
Submitted:
22 April 2023
Posted:
23 April 2023
You are already at the latest version
Abstract
During embryonic development stem cells undergo the differentiation process so that they can specialise for different functions within the organism. Complex programs of gene transcription are crucial for this process to happen. Epigenetic modifications and the architecture of chromatin in the nucleus, by the formation of specific regions of active as well as inactive chromatin, allow the coordinated regulation of the genes for each cell fate. In this mini review, we discuss the current knowledge regarding the regulation of three-dimensional chromatin structure during neuronal differentiation. We also focus on the role played in neurogenesis by the nuclear lamina that ensures the tethering of the chromatin to the nuclear envelope.
Keywords:
chromatin organization
; nuclear lamina
; epigenetics
; neuronal differentiation
1. Introduction
The DNA sequence of all cells in the body is the same even though the cells do not play the same function. Each cell type is characterised by a precise orchestration of gene expression for the production of all proteins necessary for specific cellular function. Hence, not all genes are active in all cells at any given time and their expression is tightly regulated by the epigenome. The Greek preposition “epi” means on or above, and "epigenome" takes into account all those chemical modifications which occur “on” the DNA molecule, without affecting its sequence. The epigenome acts as a film director, assigning different roles to each cell type by defining when and what genes are accessible to transcription factors to regulate their expression. Also, the epigenome contributes to gene expression by modification of the three-dimensional organization of the genetic material (chromatin). Chromatin is organised in nuclear topological domains that differ depending on the cell type and stage of differentiation. The nuclear spatial organization, or nuclear architecture, ensures the correct performance of the transcriptional programs within cells, thus, giving it an important role in cellular differentiation and development. Furthermore, nuclear architecture is dictated not only by the epigenome but also by the nuclear lamina which both physically supports the nucleus and anchors chromatin to the nuclear envelope [1]. This suggests that the nuclear lamina could also be involved in the establishment of the cell transcriptional programs. In this review, we will focus on the current knowledge about the role played by chromatin organization and nuclear lamina in mammals’ neurodevelopment. We will first give an overview of the three-dimensional organization of chromatin and the nuclear envelope structure. Afterwards, we will give a general introduction about the developmental process of the nervous system. We will then discuss examples showing the importance of chromatin organization and nuclear lamina in the development and differentiation of neuronal cells.
2. Three-dimensional organization of chromatin within the nucleus
The three-dimensional organization of chromatin within the nucleus is critical for the regulation of gene expression and cellular function. In eukaryotic cells, the DNA is organized with different levels of compaction to fit within the micron-sized nuclear space. The DNA wraps around a complex of eight histone proteins constituting the nucleosomes, resulting in the 10 nm fibre known as chromatin that can reorganise into a 30 nm fibre at least in vitro; however, chromatin is mainly found as a disorganized structure, heterogeneous and diverse in diameter and with a high bendability [2]. Indeed, chromatin can organize itself into loops due to the presence of cohesins and other architectural proteins [3]. These chromatin loops are also able to interact with each other creating a chromatin-based physical compartmentalization in megabase-scale genomic domains called “Topologically Associating Domains” (TADs) [4,5]. Although genomic regions within the same TAD can interact regardless of the distance separating them, there is usually no contact with regions outside the domain [5]. The presence of TAD boundaries, constituted by architectural proteins that interact with the TAD, restrains the interactions between regulatory sequences (i.e. enhancer, silencer) and target genes of different TADs [6]. In this way, TADs greatly contribute to gene expression regulation. In interphase nuclei, TADs belonging to the same chromosome occupy distinct regions known as Chromosomal Territories [7,8] inside of which two types of compartments can be identified. The “A” compartment is the more relaxed configuration of chromatin fibre called Euchromatin which is usually actively transcribed and mostly found at the interior of the nucleus; whereas the “B” compartment is the more compact chromatin called Heterochromatin which is mainly transcriptionally repressed and found preferentially at the nuclear periphery and nucleolus [1,9,10] (details of eu- and heterochromatin will be discussed below in this paragraph). In this regard, it has been observed that some specific TADs are associated with the nuclear lamina (see the paragraph 3. Nuclear envelope: LINC complex and nuclear lamins), an intermediate filament network lining the inner surface of the nuclear envelope. This association forms the so-called “Lamina Associated Domains” (LADs) with a repressive role in gene expression [11].
From the epigenetic point of view, chromatin compartmentalization and degree of compaction are determined by specific modifications of both histones and DNA. The relaxed configuration of euchromatin is accompanied by histone acetylation that neutralizes the positive charge of lysine residues favouring chromatin opening [12]. Also, transcriptionally active genes are usually marked by H3K4 methylation [13]. On the other hand, condensed chromatin is induced by the deacetylation of histones, DNA methylation, and methylation of specific residues of histones. Characteristic markers of heterochromatin are trimethylated H3K9, trimethylated H3K27, and trimethylated H4K20 [1,14].
3. Nuclear envelope: LINC complex and nuclear lamins
The nuclear envelope (NE) is made up of the outer nuclear membrane (ONM) and the inner nuclear membrane (INM) and acts as a physical barrier that separates the cell nucleus harboring chromatin from the cell cytoplasm [1]. Between the INM and the ONM lies the perinuclear space (PNS), with a width of 30-50 nm [15], and juxtaposed to the nucleoplasmic side of the INM there is the nuclear lamina.
