Submitted:
14 April 2023
Posted:
24 April 2023
You are already at the latest version
Abstract
As two commonly-used non-selective herbicides, glyphosate (GP) and diquat (DQ) are easily co-resided in lateritic paddy soil due to the rich iron/aluminum oxides, nevertheless there is limited information on their co-impact on microbial diversity and community structure in this type soil. In this study, the short-term effects of combined GP and DQ on soil microbial diversity and community structure shifts were investigated in lateritic paddy soil from a tropical agricultural region (Hainan, China) based on 16S rRNA and ITS high-throughput sequencing technology. The results showed that mixed herbicides promoted the abundance of Streptomyces in bacteria (0.45-1.84%) and Curvularia in fungi (0.01-5.85%), while GP and DQ had inhibitory effects on the abundance of Streptomyces (0.13-2.21%) and Curvularia (0.03-1.13%), which were significantly different with their single exposure (p < 0.05); the combined application of the two herbicides aggravate the adverse effect on the diversity of soil fungal community (p < 0.05), although their mixture did not have a greater impact on the soil bacteria abundance/diversity and fungi abundance (p > 0.05). Results suggested that the combined application of GP and DQ affected the fungal diversity although they did not cause other significant negative effects on soil microorganisms, hinting that more attention should be paid to the mixed effect caused by GP and DQ on specific fungal populations in lateritic paddy soil.
Keywords:
Herbicides
; mixed toxicity
; microbial diversity
; microbial community structure
1. Introduction
Herbicides are the most widely used class of pesticides in global agriculture [1], with up to 1 million tons in 2016 [2]. Among herbicides, glyphosate (GP) and diquat (DQ), as two non-selective herbicides, both rank in the top five highest use wide in global market, especially GP owing highest use in the global market with 0.86 million tons in 2016 [3]. At present, GP was recorded with heavy pollution in different environmental media [4,5,6], e.g., water (21.2-32.5 μg/L) [7,8,9], soil (1200-1502 μg/kg) [10,11,12] and sediment (1149 μg/kg) [13], especially in tropical soil (690-40000 μg/kg) [12,14,15,16,17]. Meanwhile, lateritic paddy soil is rich in iron and aluminum, due to its strong binding with clay particles and organic matter, DQ is easily retained in the lateritic paddy soil [18]. In practice, two or more mixed herbicides (e.g., GP and DQ) are often applied in varied agricultural scenarios, especially in the tropics [6,16,19]. Undoubtedly, the GP and DQ will inevitably co-occur in lateritic paddy soil. However, there is limited information on the co-occurrence of GP and DQ on microbial diversity and community structure shifts in lateritic paddy soil.
Microbial communities in the soil environment play an important role in the Biogeochemistry cycle of many important soil elements [20,21]. It was found that herbicides residues changed the soil microflora and had a potentially long-term effect on the function of agricultural soils [22,23,24,25]. Most studies have shown that GP has no or transient stress effect on soil microbial community [26,27,28,29,30,31], but some studies have shown significant adverse effects [32,33,34]. For instance, GP can lead to a decrease in the abundance of Pseudomonas fluorescens, Mn-transforming bacteria, and indoleacetic acid-producing bacteria in soybean rhizosphere soil [35]. The abundance of certain Bacteroidetes, Chloroflexi, Cyanobacteria, Planctomycetes, and alpha-Proteobacteria members are highly negatively correlated with GP concentrations [36]. And the decrease in richness and diversity in the presence of a high concentration of GP was only observed for eukaryotic communities [37]. There is also some other literature on the structure of specific microbial communities in different GP exposure cycles, including the effects of GP on Acidobacteria, Ammonia-oxidizing bacteria, Mycorrhiza, etc [38,39,40]. Due to the strong binding property of DQ with clay particles, it is reasonable that higher levels of DQ occurred in lateritic paddy soil, which might bring more pressure on the structure and function of the microbe. Nevertheless, there are few studies on the effects of combined exposure of GP with other herbicides on microbial populations [41], where the combined toxicology of GP with other chemicals on microorganisms mainly focuses on it with heavy metals or polyethylene microplastics [42,43,44,45,46,47]. GP contains coordination groups such as carboxyl, amino, and phosphate groups, and has a strong complexing ability to heavy metal cations and organic cations [44]. Similar to the charge characteristics of metal ions, DQ has the ability to strongly adsorb on the surface with negative charges and also the presence of a strong oxidation-reduction cycling ability that can be reduced to generate free radicals [18]. These similar characteristics coupled with the observed difference (i.e., strong oxidation-reduction cycling ability) might partly contribute to the combined toxic effects of GP and DQ differing from that of GP and heavy metal.
