
Submitted:
20 April 2023
Posted:
21 April 2023
You are already at the latest version
Abstract
Abstract: Heavy metals (HMs) contamination is one of the main among abiotic factors affecting crop productivity and also threatens human health via consuming metal contaminated crops as a food source. Over the past few years, HMs have drawn a lot of attention due to their increased use for commercial purposes and their harmful effects on plants and other life forms, thus threatening human survival. However, in recent years, several methods have been adopted to combat the harsh effects of HMs. After phytohormones, use of mineral nutrients such as selenium (Se) in the prevention of HM stress has been explored by the researchers more recently. Selenium is an important micronutrient widely known for its antioxidant properties in both plants and animals. Exogenous Se inhibits metal uptake and its translocation and also improves the antioxidant system, thus imparts resistance to HM toxicity in plants. Moreover, Se also regulates the production of various osmolytes in cells that helps in developing cell osmolarity. Selenium also induces the production of different types of secondary metabolites (SMs) that are also involved in plant's secondary defense mechanisms to different stresses. Uptake of mineral nutrients is a vital process for plant growth and development, which is also positively correlated with Se under metalloid toxicity. However, in order to understand the exact mechanism of Se in HM tolerance, different metabolic processes stimulated by Se and their pathways need to be explored. Hence, this review focuses on the role of Se on nutritional status, antioxidants metabolism, interaction with phytohormones and its role in the regulation of various genes involved in Se induced HM tolerance. Thus, this study will help researchers in the future for the improvement of HM tolerance via Se application in plants.
Keywords:
Heavy metal
; Mineral Nutrition
; Phytohormones
; Selenium
1. Introduction
Heavy metal pollution is one of the notable obstructions in determining the productivity of crops at the worldwide level [1,2,3]. Heavy metals enter the plant through the root from soil deposits and have a significant negative impact on several essential plant functions, including mineral nutrition, photosynthesis, water uptake and ion distribution [4,5,6,7]. Reports suggest that higher concentration of arsenic (As) in soil increases its uptake and accumulation in several commercial crops such as rice [8,9,10]. However, the increasing concentrations of HMs in agricultural land have negatively impacted on the productivity of crops [10,11]. The contamination of agricultural soil with innumerable HMs has become a worldwide problem which mainly arised due to human activities [4,11]. Recently, it has been reported that As toxicity reduced photosynthetic functions, N-assimilation and hence affected the overall growth of Oryza sativa [7,10]. Besides, mineral elements, plants also require a limited amount of HMs for their normal functioning and metabolic activities. In order to prevent agricultural crops from metal toxicity, use of several essential elements has been made and, among them, Se is widely used because of its various beneficial roles and its potential to trigger HM tolerance in plants. Mineral nutrition is one of the key processes that is directly involved in the growth and development of plants. Plants under stressful conditions produce different types of signaling molecules to overcome stress [12]. Selenium is a chief element for plants. Selenium is a broadly distributed trace element with both toxic and beneficial effects on plants, humans, and animals. At low concentrations, Se might be useful for plant growth and development under optimum and unfavorable conditions [13]. However, the agglomeration of Se on the surface of water or soil may become the source of toxicity for plants. For this reason, most of the research on plant Se resistance has emphasized on Se interaction with sulfur (S) metabolisms [14]. The addition of Se limits the uptake of metals by roots and their translocation to shoots by synthesizing various growth regulators, which is one of the indispensable stress tolerance mechanisms in plants [15]. The low concentration of Se can reduce metals' toxic effects on plants and develop tolerance against stress by inducing different metabolic processes [16]. However, at higher concentration Se affects plant growth and decrease the level of organogenesis, nucleic acid synthesis and protein synthesis. The uptake of Se from soil is controlled on many edaphic and plant factors. Primarily, soil is the most common factor controlling Se entry into the plant body [17]. Selenium application reduces the adverse effects of both biotic and abiotic stresses by adjusting different type of mechanisms like secondary metabolites and antioxidant system [13]. Selenium application enhanced photosynthesis by activating antioxidant enzymes involved in ROS detoxification under cadmium (Cd) stress in Capsicum annuum L. [18]. Studies have also shown that selenium decreased the levels of ROS and induced lignification of cell wall under cadmium stress in Oryza sativa [19]. However, [20] investigated the detoxification action of Se on growth of garlic plants under Cd stress. Osmolytes, which are also the important components of plant defense systems, are synthesized under stress conditions. Further, the application of Se also induced the production of some amino acids such as proline (Pro), glycine betaine (GB), etc. under metal stress [21,22]. Nanoparticle treatment of Se also regulates the accumulation of Pro, GB, and sugars in response to HMs stress [23,24]. These amino acids act as osmolytes that help in developing tolerance to different stress conditions by maintaining their osmotic balance. Moreover, plants also secrete some important compounds that participate in plant secondary defense against both biotic and abiotic stress conditions. These secretions are commonly known as SMs. Among all the classes of SMs, phenolics are one of the major classes of SMs that participate in plant secondary defense. However, the exogenous supply of Se significantly enhanced the levels of phenolic metabolites under different type of metal stresses [13,25,26,27]. Furthermore, antagonistic results of Se on the production glucosinolates (GLS) were also observed in plants when exposed to metalloid toxicity [28,29,30]. Accumulation of HMs in soil badly affects the uptake of mineral elements from soil, thereby affecting the overall growth and metabolism of plants. However, the exogenous treatment of Se enhances the uptake of the elements by interfering with metal uptake and hence improves the nutritional status of plant [31,32,33]. Furthermore, Se application also accumulates large amount of Se in grains under metal stress, thus helping in developing nutritional-rich crop variety [34]. Plant hormones play a collaborating response to the changing environment and are necessary for stress tolerance and crop production [18]. Selenium can occur in soil in four different forms as Se, Selenate, Selenite, and inorganic selenide [35]. Under HM stress conditions, the endogenous production of various signalling molecules like ABA, ETH, SA, brassinosteroid (BRs) and JA increased, while auxin (AUX), gibberellins (GA) and cytokinin (CK) levels decreased [36,37,38]. However, the interaction between ETH and ROS was found to be effective in Solanum lycopersicum under HM stress [38]. Ethylene is among the crucial plant stress hormones under adverse environmental conditions. Reports also suggest that, Se induced production of ETH is controlled by the type of Se form supplied [39,40]. However, the increased levels of ABA and GA in response to selenium nano-particles (SeNPs) treatment were also observed in Brassica napus [41]. In Cd stressed Nicotiana tabacum, exogenous Se treatment increased the levels of indole acetic acid (IAA) [42]. However, seed priming with GA and SA showed stimulatory effects on growth and metabolism of mung bean under Cd stress conditions [43]. Furthermore, role of JA and GA in inducing tolerance to Cd stress is also explored in Cicer arietinum [44]. However, the interactive role of ETH on Se mediated antioxidant system under Cd stress is studied in Solanum tuberosum [45]. Phytohormones (exogenous/endogenous) alone play a major role in HM tolerance at the molecular level by the regulation of different defense mechanism genes that contribute to developing tolerance to against HM toxicity in plants [46]. However, the information regarding the interaction of Se and phytohormones under HM stress is very scanty. Thus, the present review summarizes the mechanism of Se induced tolerance to metal stress via regulation of different processes such as Se uptake and translocation, antioxidant metabolism and mineral nutrition. This review also highlights the role of Se towards secondary metabolites production and its interaction with phytohormones under metal stress.
2. Selenium Uptake and Translocation
Selenium is a metalloid occurring naturally in sedimentary rocks present in dry areas of the world [47]. The inorganic forms of Se i.e Selenite (SeO32−) and Selenate (SeO42−) are the two main forms of Se which remain available to plants through soil. Selenite and selenate present in anoxic and oxic soils respectively [48]. Selenium chemically resembles sulfur(S), hence taken up inside the plants via S transporters located in root plasma membrane [49]. Many species of plant have been identified as Se hyper accumulators which have the capability to accumulate high concentrations of Se such as Stanleya pinnata [50]. Selenium uptake differs from species to species, soil type, soil concentrations and form of Se supplied [51]. Selenate absorption by plants is a well-established mechanism. The high-affinity sulphate transporters help in the absorption of selenate by roots. However, the selectivity of various plant species for sulphate and selenate varies [52,53]. On the other hand, nothing is understood about the process behind selenite uptake by plants. It has been observed that plant roots absorb selenite through passive diffusion [54]. Several species of plants belonging to the families Asteraceae, Brassicaceae and Fabaceae absorb higher concentrations of Se in hair like epidermal appendages, called as trichomes [55]. Metabolism of Se in higher plants is closely associated to that of S due to their chemical analogy. Both ions SeO32− and SeO42− are absorbed by the plants via roots and SeO42− is taken via sulphate transporters (SULTRs). This relationship is demonstrated by the fact that Se is taken up by the plants via sulphate transporters and SeO32− via aquaporins and phosphate transporters [48,53,56].). However, considerable differences in uptake and transport mechanisms of inorganic forms are found in plants [57]. Selenate is uptake by plant roots in an active manner and leads to increased Se concentrations in the roots. However, selenite is not normally concentrated in roots [57]. Selenium is metabolized by S assimilation enzymes resulting in the synthesis of organic forms of Se i.e., selenocysteine (SeCys) and selenomethionine (SeMT). However, Se uptake can also be increased from soil via aquaporins by maintaining the pH of apoplastic space through H+-ATPases [58]. Moreover, in rice it was observed that selenite and SeNPs were accumulated in roots, whereas selenate is more stored in shoots [59]. Selenium uptake can also be enhanced by regulating the expression of SLUTRs [60]. Thus, from the above paragraph it can be concluded that, plants uptake different forms of Se from soil by means of different transporters such as sulfur and phosphate transporters. However, plants also promote uptake of some forms of Se in energy-independent manner via aquaporins. Moreover, the organs for different forms of Se accumulation and its transport also vary based on the form of Se accumulated.
