
Submitted:
17 April 2023
Posted:
18 April 2023
You are already at the latest version
Abstract
Bone metabolism is a complex process which is influenced by the activity of bone cells (e.g., osteocytes, osteoblasts, osteoclasts), the effect of some specific biomarkers (e.g., parathyroid hormone, vitamin D, alkaline phosphatase, osteocalcin, osteopontin, osteoportegerin, osterix, RANKL, Runx2) and the characteristic signaling pathways (e.g., RANKL/RANK, Wnt/β, Notch, BMP, SMAD). Some phytochemical compounds such as flavonoids, tannins, polyphenols, anthocyanins, terpenoids, polysaccharides, alkaloids and others presented a beneficial and stimulating effect in the bone regeneration process due to pro-estrogenic activity, antioxidant and anti-inflammatory effects and modulation of bone signaling pathways. Lately, nanomedicine has emerged as an innovative concept for new treatments in bone related pathologies envisaged by incorporation of medicinal substances in nanometric systems for oral or local administration, as well as in nanostructured scaffolds with huge potential in bone tissue engineering.
Keywords:
bone regeneration
; bone cells
; bone biomarkers
; bone signaling pathways
; plants extracts
; phytochemicals compounds
; nanometric systems
; nanostructured scaffolds
1. Introduction
Bone is a complex tissue that shows great resistance, with an intense activity that ensures multiple functions and that is in permanent renewal. Bone regeneration maintains the integrity of the tissue and is coordinated by two processes: bone resorption carried out by osteoclasts, through which the removal of aged cells takes place, and the second process is the formation of new bone tissue by osteoblasts, the two processes are in balance [1]. The bone remodeling process is a continuous and cyclic one that takes place at the level of a temporary anatomical structure called: Basic Multicellular Unit (MBU) [2]. Bone remodeling at the MBU level takes place in three phases: i) Initiation: the selection of osteoclastic precursors and their differentiation into mature osteoclast cells and their activation for bone resorption; ii) Inversion includes the inhibition of osteoclast activity and their apoptosis, as well as the differentiation of osteoblasts; iii) The Terminal Phase involves the formation of new bone tissue by osteoblasts [3].
Bone formation or osteogenesis performed by the osteoblast and bone resorption or osteolysis achieved by the osteoclast are the main processes that maintain bone homeostasis. The balance between these two processes is regulated by hormonal and signaling pathways.
Parathyroid hormone (PTH) is the most important hormonal bone metabolism regulator. Normal action of PTH determines the reabsorption of calcium from the bone level. PTH binds to its receptor, type 1PTH receptor (PTHR1), which leads to the activation of protein kinase A (PKA) or protein kinase C (PKC), using these signaling pathways PTH can determine both bone formation and bone resorption [4]. PTHR1 is found on the surface of several cells: osteoblasts, osteoclasts, and renal tubule cells. The effect on osteoblasts decreases cell apoptosis, and on osteoclasts it decreases sclerostin production [5,6]. The blood levels of PTH are directly influenced by the Ca+2 and Vitamin D concentrations, low levels of Vitamin D causes hyperparathyroidism [7]. Low levels of Vitamin D led to rickets in children and osteomalacia in adults, meantime Vitamin D supplements show an increase of Calcium levels, bone density and reduce bone resorption and the fractures risk [8,9].
In order to better understand bone metabolism, we will shortly review the bone cells, biomarkers involved in bone metabolism and bone regeneration and their main regulatory pathways. Additionally, the biomaterials used for bone regeneration, the plant extracts, and the phytochemical compounds with a positive effect on the bone regeneration process as well as the pathogenetic mechanisms involved were described.
1.1. Bone metabolism:
1.1.1. The main cells of bone metabolism
Three main cell types are involved in the bone tissue constitution: osteoblast responsible with new bone formation, osteoclast which conducts the osteolysis and osteocytes, the mature osteoblasts, located in the bone matrix, responsible for bone mineralization.
Osteocytes: they are cells located at the level of the bone matrix, which derive from mature osteoblasts, producing bone matrix [10]. They represent approximately 90-95% of bone cells, have a long life of up to 25 years, are made up of the cell body and dendrites and are found at the level of the lacuno-canalicular system, which allows their connection with adjacent osteocytes, osteoblasts, osteoclasts, bone marrow, blood vessels and nerves [11]. The osteocyte plays a main role in bone homeostasis, acting as a mechanical sensor and mechanical transducer to the different stimuli to which the bone is subjected, controlling through chemical and hormonal responses both the process of osteolysis by activating the cellular differentiation process of osteoclasts, as well as that of osteogenesis through the formation of young osteocytes from osteoblasts [12,13].
Osteoblasts: they are small, mononucleated, cubic-shaped cells, but they can also appear in a flat or cylindrical form, and they derive from the mesenchymal cells located in the bone marrow. Their differentiation takes place under the action of a network of cytokines and transcription factors [14]. The mesenchymal cells located at the level of the bone marrow have a multipotent character from their level deriving several cell lines: adipocytes, chondrocytes and osteoblasts [15]. Osteoblastogenesis is the cellular differentiation process of osteoblasts from mesenchymal cells, this process takes place in three phases: proliferation, matrix maturation and mineralization. The three phases are characterized by the expression of specific osteoblastic genetic markers such as: osteocalcin (OCN), osteopontin (OPN), bone sialoprotein (BSP), collagen type I (COL1A) and alkaline phosphatase (ALP). Osteoblastogenesis is controlled by hormonal factors (parathormone and glucocorticoids), specific signaling pathways (Wnt, BMP, Hedgehog, Notch), circulating cytokines and transcription factors (Runx2 and Osx1), fibroblast growth factor (FGF), transforming growth factor β (TGFβ) [16].
Osteoclasts: are the cells responsible for bone resorption. They are large, multinucleated ones that derive from hematopoietic cells, formed by the fusion of mononuclear precursors of the monocyte-macrophage line. The cellular differentiation process requires coordination of transcription factors with co-activating and co-repressing factors [17]. In the process of cellular differentiation and maturation of osteoclasts, a central role is played by the activator receptor of nuclear factor kB ligand (RANKL). Secreted by osteoblasts, bone marrow cells and lymphocytes, RANKL is part of the tumor necrosis factor family, which interacts with the RANK receptor on the cells of preosteoclasts, promoting their differentiation into osteoresorbing mature osteoclasts [18].
1.1.2. The main biomarkers of bone metabolism
Bone metabolism represents the total of biochemical processes through which osteolysis and osteogenesis take place, and represents both the metabolism of proteins and that of the minerals that enter the composition of the bone. The regulation of this metabolism is done by a series of hormonal or enzymatic biomarkers.
Parathyroid hormone (PTH): influences the biochemical processes that take place in bone remodeling. Low PTH values cause a decrease in bone circulatory markers, both osteoforming and bone resorption markers, which causes a decrease in bone turnover, while in hypoparathyroidism bone mineral density increases. Treatment of hypoparathyroidism with human parathyroid hormone (hPTH) or recombinant human parathyroid hormone (rhPTH) increases bone turnover, while bone mineral density shows increases at the hip and spine level and decreases at the radiocarpal level [19].
Vitamin D: its deficiency determines the occurrence of rickets in children and osteomalacia in adults, being at the same time associated with the occurrence of osteoporosis and the increased risk of fractures. Recent studies have demonstrated the effect of 25-hydroxy vitamin D3 and its metabolite 1α,25 hydroxy vitamin D3 to differentiate mesenchymal stem cells of human origin towards osteoblasts [20].
Alkaline phosphatase: the bone isozyme, is a glycoprotein attached to the cell membrane of osteoblasts, from where it is released into the circulation, being the main indicator of osteogenesis. It participates in the synthesis of hydroxyapatite, providing inorganic phosphate, pyrophosphate and monophosphoesters and in the same time it hydrolyzes pyrophosphate, which is an inhibitor of the mineralization process [21,24]
Osteocalcin: is the most abundant non-collagenous protein at the bone level, dependent on vitamin K, being produced by osteoblasts. It presents in its structure three gamma-carboxyglutamic acids that have an affinity for the Ca2+ ion, which will influence the bone remodeling and mineralization processes, acting at the same time as an inhibitor of the hydroxyapatite growth and a regulator of the activity of osteoclastic precursors [22,23].
Osteopontin: is a phosphoprotein secreted at bone level, especially by osteoblasts, with a role in bone metabolism in which it participates through endocrine, neurological, and immunological processes. It acts as a parathormone regulator, low osteopontin values blocking the PTH activity of stimulating alkaline phosphatase and osteocalcin expression [24,27].
RANKL/RANK: the activator receptor of the NF-kB ligand factor (RANKL) is a homotrimeric protein secreted by osteoblasts with multiple functions: it stimulates the differentiation of preosteoblasts, the adhesion of osteoclasts to the bone tissue, as well as the reduction and maintenance of their activity [25].
Osteoportegerin (OPG): is a glycoprotein secreted specifically by osteoblasts, being a cytokine receptor of the Tumor Necrosis Factor (TNF), acting as a "bait" receptor for RANKL, inhibiting osteoclastogenesis and bone resorption [26].
Osterix: is a protein with a role in the differentiation of mesenchymal cells into osteoblasts, inhibiting the formation of chondrocytes. At the osteoblastic level, it indicates the genetic expression of: osteopotin, oteonectin, type 1a1 collagen and bone sialoprotein, which are necessary for the process of bone mineralization at the level of osteoblasts [27].
Runx2: is an essential protein in the maturation process of osteoblasts, its expression is weak in mesenchymal cells and maximum in immature osteoblasts, being essential in the differentiation of mesenchymal cells into osteoblastic cell lines [28].
1.1.3. The main signaling pathways specific to bone metabolism
Together with the regulation hormonal of bone homeostasis, an important role in this process belongs to the signaling pathways. They are involved in the processes of embryonic bone development, as well as in the processes of bone repair [29].
RANKL/RANK/OPG: RANKL is produced by osteoblasts and osteocytes, its binding to RANK at the level of osteoclastic precursor cells determines the differentiation of this cell line, favoring osteoclastogenesis. OPG secreted by osteoblasts is a "bait" receptor for RANKL, preventing its binding to RANK, thereby preventing osteoclastogenesis and bone resorption [25].
Wnt/β signaling: the Wnt pathway stimulates osteoblast activation and differentiation. Canonical Wnt causes β-catenin translocation and stabilization in the cell nucleus, which regulates gene transcription in response to Wnt signaling [30].
Notch: at the bone level, Notch receptors and their ligands are responsible for a multitude of phenomena: osteoblastic differentiation, bone matrix mineralization, osteoclast recruitment, cell fusion and osteoblast/osteoclast cell proliferation [31].
Bone morphogenetic proteins (BMPs): part of the transforming growth factor β (TGFβ) family, they are involved in the processes of bone formation, through the differentiation of osteoblasts. A central role in the BMP pathway signaling is played by SMAD 1, 5 and 8 proteins that interact with BMP receptors [32,33].
1.2. Biomaterials for bone regeneration
During many decades, a wide range of biomaterials have been developed to address the bone defects promoting the bone regeneration through various mechanisms such as: mechanical support, osteoconduction, osteoinduction, vascularization, neurotization, antibacterial effect etc. [34]. Primarily, the autologous bone grafts were considered the “gold standard” material for bone defects, but due to some disadvantages (limited quantity, long surgical procedures, and morbidity) they have been replaced by synthetic bone graft, therefore a plethora of emerging biomaterials have been designed possessing specific advantageous characteristics [35]. According to their dimensional structure and dimension size (as nanoscale), the biomaterials have been classified in zero-, one-, two-, three- and four-dimensional biomaterials [34]. Frequently, different classes of biomaterials have been combined leading to hybrid composite biomaterials with synergic properties for bone tissue regeneration.
The zero-dimensional biomaterials have the all three dimensions confined at nanoscale with a high surface-to-volume ratio including some carbon-based nanomaterials (fullerene, nanodiamonds, carbon dots) and inorganic nanoparticles (NPs) (AuNPs, AgNPs, iron oxide NPs, etc.). They presented great benefits for bone regeneration such as biomineralization, osteogenic differentiation, good mechanical performance, and biocompatibility.
The one-dimensional biomaterials (two dimensions are nanosized) presenting high length-to-diameter ratio and unique nanotopography refer to nanowires (silicon nanowire - SiNW) and nanotubes (Titanium oxide nanotubes - TiO2NTs, carbon nanotubes - CNTs). They modulated osteogenic and chondrogenic cell adhesion, proliferation, and differentiation, also facilitated mineralization and demonstrated an exceptional bone-tissue compatibility [34].