In the context, the short-term exposure (28 days) test was conducted, wherein the effects of single GP, DQ, and their mixture on the diversity and community structure of soil bacteria and fungi were investigated in lateritic paddy soil from a tropical agricultural region (Hainan, China) based on 16S rRNA and Internal Transcribed Spacer (ITS) high-throughput sequencing technology. We hope the results of the study will provide a scientific basis for the rational composite application of herbicides in lateritic paddy soil in agricultural areas.
2. Materials and Methods
2.1. Experiment Design and Sample Collection
Lateritic paddy soils (0-15 cm) were collected from a village (19°55’N, 110°25’E) in the tropical agricultural region of the Nandu River Basin in Hainan Province, China. The residues of GP and DQ were not founded in the soils (the limits of detection were < 0.03 mg kg−1) based on the analytical methods by Delhomme et al. [48] and Pizzutti et al. [49]. The texture of the soil is a typical sandy loam identified by a method from USDA [50]. In order to recover soil microorganisms, the soil samples were sieved through a 10 mesh sieve (2 mm). Then, the deionized water was added to keep the soil at 50% of the maximum water holding capacity, lasting 2 weeks for pre-incubation. Analytical grade DQ and GP (active ingredient > 98.5%) were purchased from Beijing Tanmo Quality Inspection Technology Co., Ltd. (Beijing, China). The recommended field dose of GP was 0.6 mg kg−1, and meanwhile the recommended dosage of DQ in the field was 0.4 mg kg−1 [51]. In the experiment, 10 treatments were set up based on the recommended field dose, containing a blank only with deionized water and the 9 different experimental treatments of GP, DQ, and GP+DQ at different concentrations (Table 1). During the experiment, deionized water was added regularly during the incubation process to compensate for evaporated water. Samples were collected on days 1, 7, 14 and 28 after application, and then were stored at -80 °C for further analysis. All experiments were conducted in triplicates.
2.2. DNA Extraction and Database Construction
Microbial DNA was extracted using the HiPure Soil DNA Kits (or HiPure Stool DNA Kits) (Magen, Guangzhou, China) according to the manufacturer’s protocols. The 16S rDNA target region of the ribosomal RNA gene was amplified by PCR (95 °C for 5 min, followed by 30 cycles at 95 °C for 1 min, 60 °C for 1 min, and 72 °C for 1 min and a final extension at 72 °C for 7 min) using primers listed in the Table S1. PCR reactions were performed in triplicate 50 μL mixture containing 10 μL of 5 × Q5@ Reaction Buffer, 10 μL of 5 × Q5@ High GC Enhancer, 1.5 μL of 2.5 mM dNTPs, 1.5 μL of each primer (10 μM), 0.2 μL of Q5@ High-Fidelity DNA Polymerase, and 50 ng of template DNA. Related PCR reagents were from New England Biolabs, USA.
For bacteria, after genomic DNA was extracted from the soil samples, the 16S rDNA V3 + V4 region was amplified with barcode-specific primers. The primer sequence was: 341F: CCTACGGGNGGCWGCAG; 806R: GGACTACHVGGGTATCTAAT.
For fungi, the amplified region is ITS2 region of ITS. The primer sequences were as follows: ITS3: GATGAAGAACGYAGYRAA; ITS4: TCCTCCGCTTATTGATATGC. The purified amplified products were connected to sequencing linkers to construct sequencing libraries and sequenced by Illumina.
2.3. Bioinformatic Analysis of Microbial 16S rRNA and ITS
After the raw data named “Reads” were obtained by sequencing, the low-quality data in Reads were filtered first, then the two-terminal Reads were spliced into Tag, and the Tag was filtered again. The data obtained was called Clean Tag. Next, based on Clean Tag, Usearch software was used to cluster, remove the chimeric Tag detected during the clustering process, and obtain the representative sequences and abundance of OTU. Based on the sequence and abundance data of OTU, species annotation, species composition analysis, indicator species analysis, α diversity analysis and β diversity analysis were carried out. If there was effective grouping, the differences between groups were compared and tested statistically.