The LINC complex (Linker of Nucleoskeleton and Cytoskeleton) spans the INM and ONM and provides mechanical coupling between the actin cytoskeleton and the nucleus [16,17]. This is possible thanks to both the ONM and the INM components of the LINC complex, the nesprins (nuclear envelope spectrin repeat proteins), and the SUN proteins (Sad1p and UNC-84 homology), respectively [18,19,20]. Specifically, nesprin proteins act as a bridge between the cell cytoskeleton and the INM by binding actin on one side and SUN proteins on the other side; whereas, SUN proteins interact on the nucleoplasmic side with either the nesprins in the PNS or the nuclear lamins underneath the INM, thus, indirectly connecting cell cytoskeleton with the nuclear lamina [15].
In humans, three genes encode the components of the nuclear lamina, LMNA, LMNB1 and LMNB2. The products of LMNA gene are lamins A and C, which are translated following an alternative splicing event on exon 10. LMNB1 and LMNB2 genes encode Lamin B1 and Lamin B2, respectively [15]. Lamin B1 is bound by LBR (Lamin B Receptor) which, in turn, interacts with MeCP2 (Methyl-CpG binding protein 2) and HP1α (Heterochromatin Protein 1) that are responsible for the binding of 5-Methylcytosine and H3K9me3 on the DNA respectively, both epigenetic markers of heterochromatin [1,21,22].
4. Neurogenesis: an overview
During embryogenesis, at the beginning of gastrulation, human Embryonic Stem Cells (hESCs) from the ectoderm start to give rise to human Neural Progenitor Cells (hNPCs) characterized by a radial alignment and a bipolar morphology. These cells undergo symmetric divisions for self-renewal to increase the size of the cell pool, which, by the end of gastrulation, forms the neuronal plate along the rostral-caudal midline of the upper layer of the embryo. The ridges of the neural plate then fold inward to create the neural tube [26]. At this point, the hNPCs, depending on their position, will differentiate into either neurons or glia to construct the nervous system. The rostral region of the neural tube will give rise to the brain meanwhile, the caudal region will give rise to the hindbrain and spinal column [26]. In the process mentioned above, in addition to the members of the TGF-β family (that have different roles, from maintaining the pluripotency of embryonic stem cells to the mesenchymal differentiation) both chromatin organization and epigenome are crucial players. In this review we have decided not to tackle the issue of epigenetic modifications characterizing neuronal development that has been extensively reviewed recently [27,28]. Instead, we have focused on the chromatin structure changes and their relationship with nuclear lamina during neuronal development.
5. Chromatin structure involvement in neural development
Epigenetic modifications are usually associated with the regulation of the development and differentiation processes. However, the different compartmentalization of chromatin also plays an important role in these contexts. Studies on murine Embryonic Stem cells (mESCs) as well as on hESCs revealed, by transmission electron microscopy, that undifferentiated ESCs have euchromatin-rich nuclei with prominent nucleoli, whereas differentiating ESCs are characterized by increasingly condensed heterochromatin distributed in a diffuse granular pattern and as a dense strip beneath the nuclear edge [29]. These changes are accompanied by specific histone epigenetic modifications, mainly acetylation and trimethylation of lysine 9 of histone H3 which are important to determine the developmentally regulated genes as being active euchromatin or repressed heterochromatin. This is a fundamental step towards cell fate commitment during development.
In this regard, the analysis of the three-dimensional (3D) organization of the genome during neuronal murine development showed that mESCs are characterized by open chromatin with epigenetic marks of active gene expression (i.e. high levels of H3/H4 acetylation and trimethyl H3K4, or low levels of histone trimethylation on lysine 27), and few compacted chromatin domains. Instead, murine Neural Progenitor Cells (mNPCs) have condensed chromatin with more heterochromatic domains clustered in chromocenters, bright DAPI-positive domains of constitutive heterochromatin [30,31,32,33]. Nakao’s group also showed that even mESCs are characterized by chromocenters that are smaller in mNPCs and integrated into larger foci in post-mitotic neurons (mPMNs) [31]. The number and shape of chromocenters also changes during the differentiation of the neural cell types [34,35,36,37]. Specifically, chromocenter numbers decrease in murine Purkinje cells from the day of birth till postnatal day 6, and, then, increase till mice become adults [35]. Also, the deposition of epigenetic markers is involved, such as the active histone mark trimethylated lysine 4 of histone H3 that increases at chromocenters during neuronal differentiation in the neocortex. This was accompanied by a parallel increase in the transcription of major satellites [38]. In accordance with changes in the chromatin state (open/closed), the interior of the TADs changes, however, the boundaries of TADs stay invariant during development [39]. Interestingly, by inducing mESCs to become Neuronal Committed Cells (mNCCs) with Retinoic Acid, Stachowiak’s group identified 3965 TADs in mESCs and 3953 TADs in mNCCs, with relocation of TADs in mNCCs compared to the mESCs [40]. Specifically, mNCCs increase the expression of Nuclear Fibroblast Growth Factor Receptor 1 (nFGFR1) [40], which strongly correlates with neuronal differentiation by regulating pluripotency genes [41]. In addition, nFGFR1 also works as a protein insulator leading to a reorganization of the chromatin loops and TADs [40]. Finally, CTCF insulator was reduced in comparison to the mESC [40]. From these studies, the reorganization of TADs has been found to be related to not only the loss of stem potency but also to cell differentiation.