3. Selenium and Heavy Metals
Selenium is an important nutrient for plant which is foe to some living organisms like bacteria, human and most of the chlorophyte species [61,62,63]. Even Se is crucial for oxidative stress resistance, immunity enhancement, and antipathy of heavy metal toxicity [64,65]. Metalloid toxicity is now emerging as a significant concern to both humans and the environment. [7,10]. Globally anthropogenic is the main cause for metal toxicity affects in agricultural lands Heavy metals are non-biodegradable, gets accumulated in higher trophic levels of food chain (biomagnification) and become a threat for biotic forms and its environment [66,67]. In Lactuca sativa, addition of selenite significantly reduced the agglomeration of Pb and Cd and at the same time increased the uptake of some essential elements including Se [68]. Selenium promotes the growth of plants and may act as heavy metal opponent as it is a necessary micronutrient with some anti-oxidative and physiological properties [69,70]. The cadmium existing in water, soil and in the atmosphere can cause serious problem to all organisms and its bioaccumulation in the food chain can be highly hazardous [71]. The presence of Se may affect the distribution of elements which are necessary for plant growth and metabolism. The Se functions to reduce metal toxicity and regulate micronutrient levels by modifying soil conditions like pH, adsorption, and organic matter. It plays a vital role in enhancing the metal chelation and improves ROS scavenging mechanism in [70,72,73,74]. The useful impact of Se on plants is often described as its ability to increase the anti-oxidative capacity of their cells. The application of Se enhances the cellular anti-oxidative mechanism by elevating the enzymatic activity of CAT, GR, SOD and APX which imparts the protection to the plant cellular membranes against oxidation [75,76]. An appropriate concentration of Se can delay senescence, improve photosynthesis, antioxidant capacity, induces auxin levels and promote plant growth [42,77,78]. Selenium application at optimum concentration improved the growth and biomass of Cucumis sativus plants under heavy metals stress (Cd, Pb, Ni) by detoxification of ROS induced damages via regulation of activities of various antioxidant enzymes [79]. Application of 2 μM Se improved root growth and decreased oxidative stress in Al treated Lolium perenne [80]. Exogenous treatment of Se reduced As induced toxicity in Phaseolus aureus by interfering with its uptake and also improved plant growth by the activation of antioxidant system under As toxicity [81]. However, the application of Se in Pb stressed Vicia faba plants reduced the accumulation of ROS via enhancement in the activities of various antioxidant enzymes [82]. Application of Se reduced the oxidative stress caused by combined treatment of Cd and Pb in Brassica napus via minimizing the uptake and accumulation of both Cd and Pb [83]. Selenium plays a pivotal role in neutralizing abiotic stresses in plants. However, the role of Se in response to antioxidant metabolism has been studied under HM stress (Table 1) and found to induce tolerance to different HMs discussed below:
Selenium application is involved in the recovery of chloroplast injuries caused due to Cd stress in Cucumus sativus by increasing starch content in leaves [91]. Exogenous Se alleviated Zn stress by improving growth and photosynthesis in Billbergia zebrina [92]. Application of SeMT mitigated Cd toxicity in mustard species by enhanced levels of enzymatic antioxidants and also prevented the aggregation of Cd in cell organelles [86]. Further, leaf spraying of Se (10, 20, and 40 μM) reduced Cd toxicity by limiting Cd uptake by roots and improved phtosynthetic attributes, osmoprotectants and antioxidants level in Satureja hortensis [87]. However, Se application significantly increased the accumulation of organic Se in grains but at the same time affected the uptake and translocation of other HMs in Triticum aestivum [93]. More interestingly, exogenous supplementation of Se is also involved in promoting flowering in cucumber which was delayed by Cd and maintains sex ratio in plants [94]. Foliar spraying of Se in wheat leaves inhibited metal uptake and enhanced the nutritional status of wheat by accumulating more Se in the grains [34]. Moreover, the application of SeNPs ameliorated the toxic effects of Cd by enhancement in the production of some essential SMs and osmoprotectants in Coriandrum sativum [22]. Supplementation of Se considerably reduced Cd stress in Raphanus sativus by improving photosynthetic attributes, antioxidants level and micro-essential nutrients [31]. In plants, Se has a dual function. At low concentrations, it acts as an antioxidant, but at enormous doses, it generates ROS, which impairs several cellular activities. It is mostly unknown how Se affects plant growth, physiology, and antioxidant machinery at various concentration levels [95]. Foliage application of SeNPs alleviated both Cd and Pb toxicity in Salvia officinalis and decreased lipid peroxidation of membranes [96]. Selenium treatment reduced ROS accumulation in the reproductive parts of rice and increased pollen viability and germination under Cd stress [97]. Researchers revealed that the utilization of elements with antioxidant properties can abate the hazardous effects of heavy metal stresses [98]. Figure 1 shows the mechanism of HM induced changes on growth and metabolism of plants and the role of Se in the alleviation of HM induced oxidative stress by triggering various defense processes in plants. (ROS: reactive oxygen species, PCD: programmed cell death).
Some researchers reported that Se application reduced Cd accumulation and decreased toxicity in pepper by enhancing chlorophyll contents and overall antioxidant activity [99]. In rice seedlings, Se increases the absorption of Cd in the cell wall and vacuoles and decreases the amount of Cd in its toxic form [89]. Optimal doses of Se were known to cope up various types of biotic and abiotic stresses, including exposure to HMs by the regulation of antioxidant system [100]. Selenium is also a fundamental part of some antioxidant enzymes, which shield cells from being damaged by ROS [25,101]. Additionally, reports also showed that an ideal concentration of Se (1/2 µM) enhanced the antioxidant capabilities of photosynthetic pigments and slowed the ageing of leaf tissues in Brassica oleracea [25,102]. Addition of Se reduced oxidative stress by the stimulation of ROS scavenging system, and synchronizes Cd accumulation in shoots and root in Brassica juncea [88]. Recent reports also confirm that all the forms of Se (SeNPs, Inorganic and Organic) contribute in HM tolerance by inducing the activities of various antioxidants and other biochemical parameters in Oryza sativa under Cd stress [89].
Selenium plays a significant role in the mitigation of metal induced toxicity by the improvement in overall growth and metabolism of plants. Primarily, Se reduces metal uptake and transport by changing root architecture, stimulating different types of antioxidants, PCs and produces many other types of defensive compounds in plants to increase the tolerance potential of plants against metal toxicity.
4. Selenium Interaction with Osmolytes under Heavy Metal Stress
To avoid cellular damages due to the accumulation of ROS, plants deposit some compatible solutes known as “osmolytes” to protect the cellular machinery from several environmental stresses. Among all the osmolytes, Pro, GB, polyamines, and sugars are the common osmolytes that plants accumulate under different stressful conditions and play important role in osmoregulation. However, plants synthesize different types of osmolytes under different environmental constraints and their exogenous supply may also enhance the production of other osmolytes such as proline application relieved As induced oxidative damages by improving other osmolytes concentration in mung bean crops [103]. Increment in the levels of Pro and GB with the application of Se (10µM) demonstrating its role in providing membrane stability in Solanum lycopersicum under Cd stress [21]. Increased production of osmolytes including Pro, GB and sugars protect the cells from damaging against stress condition by developing osmotic potential and regulating other defense mechanisms [88,104]. It has also been reported that Se supplementation to Cd challenged tomato plants promoted the production of compatible solutes and stress markers which relieves Cd toxicity [22]. However, application of SeNPs increased Pro accumulation in Coriandrum sativum under Cd stress [22]. However, the leaf spraying of Se elevated leaves Pro content and grain Se concentration in wheat cultivars [105]. Application of SeNPs increased the content of Pro and GB in Glycine max subjected to As stress [23]. Selenium at optimal concentration positively regulated the synthesis of important osmoprotectants such as Pro and sugars [106]. Increased level of important compatible solute Pro was also observed in Triticum aestivum in response to Se under Cd stress [24]. Osmolytes play an important part in the maintenance of osmoregulation in plants under various stress conditions. These osmolytes also called osmoprotectants are synthesized in plants and helps in developing tolerance to plants against different stresses. Among them, Pro, glycine betaine and sugars are most synthesized osmolytes in plants under stress conditions. However, it has been also observed that Se regulates the biosynthesis of these osmolytes under stress conditions. These osmolytes protect the plants against HM toxicity by maintaining osmolarity, membrane stability and strengthens antioxidant defence system to minimize ROS induced damages.
5. Selenium Interplay with Phytohormones under Heavy Metal Stress
Phytohormones have gained much attention over the last two decades due to their wide use in plants against many abiotic stresses. However, their role in amelioration of different stresses attracted researchers to more and more use of these signaling molecules. In recent years a lot of work has been carried out to elaborate the role of phytohormones against HM stress. These signaling molecules protect plants against HMs by regulating different physiological and biochemical processes that help in developing tolerance strategies against stresses [107]. Moreover, their role along with Se in the alleviation of HM stress is least studied so far. Exogenous application of AUX affected the uptake of Cd and reduced Cd stress by improving photosynthesis and antioxidant system and raised the level of endogenous nitric oxide (NO) in tomatoes [108]. Furthermore, role of GA application in the mitigation of Nickel (Ni) toxicity was also observed in mung beans by improving growth and biomass and reduced Ni uptake [109]. However, Se interferes with HMs by regulating the endogenous production of phytohormones [15]. Reports also suggest that inorganic forms of Se negatively affect the production of ethylene [39]. However, their organic forms, such as SeMT, positively regulated ethylene biosynthesis but also affected AUX transport in rice [40]. Moreover, it has been also noticed that optimal concentration of Se is involved in inducing tolerance to Cd and low phosphorous stress in Nicotiana tabacum by regulating AUX biosynthesis and transport genes [42,110]. However, pretreatment of mung bean seeds with GA and SA showed an improvement in overall growth and antioxidant system but also prevented the Cd induced alterations in roots under Cd stress conditions [43]. Furthermore, application of JA and GA mitigated Cd induced toxicity in Cicer arietinum by minimizing ROS accumulation via increased antioxidants and osmolytes production under Cd stress [44]. Besides organic and inorganic forms of Se, it can also be applied exogenously in the form of nano-particles (NPs). However, the literature available on Se so far indicates the effectiveness of Se in the removal of various abiotic stresses. Reports further enlightened that supplementation of SeNPs and ZnONPs (Zinc oxide) in combination improved germination rate and seedling growth by inducing the endogenous levels of ABA and GA in Brassica napus [41]. However, the application of these signaling molecules (Phytohormones) impart tolerance to various abiotic stresses through interaction with other signaling molecules and promoting signal transduction cascades in plants [111]. Biochemical and molecular studies have revealed that defense response facilitated by phytohormones such as ETH and JA play an important role in Se resistance and accumulation [112]. Reports suggest that selenite has been shown to decrease NO level in the concentration dependent manner in Arabidopsis root [113]. It has been also reported that application of NO and SeNPs regulated the expression of DREB1A gene in Cichorium intybus [114]. However, the endogenous production of ABA under Cd stress also play role in the regulation of various Cd responsive genes such as HsfA4c, HMA4 in roots and HMA2, HMA3, CAD, and NAS, suggesting its role in the accumulation of more Cd in apoplast region [115].
Studies also emphasized the role of Se and AUX on morphological and biochemical attributes and genotoxicity under As stress in rice plant [116]. It has been reported that presence of PGRs in the external growth medium of plant has altered ion homeostasis (K+, Na+ Ca2+) in the root tissues and mesocotyle [17]. Reports also indicated that both JA and ETH are important for Se resistance in Arabidopsis plant [14]. Exogenous GR24, a source of strigolactone also reduced Cd toxicity by inducing the production of NO and also elevated the levels of different antioxidants in Hordeum vulgare [117]. Application of IAA also increased the phytoextraction of Cd/U from soil in Helianthus annuus [118]. The roots are the foremost organ of plant that reflect heavy metal stress. Thus, the knowledge about the interaction between heavy metal stress and AUX homeostasis is of utmost significance. Involvement of phytohormones in the amelioration of HM stress and their response to different HMs has been studied (Table 2) and discussed in detail as below:
In recent times, participation of plant growth regulators (PGRs in the establishment and activation of plant defense mechanisms to HMs exposure has been largely studied [128,129,130]. Accumulating evidence indicates that exogenous ABA can stimulate the transport of Pb2+ from underground to aerial parts of herbaceous plants [120]. Exogenous ABA addition enhanced activities of enzymatic antioxidants that relieve oxidative stress by scavenging ROS produced due to Pb toxicity in Atractylodes macrocephala [131]. SA interacts with other plant hormones, such as AUX, ABA, and GA, and encourages the production of antioxidant chemicals and enzymes in response to stress, particularly HM stress [132].
Figure 2.
Supplementation of Se and phytohormones ameliorated heavy metals toxicity via reduced uptake of HMs and activation of various defence systems.
Figure 2.
Supplementation of Se and phytohormones ameliorated heavy metals toxicity via reduced uptake of HMs and activation of various defence systems.
