
Submitted:
18 April 2023
Posted:
19 April 2023
You are already at the latest version
Abstract
Food production heavily depends on ammonia-containing fertilizers to improve crop yield and profitability. However, ammonia production is challenged by huge energy demand, the release of ~2% of global CO2, and political instability. To mitigate this challenge, many research efforts have developed bioprocessing technologies to make biological ammonia. This review presents three different biological approaches that drive the biochemical mechanisms to convert nitrogen gas, bioresources, or waste to biological ammonia. The use of advanced technologies— enzyme immobilization and microbial bioengineering – enhanced bio-ammonia production. This review also highlighted some challenges and research gaps that require researchers’ attention for bio-ammonia to be industrially pragmatic.
Keywords:
ammonia
; bioprocessing
; bioengineering
; fermentation
; enzyme immobilization
1. Introduction
Ammonia has the structural formula: NH3, consisting of the two common elements— nitrogen and hydrogen. Ammonia (NH3) is an odiferous, highly irritating, and colorless gas that is dated as far back as nature. It exists in the soil and the environment as a product of ammonia-producing bacteria and a product of plant, animal, and waste decomposition, respectively. In its pure form, ammonia is anhydrous and hygroscopic as it readily absorbs moisture. Anhydrous ammonia can be a gas or liquid or solid, depending on its temperature and pressure. At room temperature, it is a highly irritating gas with a pungent and suffocating odor. Ammonia in this gaseous form is less dense than air, and its liquid is less dense than water at standard atmospheric temperature and pressure. Ammonia vapor diffuses readily in air, and liquid ammonia is soluble in water with a simultaneous release of heat. When dissolved in water ammonia gas forms ammonium hydroxide (NH4OH), a weak base and caustic solution. Under pressure, ammonia gas is easily compressed and forms a clear liquid. Generally, ammonia is corrosive and has alkaline properties. Several other physicochemical properties of ammonia are published [1,2].
Ammonia was first produced on large scale in 1913 following the evolution of an artificial nitrogen fixation process developed by German chemists Fritz Haber and Carl Bosch in 1909 [3]. This process wherein hydrogen gas reacts with nitrogen gas in the presence of an iron catalyst to produce ammonia [4] is referred to as the Haber–Bosch process (Figure 1). At the inception of the Haber–Bosch process, about 20 tons of ammonia was produced per day. In 2018, about 230 million tons of anhydrous ammonia was produced and it is projected that nearly 290 million metric tons of anhydrous ammonia will be produced in the year 2030 [5].
However, a major challenge with ammonia production is the use of fossil fuels such as natural gas, coal, and oil as feedstock. Concerns about its sustainability and more importantly, the effects of its energy intensity level on climate change and the environment have been on the increase. The Haber–Bosch process is presently one of the most prominent emitters of greenhouse gases accounting for about 1.2% of CO2 emissions produced globally. The entire ecosystem is being impacted by higher emissions of nitrous oxide (N2O) which has now been classified as the third most significant greenhouse gas with Carbon dioxide and methane at the top [6]. As a result of these challenges, scientists have been prompted to make findings on better alternatives [7].
Nonetheless, even with the numerous advantages of Haber–Bosch process-based ammonia, the need for large-scale production of ammonia from other sources is requisite. Research on green ammonia and organic ammonia has recently been on the increase. Green ammonia is a terminology used to describe ammonia produced to accomplish a zero-emission target. It is a product of a renewable and carbon-free process, thus effective in the reduction of greenhouse emissions. However, the level of ammonia production was found to be 50,000 tons/year [8] which is considerably low.
2. Economic Importance of Ammonia
Ammonia is versatile for making fertilizer, refrigerant gas, water purification, and other industrial applications such as the manufacture of explosives, fabrics, pesticides, plastics, and dyes. Domestically, it is used in the production of household cleaning agents.
2.1. Scale of production
Apart from sulfuric acid, ammonia is the highest-volume chemical commodity produced in the world [9]. East Asia, including China, is the largest producer of ammonia; followed by East Europe and Central Asia, North America, South Asia, and the rest of them (Figure 2). In the United States, the third largest ammonia producer, 14 million MT was recorded in 2021. Nutrien, Koch Industries, and CF Industries are by far the largest producer of ammonia in the United States; with CF Industries alone producing over seven million MT of ammonia in 2021 with its top three production facilities in the United States [10].
2.2. Application as fertilizer
Presently, more than 85% of ammonia is used to produce chemical fertilizer for proper plant development and growth [11]. According to the World Fertilizer Trends and Outlook 2020 report of the Food and Agriculture Organization (FAO), United Nations, the volume of ammonia produced globally is approximated at 150 million metric tons with a prediction of about 2.3% increment per year [12]. Ammonium sulfate (NH4)2SO4 is the first nitrogenous fertilizer made by BASF, and it is the leading form in which fixed nitrogen was produced before the second world war. Many forms of ammonia fertilizers are available for different soil types and vegetation. Ammonium nitrate, (NH4)NO3, which contains ~35% nitrogen used to be a leading form of solid ammonia fertilizer compounds. Urea fertilizer, also first manufactured by BASF in 1922, contains more nitrogen (46.6%) and has more advantages over other ammonia compounds-containing fertilizers produced prior, making it a leading solid nitrogen fertilizer. In today’s plant agriculture, however, mixed fertilizers are used. They range from combining two or more macronutrients such as potassium nitrate (~13.8% nitrogen) and ammonium phosphates (~10-21% nitrogen) and other granulated materials usually with selected micronutrients [4].
2.3. Fuel potential
The potential for ammonia to be used directly as fuel to replace fossil fuels in a wide range of applications is also garnering interest. It is anticipated that upon ammonia combustion in engines, the exhaust should be pure nitrogen and water that are safely released into the environment, forming a sustainable and circular cycle that is dubbed the “ammonia economy”. Relative to fossil fuels, ammonia can store chemical energy which is subsequently released by the breaking and making of chemical bonds. The net energy gain is generated from breaking N-H bonds which produces nitrogen and water in the presence of oxygen.
4NH3 + 3O2 ⇌ 2N2 + 6H2O
The energy storage mechanisms of ammonia are fundamentally the same as those of methane, which has four C-H bonds that can be broken down to release energy. But in ammonia where the central atom is nitrogen, nitrogen gas (N2) is produced instead of carbon dioxide (CO2) which results when methane and other C-H-containing gases are burnt. Ammonia can be safely stored in bulk in a liquified form that is achieved by compression to 10 times atmospheric pressure or cooling to -33oC. In its liquified form, ammonia has an energy density of about 3-3.5 kWh/liter which is less than that of ethanol and liquified natural gas at 6 kWh/liter, but comparable to them [13]. The Mid-West of the US has over 10,000 ammonia storage sites, with the highest densities in Iowa with a storage capacity of ~800,000 tons. Transportation is by multiple means including pipelines of around 3,000 miles that connect 11 states, transporting ~2 MT of ammonia per year [14]. Leakage of these pipes could result in serious health and environmental risks due to the corrosive and potential toxicity of ammonia, but it is readily detectable by smell even at very low concentrations that are below levels that could cause any lasting health issues. Nevertheless, stringent controls must be adopted at ammonia storage, transportation, and industrial sites to ensure that the risks of ammonia release are negligible. Apart from its use in nitrogen fertilizer manufacturing and as zero-carbon fuel for many engines, ammonia has also found applications in explosives, textile, and pharmaceutical industries.
3. Ammonia Classification
Ammonia production systems can be classified into three based on the carbon emissions from the production processes, but not the type of ammonia being produced: brown (or grey) ammonia, blue ammonia, and green ammonia.
3.1. Brown (or Grey) ammonia
The Haber-Bosch process is the conventional method for ammonia production. It is responsible for more than 60% of the ammonia produced globally. Due to its high energy requirement and significant contribution to CO2 emissions, the resulting ammonia from the Haber-Bosch process is termed brown ammonia. The Haber-Bosch process is the reaction of nitrogen (N2) and hydrogen (H2) in the presence of an iron catalyst and other oxide promoters such as K2O, Al2O, and CaO. The reaction runs at around 400-600oC for efficient catalysis and up to 200-400 atmospheres of gas pressure to enhance entropy, an energy-hungry reaction that sucks up about 1% of global energy production and is thermodynamically exothermic [15].
N2(g) + 3H2(g) ⇌ 2NH3(g) ∆Ho = -92 kJ
While the nitrogen in the reaction above is extracted from the air, the hydrogen comes from natural gas (methane), oil, or coal through industrial processes that release CO2. Steam methane reforming is the most used method to generate hydrogen, generating CO2 emissions. The current form of the Haber-Bosch process begins by generating hydrogen from fossil-fuel feedstocks, usually coal or oil. A reformer converts the feedstocks into a mixture of gases (syngas) which includes hydrogen. Thereafter a carbon monoxide shift converter mixes water and the carbon monoxide from the preformed syngas to form carbon dioxide (CO2) and more hydrogen. The final steps involve the separation of hydrogen from ammonia synthesis by acid gas removal. At various steps of the process, CO2 is released (Figure 2). For every molecule of natural gas (methane) used, three molecules of CO2 are generated, and 1.6 tons of CO2 is emitted per ton of ammonia produced from the most efficient ammonia production plants [16,17].
CH4 + H2O + 4N2 ↝ 8NH3 + 3CO2
3.2. Blue Ammonia
Efforts by engineers across the world to make ammonia production less energy consuming and sustainable gave rise to the concept of blue ammonia. Blue ammonia, like brown ammonia, is produced from hydrocarbon feedstocks but carbon capture utilization and storage (CCUS) technologies are integrated into ammonia production plants to sequester the resulting CO2. Of all the CCUS technologies known, amine absorption technology is the most used and commercially available [18]. Amines (or alkanolamines) are organic compounds with a basic nitrogen atom. They can be used to separate CO2 from the gas stream during ammonia production through the exothermic reaction of CO2 with an amine. Another CCUS technology is based on the principle that CO2 from any gas mixture (syngas) can be separated by cooling and condensation [19]. The technology, termed cryogenic separation, facilitates the direct production of liquid CO2 which can be transported. Although the amount of energy required for cooling in the process is relatively high and that water must be removed to prevent cooling of the blocks by gas flow, the use of membranes in the gas separation process is promising [20]. Some of the membranes known to decompose CO2 are palladium membranes, polymeric membranes, and zeolites [21].