6. Implication of nuclear lamina in neuronal development
The redistribution of chromatin is needed during development to specify the cell type’s fate and is essential to cell fitness and function. Alterations in key components of the chromatin 3D reorganization such as the nuclear lamina induce aberrations during development that potentially leads to organism death [42]. Indeed, a homozygous LMNA mutation leads to prenatal lethality in humans, whereas in mice LMNA mutation is not lethal and results in different pathologies a few weeks after birth [42]. Intriguingly, upon differentiation, some genes move towards or from the nuclear lamina [43] according to their activation or repression state. For example, mNPCs, after induction from mESC, showed relocation of the pluripotency genes towards the nuclear lamina, a position that is maintained even after further differentiation. Van Steensel’s group analysed mNPCs differentiation and discovered an increased interaction between the nuclear lamina and 633 genes. Some of which are “stemness” genes, i.e. Nanog, Klf4, and Oct4 [44]. Since these genes move to the nuclear periphery, they are usually associated with the LADs implying a heterochromatinization during differentiation [45,46]. Williams’ group analysed the positioning of Mash1, a proneuronal factor, and noticed that the Mash1 locus is mostly located at the nuclear periphery in ESCs that, upon neuronal induction, relocates to the interior part of the nucleus. Moreover, this study demonstrates that repositioning is directed in a cell-type specific manner. Indeed, other differentiated cell types were characterized by Mash1 located at the nuclear periphery similar to the ESCs [47]. However, it is important to mention that relocation is not necessarily equivalent to gene activation since transcription of some neuronal genes is associated with further differentiation [43]. This is the case for the brain Pcdh9 gene [44] and several neuronal genes that dissociate from the nuclear lamina even if they are not actively transcribed in mNPCs.
Considering the strong interdependence between chromatin organization, heterochromatin and nuclear lamina, as discussed above and in Carollo & Barra 2023 [1], it is not surprising that nuclear lamina and its mechanics have been shown to be important for neuronal development, as demonstrated by the elegant work conducted by the Young’s group. In 2011, they demonstrated that the deficiency of Lamin B1 (Lmnb1Δ/Δ) causes problems in the development of the cerebral cortex in mouse embryos, with impairments in neuronal migration as well [48]. This was accompanied by a reduction in neuronal progenitor cells and an increase in apoptotic cell death. Moreover, Lamin B1 deficiency was the cause of misshapen cell nuclei of cortical neurons, which has been correlated with the alteration of the heterochromatin:euchromatin ratio in other cell contexts [1]. In addition, KO of either Lamin B1 or Lamin B2 via Cre recombinase in mouse embryos caused both reduced cranium and cerebral cortex size. Cortex was also smaller and showed atrophy in double knockout Lamin B1 and Lamin B2 mice compared to the single KO condition. Moreover, adult mice lacking either Lamin B1 or Lamin B2 exhibited problems in the layering of cortical neurons, as demonstrated by the absence of Cux1, a marker of layer II/III, in most of the neurons. Lamin B1 KO neurons displayed nuclear blebs (one bleb/nucleus) with an asymmetric distribution of Lamin B2 [48]. Atypical nuclei can also be formed when Lamin B1 does not correctly localize in the nuclear envelope, which is the case of mouse mutants for Lamin B1 that cannot be farnesylated (Lmnb1CS/CS). The mutated Lamin B1 mislocalizes in the nucleus in a honeycomb fashion which correlates with strong defects in cell nucleus shape. Specifically, it has been observed that during in vitro migration, the NPCs of Lmnb1CS/CS mice have dumbbell-shaped nuclei and blebs. In these cells, Lamin B1 was mainly at the leading edge (towards the direction of the migration). Strikingly, the opposite side of the cell (the trailing edge) was occupied by the bulk of chromatin, called “naked chromatin” because disconnected from the nuclear lamina [49]. The authors supposed that the dumbbell-shaped nuclei form because of a weakened interaction between the nuclear lamina and the inner nuclear membrane due to the mutant Lamin B1. In detail, during neuronal migration, the nuclear lamina follows the nucleokinesis and is pulled forward by the microtubule's cytoskeleton, but it loses connection with the trailing edge of the nucleus because of Lmnb1CS. Consequently, chromatin would not be trapped in the nuclear lamina meshwork and would eventually escape through the honeycomb-like pores remaining in the trailing edge, uncoupling it from the nuclear lamina. However, it is also possible that other mechanisms are involved in this event. For example, the chromatin could not be affixed anymore to the nuclear lamina due to mutant Lamin B1. We should keep in mind that Lamin B1 connects with chromatin and binds LBR which, in turn, tethers heterochromatin to the inner nuclear membrane [50,51].
