
Submitted:
20 April 2023
Posted:
21 April 2023
You are already at the latest version
Abstract
Nitrogen (N) strongly affects plant growth and metabolism. For shikonin biosynthesis, although the ammonium toxicity phenomenon has been reported, the effects of nitrogen on the shikonin synthesis remains obscure. In this study, we carried out four different concentrations of NH4+ treatments on Arnebia euchroma hairy roots (AEHR) to clarify the influence of NH4+ on the growth and shikonin accumulation in A. euchroma and the possible mechanism. The results showed that compared with the 0% NH4+ treatment (only nitrate as nitrogen source), 10% NH4+ treatment increased the fresh weight and the dry weight of AEHR, and promoted the synthesis of shikonins; 20% NH4+ treatment started to show the inhibition effects on growth and shikonin accumulation of AEHR and 30% NH4+ treatment exhibited the strongest inhibition effects. With increased percentage of NH4+, AEHRs became shorter and thicker with more branches. To further elucidate the mechanism, we analyzed the time-course of nitrogen assimilation, gene expression level of key enzymes involved in the shikonin biosynthesis pathway, and contents of various endogenous hormones under NH4+ toxicity condition. Results indicated that auxin and cytokinin might regulate the growth and architecture of AEHR under NH4+ toxicity, and jasmonate level was reduced along with the inhibition of shikonin biosynthesis. This first comprehensive investigation into the effects of ammonium/nitrate ratio on shikonin biosynthesis not only provided valuable data for optimizing the in vitro culture and shikonin production in A. euchroma, but also suggested potential fertilizer strategies for its cultivation.
Keywords:
ammonium/nitrate ratio
; Arnebia euchroma
; shikonin
; hairy root
; ammonium toxicity
1. Introduction
Arnebia euchroma (Royle) Johnst., a species of Boraginaceae Arnebia, is one of the important resources of traditional Chinese medicine Arnebiae Radix. Its major bioactive molecules are shikonin and its derivatives, a class of compounds in most Boraginaceae plants, which process the activities of anti-inflammatory [1], antiviral [2], antioxidant [3], anti-tumor [4], and the inhibitory effect on topoisomerase I [5]. Shikonin and its derivatives are red naphthoquinone pigments, which can be used in textile dyeing, food and cosmetic industry with a high commercial value [6,7,8]. Tissue culture technology was used to produce shikonin since the 1970s, initially from Lithospermum erythrorhizon [9]. Compared to L. erythrorhizon, A. euchroma is a better source of shikonin-related compounds, harboring red naphthoquinone pigments [10]. While the wild resources of A. euchroma have become endangered because of overexploitation, the artificial cultivation technology is still immature, making the production of shikonin compounds by tissue culture necessary to meet market demand.
For in vitro production of shikonin, White medium without NH4+ was firstly applied in the suspension culture of L. erythrorhizon [11]. It was reported that when NH4+ concentration was increased up to 3% and 30% in the White medium, the synthesis of shikonin in L. erythrorhizon was completely inhibited and more than half of the cell biomass was decreased. After that, medium without NH4+ was widely used for shikonin production via in vitro cell, tissue, and organ cultures of L. erythrorhizon, A. euchroma, and Onosma paniculatum et al. To optimize the culture conditions, the effects of variable medium compositions, abiotic/biotic elicitors, and signal transduction molecules on shikonin synthesis of callus, suspension cells, and hairy root cultures have been widely studied [9,11,12,13,14,15,16]. However, the effects of nitrogen on the shikonin synthesis remains elusive. In addition, there was no mechanism research on NH4+ inhibiting shikonin synthesis published.
Nitrogen is a primary nutrient essential for plant growth. Plants can absorb two types of inorganic nitrogen from soil: nitrate-nitrogen (NO3--N) and ammonium-nitrogen (NH4+-N). Both the absolute nitrogen amount and the ratio of ammonium to nitrate in soil can change nitrogen metabolism in plants. The nitrogen contained organic compounds, nitrogen metabolism affects many physiological metabolic activities by affecting the synthesis of many structural and functional bioactive molecules, which is reflected in the growth, development, and also secondary metabolism of plants. It was generally found that the nitrogen metabolism activity of crop was the strongest under the condition of mixed ammonium nitrate nutrition and appropriate nitrogen application strategy can significantly promote crop growth and effective compounds accumulation. For example, The nitrate reductase (NR) and glutamine synthetase (GS) enzyme activity in Poncirus trifoliata (L.) Raf [17]. seedling root was the strongest under the condition of NH4+:NO3-=50:50, and the total organic acid, total alkaloid, and adenosine amount of Pinellia pedatisecta was the highest under the condition of NH4+:NO3-=75:25 [18].
In addition to being involved in nitrogen metabolism, NO3- can also act as a signaling molecule to regulate many physiological processes, with a significant regulatory effect on the expression of genes related to other metabolic pathways [19,20]. Although some studies have found that NH4+-N may also be a signaling molecule, more studies have verified that excessive NH4+-N can produce significant toxic effects on plants in the following ways: i) absorbing process of NH4+-N will cause environmental acidification and inhibit the absorption of other cations; ii) a large amount of free NH4+ will destroy transmembrane proton gradient and affect cell metabolism; iii) ineffective assimilation to avoid free NH4+ toxicity will consume lots of energy and carbohydrates, which break the carbon and nitrogen metabolic balance and affect respiration and photosynthesis; iv) causing hormone metabolism imbalance, etc.
Wild A. euchroma grows in alpine meadow soil rich in humus. We collected the inter-root soil of A. euchroma and measured the contents of inorganic nitrogen NH4+-N and NO3--N inside, which contained 17.384 mg/kg NH4+-N and 44.486 mg/kg NO3--N (unpublished). The natural high ratio of NH4+ (28%) moved us to explore what NH4+ concentration was favorable for the growth and shikonin accumulation in A. euchroma.
In this study, we investigated the effects of NH4+ on the growth and shikonin synthesis of A. euchroma hairy root (AEHR), as well as the possible mechanisms which were of great importance for the shikonin production in in vitro culture of A. euchroma, and also the fertilizer strategy for farming.
2. Materials and Methods
Material and regents
A. euchroma seeds for aseptic seedling were collected in Xinjiang, China. Agrobacterium Rhizogenes (Strain C58C1) were provided by Resource Center of Chinese Materia Medica, China Academy of Chinese Medical Sciences. AEHR was induced by C58C1 infected cotyledon of sterile plantlets of A. euchroma as described previously [8]. The AEHR was cultured in MS ammonia-free liquid medium (50 mL) for extended culture.
Both acetonitrile and methanol were LC grade, and were supplied from Merck Company. A Water Purification System from Milli-Q (Millipore, Bedford, MA) was used to acquire ultrapure water. The information of standards is shown in Supplemental data (Table S1).
Medium formulation
Based on MS medium, AEHR was treated with four nutrient solutions with different NH4+ concentrations: 0% NH4+, 10% NH4+, 20% NH4+, and 30% NH4+ under the premise of controlling total inorganic nitrogen at 20 mM. To avoid the large differences of other major elements in nutrient solutions with different NH4+ concentrations, the formula of major elements in MS medium is optimized (Table S3). The trace elements, iron salts and organic components in each NH4+ concentration nutrient solution were the same as MS medium. All mediums were adjusted to pH = 5.80.
Experimental design
Long-term treatment for 15d
About 0.05g AEHR with the same growth status cultured in MS ammonia-free solid medium for 11days were separately transferred to 20mL different mediums with 0% ,10%, 20%, and 30% NH4+. AEHRs treated for 10d, 13d, and 15d were taken out for determination of fresh weight, dry weight and content of shikonin compounds. AEHR treated for 15d was also used for determination of root morphological indexes.
Short-term treatment for 48h
AEHR (0.2 g) with the same growth status cultured in MS ammonia-free solid medium for 11days were transferred to MS ammonia-free liquid medium (20 mL). After 8 days, removed original medium and added 20 mL different mediums with 0%, 10%, 20%, and 30% NH4+ subsequently. AEHR was treated with different concentrations of NH4+for 0h, 6h, 12h, 24h, 36h, 48h and harvested. One-half of AEHR was used to determine the hormones content (fresh), quarter of AEHR was used to determine the expression level of key enzyme genes in shikonin biosynthesis (fresh), the rest of AEHR was used to determine the content of shikonin compounds. Short-term treatment was repeated, and AEHR (0.10 g) was precisely weighed for the determination of nitrate nitrogen content and ammonia nitrogen content (fresh).
Mass weight measurement and root morphology analysis
Dry the medium on the AEHR surface with absorbent paper, weigh the fresh weight, dry weight at 40°C to constant weight, and calculate drying rate = (dry weight/fresh weight) × 100%. Carefully separated AEHR with tweezers, collected images under the root scanner, and calculated the number of branches of root length, root diameter, root surface area, and root volume with WinRHIZO software (Regant, Canada). 3 biological replicates were taken from each treatment, and 3 complete AEHRs were taken from each biological replicate to get those root morphological indexes.
Quantification of shikonin compounds
UPLC was carried out with a Waters Acquity UPLC-PDA system equipped with a Waters HSS T3 column (2.1 mm × 100 mm, 1.8 μm) with absorbance at 516 nm. The column temperature was set at 40°C. For shikonin content determination in hairy roots, about 20 mg of lyophilized hairy root was extracted by ultrasonication in 1 mL of methanol. The mobile phase comprised acetonitrile (A) and water (0.1% formic acid, B) at 0.5 mL min-1 with the following gradient program (0~2.0 min, 10.0%~55.0% A; 2.0~2.5 min, 55.0%~59.0% A; 2.5~7.0 min, 59.0%~65.0% A; 7.0~8.0 min, 65.0%~65.6% A; 8.0~14.0 min, 65.6%~79.0% A; 14.0~14.1 min, 79.0%~98.0% A; 14.1~16.0 min, 98.0%~98.0% A; 16.0~16.1 min, 98.0%~10.0% A; 16.1~18.0 min, 10%~10% A).
Determination of NO3-N and NH2-N contents
Contents of NH2-N were detected by using Plant Ammonia Nitrogen Assay Kit (Youxuan BIC, Shanghai) and the NO3-N were measured with Plant Nitrate Nitrogen Assay Kit (Youxuan BIC, Shanghai).
RNA extraction and qRT-PCR analysis
Total RNA of AEHR was extracted using GK reagent (Huayueyang, Beijing) following the manufacturer’s instructions. After treatment with DNases, samples were fractionated on agarose gel to analyze RNA integrity and genomic DNA contamination. First-strand cDNAs were synthesized with the Primer Script First Strand cDNA Synthesis Kit with random primers and oligo (dT) at the same time (TaKaRa). qRT-PCR was performed using the Power SYBR Green PCR Master Mix (Applied Biosystems) and an Applied Biosystems 7500 real-time instrument [21]. The primers used are listed in Supplemental Table S2. The 18S rRNA was used as the endogenous control to normalize expression data [22]. At least three independent experiments were performed for each analysis.