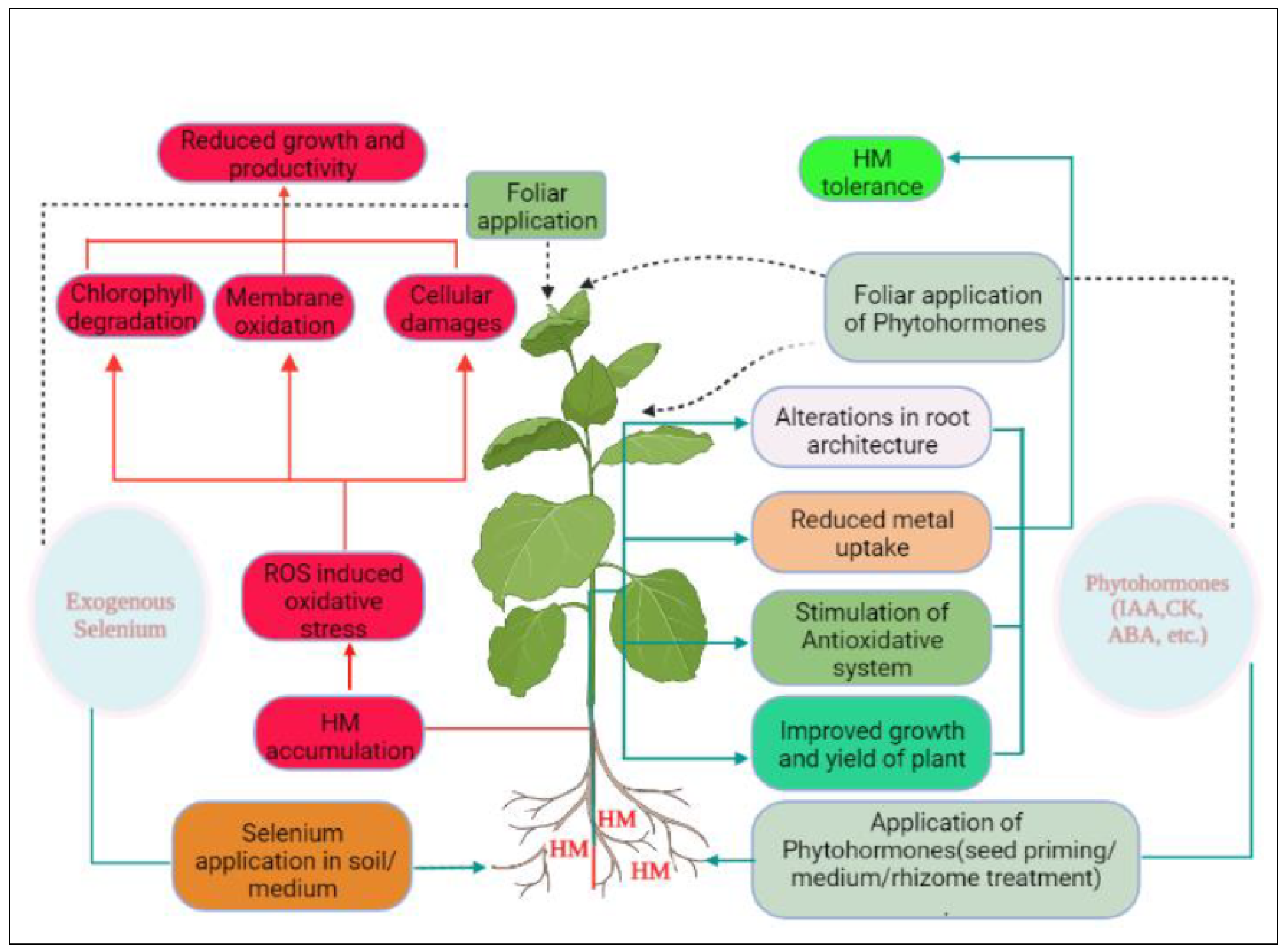
Phytohormones play crucial role in the growth and developmental processes of plants when exposed to a variety of abiotic stresses. Some of these signaling molecules are produced endogenously during stress conditions. However, their exogenous application to different abiotic stresses including HM stress shows major contribution towards developing tolerance. Primarily, PGRs interfere with metal uptake via altering root architecture, thus resisting the uptake and accumulation of metals to the plant body. Moreover, Se supplementation also induces the endogenous production of different growth regulators under different stresses and thus both Se and PGRs help in developing tolerance to stress conditions by regulating different mechanisms in plants to minimize the oxidative damage.
6. Molecular and Proteomic Interaction of Selenium under Heavy Metals
Work on Se indicated that Se plays beneficial role in enhancement of abiotic stress tolerance [13]. Proteomics is acknowledged as an effective method for understanding how plants respond to abiotic stress, comprising toxic metal such as Cu, As, Al, Cr etc. [133,134]. Analysis of more than hundred (110) proteins to understand the proteomic response of rice seedling under different level of Cr stress was carried out by [135]. In a recent study, RNA sequencing and proteome profiling along with morphological demonstration were conducted to explore molecular cross talk of Se mediated As stress amelioration [85].There are numerous transporters family that play crucial role in response to As stress in plants specifically for uptake of As in root, phloem movements, xylem loading and unloading in cells and subcellular organelles specific distribution [85]. The Germin protein family participates in numerous metabolic and stress tolerance mechanisms. Selenium helps the plants to cope with As stress by regulating various stress responsive genes [85]. When exposed to Cd, rice developed Cd responsive miRNA, and the translational levels of several targeted genes were inversely linked with the corresponding miRNA, and suggesting the molecular role under HM stress resistance in plant [136]. At molecular level, Se also plays an important role in developing HMs tolerance in plants by regulating numerous stress related genes. However, involvement of ETH and Se in the regulation of genes which encode enzymes of antioxidants has been observed in mustard subjected to As stress [119]. Significant enhancement in the regulation of WRKY1, PAL and 4CL genes was observed in Mimordica charantia when exposed to slightly higher concentrations of nano-Se [137]. Increased expression of antioxidant genes, PAL and CHS was also noticed in Cr stressed mustard [138]. Selenium treatment significantly up-regulated the expression of S transporters (SLUTs) genes involved in the transport of Se [139]. Application of Se at 25µM in broccoli has shown negative effects at both translational and transcriptional level. Proteins of BIN family which express during stress conditions and heat shock proteins (HSPs) were badly affected in Se treated plants. Moreover, down regulation of different transcription genes such as MYB (28, 34), BCAT4, MAM1, CYP79B2 and FMO2 was also detected in Se treated plants [28]. Regulation of various antioxidant genes and thiol metabolism genes in response to Se was also observed in Oryza sativa exposed to As stress [90]. Exogenous Se considerably increased the expression levels of NtPT2 gene in tobacco [110]. Selenium also up-regulated different genes responsible for seed germination and seedling growth in rapeseed [41]. In Glycine max, SeNPs treatment significantly upregulated various stress marker genes in response to As stress [23]. However, the addition of Se in rice cell culture in the presence of Cd positively up-regulated OsHMA3 and OsPAL, OsCoMT and Os4CL3 genes responsible for Cd accumulation in vacuole and lignin biosynthesis. On the other hand, Se also represses the expression of genes (OsNramp5, OsLCT1) involved in Cd uptake and transport [19]. Moreover, the expression of SBP1 protein in Arabidopsis seedlings indicated its role in alleviation of Cd induced oxidative stress by promoting its binding to Cd [140]. Furthermore, foliar treatment of Se induced genes responsible for Se accumulation and transport to shoots and differentially expressed the proteins related to Se uptake, aggregation, antioxidants and heat shock proteins [141]. Meanwhile, Se treatment upregulated genes responsible for cell wall lignification and triggered resistance to Cd by inducing the expression of BZR1, LOX3, and NCDE1 genes [18]. Selenium supplementation negatively affected the expression of genes (TaNramp5, TaHMA2) responsible for Cd uptake and translocation but also upregualted TaHMA3-gene required for Cd translocation to vacuole in both root and shoot in wheat [84].
An important areas of the plants to respond or increase tolerance potential to stresses including HM is the regulation of genes at both transcriptional and translational level. During stress conditions, activation/expression of different stress responsive proteins or genes occur, which help the plant to generate tolerance ability for the survival against stress. Selenium application in response to metalloid toxicity, down-regulates various genes that are responsible for uptake and transport of HM in plants, thus indicating its importance in plant stress tolerance. Moreover, Se also regulates the expression of various antioxidant genes that are the major players for developing the tolerance and hence protection of plants to abiotic stress.
7. Selenium Interplay with Secondary Metabolites (Phenolics, N-containing Metabolites and Terpenes)
In the era of climate change, abiotic stress hampered the growth and development of plant. Plants develop several defense mechanisms in order to cope with such conditions. Plant secondary metabolites (SMs) display their functions against stresses [142,143]. The amounts of SMs under stress conditions are slightly higher and are involved in signaling, thereby regulating the activity of genes responsible for plant defense [144]. Among different SMs, terpenes, phenolics and compounds of nitrogen are widely known for their protective affects against various environmental constraints [145]. To investigate the role of these compounds (SMs) in plant defense, the exogenous use of monoterpenoid on Quercus suber leaves indicated its role in endogenous terpene accumulation and also improved photosynthesis of plants subjected to heat stress [146]. The production of plant metabolites is regulated genetically, environmental factors and also the mineral status of plant [147]. Plants accumulate different types of metabolites under varying conditions. For instance, production of flavonoids during drought stress in Arabidopsis thaliana is thought to be involved in drought tolerance [148]. Matricaria chamomilla plants subjected to copper (Cu) and Cd stress have shown more phenolics content than the non-stressed plants [149]. Similarly, increased levels of phenolics were observed in Vaccinium corymbosum plants facing Cd stress [150]. Increased biosynthesis of phenolics was also observed in Kandelia obovata under HM stress [151]. Increased accumulation of different type of SMs was also noticed in Brassica juncea, which has higher Cd tolerance capability without any use of external mitigators [152]. In order to overcome plants from metal toxicity, use of elicitors can be done to enhance the SMs production [153]. The effect of Se on SMs production and their function in plants has been studied (Table 3) and discussed in detail as below:
Research on selenium has shown that it plays a crucial part in plant metal tolerance by posing restrictions on the uptake of metals from soil by changing root architecture system and also prevents their translocation to leaves [15]. Selenium at low concentrations show beneficial effects on plant secondary metabolites, as it enhanced the levels of carotenoids and phenolic compounds in rice [159]. Selenium nanoparticles supplementation in bittermelon induced the activity of phenylalanine ammonia-lyase (PAL), an important enzyme involved in the bio-synthesis of phenolic compounds [137]. Addition of Se upregulated PAL and Chalcone synthase genes and increased the contents of phenolics, flavonoids and anthocyanin compounds in mustard under chromium (Cr) stress [13,27]. Report showed that Se decreased the accumulation of glucosinolates (GLS) in broccoli [29]. In Se treated Arabidopsis thaliana, reduction in GLS and carotenoids content was observed in shoots by suppressing their biosynthetic genes [30]. Selenium application negatively affected the accretion of GLS by suppressing genes involved in GLS biosynthesis and their precursors in broccoli [28]. Increment in the levels of plant essential oils (secondary metabolites) in response to Se was also observed in Melissa officinalis [16]. However, the supplementation of Se at low doses increased the accumulation of GLS in Brassica oleracea [25]. In Allium sativum, treatment with Se improved PAL activity and increased the content of phenolics under salt stress [157]. Selenium application increased the levels of total phenolics and flavonoids in Zea mays and Valerianella locusta [155,158]. Recently, it has been reported that spraying of SeNPs on the leaves of Mentha suaveolens induced the accumulation of various types of essential oils under salt stress [154]. Reports also indicate that Se ameliorated Cd toxicity of Brassica juncea by improving its antioxidant system and SMs status [88]. Selenium also enhances the biosynthesis of flavonoids and tocopherols which are also involved in relieving oxidative stress of different abiotic stresses [76]. Selenium also attenuated As toxicity in Oryza sativa by inducing the aggregation of phenolics in both underground and aerial parts of plant [156].