The two-dimensional biomaterials (one dimension is in nanoscale range) being characterized by a high diameter-to-thickness ratio include graphene and its derivatives (graphene oxide, reduced graphene oxide). The exceptional osteoinductive property (in vitro and in vivo), along with enhanced mechanical properties, favorable biocompatibility, facilitated mineralization have been attributed to graphene due to various interactions with biomolecules and physical stress (affecting cytoskeletal tension and inducing cytoskeletal reorganization). Other nanofilms coatings (calcium phosphate coatings, black phosphorus (BP) nanofilms) have been applied for facilitating the biomaterial integration [34,35].
The three-dimensional biomaterials (all dimensions are larger than the nanoscale) are the most used implants in clinics including metallic (titanium and its alloy, silver, magnesium, niobium, strontium, stainless steel, cobalt, tantalum) [36], bioceramic (bioactive bioceramics: calcium phosphate ceramics, hydroxyapatite, bioglass, bioinert bioceramics: alumina, zirconia, silicon carbide) [37] and polymeric scaffolds and hydrogels. The disadvantages of metal-based scaffolds (poor biodegradability, local/systemic toxicity, higher elastic modulus) were overcome by fabricating scaffolds with tunable porosity [34].
From the group of bioceramics, calcium phosphates and bioactive glass are the most frequently used in orthopedic and dental field. Calcium phosphates (with various forms, tunable porosity, and density) resemble with the native bone tissue having a good capacity of integration, great bioactivity and osteoconductivity [34]. Hydroxyapatite (HA) (one example of calcium phosphate) is an intrinsic component of bone tissue presenting excellent biocompatibility, osseointegration, osteoconductivity, osteoinductivity, and angiogenic effects. Biodegradable polymers have been mixed with HA in order to solve its limitations (brittleness and insufficient mechanical strength). Bioactive glass (BG) with main component silicate, borate and phosphate glass presents a fabulous surface reactivity, bioactivity and osteoinductivity, but also brittleness (disadvantage solved by polymer inclusion) [34].
Polymeric scaffolds are based on natural polymers (collagen, chitosan, hyaluronic acid, silk, alginate, gelatin, cellulose, etc.) and synthetic polymers (polylactic acid (PLA), poly (glycolic acid) (PGA), poly(lactic-co-glycolic acid) (PLGA), poly(ethylene glycol) (PEG), polycaprolactone (PCL), polyurethane (PU) etc.) [[38,39]]. Both classes are ascribed with pros and cons: biocompatibility, design flexibility, supporting cell attachment, osteogenic differentiation, calcium biomineralization, easy tailoring of microstructure, hydrophilicity, pore size, porosity, mechanical characteristics, degradability; and immunogen and pathogenic impurities, poor replicability, need of crosslinking and inferior loading-bearing capacity, respectively [34].
Hydrogels are based on hydrophilic polymers (natural, synthetic or hybrid) with hydrophilic nature, high water content and permeability being substrates for supporting cell growth, therefore promoting osteogenesis, calcium biomineralization and angiogenesis [34].
The four-dimensional biomaterials are a new smart generation exhibiting dynamic self-remodeling capability and tunable stimuli-responsiveness. They may contain hydrogels, bioceramics, piezoelectric materials, etc. that undergo self-transformation of shape and functionality after stimuli exposure [34].
The biomaterials/scaffolds properties such as: pore size, stiffness, scaffold composition, surface topography, surface functional groups, surface wettability and degradation product are crucial for their effects on cellular behavior being taken into consideration for the appropriate selection of a certain biomaterial-based scaffold [40]. The techniques applied for manufacturing the bone tissue regeneration scaffolds are freeze-drying, electrospinning, 3D printing (selective laser sintering, stereolithography, fused deposition modeling), solvent casting, sol-gel, gas foaming and particulate leaching [36]. Lately, a new3D bioprinting technology (inkjet-based bioprinting, extrusion-based bioprinting, laser-based bioprinting) has emerged depositing living cells, extracellular matrices and biomaterials [41].
Bone pathology includes a range of multiple pathologies, including bone tumors, metastases, fractures, bone deformity, osteoporosis, osteoarthritis, etc. Regardless of the type of bone pathology, they are characterized by osteoblast dysfunctions and bone reabsorption. Bone tissue is made up of organic elements such as type I collagen and inorganic elements such as hydroxyapatite. These elements vary according to age, sex, related pathologies and even the daily diet. The processes of bone degradation can therefore be postponed or even blurred, taking into account these factors, of which the daily diet is the most accessible to the patient with bone pathology.
The inclusion of some plants, plant extracts or phytochemical compounds in the daily diet and in the biomaterials-based scaffolds composition can improve the bone regeneration process. Phytochemical compounds from the classes of anthocyanins, phenols and flavonoids presented a beneficial and stimulating effect in the bone regeneration process.
Thus, the purpose of this review is to summarize the recent information regarding the implications of phytochemical extracts and their mechanism of action in the bone regeneration process.
2. Research methodology
The studies considered were selected using the PRISMA 2020 flow diagram according to the author Page et al. 2021. The steps and selection criteria, followed by the number of the studies used for our review, are shown in Figure 1. Databases such as PubMed, Scopus, Science Direct, Elsevier, Google Scholar, Google Patents were accessed to search the literature. The Medical Subject Headings keywords included in the search were: "bone regeneration", “bone markers”, “bone signaling pathways”, “bone metabolism”, "bone pathology", "plant extract", “plant bone regeneration”, “natural compounds bone regeneration”, “elagic acid bone”, “bone regeneration materials”.
All information systematized in the tables was obtained from research articles (in vivo /in vitro studies) and reviews between 2012 to 2022 (last 10 years). A Prisma flow-diagram was used for description how to select the studies, articles included in the review, shown in Figure 1 [42,43].
Studies published in languages other than English were excluded. A total of 118 studies were selected and included in this review.
3. Plant extracts and phytochemical compounds with a positive effect on the bone regeneration process
3.1. Classes of phytochemical compounds involved in the bone regeneration process
Natural products exhibit a wide range of modulatory effects on various pathways involved in bone regeneration context (osteoclastogenesis inhibition, bone anabolism and bone resorption) such as: NF-κB signaling pathways, MAPKs signaling pathways, Akt signaling pathways, calcium ion (Ca2+) signaling, ROS-mediated effects, and inflammatory mediator genes (Fig. 1) [44,45]. Some phytochemical compounds have been effective for improving the bone regeneration and preventing/treating osteoporosis and osteoarthritis enhancing some mechanisms such as: mineral turnover, bone mineral-density, inhibition of bone-loss, increase of calcium and vitamin D3, prevention of inflammation and oxidative stress [46].
The phytocompounds which proved their potential in orthopedic applications belong to the following classes: flavonoids, tannins, polyphenols, anthocyanins, terpenoids, polysaccharides, alkaloids and others (Figure 1) [44,45,47]. Many researchers pointed out that a certain activity is due to the complex composition of the plant product (including the interactions such as synergistic, additive, or antagonistic effect) rather than to a single compound, solvent extraction and doses [48].
3.2. Phytocompounds used in the bone regeneration process – state of the art
Currently, many conventional treatments are known whose mechanism of action is to stop reabsorption (anti-resorptive drugs) or anabolic drugs, but these are also known for their characteristic adverse effects after long-term administration.
The treatment of osteoporosis can be carried out both with medicinal agents from the mentioned classes individually and as a combined treatment. It can be mentioned as a classic treatment against the loss of bone density the combined therapy between anabolic and antiresorptive drugs such as retiparatide, romosozumab and bisphosphonates, selective modulators of estrogen receptors, as well as the continuous administration of calcium combined with vitamin D. However, the demonstrated adverse effects suggest that these classical treatments should be administered with caution and not on long term, but bone diseases such as osteoporosis and osteoarthritis are chronic diseases, so this premise is not valid.
Some of the adverse effects in the case of long-term administration of the drug classes involved in conventional treatment are the occurrence of thrombotic events, the occurrence of breast cancer, the occurrence of kidney diseases (especially with continuous administration of calcium together with vitamin D), gastrointestinal diseases and cardiovascular events.
Taking into account these aspects, the treatment plan for bone diseases such as osteoporosis brings to the fore the treatment with natural molecules, phytochemicals, these being considered of major interest in terms of the treatment and stimulation of bone regeneration, being considered free of adverse effects for the patient.
According to the mechanism of action, natural compounds can be divided into three large classes such as: compounds with pro-estrogenic activity, compounds with antioxidant and anti-inflammatory properties and modulatory compounds of bone regeneration pathways.
Estrogen has multiple effects on bone metabolism: through its action on osteocytes, it inhibits bone remodeling, it also inhibits bone resorption through direct actions on osteoclasts [49].
Oxidative stress disturbs the balance of bone metabolism, determining the apoptosis of osteocytes and osteoblasts and favoring the cell proliferation of osteoclasts, which causes bone destruction [50]. At the bone tissue level, oxidative stress favors postmenopausal, diabetic, and glucocorticoid osteoporosis. Under the action of oxidative stress at the intracellular level, the mitochondria are deformed, which leads to the disruption of cellular metabolism and even to apoptosis [51].
Prostaglandin E2 (PGE2) is produced by both osteoblasts and osteoclasts in the initial phases of the bone healing process through a reaction catalyzed by the enzyme cyclooxygenase (COX). At the bone fracture, it promotes angiogenesis, increases the number of osteoclasts, which promotes osteolysis and increases the differentiation of osteoblasts, thus promoting osteogenesis. Both traditional non-steroidal anti-inflammatory drugs and selective COX2 inhibitors inhibit bone formation due to their effect on prostaglandins [52].
Proinflammatory cytokines regulate the inflammatory process on bone, regulating both formation and bone resorption, thus altering bone homeostasis. Proinflammatory cytokines (TNFα, IL-1, and IL-17) cause osteoclast activation, thus explaining increased bone loss during inflammation. Other cytokines such as: IL-12, IL-18, IL-33 and IFN are suppressors of osteoclast differentiation and thus inhibit bone loss. The presence of certain cytokines in bone tissue can influence osteolysis [53].
Bone regeneration involves a large number of small molecules, transmission and signaling pathways, growth factors, and physicochemical stimuli from the extracellular matrix, often interconnected and overlapping [54]. The main signaling pathways with a role in the bone regeneration process are: Wnt, TGF-b, MAPK, JNK and the Notch pathway. Small molecules act as activators or inhibitors of transcription factors and through this they can regulate the process of bone formation [55].
Several plant-derived components obtained from plant extracts proved their ability to affect the proliferation and differentiation potential of MSCs having as target various signal transduction pathways showing an osteopromotive role in bone regeneration [48,56,57,58]. The mechanism of action of some bioactive compounds are highlighted in Table 1.
The conventional administration of treatments in bone diseases (oral, systemic administration) presents multiple disadvantages such as: the low bioavailability of the administered medicinal substances or of the administered phytochemical compounds, the occurrence of side effects at the gastric and intestinal level, low absorption, the need to increase the dose of administration [109,119,120].
Taking into account these inconveniences, traditional medicine is considered to be outdated in the case of bone diseases, therefore the attention is recently directed to the targeted administration of natural compounds in nanometric form [119].
The advantages of using nanomedicine are multiple such as: the incorporation of medicinal substances, phytochemicals or their mixture in different materials (biomaterials) that are compatible with bone tissue [119,120,121]. Also, having an increased compatibility with the human body, the bioavailability of the substances administered by this route is increased, thus managing to decrease the doses administered [122,123,124].
The administration can be sustained at the level of the therapeutic plateau and thus the number of administrations per day can decrease, thus gaining increased compliance with the applied treatment [122]. The phytochemical compounds included in such systems are also protected from factors that destroy their therapeutic activity such as light, temperature and pH [121,122,123].
The nanometric systems also allow the controlled and targeted release of the contained compounds, the effectiveness of the treatment being much higher than conventional treatments [124,125,126]. Also, these systems allow local administration, at the bone level, which allows avoiding gastric and intestinal adverse effects encountered with conventional medication [127,128,129,130,131].
Some of these novel transport systems are presented in Figure 2.
New treatment opportunities and solutions are envisioned by combining herbal medicines having osteogenic, antitumor, antimicrobial, and anti-inflammatory properties with advanced materials for bone tissue engineering. Plant derived compounds have been included or added in the composition of orthopedic biomaterials (metallic, ceramic and polymeric matrix) in order to deliver the phytochemical substance at bone site leading to functionalized scaffolds with plant extracts applied in tissue engineering. A wide range of natural compounds have been proposed for incorporation within bone tissue engineering scaffolds in order to enhance bone growth, to inhibit osteoclastic bone resorption, and to prevent other bone-related complications [132,133]. Lately, an innovative concept has emerged from nanotechnology-tissue engineering combination and nanostructured scaffolds attracted a huge interest due to their promising results in improving the bone healing process.