Sample preparation for hormones determination
Powder samples ground in liquid nitrogen was extracted with isopropanol/hydrochloric acid (10 mL) extraction buffer and of 1 µg/mL internal standard solution (8 µL) were added, and the sample was shaken at 4°C for 30 min. 20 mL dichloromethane was added to the sample and shaked at 4°C for another 30 min. Centrifuged at 13000 r/min for 5 min at 4°C, the lower organic phase was taken and dried with nitrogen away from the light. Redissolved with 400 µL methanol (0.1% formic acid), and tested by 0.22 µm filter membrane before processed to the UPLC-MS/MS.
Quantification of hormones in AEHR
UPLC-MS/MS was carried out with a QTRAP 6500 mass spectrometer (ABSCIEX), connecting with an Acquity UPLC system (Waters). The UPLC system equipped with poroshell 120 SB-C18(2.1 mm×150 mm,2.7 μm) reversed phase column. The column temperature was set at 40°C. The mobile phase comprised methanol (A) and water (0.1% formic acid, B) at 0.5 mL min-1 with the following gradient program (0~1.0 min, 20% A; 1.0~9.0 min, 20%~80% A; 9.0~10.0 min, 80% A; 10.0~10.1 min, 80%~20% A; 10.1~15.0 min, 20% A). The mass spectrometric parameters: curtain gas, 15 psi; spray voltage, 4500 V; atomizer pressure, 65 psi; auxiliary gas pressure, 70 psi; atomization temperature, 400°C.
Statistical analysis
The statistical analysis of the obtained results was evaluated by One-Way ANOVA using LSD test at a significance level of p < 0.05. All statistical tests were performed using the SPSS 22.0 (Chicago, IL, USA).
3. Results
3.1. Growth of AEHR in Response to Ammonium
Medium components in this study was based on MS medium with two types of inorganic nitrogen NH4+ and NO3- (Table S3). When A. euchroma hairy roots (AEHRs) were cultured in mediums with consistent total 20 mM nitrogen content but different percentage of NH4+ (0%, 10%, 20%, 30%), the AEHRs showed obvious differences in root characteristics including surface color (representing the contents of shikonin, Figure 1).
AEHRs were harvested after being cultured for 10d, 13d, 15d. The fresh weight and dry weight of each were measured (Figure 2). The amounts of biomass significantly increased from 0d to 15d and the trend of changes varied with the NH4+ concentrations. Compared to 0% NH4+, 10% NH4+ highly increased both fresh weight (1.58-1.97 folds) and dry weight (1.19-1.67 folds) of AEHR since cultured for 10 days. 20% NH4+ also significantly increased the fresh weight of AEHR, but the promotion effect was relatively weaker than 10% NH4+. There was only a promotion at 10d in dry matter accumulation. When the NH4+ concentration increased to 30%, it showed a stronger inhibition effect on dry matter accumulation. Significant decrease in dry weight at 13d and 15d were observed, even though there was a slight increase in fresh weight. It was interesting to notice that all the drying rate of 10% NH4+ 20% NH4+ and 30% NH4+ were significantly lower than 0% NH4+, in spite of various effects on biomass accumulation. Overall, with increasing ammonium concentration, the effect of NH4+ concentration on the growth of AEHR showed a promotion (especially 10%) followed by inhibition (30%).
3.2. Accumulation of Shikonin in AEHR in Response to Ammonium
As typical pigments, the red color of the roots could roughly represent the accumulation of shikonin and its derivatives. It can be observed that the roots treated with 10%, 20% and 30% NH4+ turned red at 10d, 13d, and 15d respectively after transfer to fresh medium (Figure 1), while under 0% NH4+ condition AEHR stayed red at 5d and 8d. The results observed indicated that NH4+ concentration affected the accumulation process and content of shikonin. The contents of 6 main shikonin compounds in AEHR treated with different concentrations of NH4+ for 10d, 13d and 15d were quantified by UPLC (Figure 3). After 10 days treatment, except deoxyshikonin (DS), other 5 shikonin derivatives in AEHR under 10% NH4+ treatment were notably less than those in 0% NH4+ and closed to zero or non-detected in 20% NH4+ and 30% NH4+. But the concentration of DS, the precursor substance of other shikonin compounds, of 10% NH4+ and 0% NH4+ was comparable. After cultured to 13 days, AEHR under 10% NH4+ treatment reached the highest concentration of β-hydroxyisovalerylshikonin, acetylshikonin and β,β’-dimethacrylicalkannin (HIVS, AS and DAA), which was respectively 18.33%, 77.34%, 71.99% higher than 0% NH4+ and 162.85%, 80.99%, 82.54% higher than 20% NH4+. AEHR treated with 20% NH4+ began to accumulate shikonin components especially for DS, while accumulation of all kinds shikonin derivatives in 30% NH4+ were still strongly suppressed except for a visible increase in DS.
When processing to the end of 15 days, both 10% NH4+ and 20% NH4+ treatment promoted the synthesis of shikonin compounds in AEHR compared with 0% NH4+ treatment. In terms of the summation of 6 shikonin derivatives, 10% NH4+ treatment had the strongest promotion effect reaching 8.59 mg/g dry weight, which was 2.17-fold than 0% NH4+ treatment. The number of 20% NH4+ was also significantly higher than that of 0% NH4+ (p<0.05), which was 0.90-fold than 0% NH4+ (20% NH4+ reached 5.16 mg/g). For roots under 30% NH4+ treatment, it was kept at very low level (less than 1 mg/g) until the 15d, which indicated a fierce inhibition of shikonin biosynthesis.
3.3. Development and Morphology of AEHR in Reponse to Ammonium
The WinRHIZO root-scanning system was used to investigate the morphology characters of hairy roots harvested at 15d(Figure 4a). The AEHR treated with 0% NH4+ had the longest length (99.73 cm) and the smallest diameter (0.28 mm). With the increase of NH4+ concentration, AEHR gradually became shorter and thicker(Figure 4b, 4c). It was also reflected from the root length with the diameter range from 0.00 mm to 0.25 mm (L1) and its percentage of total root length (PTL). They were 71.57±5.62 cm, 44.20±2.59, 32.60±7.26 and 25.68±9.04 cm in length (L1) and accounted for 71.76%, 59.82%, 47.05%, and 37.12% in PTL respectively under condition of 0% NH4+, 10% NH4+, 20% NH4+, and 30% NH4+. Meanwhile, for the thicker roots, of which diameter range between 0.25 mm~0.50 mm (L2), 0.50 mm~1.00 mm (L3) and 1.00 mm~1.50 mm (L4) PTL of different NH4+ concentration group showed almost the same trends, which was 0% NH4+<10% NH4+< 20% NH4+<30% NH4+(Figure 4h, 4i).
Although the root length varied, the root surface area (SA) and root projection area (PA) of AEHR showed no significant difference under different NH4+ treatments. What’s more, the root volume (V) increased with the increase of NH4+concentration, and 30% NH4+ treatment was significantly higher than 0% NH4+ (p<0.05). The branch number (B) also increased with the increase of NH4+concentration, and the branches of AEHR of 20% NH4+ were the most and significantly higher than 0% NH4+ (p<0.05,Figure 4d-4g).
3.4. Nitrogen Assimilation in AEHR under Short-term Ammonium Treatment
In general, NO3-N and NH2-N content in plants is directly related to the level of NO3-N and NH4+-N in the environment respectively. Since NO3- absorption consumes more energy than NH4+ absorption, plants tend to absorb NH4+. Studies have shown that the presence of NH4+ inhibited NO3- absorption [23,24]. Meanwhile, in order to avoid free ammonium toxicity, they rapidly assimilate NH4+-N into NH2-N. In order to investigate the possible mechanism of NH4+ concentration affecting AEHR growth and shikonin compounds production, we have conducted a short-term ammonium treatment for 48h. The continuous contents of NO3-N and NH2-N in AEHR within 48h after treated with different concentrations of NH4+ were measured (Figure 5). Under 0% NH4+ treatment within 48h, both NO3-N and NH2-N contents in AEHR fluctuated slightly, which indicated that nitrogen metabolism was relatively balanced. Compared with 0% NH4+ treatment, all of 3 (10%, 20% and 30%) NH4+ treatments inhibited NO3-N accumulation in AEHR, but increased NH2-N content in AEHR. Under 10% NH4+ treatment, the content of NH2-N in AEHR increased sharply within 12h and then maintained stable with a slight decline. For 20% NH4+ and 30% NH4+, the content of NH2-N increased sharply until 24h and then maintained a stable high level of around 4000 μg/g, which might cause normal nitrogen metabolism and ammonium toxicity to inhibit the growth and shikonin synthesis of AEHR.
3.5. Shikonin Biosynthesis in AEHR under Short-term Ammonium Treatment
The contents of 6 main shikonin derivatives in AEHR under short-term treatments of different NH4+ concentrations were measured by UPLC (Figure 6). Compared with AEHR at 0h (before being transferred to a fresh medium), contents of shikonin derivatives in AEHR decreased with all NH4+ concentrations. This decrease is likely because it takes time for hairy roots to adapt to the fresh culture medium and they prefer to nutrient growth rather than secondary production at the early stage of the growth period. Another reason is that the shikonin derivatives is secreted to the root surface and dispersed into the fluid medium. Compared with 0% NH4+ treatment, 10% NH4+ treatment showed significantly increased contents of HIVS, AOIVA, and IBS at 36h, also HIVS and IBS at 48h (p<0.05). Meanwhile, the inhibition effects of 20% NH4+ and 30% NH4+ treatment were not that obvious except for DS and AS, which displayed a significant decrease (p<0.05) compare to 0% NH4+. While compared with 10% NH4+ treatment, after treated for 48h, the contents of all the 6 shikonin derivatives decreased at 20% NH4+ and 30% NH4+treatment, and there were significant differences in HIVS, IBS, DS and AS (p<0.05).