Secondary metabolites, an important class of plant defense compounds which are synthesized by plants as a response to different types of stress conditions that can cause harm to plants. Plant exudates such as tannins, resins, gums etc. are the common secretions which are mostly related to stress factors i.e., both biotic and abiotic. However, the root exudates also limit the uptake and translocation of HMs, thus help in protecting plants against HMs toxicity. Based on the results of the aforesaid studies on Se and secondary metabolites, it has been concluded that Se is involved in improving the antioxidant system of plants by enhancing the synthesis of different types of SMs which play an important part in plant defense system.
8. Selenium Interaction with Mineral Nutrients under Heavy Metal Stress
Selenium serves as an essential micro-nutrient for plants and plays an important role in various metabolic pathways in plant life. However, its accumulation in plants varies from species to species. Moreover, at high concentrations, Se shows toxic effects on plants, but at optimal concentration it is reported to be safe for plants. However, some plant species can accumulate higher concentration of Se in their body. Selenium is known for its both beneficial and toxic effects on plants. Apart from developing resistance to metal/metalloid uptake, it is also known to increase nutrient absorption and accumulation in plants. Reports indicated that the coordinative supply of Se and S in mung bean increased the uptake of various mineral nutrients such as NPK, Fe, Zn and S under drought stress [32]. Selenium helps plants in relieving oxidative stress by acting as an antioxidant that also benefits plants in improving their nutritional status by enhancing uptake and aggregation of mineral nutrients [160]. Significant increment in uptake of some nutrients such as Fe, Mn and Cu was also observed in radish subjected to Cd stress in presence of Se [31]. However, the supplementation of Se in the nutrient solution favoured more uptakes of P and Ca from the media, indicating its role in maintenance of mineral nutrition in Zea mays ([161]. Application of Se significantly enhanced the uptake of micro-nutrients that regulated various metabolic processes to develop tolerance against As stress [156]. Moreover, Se treatment improves the nutritional quality of crops by accumulating more Se in edible parts of plants [34]. Supplementation of Se at different concentrations showed fluctuations in the contents of both macro and micro elements in different species [162].
Furthermore, Se at higher concentration affected the uptake of micronutrients in Eruca sativa [163]. However, improvement in N and Ca content was also observed in wheat subjected to exogenous Se treatment [164]. Selenium supplementation significantly up-regulated the uptake of various essential mineral elements in Brassica campestris by modifying roots architecture to reduce Cr uptake [165]. Introduction of biosynthetic form of SeNPs to different HMs (Cd, Pb, Hg) stress improved the nutritional status of Brassica chinensis by enhancing the uptake of nutrient elements [33].
Availability of mineral nutrients is one among the chief processes in the life cycle of a plant to regulate all its growth and developmental processes. However, the uptake of these mineral elements is severely affected by the soils contaminated with metals, thus affecting the overall growth and metabolism of plants. On the other hand, Se is directly involved in the uptake of these nutrients from soil by inhibiting metal uptake and therefore helps the plants to restore the activities of all its physiological, biochemical and developmental processes to withstand against such harsh environmental constraints.
9. Conclusions
Rising concentrations of HMs in the soil and their alarming negative effects on plants and thus on other living life forms attracted scientists in the field of research more towards this environmental constraint in the present era of science. Due to its increasing threat to the agronomy field, various methods have been adopted by scientists to overcome this problem. Exogenous supplementation of Se in the form of selenite, selenate, SeCys, SeMT and SeNPs has been successfully done to cope the harsh effects of HM stress in plants. Selenium is directly involved in the alleviation of HM toxicity by either inhibiting or limiting its uptake and translocation from soil to aerial parts of plants. Primarily, Se is involved in the improvement of plants' growth and photosynthesis by improving nutritional status, osmoprotectants concentration, antioxidant metabolism and secondary metabolites under metal stress conditions. However, the interaction of Se with phytohormones under metal stress studied so far is very least. To better understand and for more effective results of Se under HM stress, exploration of interaction between plant growth regulators and Se under metal toxicity is required. Further, the crosstalk between HMs, Se and phytohormones is needed to be understood. By understanding this problem, it will become more feasible to overcome such threats and in future, crops with higher nutritional quality and yield can be produced. And hence, this approach can also become useful to reduce global food scarcity/hunger.
Authors Contribution
Conceptualization, M.A and N.A.K Validation, A.R., S.N.I., and M.A. writing-Original Draft, M.A., A.R., S.N.I., and M.A. Writing and editing, M.A and N.A.K., A.R., S.N.I., and M.A Supervision M.A and N.A.K; funding acquisition, M.A. All authors read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
“The authors declare no conflict of interest”.
References
- Hasan, M.M.; Uddin, M.N.; Ara-Sharmeen, F.I.; Alharby, H.; Alzahrani, Y.; Hakeem, K.R. Assisting phytoremediation of heavy metals using chemical amendments. Plants 2019, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Rather, B.A.; Mir, I.R.; Masood, A.; Anjum, N.A.; Khan, N.A. Nitric oxide pre- treatment advances seed germination and alleviates copper-induced photosynthetic inhibition in Indian mustard. Plants 2020, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Rai, P.; Guerriero, G.; Sharma, S.; Corpas, F.J.; Singh, V.P. Silicon induces adventitious root formation in rice under arsenate stress with involvement of nitric oxide and indole-3-acetic acid. J. Exp. Bot. 2021, 72, 4457–4471. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants—role of plant growth regulators. Protoplasma 2015, 252, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 1, 1–18. [Google Scholar] [CrossRef]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicol. Environ. Saf. 2021, 223, 112578. [Google Scholar] [CrossRef]
- Asgher, M.; Ahmed, S.; Sehar, Z.; Gautam, H.; Gandhi, S.G.; Khan, N.A. Hydrogen peroxide modulates activity and expression of antioxidant enzymes and protects photosynthetic activity from arsenic damage in rice (Oryza sativa L.). J. Hazard. Mater. 2021, 401, 123365. [Google Scholar] [CrossRef]
- Upadhyay, M.K.; Majumdar, A.; Barla, A.; Bose, S.; Srivastava, S. An assessment ofarsenic hazard in groundwater–soil–rice system in two villages of Nadia district,West Bengal, India. Environ. Geochem. Health 2019, 41, 2381–2395. [Google Scholar] [CrossRef]
- Tyagi, N.; Raghuvanshi, R.; Upadhyay, M.K.; Srivastava, A.K.; Suprasanna, P.; Srivastava, S. Elemental (As, Zn, Fe and Cu) analysis and health risk assessment of rice grains and rice based food products collected from markets from different cities of Gangetic basin, India. J. Food Compos. Anal. 2020, 93, 103612. [Google Scholar] [CrossRef]
- Asgher, M.; Sehar, Z.; Rehaman, A.; Rashid, S.; Ahmed, S.; Per, T.S.; Alyemeni, M.N.; Khan, N.A. Exogenously-applied L-glutamic acid protects photosynthetic functions and enhances arsenic tolerance through increased nitrogen assimilation and antioxidant capacity in rice (Oryza sativa L.). Environ. Pollut. 2022, 301, 119008. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Verma, S.; Vyas, D.; Per, T.S.; Khan, N.A. Ethylene and polyamines in counteracting heavy metal phytotoxicity: a crosstalk perspective. J. Plant Growth Regul. 2019, 37, 1050–1065. [Google Scholar] [CrossRef]
- Hasan, M.M.; Alharbi, B.M.; Alhaithloul, H.A.S.; Abdulmajeed, A.M.; Alghanem, S.M.; Al-Mushhin, A.A.M.; Jahan, M.S.; Corpas, F.J.; Fang, X.-W.; Soliman, M.H. Spermine-mediated tolerance to selenium toxicity in wheat (Triticum aestivum L.) depends on endogenous nitric oxide synthesis. Antioxidants 2021, 10, 1835. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Mahmud, J.A.; Nahar, K, Fujita, M. Selenium in plants: Boon or Bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Tamaoki, M.; Freeman, J.L.; Pilon-Smits, E.A.H. Cooperative ethylene and jasmonic acid signaling regulates selenite resistance in Arabidopsis. Plant Physiol. 2008, 146, 1219e1230. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; García-Caparrós, P.; Parvin, K.; Zulfiqar, F.; Ahmed, N.; Fujita, M. Selenium supplementation and crop plant tolerance to metal/metalloid toxicity. Front. Plant Sci. 2022, 12, 792770. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, S.; Enteshari, S.; Yousefifard, M. 'The effect of selenium on physiologic and morphologic properties of Melissa officinalis L.'. Iran. J. Plant Physiol. 2020, 10, 3125–3134. [Google Scholar]
- Pazurkiewicz-Kocot, K.; Galas, W.; Kita, A. The effect of selenium on the accumulation of some metals in Zea mays L. plants treated with indole-3-acetic acid. Cell. Mol. Biol. Lett. 2003, 8, 97–104. [Google Scholar]
- Li, D.; Zhou, C.; Ma, J.; Wu, Y.; Kang, L.; An, Q.; Pan, C. Nanoselenium transformation and inhibition of cadmium accumulation by regulating the lignin biosynthetic pathway and plant hormone signal transduction in pepper plants. J. Nanobiotech. 2021, 19, 1–14. [Google Scholar] [CrossRef]
- Cui, J.; Liu, T.; Li, Y.; Li, F. Selenium reduces cadmium uptake into rice suspension cells by regulating the expression of lignin synthesis and cadmium-related genes. Sci. Total Environ. 2018, 644, 602–610. [Google Scholar] [CrossRef]
- Sun, H.W.; Ha, J.; Liang, S.X.; Kang, W.J. Protective role of selenium on garlic growth under cadmium stress. Commun. Soil Sci. Plan. 2010, 41, 1195–1204. [Google Scholar] [CrossRef]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Babashpour-Asl, M.; Farajzadeh-Memari-Tabrizi, E.; Yousefpour-Dokhanieh, A. "Foliar-applied selenium nanoparticles alleviate cadmium stress through changes in physio-biochemical status and essential oil profile of coriander (Coriandrum sativum L.) leaves. Environ. Sci. Pollut. Res. 2022, 29, 80021–80031. [Google Scholar] [CrossRef]