All this strongly suggests that Lamin B1 is essential in retaining chromatin bounding to the nuclear lamina. Indeed, it has been shown that Lamin B1 is important for TAD-TAD interaction in a mESC model of TKO for LMNA, LMNB1 and LMNB2 and that, more specifically, Lamin B1 depletion causes LAD detachment from nuclear lamina with following impact on chromatin redistribution and, thus, chromatin dynamics in MDB-MB-231 breast cancer cells [52,53]. Intuitively, this can affect the three-dimensional organisation of chromatin which strongly correlates with the gene expression program as discussed above. This can be evidenced by the fact that Lmnb1CS/CS mice have severe neurodevelopmental abnormalities with the formation of a flattened cranium and reduced size of the brain [49]. In addition, the presence of a functional Lamin B1 is required to ensure genome integrity and cell viability during neuronal migrations for the development of both the cerebral cortex and retina [54,55].
7. Conclusions
Development is a complex event, with many aspects of cell regulation involved. For instance, changes in cell microenvironment can result in modification of the cell's phenotype contributing to the determination of cell fate. Nevertheless, the manual of cell differentiation is written on the DNA and thus, the plasticity of cells depends on its regulation. DNA regulation is, in fact, an intricate and intriguing event in a cell’s life. Epigenetic factors, cis-acting elements (insulators), non-coding RNAs, and DNA compaction, are important aspects of DNA regulation. Not least, the 3D organisation of chromatin and the positioning of the genes seem to be