Essential oils (from different plant sources) and other compounds (metallic nanoparticles, zinc nitrate, copper sulphate, cobalt nitrate, etc.) were used to treat polyurethane scaffolds leading to improved physical and biomedical properties of the designed scaffolds (stability, biocompatibility, bone mineralization, osteoblast cell adhesion, antimicrobial activity etc.) [46]. Polyphenols, another class of natural compounds, were incorporated in bioactive scaffolds composition conferring unique structural and functional features: bio adhesion, antioxidation, anti-inflammatory and antibacterial properties, hydrophilicity, self-healing, and biocompatibility promoting bone regeneration. The approaches addressed for polyphenol-based scaffolds fabrication included coating onto polymer scaffold, grafting or blending into biopolymers. They presented different morphologies such as hydrogels (3D cross-linked networks), films (2D materials) and nanofibers (1D materials) [134].
Allium cepa extract, chitosan and poly (DL-lactic-co-glycolic) acid have been em-ployed for the synthesis of a 3D matrix with porous morphology (50-100 um) allowing its surface mineralization with a uniform hydroxyapatite layer [135]. Cucurbitacin B was incorporated into a biomaterial scaffold based on poly (lactidecoglycolide) and β-tricalcium phosphate from where it was linearly released showing enhanced neovascularization (via VEGFR-related signaling pathways) and bone regeneration (via higher bone mineral density, bone volume and number of trabeculae). The composite presented a bio-mimic structure with a pore between 16-466 nm and improved mechanical properties [112].
Aloe vera gel was incubated on the surface of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) nanofibers generating a scaffold with promising osteoinductive potential (higher amounts of alkaline phosphatase activity, mineralization, and bone-related gene and protein expression were recorded) [136]. In another study, polycaprolactone/aloe vera/silk fibroin nanofibrous scaffolds were synthesized followed by hydroxyapatite deposition resulting in biomimetic scaffolds with increased cell proliferation, osteogenic markers expression, osteogenic differentiation and mineralization [137].
A lipid-based self-nano emulsifying drug delivery system (100-180 nm) containing Cassia occidentalis L. butanolic extract prevented the downregulation of miR29a (Runx2), miR17 and miR20a (RANKL) induced by methyl prednisone proving osteogenic and anti-resorptive mechanisms. The anti-inflammatory effect of the glucocorticoid was not affected, meanwhile the induced sarcopenia and muscle atrophy were counteracted [90]. Linum usitatissimum extract rich in phenolic compounds and flavonoids was incorporated in a nanocomposite hydrogel based on alginate and nano-hydroxyapatite which presented a porous structure (100-200 µm) and sustainedly released the natural compound. This nanocomposite showed hemocompatibility, antioxidant activity and cell proliferation, thus promoting bone regeneration [138].
Fruits (grape seed, pomegranate peel, jabuticaba peel) extracts were used as crosslinkers for anionic collagen which together with nanohydroxyapatite formed a smart scaffold with a pore size (17-230 nm) appropriate for bone growth [139]. Genipin, a natural compound from gardenia fruits, shows a promising cross-linking ability for natural biopolymers (collagen, gelatin, and chitosan) and also biosafety profile. The surface of several scaffolds was coated with genipin-crosslinked hydrogel releasing osteogenic factors, acting as an anti-infection agent or presenting water-absorbing function [140].
Epigallocatechin gallate was employed for coating a poly (L-lactic acid) nanofibers enhancing hydrophilicity and stem cell adhesion, the flavonoid serving as a protective agent of external oxidative stress for stem cells. The plant flavonoid-based scaffold promoted osteogenic differentiation of adipose-derived stem cells and reduced osteo-clastic maturation of murine macrophages [99]. In another study, epigallo-catechin gallate represented a promising tool to chemically modify gelatin sponges influencing its surface properties (hydrophilicity and negative zeta potential). This functionalization enhanced cell adhesion and calcium phosphate precipitation inducing superior bone formation in vivo [100]. In other study, gellan gelum hydrogels loaded with alkaline phosphatase were enriched with 5 types of gallotannins (three tannic acids with differing molecular weight, pentagalloyl glucose, and a gallotannin-rich extract from mango kernel (Mangifera indica L.). Those preparations promoted the mineral formation (dry mass percentage values were increased) due to the interactions between ALP and gallotannins dependent on the medium [102].
Forskolin, a triterpenoid from Coleus forskohlii, was loaded on halloysite nanotubes which then were used as dopant for the modification biopolymer scaffold (based on gelatin, chitosan, agarose) resulting in a new osteoconductive smart polymeric scaffold. The forskolin-loaded halloysite nanotubes acted as a filler increasing the compo-site volume and filling the voids, therefore facilitating the cell attachment. The mechanical properties of the scaffold together with the chemical signal of forskolin (cyclic adenosine monophosphate signaling activation in stem cells) had a synergic effect in promoting the osteodifferentiation of mesenchymal stem cells [101].
Plant-derived nanoparticles from potato containing rhamnogalacturonan-I (with relatively higher amount of gallactose) were used for poly(L-lactide-co-ɛ-caprolactone) scaffolds functionalization revealing a downregulation of pro-inflammatory gene markers and promotion of osteogenic markers [77]. The same plant-derived pectin, rhamnogalacturonan-I, was employed for nanocoating of im-plants influencing osteoblast proliferation, mineralization, and gene expression (Runt-related transcription factor 2 (Runx2), alkaline phosphate (ALP), osteocalcin (Bglap), α-1 type I collagen (Col1a1), receptor activator of NF-κB ligand (RANKL)) [78]. A plant-derived phenolic compound, sinapic acid, was used for the synthesize of sinapic acid loaded chitosan nanoparticles (100-115 nm) which furthermore were incorporated into polycaprolactone fibers (around 350 nm) resulting in a scaffold that promoted osteoblast differentiation in vitro and bone formation in vivo (via TGF-β1/BMP/Smads/Runx2 signaling pathways) due to sustained release of sinapic acid [110].
5. Conclusions
Understanding the structure of bone tissue and the mechanisms of bone formation is crucial in the development of new methods of treatment for bone defects. The purpose of this work was to summarize the main cells, biomarkers and signaling pathways involved in bone metabolism. Subsequently, the phytochemical compounds with huge potential in bone regeneration were highlighted according to their activity and mechanisms of action included in three big classes: natural compounds with pro-estrogenic activity, with antioxidant and anti-inflammatory properties and modulatory compounds of bone signaling pathways. Recently, the targeted administration of natural compounds in nanometric forms for bone regeneration is in increased demand due to their considerable advantages. Another trend of nanomedicine applied in orthopedic field consists in the combination of plant derived compounds with nanostructured biomaterials resulting in functionalized scaffolds crucial for bone tissue engineering which shown promising results for the improvement of bone healing process.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, A.H.F., F.M.G. and L.V.G.; methodology, G.A.F., L.F.; software, A.P.; validation, M.E.M., S.C.; formal analysis, E.M.; investigation, A.H.F.; resources, A.H.F., F.M.G.; data curation, L.F.; writing—original draft preparation, A.H.F.; writing—review and editing, L.F.; visualization, T.J., S.F.; supervision, M.E.M. All authors have read and agreed to the published version of the manuscript.”
Funding
“This research was funded by University of Oradea through the grant competition “Scientific Research of Excellence Related to Priority Areas with Capitalization through Technology Transfer: INO—TRANSFER—UO”, Project no. 129/2021
Conflicts of Interest
The authors declare no conflict of interest.
References
- Infante, A.; Rodríguez, C.I. Osteogenesis and Aging: Lessons from Mesenchymal Stem Cells. Stem Cell Res Ther 2018, 9, 244. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, N. Bone Cell Communication Factors Provide a New Therapeutic Strategy for Osteoporosis. Chonnam Med J 2020, 56, 94. [Google Scholar] [CrossRef]
- Hannah, S.S.; McFadden, S.; McNeilly, A.; McClean, C. “Take My Bone Away?” Hypoxia and Bone: A Narrative Review. J Cell Physiol 2021, 236, 721–740. [Google Scholar] [CrossRef]
- Wojda, S.J.; Donahue, S.W. Parathyroid Hormone for Bone Regeneration: PARATHYROID HORMONE FOR BONE REGENERATION. J. Orthop. Res. 2018, 36, 2586–2594. [Google Scholar] [CrossRef] [PubMed]
- Leder, B.Z. Parathyroid Hormone and Parathyroid Hormone-Related Protein Analogs in Osteoporosis Therapy. Curr Osteoporos Rep 2017, 15, 110–119. [Google Scholar] [CrossRef]
- Kužma, M.; Jackuliak, P.; Killinger, Z.; Payer, J. Parathyroid Hormone-Related Changes of Bone Structure. Physiol Res 2021, S3–S11. [Google Scholar] [CrossRef]
- Schwetz, V.; Trummer, C.; Pandis, M.; Grübler, M.; Verheyen, N.; Gaksch, M.; Zittermann, A.; März, W.; Aberer, F.; Lang, A.; et al. Effects of Vitamin D Supplementation on Bone Turnover Markers: A Randomized Controlled Trial. Nutrients 2017, 9, 432. [Google Scholar] [CrossRef]
- Lerchbaum, E.; Trummer, C.; Theiler-Schwetz, V.; Kollmann, M.; Wölfler, M.; Pilz, S.; Obermayer-Pietsch, B. Effects of Vitamin D Supplementation on Bone Turnover and Bone Mineral Density in Healthy Men: A Post-Hoc Analysis of a Randomized Controlled Trial. Nutrients 2019, 11, 731. [Google Scholar] [CrossRef]
- Muresan, G.C.; Hedesiu, M.; Lucaciu, O.; Boca, S.; Petrescu, N. Effect of Vitamin D on Bone Regeneration: A Review. Medicina 2022, 58, 1337. [Google Scholar] [CrossRef] [PubMed]
- Robling, A.G.; Bonewald, L.F. The Osteocyte: New Insights. Annu. Rev. Physiol. 2020, 82, 485–506. [Google Scholar] [CrossRef] [PubMed]
- Pathak, J.L.; Bravenboer, N.; Klein-Nulend, J. The Osteocyte as the New Discovery of Therapeutic Options in Rare Bone Diseases. Front. Endocrinol. 2020, 11, 405. [Google Scholar] [CrossRef] [PubMed]