- Zeeshan, M.; Hu, Y.X.; Iqbal, A.; Salam, A.; Liu, Y.X.; Muhammad, I.; Ahmad, S.; Khan, A.H; Hale, B.; Wu, H.Y.; Zhou, X.B. Amelioration of AsV toxicity by concurrent application of ZnO-NPs and Se-NPs is associated with differential regulation of photosynthetic indexes, antioxidant pool and osmolytes content in soybean seedling. Ecotoxicol. Environ. Saf. 2021, 225, 112738. [Google Scholar] [CrossRef]
- Atarodi, B.; Fotovat, A.; Khorassani, R.; Keshavarz, P. Study of wheat (Triticum aestivum L.) response to selenium application under cadmium stress. Environmental Stresses in Crop Sciences 2019, 12, 291–305. [Google Scholar]
- Saeedi, M.; Soltani, F.; Babalar, M.; Izadpanah, F.; Wiesner-Reinhold, M.; Baldermann, S. Selenium fortification alters the growth, antioxidant characteristics and secondary metabolite profiles of cauliflower (Brassica oleracea var. botrytis) cultivars in hydroponic culture. Plants 2021, 10, 1537. [Google Scholar] [CrossRef]
- Chu, J.; Yao, X.; Zhang, Z. Responses of wheat seedlings to exogenous selenium supply under cold stress. J. Biol. Trace Elem. Res. 2010, 136, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Tian, M.; Xu, X.; Liu, F.; Fan, X.; Pan, S. Untargeted metabolomics reveals predominant alterations in primary metabolites of broccoli sprouts in response to pre-harvest selenium treatment. Food Res. Int. 2018, 111, 205–211. [Google Scholar] [CrossRef]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Selenium influences glucosinolate and isothiocyanates and increases sulfur uptake in Arabidopsis thaliana and rapid-cycling Brassica oleracea. J. Agric. Food Chem. 2013, 61, 202–209. [Google Scholar] [CrossRef]
- Sams, C.E.; Panthee, D.R.; Charron, C.S.; Kopsell, D.A.; Yuan, J.S. Selenium regulates gene expression for glucosinolate and carotenoid biosynthesis in Arabidopsis. J. Am. Soc. Hortic. Sci. 2011, 136, 23–34. [Google Scholar] [CrossRef]
- Amirabad, A.S.; Behtash, F.; Vafaee, Y. Selenium mitigates cadmium toxicity by preventing oxidative stress and enhancing photosynthesis and micronutrient availability on radish (Raphanus sativus L.) cv. Cherry Belle. Environ. Sci. Pollut. Res. 2020, 27, 12476–12490. [Google Scholar] [CrossRef]
- Aqib, M.; Nawaz, F.; Majeed, S.; Ghaffar, A.; Ahmad, K.S.; Shehzad, M. A.; Usmani, M.M. Physiological insights into sulfate and selenium interaction to improve drought tolerance in mung bean. Physiol. Mol. Bio. Plants 2021, 27, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dong, Y.; Zhu, N.; Jina, H. Foliar application of biosynthetic nano-selenium alleviates the toxicity of Cd, Pb, and Hg in Brassica chinensis by inhibiting heavy metal adsorption and improving antioxidant system in plant. Ecotoxicol. Environ. Saf. 2022, 240, 113681. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Wen, X.; Yi, N.; Liu, Y.; Wu, J.; Li, H.; Liu, G. Effect of foliar application of silicon, selenium and zinc on heavy metal accumulation in wheat grains in field studies. Environ. Pollut. Bioavail. 2022, 34, 246–252. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium in higher plants: physiological role, antioxidant metabolism and abiotic stress tolerance. J. Plant Sci. 2010, 5, 354–375. [Google Scholar] [CrossRef]
- Atici, O.; Agar, G.; Battal, P. Changes in phytohormone contents in chickpea seeds germinating under lead or zinc stress. Algologia 2005, 49, 215–222. [Google Scholar] [CrossRef]
- Maksymiec, W.; Krupa, Z. The effects of short-term exposition to Cd, excess Cu ions and jasmonate on oxidative stress appearing in Arabidopsis thaliana. Environ. Exp. Bot. 2006, 57, 187–194. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Sesin, V.; Kisiala, A.; Emery, R.N. Phytohormonal roles in plant responses to heavy metal stress: implications for using macrophytes in phytoremediation of aquatic ecosystems. Environ. Toxicol. Chem. 2021, 40, 7–22. [Google Scholar] [CrossRef]
- Malheiros, R.S.P.; Costa, L.C.; Ávila, R.T.; Pimenta, T.M.; Teixeira, L.S.; Brito, F.A.L.; Ribeiro, D.M. Selenium downregulates auxin and ethylene biosynthesis in rice seedlings to modify primary metabolism and root architecture. Planta 2019, 250, 333–345. [Google Scholar] [CrossRef]
- Malheiros, R.S.P.; Gonçalves, F.C.M.; Brito, F.A.L.; Zsögön, A.; Ribeiro, D.M. Selenomethionine induces oxidative stress and modifies growth in rice (Oryza sativa L.) seedlings through effects on hormone biosynthesis and primary metabolism. Ecotoxicol. Environ. Saf. 2020, 189, 109942. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Wang, C.; Hashem, A.M.; Tabl, K.M.; Nishawy, E.; Wang, B. Selenium and zinc oxide nanoparticles modulate the molecular and morpho-physiological processes during seed germination of Brassica napus under salt stress. Ecotoxicol. Environ. Saf. 2021, 225, 112695. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wei, Y.; Sun, S.; Wang, J.; Wang, W.; Han, D.; Fu, Y. Selenium modulates the level of auxin to alleviate the toxicity of cadmium in tobacco. Int. J. Mol. Sci. 2019, 20, 3772. [Google Scholar] [CrossRef]
- Hassan, M.; Israr, M.; Mansoor, S.; Hussain, S.A.; Basheer, F.; Azizullah, A.; Ur Rehman, S. Acclimation of cadmium-induced genotoxicity and oxidative stress in mung bean seedlings by priming effect of phytohormones and proline. Plos One 2021, 16, e0257924. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P., Raja, V., Ashraf, M. et al. Jasmonic acid (JA) and gibberellic acid (GA3) mitigated Cd-toxicity in chickpea plants through restricted Cd uptake and oxidative stress management. Sci. Rep. 2021, 11, 19768. [CrossRef] [PubMed]
- Alves, L.R.; Dos Reis, A.R.; Prado, E.R.; Lavres, J.; Pompeu, G.B.; Azevedo, R.A.; Gratao, P.L. New insights into cadmium stressful-conditions: Role of ethylene on selenium-mediated antioxidant enzymes. Ecotoxicol. Environ. Saf. 2019, 186, 109747. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.U.; Li, Y.; Hussain, S.; Hussain, B.; Riaz, L.; Ashraf, M.N.; Cheng, H. Role of phytohormones in heavy metal tolerance in plants: A review. Ecol. Indic. 2023, 146, 109844. [Google Scholar] [CrossRef]
- Frankenberger, J.W.T.; Karlson, U. Campaigning for bioremediation. Chemtech. 1994, 24, 45–51. [Google Scholar]
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta - Gen. Subj. 2018, 1862, 2333–2342. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef]
- Parker, D.R.; Feist, L.J.; Varvel, T.W. Selenium phytoremediation potential of Stanleya pinnata. Plant Soil 2003, 249, 157–165. [Google Scholar] [CrossRef]
- Zayed, A.; Lytle, C.M.; Terry, N. Accumulation and volatilization of different chemical species of selenium by plants. Planta 1998, 206, 284–292. [Google Scholar] [CrossRef]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; Smith, B.M.; Thomas, B.; Broadley, M.R. Interactions between selenium and sulfur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Arvy, M.P. Selenate and selenite uptake and translocation in bean plants (Phaseolus vulgaris). J. Expt. Bot. 1993, 44, 1083–1087. [Google Scholar] [CrossRef]
- Freeman, J.L.; Quinn, C.F.; Marcus, M.A.; Fakra, S.; Pilon-Smits, E.A. Selenium-tolerant diamondback moth disarms hyperaccumulator plant defense. Current Biol. 2006, 16, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium speciation from food source to metabolites: a critical review. Anal. Bioanal. Chem. 2006, 385, 1304–1323. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A.; Shrift, A. Selenium: toxicity and tolerance in higher plants. Biol. Rev. 1982, 57, 59–84. [Google Scholar] [CrossRef]
- Zhang, L.; Chu, C. Selenium uptake, transport, metabolism, reutilization, and biofortification in rice. Rice 2022, 15. [Google Scholar] [CrossRef]
- Wang, K.; Wang, Y.; Li, K.; Wan, Y.; Wang, Q.; Zhuang, Z.; Guo, Y.; Li, H. "Uptake, translocation and biotransformation of selenium nanoparticles in rice seedlings (Oryza sativa L.).". J. Nanobiotechnol. 2020, 18, 103. [Google Scholar] [CrossRef]
- Trippe, III.; Richard, C.; Pilon-Smits, E.A.H. "Selenium transport and metabolism in plants: Phytoremediation and biofortification implications. J. Hazard. Mater. 2021, 404, 124178. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Eukaryotic selenoproteins and selenoproteomes. Biochim. Biophys. Acta - Gen. Subj. 2009, 1790, 1424–1428. [Google Scholar] [CrossRef]
- Nancharaiah, Y. V.; Lens, P.N.L. Selenium biomineralization for biotechnological applications. Trends Biotechnol. 2015, 33, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.C.; Luz, L.M.; Nascimento, V.L.; Araujo, F.F.; Santos, M.N.S.; França, C.F.M.; Silva, T.P.; Fugate, K.K.; Finger, F.L. Selenium-ethylene interplay in postharvest life of cut flowers. Front. Plant Sci. 2020, 11, 584698. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; Copat, C.; Longo, S.; Oliveri Conti, G.; Grasso, A.; Arena, G.; Ferrante, M. First data on trace elements in Haliotistuberculata (Linnaeus, 1758) from southern Italy: Safety issues. Food Chem. Toxicol. 2015, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Copat, C.; Grasso, A.; Fiore, M.; Cristaldi, A.; Zuccarello, P.; Signorelli, S.S.; Ferrante, M. Trace elements in seafood from the Mediterranean sea: An exposure risk assessment. Food Chem. Toxicol. 2018, 115, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhao, R.; Zhao, W.; Fu, R.; Guo, J.; Bi, N.; Zhang, J. Effects of arbuscular mycorrhizal fungi on maize (Zea mays L.) and sorghum (Sorghum bicolor L. Moench) grown in rare earth elements of mine tailings. Appl. Soil Ecol. 2013, 72, 85–92. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: an overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Lv, X.; Wang, G. Effects of Se and Zn supplementation on the antagonism against Pb and Cd in vegetables. Environ. Int. 2004, 30, 167–172. [Google Scholar] [CrossRef]
- Nawaz, F.; Ashraf, M.Y.; Ahmad, R.; Waraich, E.A.; Shabbir, R.N. Selenium (Se) regulates seedling growth in wheat under drought stress. Adv. Chem. 2014, 143567. [Google Scholar] [CrossRef]
- Gui, J.Y.; Rao, S.; Huang, X.; Liu, X.; Cheng, S.; Xu, F. Interaction between selenium and essential micronutrient elements in plants: A systematic review. Sci. Total Environ. 2022, 853, 158673. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255e277. [Google Scholar] [CrossRef]