Another notable CCUS technology uses an adsorption device rotary concentrator on solid. The solid materials used for the adsorption include activated carbons, activated aluminum oxide (Al 2O3), clays, zeolites, and silicon dioxide (SiO2). A modified version of this technology is the pressurized swing adsorption (PSA) in which the gas mixture flows in the direction of the packed bed of the adsorbent at high pressure until the concentration of the desired gas to be separated reaches equilibrium [22]. The captured CO2 can be stored by several methods and used for a variety of production processes including increased oil recovery, coal bed methane extraction, and deep ocean injection, among others [17,23]. In the long run, CCUS blue ammonia production technology will not be beneficial as high energy is still being used to drive the process and the lack of CCUS infrastructure as well as transportation of CO2 poses yet another challenge.
3.3. Green Ammonia
The ammonia production process targeted at reducing or completely removing carbon dioxide emissions birthed the concept of green ammonia. To achieve zero-carbon emissions during ammonia production, renewable feedstocks coupled with reduced energy usage are harnessed. At present, the most desirable but expensive green ammonia production method generates hydrogen from water electrolysis powered by solar, wind, hydro, and or geothermal energy [24]. This approach is also known as electrochemical ammonia synthesis (EAS). Electrolytes used for the EAS are diverse; they include solid electrolytes such as polymer electrolyte membrane (PEM) and anion exchange membrane (AEM), chlorine salts, melt hydroxides, and acidic electrolytes in liquid form [25]. Electrolysis in the latter is mostly by deposition of ammonium salts in solution to cause rapid changes in the pH of the solution. The low solubility of nitrogen often hinders electrolysis in their solutions. Thus, gas diffusion in electrodes is required for high efficiency and production rates [17].
In the EAS method, electrocatalysts commonly used based on the physical state and pH of the electrolyte include precious metals, metal nitrides, and metal oxides [26]. Transition metal-free catalysts such as black phosphorus and nitrogen-doped carbons are also known catalysts. The use of these catalysts minimizes the loss of nitrogen and improves process efficiency for high ammonia synthesis [27]. One of two reactors could be used to conduct the electrolysis: hydrogen generation reactors or nitrogen reduction reactors. The latter is preferred for low-temperature applications and gives more yield in the downstream ammonia synthesis. Higher ammonia production efficiency can, however, be achieved in the hydrogen generation reactor by adding ZrO2 to the Ruthenium catalyst. Similarly, reducing the number of protons on the catalyst surface by using high-pH electrolytes has also been shown to solve the underproduction problem in the hydrogen generation reactor [28]. The source of nitrogen is also crucial in the EAS method. The moisture content of the air used as the nitrogen source is an important parameter that affects the ammonia conversion rate. Using high-purity nitrogen from the air with reduced moisture will significantly increase ammonia synthesis [29].
Albeit the innovation of EAS to produce ammonia in an environmentally friendly manner, energy consumption is still unacceptably high due to the high current density utilized for hydrogen production, and the process occurs at a low capacity (10-9 to 10-11 mol cm-2 s-1) [30]. The water electrolyzer used in EAS requires a continuous supply of high-purity pretreated water for its operation. Consequently, nine tons of water is consumed for every ton of hydrogen produced, and for the production of an amount of ammonia by EAS through water electrolysis, approximately double the amount of water is required, deepening the worldwide water crisis [24].
4. Biological Ammonia Production
Biological approaches are considered eco-friendly as they are natural processes that do not produce any harmful by-products. There are several approaches for biological ammonia production including nitrogen fixation, nitrification, nitrate/nitrite reduction, urea hydrolysis, metabolic engineering of microorganisms, and in vitro ruminal microbial fermentation of protein biomass, but the most reported methods are biological nitrogen fixation (BNF) and metabolic engineering of microorganism methods. Biological ammonia production by rumen bacteria fermentation of protein biomass experimented on in this review is a relatively new approach and has shown the potential to complement ammonia bioproduction.
4.Biological Nitrogen Fixation by Nitrogenase
Biological nitrogen fixation (BNF) is a natural process that converts atmospheric molecular nitrogen (N2) to ammonia (NH3). BNF, an ATP-dependent reduction reaction catalyzed by nitrogenase enzyme, is responsible for approximately half of the bioavailable nitrogen that supports all life forms [31]. Relative to the Haber-Bosch process which requires high temperature and pressure conditions to break down molecular nitrogen, nitrogen-fixing microorganisms produce ammonia at ambient temperature and pressure. Nitrogen-fixing microbes are robust and have been explored to produce biofertilizers in commercial quantities [32,33]. Researchers are actively making attempts to mimic the natural process of BNF by isolating nitrogen-fixing (Figure 3) bacteria and nitrogenase for synthetic ammonia production. The major challenge with this research effort is that nitrogenase catalysis is highly energy dependent, making its reaction rate slower than most enzymes in nature [34].
The main microorganisms that possess nitrogenase and carry out nitrogen fixation are the genus Rhizobia which colonizes the root of legumes, and species in the genera Azotobacter and Klebsiella that can fix nitrogen without parasitizing plant roots. The latter group is the main focus of research for synthetic BNF [35,36]. Nitrogenase requires up to eight molecules of ATP to produce a molecule of ammonia in an anoxic condition. Although the reaction mechanism of nitrogenase is unclear due to its interrelated multiple subunits, scientists have attempted to construct a heterologous expression system for Klebsiella nitrogenase subunits in E. coli [37]. Similarly, an heterologous expression of the Klebsiella nitrogenase gene cluster has been constructed in E. coli and yeast to understand the mechanism by which nitrogenase functions without oxygen as well as to increase its activity [38,39]. Various studies have also investigated how nitrogen-fixing bacteria can function under aerobic conditions without inactivating nitrogenase. Such researches involve the use of polysaccharide membrane to protect nitrogenase from oxygen exposure [39,40].
For industrial applications of nitrogen-fixing bacteria, companies such as Pivot Bio, Inc. and Ginkgo Bioworks engineered Enterobacter sp. lacking glutamine due to low expression of the transcription factor GlnR to increase intracellular glutamine and, consequently, synthesize ammonia in the presence of nitrogenase [41]. Steady nutrient supply through BNF has also been successful with non-leguminous crop plants such as corn. The use of anaerobic microflora is also a known strategy for ammonia bioproduction by BNF and a plethora of methodologies for ammonia recovery have been established. A notable one is the evaporation of solution following fermentation and pH increase [42,43,44].
4.2. Cell and Metabolic Engineering for Ammonia Production
Various biomass including food waste, microbial biomass, and protein-rich crop residues can be fermented by engineered microorganisms whose metabolisms are well understood for ammonia bioproduction. In a metabolic engineering study on the conversion of protein wastes into biofuels and ammonia using microbes, codY gene (a transcriptional regulator), in Bacillus subtilis was knocked out. codY gene regulates the activity of several other genes involved in different processes, such as producing branched-chain amino acids (ilvABHCD and leuABCD), removing amino groups from other molecules (ybgE, ald, yhdC, appBC, and dppBC), and inhibiting the expression of genes that cause protein break down and uptake (yhdG, appBC, and dppBC). In bacteria, proteins are encoded for amino acid biosynthesis by the ilv-leu operon. The deletion of codY gene removed regulatory constraints on this operon causing a significant increase in the production and uptake of branched-chain amino acids (BCAA) due to the derepression of ilv-leu operon and subsequent upregulation of genes responsible for BCAA synthesis.
In addition to the deletion of the codY gene, the BkdB gene in Bacillus subtilis was also knocked out. BkdB is a lipoamide acyltransferase enzyme that helps in the biosynthesis of branched-chain fatty acids by converting branched-chain keto acids into their acyl-CoA derivatives. This conversion inhibits the production of biofuels and ammonia. BkdB gene knockout had a significant impact on the production of branched-chain fatty acids in Bacillus subtilis. Obstruction of the production resulted in increased availability of metabolic precursors for the production of biofuels and NH3. To completely transform B. subtilis to favor ammonia synthesis, an alcohol dehydrogenase gene, leuDH, and two-keto-acid decarboxylase were overexpressed. LeuDH is an alcohol dehydrogenase gene that plays an important role in the conversion of amino acids to alpha-keto acids while two-keto-acid decarboxylase is an enzyme that catalyzes the decarboxylation of alpha-keto acids, which are important metabolic intermediates in amino acid biosynthesis. Overexpression of LeuDH increased the rate of amino-acid nitrogen reflux which helped to increase the efficiency of protein conversion. Similarly, overexpressing two-keto-acid decarboxylase led to the increased availability of metabolic precursors such as alpha-ketoisocaproate (KIC) and alpha ketoglutarate (AKG) for the production of ammonia. The resulting final strain of B. subtilis was employed in the fermentation of protein biomass obtained from E.coli cells. This process produced ammonia with a theoretical yield of about 50% [45].
A similar study on ammonia production from amino acid-based biomass-like sources using engineered E. coli has been reported [46]. Since E.coli assimilates ammonia intracellularly [47], the two genes involved in the ammonia assimilation pathway, glnA and gdhA which are both glutamine assimilation genes, were knocked out to enhance ammonia production. glnA encodes for enzyme glutamine synthetase (GS) and catalyzes the conversion of glutamate and ammonia to glutamine while gdhA encodes for enzyme glutamate dehydrogenase (GDH) and catalyzes the reversible conversion of glutamate and ammonia to alpha-ketoglutarate. Deletion of glnA promotes the extracellular leaching of ammonia while the deletion of gdhA increases ammonia flux to produce more glutamate, a known precursor of ammonia. In this study, deleting the two genes redirected the nitrogen assimilation pathways in E. coli toward ammonia production, resulting in a peak titer yield of 458 mg/L equivalent to an overall yield of 47.8% [46].
Further studies on the metabolic engineering of E. coli for ammonia production converted different food wastes including soy sauce cake, mirin cake, and tomato peel to ammonia. Using metabolic profiling to assess the correlation between substances in the media (amino acids, sugars, and organic acids) and ammonia production, glucose was implicated as an inhibitor of ammonia production. When glucose was added to the amino acid-containing medium at different concentrations, a negative correlation with ammonia production was obtained. Thus, E coli was engineered to hinder the inhibitory effect of glucose by knocking out the transporter gene, ptsG, and the phosphotransferase system which transports glucose and other sugars. Briefly, the polymerase chain reaction (PCR) technique was used to amplify and copy specific fragments of genes that encoded resistance to pts'G-Kim and glnA-Km (amplified from pKD13) using primers ptsGF and ptsGR. The amplified DNA fragments were then transferred into E. coli cells through electroporation. Following the transfer, E. coli cells were grown on LB agar containing specific antibiotics – ampicillin and kanamycin. This allowed only the cells that had taken up the amplified DNA fragments to survive and grow, while the others died off. By repeating this process with different combinations of DNA fragments and antibiotics, more varieties of E. coli strains with different genetic modifications, such as AptsG and AglnA were created. To ensure that the modified DNA fragments had been inserted into the correct location in the E. coli genome, PCR was used to amplify and sequence the insertion region using insertion-checking primers. The resulting E. coli strain succeeded in producing ammonia in a glucose-containing amino acid medium, with up to 73% yield [48]. In the studies described above, ammonia was, however, produced intracellularly. As a result, the produced ammonia can still be used up by these microbes for growth [47]. Therefore, a system that can produce ammonia extracellularly without impeding microbial growth may improve productivity.