3.6. Gene Expression of Key Enzymes in Shikonin Biosynthesis Pathway
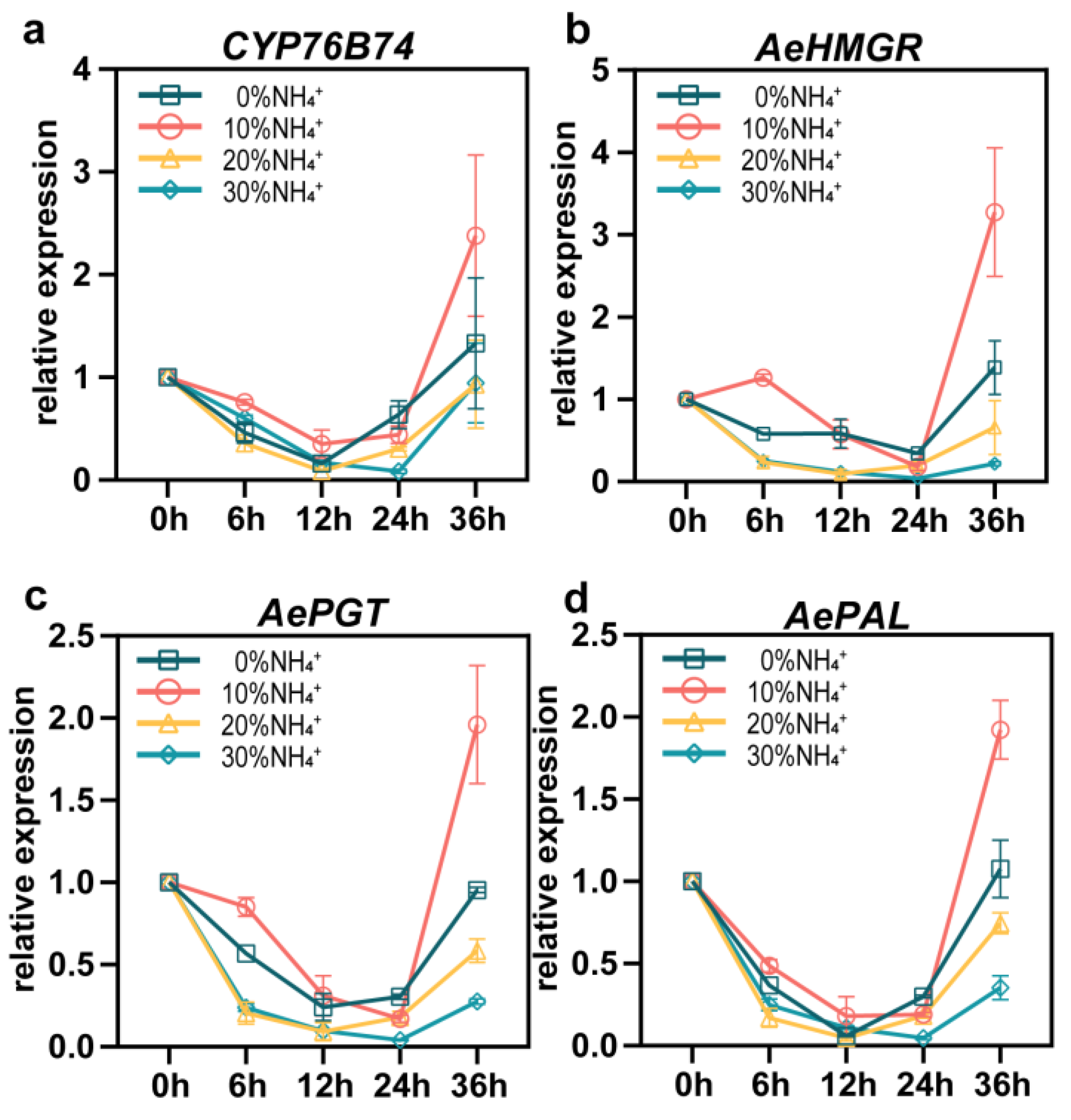
The biosynthesis pathway of shikonin has been well dissected [8,25,26]. AeHMGR, AePAL, AePGT, and CYP76B74 were proven to be key enzymes regulating the accumulation of shikonin in previous studies [8,26]. To reveal the effect of NH4+ on shikonin biosynthesis at the gene expression level, the time-course expression levels of them treated with the different NH4+ concentration were quantified with qRT-PCR (Figure 7). Compared with 0% NH4, 10% NH4+ increased the expression levels of AeHMGR, AePAL, AePGT and CYP76B74 after 6h treatment, while the relative expression levels of the AeHMGR, AePAL, AePGT were down-regulated by 20% NH4+ and 30% NH4+ treatments. After treated for 36h, expression levels of them appeared significant difference. The expression level of AeHMGR, AePAL, AePGT and CYP76B74 treated with 10% NH4+ were up-regulated about 1.36-fold, 0.79-fold, 1.05-fold and 0.78-fold, respectively, more than 0% NH4+. These corresponding genes treated with 30% NH4+ were down-regulated 0.84-fold, 0.67-fold, 0.71-fold and 0.29-fold, respectively. The 20% NH4+ also showed a down-regulated gene expression of AeHMGR, AePAL, AePGT, and CYP76B74 compare to 0% NH4, but a higher gene expression level of AeHMGR, AePAL, AePGT but CYP76B74 than 30% NH4+. The consistent results of gene expression and metabolite accumulation under different NH4+ conditions indicated that 10% and 30% NH4+ influenced the accumulation of shikonin compounds by regulating the gene expression involved in the biosynthesis pathway.
3.7. Hormone Metabolism of AEHR under NH4+ Toxicity
NH4+ toxicity is common in plants but the threshold for responding to it varies between species [27]. Studies indicate that NH4+ toxicity is associated with hormonal disruption in plants [28]. As a plant sensitive to the toxicity of NH4+, 8 kinds of hormone in AEHR treated with 0% NH4+ and 30% NH4+ for 48h were determined in this study, to profile the possible hormonal disruption of NH4+ toxicity in AEHR (Figure 6). Compared with 0% NH4+, 30% NH4+ treatment promoted the synthesis of endogenous cytokinin ZT and TZR, and auxin IAA, but not IPA. 30% NH4+treatment had a stronger effect on cytokinin than auxin, and the contents of ZT were significantly higher than 0% NH4+ at 12h, 36h, and 48h (p<0.05), and relevant contents of TZR were extremely significantly higher (p<0.01). For endogenous auxin, there was no significant difference and consistent tendency between 0% NH4+ and 30% NH4+ treatment within 48h. The increased cytokinin might be responsible for shorter root length and thicker diameter of AEHR under 30% NH4+ treatment.
Figure 8.
Impact of 30% NH4+ treatment on auxin and cytokinin in AEHR within 48h. (a) IAA, Auxin; (b) IPA, Indolepropionic acid; (c) ZT, Zeatin; (d) TZR, trans-Zeatin-riboside; (e) GA4, Gibberellin4; (f) ABA, Abscisic acid; (g) MeJA, (h) SA. Vertical bars represent mean ± SE (n = 3).
Figure 8.
Impact of 30% NH4+ treatment on auxin and cytokinin in AEHR within 48h. (a) IAA, Auxin; (b) IPA, Indolepropionic acid; (c) ZT, Zeatin; (d) TZR, trans-Zeatin-riboside; (e) GA4, Gibberellin4; (f) ABA, Abscisic acid; (g) MeJA, (h) SA. Vertical bars represent mean ± SE (n = 3).
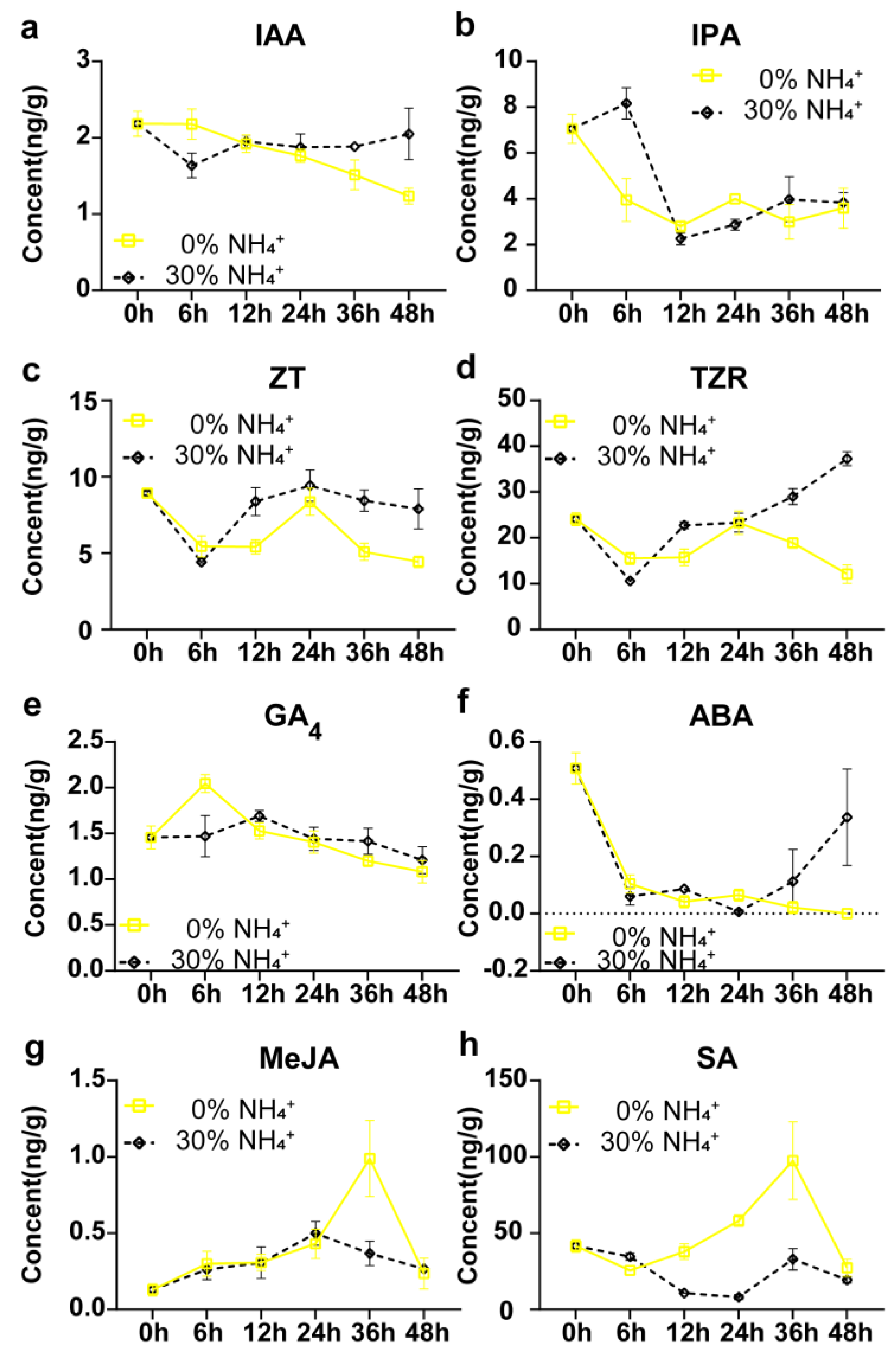
Previous studies demonstrated that different exogenous hormones have various effects on shikonin accumulation. For instance, GA [29] and ABA have significantly negative impacts on shikonin biosynthesis, while MeJA and SA [12,16,30,31]can effectively promote it. In our study, the content of endogenous GA4 in AEHR under 30% NH4+ treatment did not change significantly within 48h, suggesting that GA may not be involved in the regulation process of NH4+ toxicity in AEHR. As for ABA, the contents in 30% NH4+ treatment decreased rapidly and kept at a low and steady level till 24h as well as 0% NH4+, which might be related to the adaptation of the hairy roots to the new medium. Then, it began to rise after being treated for 36h, and reached a high level of 0.11 ng/g at 48h. It was contrary to the trend of shikonin accumulation and the expression of key enzymes involved in shikonin biosynthesis pathway, indicating ABA as a negative regulator.