- Filek, M.; Koscielniak, J.; Labanowska, M.; Bednarska, E.; Bidzinska, E. Selenium-induced protection of photosynthesis activity in rape (Brassica napus) seedlings subjected to cadmium stress:Fluorescence and EPR measurements. Photosynth. Res. 2010, 105, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Tobiasz, A.; Walas, S.; Filek, M.; Mrowiec, H.; Samsel, K.; Sieprawska, A.; Hartikainen, H. Effect of selenium on distribution of macro- and micro-elements to different tissues during wheat. Biol. Plant. 2014, 58, 370–374. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Elem. Res. 2012, 49, 248–61. [Google Scholar] [CrossRef] [PubMed]
- Lanza, M.G.D.B.; Reis, A.R.dos. Roles of selenium in mineral plant nutrition: ROS scavenging responses against abiotic stresses. Plant Physiol. Biochem. 2021, 164, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron. Soil Sci. 2014, 60, 1593–1624. [Google Scholar] [CrossRef]
- Cakir, O.; Candar-Cakir, B.; Zhang, B.H. Small RNA and degradome sequencing reveals important microRNA function in Astragalus chrysochlorus response to selenium stimuli. Plant Biotechnol J. 2016, 14, 543–556. [Google Scholar] [CrossRef]
- Aroiee, H.; Shekari, L.; Mirshekari, A. Effects of selenium on damage of heavy metals in germination, growth and antioxidant activities of cucumber (Cucumis sativus L.) seedling. Iran. J. Seed Sci. Res. 2019, 6, 269–286. [Google Scholar] [CrossRef]
- Cartes, P.; Jara, A.A.; Pinilla, L.; Rosas, A.; Mora, M.L. Selenium improves the antioxidant ability against aluminium-induced oxidative stress in ryegrass roots. Ann. Appl. Biol. 2010, 156, 297–307. [Google Scholar] [CrossRef]
- Malik, J.A.; Kumar, S.; Thakur, P.; Sharma, S.; Kaur, N.; Kaur, R.; Pathania, D.; Bhandhari, K.; Kaushal, N.; Singh, K.; Srivastava, A.; Nayyar, H. Promotion of growth in mungbean (Phaseolus aureus Roxb.) by selenium is associated with stimulation of carbohydrate metabolism. Biol. Trace Elem. Res. 2011, 143, 530–539. [Google Scholar] [CrossRef]
- Mroczek-Zdyrska, M.; Strubińska, J.; Hanaka, A. selenium improves physiological parameters and alleviates oxidative stress in shoots of lead-exposed Vicia faba L. minor plants grown under phosphorus-deficient conditions. J. Plant Growth Regul. 2017, 36, 186–199. [Google Scholar] [CrossRef]
- Wu, Z.; Yin, X.; Bañuelos, G.S.; Lin, Z.Q.; Liu, Y.; Li, M.; Yuan, L. Indications of selenium protection against cadmium and lead toxicity in oilseed rape (Brassica napus L.). Front. Plant Sci. 2016, 7, 1875. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhao, P.; Nie, Z.; Shi, H.; Li, C.; Wang, Y.; Qin, S.; Qin, X.; Liu, H. Selenium supply alters the subcellular distribution and chemical forms of cadmium and the expression of transporter genes involved in cadmium uptake and translocation in winter wheat (Triticum aestivum). BMC Plant Boil. 2020, 20, 1–12. [Google Scholar]
- Chauhan, R.; Awasthi, S.; Indoliya, Y.; Chauhan, A.S.; Mishra, S.; Agrawal, L.; Tripathi, R.D. Transcriptome and proteome analyses reveal selenium mediated amelioration of arsenic toxicity in rice (Oryza sativa L.). J. Hazard. Mater. 2020, 122122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Dong, Y.Y.; Feng, L.Y.; Deng, Z.L.; Xu, Q.; Tao, Q.; Wang, C.Q.; Chen, Y.E.; Yuan, M.; Yuan, S. Selenium enh;nces cadmium accumulation capability in two mustard family species—Brassica napus and B. Juncea. Plants 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Azizi, I.; Esmaielpour, B.; Fatemi, H. Effect of foliar application of selenium on morphological and physiological indices of savory ( Satureja hortensis ) under cadmium stress. Food Sci. Nutr. 2020, 8, 6539–6549. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Abd Allah, E.F.; Hashem, A.; Sarwat, M.; Gucel, S. Exogenous application of selenium mitigates cadmium toxicity in Brassica juncea L. (Czern and Cross) by up-regulating antioxidative system and secondary metabolites. J. Plant Growth Regul. 2016, 35, 936–950. [Google Scholar] [CrossRef]
- Xu, H.; Yan, J.; Qin, Y.; Xu, J.; Shohag, M.J.I.; Wei, Y.; Gu, M. Effect of different forms of selenium on the physiological response and the cadmium uptake by rice under cadmium stress. Int. J. Environ. Res. Public Health 2020, 17, 6991. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, R.P.; Singh, P.K.; Awasthi, S.; Chakrabarty, D.; Trivedi, P.K.; Tripathi, R.D. Selenium ameliorates arsenic induced oxidative stress through modulation of antioxidant enzymes and thiols in rice (Oryza sativa L.). Ecotoxicol. 2014, 23, 1153–1163. [Google Scholar] [CrossRef]
- Sun, H.; Wang, X.; Li, H.; Bi, J.; Yu, J.; Liu, X.; Rong, Z. Selenium modulates cadmium-induced ultrastructural and metabolic changes in cucumber seedlings. RSC Adv. 2020, 10, 17892–17905. [Google Scholar] [CrossRef]
- Martins, J.P.R.; Souza, A.F.C.; Rodrigues, L.C.A.; Braga, P.C.S.; Gontijo, A.B.P.L.; Falqueto, A.R. Zinc and selenium as modulating factors of the anatomy and physiology of Billbergia zebrina (Bromeliaceae) during in vitro culture. Photosynthetica 2020, 58, 1068–1077. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, S.; Jiang, Z.; Wang, Y.; Zhang, Z. Selenium biofortification modulates plant growth, microelement and heavy metal concentrations, selenium uptake, and accumulation in black-grained wheat. Front. Plant Sci. 2021, 12, 748523. [Google Scholar] [CrossRef]
- Shekari, L.; Aroiee, H.; Mirshekari, A.; Nemati, H. Protective role of selenium on cucumber (Cucumis sativus L.) exposed to cadmium and lead stress during reproductive stage role of selenium on heavy metals stress. J. Plant Nutr. 2019, 42, 529–542. [Google Scholar] [CrossRef]
- Naseem, M.; Anwar-ul-Haq, M.; Wang, X.; Farooq, N.; Awais, M.; Sattar, H.; Ahmed Malik, H.; Mustafa, A.; Ahmad, J.; El-Esawi, M.A. Influence of selenium on growth, physiology, and antioxidant responses in maize varies in a dose-dependent manner. J. Food Qual. 2021, 1–9. [Google Scholar] [CrossRef]
- Bakhtiari, M.; Raeisi Sadati, F.; Raeisi Sadati, S.Y. Foliar application of silicon, selenium, and zinc nanoparticles can modulate lead and cadmium toxicity in sage (Salvia officinalis L.) plants by optimizing growth and biochemical status. Environ. Sci. Pollut. Res 2023, 1–11. [Google Scholar] [CrossRef]
- Barman, F.; Kundu, R. Foliar application of selenium affecting pollen viability, grain chalkiness, and transporter genes in cadmium accumulating rice cultivar: A pot study. Chemosphere 2023, 313, 137538. [Google Scholar] [CrossRef]
- Rostami, M.; Abbaspour, H. Effect of selenium on growth and physiological traits of basil plant (Ocimum basilicum L.) under arsenic stress conditions. Rev. Agric. Neotrop. 2019, 6, 30–37. [Google Scholar] [CrossRef]
- Mozafariyan, M.; Shekari, L.; Hawrylak-Nowak, B.; Kamelmanesh, M.M. Protective role of selenium on pepper exposed to cadmium stress during reproductive stage. Biol. Trace Elem. Res. 2014, 160, 97–107. [Google Scholar] [CrossRef]
- Feng, R.; Wang, L.; Yang, J.; Zhao, P.; Zhu, Y.; Li, Y. Underlying mechanisms responsible for restriction of uptake and translocation of heavy metals (metalloids) by selenium via root application in plants. J. Hazard. Mater. 2020, 402, 23570. [Google Scholar] [CrossRef]
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. J. Mol. 2019, 24, 1298. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Alam, M.Z.; McGee, R.; Hoque, M.A.; Ahammed, G.J.; Carpenter-Boggs, L. Effect of arbuscular mycorrhizal fungi, selenium and biochar on photosynthetic pigments and antioxidant enzyme activity under arsenic stress in mung bean (Vigna radiata). Front. Physiol. 2019, 10, 193. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M. Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L) as influenced by potassium supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef]
- Sajedi, N. Evaluation of selenium and salicylic acid effect on physiological and qualitative characteristics of dry-land wheat cultivars. Iran Agric. Res. 2017, 36, 91–100. [Google Scholar]
- Khalofah, A.; Migdadi, H.; El-Harty, E. Antioxidant enzymatic activities and growth response of quinoa (Chenopodium quinoa Willd) to exogenous selenium application. Plants 2021, 10, 719. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone priming: Regulator for heavy metal stress in plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Khan, M.Y.; Prakash, V.; Yadav, V.; Chauhan, D.K.; Prasad, S.M.; Ramawat, N.; Sharma, S. Regulation of cadmium toxicity in roots of tomato by indole acetic acid with special emphasis on reactive oxygen species production and their scavenging. Plant Physiol. Biochem. 2019, 142, 193–201. [Google Scholar] [CrossRef]
- Ali, M.A.; Asghar, H.N.; Khan, M.Y.; Saleem, M.; Naveed, M.; Niazi, N.K. Alleviation of nickel-induced stress in mungbean through application of gibberellic acid. Int. J. Agric. Biol. 2015, 17, 990–994. [Google Scholar] [CrossRef]
- Jia, H.; Song, Z.; Wu, F.; Ma, M.; Li, Y.; Han, D.; Yang, Y.; Zhang, S.; Cui, H. Low selenium increases the auxin concentration and enhances tolerance to low phosphorous stress in tobacco. Environ. Exp. Bot. 2018, 153, 127–134. [Google Scholar] [CrossRef]
- Alhaithloul, H.A.S.; Abu-Elsaoud, A.M.; Soliman, M.H. Abiotic stress tolerance in crop plants: Role of phytohormones. In Abiotic stress in plants, Fahad, S.; Saud, S.; Chen, Y.; Wu, C.; Wang, D. (Eds.), IntechOpen: 2020. [CrossRef]
- Tamaoki, M.; Freeman, J.L.; Marquès, L.; Pilon-Smits, E.A.H. New insights into the roles of ethylene and jasmonic acid in the acquisition of selenium resistance in plants. Plant Signal. Behav. 2008, 3, 865–867. [Google Scholar] [CrossRef]
- Kolbert, Z.; Ortega, L.; Erdei, L. Involvement of nitrate reductase (NR) in osmotic stress-induced NO generation of Arabidopsis thaliana L. roots. J. Plant Physiol. 2010, 167, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Abedi, S.; Iranbakhsh, A.; Oraghi-Ardebili, Z.; Ebadi, M. Nitric oxide and selenium nanoparticles confer changes in growth, metabolism, antioxidant machinery, gene expression, and flowering in chicory (Cichorium intybus L.): potential benefits and risk assessment. Environ. Sci. Pollut. Res. Int. 2021, 28, 3136–3148. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Chen, S.; Li, Y.; Zheng, F.; He, B.; Gu, M. Exogenous abscisic acid (ABA) promotes cadmium (Cd) accumulation in Sedum alfredii Hance by regulating the expression of Cd stress response genes. Environ. Sci. Pollut. Res. 2020, 27, 8719–8731. [Google Scholar] [CrossRef] [PubMed]
