
Submitted:
21 April 2023
Posted:
23 April 2023
You are already at the latest version
Abstract
Plant stem cell cultures have so far been established in only a few plant species using cambial meristematic cells. The presence of stem cells or stem cell-like cells in other organs and tissues of the plant body, as well as the possibility of de novo generation of meristematic cells from differentiated cells, allows to consider the establishment of stem cell cultures in a broader range of species. Therefore, this study aimed to establish a stem cell culture of the medicinal plant Calendula officinalis L. Callus tissues were induced from explants taken from leaves and roots. Appropriate combinations of plant growth regulators and light parameters for in vitro cultivation were selected. Already at this stage, stem and dedifferentiated cells could be identified, and cell suspension cultures were established from specific parts of the formed callus. Cell suspension cultures with a high proportion of stem cells originating from roots (92–93%) or leaves (72–73%) were developed. The amount of stem cells derived from the roots in the cell suspension reached a plateau at a level of 5.60–5.72105.
Keywords:
marigold
; callus
; cell suspension
; growth parameters
; cell viability
; stem cell
1. Introduction
Calendula officinalis L. (calendula), commonly known as the pot marigold or marigold, is cultivated as an ornamental annual or naturally grown as a perennial flowering plant of the family Asteraceae, genus Calendula. It has yellow or orange-colored edible florets that are also used as a dye in the production of foods and cosmetics. However, more interesting are the various pharmacological properties and therapeutic potential attributed to flowers and leaves [1]. Calendula possesses many phytochemicals and pharmacological activities to be considered an excellent source of new drugs [2]. Many of these bioactive substances find application as antioxidants in cosmetics [3]. Extracts from calendula protect cells against UV radiation, stimulate skin cell regeneration, and improve skin elasticity [4]. In addition, anti-inflammatory, antimicrobial, immunostimulatory, as well as spasmolytic, hypolipidemic, antidiabetic, cardioprotective, hepatoregenerative, pancreas-regenerating, neuroprotective, anticancer, and other health-promoting effects, determine its applications in dermatology and medicine [5]. The Cosmetic Ingredient Review Expert Panel concluded that all ingredients from calendula are safe for applications in cosmetics under current practices [6]. Because of its interesting phytochemical composition, the traditional field growing of calendula plants is being replaced by the in vitro production of calendula cells and extracts from those cells as active cosmetic ingredients [7].
The in vitro cultivation systems in calendula can produce plantlets by micropropagation [8], but callus, cell suspension, and hairy root cultures can provide a continuous supply of calendula bioactive metabolites [7,9,10]. The induction of callus and its growth are fundamental steps for the establishment and applications of other plant tissue cultures. Callus tissues themselves can be used to prepare extracts with properties like antibiotics [11], establish cell suspensions [12], and protoplast cultures [13]. Hairy root cultures of calendula, induced by genetic transformation with the Agrobacterium rhizogenes, can also effectively produce secondary metabolites [10,14,15,16].
Different factors are responsible for the induction of callus in calendula, including genotype, explant type, culture medium, exogenous growth hormones, and others [17,18,19]. These factors affect not only the callus induction itself but also the growth parameters, morphogenetic capacity, and production potential of the callus. Cultivated calli and cell suspension cultures derived from calli are currently the most used platforms for in vitro biosynthesis of complex molecules or natural products, especially those with high economic value [20]. These in vitro production systems exploit mainly dedifferentiated cells developed from the differentiated cells of initial explants or, most often, a heterogeneous mixture of different types of cells. This approach has several limitations due to cellular heterogeneity and genetic and epigenetic instability. This is manifested by the slow growth rate of dedifferentiated plant cells, the lower yield of secondary metabolites, the aggregation of cells in the bioreactor, and mainly by undesirable variation in these parameters [21]. To overcome these problems and bypass the dedifferentiation step, a new platform in plant cell cultures has been described. It tries to use the potential of innately undifferentiated cambial meristematic cells (CMC) [22]. CMCs may provide a robust, cost-effective, environmentally friendly platform and a sustainable source of plant cells and plant-derived natural products [23]. CMCs have plant stem cell properties [24]. They can actively divide, leading to the formation of different cells that eventually go through a differentiation process and at the same time produce new stem cells [25]. Populations of stem cells can be considered immortal. They are theoretically able to divide an unlimited number of times [26] without being adversely limited by plant source, location, harvest period, or prevailing environmental conditions. The growth of stem cells (SCs) or stem cell-like cells (SCLCs) in culture in vitro is far superior to that of dedifferentiated cells in solid format (callus culture) as well as in liquid format (suspension culture) [26]. Moreover, SCs and SCLCs in suspension culture can tolerate shear stress, and the presence of small and abundant vacuoles avoid or reduce cell aggregation in bioreactors [27].
To date, several studies on CMCs have been reported, but only in a limited number of plant species, mainly medicinal ones e.g., Taxus cuspidata, Ginkgo biloba, Solanum lycopersicon [22], Panax ginseng [22,28], Catharanthus roseus [29], Tripterygium wilfordii [30], Camptotheca acuminate [31], Ocimum basilicum [32], Fraxinus mandshurica [33]. However, no study has been presented on the establishment and cultivation of stem cell or stem-like cell suspension cultures derived from Calendula officinalis L. Establishment of stem cell cultures in small volumes or in bioreactors should ensure higher production of cells and secondary metabolites in comparison with the cultivation of dedifferentiated cells in suspension cultures. Therefore, the aims of this work were to: i) induce and characterize callus cultures; ii) establish cell suspension culture and determine its basic parameters; and iii) establish and characterize a line of stem cells or stem cell-like cells from Calendula officinalis L.
2. Results
2.1. Callus Formation
The frequency of callus formation from leaf explants during the first 4 weeks was 100% using all combinations of auxin and cytokinin in the callus induction medium. The root explants also developed callus tissue at 100% frequency, but only in media containing 2,4-D as auxin. Lower frequencies (67–92%, rounded) were in media containing IAA as auxin (Table S1). Callus formation on leaf explants had already been initiated earlier, 12 days after they were placed on the induction medium. Root explants formed calli after 15 days. Callus formation after 4 weeks occurred along the entire length of root segments, while in the leaf segments, calli were visible only on the cutting wounds, but later calli overgrew the whole leaf explant. Significant differences (p < 0.05) in the percentage of callus formation were observed between the leaf- and root-derived explants.
The color of the calli that developed after 4 weeks varied according to the combinations of auxin and cytokinin used. The calli were multicolored. However, lighter colors prevailed, mainly yellow. During subsequent cultivation, the color gradually changed. After 16 weeks of cultivation (i.e., after three passages), most of them partially or completely darkened to brown and black, and necrotization was also observed. The structure of a callus usually relates to its quality, regarding the processes of morphogenesis and plant regeneration. After 4 weeks of cultivation, all calli induced both from leaf and root explants, regardless of combinations of growth regulators, were compact. The compact callus had a firm texture that could not be easily cut. But, already after the first and subsequent passages, its structure changed to watery soft and watery soft friable, especially if the 2iP was used as a cytokinin (Table S1).
Some combinations of growth regulators in the medium also induced the development of roots, already after 4 weeks of cultivation. This was observed in the combinations IAA + 2iP and IAA + KIN in leaf explants and IAA + 2iP and IAA + TDZ in root explants.
2.2. Callus Growth
The highest average fresh weight of callus biomass derived from leaves (˃ 6 g), after 4 weeks of cultivation, was on the medium with the combination IAA + 2iP (Figure 1). Unfortunately, these calluses developed roots, which undesirably increased their weight. After 8, 12, and 16 weeks, this combination of growth regulators no longer produced the highest callus biomass weight. For this reason, this combination of growth regulators was excluded from the following experiments aimed at analysing of the stem cells presence. The fresh weight of callus biomass derived from leaves using all other combinations of growth regulators was in the range 1.24–3.11 g. The range of average fresh weight of callus biomass derived from root explants was similar, from 0.80 g to 3.58 g (Figure 1).
An important growth parameter of callus biomass was an increase in the fresh weight determined after three subsequent passages (i.e., after 16 weeks) on the fresh medium. Continuous increase in the fresh weight of callus biomass was recorded up to 16 weeks of cultivation in media with all combinations of growth regulators and with both types of explants (Figure 1).
The highest increase in fresh biomass weight after 16 weeks of cultivation was in calli derived from leaf tissue, in medium containing combinations IAA + BAP (from 2.44 g to 22.87 g) and 2,4-D + BAP (from 2.23 g to 16.91 g). In the case of callus biomass derived from roots, the same combination of IAA + BAP (from 1.42 g to 16.20 g) was most effective (Figure 2).
2.3. Stem Cells in Callus
The cells with the properties of stem cells or stem cell-like cells should be present in fast growing multicellular callus biomass along with dedifferentiated and differentiated cells. The presence of SCs or cells similar to them in the growing cell biomass should be supported by the morphological features of cells developed on the callus surface. After four weeks of cultivation, different types of callus cell biomass, containing SCs, cells similar to them (stem cell-like cells), as well as DDCs (dedifferentiated cells), could be visually observed. Calli with potentially proliferating SCs were light yellow, dim, and soft, while DDCs were light white, compact, and with an irregular structure (Figure 3). However, these parameters can be ambiguous as they may vary depending on the plant species, the explant, and the callus induction conditions.