Studies on yeast for extracellular ammonia production have been attempted. Prominent among such studies is the use of yeast cell surface engineering (YCSE) systems to avoid ammonia toxicity and assimilation. In YCSE, the protein to be converted to ammonia is displayed on the cell surface usually by the attachment of a secretory signal to the N-terminus of the target protein and a signal sequence, an α-agglutin containing a glycosylphosphatidylinositol anchor, on its C-terminus. Briefly, the plasmid for yeast cell surface display of L-amino acid oxidase was constructed by synthesizing and inserting the codon-optimized sequence of the HcLAAO (L-amino acid oxidase) into pULDl, resulting in a plasmid named pULDl-HcLAAO. A strep-tag negative control plasmid called pULDl-s was also constructed. The yeast strain Saccharomyces cerevisiae BY4741/sedlA was utilized to display HcLAAO on the cell surface. The constructed plasmid was then introduced into the yeast strain. Yeast cells were then transformed and grown in a synthetic dextrose medium and cultured in SDC buffer at pH 7.0. Using this approach, up to106 target proteins could be displayed on the yeast cell surface which are then used as biocatalysts for enzyme immobilization [47,49,50].
Ammonia production from soybean residues has been successful with the YCSE technique [51]. Amino acid catabolic enzymes that produce ammonia from amino acid precursors such as ammonia lyases have attracted interest for their efficiency in being displayed on the yeast cell surface because their catalysis does not require cofactors, unlike nitrogenases. With yeast cells displaying glutamine ammonia-lyases, ammonia was produced from glutamine solution reaching a titer of up to 3.34g/L and efficiency of 83.2% [51]. The limitation of this approach is that only glutamine of the 20 amino acids can be utilized. Interestingly, L-amino acid oxidase with a broad substrate specificity can be displayed for ammonia production from several amino acids [52,53]. These are lab-scale studies that may be difficult to transition to an industrial scale for eco-friendly biological ammonia production. Table 1 shows a summary of the metabolic engineering route for biological ammonia production.
4.3. Hyper ammonia-producing bacteria route
The digestive compartment of ruminant animals, the rumen, is a biorefinery for ammonia production. Ruminal microorganisms can break down plant materials containing carbohydrates and proteins in their feeds for energy. The products of protein degradation including peptides and amino acids are metabolized to protein and/or ammonia. The microbial protein thus formed is required for animal products, but the ammonia is absorbed from the rumen, metabolized, and excreted in the urine. This is an inefficient use of dietary proteins with devastating consequences on the environment through environmental nitrogen pollution [54].
Several studies in the animal sciences have sought strategies to promote microbial protein synthesis and regulate ammonia production. These studies revealed the identity of a certain group of bacteria whose rate of ammonia production is much higher than can be used up by the ruminal microbes for other functions including microbial protein synthesis [55,56,57]. This group of bacteria, known as the hyper ammonia-producing bacteria (HAB) can effectively convert dietary protein to surplus ammonia [58,59]. This type of natural ammonia is produced when the digestive system of man and animal undergoes a biochemical reaction leading to the breakdown of nitrogen-containing amine (NH2) of proteins into ammonia or the ionic form (ammonium) is referred to as biological ammonia.
The first step towards the degradation of amino acids is deamination which is the removal of an amine group to convert it to ammonia. It has been reported that amino acid deamination in the rumen produces more ammonia than can be utilized by the bacteria [60]. Deamination may occur through oxidation, reduction, hydrolysis, or removal of elements. It helps to free the carbon skeleton by removing the amine group from the amino acid. Furthermore, deamination could be carried out on either a single amino acid, pairs of amino acids as in the case of Stickland reaction, or a combination of amino acids and a non-nitrogenous compound with all resulting into ammonia and keto-acids as major products [61].
The next biochemical reaction is called ammonification which is the second stage of mineralization [62]. Useful energy can also be derived metabolically by bacteria and related microorganisms through ammonification. Ammonium (NH4+) is thus produced by microorganisms and if in excess, it is excreted into the environment as nutrients for uptake by plants or as feedstock for further nitrification [62]. HABs have been implicated in converting ~50% of ruminal dietary protein to ammonia [63,64].
HABs are found in cattle rumen or swine manure stored in the pit [65,66,67]. Additionally, HABs thrive in the rumen of hay-fed cattle compared to grain-fed cattle [68] because the pH of hay-fed cattle rumen environment is relatively neutral, thus providing a favorable condition for their growth compared to the slightly acidic pH (<6.0) observed in grain-fed cattle [69]. HABs are capable of producing up to 40 mM (0.6812 mg/L) of ammonia in peptone-amino acid medium depending on energy and carbon source [66,70]. HABs can operate in both anaerobic and aerobic environments, but anaerobic-HAB are more prominent and of major concern because they convert a large percentage of dietary protein in the rumen to ammonia. Although HABs are detrimental to ruminant metabolism due to excess ammonia generation causing toxicity to rumen microbes and hyperammonemia in farm animals [71], they can be harnessed as a sustainable source for large-scale ammonia production with low energy requirements and zero emissions.
There are several strains of hyper-ammonia-producing bacteria (HAB) with different biological ammonia-production capacities. Selenomonas ruminantium, Peptostreptococcus elsdenii, and Bacteroides ruminicola are HAB strains that are capable of producing at least 1µM of biological ammonia on a lab scale through deamination. S. ruminantium catabolizes cysteine hydrolysate, while P. elsdenii breaks down casein hydrolysate and specific amino acids (L-serine, L-threonine, and L-cysteine) to produce biological ammonia [72,73]. Depending on HAB strain and environmental conditions, it is also possible to produce much higher concentrations of biological ammonia (> 24 mM) [70]. HAB can operate in both anaerobic and aerobic environments, but anaerobic-HAB are more prominent and of major concern because they convert a large percentage of dietary protein in the rumen to ammonia [65].
5. Developing Biological Ammonia Biomanufacturing
5.1. Conceptual bioprocess flow
There have been a few attempts to employ HAB in the fermentation of dietary proteins to produce biological ammonia. For instance, in a study to investigate the affinity of hyper ammonia-producing bacteria (HABs) to produce biological ammonia, three hyper ammonia-producing ruminant bacteria (Clostridium aminophilum, Peptostreptococcus anaerobius, and Clostridium sticklandii) were anaerobically cultured using five different organic nitrogen substrates: soy protein isolate (SPI), blood meal (BM), feather meal (FM), dried fish meal (DFM), and yeast extract (YE). The study examined the affinity of these HABs to produce biological ammonia. The ability of these bacteria to produce ammonia when grown in pure culture with various protein sources was then assessed to determine which bacteria species and protein substrate produce the highest concentrations of ammonia. Results showed that all three bacteria produced ammonia at various rates, depending on the organic nitrogen substrate used. Clostridium aminophilum and SPI produced the highest biological ammonia concentration of about 7.23 mM [74]. With this finding, the fermentation of proteins using HABs could be harnessed for sustainable biological ammonia production.
As earlier stated, there have been few studies on the fermentation of hydrolyzed proteins, peptides, and amino acids. HABs synthesize varying amounts of biological ammonia from different peptides and amino acids [65,74]. In fact, before fermentation, HABs inherently secrete proteases to initially hydrolyze protein [65]. The protein hydrolysates, which comprise oligopeptides, peptides, and amino acids, are easily absorbed by HABs [65]. However, there has not been any study that compares hydrolyzed and unhydrolyzed protein during biological ammonia production. Therefore, to understand the importance of pre-fermentation hydrolysis, future works should investigate the effect of pre-fermentation enzymatic hydrolysis of protein on biological ammonia production. Hence, we include protein hydrolysis as an important step in the conceptual bioprocess flow for biological ammonia biomanufacturing (Figure 4).
5.2. Protein Hydrolysis
Proteins are large polymers of amino acids joined together, primarily, by peptide bonds. Polypeptides and peptides are long and short chains of amino acids, respectively. Most of the proteins present in soybean and soybean meals are in the form of storage globulins [75]. The polypeptide chains of the proteins are entangled into a three-dimensional complex structure by several hydrogen and disulfide bonds, amounting to a molecular weight of up to 600,000 kDa. Soybean proteins are insoluble in water at their isoelectric point. The two major types of proteins in soybean seeds are glycinin and conglycinin that form ~80% of the total soybean protein. Glycinin and conglycinin have similar secondary structures with 57% random coils, 38% beta-sheet, and 6% alpha-helix. In glycinin, glycine, tyrosine, and tryptophan are wrapped inside the globular structure; whereas in conglycinin tryptophan is exposed. Also, glycinin has two major sub-proteins: acidic glycinin of ~40 kDa and basic glycinin of 20 kDa, whereas conglycinin has three sub-proteins: alpha-conglycinin, beta-conglycinin, and gamma-conglycinin with 68 kDa, 175 kDa, and up to 200 kDa, respectively [76]. These protein polymers must be deconstructed and utilized by microbes for ammonia bioproduction. Products of protein hydrolysis— amino acids and peptides— are referred to as protein hydrolysates. Relative to intact proteins, protein hydrolysates have increased solubility which could enhance their bioavailability. Complete protein degradation generates amino acids that are then further catabolized and deaminated to release ammonia. Peptides, products of partial protein degradation with short chains of amino acids, are also utilized by ruminal microbes for ammonia biosynthesis. Catabolism of peptides by ruminal microbes involves the activity of peptidases secreted by these microbes. Peptidases abound in the rumen and have different substrate specificities [77]. However, several studies have shown that external enzymes can be highly effective in enhancing soybean protein degradation and utilization of the resulting products— short peptides and amino acids in the rumen [78,79]. In vitro digestion of proteins for various applications has been extensively studied. The most common methods for protein hydrolysis are biological (enzymatic) and thermochemical hydrolysis.