- Li, M.C.M.; Chow, S.K.H.; Wong, R.M.Y.; Qin, L.; Cheung, W.H. The Role of Osteocytes-Specific Molecular Mechanism in Regulation of Mechanotransduction – A Systematic Review. Journal of Orthopaedic Translation 2021, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular Mechanosensors in Osteocytes. Bone Res 2020, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of Osteoblast Differentiation by Cytokine Networks. IJMS 2021, 22, 2851. [Google Scholar] [CrossRef] [PubMed]
- Al-Bari, A.A.; Al Mamun, A. Current Advances in Regulation of Bone Homeostasis. FASEB BioAdvances 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Ghorbaninejad, M.; Khademi-Shirvan, M.; Hosseini, S.; Baghaban Eslaminejad, M. Epidrugs: Novel Epigenetic Regulators That Open a New Window for Targeting Osteoblast Differentiation. Stem Cell Res Ther 2020, 11, 456. [Google Scholar] [CrossRef] [PubMed]
- Astleford, K.; Campbell, E.; Norton, A.; Mansky, K.C. Epigenetic Regulators Involved in Osteoclast Differentiation. IJMS 2020, 21, 7080. [Google Scholar] [CrossRef]
- Hayashibara, T.; Hiraga, T.; Sugita, A.; Wang, L.; Hata, K.; Ooshima, T.; Yoneda, T. Regulation of Osteoclast Differentiation and Function by Phosphate: Potential Role of Osteoclasts in the Skeletal Abnormalities in Hypophosphatemic Conditions. J Bone Miner Res 2007, 22, 1743–1751. [Google Scholar] [CrossRef]
- Bilezikian, J.P. Hypoparathyroidism. The Journal of Clinical Endocrinology & Metabolism 2020, 105, 1722–1736. [Google Scholar] [CrossRef]
- Lou, Y.-R.; Toh, T.C.; Tee, Y.H.; Yu, H. 25-Hydroxyvitamin D3 Induces Osteogenic Differentiation of Human Mesenchymal Stem Cells. Sci Rep 2017, 7, 42816. [Google Scholar] [CrossRef]
- Marcus and Feldman’s Osteoporosis. Volume 1; Dempster, D.W., Cauley, J.A., Bouxsein, M.L., Cosman, F., Eds.; Fifth edition.; Academic Press: London, 2020; ISBN 978-0-12-813074-2. [Google Scholar]
- Moriishi, T.; Ozasa, R.; Ishimoto, T.; Nakano, T.; Hasegawa, T.; Miyazaki, T.; Liu, W.; Fukuyama, R.; Wang, Y.; Komori, H.; et al. Osteocalcin Is Necessary for the Alignment of Apatite Crystallites, but Not Glucose Metabolism, Testosterone Synthesis, or Muscle Mass. PLoS Genet 2020, 16, e1008586. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Ghosh, A.; Guo, X.; Wang, S.; Hou, Y.; Li, S.; Liu, J. Roles for Osteocalcin in Brain Signalling: Implications in Cognition- and Motor-Related Disorders. Mol Brain 2019, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Hou, W.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med Sci Monit 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling. BioMed Research International 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Baud’huin, M.; Duplomb, L.; Teletchea, S.; Lamoureux, F.; Ruiz-Velasco, C.; Maillasson, M.; Redini, F.; Heymann, M.-F.; Heymann, D. Osteoprotegerin: Multiple Partners for Multiple Functions. Cytokine & Growth Factor Reviews 2013, 24, 401–409. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Wang, S.; Xiao, Z.; Xiong, Y.; Wang, G. Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation. Front. Cell Dev. Biol. 2020, 8, 601224. [Google Scholar] [CrossRef]
- Komori, T. Regulation of Proliferation, Differentiation and Functions of Osteoblasts by Runx2. IJMS 2019, 20, 1694. [Google Scholar] [CrossRef]
- Majidinia, M.; Sadeghpour, A.; Yousefi, B. The Roles of Signaling Pathways in Bone Repair and Regeneration. J Cell Physiol 2018, 233, 2937–2948. [Google Scholar] [CrossRef]
- Karner, C.M.; Long, F. Wnt Signaling and Cellular Metabolism in Osteoblasts. Cell. Mol. Life Sci. 2017, 74, 1649–1657. [Google Scholar] [CrossRef]
- Ballhause, T.M.; Jiang, S.; Baranowsky, A.; Brandt, S.; Mertens, P.R.; Frosch, K.-H.; Yorgan, T.; Keller, J. Relevance of Notch Signaling for Bone Metabolism and Regeneration. IJMS 2021, 22, 1325. [Google Scholar] [CrossRef]
- Cao, X.; Chen, D. The BMP Signaling and in Vivo Bone Formation. Gene 2005, 357, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lowery, J.W.; Rosen, V. The BMP Pathway and Its Inhibitors in the Skeleton. Physiological Reviews 2018, 98, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Zhu, D.; Yang, Q.; Chen, Y.; Zhang, C.; Gao, J.; Gao, Y. Emerging Zero-Dimensional to Four-Dimensional Biomaterials for Bone Regeneration. J Nanobiotechnol 2022, 20, 26. [Google Scholar] [CrossRef] [PubMed]
- Battafarano, G.; Rossi, M.; De Martino, V.; Marampon, F.; Borro, L.; Secinaro, A.; Del Fattore, A. Strategies for Bone Regeneration: From Graft to Tissue Engineering. IJMS 2021, 22, 1128. [Google Scholar] [CrossRef] [PubMed]
- Girón, J.; Kerstner, E.; Medeiros, T.; Oliveira, L.; Machado, G.M.; Malfatti, C.F.; Pranke, P. Biomaterials for Bone Regeneration: An Orthopedic and Dentistry Overview. Braz J Med Biol Res 2021, 54, e11055. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Yu, T.; Hu, B.; Wu, H.; Ouyang, H. Current Biomaterial-Based Bone Tissue Engineering and Translational Medicine. IJMS 2021, 22, 10233. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liang, Z.; Yang, L.; Du, W.; Yu, T.; Tang, H.; Li, C.; Qiu, H. The Role of Natural Polymers in Bone Tissue Engineering. Journal of Controlled Release 2021, 338, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaz, A.; Jayasuriya, A.C. Recent Trends in the Application of Widely Used Natural and Synthetic Polymer Nanocomposites in Bone Tissue Regeneration. Materials Science and Engineering: C 2020, 110, 110698. [Google Scholar] [CrossRef]
- Zhu, Y.; Goh, C.; Shrestha, A. Biomaterial Properties Modulating Bone Regeneration. Macromol. Biosci. 2021, 21, 2000365. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, G.; Johnson, B.N.; Jia, X. Three-Dimensional (3D) Printed Scaffold and Material Selection for Bone Repair. Acta Biomaterialia 2019, 84, 16–33. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, n71. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Moher, D. Updating Guidance for Reporting Systematic Reviews: Development of the PRISMA 2020 Statement. Journal of Clinical Epidemiology 2021, 134, 103–112. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Hao, D.; Zhang, Q.; Chen, B.; Zhang, R.; Wang, Y.; Yang, H. Natural Products for Treatment of Bone Erosive Diseases: The Effects and Mechanisms on Inhibiting Osteoclastogenesis and Bone Resorption. International Immunopharmacology 2016, 36, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Torre, E. Molecular Signaling Mechanisms behind Polyphenol-Induced Bone Anabolism. Phytochem Rev 2017, 16, 1183–1226. [Google Scholar] [CrossRef] [PubMed]
- Chircov, C.; Miclea, I.I.; Grumezescu, V.; Grumezescu, A.M. Essential Oils for Bone Repair and Regeneration—Mechanisms and Applications. Materials 2021, 14, 1867. [Google Scholar] [CrossRef]
- Schilling, T.; Ebert, R.; Raaijmakers, N.; Schütze, N.; Jakob, F. Effects of Phytoestrogens and Other Plant-Derived Compounds on Mesenchymal Stem Cells, Bone Maintenance and Regeneration. The Journal of Steroid Biochemistry and Molecular Biology 2014, 139, 252–261. [Google Scholar] [CrossRef]
- Singh, P.; Gupta, A.; Qayoom, I.; Singh, S.; Kumar, A. Orthobiologics with Phytobioactive Cues: A Paradigm in Bone Regeneration. Biomedicine & Pharmacotherapy 2020, 130, 110754. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the Skeleton. Trends in Endocrinology & Metabolism 2012, 23, 576–581. [Google Scholar] [CrossRef]
- Domazetovic, V. Oxidative Stress in Bone Remodeling: Role of Antioxidants. ccmbm 2017, 14, 209. [Google Scholar] [CrossRef]
- Zhu, C.; Shen, S.; Zhang, S.; Huang, M.; Zhang, L.; Chen, X. Autophagy in Bone Remodeling: A Regulator of Oxidative Stress. Front. Endocrinol. 2022, 13, 898634. [Google Scholar] [CrossRef]
- Vuolteenaho, K.; Moilanen, T.; Moilanen, E. Non-Steroidal Anti-Inflammatory Drugs, Cyclooxygenase-2 and the Bone Healing Process. Basic Clin Pharmacol Toxicol 2007, 0, 071031053307004. [Google Scholar] [CrossRef] [PubMed]
- Schett, G. Effects of Inflammatory and Anti-Inflammatory Cytokines on the Bone: CYTOKINE EFFECTS ON BONE. European Journal of Clinical Investigation 2011, 41, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, U.; Jaffery, H.; Salmeron-Sanchez, M.; Dalby, M.J. An Ossifying Landscape: Materials and Growth Factor Strategies for Osteogenic Signalling and Bone Regeneration. Current Opinion in Biotechnology 2022, 73, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.-Q.; Du, Y.; Yang, P.-S. The Role of Small Molecules in Bone Regeneration. Future Medicinal Chemistry 2013, 5, 1671–1684. [Google Scholar] [CrossRef]
- Maeda, A. Recruitment of Mesenchymal Stem Cells to Damaged Sites by Plant-Derived Components. Front. Cell Dev. Biol. 2020, 8, 437. [Google Scholar] [CrossRef]
- Kornicka, K.; Kocherova, I.; Marycz, K. The Effects of Chosen Plant Extracts and Compounds on Mesenchymal Stem Cells-a Bridge between Molecular Nutrition and Regenerative Medicine- Concise Review. Phytother Res 2017, 31, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Huang, G.; Chen, H.; Xu, L.; Qin, S.; Li, A. Research Progress of the Role of Anthocyanins on Bone Regeneration. Front Pharmacol 2021, 12, 773660. [Google Scholar] [CrossRef]
- Martiniakova, M.; Babikova, M.; Omelka, R. Pharmacological Agents and Natural Compounds: Available Treatments for Osteoporosis. J Physiol Pharmacol 2020, 71. [Google Scholar] [CrossRef]
- Sarkar, N.; Bose, S. Controlled Release of Soy Isoflavones from Multifunctional 3D Printed Bone Tissue Engineering Scaffolds. Acta Biomater 2020, 114, 407–420. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, Y.; Dai, H.; Ma, Y.; Bing, H. Engineered 3D-Printed Polyvinyl Alcohol Scaffolds Incorporating β-Tricalcium Phosphate and Icariin Induce Bone Regeneration in Rat Skull Defect Model. Molecules 2022, 27, 4535. [Google Scholar] [CrossRef]
- Burim, R.A.; Sendyk, D.I.; Hernandes, L.S.; de Souza, D.F.M.; Correa, L.; Deboni, M.C.Z. Repair of Critical Calvarias Defects With Systemic Epimedium Sagittatum Extract. J Craniofac Surg 2016, 27, 799–804. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Jing, J.; Tian, D.; Qian, J.; Yu, G. Dioscin Stimulates Differentiation of Mesenchymal Stem Cells towards Hypertrophic Chondrocytes in Vitro and Endochondral Ossification in Vivo. Am J Transl Res 2016, 8, 3930–3938. [Google Scholar] [PubMed]