- Pandey, C.; Gupta, M. Selenium and auxin mitigates arsenic stress in rice (Oryza sativa L.) by combining the role of stress indicators, modulators and genotoxicity assay. J. Hazard. Mater. 2015, 287, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.W.; Zhang, C.; Wang, N.H.; Mao, W.; Wu, F. Strigolactone GR24 improves cadmium tolerance by regulating cadmium uptake, nitric oxide signaling and antioxidant metabolism in barley (Hordeum vulgare L.). Environ. Pollut. 2021, 273, 116486. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, W.; Long, C.; Wang, D. Exogenous plant growth regulator alleviate the adverse effects of U and Cd stress in sunflower (Helianthus annuus L.) and improve the efficacy of U and Cd remediation. Chemosphere 2020, 127809. [Google Scholar] [CrossRef]
- Sehar, Z.; Iqbal, N.; Fatma, M.; Rather, B.A.; Albaqami, M.; Khan, N.A. Ethylene suppresses abscisic acid, modulates antioxidant system to counteract arsenic-inhibited photosynthetic performance in the presence of selenium in mustard. Front. Plant Sci. 2022, 13, 852704. [Google Scholar] [CrossRef]
- Shi, W.G.; Liu, W.; Yu, W.; Zhang, Y.; Ding, S.; Li, H.; Mrak, T.; Kraigher, H.; Luo, Z.B. Abscisic acid enhances lead translocation from the roots to the leaves and alleviates its toxicity in Populus × canescens. J. Hazard. Mater. 2019, 362, 275–285. [Google Scholar] [CrossRef]
- Poonam, S.; Kaur, H.; Geetika, S. Effect of jasmonic acid on photosynthetic pigments and stress markers in Cajanus cajan (L.) Mill sp. seedlings under copper stress. Am. J. Plant Sci. 2013, 4, 817–823. [Google Scholar] [CrossRef]
- Singh, S.; Prasad, S.M. Effects of 28-homobrassinoloid on key physiological attributes of Solanum lycopersicum seedlings under cadmium stress: Photosynthesis and nitrogen metabolism. Plant Growth Regul. 2017, 82, 161–173. [Google Scholar] [CrossRef]
- Šípošová, K.; Labancová, E.; Kučerová, D.; Kollárová, K.; Vivodová, Z. Effects of exogenous application of indole-3-butyric acid on maize plants cultivated in the presence or absence of cadmium. Plants 2021, 10, 2503. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Asgher, M.; Per, T.S.; Masood.; Fatma, M.; Khan, M.I. Ethylene potentiates sulfur-mediated reversal of cadmium inhibited photosynthetic responses in mustard. Front. Plant Sci. 2016, 7, 1628. [Google Scholar] [CrossRef] [PubMed]
- Tai, Z.; Yin, X.; Fang, Z.; Shi, G.; Lou, L.; Cai, Q. Exogenous GR24 alleviates cadmium toxicity by reducing cadmium uptake in switchgrass (Panicum virgatum) seedlings. Int. J. Environ. Res. Public Health 2017, 14, 852. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, A.; Bhardwaj, R. Plant steroidal hormone epibrassinolide regulate – Heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ. Exp. Bot. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between plant hormones and heavy metals responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef]
- Rajewska, I.; Talarek, M.; Bajguz, A. Brassinosteroids and response of plants to heavy metals action. Front. Plant Sci. 2016, 7, 629. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Pan, K. Effect of exogenous abscisic acid on the level of antioxidants in Atractylodes macrocephala Koidz under lead stress. Environ. Sci. Pollut. Res. 2013, 20, 1441–1449. [Google Scholar] [CrossRef]
- Sharma, A.; Sidhu, G.P.S.; Araniti, F.; Bali, A.S.; Shahzad, B.; Tripathi, D.K.; Landi, M. The role of salicylic acid in plants exposed to heavy metals. Molecules 2020, 25, 540. [Google Scholar] [CrossRef]
- Ahsan, N.; Renaut, J.; Komatsu, S. Recent developments in the application of proteomics to the analysis of plant responses to heavy metals. Proteomics 2009, 9, 2602–2621. [Google Scholar] [CrossRef]
- Hossain, Z.; Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front. Plant Sci. 2013, 3, 310. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Wu, X.; Qiu, B.; Wu, F.; Jiang, L.; Zhang, G. Physiological and proteomic alterations in rice (Oryza sativa L.) seedlings under hexavalent chromium stress. Planta 2014, 240, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Thapa, G.; Sadhukhan, A.; Panda, S.K. Molecular mechanistic model of plant heavy metal tolerance. Biometals 2012, 25, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Behbahani, S.R.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Red elemental selenium nanoparticles mediated substantial variations in growth, tissue differentiation, metabolism, gene transcription, epigenetic cytosine DNA methylation, and callogenesis in bittermelon (Momordica charantia); an in vitro experiment. PLoS ONE 2020, 15, e0235556. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Thukral, A.K.; Arora, S.; Bhardwaj, R. Role of Se(VI) in counteracting oxidative damage in Brassica juncea L. under Cr(VI) stress. Acta Physiol. Plant. 2017, 39, 39. [Google Scholar] [CrossRef]
- Schiavon, M.; Berto, C.; Malagoli, M.; Trentin, A.; Sambo, P.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Selenium biofortification in radish enhances nutritional quality via accumulation of methyl-selenocysteine and promotion of transcripts and metabolites related to glucosinolates, phenolics, and amino acids. Front. Plant Sci. 2016, 7, 1371. [Google Scholar] [CrossRef]
- Dutilleul, C.; Jourdain, A.; Bourguignon, J.; Hugouvieux, V. The Arabidopsis putative selenium-binding protein family: Expression study and characterization of SBP1 as a potential new player in cadmium detoxification processes. Plant Physiol. 2008, 147, 239–251. [Google Scholar] [CrossRef]
- Feng, X.; Ma, Q. Transcriptome and proteome profiling revealed molecular mechanism of selenium responses in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2021, 21, 1–16. [Google Scholar] [CrossRef]
- Kumar, I.; Sharma, R.K. Production of secondary metabolites in plants under abiotic stress: an overview. Significances Bioeng. Biosci. 2018, 2, 196–200. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Akhi, M.Z.; Haque, M.M.; Biswas, M.S. Role of secondary metabolites to attenuate stress damages in plants. In Antioxidants-Benefits, Sources, Mechanisms of Action. IntechOpen: 2021.
- Ahanger, M.A.; Bhat, J.A.; Siddiqui, M.H.; Rinklebe, J.; Ahmad, P. Silicon and secondary metabolites integration in plants: A significant association in stress tolerance. J. Exp. Bot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Delfine, S.; Csiky, O.; Seufert, G; Loreto, F. Fumigation with exogenous monoterpenes of a non-isoprenoid-emitting oak (Quercus suber): monoterpene acquisition, translocation, and effect on the photosynthetic properties at high temperatures. New Phytol. 2000, 146, 27–36. [Google Scholar] [CrossRef]
- Hassan, A. Effects of mineral nutrients on physiological and biochemical processes related to secondary metabolites production in medicinal herbs. Med. Arom. Plant Sci. Biotechnol. 2012, 6, 105–110. [Google Scholar]
- Nakabayashi, R.; Mori, T.; Saito, K. Alternation of flavonoid accumulation under drought stress in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e29518. [Google Scholar] [CrossRef]
- Kováčik, J.; Grúz, J.; Bačkor, M.; Strnad, M.; Repčák, M. Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep. 2009, 28, 135–143. [Google Scholar] [CrossRef]
- Manquián-Cerda, K.M.; Escudey, G.; Zúñiga, N.; Arancibia-Miranda, M.; Molina, E. C. "Effect of cadmium on phenolic compounds, antioxidant enzyme activity and oxidative stress in blueberry (Vaccinium corymbosum L.) Plantlets grown in vitro.". Ecotoxicol. Environ. Saf. 2016, 133, 316–326. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef]
- Tan, P.; Zeng, C.; Wan, C.; Liu, Z.; Dong, X.; Peng, J.; Lin, H.; Li, M.; Liu, Z.; Yan, M. Metabolic profiles of Brassica juncea roots in response to cadmium stress. Metabolites 2021, 11, 383. [Google Scholar] [CrossRef]
- Anjitha, K.S.; Sameena, P.P.; Puthur, J.T. Functional aspects of plant secondary metabolites in metal stress tolerance and their importance in pharmacology. Plant Stress 2021, 2, 100038. [Google Scholar] [CrossRef]
- Kiumarzi, F.; Morshedloo, M.R.; Zahedi, S.M.; Mumivand, H.; Behtash, F.; Hano, C.; Chen, J.T.; Lorenzo, J.M. Selenium nanoparticles (Se-NPs) alleviates salinity damages and improves phytochemical characteristics of pineapple mint (Mentha suaveolens Ehrh.). Plants 2022, 11, 1384. [Google Scholar] [CrossRef] [PubMed]
- Gul, H.; Kinza, S.; Shinwari, Z.K.; Hamayun, M. Effect of selenium on the biochemistry of Zea mays under salt stress. Pak. J. Bot. 2017, 49, 25–32. [Google Scholar]
- Chauhan, R.; Awasthi, S.; Tripathi, P.; Mishra, S.; Dwivedi, S.; Niranjan, A.; Tripathi, R.D. Selenite modulates the level of phenolics and nutrient element to alleviate the toxicity of arsenite in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2017, 138, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. Effect of selenium application on phenylalanine ammonia-lyase (PAL) activity, phenol leakage and total phenolic content in garlic (Allium sativum L.) under NaCl stress. Inf. Process. Agric. 2018, 5, 339–344. [Google Scholar] [CrossRef]
- Skrypnik, L.; Styran, T.; Savina, T.; Golubkina, N. Effect of selenium application and growth stage at harvest on hydrophilic and lipophilic antioxidants in lamb’s lettuce (Valerianella locusta L. Laterr.). Plants 2021, 10, 2733. [Google Scholar] [CrossRef] [PubMed]
- Chomchan, R.; Siripongvutikorn, S.; Puttarak, P.; Rattanapon, R. Influence of selenium bio-fortification on nutritional compositions, bioactive compounds content and anti-oxidative properties of young ricegrass (Oryza sativa L.). Funct. Foods Health Dis. 2017, 7, 195–209. [Google Scholar] [CrossRef]
- Farman, M.; Nawaz, F.; Majeed, S.; Ahmad, K.S.; Rafeeq, R.; Shehzad, M.A; Shabbir, R.N.; Usmani, M.M. Interplay between selenium and mineral elements to improve plant growth and development. In Handbook of Bioremediation, Academic Press, 2021, 221-236.
- Hawrylak-Nowak, B. Effect of selenium on selected macronutrients in maize plants. J. Elem. 2008, 13, 513–519. [Google Scholar]
- Drahoňovský, J.; Száková, J.; Mestek, O.; Tremlová, J.; Kaňa, A.; Najmanová, J.; Tlustoš, P. Selenium uptake, transformation and inter-element interactions by selected wildlife plant species after foliar selenate application. Environ. Exp. Bot. 2016, 125, 12–19. [Google Scholar] [CrossRef]
- Khattab, H. Metabolic and oxidative responses associated with exposure of Eruca sativa (Rocket) plants to different levels of selenium. Int. J. Agric. Biol. 2004, 6, 1101–1106. [Google Scholar]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, C.; Wang, X.; Qing, X.; Wang, P.; Zhang, Y.; Zhang, X.; Zhao, X. Selenium alleviated chromium stress in chinese cabbage (Brassica campestris L. ssp. Pekinensis) by regulating root morphology and metal element uptake. Ecotoxicol. Environ. Saf. 2019, 173, 314–321. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of selenium in the mitigation of heavy metals stress.