Microscopic analysis revealed typical differences in the morphology of vacuoles between SCs and DDCs stained with the Neutral Red assay. Abundant and small spheric vacuoles, or vacuole-like structures, were observed in SCs, while only one large vacuole was present in DDCs (Figure 4). These differences in the parameters of the vacuoles are considered characteristic.
The relative ratio between SCs and other cells in the growing cell biomass was determined subjectively using microscopic observations. The differences in ratio between SCs and DDCs were related to the explant source. A higher proportion of SCs was subjectively evaluated in callus biomass derived from leaves (Figure 5a, left). The callus derived from root explants contained a higher proportion of DDCs and other cell types (Figure 5b, right).
Two combinations of growth regulators that most effectively promoted the growth of leaf-derived callus cell biomass were subjectively compared in amount of SCs. Apparently higher relative amounts of SCs over DDCs were found in medium containing 1.0 mg/L 2.4-D with 0.5 mg/L BAP (Figure 6, left). On the contrary, the combination of 1.0 mg/L IAA with 0.5 mg/L BAP generated predominantly DDCs (Figure 6, right).
2.4. Cell Suspension Cultures
Cell suspension cultures were derived from callus tissues induced either from leaf or root explants in the same composition of nutrient medium, either in continuous darkness or under photoperiod (16 h light/8 h darkness). The growth characteristics of cells in suspensions were different (Figure 7).
The highest fresh cell weight (FCW, g/L) was achieved between the 11th and 13th day after the inoculation of cells into the fresh liquid medium. The optimal interval for passaging of cells into fresh liquid medium was 12–13 days in cell suspensions cultivated in darkness and 11 days in cell suspensions cultivated under the photoperiod. Cultivation in continuous darkness produced 3.8 times more root-derived cells and 3.2 times more leaf-derived cells in comparison to cultivation under the photoperiod. Therefore, only the darkness regime of cultivation was used in the following experiments. Statistically significant (p < 0.05) the highest production of fresh cells (64.3 g of cells/L of cultivation medium) was achieved in cell suspension derived from leaves. It was 1.42 times higher than in root-derived cell suspension.
Determination of cell viability revealed that cell suspensions developed from root-derived callus (CR) were more viable than those from leaf-derived callus (CL). The maximum number of viable root-derived cells (5.08×105 cells/mL) was on the fifth day after initiation of cell suspension culture. In leaf-derived cells, the maximum number of viable cells was lower by 56% (2.24×105 cells/mL) and reached later, on the 11th day of cultivation. It has been shown that both stains, Evans blue and Trypan blue, are essentially equally appropriate for cell viability detection (Figure 8).
2.5. Stem Cell Suspension Cultures
The cell suspension cultures were used for selection and establishment of a culture of stem cells i.e., a culture with the highest possible proportion of SCs versus DDCs. The Neutral Red assay was much easier in cell suspension compared to callus tissue. SCs had typical very small and abundant vacuoles. Prolonged cultivation time and repeated passages of cells into fresh liquid medium eliminated or fundamentally reduced aggregation of cells, and the single-cell character of the cell suspension began to dominate. At the same time, the fewer aggregates and more individual cells were in the cell suspension, the higher the proportion of stem cells (Figure 9).
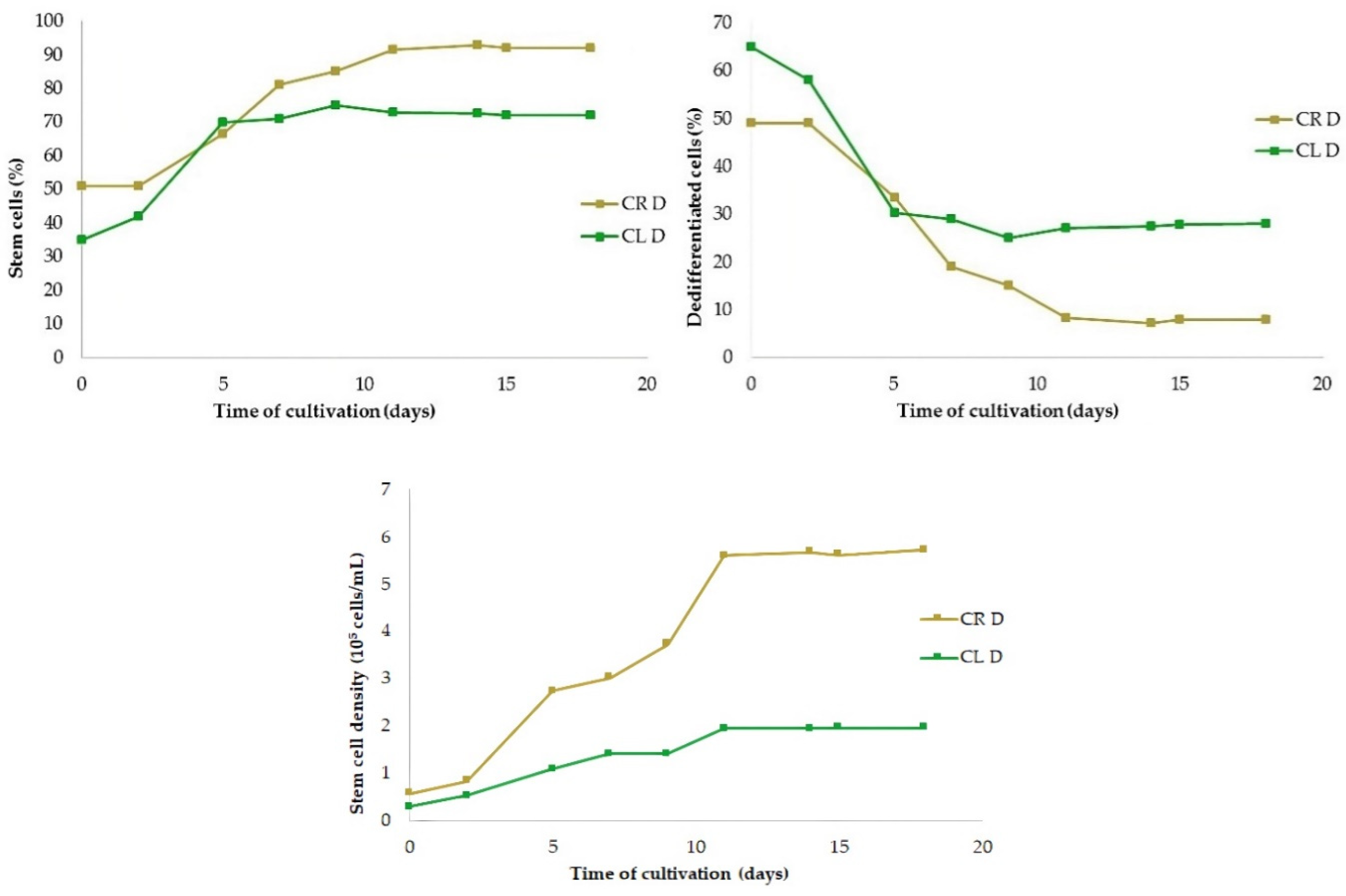
The ratio of SCs to DDCs in suspension culture had opposite tendencies. As the ratio of SCs increased (Figure 10, top left), the ratio of DDCs decreased adequately (Figure 10, top right). The ratio of SCs derived from roots exceeded the level of 91.6% on the 11th day and was maintained in the range 92–93%, in contrast to root-derived DDCs that gradually decreased to less than 8%. A similar course had a proportion of leaf-derived SCs that reached a maximum also on the 11th day, but at a level of 72.9%, which stabilized subsequently at a level of 72–73%. The share of leaf-derived DDCs decreased adequately to 27–28%. The number and percentage of SCs did not decrease at all, even on the 18th day of cultivation.
From the point of view of the amount of SCs produced, root cells were more productive than leaf cells (Figure 10, bottom). Their multiplication reached a plateau on the 11th day of cultivation, and the number of SCs then stabilized at a level 5.60–5.72×105 cells/mL. The growth of leaf-derived stem cells reached a plateau also on the 11th day of cultivation at a level 2.2×105 cells/mL i.e., about 2.5 times less.
3. Discussion
The plant species, type of explant, and composition of the cultivation medium, especially the content of growth regulators, are well-known crucial factors affecting the initiation of callus development and growth in vitro. All combinations of auxins and cytokinins used in our experiments were able to induce callogenesis in both types of C. officinalis L. explants, the leaf as well as root segments. Inducing the formation and growth of callus tissue is generally not a problem in plant tissue cultures. This was also the case in this study. The most effective combinations of growth hormones for the induction and growth of callus biomass can be proven to be the same, also between laboratories. Sometimes other combinations of growth hormones prove to be the most effective, even for the same plant species and the same type of explant. This is mainly related to the specific genotypes used. In the presented work, the most effective combination of auxin and cytokinin was IAA + BAP. The combinations 2.4-D + KIN and 2.4-D + 2iP used previously [12] induced calli with much lower efficiency. Other growth hormones have proven effective for other explants such as hypocotyls, cotyledons, cotyledon nodes [11], and floret explants [34]. Calli initiated from different types of explants of C. officinalis L. tend to be variable in their color and texture, which also changes during cultivation depending on the growth regulators used for callus initiation. The coloration of the calli in this study ranged from white to yellow, orange, brown, and black, and the calli were often multicolored. Similar coloration and texture were also found in calli derived from hypocotyls [11], but the calli derived from the leaves and cotyledons were in shades of green [12]. The color of calli induced from leaves and florets also depended on sucrose concentrations in medium and was light green at lower concentrations and cream-yellow at higher concentrations [34]. In general, callus cultures of calendula can be easily induced from different types of explants, using culture media containing a mixture of auxin and cytokinin. Calli also differ in their texture, from firm and compact to friable and watery. Moreover, sometimes they are very heterogeneous in these characteristics.