- Adhikary, S.; Choudhary, D.; Ahmad, N.; Karvande, A.; Kumar, A.; Banala, V.T.; Mishra, P.R.; Trivedi, R. Dietary Flavonoid Kaempferol Inhibits Glucocorticoid-Induced Bone Loss by Promoting Osteoblast Survival. Nutrition 2018, 53, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, J.; Xu, B.; Yuan, Z.; Leng, Y.; Min, J.; Lan, X.; Luo, J. Kaempferol Promotes Bone Formation in Part via the MTOR Signaling Pathway. Mol Med Rep 2019, 20, 5197–5207. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, C.; Sharma, S.; Bora, P.S.; Verma, S.; Rajput, S.; Porwal, K.; Rath, S.K.; Gayen, J.R.; Sharma, U.; Chattopadhyay, N. A Novel Extraction Method Enhanced the Osteogenic and Anti-Osteoporosis Effect of Tea Extract without Any Hepatotoxicity in Ovariectomized Rats. Frontiers in Endocrinology 2022, 13. [Google Scholar] [CrossRef]
- Wei, Y.; Fu, J.; Wu, W.; Ma, P.; Ren, L.; Yi, Z.; Wu, J. Quercetin Prevents Oxidative Stress-Induced Injury of Periodontal Ligament Cells and Alveolar Bone Loss in Periodontitis. Drug Des Devel Ther 2021, 15, 3509–3522. [Google Scholar] [CrossRef]
- Khedgikar, V.; Kushwaha, P.; Ahmad, N.; Gautam, J.; Kumar, P.; Maurya, R.; Trivedi, R. Ethanolic Extract of Dalbergia Sissoo Promotes Rapid Regeneration of Cortical Bone in Drill-Hole Defect Model of Rat. Biomed Pharmacother 2017, 86, 16–22. [Google Scholar] [CrossRef]
- Gong, W.; Zhang, N.; Cheng, G.; Zhang, Q.; He, Y.; Shen, Y.; Zhang, Q.; Zhu, B.; Zhang, Q.; Qin, L. Rehmannia Glutinosa Libosch Extracts Prevent Bone Loss and Architectural Deterioration and Enhance Osteoblastic Bone Formation by Regulating the IGF-1/PI3K/MTOR Pathway in Streptozotocin-Induced Diabetic Rats. Int J Mol Sci 2019, 20, 3964. [Google Scholar] [CrossRef]
- Tan, B.; Wu, Y.; Wu, Y.; Shi, K.; Han, R.; Li, Y.; Qian, Z.; Liao, J. Curcumin-Microsphere/IR820 Hybrid Bifunctional Hydrogels for In Situ Osteosarcoma Chemo-Co-Thermal Therapy and Bone Reconstruction. ACS Appl Mater Interfaces 2021, 13, 31542–31553. [Google Scholar] [CrossRef]
- Verma, A.H.; Kumar, T.S.S.; Madhumathi, K.; Rubaiya, Y.; Ramalingan, M.; Doble, M. Curcumin Releasing Eggshell Derived Carbonated Apatite Nanocarriers for Combined Anti-Cancer, Anti-Inflammatory and Bone Regenerative Therapy. J Nanosci Nanotechnol 2019, 19, 6872–6880. [Google Scholar] [CrossRef]
- Li, Y.; Dånmark, S.; Edlund, U.; Finne-Wistrand, A.; He, X.; Norgård, M.; Blomén, E.; Hultenby, K.; Andersson, G.; Lindgren, U. Resveratrol-Conjugated Poly-ε-Caprolactone Facilitates in Vitro Mineralization and in Vivo Bone Regeneration. Acta Biomater 2011, 7, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Murgia, D.; Mauceri, R.; Campisi, G.; De Caro, V. Advance on Resveratrol Application in Bone Regeneration: Progress and Perspectives for Use in Oral and Maxillofacial Surgery. Biomolecules 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, G.; Poudel, S.B.; Kook, S.-H.; Lee, J.-C. Resveratrol Prevents Alveolar Bone Loss in an Experimental Rat Model of Periodontitis. Acta Biomater 2016, 29, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Yoon, D.S.; Lee, K.-M.; Choi, S.M.; Lee, M.-H.; Park, K.H.; Han, S.H.; Lee, J.W. Enhancement of Mesenchymal Stem Cell-Driven Bone Regeneration by Resveratrol-Mediated SOX2 Regulation. Aging Dis 2019, 10, 818–833. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Takanche, J.S.; Kim, J.-E.; Jeong, S.-H.; Han, S.-H.; Yi, H.-K. Schisandra Chinensis Extract Ameliorates Age-Related Muscle Wasting and Bone Loss in Ovariectomized Rats. Phytotherapy Research 2019, 33, 1865–1877. [Google Scholar] [CrossRef] [PubMed]
- Suliman, S.; Mieszkowska, A.; Folkert, J.; Rana, N.; Mohamed-Ahmed, S.; Fuoco, T.; Finne-Wistrand, A.; Dirscherl, K.; Jørgensen, B.; Mustafa, K.; et al. Immune-Instructive Copolymer Scaffolds Using Plant-Derived Nanoparticles to Promote Bone Regeneration. Inflamm Regen 2022, 42, 12. [Google Scholar] [CrossRef]
- Folkert, J.; Meresta, A.; Gaber, T.; Miksch, K.; Buttgereit, F.; Detert, J.; Pischon, N.; Gurzawska, K. Nanocoating with Plant-Derived Pectins Activates Osteoblast Response in Vitro. Int J Nanomedicine 2016, 12, 239–249. [Google Scholar] [CrossRef]
- SOARES, I.M.V.; FERNANDES, G.V. de O.; CAVALCANTE, L.C.; LEITE, Y.K.P. de C.; BEZERRA, D. de O.; de CARVALHO, M.A.M.; CARVALHO, C.M.R.S. The Influence of Aloe Vera with Mesenchymal Stem Cells from Dental Pulp on Bone Regeneration: Characterization and Treatment of Non-Critical Defects of the Tibia in Rats. J Appl Oral Sci 2019, 27, e20180103. [Google Scholar] [CrossRef]
- Lin, Z.; Lin, C.; Fu, C.; Lu, H.; Jin, H.; Chen, Q.; Pan, J. The Protective Effect of Ellagic Acid (EA) in Osteoarthritis: An in Vitro and in Vivo Study. Biomed Pharmacother 2020, 125, 109845. [Google Scholar] [CrossRef]
- Fikry, E.M.; Gad, A.M.; Eid, A.H.; Arab, H.H. Caffeic Acid and Ellagic Acid Ameliorate Adjuvant-Induced Arthritis in Rats via Targeting Inflammatory Signals, Chitinase-3-like Protein-1 and Angiogenesis. Biomed Pharmacother 2019, 110, 878–886. [Google Scholar] [CrossRef]
- Ding, L.; Gu, S.; Zhou, B.; Wang, M.; Zhang, Y.; Wu, S.; Zou, H.; Zhao, G.; Gao, Z.; Xu, L. Ginsenoside Compound K Enhances Fracture Healing via Promoting Osteogenesis and Angiogenesis. Front Pharmacol 2022, 13, 855393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, J.; Zhang, W. Berberine for Bone Regeneration: Therapeutic Potential and Molecular Mechanisms. J Ethnopharmacol 2021, 277, 114249. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Liu, X.; Li, J.; Wang, B.; Yin, J.; Zhang, H.; Liu, W. Berberine Encapsulated in Exosomes Derived from Platelet-Rich Plasma Promotes Chondrogenic Differentiation of the Bone Marrow Mesenchymal Stem Cells via the Wnt/β-Catenin Pathway. Biol Pharm Bull 2022, 45, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Shaban, N.Z.; Kenawy, M.Y.; Taha, N.A.; Abd El-Latif, M.M.; Ghareeb, D.A. Cellulose Acetate Nanofibers: Incorporating Hydroxyapatite (HA), HA/Berberine or HA/Moghat Composites, as Scaffolds to Enhance In Vitro Osteoporotic Bone Regeneration. Polymers (Basel) 2021, 13, 4140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, J.; Wu, J.; Xiao, L.; Miao, L.; Qi, X.; Li, Y.; Sun, W. Berberine Promotes Osteogenic Differentiation of Mesenchymal Stem Cells with Therapeutic Potential in Periodontal Regeneration. European Journal of Pharmacology 2019, 851, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Talebi, A.; Hayati Roodbari, N.; Reza Sameni, H.; Zarbakhsh, S. Impact of Coadministration of Apigenin and Bone Marrow Stromal Cells on Damaged Ovaries Due to Chemotherapy in Rat: An Experimental Study. Int J Reprod Biomed 2020, 18, 551–560. [Google Scholar] [CrossRef]
- Mao, Y.-W.; Lin, R.-D.; Hung, H.-C.; Lee, M.-H. Stimulation of Osteogenic Activity in Human Osteoblast Cells by Edible Uraria Crinita. J Agric Food Chem 2014, 62, 5581–5588. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Kumar, P.; Ramakrishna, E.; Kumar, S.; Porwal, K.; Kumar, B.; Arya, K.R.; Maurya, R.; Chattopadhyay, N. Extract and Fraction of Cassia Occidentalis L. (a Synonym of Senna Occidentalis) Have Osteogenic Effect and Prevent Glucocorticoid-Induced Osteopenia. J Ethnopharmacol 2019, 235, 8–18. [Google Scholar] [CrossRef]
- Pal, S.; Mittapelly, N.; Husain, A.; Kushwaha, S.; Chattopadhyay, S.; Kumar, P.; Ramakrishna, E.; Kumar, S.; Maurya, R.; Sanyal, S.; et al. A Butanolic Fraction from the Standardized Stem Extract of Cassia Occidentalis L Delivered by a Self-Emulsifying Drug Delivery System Protects Rats from Glucocorticoid-Induced Osteopenia and Muscle Atrophy. Sci Rep 2020, 10, 195. [Google Scholar] [CrossRef]
- Zhou, R.P.; Lin, S.J.; Wan, W.B.; Zuo, H.L.; Yao, F.F.; Ruan, H.B.; Xu, J.; Song, W.; Zhou, Y.C.; Wen, S.Y.; et al. Chlorogenic Acid Prevents Osteoporosis by Shp2/PI3K/Akt Pathway in Ovariectomized Rats. PLoS One 2016, 11, e0166751. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, X. Mechanism of Chlorogenic Acid Treatment on Femoral Head Necrosis and Its Protection of Osteoblasts. Biomed Rep 2016, 5, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Na, W.; Kang, M.-K.; Park, S.-H.; Kim, D.Y.; Oh, S.Y.; Oh, M.-S.; Park, S.; Kang, I.-J.; Kang, Y.-H. Aesculetin Accelerates Osteoblast Differentiation and Matrix-Vesicle-Mediated Mineralization. Int J Mol Sci 2021, 22, 12391. [Google Scholar] [CrossRef] [PubMed]
- Kurian, I.G.; Dileep, P.; Ipshita, S.; Pradeep, A.R. Comparative Evaluation of Subgingivally-Delivered 1% Metformin and Aloe Vera Gel in the Treatment of Intrabony Defects in Chronic Periodontitis Patients: A Randomized, Controlled Clinical Trial. J Investig Clin Dent 2018, 9, e12324. [Google Scholar] [CrossRef] [PubMed]
- Tanık, A.; Güler Doğru, A.; Akpolat, V.; Acun Kaya, F.; Sarıbaş, E.; Gül, M.; İrtegün Kandemir, S.; Deveci, E. Investigation of the Effect of Combined Use of Alloplastic-Based Tricalcium Phosphate Bone Graft and Antihemorrhagic Plant Extract (ABS) on Bone Regeneration in Surgically Induced Bone Defects in Nondiabetic Rats: An Experimental Animal Study. Turk J Med Sci 2018, 48, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Khedgikar, V.; Kushwaha, P.; Gautam, J.; Verma, A.; Changkija, B.; Kumar, A.; Sharma, S.; Nagar, G.K.; Singh, D.; Trivedi, P.K.; et al. Withaferin A: A Proteasomal Inhibitor Promotes Healing after Injury and Exerts Anabolic Effect on Osteoporotic Bone. Cell Death Dis 2013, 4, e778. [Google Scholar] [CrossRef]
- Yan, C.-P.; Wang, X.-K.; Jiang, K.; Yin, C.; Xiang, C.; Wang, Y.; Pu, C.; Chen, L.; Li, Y.-L. β-Ecdysterone Enhanced Bone Regeneration Through the BMP-2/SMAD/RUNX2/Osterix Signaling Pathway. Frontiers in Cell and Developmental Biology 2022, 10. [Google Scholar] [CrossRef]
- Li, F.; Yang, X.; Yang, Y.; Guo, C.; Zhang, C.; Yang, Z.; Li, P. Antiosteoporotic Activity of Echinacoside in Ovariectomized Rats. Phytomedicine 2013, 20, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Madhurakkat Perikamana, S.K.; Lee, S.M.; Lee, J.; Ahmad, T.; Lee, M.S.; Yang, H.S.; Shin, H. Oxidative Epigallocatechin Gallate Coating on Polymeric Substrates for Bone Tissue Regeneration. Macromol Biosci 2019, 19, e1800392. [Google Scholar] [CrossRef]
- Sasayama, S.; Hara, T.; Tanaka, T.; Honda, Y.; Baba, S. Osteogenesis of Multipotent Progenitor Cells Using the Epigallocatechin Gallate-Modified Gelatin Sponge Scaffold in the Rat Congenital Cleft-Jaw Model. Int J Mol Sci 2018, 19, 3803. [Google Scholar] [CrossRef]
- Naumenko, E.; Guryanov, I.; Zakirova, E.; Fakhrullin, R. Forskolin-Loaded Halloysite Nanotubes as Osteoconductive Additive for the Biopolymer Tissue Engineering Scaffolds. Polymers 2021, 13, 3949. [Google Scholar] [CrossRef]