2.5. Data Analysis and Statistical Analysis
Gene abundance data were analyzed using One-way Analysis of Variance (ANOVA) by using the SPSS Statistical Package (version 19.0, IBM, Armonk, New York, USA). Duncan's multi-range test and Spearman's rank statistical analysis were used to calculate the correlation between samples, bacteria and fungi and BDGs/PDGs. All data are mean ± standard error (SE) of three replicates. The data was considered to be significant when p < 0.05.
3. Results
3.1. Effects of Combined Pollution of GP and DQ on Composition and Diversity of Soil Bacterial Community
3.1.1. Bacterial Community Composition
In general, it was observed that GP decreased the relative abundance of Actinobacteria, nevertheless DQ had no significant effect on Actinobacteria. Particularly, combined pollution of GP and DQ increased the relative abundance of Actinobacteria Figure 1a. Specifically, low concentration of mixed herbicides caused the relative abundance of Actinobacteria to increase by 4.91%, 6.78% and 4.25% on the 7th, 14th and 28th day respectively; medium concentration of mixed herbicides caused the relative abundance of Actinobacteria to increase by 3.98% and 5.21% on the 14th and 28th day respectively; high concentration of mixed herbicides caused the relative abundance of Actinobacteria to increase by 4.38% and 2.37% on the 7th and 14th day, respectively. Notably, the mixed herbicides enhanced the inhibitory effect of DQ on Proteobacteria although the promotion of the abundance of Proteobacteria by the GP and the adverse effect on the abundance of Proteobacteria by the DQ was observed Figure 1b. Specifically, low concentration of mixed herbicides caused the relative abundance of Proteobacteria to decreased by 3.58% and 5.70% on the 7th and 14th day respectively; medium concentration of mixed herbicides caused the relative abundance of Proteobacteria to decreased by 4.93% and 2.76% at the 14th and 28th day respectively; high concentration of mixed herbicides caused the relative abundance of Proteobacteria to decreased by 4.97%, 2.49% and 0.92% on the 7th, 14th and 28th day, respectively. Taken together, the results indicated that combined pollution of GP and DQ could increase the relative abundance of Actinobacteria and decrease the relative abundance of Proteobacteria compared with a single herbicide.
The stacking chart of the distribution of bacterial community composition at the genus level was shown in Figure 2a–d. Overall, it could be observed that GP increased the relative abundance of Sphingomonas while DQ decreased the abundance of Sphingomonas, whereas the inhibition of mixed herbicides on Sphingomonas was stronger than that of DQ, but the inhibitory effect weakened over time. Specifically, low concentration of mixed herbicides caused the relative abundance of Sphingomonas to decrease by 4.36%, 3.39% and 1.40% on the 7th, 14th and 28th day respectively; medium concentration of mixed herbicides caused the relative abundance of Sphingomonas to decrease by 5.13%, 3.11% and 1.92% on the 7th, 14th and 28th day respectively; while the high concentration of mixed herbicides caused the relative abundance of Sphingomonas to decreased by 4.30%, 1.41% and 0.64% on the 7th, 14th and 28th day, respectively. Remarkably, the mixed herbicides increased the relative abundance of Streptomyces although the inhibiting effect on the Streptomyces by GP and DQ was observed. Specifically, low concentration of mixed herbicides caused the relative abundance of Streptomyces to increase by 0.69%, 0.86% and 0.81% on the 7th, 14th and 28th day respectively; while the medium concentration of mixed herbicides caused the relative abundance of Streptomyces to increase by 0.85%, 0.69% and 1.29% on the 7th, 14th and 28th day, respectively. In addition, it was observed that GP increased the relative abundance of Phenylobacterium, nevertheless, DQ had little effect on Phenylobacterium. Particularly, the mixed herbicides decreased the relative abundance of Phenylobacterium to some extent. On the 7th day, a low concentration of mixed herbicides caused the abundance of Phenylobacterium to decrease by 0.59%. However, the medium concentration of mixed herbicide reduced the Phenylobacterium abundance by 0.74% on day 7. The results showed that the mixed herbicides inhibited the growth of Sphingomonas and Phenylobacterium, and promoted the growth of Streptomyces at low and high concentrations as compared with a single herbicide.