Callogenesis can also run simultaneously with organogenesis, and then this type of tissue culture can be suitable for the micropropagation of calendula plants [8,35]. Moreover, the long-term cultivation and multiplication of calendula callus in vitro, either unelicited or elicited, is important in the production of useful compounds such as carotenoid pigments [34], salicylic acid [36], and extracts with antibacterial activity [11]. The callus induction stage is already important for the subsequent establishment of cell suspension culture. From the point of view of callus disintegration, needed for establishing a fine cell suspension culture in a liquid medium, its friable structure is more advantageous. However, the character of cells in callus is especially important in establishing a fast-growing, long-term cultivable, and more productive cell suspension culture. The callus is initiated either from explants containing meristems with pluripotent stem cells or from explants containing essentially only somatic cells. Callus is formed predominantly from a pre-existing population of stem cells [37] and from dedifferentiated cells arising through transformation from differentiated cells. Dedifferentiated cells are in a state of less differentiation or in a stem-like state (stem cell-like cells), which gives them the properties of totipotency [38,39]. Depending on the stem cells already present in the explant used for callus induction as well as the growth regulators in the callus induction medium, the callus biomass has typically improved growth parameters [22,33]. The presumed presence of calendula stem cells was experimentally confirmed by microscopic analyzes in calluses that had the fastest and largest increase in callus biomass.
The origin and character of the cells already present in the primary explant, interacting with the growth regulators in the callus-induction medium, are inevitably reflected in the parameters of cell suspension culture derived from them. The amount of stem cells present in the callus should be reflected in many parameters of the cell suspension culture, such as a rapid cell growth rate, an exponential increase in fresh cell weight, a lower aggregation rate, cell size and shape, and others [22,30,32,33]. Homogenization of the cell suspension culture into a single-cell culture was done without removing cell aggregates by sieving or gravity. This was achieved by several passages of the cells into fresh cultivation medium at the late exponential phase, which was around the 13th day after initiation. An increased homogeneity of the suspension culture toward the single-cell culture correlated with the increasing proportion of SCs versus DDCs in the culture. The cultures of stem cell lines with a very high proportion of stem cells were established in this way. Until now, stem cell cultures have been established exclusively from isolated and cultured innately undifferentiated cambial meristematic cells from stem segments [22,29,30,31,32,33]. However, the use of cambial meristematic cells from stem explants may not be the only way to establish an in vitro culture of stem cells. A callus resembling the root tip meristem can be developed from pericycle cells of mature roots that retain some meristematic cell-specific features, and this is also possible from cells surrounding vasculature bundles in other aerial organs [40]. Meristematic features of some pericycle cells, such as three or more vacuoles and dense cytoplasm [41] are considered typical features of plant stem cells. Also, leaves maintain their own meristems. Leaf mesophyll cells as well as the initial cells of stomata and veins are produced by leaf meristems classified as intercalary meristems [42]. The leaf meristems contain the plate meristem, which extends the leaf two-dimensionally by active cell proliferation [43] as well as the marginal meristem restricted to the leaf margin [44]. The plate and marginal meristems, both active in the basal area of leaf primordia, represent two zones of a leaf meristem, analogous to the central and peripheral zones of the shoot apical meristem [45]. Leaves exhibit determinate growth, indicating that the potential leaf meristem, if it exists, has only transient meristematic activity, but the leaf marginal and submarginal regions maintain active cell divisions during early leaf development and are responsible for leaf lamina initiation [46]. Other studies suggested that marginal meristem activity contributes little to the growth of the leaf blade, but the blade growth in angiosperm is sustained by cell proliferation in a plate meristem region [43].
Another possibility is related to de novo organogenesis and plant regeneration in plant tissue cultures. Plant somatic cells, after reprogramming, can form a mass of pluripotent cells in auxin-induced callus tissue [47]. These pluripotent and regeneration-competent cells in the root are termed vasculature-associated pluripotent cells, and the callus originates from xylem-pole pericycle and pericycle-like cells [48]. The regeneration-competent cells were found in leaf explants, throughout the mature leaf along the vasculature in dicot plants, and in the leaf base in monocot plants [49]. Just cells acquiring pluripotency are crucial for the formation of the callus and the promeristem, and finally for de novo shoot regeneration by indirect organogenesis [47,50]. Shoots and roots can be regenerated through de novo formed meristems [51]. Meristems can also be formed de novo by gene editing after the concomitant expression of developmental regulators and gene-editing reagents [52]. Thus, the use of explants derived from roots and leaves provides a prerequisite for the possibility of establishing stem and stem cell cultures.
The latest findings change opinions not only about the nature and composition of callus tissues but also about meristems, stem cells, and the processes of their de novo formation. Views on the capabilities of differentiated cells and the processes of their dedifferentiation are also changing. This opens the way not only to the isolation of plant stem cells but also to their propagation, the establishment of in vitro stem cell cultures, the production of stem cell biomass, and the more efficient production of secondary metabolites. This opens interesting perspectives for plant biotechnologies and their use in obtaining additional theoretical knowledge as well as practical applications.
4. Materials and Methods
4.1. Plant Material
Seeds of Calendula officinalis L. were obtained from the breeding company Zelseed spol. s r. o. (Horná Potôň, Slovakia). Surface sterilization of mature seeds was performed with 96% ethanol (v/v) for 30 seconds, followed by soaking in a 4.7% (v/v) solution of sodium hypochlorite for 5 minutes with constant stirring and rinsing with sterile distilled water five times. Seed were germinated in vitro in culture vessels containing Murashige and Skoog (MS) medium [53] with a reduced concentration of all components (½MS) and 0.8% agar (w/v), adjusted to pH 5.7. Germinated seeds and seedlings were cultivated at 23 ± 2 °C under a photoperiod of 16 h of light and 8 h of darkness.
4.2. Callus Cultures
Explants were taken from the leaves and roots of 4–5 weeks-old aseptic plants. The leaf explants were approximately 0.5x0.5 cm in size, and the length of the root explants was approximately 0.5 cm. The culture medium used was Murashige and Skoog (MS) medium (1962) which contained 30 g/L (w/v) sucrose, 8 g/L (w/v) of plain agar powder, and complete nutrients in the form of macro, micro, trace elements, and vitamins, supplemented with plant growth hormones and regulators, respectively. Combinations of two auxins: 2,4-D (2,4-dichlorophenoxyacetic acid) or indole-3-acetic acid (IAA) with four cytokinins—6-(γ,γ-dimethylallylamino)purine (2iP), kinetin (KIN), 6-benzylaminopurine (BAP), or thidiazuron (TDZ) in a ratio 2:1 (1 mg/L auxin and 0.5 mg/L cytokinin) were tested for callus induction (Table S1). Twenty-five explants have been used for each combination of plant growth regulators. Cultures were incubated at 23 ± 2 °C in the dark and sub-cultured at 28-day intervals on the fresh medium for four months. The fresh weight of the callus biomass was determined as the total fresh weight of the same number of calli grown on individual combinations of growth hormones after 4, 8, 12, and 16 weeks, respectively.
4.3. Cell Suspension Cultures
Cell suspension cultures were derived from calli developed from leaf and root explants on the liquid MS medium supplemented with 1.0 mg/L 2.4-D + 0.5 mg/L BAP. Cultivation was at the same temperature (23 ± 2 °C) either in darkness or under the photoperiod (16 hours of light and 8 hours of darkness) with a light intensity of 50 μmol.m-2.s-1. Cells were cultivated under constant stirring (VS-202P, Vision Scientific Co., Ltd., Daejeon, Korea) in 100 ml Erlenmayer flasks at 110 rpm. The optimal sub-cultivation interval was identified by observation of the cell growth rate by regular cell samplings during 18 days of cultivation and determination of fresh cell weight and dry cell weight.
The fresh weight of cells was calculated by weighing the cell residue on filter paper after filtering the entire volume of the flask and subtracting the weight of the filter paper. The dry weight was determined by drying the filtered fresh cells in an oven at 65 °C for 2 hours.
Cell viability was determined by staining procedures using two stains. Cell suspension was mixed with a 0.1% (w/v) solution of Evan’s blue in a ratio 5:1 or with a 0.4% solution of Trypan blue (both from Merck KGaA, Darmstadt, Germany) in a ratio 1:1. Viable or dead cells were detected microscopically (Leica DM6000 Upright Optical Microscope, Leica Microsystems GmbH, Wetzlar, Germany). Observations were performed in three replicates.
4.4. Stem Cell Cultures
Microscopic analysis of cells was performed in developed callus tissues by dyeing vacuoles using the modified method of Lee et al. (2010). Cells taken from callus biomass were stained with 0.01% (w/v) Neutral Red (3-amino-7-dimethyl-amino-2-methylphenazine hydrochloride, Merck KGaA, Darmstadt, Germany) for 10 min and washed with 0.1 M phosphate buffer, pH 7.2. Samples were prepared by the squash smear technique and observed using the Leica DM6000 Upright Optical Microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Stem cells in suspension culture were identified by the same method as in callus cultures. However, cell suspension was mixed with Neutral Red in a ratio 1:1, centrifuged for 5 min at 2,500 rpm, washed with 0.1 M phosphate buffer, pH 7.2, and counted in the Fuchs-Rosenthal Counting Chamber (Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany) under the same microscope. The experiments were performed in three replicates.