involved in the regulation as well (Figure 1). During neuronal differentiation, loss of cell stemness is usually associated with the repositioning of key pluripotency-related genes, to repressed chromatin. Differentiation genes are, instead, characterized by loosened chromatin and are relocated to the active chromatin in the transcriptional factories [56]. Finally, the existence of TADs and chromosomes’ territories, whose positions inside the nucleus could change based on cell type, shows how the DNA 3D organisation would be a common mechanism of DNA regulation that is not restricted to cell differentiation and development. In this regard, the nuclear lamina plays a key role given its ability to bind heterochromatin, which allows it to act as a regulator or stabilizer of DNA organisation inside the nucleus. As a result, the alteration of lamins induces several neuronal defects, such as migration problems and nuclear aberrations that lead to dysfunctions in the nervous system. Additional data is needed to confirm in other contexts of cell differentiation and refine this scenario of chromatin architecture and nuclear lamina collaboration in cell differentiation. This can provide insights into the process of cell differentiation and more widely of DNA regulation.
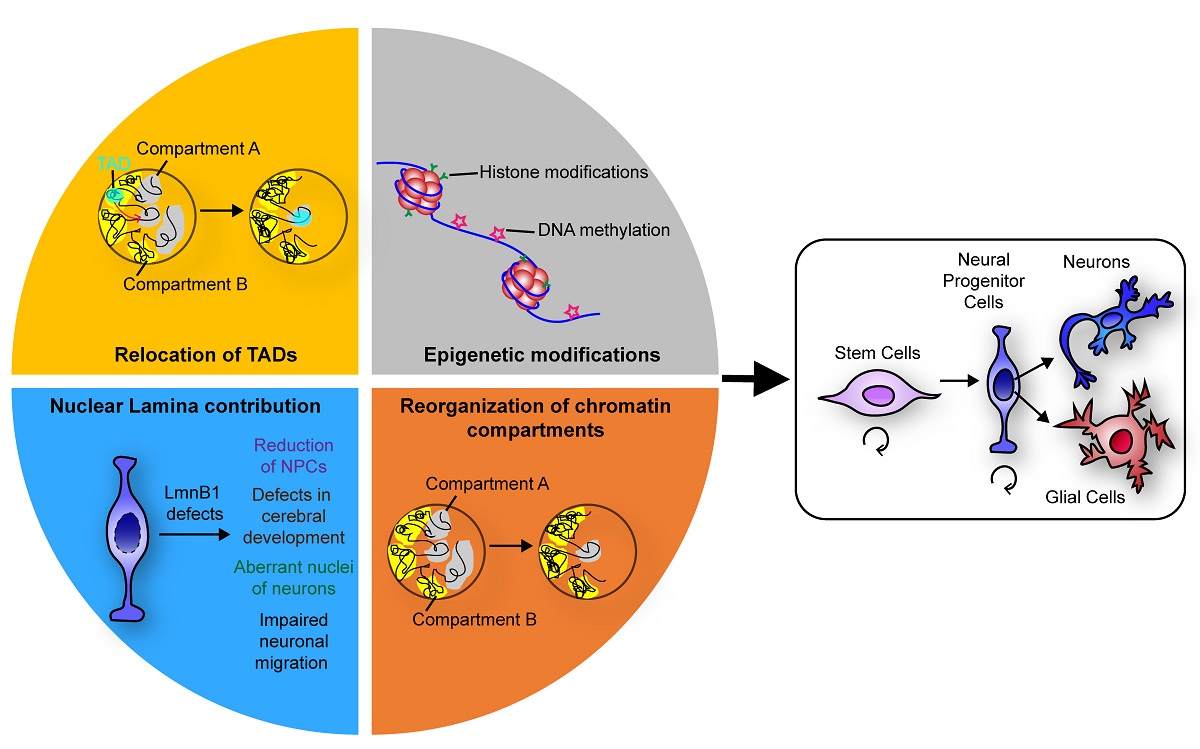
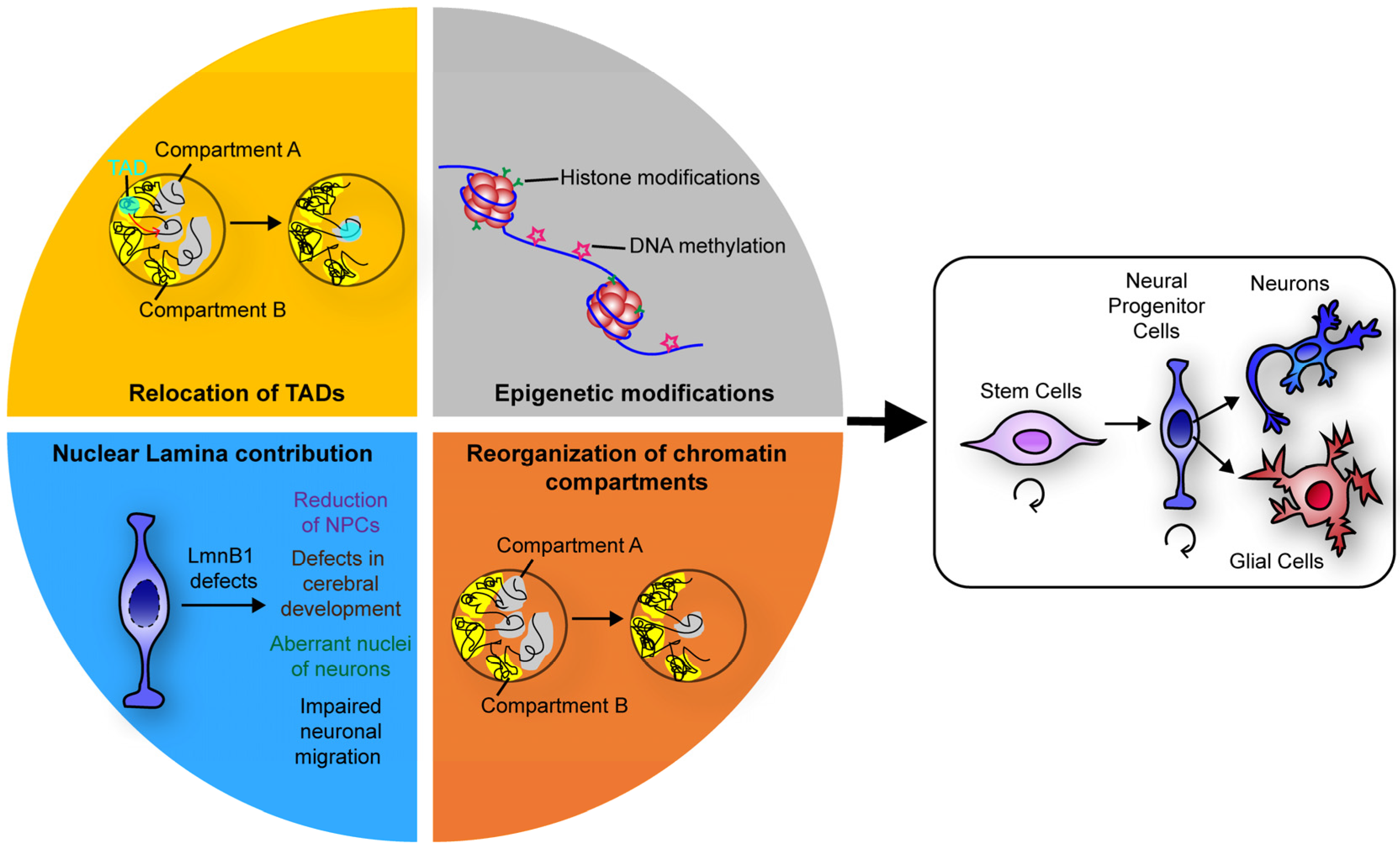
Figure 1.
Schematics summarising the events that, by regulating chromatin three-dimensional organisation, ensure a faithful neuronal differentiation.
Figure 1.
Schematics summarising the events that, by regulating chromatin three-dimensional organisation, ensure a faithful neuronal differentiation.