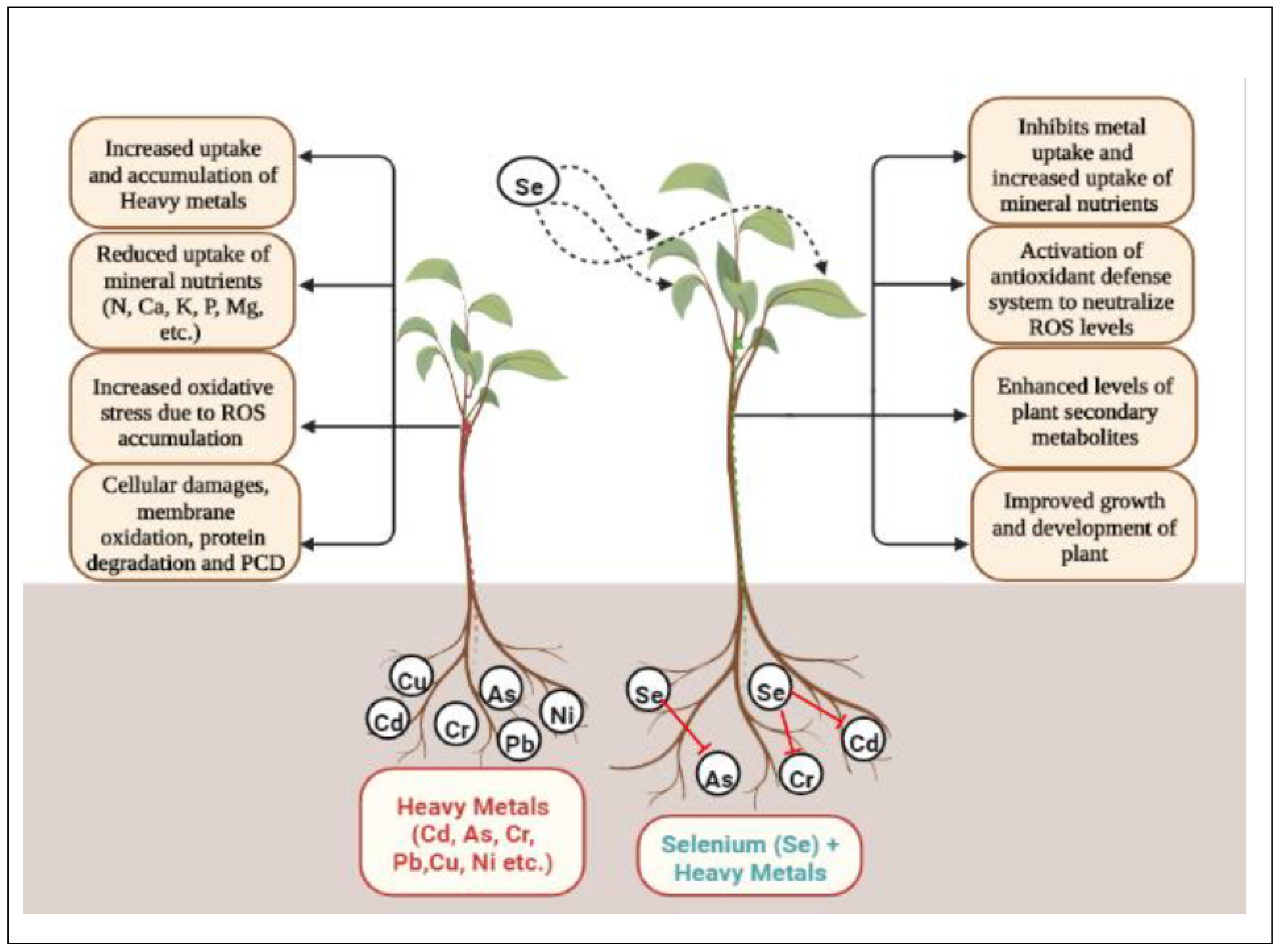

Table 1.
Role of Selenium under HM stress conditions.
| Species | Heavy metal conc. | Se conc. | Effect of Se on antioxidants metabolism | Reference(s) |
|---|---|---|---|---|
| Glycine max | As |
25 μM |
Reduced As toxicity by improving photosynthesis, antioxidants, and regulation of some defense genes | [23] |
| Triticum aestivum | Cd | 5 and 10 μM | Down-regulation of genes of Cd uptake and transport | [84] |
| Oryza sativa | As |
25 μM |
Up-regulated various As tolerant genes and also induced the antioxidant expression | [85] |
| Cucumus sativus | 25 µM Cd 200 µM Ni 100 µM Pb |
8 mg/L |
Stimulated antioxidant system by enhancing the activity of CAT, APX and GPOX | [79] |
| Vicia faba | 50 µM Pb |
6 µM |
Up regulated CAT, GPOX and GSH-Px levels | [82] |
| Phaseolus aureus | 10 µM As | 5 µM | Reduced As induced oxidative stress by increasing CAT, APX GR, AsA and GSH levels | [81] |
| Brassica spp. | 50 µmol/L Cd |
3 µmol/L | Induced ROS detoxification, maintained the levels of SOD, CAT and POD | [86] |
| Satureja hortensis | 150 µM Cd |
40 µM | Elevated the levels of CAT and POD and reduced Cd toxicity. | [87] |
| Brassica juncea | 300 µM Kg-1 Cr | 4 µM Kg-1 | Strengthen the inheretent defense system by inducing the expression of SOD, CAT, APX, GPOX, GR, GST, DHAR M-DHAR, AsA and GSH against Cr stress. | [27] |
| Brassica juncea | 100 or 200 mg/L Cd | 50 µM | Increment in CAT, APX and GR activity was observed | [88] |
| Lolium perenne | 0.2 mM Al |
5 µM | Reduced lipid peroxidation by Increasing SOD and APX activity | [80] |
| Oryza sativa | 20 µmol/L Cd |
1 µmol/L |
Up regulated CAT and GSH-Px activity and reduced lipid oxidation | [89] |
| Oryza sativa | 25 µM As |
25 µM | Positively enhanced the activities of CAT, APX, GSH-Px, GR, GST and GSH | [90] |
| Brassica napus | 5 mg/kg Cd 500 mg/kg Pb |
15 mg/kg |
Enhanced SOD and GSH-Px levels to minimize Cd/Pb induced oxidative stress | [83] |
Table 2.
Role of phytohormones under heavy metals stress.
| Species | HM conc. | Phytohormones Conc. |
Response | Reference(s) |
|---|---|---|---|---|
| Brassica juncea | 24 mg kg−1 As |
200 μl/l Ethephon |
Improvement of photosynthetic attributes by reduced As and ABA accumulation, increment in antioxidant activity | [119] |
| Cicer arietinum | Cd | ABA, JA | Improvement in accumulation of antioxidants and osmolytes,ROS detoxification, decreased Cd accumulation | [44] |
| Vigna radiata | Cd | GA, SA | Improved growth and metabolism, stimulation of antioxidants; reduced MDA content | [43] |
| Sedum alfredii | 100 μmol/L Cd | 0.2 mg/L ABA |
Exogenous ABA enhanced endogenous ABA production which is involved in Cd tolerance by regulating the expression of Cd tolerance genes | [115] |
|
Populus × Canescens |
3mM Pb |
10μM ABA |
Ameliorated the toxic effects of Pb by minimizing oxidative stress and induced expression of genes responsible for Pb resistance | [120] |
| Oryza sativa | 150 μM As |
3μM IAA |
Improved growth by accumulating more amino acids, proteins etc. | [116] |
| Cajanus cajan | 5mM Cu2+ |
1nM JA |
Improved photosynthesis, antioxidative system and reduced oxidative stress | [121] |
| Solanum lycopersicum | 3mg/kg Cd |
10nM HBL |
Improved overall growth and productivity of plants, positively regulated N-metabolism. | [122] |
| Zea mays | 50mM Cd |
10−9M IBA |
Reduced Cd toxicity by inducing ROS detoxification and improved nutrient status of plants | [123] |
| Vigna radiata | 20-60 mg/kg Ni |
10−4M GA3 |
Improved growth and biomass of plants by reducing the uptake of Ni from soil | [109] |
| Helianthus annuus | 3.25/0.27 mg kg1 U/Cd |
500 mg/l IAA |
ROS detoxification by inducing antioxidants, increased uptake of U/Cd from soil | [118] |
| Brassica juncea | 50 μM Cd |
200 μl/l Ethephon |
Improved growth, induced antioxidants and amino acids accumulation | [124] |
| Brassica juncea | 1.2mM Cr |
200 μl/l Ethephon |
Mitigated Cr stress by improving photosynthesis, reduced oxidative stress and also enhanced proline accumulation | [11] |
| Hordeum vulgare | 10 μM Cd |
1 μM GR24 (strigol analogue) |
Reduced Cd toxicity by improving photosynthesis, uptake of essential nutrients, inducing stress markers | [117] |
| Panicum virgatum | 10 μmol/l Cd |
1 μmol/l GR24 (strigol analogue) |
Increment in photosynthetic parameters, stimulation of antioxidant system and improvement in mineral status of plants | [125] |
| Solanum lycopersicum | 150 μM Cd | 100 μM NO |
Positive effect on photosynthesis and improved growth, antioxidant system, osmoprotectants and secondary metabolites accumulation, hence involved in Cd tolerance. | [126] |
| Oryza sativa | 0.5mM Cr |
0.1nM 24-EBL |
Induced Cd detoxification by enhancing photosynthesis and regulating the genes expression | [127] |
Table 3.
Effect of Se on the production of secondary metabolites.
| Plant species | Se dose | Secondary metabolites | Response | Reference(s) |
|---|---|---|---|---|
| Mentha suaveolens | 10 mg L−1 | Production of essential oils such as piperitenone oxide, limonene, jasmone etc. | Enhanced growth and secondary metabolites production | [154] |
| Brassica juncea | 4 µM Kg−1 | Enhanced levels of phenols, flavonoids and anthocyanins | Improvement in the photosynthesis thereby growth of plant by inducing the accumulation of antioxidants, SMs, etc. against Cr stress | [27] |
| Brassica oleracea | 25 µM | Affected the production or accumulation of glucoraphanin an important glucosinolate | Reduced content of glucosinlates precursors, Suppressed the expression of genes involved in glucosinlates biosynthesis | [28] |
| Melissa officinalis | 5 µM | Increased the accretion of essential oils such as z-citral, citral and geranyl acetate | Positive effect on the growth at low concentration | [16] |
| Brassica oleracea | 10mg/L | Increment in the levels of phenolic compounds and glucosinolates | Improved growth and yield characteristics of plants by inducing antioxidant and SMs levels | [25] |
| Zea mays | 10mM | Enhancement of phenols and flavonoids content | Induced accumulation of proteins, sugars and SMs | [155] |
| Oryza sativa | 25 µM | Induced the accumulation of important phenolics such as gallic, protocatechuic and ferulic (acids) | Increased the uptake of nutrients from soil and regulated SMs production, hence reduced As toxicity | [156] |
| Brassica juncea | 50 µM | Enhanced phenolic content | Reduced Cd stress by inducing the antioxidant system and SMs status of plants | [88] |
| Allium sativum | 4/8 mg Se L−1 | Increased total phenolic content | Increased tolerance ability of garlic to salt stress by preventing membrane oxidation, phenols accumulation and regulation of phenylalanine ammonia lyase activity | [157] |
| Vallerianella locusta | 5 µM | Enhanced the endogenous levels of flavonoids and phenolics | Improved growth, antioxidant activity and accumulation of SMs | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