4.5. Data Analysis
The effects of used growth regulators for callus induction and growth were analyzed in five replications, each with five leaf- or root-derived explants, respectively. Evaluated parameters included the day of the initial callus formation, the fresh weight of the callus, the percentage of callus formation, as well as the morphology parameters, including color and structure of the callus. Obtained data were evaluated by the analysis of variance (ANOVA) using Statgraphics software version 19.2.01 (Statgraphics Technologies, Inc., The Plains, VA, USA). Significant differences between means were compared using the least significant difference (LSD) test at the 5% level of significance (p < 0.05).
5. Conclusions
Meristematic cells contained in primary explants or formed de novo in tissue cultures can determine the establishment of high-effective production of cell biomass through callus tissue and cell suspension culture. However, much more interesting and important is the possibility to establish the formation, respectively to select stem cells and cultivate them in a liquid medium as a cell suspension with a very high proportion of stem cells. In the presented work, such a culture of stem cells was established from segments of leaves and roots of C. officinalis L. In the case of this medicinal plant, it seems that the way to establish stem cell culture systems is neither impossible nor too difficult. It can open new approaches to the production of cell biomass and secondary metabolites for different ways of their application.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Effects of explant types and plant growth regulators on induction and growth of callus biomass of Calendula officinalis L.
Author Contributions
Conceptualization, Š.K. and J.K.; methodology, Š.K., M.G; software, Š.K. and J.K.; validation, J.K.; investigation, Š.K. and K.L.; data curation, Š.K. and J.K.; writing—original draft preparation, Š.K.; writing—review and editing, J.K.; visualization, Š.K. and J.K.; supervision, J.K.; funding acquisition, J.K. and M.G. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ashwlayan, V.D.; Kumar, A.; Verma, M.; Garg, V.K.; Gupta, S.K. Therapeutic potential of Calendula officinalis. Pharm. Pharmacol. Int. J. 2018, 6, 149–155. [Google Scholar] [CrossRef]
- Jan, N.; Andrabi, K.I.; John, R. Calenula officinalis—An important medicinal plant with potential biological properties. Proc. Indian Natl. Sci. Acad. 2017, 4, 769–787. [Google Scholar]
- Xuan, S.H.; Kim, G.Y.; Yu, J.Y.; Kim, J.W.; Yang, Y.R.; Jeon, Y.H; Jeong, Y.J.; Kim, A.R.; Park, S.N. Antioxidant and cellular protective effects against oxidative stress of Calendula officinalis flowers extracts in human skin cells. Appl. Chem. Eng. 2016, 27, 620–626. [Google Scholar] [CrossRef]
- Akhtar, N.; Zaman, S.U.; Khan, B.A.; Amir, M.N.; Ebrahimzadeh, M.A. Calendula extract: Effects on mechanical parameters of human skin. Acta Pol. Pharm. 2011, 68, 603–701. [Google Scholar]
- Szopa, A.; Klimek-Szczykutowicz, M.; Jafernik, K.; Koc, K.; Ekiert, H. Pot marigold (Calendula officinalis L.)—A position in classical phytotherapy and newly documented activities. Acta Sci. Pol. Hortorum Cultus 2020, 19, 47–61. [Google Scholar] [CrossRef]
- Andersen, F.A.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks Jr, J.G.; Shank, R.C.; Slaga, T.J.; Snyder, P.W. Final Report of the Cosmetic Ingredient Review Expert Panel amended safety assessment of Calendula officinalis–derived cosmetic ingredients. Int. J. Toxicol. 2010, 29S4, 221S–243S. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, V.; Slavov, A.; Vasileva, I.; Pavlov, A. Plant cell culture as emerging technology for production of active cosmetic ingredients. Eng. Life Sci. 2018, 18, 779–798. [Google Scholar] [CrossRef]
- Çöçü, S.; Uranbey, S.; Ipek, A.; Khawar, K.M.; Sarihan, E.O.; Kaya, M.D.; Parmaksız, I.; Özcan, S. Adventitious shoot regeneration and micropropagation in Calendula officinalis L. Biol. Plant. 2004, 48, 449–451. [Google Scholar] [CrossRef]
- Wiktorowska, E.; Dlugosz, M.; Janiszowska, W. Significant enhancement of oleanolic acid accumulation by biotic elicitors in cell suspension cultures of Calendula officinalis L. Enzyme Microb. Technol. 2010, 46, 14–20. [Google Scholar] [CrossRef]
- Długosz, M.; Wiktorowska, E.; Wiśniewska, A.; Pączkowski, C. Production of oleanolic acid glycosides by hairy root established cultures of Calendula officinalis L. Acta Biochim. Pol. 2013, 60, 467–473. [Google Scholar] [CrossRef]
- Çetin, B.; Kalyoncu, F.; Kurtuluş, B. Antibacterial activities of Calendula officinalis callus extract. Int. J. Sec. Metabolite 2017, 4, 257–263. [Google Scholar] [CrossRef]
- Grzelak, A.; Janiszowska, W. Initiation and growth characteristics of suspension cultures of Calendula officinalis cells. Plant Cell, Tissue Organ Cult. 2002, 71, 29–40. [Google Scholar] [CrossRef]
- Auguścińska, E.; Kasprzyk, Z. Studies on the labelling of terpenoids in shoots and cells or protoplasts from Calendula officinalis leaves. Acta Biochim. Pol. 1982, 29, 7–13. [Google Scholar] [PubMed]
- Długosz, M.; Markowski, M.; Pączkowski, C. Source of nitrogen as a factor limiting saponin production by hairy root and suspension cultures of Calendula officinalis L. Acta Physiol. Plant. 2018, 40, 35. [Google Scholar] [CrossRef]
- Alsoufi, A.S.M.; Pączkowski, C.; Szakiel, A.; Długosz, M. Effect of jasmonic acid and chitosan on triterpenoid production in Calendula officinalis hairy root cultures. Phytochem. Lett. 2019, 31, 5–11. [Google Scholar] [CrossRef]
- Rogowska, A.; Paczkowski, C.; Szakiel, A. Modulation of steroid and triterpenoid metabolism in Calendula officinalis plants and hairy root cultures exposed to cadmium stress. Int. J. Mol. Sci. 2022, 23, 5640. [Google Scholar] [CrossRef] [PubMed]
- Mehrabi, A.A.; Khodadadi, E.; Sadeghi, Z.; Lia shooshtari. An investigation of tissue culture and co-cultures of different explants in Calendula officinalis. Int. J. Biosci. 2013, 12, 201–205. [Google Scholar] [CrossRef]
- Kaya, N.; Aki, C. In vitro effects of plant growth regulators on callus formation in Calendula officinalis L. and Calendula arvensis L. species. Ann. Biol. Res. 2017, 8, 1–7. [Google Scholar]
- Al-Abasi, I.N.; Bashi, B.Z.K.; Al-Mallah, M.K. Design of culture medium and leaf clones are determinant factors in callus induction of Calendula officinalis L. Eur. Acad. Res. 2018, 6, 1901–1913. [Google Scholar]