5.2.1. Biological/Enzymatic Protein Hydrolysis
Enzymatic hydrolysis is a safe and effective approach to processing proteins to hydrolysates with improved functional properties. Enzymes used for protein hydrolysis are also called proteases. The operational conditions for enzymatic hydrolysis are environmentally friendly and reduce the formation of by-products [80]. Common enzymes used include pepsin, trypsin, chymotrypsin, papain, and various extracted fungal and bacterial proteases such as flavourzyme, alcalase, neutrase, and protamex. The form of soybean proteins (toasted or untoasted soybean meal (SM) protein, processed SM, heated SM, defatted SM, native SPI, etc.) and the enzymes/proteases impact the degree of hydrolysis (DH) and functional properties of the resulting hydrolysates [81]. The extent of protein denaturation that gives rise to different protein forms determines the level of resistance of the protein to proteolysis. In toasted SM, for instance, beta-conglycinin is effectively hydrolyzed by many different endo-proteases [82]. On the other hand, high resistance to proteolysis was observed with beta-conglycinin than with glycinin by rumen bacteria [83]. These results informed the need to explore the potential of using multiple enzymes together or sequentially to achieve a complete breakdown of any protein form.
5.2.2. Multi-enzymatic Hydrolysis
Multi-enzymatic hydrolysis of protein is the combination of different enzymes to digest protein molecules into peptides and amino acids. Based on their catalytic mechanisms and an amino acid sequence, proteases are diverse and have different activity and specificity. While serine proteases use serine residue in their active sites to cleave peptide bonds, cysteine proteases, and metalloproteases use cysteine residue and metal ions to cleave peptide bonds, respectively. Some proteases such as trypsin cleave specific peptide bonds only (after lysine and arginine residues); yet other proteases have broader specificity, especially the industrial proteases. Proteases also differ in their site of catalytic action. They could either cut from within (endoproteases) or from the terminal amino acid residues (exopeptidase). Industrial proteases are usually a mixture of different enzymes purified from bacteria or fungi. Thus, a single industrial enzyme could be both an endo- and exoprotease. For example, flavourzyme is a mixture of proteases with both endo- and exopeptidase activities produced and purified from Aspergillus oryzae [84].
Studies have assessed the potential of different industrial proteases in simultaneous or sequential combinations for the hydrolysis of different protein isolates. These studies revealed that proteases used in combination increase the functional properties of the resulting protein hydrolysates [85,86,87]. The concentrations of the constituent enzymes in combination have also been shown to increase the DH of protein isolates. In a study, DH was shown to increase significantly when a higher concentration of alcalase in an alcalse-flavourzyme combination and alcalse-corolase combination was used to hydrolyze potato pulp protein [88]. The alcalase-flavourzyme combination gave the highest DH in both the hydrolysis of potato pulp protein and poultry meal [88,89]. Proteases combination for hydrolysis in any bioconversion is process specific. Several studies have revealed the different combinations that work for specific bioconversion processes [87,90,91]. There is a paucity of data on protein hydrolysis for biological ammonia production. Thus, research efforts in this area would enhance the development of optimal parameters for biological ammonia production.
5.3. Leading HABs for Biological Ammonia Production
Clostridium aminophilum (amino and philos meaning loving amino acids) is a species of bacteria in the family Clostridiaceae. It is an atypical gram-positive bacterium with a length and width of 1.0 µm and 1.5 µm, respectively. The cellular spores are non-motile and can withstand a temperature as high as 80°C for 10min. Classified as obligately anaerobic with an optimum growth temperature of 25°C to 45°C, they utilize glutamine, glutamate, serine, and histidine as carbon sources [92]. Their end products of fermentation include ammonia, acetate, and butyrate with tinctures of lactate and succinate.
Clostridium sticklandii is also a specie of bacteria in the genus Acetoanaerobium and the family Peptostreptococcaceae. It is also a gram-positive obligate bacterium that utilizes threonine, arginine, serine, threonine, cysteine, proline, and glycine as preferable carbon sources. [93], in their research on the biology of bacteria, concluded that C. sticklandii can utilize pairs of amino acids through the Stickland reaction to bring forth fermentation products such as acetate, butyrate, and ammonia.
Peptostreptococcus anaerobius unlike Clostridium aminophilum is a non-spore-forming obligate bacterium. It is, however, gram-positive [94]. Their common carbon sources include leucine, serine, threonine, glycine, and phenylalanine [95]. P. anaerobius can also break down amino acids and peptones into ammonia, acetic, butyric, and isobutyric acid. It is however worthy of note that there is very little or no information in the literature on the genetic makeup and characteristics of HAB.
5.4. Factors affecting biological ammonia production
Several bacteria species have over time been identified with a speedy rate of ammonia production [96]. However, the ability of these bacteria to produce ammonia at an optimum rate is dependent on several factors. Research on the isolation and characterization of these bacteria has been carried out extensively to understand the factors responsible for ammonia production.
5.4.1. Effect of diet, substrate, and substrate combination
According to [97] , the rate at which ammonia is produced changes based on diet. The rate at which organic ammonia proceeds is also dependent on the substrate, its concentration, and the method of ammonia production utilized [97]. Isolation of bacteria by [95] identified Peptostreptococcus and Clostridium as ammonia-producing bacteria when soybean meal was a major ingredient in ruminant feed. Also, in a study to appraise the rate of ammonia production by HAB, pure cultures of P. anaerobius, C. sticklandii, and C. aminophilum were grown invitro utilizing different substrates and in different combinations. Five different diets rich in organic nitrogen sources (soy protein isolate (SP), blood meal (BM), feather meal (FM), dried fish (DF), and yeast extract (YE) were utilized to discover the best protein substrate and bacteria specie yielding maximal quantity of ammonia. Results showed that with the combination of all substrates, P. anaerobius produced the least amount of ammonia followed by C. sticklandii and then C. aminophilum. The same quantity of organic ammonia was produced for combinations of BM alone, BM and YE, SP and YE, FM and YE, DF alone, DF and YE, and YE alone. The results recorded is in alignment with [92] where C. aminophilum was best in the production of organic ammonia. Moreover, C. aminophilum yielded the highest organic ammonia concentration (7.23 mM) when cultured on soy protein isolate alone [74]. The substrates were particularly used due to their richness in certain specific amino acids that aid the rapid multiplication of HAB species [74]. A careful study of the amino acid profile of the best experimental substrate (Soy protein isolate) used by [74] revealed the presence of nine standard and essential amino acids which can only be synthesized from food sources. They include histidine, isoleucine, methionine, phenylalanine, lysine, valine, threonine, leucine, and tryptophan [98,99]. [74] therefore concluded that the effect of substrate on HAB species was highly substantial to the extent that it accounted for about 17.0% of the variation, indicating that all bacteria responded differently to the substrate.
Moreover, following the growth of swine manure isolates and ruminal HAB on different sources of amino acid, there were pronounced variations in the growth and ammonia production of HAB [100]. As described in a previous study, HAB thrived well on tryptone, casamino acids, and a mixture of both producing ammonia in high concentrations [70,95,101].
Similarly, in a study of ruminal ammonia-producing bacterial species, three experimental cows A, B, and C were put on dissimilar diets of timothy hay, grain mixture, and alfalfa hay, respectively [102]. Animals were later fed with the feed mixture in varying proportions. Following the collection of rumen samples, bacteria strains were isolated and subsequently labeled as A, B, and C based on the source (animal) from which the samples were collected. Results showed a difference in the concentration of ammonia produced. Variation in the amount of organic ammonia produced has been attributed to diet. In addition, [68] conducted research aimed at calculating and equating the ammonia production of mixed ruminal bacteria with their deamination rates and that of carbohydrate-fermenting ruminal bacteria, thus evolving a mathematical model of ammonia production and figuring out the population of HAB in the rumen of cattle fed predominantly with hay or grain-based diets. Results showed that the rate of ammonia production from mixed ruminal bacteria of cattle fed with hay doubled that of cattle fed predominantly with grains. In furtherance to this research, a mathematical model supporting the results was developed. The model showed that the hyper ammonia-producing ruminal bacteria of cattle fed with hay was four times more thus indicating that HAB of cattle-fed hay had a high maximal velocity of ammonia production.
5.4.2. Effect of pH
Microorganisms are generally sensitive to their ambient hydrogen ion concentration (pH). Most ruminal bacteria are anaerobes and thus act optimally at neutrality (pH range of 6.5-7.5). In research to determine ammonia production from HAB, a pH of 7.0 was utilized [68]. Likewise, in research on ammonia production by ruminal microorganisms, ammonia production was determined utilizing filtered ruminal fluid from sheep using the method proposed by [103]. Substrates such as casein, Trypticase Peptone, soluble soybean protein, and Soya Peptone were utilized. The pH of the resulting liquid (supernatant) was brought to 7 by neutralization with KOH and the rates of ammonia production were determined through linear regression. In addition, all incubations were also carried out at pH of 6.7 to 7 in a similar research on ammonia production by monensin-sensitive ammonia producing bacteria [70]. However, other studies have indicated that there might be no direct relationship between rumen pH and the percentage of HAB organisms present in the ruminants sampled [96] suggesting that pH may not have any effect on the rate of ammonia production.
5.4.3. Effect of Temperature
Temperature plays a very significant role in ammonia production by HAB. Different bacteria have different temperature ranges for optimal performance. In a research on the isolation and identification of HAB from swine manure storage pits, all bacterial isolates were grown at 37°C [100]. Likewise, the measurement of ammonia produced in ruminal fluid in vitro involving the dissolution of soluble soybean protein in water was also done at room temperature [97].
Furthermore, in a research on the estimations of hyper-ammonia-producing ruminal bacteria by [68], ruminal bacteria were anaerobically grown at a temperature of 39oC in a basal medium. Isolated ruminal hyper ammonia-producing bacteria have also been previously cultured in a growth medium at 39oC [104]. Likewise, in a study on the characterization of ruminal microorganisms responsible for ammonia production, sheep-filtered ruminal fluid was grown in vitro at a temperature of 39°C [97]. These results show consistency in temperature values and thus suggest that HAB are generally mesophilic, acting optimally at temperatures between 20 - 45 oC.
5.4.4. Effect of Agitation
Agitation is generally referred to as the forceful mechanical flow of fluids in a container. The flow could be in a circulatory pattern or otherwise to attain mixture homogenization. Presently, it appears that information on the study of agitation to produce ammonia by HAB is not available in the literature.