- Douglas, T.E.L.; Keppler, J.K.; Vandrovcová, M.; Plencner, M.; Beranová, J.; Feuereisen, M.; Parakhonskiy, B.V.; Svenskaya, Y.; Atkin, V.; Ivanova, A.; et al. Enhancement of Biomimetic Enzymatic Mineralization of Gellan Gum Polysaccharide Hydrogels by Plant-Derived Gallotannins. International Journal of Molecular Sciences 2020, 21, 2315. [Google Scholar] [CrossRef] [PubMed]
- Porwal, K.; Pal, S.; Dev, K.; China, S.P.; Kumar, Y.; Singh, C.; Barbhuyan, T.; Sinha, N.; Sanyal, S.; Trivedi, A.K.; et al. Guava Fruit Extract and Its Triterpene Constituents Have Osteoanabolic Effect: Stimulation of Osteoblast Differentiation by Activation of Mitochondrial Respiration via the Wnt/β-Catenin Signaling. J Nutr Biochem 2017, 44, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chen, L.; Chen, Y.; Zhang, Z.; Wang, X.; Zhou, B. Cyanidin-3-Glucoside Regulates Osteoblast Differentiation via the ERK1/2 Signaling Pathway. ACS Omega 2021, 6, 4759–4766. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Boonanantanasarn, K.; Kang, M.; Kim, I.; Woo, K.M.; Ryoo, H.-M.; Baek, J.-H. Morinda Citrifolia Leaf Extract Enhances Osteogenic Differentiation Through Activation of Wnt/β-Catenin Signaling. J Med Food 2018, 21, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Boonanantanasarn, K.; Janebodin, K.; Suppakpatana, P.; Arayapisit, T.; Rodsutthi, J.; Chunhabundit, P.; Boonanuntanasarn, S.; Sripairojthikoon, W. Morinda Citrifolia Leaves Enhance Osteogenic Differentiation and Mineralization of Human Periodontal Ligament Cells. Dent Mater J 2014, 33, 157–165. [Google Scholar] [CrossRef]
- Shalan, N.A.A.M.; Mustapha, N.M.; Mohamed, S. Noni Leaf and Black Tea Enhance Bone Regeneration in Estrogen-Deficient Rats. Nutrition 2017, 33, 42–51. [Google Scholar] [CrossRef]
- Kalalinia, F.; Ghasim, H.; Amel Farzad, S.; Pishavar, E.; Ramezani, M.; Hashemi, M. Comparison of the Effect of Crocin and Crocetin, Two Major Compounds Extracted from Saffron, on Osteogenic Differentiation of Mesenchymal Stem Cells. Life Sci 2018, 208, 262–267. [Google Scholar] [CrossRef]
- Kim, S.S.; Kim, N.K.; Seo, S.R. Cynanchi Atrati and Its Phenolic Constituent Sinapic Acid Target Regulator of Calcineurin 1 (RCAN1) to Control Skin Inflammation. Antioxidants (Basel) 2022, 11, 205. [Google Scholar] [CrossRef]
- Balagangadharan, K.; Trivedi, R.; Vairamani, M.; Selvamurugan, N. Sinapic Acid-Loaded Chitosan Nanoparticles in Polycaprolactone Electrospun Fibers for Bone Regeneration in Vitro and in Vivo. Carbohydr Polym 2019, 216, 1–16. [Google Scholar] [CrossRef]
- Abiramasundari, G.; Gowda, C.M.M.; Pampapathi, G.; Praveen, S.; Shivamurugan, S.; Vijaykumar, M.; Devi, A.; Sreepriya, M. Ethnomedicine Based Evaluation of Osteoprotective Properties of Tinospora Cordifolia on in Vitro and in Vivo Model Systems. Biomed Pharmacother 2017, 87, 342–354. [Google Scholar] [CrossRef]
- Cheng, W.-X.; Liu, Y.-Z.; Meng, X.-B.; Zheng, Z.-T.; Li, L.-L.; Ke, L.-Q.; Li, L.; Huang, C.-S.; Zhu, G.-Y.; Pan, H.-D.; et al. PLGA/β-TCP Composite Scaffold Incorporating Cucurbitacin B Promotes Bone Regeneration by Inducing Angiogenesis. Journal of Orthopaedic Translation 2021, 31, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Jiang, Z.; Yang, R.; Ye, Y.; Pei, L.; Xiong, S.; Wang, S.; Wang, L.; Liu, S. Polysaccharide-Rich Extract from Polygonatum Sibiricum Protects Hematopoiesis in Bone Marrow Suppressed by Triple Negative Breast Cancer. Biomed Pharmacother 2021, 137, 111338. [Google Scholar] [CrossRef] [PubMed]
- Wardhana, A.S.; Nirwana, I.; Budi, H.S.; Surboyo, M.D.C. Role of Hydroxyapatite and Ellagic Acid in the Osteogenesis. Eur J Dent 2021, 15, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xu, J.; Wang, T.; Liu, W.; Wei, H.; Yang, X.; Yan, W.; Zhou, W.; Xiao, J. Ellagic Acid and Sennoside B Inhibit Osteosarcoma Cell Migration, Invasion and Growth by Repressing the Expression of c-Jun. Oncol Lett 2018, 16, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Nirwana, I.; Munadziroh, E.; Yuliati, A.; Fadhila, A.I.; Nurliana; Wardhana, A. S.; Shariff, K.A.; Surboyo, M.D.C. Ellagic Acid and Hydroxyapatite Promote Angiogenesis Marker in Bone Defect. J Oral Biol Craniofac Res 2022, 12, 116–120. [Google Scholar] [CrossRef]
- Primasari, D.N.; Nirwana, I.; Budi, H.S.; Wardhana, A.S.; Sari, A.F.; Novita, N.; Setyawan, A.P.; Surboyo, M.D.C.; Shariff, K.A. The Cytokine and Bone Protein Expression by Ellagic Acid-Hydroxyapatite in Bone Remodelling Model. ScientificWorldJournal 2022, 2022, 6740853. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhong, Z.; Jiang, F.; Yang, J.; Nie, H. Juglans Regia L. Extract Promotes Osteogenesis of Human Bone Marrow Mesenchymal Stem Cells through BMP2/Smad/Runx2 and Wnt/β-Catenin Pathways. J Orthop Surg Res 2022, 17, 88. [Google Scholar] [CrossRef]
- Vandenbroucke, A.; Luyten, F.P.; Flamaing, J.; Gielen, E. Pharmacological Treatment of Osteoporosis in the Oldest Old. Clin Interv Aging 2017, 12, 1065–1077. [Google Scholar] [CrossRef]
- Brown, J.P. Long-Term Treatment of Postmenopausal Osteoporosis. Endocrinol Metab (Seoul) 2021, 36, 544–552. [Google Scholar] [CrossRef]
- Du, Z.; Wang, C.; Zhang, R.; Wang, X.; Li, X. Applications of Graphene and Its Derivatives in Bone Repair: Advantages for Promoting Bone Formation and Providing Real-Time Detection, Challenges and Future Prospects. Int J Nanomedicine 2020, 15, 7523–7551. [Google Scholar] [CrossRef]
- Masne, N.; Ambade, R.; Bhugaonkar, K. Use of Nanocomposites in Bone Regeneration. Cureus 14, e31346. [CrossRef]
- Qaseem, A.; Forciea, M.A.; McLean, R.M.; Denberg, T.D.; Clinical Guidelines Committee of the American College of Physicians; Barry, M. J.; Cooke, M.; Fitterman, N.; Harris, R.P.; Humphrey, L.L.; et al. Treatment of Low Bone Density or Osteoporosis to Prevent Fractures in Men and Women: A Clinical Practice Guideline Update From the American College of Physicians. Ann Intern Med 2017, 166, 818–839. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Yang, H.; Zhang, Q.; Liu, C.; Zhao, J.; Zhang, L.; Chen, B. Natural Products for Treatment of Osteoporosis: The Effects and Mechanisms on Promoting Osteoblast-Mediated Bone Formation. Life Sci 2016, 147, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Raut, N.; Wicks, S.M.; Lawal, T.O.; Mahady, G.B. Epigenetic Regulation of Bone Remodeling by Natural Compounds. Pharmacol Res 2019, 147, 104350. [Google Scholar] [CrossRef] [PubMed]
- Shanmugavadivu, A.; Balagangadharan, K.; Selvamurugan, N. Angiogenic and Osteogenic Effects of Flavonoids in Bone Regeneration. Biotechnol Bioeng 2022, 119, 2313–2330. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Xu, L.; Tang, J.; Wang, Q.; Lim, K.S.; Hooper, G.; Woodfield, T.B.F.; Liu, G.; Tian, K.; Zhang, W.; et al. The Advances in Nanomedicine for Bone and Cartilage Repair. J Nanobiotechnology 2022, 20, 141. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.-Z. Current Nanoparticle-Based Technologies for Osteoarthritis Therapy. Nanomaterials (Basel) 2020, 10, 2368. [Google Scholar] [CrossRef] [PubMed]
- Mu, P.; Feng, J.; Hu, Y.; Xiong, F.; Ma, X.; Tian, L. Botanical Drug Extracts Combined With Biomaterial Carriers for Osteoarthritis Cartilage Degeneration Treatment: A Review of 10 Years of Research. Front Pharmacol 2021, 12, 789311. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, X.; Zhao, Z.; Qian, J.; Wang, Y.; Cui, J.; Weng, W.; Cao, L.; Chen, X.; Hu, Y.; et al. Reversal of Osteoporotic Activity by Endothelial Cell-Secreted Bone Targeting and Biocompatible Exosomes. Nano Lett 2019, 19, 3040–3048. [Google Scholar] [CrossRef]
- Paesa, M.; Alejo, T.; Garcia-Alvarez, F.; Arruebo, M.; Mendoza, G. New Insights in Osteoarthritis Diagnosis and Treatment: Nano-Strategies for an Improved Disease Management. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2023, 15, e1844. [Google Scholar] [CrossRef]
- Gao, Z.-R.; Feng, Y.-Z.; Zhao, Y.-Q.; Zhao, J.; Zhou, Y.-H.; Ye, Q.; Chen, Y.; Tan, L.; Zhang, S.-H.; Feng, Y.; et al. Traditional Chinese Medicine Promotes Bone Regeneration in Bone Tissue Engineering. Chin Med 2022, 17, 86. [Google Scholar] [CrossRef]
- Bose, S.; Sarkar, N.; Banerjee, D. Natural Medicine Delivery from Biomedical Devices to Treat Bone Disorders: A Review. Acta Biomater 2021, 126, 63–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Z.; Yang, P.; Duan, G.; Liu, X.; Gu, Z.; Li, Y. Polyphenol Scaffolds in Tissue Engineering. Mater. Horiz. 2021, 8, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Monárrez-Cordero, B.E.; Rodríguez-González, C.A.; Valencia-Gómez, L.E.; Hernández-Paz, J.F.; Martel-Estrada, S.A.; Camacho-Montes, H.; Olivas-Armendáriz, I. The Effect of Allium Cepa Extract on the Chitosan/PLGA Scaffolds Bioactivity. Journal of Applied Biomaterials & Functional Materials 2021, 19, 2280800021989701. [Google Scholar] [CrossRef]
- Tahmasebi, A.; Shapouri Moghadam, A.; Enderami, S.E.; Islami, M.; Kaabi, M.; Saburi, E.; Daei Farshchi, A.; Soleimanifar, F.; Mansouri, V. Aloe Vera-Derived Gel-Blended PHBV Nanofibrous Scaffold for Bone Tissue Engineering. ASAIO J 2020, 66, 966–973. [Google Scholar] [CrossRef]
- Shanmugavel, S.; Reddy, V.J.; Ramakrishna, S.; Lakshmi, B.; Dev, V.G. Precipitation of Hydroxyapatite on Electrospun Polycaprolactone/Aloe Vera/Silk Fibroin Nanofibrous Scaffolds for Bone Tissue Engineering. J Biomater Appl 2014, 29, 46–58. [Google Scholar] [CrossRef]
- Mohammadpour, M.; Samadian, H.; Moradi, N.; Izadi, Z.; Eftekhari, M.; Hamidi, M.; Shavandi, A.; Quéro, A.; Petit, E.; Delattre, C.; et al. Fabrication and Characterization of Nanocomposite Hydrogel Based on Alginate/Nano-Hydroxyapatite Loaded with Linum Usitatissimum Extract as a Bone Tissue Engineering Scaffold. Mar Drugs 2021, 20, 20. [Google Scholar] [CrossRef]
- Garcia, C.F.; Marangon, C.A.; Massimino, L.C.; Klingbeil, M.F.G.; Martins, V.C.A.; Plepis, A.M. de G. Development of Collagen/Nanohydroxyapatite Scaffolds Containing Plant Extract Intended for Bone Regeneration. Materials Science and Engineering: C 2021, 123, 111955. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, H.; Luo, W.; Cai, T.; Li, Z.; Liu, Y.; Gao, W.; Wan, Q.; Wang, X.; Wang, J.; et al. Regeneration of Skeletal System with Genipin Crosslinked Biomaterials. J Tissue Eng 2020, 11, 2041731420974861. [Google Scholar] [CrossRef]
Figure 1.
Prisma flow-diagram for description of the selection process of the bibliographic sources.
Figure 1.
Prisma flow-diagram for description of the selection process of the bibliographic sources.