- Efferth, T. Biotechnology applications of plant callus cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Kolewe, M.E.; Gaurav, V.; Roberts, S.C. Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol. Pharm. 2008, 5, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.K.; Jin, Y.W.; Park, J.H.; Yoo, Y.M.; Hong, S.M.; Amir, R.; Yan, Z.; Kwon, E.; Elfick, A.; Tomlinson, S.; Halbritter, F.; Waibel, T.; Yun, B.-W.; Loake, G.J. Cultured cambial meristematic cells as a source of plant natural products. Nat. Biotechnol. 2010, 28, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Villarreal, M.; Howat, S.; Jang, M.O.; Kim, I.S.; Jin, Y.-W.; Lee, E.-K.; Loake, G.J. Cambial meristematic cells: a platform for the production of plant natural products. New Biotechnol., 2015, 32, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.H. Vascular tissue differentiation and pattern formation in plants. Annu. Rev. Plant Biol. 2002, 53, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Laux, T. The stem cell concept in plants: a matter of debate. Cell 2003, 113, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.W.; Yan, Z.; Amir, R.; Hong, S.; Jin, Y.W.; Lee, E.K.; Loake, G.J. Plant natural products: history, limitations and the potential of cambial meristematic cells. Biotechnol. Genet. Eng. Rev. 2012, 28, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.B.; Elias, C.B.; Patole, M.S. Role of hydrodynamic shear in the cultivation of animal, plant and microbial cells. Chem. Eng. J., 1996, 62, 121–141. [Google Scholar] [CrossRef]
- Lee, S.B.; Cho, H.I.; Jin, Y.W.; Lee, E.K.; Ahn, J.Y.; Lee, S.M. Wild ginseng cambial meristematic cells ameliorate hepatic steatosis and mitochondrial dysfunction in high-fat diet-fed mice. J. Pharm. Pharmacol. 2016, 68, 119–127. [Google Scholar] [CrossRef]
- Moon, S.H.; Venkatesh, J.; Yu, J.W.; Park, S.W. Diferential induction of meristematic stem cells of Catharanthus roseus and their characterization. C. R. Biol. 2015, 338, 745–756. [Google Scholar] [CrossRef]
- Song, Y.; Chen, S.; Wang, X.; Zhang, R.; Tu, L.; Hu, T.; Liu, X.; Zhang, Y.; Huang, L.; Gao, W. A novel strategy to enhance terpenoids production using cambial meristematic cells of Tripterygium wilfordii Hook. f. Plant Methods 2019, 15, 129. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, J.; Qin, N.; Zhang, Q.; Yan, C. Biotransformation of 4-methylcoumarins by cambial meristematic cells of Camptotheca acuminate. RSC Adv. 2019, 9, 9449. [Google Scholar] [CrossRef] [PubMed]
- Mehring, A.; Haffelder, J.; Chodorski, J.; Stiefelmaier, J.; Strieth, D.; Ulber, R. Establishment and triterpenoid production of Ocimum basilicum cambial meristematic cells. Plant Cell Tissue Organ Cult. 2020, 143, 573–581. [Google Scholar] [CrossRef]
- He, L.; Zhang, J.; Guo, D.; Tian, H.; Cao, Y.; Ji, X.; Zhan, Y. Establishment of the technology of cambial meristematic cells (CMCs) culture from shoots and high expression of FmPHV (PHAVOLUTA) functions in identification and differentiation of CMCs and promoting the shoot regeneration by hypocotyl in Fraxinus mandshurica. Plant Physiol. Biochem. 2021, 160, 352–364. [Google Scholar] [CrossRef]
- Legha, M.R.; Prasad, K.V.; Singh, S.K.; Kaur, C.; Arora, A.; Kumar, S. Induction of carotenoid pigments in callus cultures of Calendula officinalis L. in response to nitrogen and sucrose levels. In Vitro Cell. Dev. Biol.—Plant 2012, 48, 99–106. [Google Scholar] [CrossRef]
- Leal, F.; Rodrigues, A.; Fernandes, D.; Nunes, F.M.; Cipriano, J.; Ramos, J.; Teixeira, S.; Vieira, S.; Carvalho, L.M.; Pinto-Carnide, O. In vitro multiplication of Calendula arvensis for secondary metabolites extraction. In Proceedings of the IIIrd International Symposium on Acclimatization and Establishment of Micropropagated Plants, Faro, Portugal, 28 February 2009. [Google Scholar]
- Ibrahim, M.M.; Abed, R.M.; Ali, F.Q. Influence of biotic elicitor Aspergillus niger on salicylic acid products in callus cultures of Calendula officinalis L. plant. J. Phys.: Conf. Ser. 2019, 1294, 062016. [Google Scholar] [CrossRef]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: dedifferentiation, transdifferentiation, or just differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef]
- Grafi, G. How cells dedifferentiate: a lesson from plants. Dev. Biol. 2004, 268, 1–6. [Google Scholar] [CrossRef]
- Verdeil, J.-L.; Alemanno, L.; Niemenak, N.; Tranbarger, T.J. Pluripotent versus totipotent plant stem cells: dependance versus autonomy? Trends Plant Sci. 2007, 12, 245–252. [Google Scholar] [CrossRef]
- Sugimoto, K.; Jiao, Y.; Meyerowitz, E.M. Arabidopsis regeneration from multiple tissues occurs via a root development pathway. Dev. Cell 2010, 18, 463–471. [Google Scholar] [CrossRef]
- Parizot, B.; Laplaze, L.; Ricaud, L.; Boucheron-Dubuisson, E.; Bayle, V.; Bonke, M.; De Smet, I.; Poethig, S.R.; Helariutta, Y.; Haseloff, J.; Chriqui, D.; Beeckman, T.; Nussaume, L. Diarch symmetry of the vascular bundle in Arabidopsis root encompasses the pericycle and is reflected in distich lateral root initiation1[W]. Plant Physiol. 2008, 146, 140–148. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Tsukaya, H. Behavior of leaf meristems and their modification. Front. Plant Sci. 2015, 6, 1060. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, P.M.; Bonetta, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell cycling and cell enlargement in developing leaves of Arabidopsis. Dev. Biol. 1999, 215, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Maksymowych, R.; Erickson, R.O. Development of the lamina in Xanthium italicum represented by the plastochron index. Am. J. Bot. 1960, 47, 451–459. [Google Scholar] [CrossRef]
- Alvarez, J.P.; Furumizu.; Efroni.; Eshed Y.; Bowman J.L. Active suppression of a leaf meristem orchestrates determinate leaf growth. eLife 2016, 5, e15023. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Guan, C.; Jiao, Y. Molecular mechanisms of leaf morphogenesis. Mol. Plant 2018, 11, 1117–1134. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Seo, P.J. De novo shoot organogenesis during plant regeneration. J Exp. Bot. 2020, 71, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J. 2009, 57, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Qi, M.; Xu, L.; Zhang, G.; Li, J.; Qin, P.; Ruan, Y.; Huang, H.; Zhang, Y.; Xu, L. Divergent regeneration-competent cells adopt a common mechanism for callus initiation in angiosperms. Regeneration 2017, 3, 132–139. [Google Scholar] [CrossRef]
- Müller-Xing, R.; Xing, Q. The plant stem-cell niche and pluripotency: 15 years of an epigenetic perspective. Front. Plant Sci. 2022, 13, 1018559. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Seth Favero, D.; Sakamoto, Y.; Iwase, A.; Coleman, D.; Rymen, B.; Sugimoto, K. Molecular mechanisms of plant regeneration. Annu. Rev. Plant Biol. 2019, 70, 3.1–3.30. [Google Scholar] [CrossRef]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
Figure 1.
Fresh weight of calli induced from leaf and root explants using different combinations of growth regulators, after 4, 8, 12, and 16 weeks (graphs from top to bottom) of cultivation. Data are presented as means ± SDs (n = 5).
Figure 1.
Fresh weight of calli induced from leaf and root explants using different combinations of growth regulators, after 4, 8, 12, and 16 weeks (graphs from top to bottom) of cultivation. Data are presented as means ± SDs (n = 5).
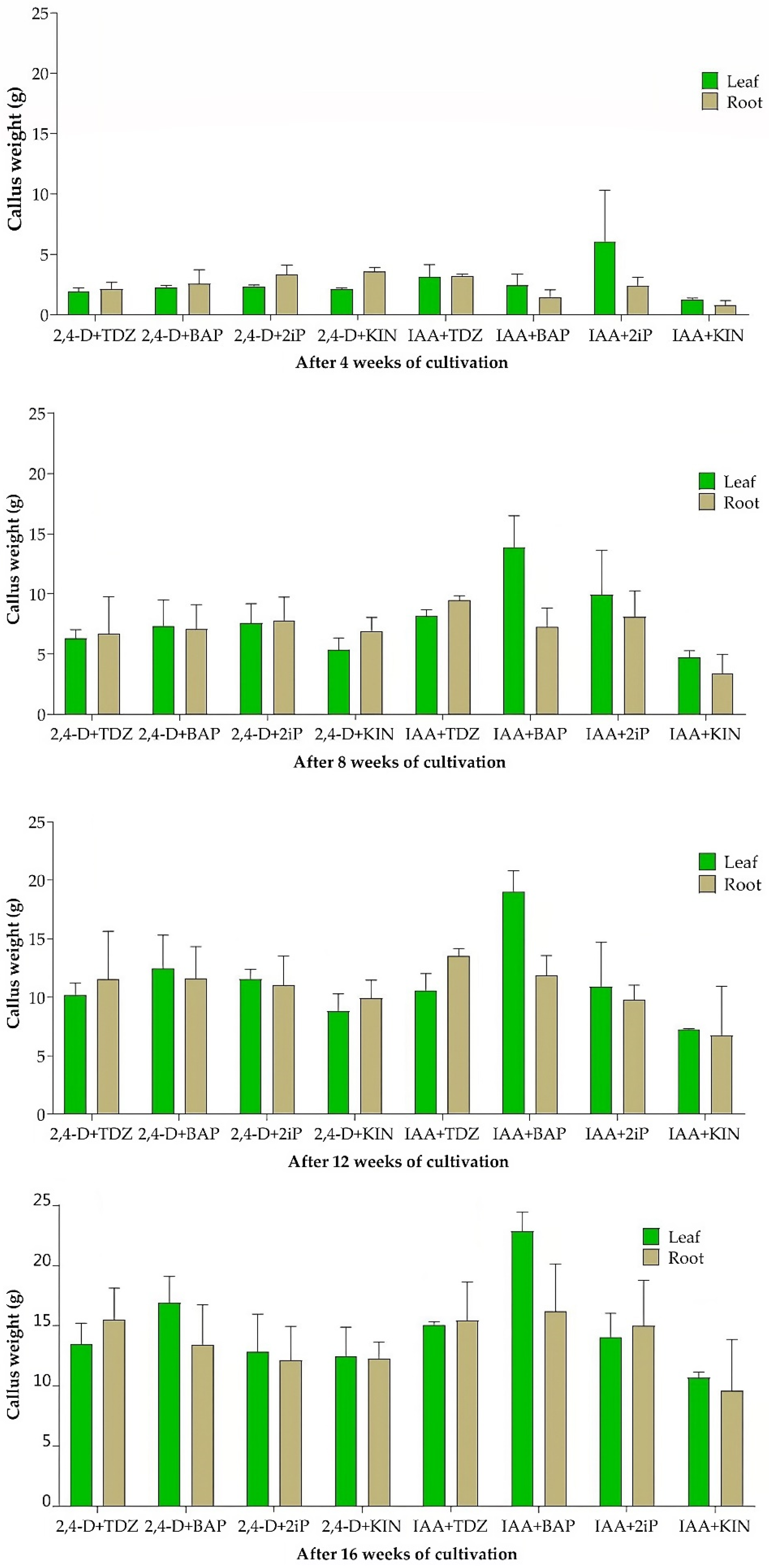
Figure 2.