The contents of endogenous MeJA grew in pace until 24h after treated with 0% and 30% NH4+, then the latter decreased and the former (0% NH4+) reached the highest level at 36h before decreasing. The trends of SA treated with 0% NH4+ was similar with MeJA, keep increasing until 36h then decrease, while it decreased slowly before 24h and followed a little fluctuation with 30% NH4+. The content of SA in AERH treated with 0% NH4+ at 12h, 24h and 36h were significantly (p<0.01) higher than 30% NH4+. These were consistent with the tendency of shikonin accumulation and relative gene expression. The inhibition of high concentration NH4+ on shikonin production was likely associated with the decrease of endogenous MeJA and SA, especially with SA.
4. Discussion and Conclusion
4.1. Arnebiae Euchroma is an Ammonium-Sensitive Species
Nitrogen is an essential element in the growth and development of plants. It was discovered in 1882 that ammonium can be toxic to plants, leading to a variety of issues such as stunted rhizomes, shortened roots, yellowing leaves, and even death. Different species and varieties of plants have varying levels of tolerance to ammonium. In this study, when the concentration of ammonium was raised to 20% NH4+ (4 mM), inhibition began to occur. Both the growth of AEHR and shikonin production could be strongly inhibited by 30% NH4+ (6 mM). These results demonstrated A. euchroma as an ammonium-sensitive type, exhibiting similar ammonium sensitivity to other ammonium-sensitive species which would display ammonium toxicity when exposed to concentrations of less than 10 mM, such as Arabidopsis, citrus, wheat, and L. erythrorhizon. Arabidopsis is a highly ammonium-sensitive plant, 1 mM of ammonium was found to impede the growth of the taproot, as well as reduce the expansion of cells and cell yield [32]. Furthermore, ammonium was also found to inhibit the geotropism of roots. Citrus plants, exposed to high ammonium (8 mM), exhibited growth inhibition, metabolic disorder, decreased biomass, abnormal morphology, and a reduced growth rate [33]. As for wheat, the plant biomass, total root length, surface area, and root volume were significantly reduced by 5 mM ammonium [34]. Cell cultures of L. erythrorhizon, another closely related shikonin-producing species with A. euchroma, were reported to be ammonium-sensitive. When NH4+ concentration was increased up to 3% and 30% in the White medium, the synthesis of shikonin was completely inhibited and more than half of the cell biomass was decreased respectively. Similar results were obtained in the present study, both the growth and shikonin production of AEHR increased as the NH4+ proportion increased to 20% and 30%, but not as extremely sensitive as L. erythrorhizon. Different from ammonium-sensitive plants, Spartina alterniflora is highly tolerant to ammonium and this can even alleviate salt stress [35,36]. Additionally, ammonium nitrogen has been found to be more beneficial for the growth and nutrient accumulation of Pinus massoniana (Masson’s pine) tissue culture seedlings than nitrate nitrogen. Therefore, it is recommended to apply ammonium nitrogen fertilizer when cultivating Masson’s pines seedlings. The preference of different species may be influenced by the ratio of ammonium+/nitrate - found in their native habitats, and the optimization studies can facilitate the development of fertilization schedules for the cultivation process.
4.2. Compare to the Sole Nitrate Nitrogen source, A. euchroma Prefers an Appropriate Ammonium/Nitrate ratio
For most plants, mixed nitrate and ammonium nitrogen source is superior to ammonium or nitrate nitrogen sources alone in terms of plant growth and chemical component accumulation [34,37,38]. The optimal ratio of ammonium/nitrate for plant growth depends on the plant species, environmental conditions, developmental stage, and the total concentration of supplied nitrogen [39]. Ammonium nitrogen can promote the growth of aboveground parts of coffee plant, but an excessive concentration of it (ammonium/nitrate ratio above 70:30) can decrease the chlorophyll content and inhibit the growth of the aboveground parts. On the other hand, an increase in nitrate nitrogen is beneficial for the growth of roots and underground parts, but it can have an adverse effect on the growth of leaves, stems, and other aboveground parts. As a result, 50:50 ammonium/nitrate ratio leads to higher biomass yield of coffee seedlings [40]. In the present study, it was clarified that 10% NH4+ (2 mM) was beneficial for both growth of AEHR and the accumulation of shikonin compounds compared to ammonium as the sole nitrogen source (0% NH4+), even though A. euchroma was an ammonium-sensitive plant.
We listed the medium used for in vitro cultures of A. erchroma and L. erythorhizon from literature, the concentrations of NH4+ and NO3-, the total content of nitrogen, and the NH4+/NO3- ratio were compared (Table 3). A two-step method is widely used for in-vitro cultures of L. erythorhizon, i.e. biomass is first accumulated on growth medium, and then inoculated on shikonin-production medium for shikonin accumulation. That is because of the different nutrients necessary for two stages of L. erythorhizon cultures, especially nitrogen. For in vitro cultures of A. erchroma, studies were mostly performed in one-step medium. The total content of nitrogen varied from around 10 mM to 60 mM, NH4+/NO3- ratio from 0% to 34.34%, in which most studies used medium without NH4+ to avoid ammonium toxicity. In present study, the optimum ratio of NH4+/NO3- for both growth and shikonin production is 10%, we highly recommended B5 and SH medium for in vitro cultures of A. erchroma, especially for hairy roots.

Table 2.
Mediums used for in vitro cultures of A. erchroma, L. erythorhizon and Onosma paniculatum from literature.
Table 2.
Mediums used for in vitro cultures of A. erchroma, L. erythorhizon and Onosma paniculatum from literature.
| Varieties | Step | Medium | NH4+(mM) | NO3-(mM) | Total content (mM) | NH4+/NO3- ratio | Reference |
| L.erythorhizon.乐suspension乐cell cultures | Two-step for growth | LS | 20.61 | 39.4 | 60.01 | 34.34% | [11] |
| MG-5 | 6.25 | 54.22 | 60.47 | 10.34% | [6] | ||
| Two-step for shikonin production | White | 0 | 3.33 | 3.33 | 0.00% | [11] | |
| M9 | 0 | 6.67 | 6.67 | 0.00% | [11] | ||
| L. erythorhizon. hairy roots | Two-step for growth | B5 | 2.02 | 24.73 | 26.75 | 7.55% | [16,31] |
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [16,31] | |
| A. erchroma suspension cell cultures | One-step | AG-7 | 7 | 18.79 | 25.79 | 27.14% | [41,42] |
| Two-step for growth | AG-7 | 7 | 18.79 | 25.79 | 27.14% | [42] | |
| LS | 20.61 | 39.4 | 60.01 | 34.34% | [43] | ||
| MS | 20.61 | 39.4 | 60.01 | 34.34% | [44,45] | ||
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [42,46,47,48,49,50,51] | |
| M10 | 0 | 10.97 | 10.97 | 0.00% | [43] | ||
| APM | 0 | 17.74 | 17.74 | 0.00% | [44,45] | ||
| A. erchroma hairy roots | Two-step for growth | SH | 2.61 | 24.73 | 27.34 | 9.55% | [51] |
| SH without NH4+ | 0 | 24.73 | 24.73 | 0.00% | [47,50,52] | ||
| MS without NH4+ | 0 | 18.79 | 18.79 | 0.00% | [47,52] | ||
| B5 without NH4+ | 0 | 24.73 | 24.73 | 0.00% | [47] | ||
| 1/2MS without NH4+ | 0 | 9.395 | 9.395 | 0.00% | [46,52] | ||
| LS without NH4+ | 0 | 18.79 | 18.79 | 0.00% | [47] | ||
| MG-5 without NH4+ | 0 | 47.97 | 47.97 | 0.00% | [52] | ||
| MS | 20.61 | 39.4 | 60.01 | 34.34% | [47] | ||
| B5 | 2.02 | 24.73 | 26.75 | 7.55% | [49] | ||
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [47,50] | |
| Onosma paniculatum suspension cell cultures | Two-step for growth | B5 | 2.02 | 24.73 | 26.75 | 7.55% | [53] |
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [53] |
4.3. Auxin and Cytokinin Might Regulate the Growth and Architecture of AEHR under NH4+ Toxicity
Studies have demonstrated that NO3- and NH4+ are involved in hormone regulation of plant root formation and leaf development. Compared with NO3-, NH4+ is more likely to cause imbalance of plant hormone metabolism, which is one of the mechanisms of ammonium toxicity. Under NH4+ toxicity, we found both growth and architecture changed. Compared with the sole nitrate nitrogen source, AEHR in 30% NH4+ treatment was shorter and thicker with more branches. These changes were linked to alterations in hormonal balance. We speculated that it was linked to increased auxin and cytokinin production in roots. Under ammonium toxicity, AEHR in this study showed significant changes in both endogenous auxin (IAA, IPA) and cytokinin (ZT, TZR). Ammonium toxicity increased IPA content at 6 h, and IAA, ZT and TZR contents at 12h, 36h, and 48 h. This was consistent with the results of Yang et al. on Arabidopsis thaliana in a single ammonium nitrogen source using matrix transcriptomes [54]. In the taproot grown in (NH4)2SO4, the nutrient and metabolic imbalance induced by ammonium was partially overcome by the increase of auxin level. Dziewit et al. [55] studied the temporal and spatial distribution of auxin in Arabidopsis thaliana under long-term ammonium toxicity and found that auxin accumulated in leaves and roots, but not in root tips. The apparent auxin pattern in tissues is associated with the developmental adaptation of ammonium-growing plants with short branches and highly branched roots. Ammonium toxicity is known to affect auxin activity, which serves as an important positive regulator of lateral root (LR) development [56]. In the general NH4+ toxicity research, it was suggested that more prolific root branching results from the increased strength of the root tissue as a carbon sink under NH4+ nutrition, which would facilitate more auxin delivery to the root. The increased number of root tips, which has been often observed, could then lead to increased production of cytokinins in ammonium-grown plants. The increased ZT and TZR contents at 12h, 36h, and 48 h were in accord with this theory. The study of NH4⁺ toxicity alleviation by Si also suggested that it was dependent on the increase in trans-ZT content in shoots. In cytokinin-deficient plants, Si did not alleviate NH4⁺ toxicity [57]. However, in some studies, the cytokinin showed the opposite performance. Walch-Liu et al. reported that NH4+ inhibition of plant growth was associated with a sharp decline in cytokinin concentration [58].
4.4. Shikonin Synthesis of AEHR under NH4+ Toxicity and Its Possible Hormones Regulation Mechanism
Plant hormones, especially MeJA and SA, play an important role in regulating plant secondary metabolism and environmental response. Apart from the effect on growth, results of shikonin biosynthesis regulation were various according to different forms of auxin or cytokinin and different plant species. It was reported that IAA could positively regulate shikonin accumulation in L. erythorhizon, while IBA inhibited it in A. euchroma cells. Synthetic auxin 2,4-D and NAA had consistent negative regulatory effects on the shikonin biosynthesis in cells of L. erythorhizon, A. euchroma, and O. paniculatum [59,60]. Among the cytokinin, KT and 6-BA promoted the shikonin biosynthesis of L. erythorhizon callus but showed inhibition in A. euchroma cells [59,61].