Author Contributions
Conceptualization, V.B. and P.S.C.; writing—original draft preparation, S.M.; writing—review and editing, V.B., P.S.C. and S.M.; supervision, V.B.. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was partly founded by University of Palermo (Italy), grant number: FFR-2020 (to V.B.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Prof. Di Leonardo (University of Palermo) for the critical reading of the manuscript and the helpful comments, and Dr. Maha Said (Université Sorbonne Paris Nord) for proofreading the manuscript and the useful suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carollo, P.S.; Barra, V. Chromatin Epigenetics and Nuclear Lamina Keep the Nucleus in Shape: Examples from Natural and Accelerated Aging. Biol. Cell 2023, 115, 2200023. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, K.; Ide, S.; Babokhov, M. Dynamic Chromatin Organization without the 30-Nm Fiber. Curr. Opin. Cell Biol. 2019, 58, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Cubeñas-Potts, C.; Corces, V.G. Architectural Proteins, Transcription, and the Three-Dimensional Organization of the Genome. FEBS Lett. 2015, 589, 2923–2930. [Google Scholar] [CrossRef] [PubMed]
- Nora, E.P.; Lajoie, B.R.; Schulz, E.G.; Giorgetti, L.; Okamoto, I.; Servant, N.; Piolot, T.; van Berkum, N.L.; Meisig, J.; Sedat, J.; et al. Spatial Partitioning of the Regulatory Landscape of the X-Inactivation Centre. Nature 2012, 485, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological Domains in Mammalian Genomes Identified by Analysis of Chromatin Interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef]
- Dixon, J.R.; Gorkin, D.U.; Ren, B. Chromatin Domains: The Unit of Chromosome Organization. Mol. Cell 2016, 62, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.M.; Foeger, N.; Kill, I.R.; Herrmann, H. The Nuclear Lamina. FEBS J. 2007, 274, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.; Cremer, M.; Dietzel, S.; Müller, S.; Solovei, I.; Fakan, S. Chromosome Territories – a Functional Nuclear Landscape. Curr. Opin. Cell Biol. 2006, 18, 307–316. [Google Scholar] [CrossRef]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive Mapping of Long Range Interactions Reveals Folding Principles of the Human Genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef]
- Barra, V.; Chiavetta, R.F.; Titoli, S.; Provenzano, I.M.; Carollo, P.S.; Di Leonardo, A. Specific Irreversible Cell-Cycle Arrest and Depletion of Cancer Cells Obtained by Combining Curcumin and the Flavonoids Quercetin and Fisetin. Genes 2022, 13, 1125. [Google Scholar] [CrossRef]
- Guelen, L.; Pagie, L.; Brasset, E.; Meuleman, W.; Faza, M.B.; Talhout, W.; Eussen, B.H.; de Klein, A.; Wessels, L.; de Laat, W.; et al. Domain Organization of Human Chromosomes Revealed by Mapping of Nuclear Lamina Interactions. Nature 2008, 453, 948–951. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Ruthenburg, A.J.; Allis, C.D.; Wysocka, J. Methylation of Lysine 4 on Histone H3: Intricacy of Writing and Reading a Single Epigenetic Mark. Mol. Cell 2007, 25, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, X.G.; Barra, V. Losing DNA Methylation at Repetitive Elements and Breaking Bad. Epigenetics Chromatin 2021, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.; Lammerding, J. The Driving Force: Nuclear Mechanotransduction in Cellular Function, Fate, and Disease. Annu. Rev. Biomed. Eng. 2019, 21, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Crisp, M.; Liu, Q.; Roux, K.; Rattner, J.B.; Shanahan, C.; Burke, B.; Stahl, P.D.; Hodzic, D. Coupling of the Nucleus and Cytoplasm: Role of the LINC Complex. J. Cell Biol. 2006, 172, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Starr, D.A.; Han, M. Role of ANC-1 in Tethering Nuclei to the Actin Cytoskeleton. Science 2002, 298, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Skepper, J.N.; Yang, F.; Davies, J.D.; Hegyi, L.; Roberts, R.G.; Weissberg, P.L.; Ellis, J.A.; Shanahan, C.M. Nesprins: A Novel Family of Spectrin-Repeat-Containing Proteins That Localize to the Nuclear Membrane in Multiple Tissues. J. Cell Sci. 2001, 114, 4485–4498. [Google Scholar] [CrossRef]
- Lygerou, Z.; Christophides, G.; Séraphin, B. A Novel Genetic Screen for SnRNP Assembly Factors in Yeast Identifies a Conserved Protein, Sad1p, Also Required for Pre-MRNA Splicing. Mol. Cell. Biol. 1999, 19, 2008–2020. [Google Scholar] [CrossRef]
- Malone, C.J.; Fixsen, W.D.; Horvitz, H.R.; Han, M. UNC-84 Localizes to the Nuclear Envelope and Is Required for Nuclear Migration and Anchoring during C. Elegans Development. Dev. Camb. Engl. 1999, 126, 3171–3181. [Google Scholar] [CrossRef]