Increase in fresh weight of callus biomass derived from leaf (green curves) and root (brown curve) explants within 16 weeks of cultivation on the most effective combinations of growth regulators.
Figure 2.
Increase in fresh weight of callus biomass derived from leaf (green curves) and root (brown curve) explants within 16 weeks of cultivation on the most effective combinations of growth regulators.
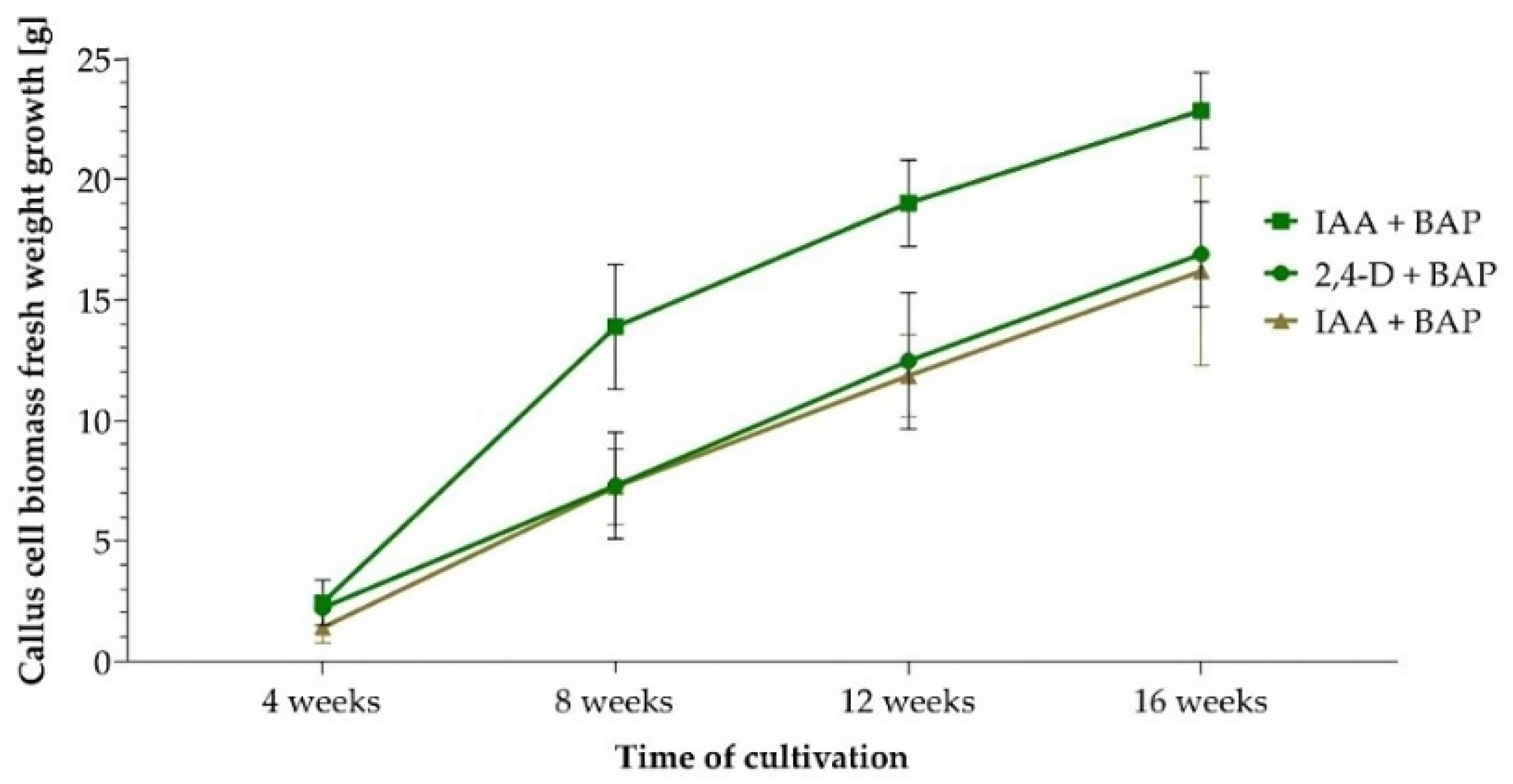
Figure 3.
Different types of calli (color, texture) on the same leaf explant (SCs: red arrows, DDCs: black arrows).
Figure 3.
Different types of calli (color, texture) on the same leaf explant (SCs: red arrows, DDCs: black arrows).

Figure 4.
Abundant very small vacuoles in SCs (a) and one large vacuole in DDC (b).
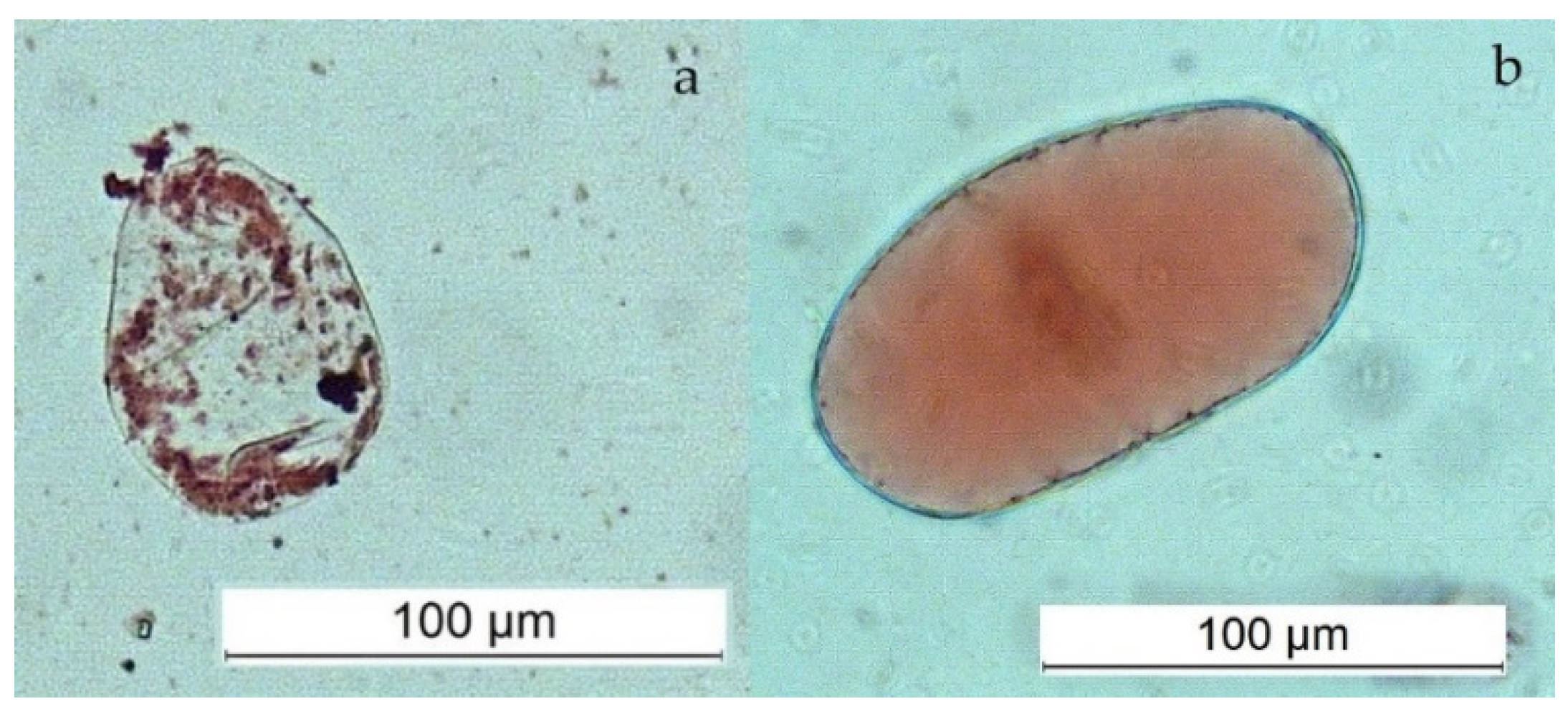
Figure 5.
SCs (red arrows), DDCs (black arrows), and other cells in cell biomass derived from leaf (a) or root (b) explants formed in medium containing 1.0 mg/L 2.4-D + 0.5 mg/L BAP.
Figure 5.
SCs (red arrows), DDCs (black arrows), and other cells in cell biomass derived from leaf (a) or root (b) explants formed in medium containing 1.0 mg/L 2.4-D + 0.5 mg/L BAP.