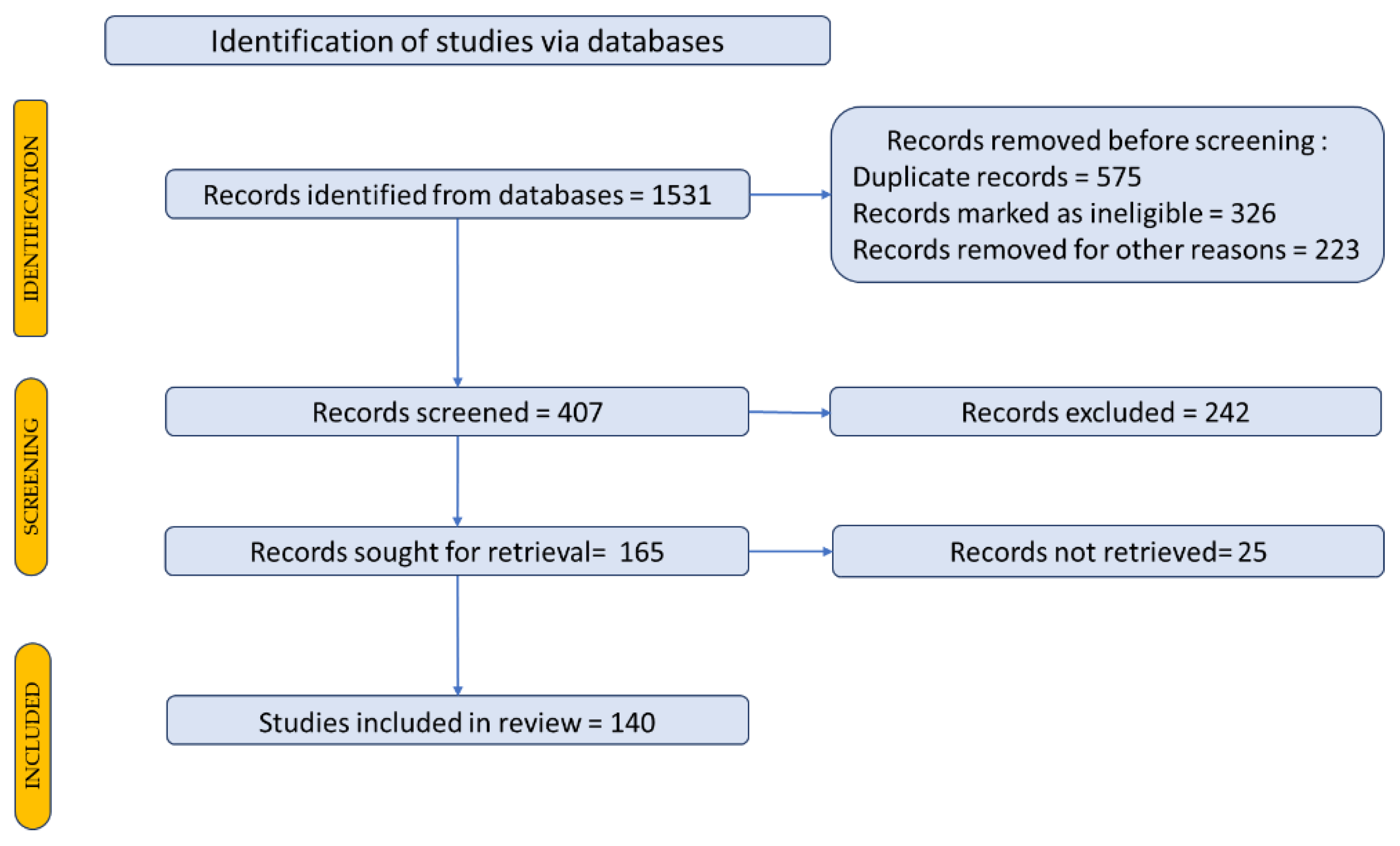
Figure 1.
The main classes of phytocompounds and the main regulatory mechanisms involved in the bone regeneration process, the stages of bone density restoration in different pathologies.
Figure 1.
The main classes of phytocompounds and the main regulatory mechanisms involved in the bone regeneration process, the stages of bone density restoration in different pathologies.
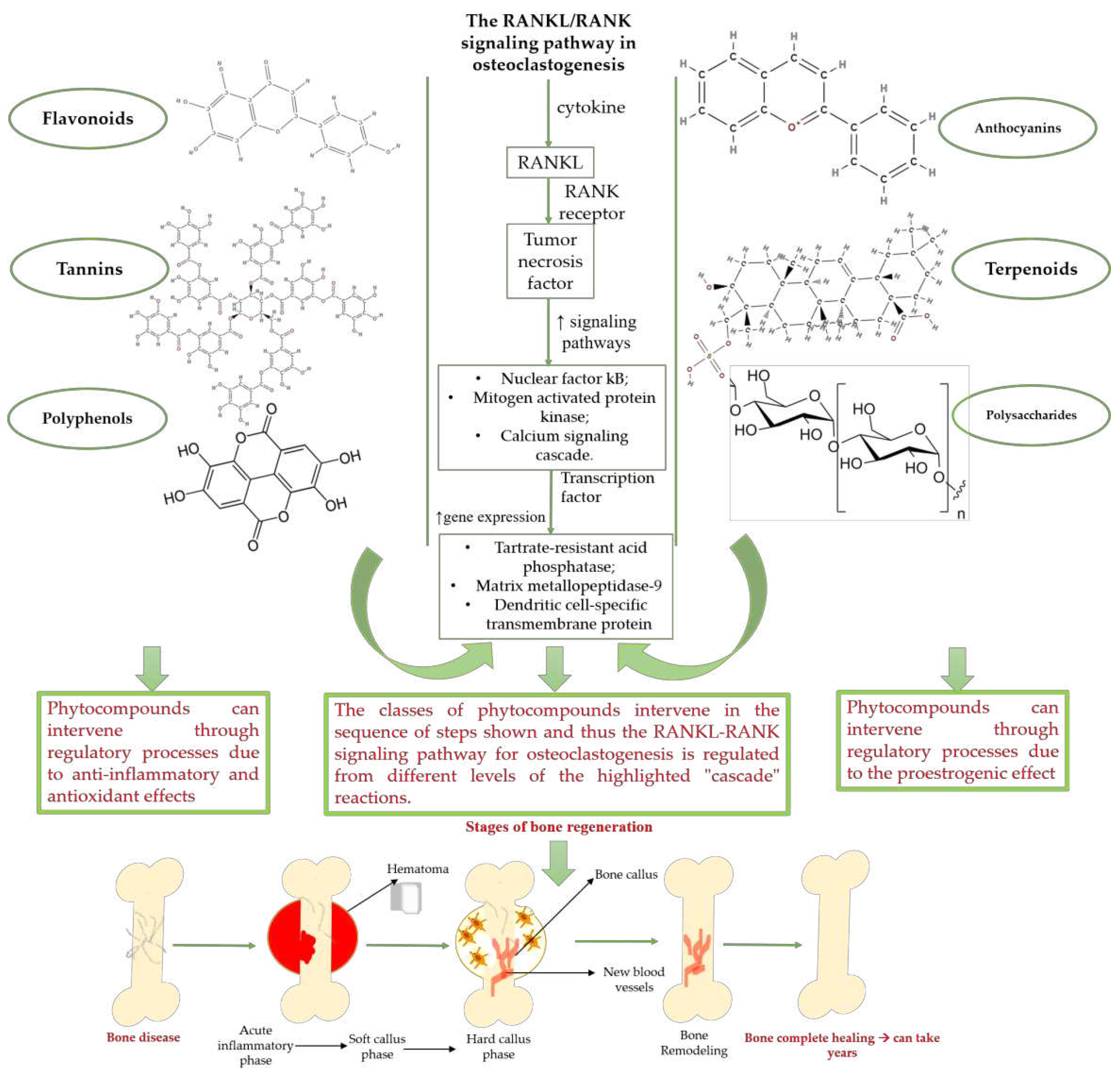
Figure 2.
The diversity of nanometric systems used for the innovative application of phytocompounds for the purpose of bone regeneration.
Figure 2.
The diversity of nanometric systems used for the innovative application of phytocompounds for the purpose of bone regeneration.
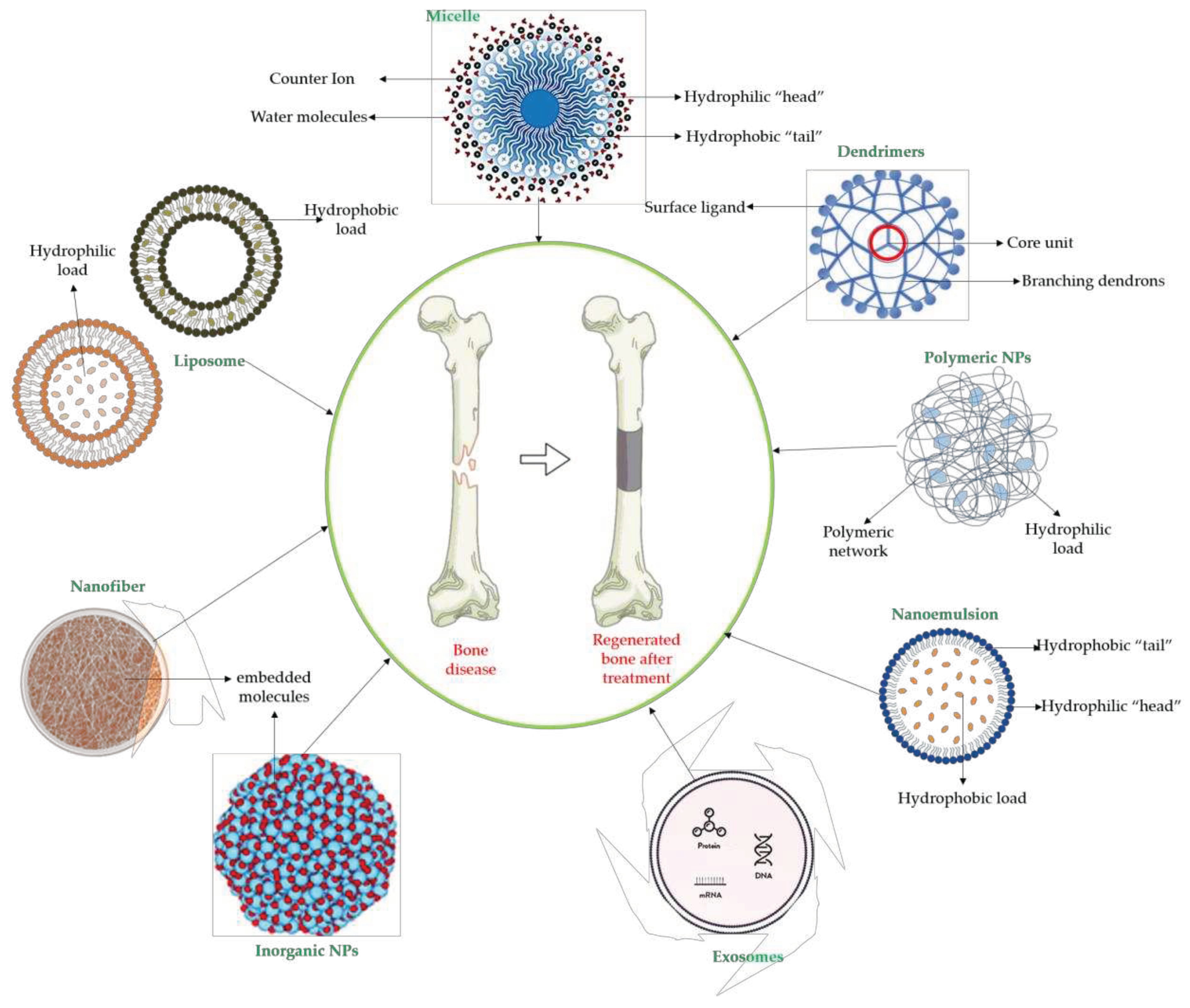

Table 1.
Phytochemical compounds involved in bone regeneration process including their mechanism of action and natural sources.
Table 1.