5.4.5. Effect of time
The effect of time in the production of organic ammonia by HAB cannot be over-emphasized. In the experiment on the potential of HAB to produce organic ammonia, results indicated that C. aminophilum yielded the most organic ammonia when cultured on soy protein isolate substrate for 96h [74]. The main effect of time on organic ammonia production accounted for about 9.7% of the variation. It was however observed that P. anaerobius had very low growth rates and produced the lowest organic ammonia concentrations and was unaffected by time. [102] in their study of bacteria species from the rumen found out that the incubation period for ammonia production gradually increased from 72 h. In the long run, a 96h standard time was chosen.
5. Conclusions
In this review, we have presented numerous attempts towards achieving sustainable, CO2 emission-free biological ammonia production, and although several promising approaches have been presented, their respective limitations have also been acknowledged. In addition to these renewable alternatives, we have proposed a novel strategy for generating biological ammonia through the fermentation of dietary proteins via the rumen hyper ammonia-producing bacteria pathway. We recommend further exploration and investigation of this approach for adapting rumen microbial fermentation to laboratory-scale studies, with the ultimate goal of advancing this approach toward commercialization.
Funding
This research and the APC were funded by North Dakota Soybean Council and State Board of Agricultural Research and Education with grant numbers FARG0034901 and FARG090432, respectively
References
- MDA Physical and Chemical Properties of Anhydrous Ammonia.
- IIAR Ammonia Data Book; 2nd Editio.; International Institute of Ammonia Refrigeration: Alexandria, VA, 2008; ISBN 1635473500, 9781635473506.
- Smith, C.; Hill, A.K.; Torrente-Murciano, L. Current and Future Role of Haber-Bosch Ammonia in a Carbon-Free Energy Landscape. Energy and Environmental Science 2020, 13, 331–344. [Google Scholar] [CrossRef]
- Smil, V. Enriching the Earth: Fritz Haber, Carl Bosch, and the Transformation of World Food Production; MIT press, 2004. F: the Earth.
- Liu, B.; Manavi, N.; Deng, H.; Huang, C.; Shan, N.; Chikan, V.; Pfromm, P. Activation of N2 on Manganese Nitride-Supported Ni3 and Fe3 Clusters and Relevance to Ammonia Formation. The Journal of Physical Chemistry Letters 2021, 12, 6535–6542. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.; Lamprecht, R.E.; Marushchak, M.E.; Lind, S.E.; Novakovskiy, A.; Aurela, M.; Martikainen, P.J.; Biasi, C. Warming of Subarctic Tundra Increases Emissions of All Three Important Greenhouse Gases–Carbon Dioxide, Methane, and Nitrous Oxide. Global Change Biology 2017, 23, 3121–3138. [Google Scholar] [CrossRef] [PubMed]
- Norskov, J.; Chen, J.; Bullock, M.; Chirik, P.; Chorkendorff, I. Sustainable Ammonia Synthesis. DOE Roundtable Report 2016, 2–2016. [Google Scholar]
- Zhang, H.; Wang, L.; Van, J.; Maréchal, F.; Desideri, U. Techno-Economic Comparison of Green Ammonia Production Processes. Applied Energy 2019, 114135. [Google Scholar] [CrossRef]
- Ghavam, S.; Vahdati, M.; Wilson, I.G.; Styring, P. Sustainable Ammonia Production Processes. Frontiers in Energy Research 2021, 9, 580808. [Google Scholar] [CrossRef]
- Fernandez, L. Ammonia Production Worldwide from 2010 to 2021 Available online: https://www.statista.com/statistics/1266378/global-ammonia-production/#:~:text=In 2021%2C the global production,approximately 64.6 million metric tons.
- Pfromm, P.H. Towards Sustainable Agriculture : Fossil-Free Ammonia Towards Sustainable Agriculture : Fossil-Free Ammonia. 2017, 034702. [CrossRef]
- FAO, F. World Fertilizer Trends and Outlook to 2020. Food and Agriculture Organization of the United Nations 2017.
- Bird, F.; Clarke, A.; Davies, P.; Surkovic, E. Ammonia: Zero-Carbon Fertiliser, Fuel and Energy Store; KBR Inc.: London, 2020; ISBN 9781782524489. [Google Scholar]
- Papavisasam, S. Oil and Gas Industry Network. In Corrosion Control in the Oil and Gas Industry; 2014; pp. 41–131.
- Paschkewitz, T.M. Ammonia Production at Ambient Temperature and Pressure, University of Iowa, 2012.
- Boerner, L.K. Industrial Ammonia Production Emits More CO2 than Any Other Chemical-Making Reaction. Chemists Want to Change That. Chem. Eng. News 2019, 97, 1–9. [Google Scholar]
- Yüzbaşıoğlu, A.E.; Tatarhan, A.H.; Gezerman, A.O. Decarbonization in Ammonia Production, New Technological Methods in Industrial Scale Ammonia Production and Critical Evaluations. Heliyon 2021, 7, e08257. [Google Scholar] [CrossRef]
- Hussin, F.; Aroua, M.K. Recent Trends in the Development of Adsorption Technologies for Carbon Dioxide Capture: A Brief Literature and Patent Reviews (2014–2018). Journal of Cleaner Production 2020, 253, 119707. [Google Scholar] [CrossRef]
- Abdelkareem, M.A.; Lootah, M.A.; Sayed, E.T.; Wilberforce, T.; Alawadhi, H.; Yousef, B.A.A.; Olabi, A.G. Fuel Cells for Carbon Capture Applications. Science of the Total Environment 2021, 769, 144243. [Google Scholar] [CrossRef] [PubMed]
- Kamble, A.R.; Patel, C.M.; Murthy, Z.V.P. A Review on the Recent Advances in Mixed Matrix Membranes for Gas Separation Processes. Renewable and Sustainable Energy Reviews 2021, 145, 111062. [Google Scholar] [CrossRef]
- Mutch, G.A.; Qu, L.; Triantafyllou, G.; Xing, W.; Fontaine, M.L.; Metcalfe, I.S. Supported Molten-Salt Membranes for Carbon Dioxide Permeation. Journal of Materials Chemistry A 2019, 7, 12951–12973. [Google Scholar] [CrossRef]
- Siqueira, R.M.; Freitas, G.R.; Peixoto, H.R.; Nascimento, J.F.D.; Musse, A.P.S.; Torres, A.E.B.; Azevedo, D.C.S.; Bastos-Neto, M. Carbon Dioxide Capture by Pressure Swing Adsorption. Energy Procedia 2017, 114, 2182–2192. [Google Scholar] [CrossRef]
- Bello, I.; Rasaq, N.; Adeniyi, A.; Hammed, A. Enzyme Aided Processing of Oil. International Journal of Halal Research 2021, 3, 60–72. [Google Scholar] [CrossRef]
- Ghavam, S.; Vahdati, M.; Wilson, I.A.G.; Styring, P. Sustainable Ammonia Production Processes. Frontiers in Energy Research 2021, 9, 580808. [Google Scholar] [CrossRef]
- Miller, H.A.; Bouzek, K.; Hnat, J.; Loos, S.; Bernäcker, C.I.; Weißgärber, T.; Röntzsch, L.; Meier-Haack, J. Green Hydrogen from Anion Exchange Membrane Water Electrolysis: A Review of Recent Developments in Critical Materials and Operating Conditions. Sustainable Energy and Fuels 2020, 4, 2114–2133. [Google Scholar] [CrossRef]
- Yang, J.; Weng, W.; Xiao, W. Electrochemical Synthesis of Ammonia in Molten Salts. Journal of Energy Chemistry 2020, 43, 195–207. [Google Scholar] [CrossRef]
- Zhang, L.; Ji, X.; Ren, X.; Ma, Y.; Shi, X.; Tian, Z.; Asiri, A.M.; Chen, L.; Tang, B.; Sun, X. Electrochemical Ammonia Synthesis via Nitrogen Reduction Reaction on a MoS2 Catalyst: Theoretical and Experimental Studies. Advanced Materials 2018, 30, 1–6. [Google Scholar] [CrossRef]
- Casallas, C.; Dincer, I. Assessment of an Integrated Solar Hydrogen System for Electrochemical Synthesis of Ammonia. International Journal of Hydrogen Energy 2017, 42, 21495–21500. [Google Scholar] [CrossRef]
- Giddey, S.; Badwal, S.P.S.; Kulkarni, A. Review of Electrochemical Ammonia Production Technologies and Materials. International Journal of Hydrogen Energy 2013, 38, 14576–14594. [Google Scholar] [CrossRef]
- Frattini, D.; Cinti, G.; Bidini, G.; Desideri, U.; Cioffi, R.; Jannelli, E. A System Approach in Energy Evaluation of Different Renewable Energies Sources Integration in Ammonia Production Plants. Renewable Energy 2016, 99, 472–482. [Google Scholar] [CrossRef]
- Boyd, E.S.; Peters, J.W. New Insights into the Evolutionary History of Biological Nitrogen Fixation. Frontiers in Microbiology 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Murugeasn, S.; Dhar, D. Biofertilizers for Higher Pulse Production in India : Scope, Accessibility and Challenges. Indian Journal of Agronomy 2016, 61, 173–181. [Google Scholar]
- Khosro, M.; Yousef, S. Bacterial Biofertilizers for Sustainable Crop Production : A Review. Journal of Agricultural and Bological Science 2012, 7, 307–316. [Google Scholar]
- Rapson, T.D.; Wood, C.C. Analysis of the Ammonia Production Rates by Nitrogenase. Catalysts 2022, 12, 844. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of Nitrogen Fixation in Rhizobia. Microbial Biotechnology 2020, 13, 1314–1335. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nature Reviews Microbiology 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Temme, K.; Zhao, D.; Voigt, C.A. Refactoring the Nitrogen Fixation Gene Cluster from Klebsiella Oxytoca. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 7085–7090. [Google Scholar] [CrossRef]
- López-Torrejón, G.; Burén, S.; Veldhuizen, M.; Rubio, L.M. Biosynthesis of Cofactor-Activatable Iron-Only Nitrogenase in Saccharomyces Cerevisiae. Microbial Biotechnology 2021, 14, 1073–1083. [Google Scholar] [CrossRef]
- Takimoto, R.; Tatemichi, Y.; Aoki, W.; Kosaka, Y.; Minakuchi, H.; Ueda, M.; Kuroda, K. A Critical Role of an Oxygen-Responsive Gene for Aerobic Nitrogenase Activity in Azotobacter Vinelandii and Its Application to Escherichia Coli. Scientific Reports 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Wang, D.; Xu, A.; Elmerich, C.; Ma, L.Z. Biofilm Formation Enables Free-Living Nitrogen-Fixing Rhizobacteria to Fix Nitrogen under Aerobic Conditions. ISME Journal 2017, 11, 1602–1613. [Google Scholar] [CrossRef]
- Bhatti, M.; Feng, P.C.C.; Pitkin, J. Methods and Compositions for Improving Plant Health 2014.