3.1.2. Alpha Diversity of Bacterial Community
The results of bacterial richness and diversity analysis were shown in Table 2. Generally, the Chao1 index was used to evaluate microbial richness, while the Shannon index was used to evaluate microbial diversity. The larger Chao1 indices is, the higher the microbial richness will be. And the larger the Shannon index is, the higher the microbial diversity will be. On day 1 and day 28, it was observed that the medium concentration of GP increased bacterial richness (risen by 413.413/146.912) while the same concentration of DQ inhibited bacterial richness (reduced by 195.023/48.648). Particularly, the medium concentration of mixed herbicides promoted bacterial richness (risen by 300.247/63.717). It is worth noting that medium concentrations of GP and DQ promoted bacterial diversity on days 7 and 14, but by day 28, the impact of single and mixed herbicides on soil bacterial diversity was not significantly different. In general, the effects of mixed herbicides on soil bacterial richness and diversity were not significantly different from those of a single herbicide.
3.1.3. Beta Diversity of Bacterial Community
The principal coordinate analysis based on the Bray-curtis distance for the bacterial communities at the phylum level and genus level was shown in Figure 3a,b and Figure S1. The results showed that the difference in community structure between the control and the composite herbicide was less than that between the control and the single herbicide (Figure S1). In addition, on day 1 (Figure 3a) and day 14 (Figure 3b), the difference in community structure between the mixed herbicide treatment and the control treatment was less than that between the the single herbicide treatment and the control treatment (R2=0.723, p=0.001; R2=0.405, p=0.044). From the above analysis, it can be concluded that the effect of mixed herbicides on soil bacterial community structure was less than that of single herbicides at low concentrations, although there was no significant difference in the effects between mixed herbicides and single herbicides on soil bacterial community structure at medium or high concentrations.
3.1.4. LEfse Analysis of Bacterial Community
Linear discriminant analysis (LDA ≥ 2) of bacterial community was shown in Figure 4a, Figure S2a–h. It can be observed that at the phylum level, Cyanobacteria were the significantly different biomarker taxa between GP treatment and other herbicide treatments on day 1 (p ≤ 0.038). On the 14th day, the significantly different biomarker taxa between GP treatment and other herbicide treatments became Bacteroides (p ≤ 0.040) instead of Cyanobacteria. It is worth noting that Chloroflex is sensitive to various herbicides treatments on day 14. Meanwhile, at the genus level, Streptomyces showed a significant difference between mixed herbicide treatments and other herbicides treatments on day 14 (p ≤ 0.034). However, on day 28, the significantly different biomarker taxa between GP treatment and other herbicide treatments became Rickettsia (p ≤ 0.013) rather than Streptomyces. In particular, Streptomyces was sensitive to mixed herbicide responses on day 14, while Rickettsia was sensitive to GP responses on days 14 and 28. From the above analysis, it can be concluded that the relative abundance of Cyanobacteria, Bacteroides, and Rickettsia is suppressed after adding DQ to the GP. It is worth noting that compared to the single herbicide, mixed herbicides significantly increased the abundance of Streptomyces (p < 0.05).
3.2. Effects of Combined Pollution of GP and DQ on Composition and Diversity of Soil Fungal Community
3.2.1. Fungal Community Composition
The stacking chart of the species distribution of fungi at the phylum level is shown in Figure 5. Notably, at the phylum level, the low concentration of mixed herbicides inhibited the abundances of Basidomycota but promoted the abundances of Ascomycota at the 7th and 28th day compared with single herbicide. Specifically, on day 7, low concentration mixed herbicide reduced the abundance of Basidomycota by 3.3%, while increased Ascomycota abundance by 4.86%; in addition, the abundances of Basidomycota decreased by 3.21% and Ascomycota increased by 2.33% at day 14. Taken together, the results showed that Basidomycota and Ascomycota were sensitive to exposure to mixed herbicides at certain exposure times, especially on days 7 and 14.