Phytochemical compounds involved in bone regeneration process including their mechanism of action and natural sources.
| Compounds | Type of activity |
Mechanism of action |
Extract source |
Ref. |
|---|---|---|---|---|
| Genistein | Proestrogenic activity |
↑ alkaline phosphatase level ↓ urinary excretion of calcium and phosphate, → serum concentration at the appropriate normal level |
Erythrina variegate | [59] |
| Daidzein | ↑ osteoclast apoptosis through the mediation of estrogen receptors ↓ the loss of bone density activates tyrosine phosphatase → ↓ membrane depolarization producing changes in intracellular Ca2+ |
[60] | ||
| Icariin | ↓ bone loss in the median bone area by regulating the ratio between osteoprotegerin and RANKL, which are key mediators of osteoclast genesis. ↑ proliferation, differentiation of osteoblasts, bone mineralization ↓ cell apoptosis direct osteoblast stimulation: activation of the bone morphogenetic protein (BMP) cascade through (promoting Runx2/Cbfa1 expression and the production of BMP-4, BMP-2, and SMAD4 and nitrous oxide release; high levels of ALP suppression of p38 and JNK pathways in the osteoclasts, ↓ release of prostaglandin E2 by osteoblasts => inhibition of osteoclast differentiation |
Epimedium | [61] [62] |
|
| Dioscin | ↑the proliferation of bone tissue ↓ cell apoptosis by mediating signaling pathways ↓RANKL expression ↓osteoprotegerin /RANKL → inhibits bone reabsorption |
Dioscoreaceae family | [63] | |
| Kaempferol | ↑ osteoprotegerin and ↓ RANKL expression → osteoclastogenesis decreases ↑ antiapoptotic expression maintaining bone mass, microarchitecture, and bone strength of the trabecular bones |
Ginkgo biloba Camellia sinensis |
[64,65] [66] |
|
| Quercetin | ↑ the proliferation of bone tissue ↓osteoprotegerin /RANKL → inhibits bone reabsorption |
[59,67] | ||
| Ginkgolic acid | ↑ proliferation, differentiation of osteoblasts, bone mineralization | Ginkgo biloba | [59] | |
| caviunin 7-O-[A-D-apiofuranosyl-(1Y6)-A-D-glucopyranoside | stimulates BMP-2/Wnt-βcatenin pathway | Dalbergia sissoo | [68] | |
| Acteoside | Antioxidant and anti-inflammatory effect | ↓ the level of pro-inflammatory cytokines such as TNF-α and IL-6, ↓ the differentiation of osteoclasts by reducing free radicals and fighting oxidative stress ↑ cell proliferation ↓ bone demineralization |
Verbascum sp. Cistanche sp. |
[69] |
|
Curcumin |
↓the level of inflammation by decreasing the inflammatory cytokines TNF-a and IL-6 ↓bone loss and demineralization, inhibiting osteoclastogenesis ↑the level of alkaline phosphatase, which leads to an increase in the mineralization process interaction with transcription and growth factors, protein kinases, cytokines and enzymes => apoptosis of cancer cell |
Curcuma longa | [70] [71] |
|
| Resveratrol | ↓the level of free radicals from the bone level, neutralizing them ↓bone loss inhibits osteoclastogenesis and the RANKL marker influences the response of estrogen receptors to oxidative stress factors ↑bone differentiation → ↑ bone density ↑the level of morphogenetic protein at the bone level ↓decreases the level of alkaline phosphatase ↓the level of osteocalcin. allows mass production of MSCs; mRNA levels of RUNX2, Collagen Type I Alpha 1 (COL1A1), PPARγ, Adiponectin (APN) were highly expressed, ↑ SIRT1 and SOX2 levels |
- | [72,73,74] [75] |
|
| Gomisin A, Schisandrin C | down-regulation of inflammatory molecules, ROS, and up-regulation of antioxidant molecules | Schisandra chinensis | [76] | |
| rhamnogalacturonan-I | ↓ intracellular accumulation of galectin-3 down-regulation of RANKL, TNFα, IL-6, and IL-1β |
Solanum tuberosum | [77,78] | |
| Acemannan | tissue regeneration, cell proliferation, extracellular matrix synthesis, mineralization. ↑ expression of growth factors; stimulation of bone cementum and periodontal ligament regeneration; induction of bone formation, osteoblast proliferation and differentiation |
Aloe vera | [79] | |
|
Ellagic acid Caffeic acid and Ellagic acid |
- inhibition of iNOS, COX-2, NO, TNF-α, PGE2 and IL-6 - down-regulation of IL-1β-stimulated matrix metalloproteinase-13 and thrombospondin motifs 5 - up-regulation of collagen of type II and aggrecan - suppression of NF-κB signaling - ↓ chitinase-3-like protein-1, IL-1β, NF-κB, caspase-3; lipid peroxides, NO - ↑ reduced glutathione |
[80] [81] |
||
| Ginsenoside | Modulatory compounds of bone regeneration pathways | ↑ calcium absorption at the intestinal level → thus prevents bone loss ↑ the level of trabecular calcium ↓ C-terminal telopeptide of type I collagen → ↓ resistance to tartrate acid phosphatase at the femoral level |
Orchidaceae family | [59,82] |
| Berberine | ↓ bone loss by preventing decalcification and demineralization inhibits osteoclastogenesis suppresses the activity of the markers involved in the differentiation of acid phosphatase-resistant tartrate bone cells and cathepsin K ↓ the differentiation rate of osteoclasts restore downregulation of osteogenesis-related genes expression; ↑ expression of osteogenesis-related genes such as OSX, COLⅠ, ALP, OCN and OPN ↑ total β-catenin and nuclear β-catenin; activation of the Wnt/β-catenin signaling pathway |
Coptis species. Berberis species. Coptidis Rhizoma, Coptis chinensis, Coptis teeta. |
[83,84,85] [86] |
|
| Apigenin | ↑ the proliferation capacity of osteoblasts inhibits decalcification and osteoclastogenesis modulates intracellular signals → ↓bone loss induced by estrogen hormones ↓ the level of bone inflammation. ↑ mRNA levels of osteogenic genes BMP-2, Runx2 and COL1 downregulation of miR29a, miR17 and miR20a |
Olea europaea. Cassia occidentalis |
[87,88] [89,90] |
|
| Chlorogenic acid | ↑ the level of favorable markers for bone formation ↑ the level of bone morphogenetic protein →↑ the activity of osteoblasts ↓ the level of pro-inflammatory factors ↑ the level of glutathione peroxidase →strong antioxidant effect ↑ the serum activity of alkaline phosphatase, osteoprotegerin ↓ the production of RANKL decreases |
Prunus domestica L. | [91,92] | |
| Aesculetin | ↑ expression of bone morphogenetic protein-2, collagen type 1, osteoprotegerin; ALP activation; transcription of Runt-related transcription factor 2; induction of: non-collagenous proteins of bone sialoprotein II, osteopontin, osteocalcin, and osteonectin, of annexin V and PHOSPHO 1. ↑ the production of thrombospondin-1 and tenascin C |
- | [93] | |
| Acemannan | ↑ mRNA expression of bone morphogenetic protein 2 ↑ mineral deposition |
Aloe vera | [94] | |
| Antihemorrhagic plant extract | ↑ osteoblastic activity and new bone formation; ↑ osteonectin and osteopontin expression ↓ inflammatory cell infiltration, vascular dilatation and hemorrhage |
Glycyrrhiza glabra, Vitis vinifera, Alpinia officinarum Urtica dioica, Thymus vulgaris | [95] | |
| withaferin A | ↑ expression of osteoblast-specific transcription factor and mineralizing genes, osteoblast survival, ↓ inflammatory cytokines. |
Withania somnifera | [96] | |
| β-ecdysterone | ↑ gene expression of the BMP-2/Smad/Runx2/ Osterix signaling pathway, stimulates MC3T3-E1 cell proliferation | [97] | ||
| Echinacoside | ↑ the uterine weight and serum E2 levels, ↓ body weight and hydroxyproline serum levels |
Cistanche tubulosa | [98] | |
| Epigallocatechin gallate | activation of β-catenin of the Wnt signaling pathway ↑ expression of osteogenic genes, ALP activity, and mineralization in bone marrow-derived mesenchymal stem cells |
Grean tea | [99], [100] |
|
| Essential oils | blocking nuclear factor kappa B, p38, and c-Jun N-terminal kinase signaling ↓ production of nitric oxide in RAW264.7 cells, inhibited EAhy926 cell proliferation ↑ serum C-telopeptide collagen type I and osteocalcin ↑ plasma calcium and vitamin D3, bone mineral-density Prevention of inflammation and oxidative stress |
Hypericum perforatum; Cinnamomum burmanini; Thymus vulgari; Rosmarinus officinalis. Populus alba; |
[46] |
|
| Forskolin | activation of cyclic adenosine monophosphate (c-AMP) signalling in stem cells | Coleus forskohlii | [101] | |
| Gallotannin | interaction with ALP growth of Saos-2 cells |
Mangifera indica L. | [102] | |
|
Triterpenes (ursolic acid, 2α-hydroxyursolic acid) |
↑ trabecular parameters (BV/TV, Tb.Th and conn.D) ↓ SMI ↑ALP activity, osteogenic genes (Runx2, BMP-2, type 1 Col1 and Wnt3a) stimulates Wnt/β-catenin signalling osteoblast differentiation (activation of mitochondrial respiration) |
Psidium guajava | [103] | |
| malvidin, cyanidin and delphinidin | inhibition of MSC adipogenesis and downregulation of FABP4 and adiponectin genes. ↑ accumulation of calcium deposits upregulation of osteocyte-specific gene BMP-2 and Runx-2 expression |
Berries | [104] | |
| rutin and kaempferol-3-O-rutinoside | activation of Wnt/b-Catenin Signaling ↑ activity of ALP, Runx2, osterix, osteocalcin, bone morphogenetic protein 2, Wnt3a, and b-catenin |
Morinda citrifolia (Noni) | [105,106,107] | |
| rhamnogalacturonan-I | ↓ intracellular accumulation of galectin-3 up-regulation of collagen type I alpha 1 (COL-Iα1), osteocalcin, sialoprotein. down-regulation of RANKL, TNFα, IL-6, and IL-1β |
Solanum tuberosum | [77,78] | |
| Crocin, Crocetin | ↑ ALP activity and ALP mRNA expression in MSCs | Crocus sativus L. | [108] | |
| Sinapic acid | activation of TGF-β1/BMP/Smads/Runx2 signaling pathways => osteoblast differentiation | Cynanchi atrati | [109] [110] |
|
| beta ecdysone | ↑ collagen deposition, ↑ levels of osteocalcin, ↑ expression of osteogenic genes | Tinospora cordifolia | [111] | |
| cucurbitacin B | ↑ expression of ALP and OPN genes, mineralization up-regulation of VEGFR2 and VEGFR-related signaling pathways (induction of angiogenesis) |
Cucurbitaceae family plants | [112] | |
| Polysaccharides | hematopoiesis protection: ↓ myeloid cells within tumor-infiltrating immune cells Inhibition of hematopoietic cell expansion in the spleen ↑ HSPCs (hematopoietic stem and progenitor cells) and common lymphoid progenitors in the bone marrow |
Polygonatum sibiricum | [113] | |
|
Ellagic acid Ellagic acid and Sennoside B |
↑ number of osteoblasts and expression of OCN and OPG ↓ number of osteoclasts and the expression of RANKL - repression of c-Jun expression at the mRNA level |
[114] [115] |
||
| Ellagic acid and hydroxyapatite | ↑ in the expression of FGF-2, VEGF and ALP ↑ IL-10, BMP-4 and OPN ↓ TNF-α and increasing the expression of |
[116], [117] |
||
| Melibiose, melezitose, ganolactone B, methylophiopogonanone A, tubuloside A, 2-Acetylacteoside Emodin | ↑ expression of ALP, osteocalcin, osterin, osteoprotegerin, and autophagy marker proteins activation of BMP2/Smad/Runx2 and Wnt/β-catenin signaling |
Juglans regia | [118] |
RANK: receptor activator of nuclear factor kappa B; RANKL: receptor activator of nuclear factor kappa B ligand; BMP: bone morphogenetic protein; ALP: alkaline phosphatase; MSCs: Mesenchymal stem cells; APN: Adiponectin; COL-Iα1: collagen type I alpha 1; FABP4: fatty acid binding protein 4; RUNX2: runx-related transcription factor 2; ROS: reactive oxygen species; IL: interleukin; TGF-β: Transforming growth factor-beta; VEGF: vascular endothelial growth factor; JNK: c-jun-N-terminal kinase; E2: endogenous 17- β-estradiol; Wnt: the Wingless-type MMTV integration site family ; PPARγ2: peroxisome proliferator-activated receptor γ2; SIRT 1: sirtuin 1; OSX: Osterix; OPN: osteopontin; SMAD: Small mother against decapentaplegic; iNOS: inducible nitric oxide synthase, COX-2: cyclooxygenase-2, NO: Nitric oxide, PGE2: prostaglandin E2, NF-κB: nuclear factor kappa B, OCN: osteocalcin protein, OPG: osteoprotegerin , OPN: osteopontin.4. The innovative administration and application of plant extracts in the process of bone regeneration.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