- Yenigün, O.; Demirel, B. Ammonia Inhibition in Anaerobic Digestion: A Review. Process Biochemistry 2013, 48, 901–911. [Google Scholar] [CrossRef]
- Whelan, M.J.; Everitt, T.; Villa, R. A Mass Transfer Model of Ammonia Volatilisation from Anaerobic Digestate. Waste Management 2010, 30, 1808–1812. [Google Scholar] [CrossRef]
- Walker, M.; Iyer, K.; Heaven, S.; Banks, C.J. Ammonia Removal in Anaerobic Digestion by Biogas Stripping: An Evaluation of Process Alternatives Using a First Order Rate Model Based on Experimental Findings. Chemical Engineering Journal 2011, 178, 138–145. [Google Scholar] [CrossRef]
- Choi, K.; Wernick, D.G.; Tat, C.A.; Liao, J.C. Consolidated Conversion of Protein Waste into Biofuels and Ammonia Using Bacillus Subtilis. Metabolic Engineering 2014, 23, 53–61. [Google Scholar] [CrossRef]
- Mikami, Y.; Yoneda, H.; Tatsukami, Y.; Aoki, W.; Ueda, M. Ammonia Production from Amino Acid - Based Biomass - like Sources by Engineered Escherichia Coli. AMB Express 2017, 7, 83. [Google Scholar] [CrossRef]
- Watanabe, Y.; Aoki, W.; Ueda, M. Improved Ammonia Production from Soybean Residues by Cell Surface-Displayed l-Amino Acid Oxidase on Yeast. Bioscience, Biotechnology, and Biochemistry 2021, 85, 972–980. [Google Scholar] [CrossRef]
- Tatemichi, Y.; Kuroda, K.; Nakahara, T.; Ueda, M. Efficient Ammonia Production from Food by - Products by Engineered Escherichia Coli. AMB Express 2020, 10, 150. [Google Scholar] [CrossRef]
- Ueda, M. Establishment of Cell Surface Engineering and Its Development. Bioscience, biotechnology, and biochemistry 2016, 80, 1243–1253. [Google Scholar] [CrossRef]
- Kuroda, K.; Ueda, M. Cell Surface Engineering of Yeast for Applications in White Biotechnology. Biotechnology letters 2011, 33, 1–9. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kuroda, K.; Tatemichi, Y.; Nakahara, T.; Aoki, W.; Ueda, M. Construction of Engineered Yeast Producing Ammonia from Glutamine and Soybean Residues ( Okara ). AMB Express 2020, 10, 70. [Google Scholar] [CrossRef]
- Lu, P.; Ma, D.; Chen, Y.; Guo, Y.; Chen, G.Q.; Deng, H.; Shi, Y. L-Glutamine Provides Acid Resistance for Escherichia Coli through Enzymatic Release of Ammonia. Cell Research 2013, 23, 635–644. [Google Scholar] [CrossRef]
- Bloess, S.; Beuel, T.; Krüger, T.; Sewald, N.; Dierks, T.; Fischer von Mollard, G. Expression, Characterization, and Site-Specific Covalent Immobilization of an L-Amino Acid Oxidase from the Fungus Hebeloma Cylindrosporum. Applied Microbiology and Biotechnology 2019, 103, 2229–2241. [Google Scholar] [CrossRef]
- Puniya, A.K.; Singh, R.; Kamra, D.N. Rumen Microbiology: From Evolution to Revolution. Rumen Microbiology: From Evolution to Revolution 2015, 1–379. [CrossRef]
- Eschenlauer, S.C.P.; McKain, N.; Walker, N.D.; McEwan, N.R.; Newbold, C.J.; Wallance, R.J. Ammonia Production by Ruminal Microorganisms and Enumeration, Isolation, and Characterization of Bacteria Capable of Growth on Peptides and Amino Acids from the Sheep Rumen. Applied and Environmental Microbiology 2002, 68, 4925–4931. [Google Scholar] [CrossRef]
- Rychlik, J.L.; Russell, J.B. Bacteriocin-Like Activity of Butyrivibrio Fibrisolvens JL5 and Its Effect on Other Ruminal Bacteria and Ammonia Production. Applied 2002, 68, 1040–1046. [Google Scholar] [CrossRef]
- Pengpeng, W.; Tan, Z. Ammonia Assimilation in Rumen Bacteria: A Review. Animal Biotechnology 2013, 24, 107–128. [Google Scholar] [CrossRef]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen Metabolism in the Rumen. Journal of dairy science 2005, 88, E9–E21. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J. Nitrogen Metabolism. Nutritional ecology of the ruminant 1994. [Google Scholar]
- Rychlik, J.L.; Lavera, R.; Russell, J.B. Amino Acid Deamination by Ruminal Megasphaera Elsdenii Strains. 2002, 45, 340–345. [CrossRef]
- Doelle, H.W. Nitrogen Metabolism as an Energy Source for Anaerobic Microorganisms (Clostridium). Bacterial Metabolism; Elsevier: Amsterdam, The Netherlands 1969. [Google Scholar]
- Strock, J.S. Nitrogen Cycle. 2008, 162–165.
- Nolan, J.V. Quantitative Models of Nitrogen Metabolism in Sheep. Digenstion and Metabolism in the Ruminant 1975. [Google Scholar]
- Valente, T.N.P.; da Silva Lima, E.; dos Santos, W.B.R.; Cesário, A.S.; Tavares, C.J.; de Freitas, M.A.M. Ruminal Microorganism Consideration and Protein Used in the Metabolism of the Ruminants: A Review. African Journal of Microbiology Research 2016, 10, 456–464. [Google Scholar]
- Nagaraja, T.G. Microbiology of the Rumen. Rumenology 2016, 39–61. [Google Scholar]
- Whitehead, T.R.; Cotta, M.A. Isolation and Identification of Hyper-Ammonia Producing Bacteria from Swine Manure Storage Pits. Current Microbiology 2004, 48, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Adeniyi, A.; Bello, I.; Mukaila, T.; Hammed, A. A Review of Microbial Molecular Profiling during Biomass Valorization. Biotechnology and Bioprocess Engineering 2022. [Google Scholar] [CrossRef]
- Rychlik, J.L.; Russell, J.B. Mathematical Estimations of Hyper-Ammonia Producing Ruminal Bacteria and Evidence for Bacterial Antagonism That Decreases Ruminal Ammonia Production 1. 2000, 32, 121–128.
- Slyter, L.L. Influence of Acidosis on Rumen Function. Journal of animal science 1976, 43, 910–929. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Russell, J.B. More Monensin-Sensitive, Ammonia-Producing Bacteria from the Rumen. Applied and environmental microbiology 1989, 55, 1052–1057. [Google Scholar] [CrossRef]
- McDonald, I.W. The Absorption of Ammonia from the Rumen of the Sheep. Biochemical Journal 1948, 42, 584. [Google Scholar] [CrossRef]
- Bryant, M.P. The Characteristics of Strains of Selenomonas Isolated from Bovine Rumen Contents. Journal of Bacteriology 1956, 72, 162–167. [Google Scholar] [CrossRef]
- Bryant, M.P.; Small, N. The Anaerobic Monotrichous Butyric Acid-Producing Curved Rod-Shaped Bacteria of the Rumen. Journal of Bacteriology 1956, 72, 16–21. [Google Scholar] [CrossRef]
- Ward, B.K.; Dufault, R.J.; Hassell, R.; Cutulle, M.A. Affinity of Hyperammonia-Producing Bacteria to Produce Bioammonium/Ammonia Utilizing Five Organic Nitrogen Substrates for Potential Use as an Organic Liquid Fertilizer. ACS omega 2018, 3, 11817–11822. [Google Scholar] [CrossRef]
- Fischer, M. Limiting Factors for the Enzymatic Accessibility of Soybean Protein; Wageningen University and Research, 2006.
- Sun, X.S. Bio-Based Polymers and Composites. In Thermal and Mechanical Properties of Soy Proteins; Elsevier Inc., 2005; pp. 292–326.