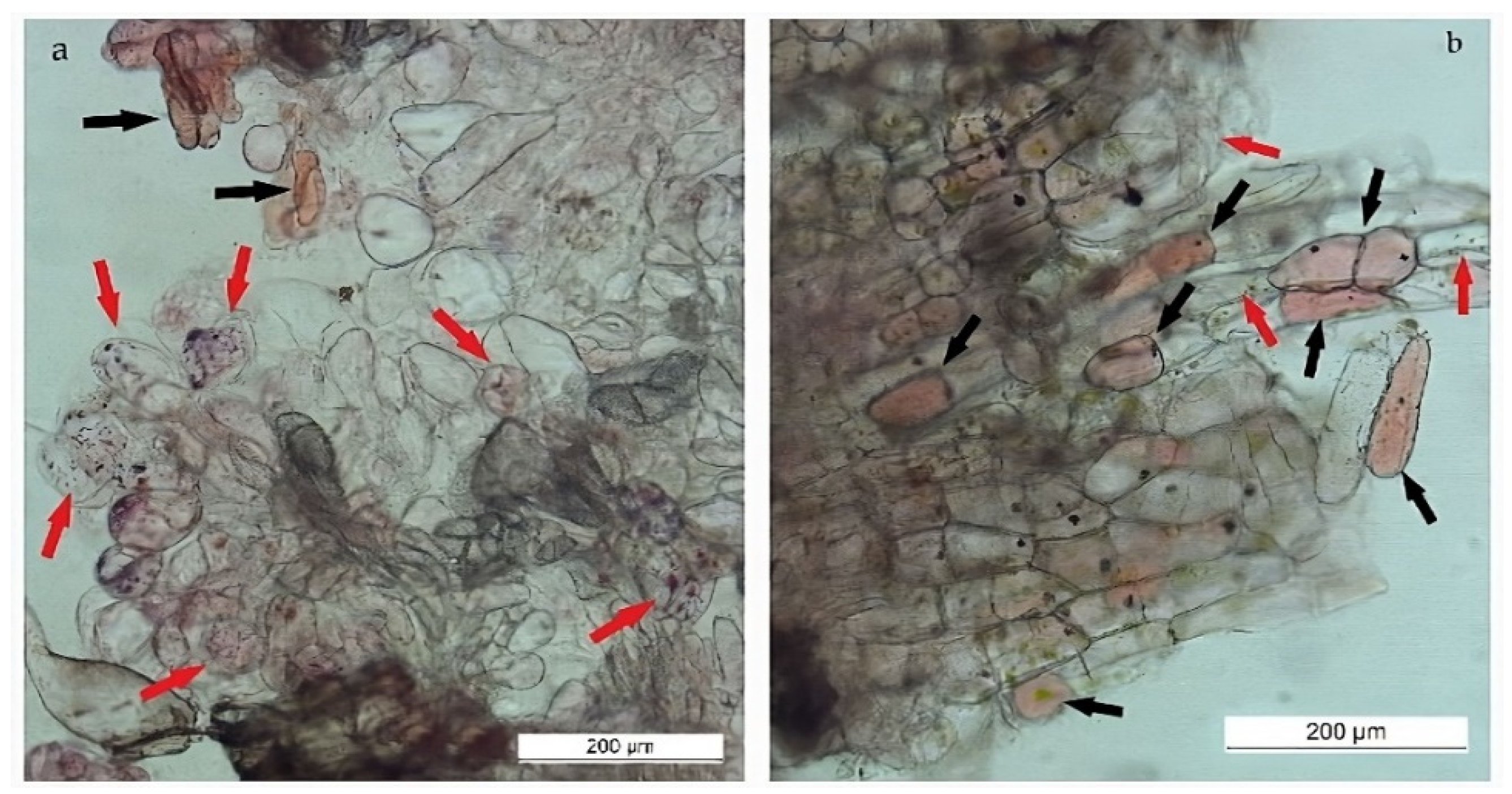
Figure 6.
SCs and DDCs in callus biomass derived from leaf and cultivated in MS medium containing 1.0 mg/L 2.4-D + 0.5 mg/L BAP (left) or 1.0 mg/L IAA + 0.5 mg/L BAP (right). The red and black arrows point to SCs and DDCs, respectively.
Figure 6.
SCs and DDCs in callus biomass derived from leaf and cultivated in MS medium containing 1.0 mg/L 2.4-D + 0.5 mg/L BAP (left) or 1.0 mg/L IAA + 0.5 mg/L BAP (right). The red and black arrows point to SCs and DDCs, respectively.
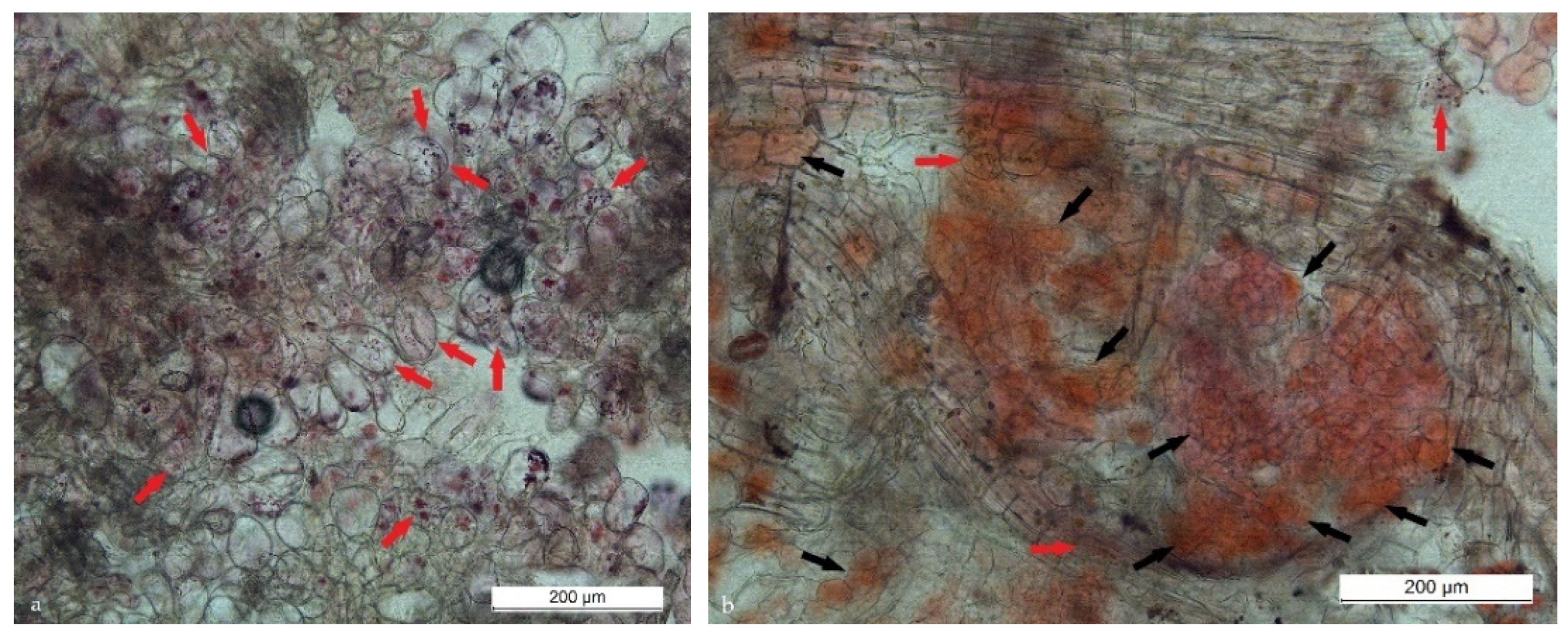
Figure 7.
Fresh weight of cells (g/L) in suspension culture originated from leaf-derived (CL) and root-derived (CR) callus, cultivated in continuous darkness (left) or under the photoperiod (right).
Figure 7.
Fresh weight of cells (g/L) in suspension culture originated from leaf-derived (CL) and root-derived (CR) callus, cultivated in continuous darkness (left) or under the photoperiod (right).
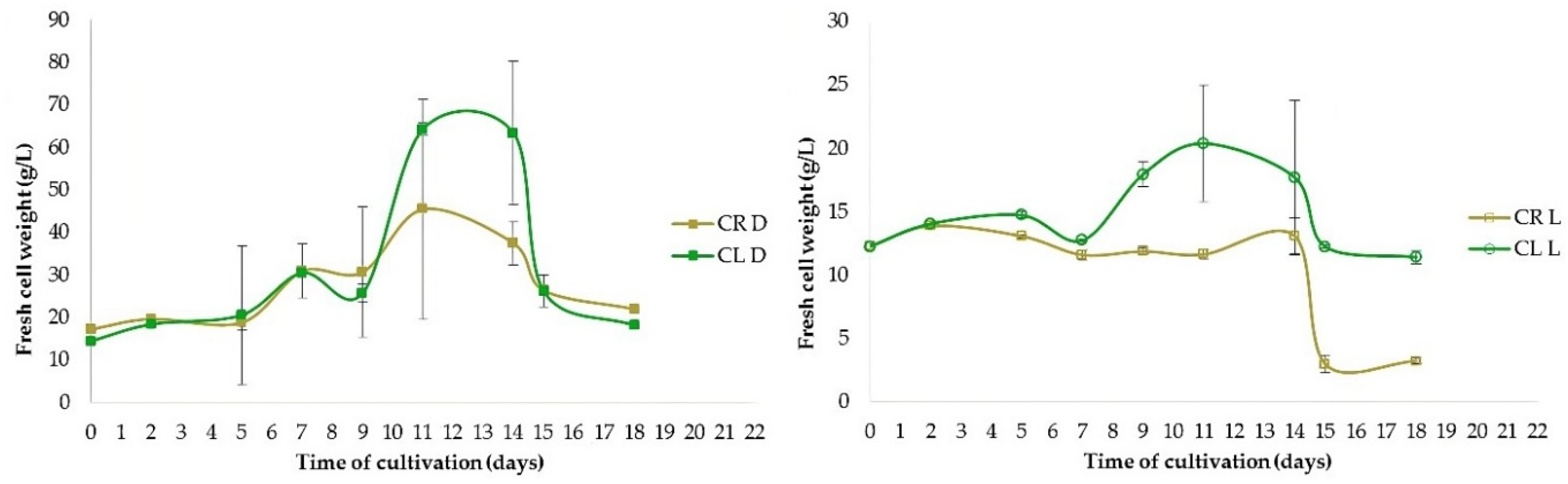
Figure 8.