Studies have shown a consistent promotional effect for shikonin biosynthesis in L. erythorhizon, A. euchroma [21], L. officinale [62], and O. paniculatum [63]. Exogenous MeJA promoted the accumulation of shikonin derivatives, shikonofuran derivatives, and rosmarinic acid in the shikonin-proficiency cells, and caused a rapid and massive increase in the expression of genes involved in the shikonin biosynthesis pathway, such as HMGR and PGT [21].Zhao et al. [16,31] overexpressed LeMYB1 in hairy roots of L. erythorhizon. The content of shikonin compounds increased significantly, and at the same time expression of LeMYB1 showed the same positive response to JA. Additionally, the endogenous JA content was significantly increased when the L. erythorhizon cell was transferred from the B5 medium (for the growth step) to the M9 medium (for the shikonin production step)[64]. While the responses to the exogenous SA were not as consistent as JA. Kumar et al. reported that exogenous SA can inhibit the synthesis of shikonin in A. euchroma cells, i.e. 10 µM SA reduced nearly 80% of shikonin production and 100 µM SA completely inhibited shikonin production. They found the addition of exogenous SA caused a decreased activity of PHB geranyltransferase, one of the key regulatory enzymes in shikonin biosynthesis [30]. In another study, Arghavani et al. reported higher enhancing effects of SA compared to JA [12]. There is no report on the relationship between the endogenous content of SA and shikonin production. It is clear that the SA could improve the enzyme activity of PAL in the phenylpropane pathway [30], where the p-hydroxybenzoic acid, one of the precursors to form shikonin, is derived. In our study, the endogenous MeJA and SA were decreased by 30%NH4+ treatment within 48h, which might be related to the inhibition of shikonin biosynthesis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The information of standards used for quantification; Table S2: Primers for qRT-PCR; Table S3: Mass element formulation of different NH4+ concentration mediums (mM).
Author Contributions
Conceptualization, S.W. and L.G.; sample preparation and measurement, L.S., J.L., B.Y., C.L.; data analysis, L.S., J.L., X.W., Q.L., M.C.; writing—original draft preparation, L.S. and J.L.; writing—review and editing, S.W., R.W., Y.Z.; supervision, L.G.; funding acquisition, S.W., R.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No.82173934, No.81473307), the Fundamental Research Funds for the Central Public Welfare Research Institutes (ZZXT201901, ZZ13-YQ-084), and CACMS Innovation Fund (CI2021A03904).
Data Availability Statement
Not applicable.
Acknowledgments
We thank Dr. Chunjuan Pu for hers support in using the WinRHIZO.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A. Supplementary Data
The following is the Supplementary data to this article:
Table S1: The information of standards used for quantification.
Table S2: Primers for qRT-PCR.
Table S3: Mass element formulation of different NH4+ concentration mediums (mM).
References
- Shindo, S.; Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Matsuo, T. Shikonin Inhibits Inflammatory Cytokine Production in Human Periodontal Ligament Cells. Inflammation 2016, 39, 1124-1129. [CrossRef]
- Cocchi, F.; DeVico, A.L.; Yarchoan, R.; Redfield, R.; Cleghorn, F.; Blattner, W.A.; Garzino-Demo, A.; Colombini-Hatch, S.; Margolis, D.; Gallo, R.C. Higher macrophage inflammatory protein (MIP)-1alpha and MIP-1beta levels from CD8+ T cells are associated with asymptomatic HIV-1 infection. Proceedings of the National Academy of Sciences of the United States of America 2000, 97, 13812-13817. [CrossRef]
- Tong, Y.; Bai, L.; Gong, R.; Chuan, J.; Duan, X.; Zhu, Y. Shikonin Protects PC12 Cells Against β-amyloid Peptide-Induced Cell Injury Through Antioxidant and Antiapoptotic Activities. Scientific reports 2018, 8, 26. [CrossRef]
- Wang, B.; Wu, Z.; Wang, J.; Li, W.; Liu, G.; Zhang, B.; Tang, Y. Insights into the mechanism of Arnebia euchroma on leukemia via network pharmacology approach. BMC complementary medicine and therapies 2020, 20, 322. [CrossRef]
- Ahn, B.Z.; Baik, K.U.; Kweon, G.R.; Lim, K.; Hwang, B.D. Acylshikonin analogues: synthesis and inhibition of DNA topoisomerase-I. Journal of medicinal chemistry 1995, 38, 1044-1047. [CrossRef]
- Yazaki, K. Lithospermum erythrorhizon cell cultures: Present and future aspects. Plant biotechnology (Tokyo, Japan) 2017, 34, 131-142. [CrossRef]
- Papageorgiou, V.P.; Assimopoulou, A.N.; Couladouros, E.A.; Hepworth, D.; Nicolaou, K.C. The Chemistry and Biology of Alkannin, Shikonin, and Related Naphthazarin Natural Products. Angew Chem Int Ed Engl 1999, 38, 270-301. [CrossRef]
- Wang, S.; Wang, R.; Liu, T.; Lv, C.; Liang, J.; Kang, C.; Zhou, L.; Guo, J.; Cui, G.; Zhang, Y.; et al. CYP76B74 Catalyzes the 3''-Hydroxylation of Geranylhydroquinone in Shikonin Biosynthesis. Plant physiology 2019, 179, 402-414. [CrossRef]
- Tabata, M.; Mizukami, H.; Hiraoka, N.; Konoshima, M. Pigment formation in callus cultures of Lithospermum erythrorhizon. Phytochemistry 1974, 13, 927-932. [CrossRef]
- Fu, S.L.; Shang, T.M.; Xiao, P.G. Analysis of naphthoquinone pigments in several Arnebiae Radix. Acta Pharmaceutica Sinica 1984, 921-925. [CrossRef]
- Fujita, Y.; Hara, Y.; Ogino, T.; Suga, C. Production of shikonin derivatives by cell suspension cultures of Lithospermum erythrorhizon : I. Effects of nitrogen sources on the production of shikonin derivatives. Plant cell reports 1981, 1, 59-60. [CrossRef]
- Arghavani, P.; Haghbeen, K.; Mousavi, A. Enhancement of Shikalkin Production in Arnebia euchroma Callus by a Fungal Elicitor, Rhizoctonia solani. Iranian journal of biotechnology 2015, 13, 10-16. [CrossRef]
- Fang, R.; Zou, A.; Zhao, H.; Wu, F.; Zhu, Y.; Zhao, H.; Liao, Y.; Tang, R.J.; Pang, Y.; Yang, R.; et al. Transgenic studies reveal the positive role of LeEIL-1 in regulating shikonin biosynthesis in Lithospermum erythrorhizon hairy roots. BMC Plant Biol 2016, 16, 121. [CrossRef]
- Srinivasan, V.; Ryu, D.D. Improvement of shikonin productivity in Lithospermum erythrorhizon cell culture by alternating carbon and nitrogen feeding strategy. Biotechnology and bioengineering 1993, 42, 793-799. [CrossRef]
- Tani, M.; Takeda, K.; Yazaki, K.; Tabata, M. Effects of oligogalacturonides on biosynthesis of shikonin in Lithospermum cell cultures. Phytochemistry 1993, 34, 1285-1290. [CrossRef]
- Zhao, H.; Baloch, S.K.; Kong, L.R.; Zhang, W.J.; Zou, A.L.; Wang, X.M.; Qi, J.L.; Yang, Y.H. Molecular cloning, characterization, and expression analysis of LeMYB1 from Lithospermum erythrorhizon. Biologia plantarum 2014, 58, 436-444. [CrossRef]
- Sun, M.H.; Lu, X.P.; Cao, X.J.; Li, J.; Xiong, J.; Xie, X.X. Effect of different forms of nitrogen on the activity of nitrate reductase and expression of the relative genes in Citrus sinensis ×Poncirus trifoliate. Journal of Fruit Science 2017, 34, 410-417. [CrossRef]
- Wang, Q.; Wang, K.C.; Zheng, C.X.; Zhu, B.C.; Wang, Y.L.; Yang, J.W.; Wang, W. Effect of different nitrogenous forms on growth and chemical component in tuber of Pinellia pedatisecta Schott. . Journal of Plant Nutrition and Fertilizers 2014, 20, 1038-1043. [CrossRef]
- Crawford, N.M.; Glass, A.D.M. Molecular and physiological aspects of nitrate uptake in plants. Trends in plant science 1998, 3, 389-395. [CrossRef]
- Migocka, M.; Warzybok, A.; Kłobus, G. The genomic organization and transcriptional pattern of genes encoding nitrate transporters 1 (NRT1) in cucumber. Plant and Soil 2012, 364, 245-260. [CrossRef]
- Wang, S.; Guo, L.P.; Xie, T.; Yang, J.; Tang, J.F.; Li, X.; Wang, X.; Huang, L.Q. Different secondary metabolic responses to MeJA treatment in shikonin-proficient and shikonin-deficient cell lines from Arnebia euchroma (Royle) Johnst. Plant Cell, Tissue and Organ Culture (PCTOC) 2014, 119, 587-598. [CrossRef]
- Kuchipudi, S.V.; Tellabati, M.; Nelli, R.K.; White, G.A.; Perez, B.B.; Sebastian, S.; Slomka, M.J.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; et al. 18S rRNA is a reliable normalisation gene for real time PCR based on influenza virus infected cells. Virol J 2012, 9, 230. [CrossRef]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937-948. [CrossRef]
- Xu, Q.F.; Tsai, C.L.; Tsai, C.Y. Interaction of potassium with the form and amount of nitrogen nutrition on growth and nitrogen uptake of maize. Journal of Plant Nutrition 1992, 15, 23-33. [CrossRef]
- Song, W.; Zhuang, Y.; Liu, T. CYP82AR Subfamily Proteins Catalyze C-1' Hydroxylations of Deoxyshikonin in the Biosynthesis of Shikonin and Alkannin. Organic letters 2021, 23, 2455-2459. [CrossRef]
- Yazaki, K.; Kunihisa, M.; Fujisaki, T.; Sato, F. Geranyl diphosphate:4-hydroxybenzoate geranyltransferase from Lithospermum erythrorhizon. Cloning and characterization of a ket enzyme in shikonin biosynthesis. The Journal of biological chemistry 2002, 277, 6240-6246. [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: a critical review. Journal of Plant Physiology 2002, 159, 567-584. [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant science : an international journal of experimental plant biology 2016, 248, 92-101. [CrossRef]