- Guarda, A.; Bolognese, F.; Bonapace, I.M.; Badaracco, G. Interaction between the Inner Nuclear Membrane Lamin B Receptor and the Heterochromatic Methyl Binding Protein, MeCP2. Exp. Cell Res. 2009, 315, 1895–1903. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Callebaut, I.; Pezhman, A.; Courvalin, J.-C.; Worman, H.J. Domain-Specific Interactions of Human HP1-Type Chromodomain Proteins and Inner Nuclear Membrane Protein LBR *. J. Biol. Chem. 1997, 272, 14983–14989. [Google Scholar] [CrossRef] [PubMed]
- Makhija, E.; Jokhun, D.S.; Shivashankar, G.V. Nuclear Deformability and Telomere Dynamics Are Regulated by Cell Geometric Constraints. Proc. Natl. Acad. Sci. 2016, 113, E32–E40. [Google Scholar] [CrossRef] [PubMed]
- Alisafaei, F.; Jokhun, D.S.; Shivashankar, G.V.; Shenoy, V.B. Regulation of Nuclear Architecture, Mechanics, and Nucleocytoplasmic Shuttling of Epigenetic Factors by Cell Geometric Constraints. Proc. Natl. Acad. Sci. 2019, 116, 13200–13209. [Google Scholar] [CrossRef]
- Jain, N.; Iyer, K.V.; Kumar, A.; Shivashankar, G.V. Cell Geometric Constraints Induce Modular Gene-Expression Patterns via Redistribution of HDAC3 Regulated by Actomyosin Contractility. Proc. Natl. Acad. Sci. 2013, 110, 11349–11354. [Google Scholar] [CrossRef] [PubMed]
- Stiles, J.; Jernigan, T.L. The Basics of Brain Development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Christian, K.M.; He, C.; Jin, P.; Ming, G.-L.; Song, H. Epigenetic Mechanisms in Neurogenesis. Nat. Rev. Neurosci. 2016, 17, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Nothof, S.A.; Magdinier, F.; Van-Gils, J. Chromatin Structure and Dynamics: Focus on Neuronal Differentiation and Pathological Implication. Genes 2022, 13, 639. [Google Scholar] [CrossRef]
- Golob, J.L.; Paige, S.L.; Muskheli, V.; Pabon, L.; Murry, C.E. Chromatin Remodeling during Mouse and Human Embryonic Stem Cell Differentiation. Dev. Dyn. 2008, 237, 1389–1398. [Google Scholar] [CrossRef]
- Fujita, Y.; Yamashita, T. Spatial Organization of Genome Architecture in Neuronal Development and Disease. Neurochem. Int. 2018, 119, 49–56. [Google Scholar] [CrossRef]
- Aoto, T.; Saitoh, N.; Ichimura, T.; Niwa, H.; Nakao, M. Nuclear and Chromatin Reorganization in the MHC-Oct3/4 Locus at Developmental Phases of Embryonic Stem Cell Differentiation. Dev. Biol. 2006, 298, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Le Gros, M.A.; Clowney, E.J.; Magklara, A.; Yen, A.; Markenscoff-Papadimitriou, E.; Colquitt, B.; Myllys, M.; Kellis, M.; Lomvardas, S.; Larabell, C.A. Soft X-Ray Tomography Reveals Gradual Chromatin Compaction and Reorganization during Neurogenesis in Vivo. Cell Rep. 2016, 17, 2125–2136. [Google Scholar] [CrossRef] [PubMed]
- Meshorer, E.; Yellajoshula, D.; George, E.; Scambler, P.J.; Brown, D.T.; Misteli, T. Hyperdynamic Plasticity of Chromatin Proteins in Pluripotent Embryonic Stem Cells. Dev. Cell 2006, 10, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Billia, F.; Baskys, A.; Carlen, P.L.; De Boni, U. Rearrangement of Centromeric Satellite DNA in Hippocampal Neurons Exhibiting Long-Term Potentiation. Mol. Brain Res. 1992, 14, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Solovei, I.; Grandi, N.; Knoth, R.; Volk, B.; Cremer, T. Positional Changes of Pericentromeric Heterochromatin and Nucleoli in Postmitotic Purkinje Cells during Murine Cerebellum Development. Cytogenet. Genome Res. 2004, 105, 302–310. [Google Scholar] [CrossRef]
- Solovei, I.; Kreysing, M.; Lanctôt, C.; Kösem, S.; Peichl, L.; Cremer, T.; Guck, J.; Joffe, B. Nuclear Architecture of Rod Photoreceptor Cells Adapts to Vision in Mammalian Evolution. Cell 2009, 137, 356–368. [Google Scholar] [CrossRef]
- Clowney, E.J.; LeGros, M.A.; Mosley, C.P.; Clowney, F.G.; Markenskoff-Papadimitriou, E.C.; Myllys, M.; Barnea, G.; Larabell, C.A.; Lomvardas, S. Nuclear Aggregation of Olfactory Receptor Genes Governs Their Monogenic Expression. Cell 2012, 151, 724–737. [Google Scholar] [CrossRef]
- Kishi, Y.; Kondo, S.; Gotoh, Y. Transcriptional Activation of Mouse Major Satellite Regions during Neuronal Differentiation. Cell Struct. Funct. 2012, 37, 101–110. [Google Scholar] [CrossRef]
- Fraser, J.; Ferrai, C.; Chiariello, A.M.; Schueler, M.; Rito, T.; Laudanno, G.; Barbieri, M.; Moore, B.L.; Kraemer, D.C.A.; Aitken, S.; et al. Hierarchical Folding and Reorganization of Chromosomes Are Linked to Transcriptional Changes in Cellular Differentiation. Mol. Syst. Biol. 2015, 11, 852. [Google Scholar] [CrossRef]