The viability of cells in leaf-derived (CL) and root-derived (CR) suspension cultures cultivated in continuous darkness assayed by Evans blue and Trypan blue.
Figure 8.
The viability of cells in leaf-derived (CL) and root-derived (CR) suspension cultures cultivated in continuous darkness assayed by Evans blue and Trypan blue.
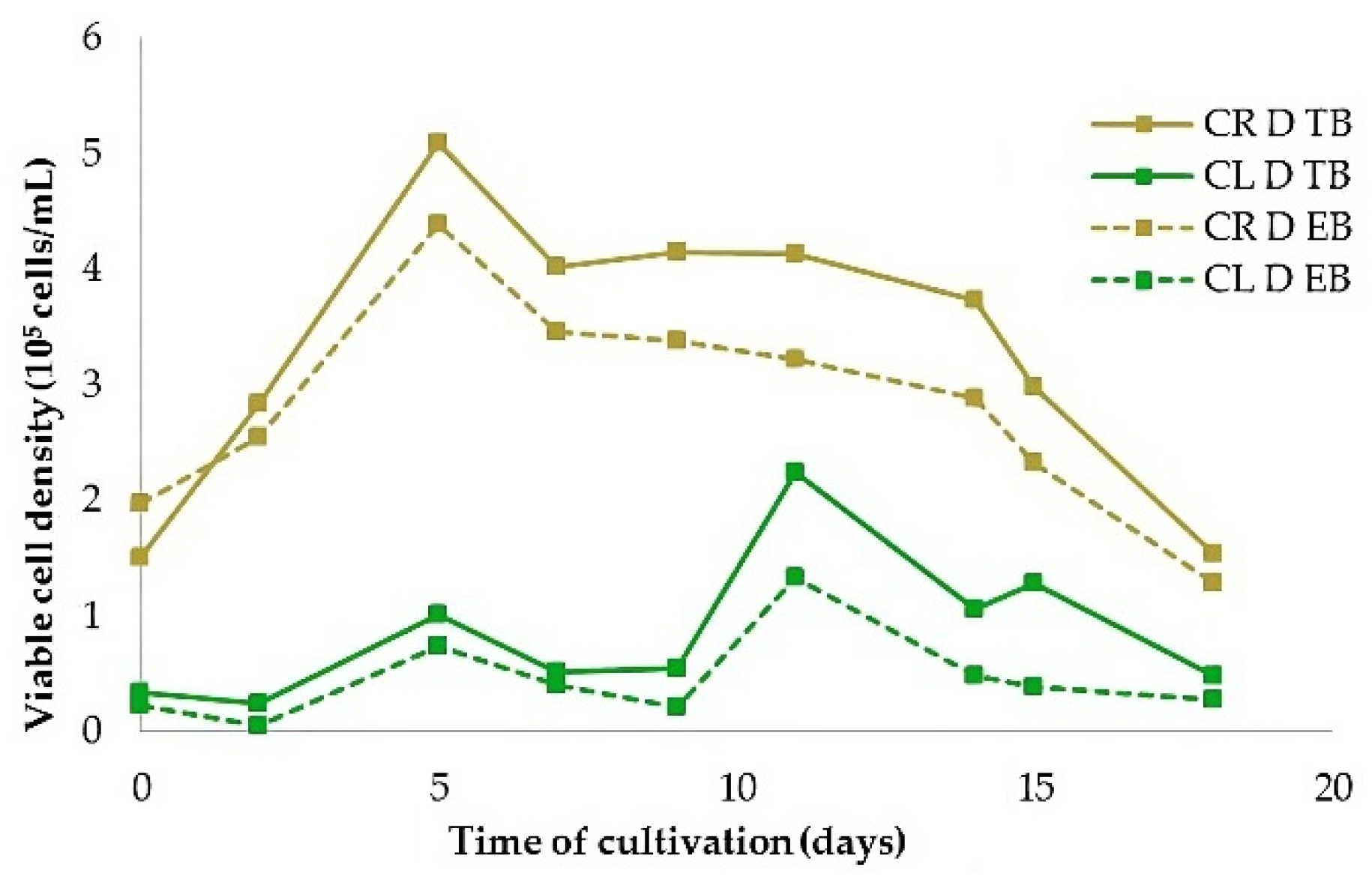
Figure 9.
SCs and a small amount of DDCs (black arrow) in stem cell suspension culture derived from root-derived callus.
Figure 9.
SCs and a small amount of DDCs (black arrow) in stem cell suspension culture derived from root-derived callus.
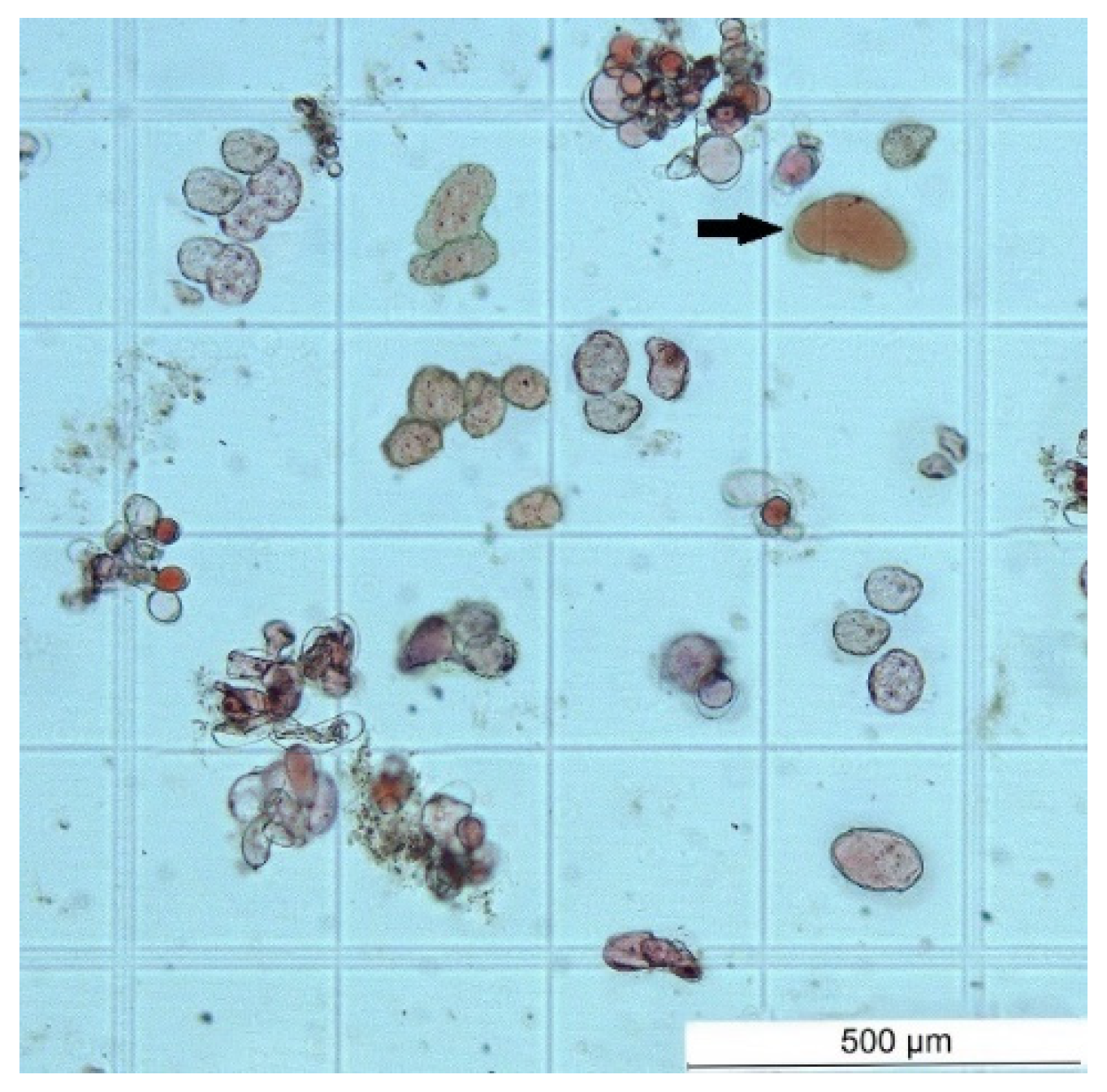
Figure 10.
Proportion of stem cells (top left) and dedifferentiated cells (top right) in cell suspension cultures derived from leaves (CL) and roots (CR) and the number of both cell types (bottom) cultivated in continuous darkness.
Figure 10.
Proportion of stem cells (top left) and dedifferentiated cells (top right) in cell suspension cultures derived from leaves (CL) and roots (CR) and the number of both cell types (bottom) cultivated in continuous darkness.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