- Wang, L.; Liu, K.; Wang, Z.; Bai, X.; Peng, Q.; Jin, L. Bacterial Community Diversity Associated with Different Utilization Efficiencies of Nitrogen in the Gastrointestinal Tract of Goats. Frontiers in Microbiology 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Thorpe, J.; Beal, J.D. Vegetable Protein Meals and the Effects of Enzymes. In Enzymes in farm animal nutrition; Bedford, M.R., Partridge, G.G., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 125–143. [Google Scholar]
- Bedford, M.R. Exogenous Enzymes in Monogastric Nutrition - Their Current Value and Future Benefits. Animal Feed Science and Technology 2000, 86, 1–13. [Google Scholar] [CrossRef]
- Xue, P.; Sun, N.; Li, Y.; Cheng, S.; Lin, S. Targeted Regulation of Hygroscopicity of Soybean Antioxidant Pentapeptide Powder by Zinc Ions Binding to the Moisture Absorption Sites. Food Chemistry 2018, 242, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Huang, Y.; Islam, S.; Fan, B.; Tong, L.; Wang, F. Influence of the Degree of Hydrolysis on Functional Properties and Antioxidant Activity of Enzymatic Soybean Protein Hydrolysates. Molecules 2022, 27, 6110. [Google Scholar] [CrossRef]
- Marsman, G.J.P.; Gruppen, H.; Mul, A.J.; Voragen, A.G.J. In Vitro Accessibility of Untreated, Toasted, and Extruded Soybean Meals for Proteases and Carbohydrases. Journal of Agricultural and Food Chemistry 1997, 45, 4088–4095. [Google Scholar] [CrossRef]
- Lallés, J.P.; Huet, A.; Quillien, L.; Plumb, G.W.; Mills, E.N.C.; Morgen, M.R.A.; Toullec, R. Duodenal Passage of Immunoreactive Glycinin and B−conglycinin from Soya Bean in Preruminant Calves. In Recent advances of research in antinutritional factors in legume seeds and rapeseed; Jansman, A.J.M., Hill, G.D., Huisman, J., van der Poel, A.F.B., Eds.; Wageningen Press: Wageningen, 1998; pp. 255–258. [Google Scholar]
- Merz, M.; Eisele, T.; Berends, P.; Appel, D.; Rabe, S.; Blank, I.; Stressler, T.; Fischer, L. Flavourzyme, an Enzyme Preparation with Industrial Relevance: Automated Nine-Step Purification and Partial Characterization of Eight Enzymes. Journal of Agricultural and Food Chemistry 2015, 63, 5682–5693. [Google Scholar] [CrossRef] [PubMed]
- Nchienzia, H.A.; Morawicki, R.O.; Gadang, V.P. Enzymatic Hydrolysis of Poultry Meal with Endo- and Exopeptidases. Poultry Science 2010, 89, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Clemente, A.; Vioque, J.; Sánchez-Vioque, R.; Pedroche, J.; Bautista, J.; Millán, F. Protein Quality of Chickpea (Cicer Arietinum L.) Protein Hydrolysates. Food Chemistry 1999, 67, 269–274. [Google Scholar] [CrossRef]
- Chae, H.J.; In, M.J.; Kim, M.H. Process Development for the Enzymatic Hydrolysis of Food Protein: Effects of Pre-Treatment and Post-Treatments on Degree of Hydrolysis and Other Product Characteristics. Biotechnology and Bioprocess Engineering 1998, 3, 35–39. [Google Scholar] [CrossRef]
- Kamnerdpetch, C.; Weiss, M.; Kasper, C.; Scheper, T. An Improvement of Potato Pulp Protein Hydrolyzation Process by the Combination of Protease Enzyme Systems. Enzyme and Microbial Technology 2007, 40, 508–514. [Google Scholar] [CrossRef]
- Nchienzia, H.A.; Morawicki, R.O.; Gadang, V.P. Enzymatic Hydrolysis of Poultry Meal with Endo- and Exopeptidases. Poultry Science 2010, 89, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Hunsakul, K.; Laokuldilok, T.; Sakdatorn, V.; Klangpetch, W.; Brennan, C.S.; Utama-ang, N. Optimization of Enzymatic Hydrolysis by Alcalase and Flavourzyme to Enhance the Antioxidant Properties of Jasmine Rice Bran Protein Hydrolysate. Scientific Reports 2022, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Bu, G.; Chen, F. The Influence of Composite Enzymatic Hydrolysis on the Antigenicity of β-Conglycinin in Soy Protein Hydrolysates. Journal of Food Biochemistry 2018, 42, 1–8. [Google Scholar] [CrossRef]
- Paster, B.J.; Russell, J.B.; Yang, C.M.J.; Chow, J.M.; Woese, C.R.; Tanner, R. Phylogeny of the Ammonia-Producing Ruminal Bacteria Peptostreptococcus Anaerobius, Clostridium Sticklandii, and Clostridium Aminophilum Sp. Nov. International Journal of Systematic and Evolutionary Microbiology 1993, 43, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Balows, A.; Trüper, H.G.; Dworkin, M.; Harder, W.; Schleifer, K.-H. The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Springer, 1992; Vol. 3;
- Gano, J.M. Amino Acid-Fermenting Bacteria from the Rumen of Dairy Cattle-Enrichment, Isolation, Characterization, and Interaction with Entodinium Caudatum. PhD Thesis, The Ohio State University, 2013.
- Chen, G.J.; Russell, J.B. Fermentation of Peptides and Amino Acids by a Monensin-Sensitive Ruminal Peptostreptococcus. Applied and Environmental Microbiology 1988, 54, 2742–2749. [Google Scholar] [CrossRef] [PubMed]
- Attwood, G.T.; Klieve, A.V.; Ouwerkerk, D.; Patel, B.K.C. Ammonia-Hyperproducing Bacteria from New Zealand Ruminants. Applied and Environmental Microbiology 1998, 64, 1796–1804. [Google Scholar] [CrossRef]
- Newbold, C.J.; Wallace, R.J.; Eschenlauer, S.C.P.; Mckain, N.; Walker, N.D.; Mcewan, N.R.; Newbold, C.J. Ammonia Production by Ruminal Microorganisms and Enumeration, Isolation, and Characterization of Bacteria Capable of Growth on Peptides and Amino Acids from the Sheep Rumen Ammonia Production by Ruminal Microorganisms and Enumeration, Isolation, and C. 2002. [CrossRef]
- Kuiken, K.A.; Lyman, C.M.; Bradford, M.; Trant, M.; Dieterich, S. Essential Amino Acid Composition of Soy Bean Meals Prepared from Twenty Strains of Soy Beans. Journal of Biological Chemistry 1949, 177, 29–36. [Google Scholar] [CrossRef]
- Bello, I.; Adeniyi, A.; Mukaila, T.; Hammed, A. Optimization of Soybean Protein Extraction with Ammonium Hydroxide (NH4OH) Using Response Surface Methodology. Foods 2023, 12, 1515. [Google Scholar] [CrossRef]
- Whitehead, T.R.; Cotta, M.A. Isolation and Identification of Hyper-Ammonia Producing Bacteria from Swine Manure Storage Pits. 2004, 48, 20–26.
- Attwood, G.T.; Klieve, A.V.; Ouwerkerk, D.; Patel, B.K. Ammonia-Hyperproducing Bacteria from New Zealand Ruminants. Applied and environmental microbiology 1998, 64, 1796–1804. [Google Scholar] [CrossRef]
- Bladen, H.A.; Bryant, M.P.; Doetsch, R.N. A Study of Bacterial Species from the Rumen Which Produce Ammonia from Protein Hydrolyzate. Applied Microbiology 1961, 9, 175–180. [Google Scholar] [CrossRef]
- Chen, G.J.; Russell, J.B. Effect of Monensin and a Protonophore on Protein Degradation, Peptide Accumulation, and Deamination by Mixed Ruminal Microorganisms in Vitro. Journal of Animal Science 1991, 69, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Al, R.E.T. Effect of Carbohydrate Limitation on Degradation and Utilization of Casein by Mixed Rumen Bacteria. Journal of Dairy Science 1983, 66, 763–775. [Google Scholar] [CrossRef]
Figure 1.
Haber-Bosch process flow chat.
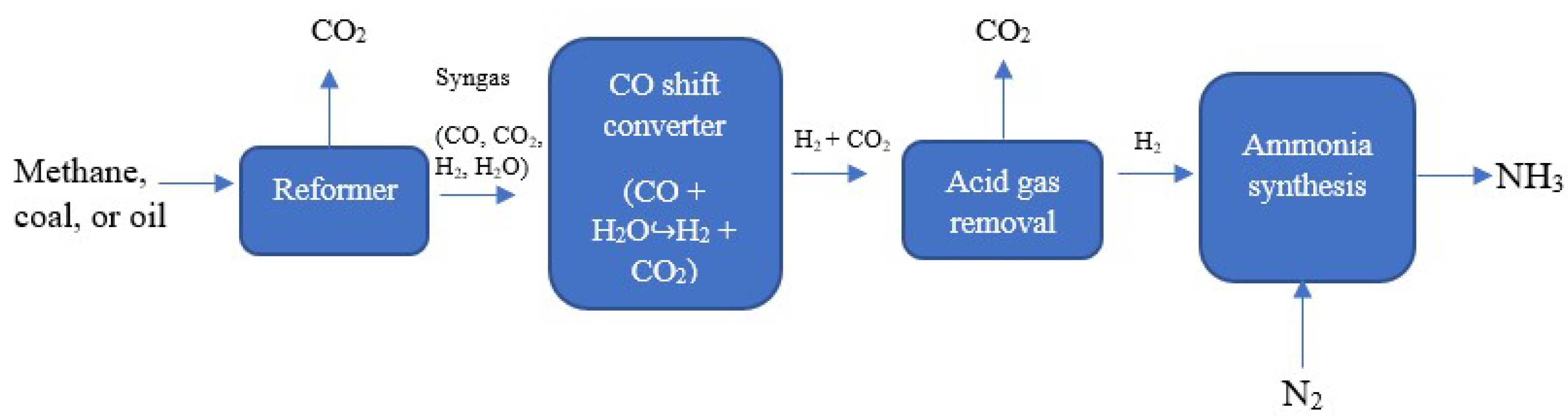
Figure 2.
Regional Ammonia Production Capacities in 2020.
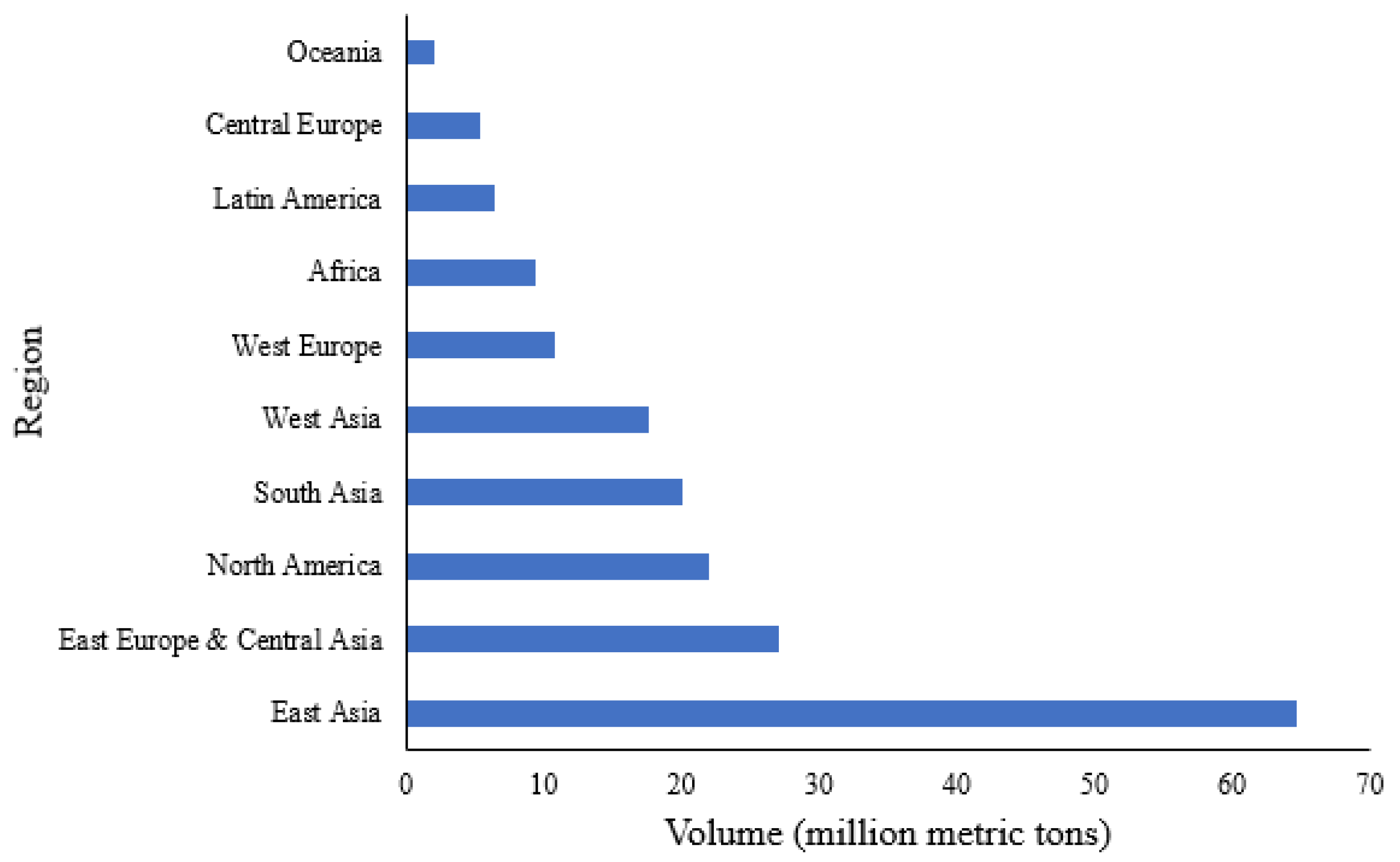
Figure 3.