Figure 2e–h shows a stacked distribution of fungi at the genus level. In general, it was observed that GP inhibited the abundance of Talaromyces at low and middle concentrations on the 7th and 28th day, nevertheless the same concentrations of DQ promoted the abundance of Talaromyces at the same time. Notably, the addition of GP enhanced the effect of DQ on the abundance of Talaromyces. Specifically, low concentration mixed herbicides increased the abundance of Talaromyces by 1.16% and 1.43% respectively on the 7th and 28th day; while the medium concentration of mixed herbicides increased the abundance of Talaromyces by 4.91% and 2.95% respectively on the 7th and 28th day. However, on the 7th and 14th day, the mixed herbicides promoted the abundance of Curvularia although the single herbicide inhibited the abundance of Curvularia. Specifically, the low concentration of mixed herbicides increased the abundance of Curvularia by 2.60% on the 7th and 14th day; the medium concentration of mixed herbicides increased the abundance of Curvularia by 5.85% and 0.86% on the 7th and 14th day, respectively; while the medium concentration of mixed herbicides increased the abundance of Curvularia by 0.30% and 0.97% on the 7th and 14th day, respectively. Particularly, on the 7th, 14th and 28th days, the mixed herbicides inhibited the abundance of Conlarium significantly compared with single herbicides at low and medium concentrations, and the inhibitory effect became more significant over time. Specifically, the low concentration mixed herbicides respectively reduced the abundance of Conlarium by 0.42%, 1.32% and 5.01% on the 7th, 14th and 28th day, while the medium concentration of mixed herbicides reduced the abundance of Conlarium by 1.26%, 1.60% and 2.97% at the 7th, 14th and 28th day, respectively. The results showed that, compared with single herbicides, the mixed herbicides promoted the abundance of Talaromyces and Curvularia at low and medium concentrations, but inhibited the abundance of Conlarium.
3.2.2. Alpha Diversity of Fungal Community
The fungal richness and diversity data were presented in Table 3. It is observed that the diversity of fungal communities increased with time, but the richness decreased over time. On the first day, the fungal diversity of low and medium concentration GP treatments was significantly higher than that of mixed herbicide treatments at the same concentration (p < 0.05). Particularly, on the 14th day, low concentrations of single herbicides had little impact on fungal diversity, while low concentrations of mixed herbicides significantly inhibited fungal diversity (p < 0.05). It could be found that the high concentration of mixed herbicides has little impact on fungal diversity, nervertheless at the same concentration the fungal diversity of GP treatment is significantly lower than that of the control treatment (p < 0.05), while the fungal diversity of DQ treatment is significantly higher than that of control treatment (p < 0.05). It is worth noting that on day 28, high concentrations of mixed herbicides significantly inhibited fungal diversity (p < 0.05), while single herbicides at the same concentration had no significant impact on fungal diversity. It can be observed that compared to the single herbicides, mixed herbicides inhibit the diversity of fungal communities in soil, but have no significant impacts on the richness of fungal.
3.2.3. Beta Diversity of Fungal Community
The principal coordinate analysis based on the Bray-curtis distance for the fungal communities at the genus level was shown in Figure 6a–d. It can be observed that the differences in fungal community structure among different treatments increase over time (R2 ≥ 0.847, p=0.001). It is worth noting that under low and high concentrations, the difference in community structure between the control treatment and the mixed herbicide treatment is smaller than the difference between the single herbicide treatment and the control (R2 ≥ 0.534, p=0.001). From the above analysis, it can be concluded that the impact of mixed herbicides on the structure of soil fungal communities is less than that of single herbicides.
3.2.4. LEfSe Analysis of Fungal Community
Linear discriminant analysis (LDA ≥ 2) of fungi was shown in Figure 4b, Figure S3. In particular, at the phylum level, Ascomycota was extremely sensitive to the response of each herbicide treatment at day 28. It is worth noting that at the genus level, Thielavia had a sensitive response to GP on day 28, for Thielavia was a significantly different species between GP treatment and other herbicides treatments. It can be concluded that GP significantly promoted the abundance of Thielavia, while there was no definite significant difference in species from mixed herbicide treatments when compared with the single herbicide treatments.