- Yoshikawa, N.; Fukui, H.; Tabata, M. Effect of gibberellin A3 on shikonin production in Lithospermum callus cultures. Phytochemistry 1986, 25, 621-622. [CrossRef]
- Kumar, P.; Saini, M.; Bhushan, S.; Warghat, A.R.; Pal, T.; Malhotra, N.; Sood, A. Effect of salicylic acid on the activity of PAL and PHB geranyltransferase and shikonin derivatives production in cell suspension cultures of Arnebia euchroma (Royle) Johnst--a medicinally important plant species. Appl Biochem Biotechnol 2014, 173, 248-258. [CrossRef]
- Zhao, H.; Chang, Q.S.; Zhang, D.X.; Fang, R.J.; Zhao, H.; Wu, F.Y.; Wang, X.M.; Lu, G.H.; Qi, J.L.; Yang, Y.H. Overexpression of LeMYB1 enhances shikonin formation by up-regulating key shikonin biosynthesis-related genes in Lithospermum erythrorhizon. Biologia plantarum 2015, 59, 429-435. [CrossRef]
- Liu, Y.; Lai, N.; Gao, K.; Chen, F.; Yuan, L.; Mi, G. Ammonium inhibits primary root growth by reducing the length of meristem and elongation zone and decreasing elemental expansion rate in the root apex in Arabidopsis thaliana. PLoS One 2013, 8, e61031. [CrossRef]
- Chen, H.; Jia, Y.; Xu, H.; Wang, Y.; Zhou, Y.; Huang, Z.; Yang, L.; Li, Y.; Chen, L.-S.; Guo, J. Ammonium nutrition inhibits plant growth and nitrogen uptake in citrus seedlings. Scientia Horticulturae 2020, 272, 109526. [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate Addition Alleviates Ammonium Toxicity Without Lessening Ammonium Accumulation, Organic Acid Depletion and Inorganic Cation Depletion in Arabidopsis thaliana Shoots. Plant and Cell Physiology 2012, 53, 577-591. [CrossRef]
- Hessini, K.; Kronzucker, H.J.; Abdelly, C.; Cruz, C. Drought stress obliterates the preference for ammonium as an N source in the C(4) plant Spartina alterniflora. J Plant Physiol 2017, 213, 98-107. [CrossRef]
- Hill, T.D.; Sommer, N.R.; Kanaskie, C.R.; Santos, E.A.; Oczkowski, A.J. Nitrogen uptake and allocation estimates for Spartina alterniflora and Distichlis spicata. J Exp Mar Biol Ecol 2018, 21, 466-472. [CrossRef]
- Garnica, M.; Houdusse, F.; Zamarreno, A.M.; Garcia-Mina, J.M. The signal effect of nitrate supply enhances active forms of cytokinins and indole acetic content and reduces abscisic acid in wheat plants grown with ammonium. J Plant Physiol 2010, 167, 1264-1272. [CrossRef]
- Petersen, F.; Demann, J.; Restemeyer, D.; Ulbrich, A.; Olfs, H.W.; Westendarp, H.; Appenroth, K.J. Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina. Plants (Basel) 2021, 10, 1741. [CrossRef]
- Mantovani, C.; Prado, R.M.; Pivetta, K.F.L. Impact of Nitrate and Ammonium ratio on Nutrition and Growth of two Epiphytic Orchids. An Acad Bras Cienc 2018, 90, 3423-3431. [CrossRef]
- Carr, N.F.; Boaretto, R.M.; Mattos, D., Jr. Coffee seedlings growth under varied NO(3)(-):NH(4)(+) ratio: Consequences for nitrogen metabolism, amino acids profile, and regulation of plasma membrane H(+)-ATPase. Plant Physiol Biochem 2020, 154, 11-20. [CrossRef]
- Hao, H.; Lei, C.; Dong, Q.; Shen, Y.; Chi, J.; Ye, H.; Wang, H. Effects of exogenous methyl jasmonate on the biosynthesis of shikonin derivatives in callus tissues of Arnebia euchroma. Appl Biochem Biotechnol 2014, 173, 2198-2210. [CrossRef]
- Kang, Q.S.; Li, H.L.; Wu, Y.L.; Lu, D.Y. Studies on cell growth and Product Synthesis in Production Stage Culture of Arnebia euchroma Cell II: Fermentation culture. Natural Product Research and Development 2003, 15, 429-432. [CrossRef]
- Ge, F.; Yuan, X.; Wang, X.; Zhao, B.; Wang, Y. Cell growth and shikonin production of Arnebia euchroma in a periodically submerged airlift bioreactor. Biotechnology letters 2006, 28, 525-529. [CrossRef]
- Malik, S.; Bhushan, S.; Sharma, M.; Singh Ahuja, P. Physico-chemical factors influencing the shikonin derivatives production in cell suspension cultures of Arnebia euchroma (Royle) Johnston, a medicinally important plant species. Cell biology international 2011, 35, 153-158. [CrossRef]
- Malik, S.; Bhushan, S.; Verma, S.C.; Sharma, N.; Sinha, A.K.; Sharma, M.; Ahuja, P.S. Production of naphthoquinone pigments in cell suspension cultures of Arnebia euchroma (Royle) Johnston: influence of pH on growth kinetics and acetylshikonin. Med Aromat Plant Sci Biotechnol 2008, 2, 43-49.
- Liang, J.W.; Li, T.; Wang, R.S.; Zhou, L.; Yang, Q.; Wang, S.; Guo, L.P. Establishment of RNA interfered hairy root system of two CYP450 genes in Arnebia euchroma and its influence. China Journal of Chinese Materia Medica 2020, 45, 3422-3431. [CrossRef]
- Lu, W.H.; Chen, Y.F.; Wang, F.; Dai, N.B.; Hao, A.H.; Li, C.F.; Jia, S.E. Effects of physical and chemical conditions on hairy root culture and content of Arnebia euchromu. Journal of Huazhong Agricultural University 2012, 31, 50-54. [CrossRef]
- Zhang, P.; Wang, F.; Zhu, C. Influence of fungal elicitor and macroporous resin on shikonin accumulation in hairy roots of Arnebia euchroma (Royle) Johnst. Chinese Journal of Biotechnology 2013, 29, 214-223. [CrossRef]
- Yang, Y.H.; Lu, J.; Zhao, Q.H.; Cao, R.Q. Effect of water extract of algae on growth and pigment formation in Lithospermuml erythrorhizon and Arnebia euchromu cell cultures. Journal of Plant Resources and Environment 1992, 1, 39-44.
- Ge, S.N.; Zhang, S.X.; Wang, F.; Wei, H.; Xie, W.L.; Xie, J.; Li, C.; Zhao, H. Effects of Exogenous Substances on the Content of Secondary Metabolites in Arnebia euchroma Johnst Hairy Roots. Food Science 2016, 37, 160-164. [CrossRef]
- Li, M.; Pan, Q.; Wang, F.; Zhang, P. Research of Effects on Arnebia euchroma (Royle)Johnst Hairy Roots Growth Factors. Xinjiang Agricultural Sciences 2012, 49, 2062-2068. [CrossRef]
- Lu, W.H.; Pan, Q.; Wang, F.; Chen, Y.F.; Dai, N.B. Influence of Culture Conditions on Arnebia euchromaHairy Roots Growth and Shikonin Content. Acta Botanica Boreali-Occidentalia Sinica 2012, 32, 1686-1691. [CrossRef]
- Yang, Y.; Zhang, H.; Cao, R. Effect of Brassinolide on Growth and Shikonin Formation in Cultured Onosma paniculatum Cells. J Plant Growth Regul 1999, 18, 89-92. [CrossRef]
- Yang, H.; von der Fecht-Bartenbach, J.; Friml, J.; Lohmann, J.U.; Neuh User, B.; Ludewig, U. Auxin-modulated root growth inhibition in Arabidopsis thaliana seedlings with ammonium as the sole nitrogen source. Funct Plant Biol 2015, 42, 239-251. [CrossRef]
- Dziewit, K.; Pěnčík, A.; Dobrzyńska, K.; Novák, O.; Szal, B.; Podgórska, A. Spatiotemporal auxin distribution in Arabidopsis tissues is regulated by anabolic and catabolic reactions under long-term ammonium stress. BMC Plant Biol 2021, 21, 602. [CrossRef]
- Péret, B.; De Rybel, B.; Casimiro, I.; Benková, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: an emerging story. Trends in plant science 2009, 14, 399-408. [CrossRef]
- Barreto, R.F.; de Mello Prado, R.; Lúcio, J.C.B.; López-Díaz, I.; Carrera, E.; Carvalho, R.F. Ammonium Toxicity Alleviation by Silicon is Dependent on Cytokinins in Tomato cv. Micro-Tom. Journal of Plant Growth Regulation 2021, 41, 417-428. [CrossRef]
- Walch-Liu, P.; Neumann, G.; Bangerth, F.; Engels, C. Rapid effects of nitrogen form on leaf morphogenesis in tobacco. Journal of experimental botany 2000, 51, 227-237. [CrossRef]
- Fang, D.Q.; Hou, C.S.; Li, X.M.; Ye, H.C.; Li, G.F. Effects of pH and hormone on cell growth and synthesis of Shikonin derivatives in suspension culture of Arnebia euchroma. Plant Science Journal 1994, 159-164.
- Zhu, W.H.; Fan, H.X.; Hu, Q.; Zhu, H.Q. Induction culture of callus and selection of superior clones of Onosma paniculatum. Journal of Chinese Medicinal Materials 1990, 6-9. [CrossRef]
- Yan, H.Y.; Cao, R.Q. Influencing factors of Shikonin formation in callus culture of Lithospermum erythorhizon Acta Agriculturae Boreali-Sinica 2002, 116-120. [CrossRef]
- Ahmad, M.; Varela Alonso, A.; Koletti, A.E.; Rodić, N.; Reichelt, M.; Rödel, P.; Assimopoulou, A.N.; Paun, O.; Declerck, S.; Schneider, C.; et al. Dynamics of alkannin/shikonin biosynthesis in response to jasmonate and salicylic acid in Lithospermum officinale. Scientific reports 2022, 12, 17093. [CrossRef]
- Ding, J.; Shi, S.; Jiang, B.-H.; Yang, Y.-H.; Huang, J.; Shen, H.-G.; Xia, K.; Zhang, J.; Jiang, X. Effects of Methyl jasmonate with indole-3-acetic acid and 6-benzylaminopurine on the secondary metabolism of cultured Onosma paniculatum cells. In Vitro Cellular & Developmental Biology - Plant 2004, 40, 581-585. [CrossRef]
- Zhu, Y. Cloning, expression and functional analysis of LeTCP4 gene in Lithospermum erythrorhizon. Master's degree, Nanjing University,Nanjing, 2012.
Figure 1.
Continuous growth of AEHR in 15 days after treated by different concentrations of NH4+ (0%, 10%, 20%, 30%).
Figure 1.
Continuous growth of AEHR in 15 days after treated by different concentrations of NH4+ (0%, 10%, 20%, 30%).

Figure 2.
Biomass differences of AEHR under different concentrations of NH4+.Values given are means ± SE (n = 3 replicates). Significant differences (p<0.05) were indicated by different letters for each variable.
