | Tongue | SCC of tongue | T2N1M0 | 2 | Malignant | ++ | ++ |
| 35 | 62 | M | Tongue | SCC of tongue | T2N0M0 | 2 | Malignant | +++ | ++ |
| 36 | 67 | F | Tongue | SCC of tongue | T2N0M0 | 2 | Malignant | +++ | ++ |
| 37 | 47 | F | Tongue | SCC of tongue | T2N0M0 | 2 | Malignant | +++ | +++ |
| 38 | 37 | M | Tongue | SCC of tongue | T2N1M0 | 2 | Malignant | - | - |
| 39 | 55 | F | Tongue | SCC of tongue | T2N0M0 | 2 | Malignant | ++ | + |
| 40 | 56 | F | Bucca cavioris | SCC of bucca cavioris | T2N0M0 | 2 | Malignant | +++ | + |
| 41 | 49 | M | Bucca cavioris | SCC of bucca cavioris | T1N0M0 | 2 | Malignant | - | - |
| 42 | 45 | M | Bucca cavioris | SCC of bucca cavioris | T2N0M0 | 2 | Malignant | - | - |
| 43 | 42 | M | Bucca cavioris | SCC of bucca cavioris | T3N0M0 | 2 | Malignant | +++ | ++ |
| 44 | 44 | M | Jaw | SCC of right drop jaw | T1N0M0 | 2 | Malignant | ++ | +++ |
| 45 | 40 | F | Tongue | SCC of base of tongue | T2N0M0 | 2 | Malignant | - | ++ |
| 46 | 49 | M | Bucca cavioris | SCC of bucca cavioris | T1N0M0 | 2 | Malignant | +++ | +++ |
| 47 | 56 | F | Tongue | SCC of base of tongue | T2N0M0 | 3 | Malignant | - | + |
| 48 | 42 | M | Bucca cavioris | SCC of bucca cavioris | T3N0M0 | 3 | Malignant | +++ | +++ |
| 49 | 87 | F | Face | SCC of left face | T2N0M0 | 3 | Malignant | + | + |
| 50 | 50 | M | Gum | SCC of gum | T2N0M0 | 3 | Malignant | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