4. Discussion
Pesticide use has harmful impact on soil biological activity, such as microbial abundance, diversity, and activities, all of which influence nutrient transformation and therefore, health and quality of soil [52]. This study found that the effect of combined pollution of GP and DQ on bacterial community structure at low concentrations was less than that of single herbicides. However, that was no difference at medium and high concentration between mixed herbicide and single herbicides. In addition, the effect of mixed herbicide on fungal community structure was less than that of single herbicide treatment. Most reports showed that herbicides have little or no short-term effect on microbial community structure [29,30,31], Previous studies have revealed that the microbial communities were negatively affected by GP [53] and the activity of total microbial community was also affected by GP [54]. In contrast, the ester-linked fatty acid methyl ester extraction (EL-FAME) analysis of agricultural soil exposed to repetitive application of GP displayed no significant changes in structure of the soil microbial community [55]. In this study, the short-term effect of combined pollution of GP and DQ on microbial community structure was less than single herbicide. Further researches will be needed on the long-term effects of combined pollution of GP and DQ on soil microbial community structure.
The present study found that, compared with a single herbicide, the combined pollution of GP and DQ has a certain promoting effect on Actinobacteria and a certain inhibiting effect on Proteobacteria at phylum level. It is well known that Actinobacteria and Proteobacteria are common bacteria taxa in soil [56], which may be sensitive to herbicide contamination, these taxa can have a variety of effects on soil and vegetation health, including beneficial and pathogenic effects [57,58]. At the genus level, the single herbicide decreased the relative abundance of Streptomyces. However, the combined pollution of GP and DQ has a certain promoting effect on Streptomyces. It was found that the combination of GP and Cu could reduce the toxicity of heavy metals to photoluminescent bacteria [44], it may be that DQ and Cu Combine with GP in a similar way to reduce the stress of some bacteria. Although GP promoted the abundance of Sphingomonas, however, the addition of GP enhanced the inhibitory effect of DQ on the abundance of Sphingomonas. In addition, single herbicides promoted or had no effect on the abundance of Phenylobacterium, while mixed herbicides had an inhibitory effect on the abundance of Phenylobacterium. Some researchers have suggested that the combined use of GP and Cd may aggravate the effects on E. coli [42], perhaps Phenylobacterium is as sensitive as E. coli to the stress of GP combined contamination. For fungi, although GP inhibited the abundance of Talaromyces at low and medium concentrations, the addition of GP enhanced the promotion of DQ on the abundance of Talaromyces. The single herbicide inhibited the abundance of Curvularia, while the mixed herbicides promoted the abundance of Curvularia. In addition, the abundance of Conlarium was significantly inhibited by mixed herbicides compared with single herbicides. Some studies have shown that fungal diversity and abundance respond strongly to high concentrations of GP, and herbicide combined pollution may make the stress response of different fungi more obvious.
The effects of combined pollution of GP and DQ on bacterial community richness and diversity were not significantly different from those of single herbicides. It was found that a single herbicide had a transient promoting or inhibiting effect on bacterial population abundance and community diversity in soil. Previous studies had also found that GP has an adverse effect on the interactions of manganese redox bacteria, Pseudomonas fluorescens, acetogenic rhizosphere bacteria and Fusarium in the rhizosphere soil of soybean, resulting in an increase in the number of Fusarium species, the abundances of Pseudomonas fluorescens, manganese redox bacteria and acetogenic rhizosphere bacteria decreased [35]. However, the combined application of the two herbicides did not affect the richness and diversity of soil bacteria. This may be because some bacteria produce free radical scavenging molecules when they coexist with GP, and eliminate the Free Radical produced by DQ, which makes the stress response of bacteria to GP and DQ reduced or unchanged [59]. Compared with single herbicide, mixed herbicide had no significant effect on the richness of soil fungal community, but could inhibit the diversity of the fungal community. Some scholars had shown that GP can stimulate soil fungal biomass in the early and short-term, and it has an adverse effect on both fungal community diversity and species richness after long-term application of GP [60]. Thus, mixed herbicides aggravated the adverse effects on soil fungal community diversity, but the cause of the short period of this study, the effects of various herbicides on fungal population abundance were not significant.