Figure 2.
Biomass differences of AEHR under different concentrations of NH4+.Values given are means ± SE (n = 3 replicates). Significant differences (p<0.05) were indicated by different letters for each variable.
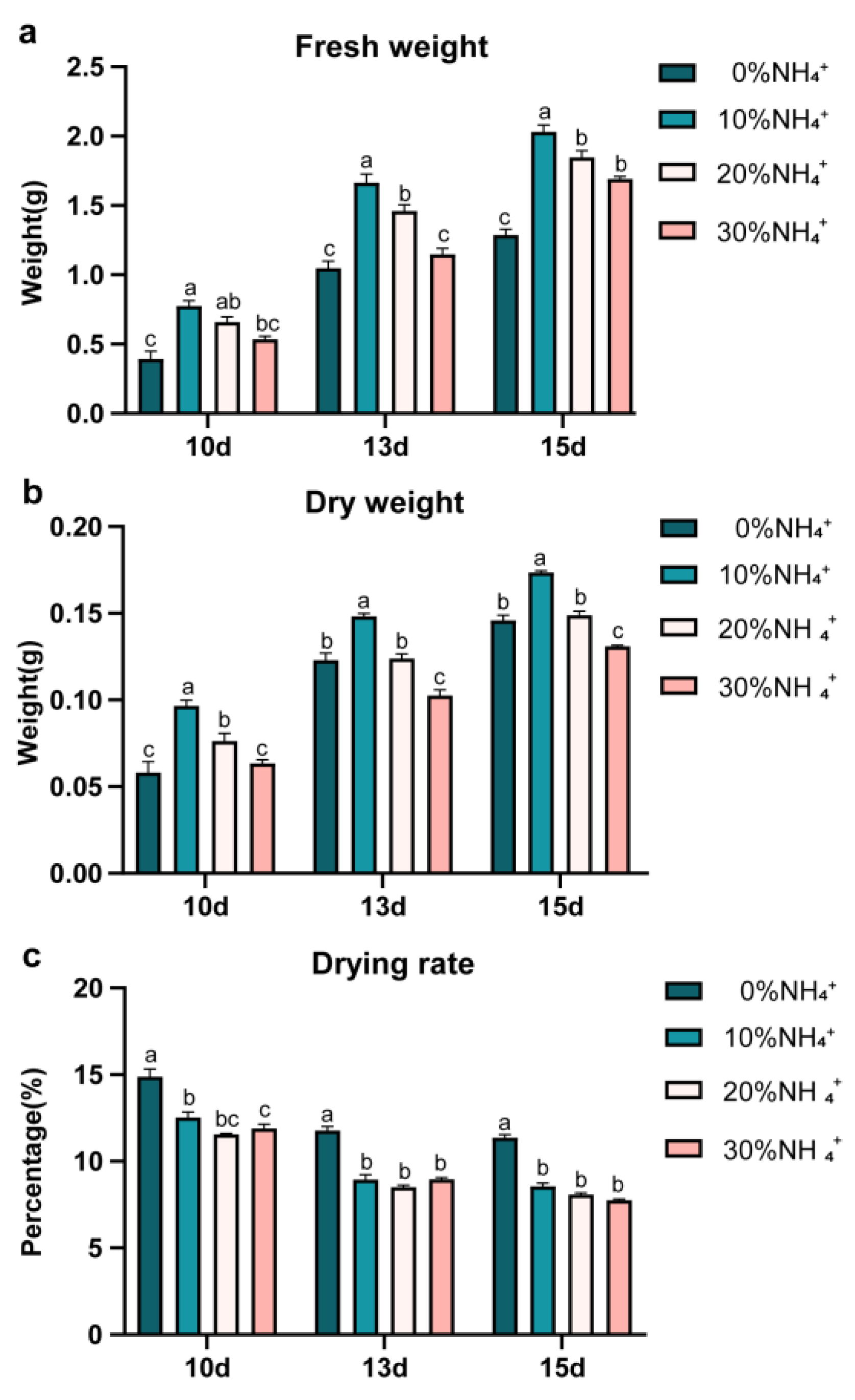
Figure 3.
Effects of different concentrations of NH4+ on synthesis of Shikonin within 15 days. Vertical bars represent mean ± SE (n = 3). (a) HIVS, β-hydroxyisovalerylshikonin; (b) AS, acetylshikonin; (c) DS, deoxyshikonin; (d) AOIVA, β-acetoxyisovalerylalkannin; (e) IBS, isobutyrylshikonin; (f) DAA, β,β’-dimethacrylicalkannin.
Figure 3.
Effects of different concentrations of NH4+ on synthesis of Shikonin within 15 days. Vertical bars represent mean ± SE (n = 3). (a) HIVS, β-hydroxyisovalerylshikonin; (b) AS, acetylshikonin; (c) DS, deoxyshikonin; (d) AOIVA, β-acetoxyisovalerylalkannin; (e) IBS, isobutyrylshikonin; (f) DAA, β,β’-dimethacrylicalkannin.
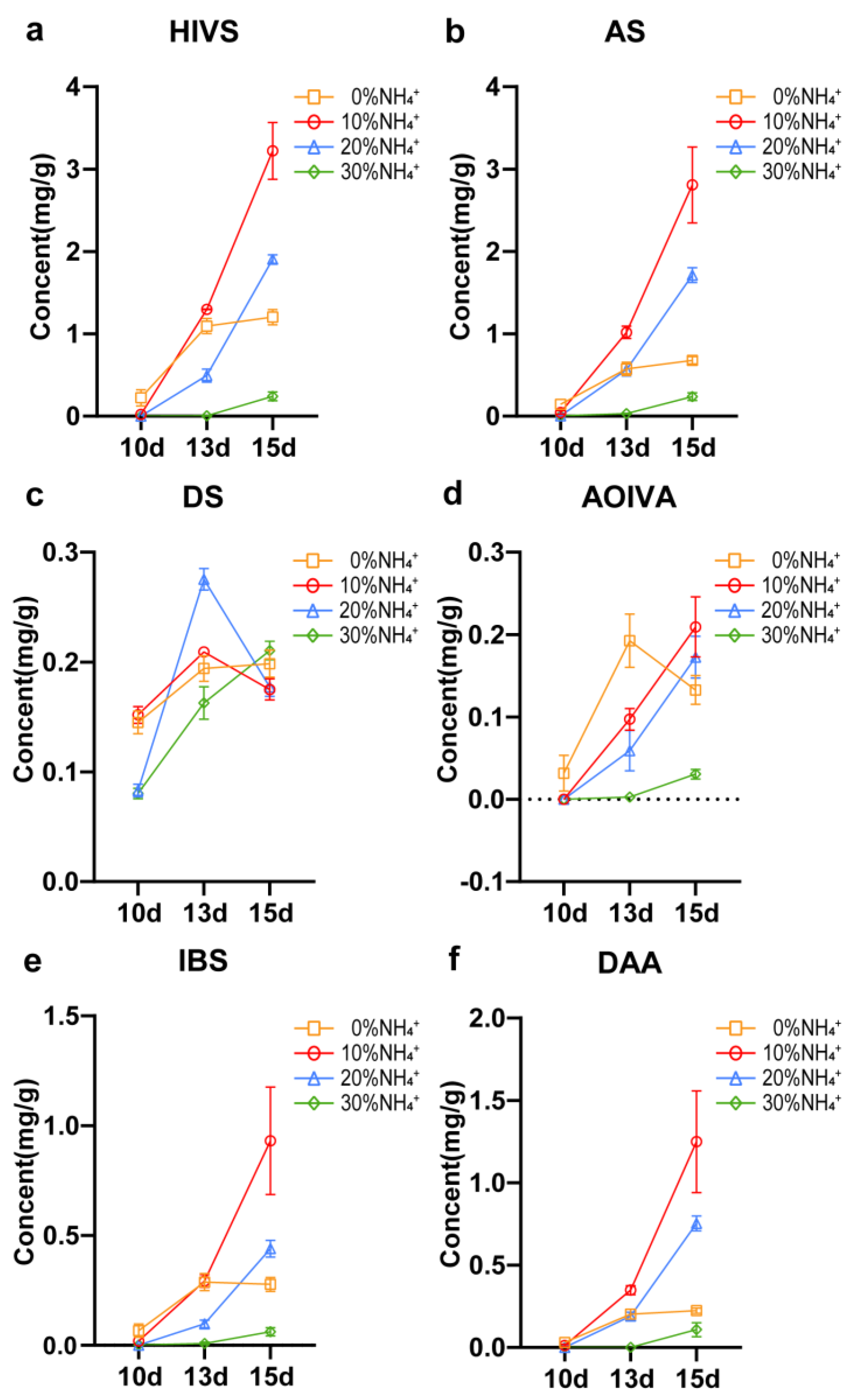
Figure 4.
Morphology characters of AEHR after treated for 15 days with different NH4+ concentrations. (a) morphology characters of AEHR after treated for 15 days with different NH4+ concentrations. (b-c) average length and average diameter of AEHR. (d) the root surface area. (e) the root projected area. (f) mean root volume. (g) branch number. (h-i) the AEHR length (L) within different diameter ranges and its percentage of total root length (PTL). Different letters represent significant differences (n=3, mean ± SE, p<0.05).
Figure 4.
Morphology characters of AEHR after treated for 15 days with different NH4+ concentrations. (a) morphology characters of AEHR after treated for 15 days with different NH4+ concentrations. (b-c) average length and average diameter of AEHR. (d) the root surface area. (e) the root projected area. (f) mean root volume. (g) branch number. (h-i) the AEHR length (L) within different diameter ranges and its percentage of total root length (PTL). Different letters represent significant differences (n=3, mean ± SE, p<0.05).