- Decker, B.; Liput, M.; Abdellatif, H.; Yergeau, D.; Bae, Y.; Jornet, J.M.; Stachowiak, E.K.; Stachowiak, M.K. Global Genome Conformational Programming during Neuronal Development Is Associated with CTCF and Nuclear FGFR1—The Genome Archipelago Model. Int. J. Mol. Sci. 2020, 22, 347. [Google Scholar] [CrossRef]
- Stachowiak, M.K.; Stachowiak, E.K. Evidence-Based Theory for Integrated Genome Regulation of Ontogeny--An Unprecedented Role of Nuclear FGFR1 Signaling. J. Cell. Physiol. 2016, 231, 1199–1218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shin, J.-Y.; Nakanishi, K.; Homma, S.; Kim, G.J.; Tanji, K.; Joseph, L.C.; Morrow, J.P.; Stewart, C.L.; Dauer, W.T.; et al. Postnatal Development of Mice with Combined Genetic Depletions of Lamin A/C, Emerin and Lamina-Associated Polypeptide 1. Hum. Mol. Genet. 2019, 28, 2486–2500. [Google Scholar] [CrossRef] [PubMed]
- Lochs, S.J.A.; Kefalopoulou, S.; Kind, J. Lamina Associated Domains and Gene Regulation in Development and Cancer. Cells 2019, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Peric-Hupkes, D.; Meuleman, W.; Pagie, L.; Bruggeman, S.W.M.; Solovei, I.; Brugman, W.; Gräf, S.; Flicek, P.; Kerkhoven, R.M.; van Lohuizen, M.; et al. Molecular Maps of the Reorganization of Genome-Nuclear Lamina Interactions during Differentiation. Mol. Cell 2010, 38, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Shevelyov, Y.Y.; Ulianov, S.V. The Nuclear Lamina as an Organizer of Chromosome Architecture. Cells 2019, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, I.; Kind, J. Spatial Chromatin Organization and Gene Regulation at the Nuclear Lamina. Curr. Opin. Genet. Dev. 2019, 55, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.R.E.; Azuara, V.; Perry, P.; Sauer, S.; Dvorkina, M.; Jørgensen, H.; Roix, J.; McQueen, P.; Misteli, T.; Merkenschlager, M.; et al. Neural Induction Promotes Large-Scale Chromatin Reorganisation of the Mash1 Locus. J. Cell Sci. 2006, 119, 132–140. [Google Scholar] [CrossRef]
- Coffinier, C.; Jung, H.-J.; Nobumori, C.; Chang, S.; Tu, Y.; Barnes, R.H.; Yoshinaga, Y.; de Jong, P.J.; Vergnes, L.; Reue, K.; et al. Deficiencies in Lamin B1 and Lamin B2 Cause Neurodevelopmental Defects and Distinct Nuclear Shape Abnormalities in Neurons. Mol. Biol. Cell 2011, 22, 4683–4693. [Google Scholar] [CrossRef]
- Jung, H.-J.; Nobumori, C.; Goulbourne, C.N.; Tu, Y.; Lee, J.M.; Tatar, A.; Wu, D.; Yoshinaga, Y.; de Jong, P.J.; Coffinier, C.; et al. Farnesylation of Lamin B1 Is Important for Retention of Nuclear Chromatin during Neuronal Migration. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, E1923–1932. [Google Scholar] [CrossRef]
- Pascual-Reguant, L.; Blanco, E.; Galan, S.; Le Dily, F.; Cuartero, Y.; Serra-Bardenys, G.; Di Carlo, V.; Iturbide, A.; Cebrià-Costa, J.P.; Nonell, L.; et al. Lamin B1 Mapping Reveals the Existence of Dynamic and Functional Euchromatin Lamin B1 Domains. Nat. Commun. 2018, 9, 3420. [Google Scholar] [CrossRef]
- Lukášová, E.; Kovařík, A.; Kozubek, S. Consequences of Lamin B1 and Lamin B Receptor Downregulation in Senescence. Cells 2018, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Hu, J.; Yue, S.; Kristiani, L.; Kim, M.; Sauria, M.; Taylor, J.; Kim, Y.; Zheng, Y. Lamins Organize the Global Three-Dimensional Genome from the Nuclear Periphery. Mol. Cell 2018, 71, 802–815.e7. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Li, M.; Shao, S.; Li, C.; Ai, S.; Xue, B.; Hou, Y.; Zhang, Y.; Li, R.; Fan, X.; et al. Nuclear Peripheral Chromatin-Lamin B1 Interaction Is Required for Global Integrity of Chromatin Architecture and Dynamics in Human Cells. Protein Cell 2022, 13, 258–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.Y.; Yang, Y.; Weston, T.A.; Belling, J.N.; Heizer, P.; Tu, Y.; Kim, P.; Edillo, L.; Jonas, S.J.; Weiss, P.S.; et al. An Absence of Lamin B1 in Migrating Neurons Causes Nuclear Membrane Ruptures and Cell Death. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 25870–25879. [Google Scholar] [CrossRef]
- Razafsky, D.; Ward, C.; Potter, C.; Zhu, W.; Xue, Y.; Kefalov, V.J.; Fong, L.G.; Young, S.G.; Hodzic, D. Lamin B1 and Lamin B2 Are Long-Lived Proteins with Distinct Functions in Retinal Development. Mol. Biol. Cell 2016, 27, 1928–1937. [Google Scholar] [CrossRef]
- Papantonis, A.; Cook, P.R. Transcription Factories: Genome Organization and Gene Regulation. Chem. Rev. 2013, 113, 8683–8705. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