Along with the persistence, concentration, toxicity, and bioavail-ability of the sprayed pesticide, variety of environmental factors influence the toxic impact of pesticides on microbial diversity [61,62]. Here, we focused on the relative abundances and diversity of soil microbial community diversity only at the concentration of herbicides. Future work should focus on the effects of combined pollution of GP and DQ in different soil environmental on microbial community composition and diversity
5. Conclusions
This study examined the effects of combined pollution of GP and DQ on composition and diversity of microbial community in lateritic paddy soil at the relative field application doses. Compared with single herbicide, the mixed herbicide (GP+DQ) had no significant impact on the richness of bacteria and fungal communities, but inhibited the diversity of fungal communities in the soil. Actinobacteria and Proteobacteria were the most sensitive microbial phylum to application of mixed herbicide. Which could increase the abundance of Actinobacteria but significanltly inhibit the abundance of Proteobacteria at phylum level, especially at low and high concentrations; and the mixed herbicide could enhanced the abundance of Streptomyces while decrease the abundance of Sphingomonas and Phenylobacterium at genus level. In general, the combination of GP and DQ had no more adverse effect on soil microorganisms, therefore, the two herbicides can be reasonably applied in the actual field work.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Principal coordinate analysis for the bacterial communities at the phylum level; Figure S2: Cladograms of line discriminant analysis effect size (LEfSe) analyses of bacteria; Figure S3: Cladograms of line discriminant analysis effect size (LEfSe) analyses of fungus; Table S1: Primer information.
Author Contributions
Conceptualization, methodology, writing—original draft preparation, X.Y.H.; investigation, funding acquisition, C.Y.W.; writing—review and editing, funding acquisition, H.D.T.; writing—review and editing, funding acquisition, X.D.; resources, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research were funded by the National Natural Science Foundation of China, grant number “42177402”, the Natural Science Foundation of Hainan Province, grant number “420QN316”and the Hainan Province Science and Technology Special Fund of China “ZDYF2021XDNY137”.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage. US EPA, Washington, DC, 2011.
- Yang, Y.J.; Zhang, B. Market forecast of China's pesticide industry in 2020. The world pesticide 2020, 42, 1–8. (in Chinese). [Google Scholar]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, M.T.; Poulsen, H.D.; Katholm, C.L.; Højberg, O. Review: Feed residues of glyphosate – potential consequences for livestock health and productivity. Animal 2021, 15, 100026. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, G.; Atreya, K.; Scheepers, P.T.J.; Geissen, V. Concentration and distribution of pesticide residues in soil: Non-dietary human health risk assessment. Chemosphere 2020, 253, 126594. [Google Scholar] [CrossRef] [PubMed]
- Horth, H.; Blackmore, K. Survey of glyphosate and AMPA in groundwaters and surface waters in Europe. WRC report, 2009, no. UC8073, 2.
- Demonte, L.D.; Michlg,N. ; Gaggiotti, M. Determination of glyphosate, AMPA and glufosinate in dairy farm water from Argentina using a simplified UHPLC-MS/MS method. Sci. Total Environ. 2018, 645, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Jiang, L.; Zhang, D. Glyphosate, aminomethyl-phosphonic acid, and glufosinate ammonium in agricultural groundwater and surface water in China from 2017 to 2018: Occurrence, main drivers, and environmental risk assessment. Sci. Total Environ. 2021, 769, 144396. [Google Scholar] [CrossRef]
- Aparicio, V.C.; De Gerónimo, E.; Marino, D.; Primost, J.; Carriquiriborde, P.; Costa, J.L. Environmental fate of glyphosate and aminomethylphosphonic acid in surface waters and soil of agricultural basins. Chemosphere 2013, 93, 1866–1873. [Google Scholar] [CrossRef]