Three groups of nitrogen-fixing microorganisms .
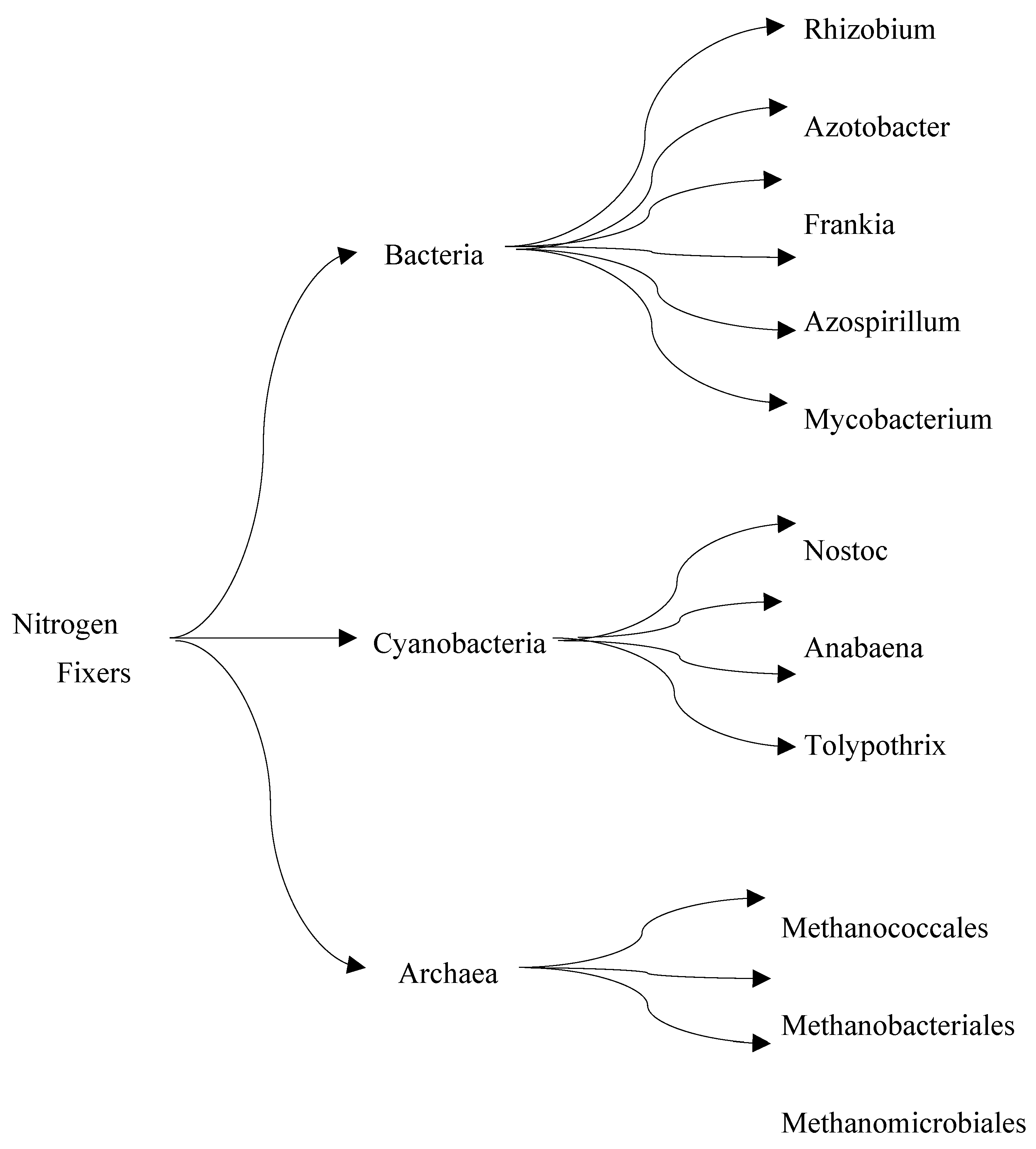
Figure 4.
Conceptual bioprocess flow for biological ammonia production.
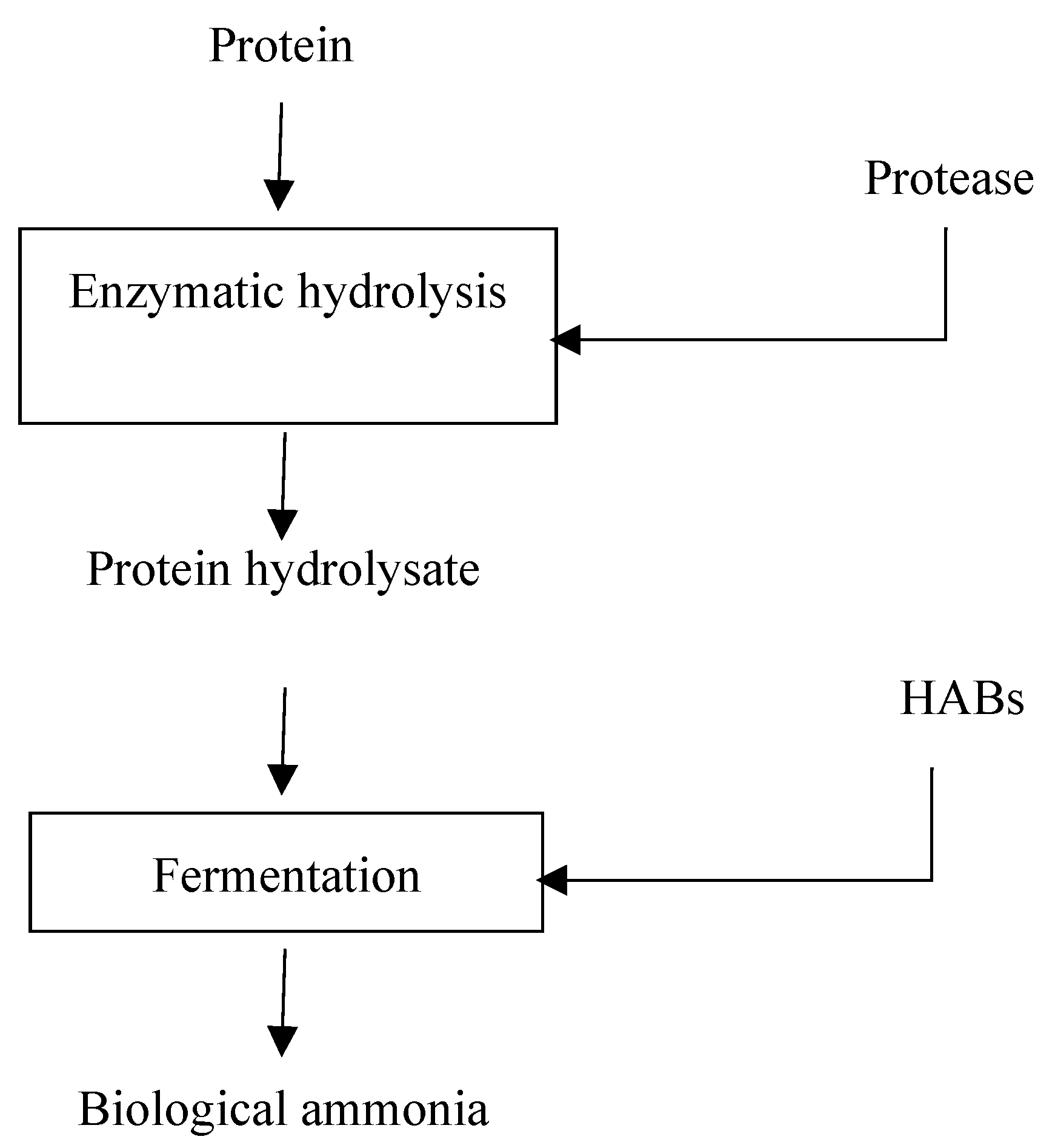

Table 1.
Summary of metabolic engineering approaches for biological ammonia production.
| Approach | Description | Host | Substrates | Ref |
| Gene knockout | Deletion of CodY gene which regulates genes:
|
Bacillus subtilis | Amino acid | [45]. |
| Gene knockout | Deletion of gene BkdB which helps in the biosynthesis of branched chain fatty acids | Bacillus subtilis | Amino acid | [45]. |
| Gene overexpression | Over expression of proteins leuDH, and two-keto-acid decarboxylase which respectively converts amino acids to important metabolic intermediates and increases the availability of metabolic precursors for ammonia production | Bacillus subtilis | Amino acid | [45]. |
| Gene knockout | Deletion of genes glnA and gdhA which aids ammonia assimilation | Eschericia coli | Amino acid | [46]. |
| Gene knockout | Deletion of ptsG (glucose transporter gene) and deletion of phosphoenol pyruvate (glucose transporter) | Eschericia coli | Soybean residue and food waste | [48] |
| Cell surface engineering | HcLAAO (L-amino acid oxidase) display on yeast cell surface by gene insertion | Yeast cells | Amino acids from soybean residue | [47] |
| Cell surface engineering | Glutaminase gene (Ybas) display on yeast cell surface by gene insertion. | Yeast cells | Soybean residue and glutamine | [51]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