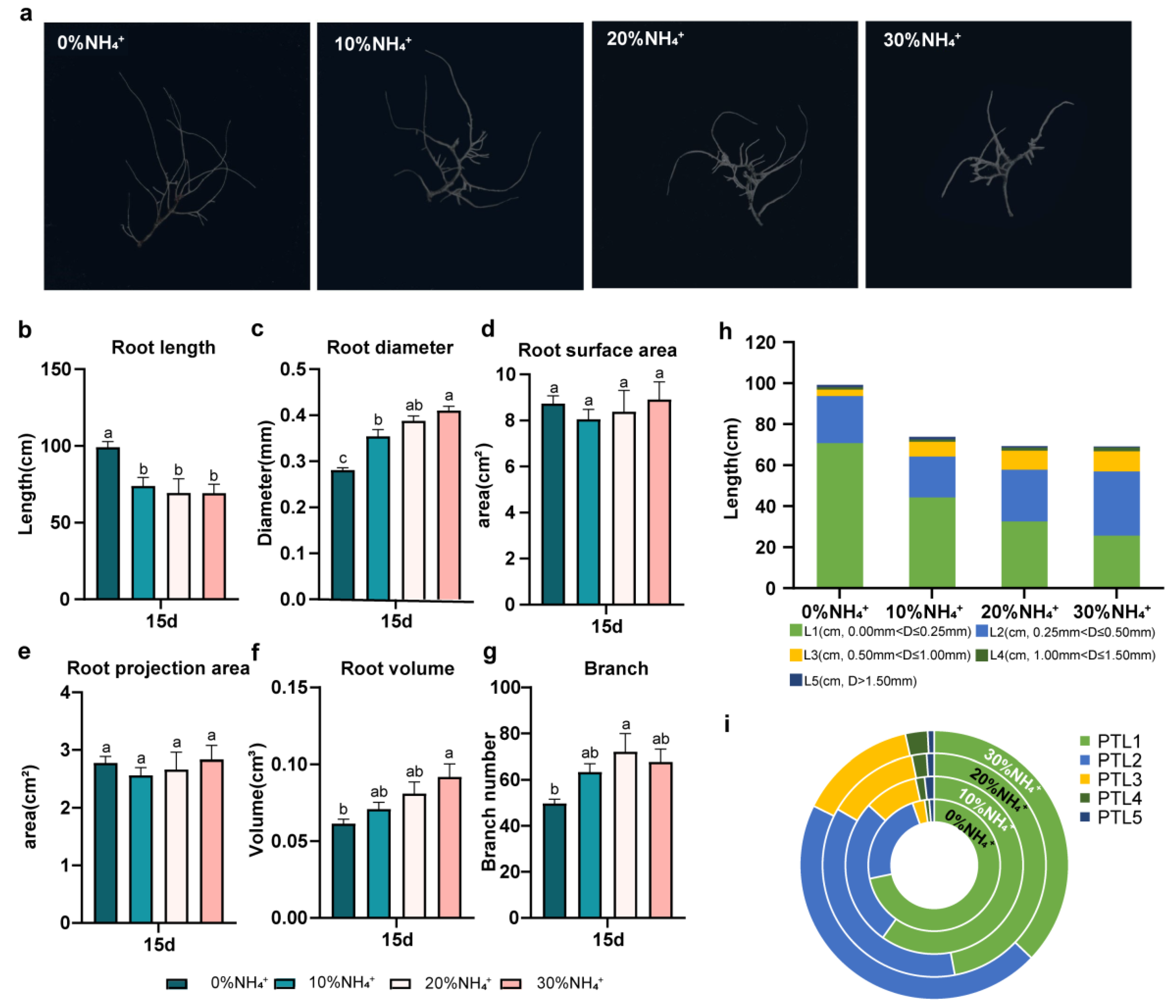
Figure 5.
Contents of NO3-N and NH2-N in AERH with different NH4+ concentration treatments within 48h. Vertical bars represent mean ± SE (n = 3).
Figure 5.
Contents of NO3-N and NH2-N in AERH with different NH4+ concentration treatments within 48h. Vertical bars represent mean ± SE (n = 3).
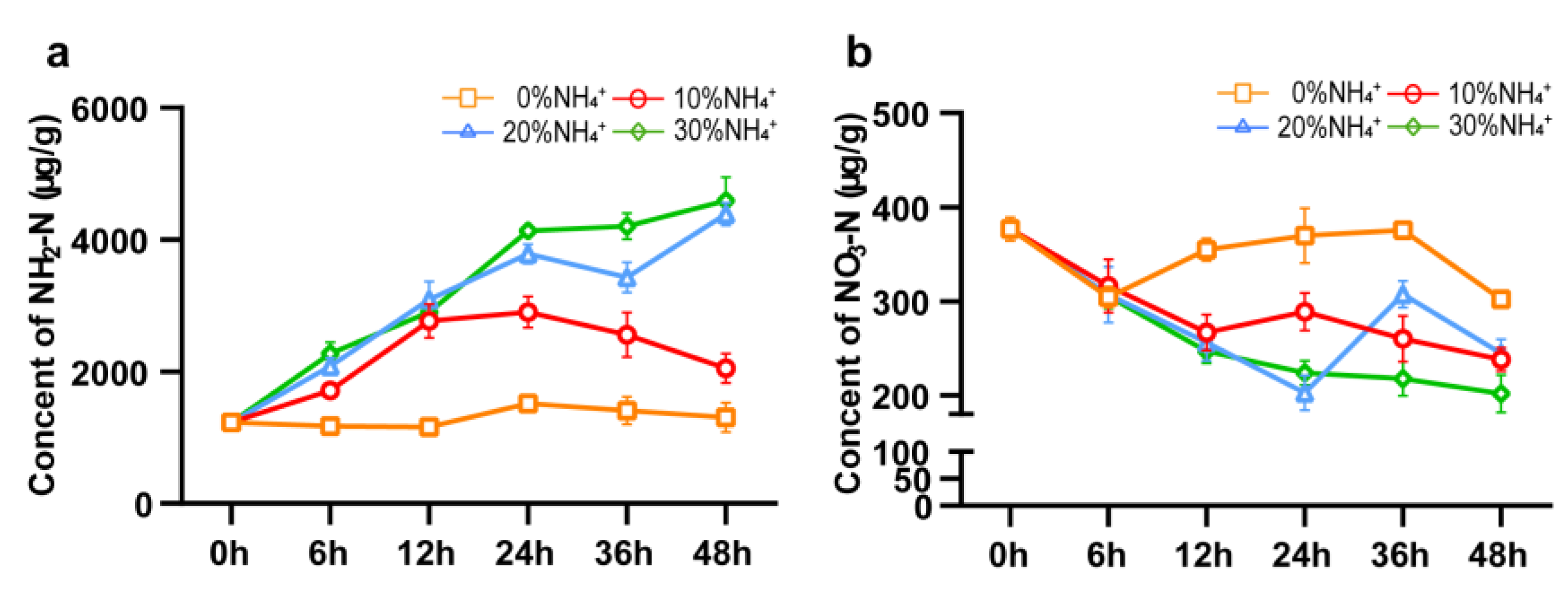
Figure 6.
Effects of different concentrations of NH4+ on synthesis of shikonin within 48 h. Vertical bars represent mean ± SE (n = 3). (a) HIVS, β-hydroxyisovalerylshikonin; (b) AS, acetylshikonin; (c) DS, deoxyshikonin; (d) AOIVA, β-acetoxyisovalerylalkannin; (e) IBS, isobutyrylshikonin; (f) DAA, β,β’-dimethacrylicalkannin.
Figure 6.
Effects of different concentrations of NH4+ on synthesis of shikonin within 48 h. Vertical bars represent mean ± SE (n = 3). (a) HIVS, β-hydroxyisovalerylshikonin; (b) AS, acetylshikonin; (c) DS, deoxyshikonin; (d) AOIVA, β-acetoxyisovalerylalkannin; (e) IBS, isobutyrylshikonin; (f) DAA, β,β’-dimethacrylicalkannin.
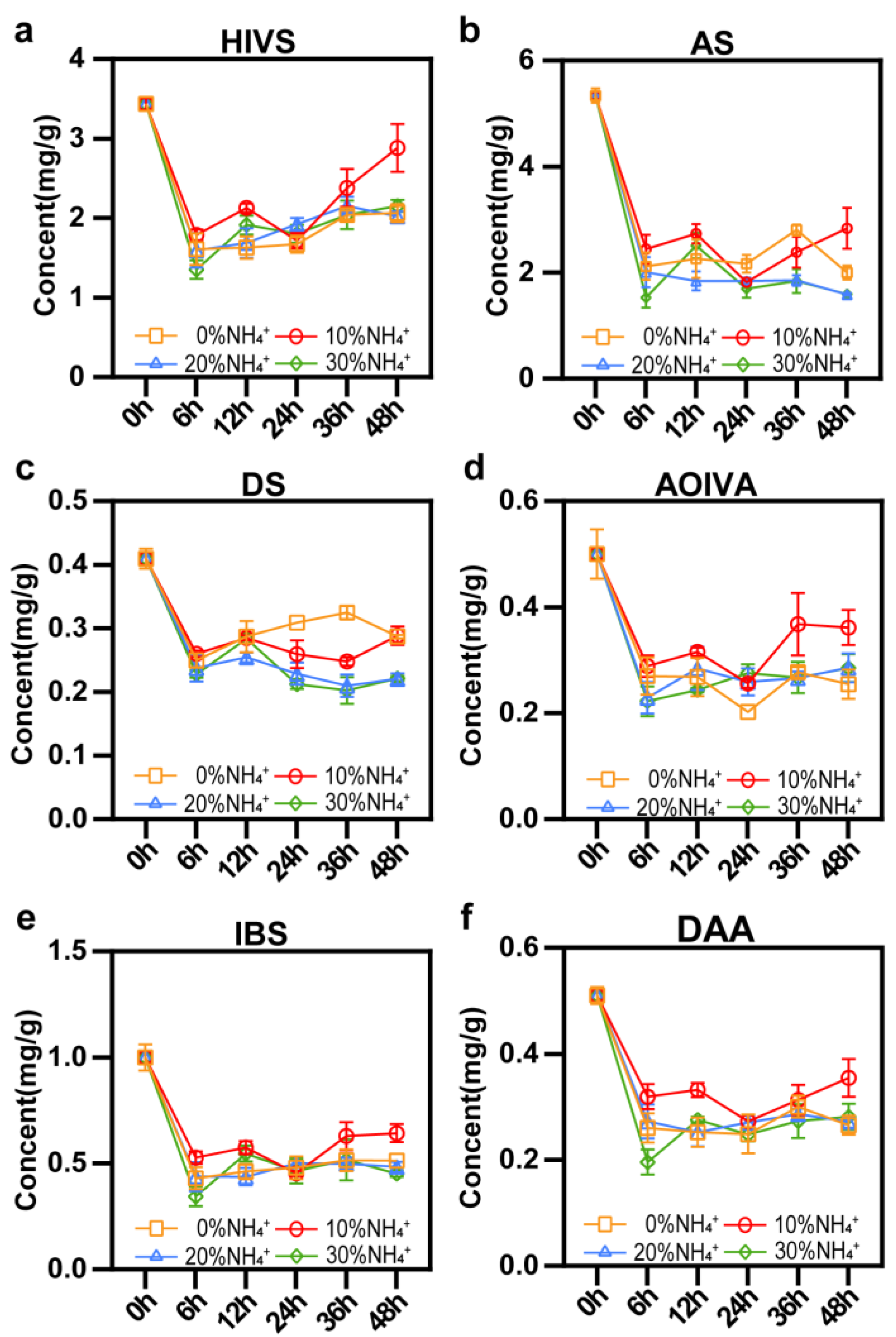
Figure 7.
Effects of different concentrations of NH4+ treatment on the gene expression of key enzymes from shikonin biosynthesis pathway within 36h. Vertical bars represent mean ± SE (n = 3).
Figure 7.
Effects of different concentrations of NH4+ treatment on the gene expression of key enzymes from shikonin biosynthesis pathway within 36h. Vertical bars represent mean ± SE (n = 3).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