- Jing, X.; Zhang, W.; Xie, J. Monitoring and risk assessment of pesticide residue in plant soil-groundwater system about medlar planting in Golmud. Environ. Sci. Pollut. R. 2021, 28, 26413–26426. [Google Scholar] [CrossRef]
- Zhang, P.; Rosee, M.; Van Zwieten, L. Direct determination of glyphosate and its metabolite AMPA in soil using mixed-mode solid-phase purification and LC-MS/MS determination on a hypercarb column. J AOAC Int. 2019, 102, 952–965. [Google Scholar] [CrossRef]
- Shen, L.Y.; Peng, Z.R.; He, W.H.; Feng, M.J.; Dai, X.L. Assessment of glyphosate residues and ecological risk in P. rosii cultured ponds. Journal of Shanghai Ocean University 2021, 5, 821–827. [Google Scholar]
- Botero-coy, A M. ; Ibáñez, M,; Sancho, J.V.; Hernandez, F. Improvements in the analytical methodology for the residue determination of the herbicide glyphosate in soils by liquid chromatography coupled to mass spectrometry. J. Chromatogr. A 2013, 1292, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Primost, J.E.; Marino, D.J.; Aparicio, V.C.; Costa, J.L; Carriquiriborde, P. Glyphosate and AMPA, "pseudo-persistent" pollutants under real-world agricultural management practices in the mesopotamic pampas agroecosystem, Argentina. Environ. Pollut. 2017, 229, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Gunarathna, S.; Gunawardana, B.; Jayaweera, M. Glyphosate and AMPA of agricultural soil, surface water, groundwater and sediments in areas prevalent with chronic kidney disease of unknown etiology, Sri Lanka. J. Environ. Sci. Heal. B 2018, 53, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Karasali, H.; Pavlidis, G. Marousopoulou A. Investigation of the presence of glyphosate and its major metabolite AMPA in Greek soils. Environ. Sci. Pollut. R. 2019, 26, 36308–36321. [Google Scholar] [CrossRef] [PubMed]
- Roede, J.R.; Miller, G.W. Diquat. Encyclopedia of Toxicology, 2014, 202–204.
- Silva, V.; Mol, H. G. J.; Zomer, P.; Tienstra, M.; Ritsema, C. J.; Geissen, V. Pesticide residues in European agricultural soils – A hidden reality unfolded. Sci. Total Environ. 2018, 653, 1532–1545. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Duke, S.O.; Lydon, J.; Koskinen, W.C.; Moorman, T.B.; Chaney, R.L.; Hammerschmidt, R. Glyphosate effects on plant mineral nutrition, crop rhizosphere microbiota, and plant disease in glyphosate-resistant crops. J. Agric. Food Chem. 2012, 60, 10375–10397. [Google Scholar] [CrossRef]
- Nguyen, D.B.; Rose, M.T.; Rose, T.J.; Morris, S.G.; Zwieten, L.V. Impact of glyphosate on soil microbial biomass and respiration: A meta-analysis. Soil Biol. Biochem. 2016, 92, 50–57. [Google Scholar] [CrossRef]
- Padilla, J.T.; Selim, H.M. Environmental behavior of Glyphosate in soils. Adv. Agron. 2020, 159, 1–34. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; Finckh, M.R.; He, M.; Ritsema, C.J.; Harkes, P.; Knuth, D.; Geissen, V. Indirect effects of the herbicide glyphosate on plant, animal and human health through its effects on microbial communities. Front. Environ. Sci. 2021, 9, 763917. [Google Scholar] [CrossRef]
- Weaver, M.A.; Krutz, L.J.; Zablotowicz, R.M.; Reddy, K.N. Effects of glyphosate on soil microbial communities and its mineralization in a Mississippi soil. Pest Manag. Sci. 2007, 63, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Schmidt, D.J.E.; Yarwood, S.A.; Cavigelli, M.A.; Reddy, K.N.; Duke, S.O.; Bradley, C.A.; Williams, M.M., Jr.; Buyer, J.S.; Maul, J.E. Soil microbial communities in diverse agroecosystems exposed to the herbicide glyphosate. Appl. Environ. Microbiol. 2020, 86, e01744–e19. [Google Scholar] [CrossRef] [PubMed]
- Lupwayi, N.Z.; Fernandez, M.R.; Kanashiro, D.A.; Petri, R.M. Profiles of wheat rhizobacterial communities in response to repeated glyphosate applications, crop rotation, and tillage. Can. J. Soil Sci. 2021, 101, 157–167. [Google Scholar] [CrossRef]
- Accinelli, C.; Koskinen, W.C.; Seebinger, J.D.; Vicari, A.; Sadowsky, M.J. Effects of incorporated corn residues on Glyphosate mineralization and sorption in soil. J. Agric. Food Chem. 2005, 53, 4110–4117. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Ferreras, L.; Lovotti, L.; Fernández, E. Impact of Glyphosate application on microbial biomass and metabolic activity in a Vertic Argiudoll from Argentina. Eur. J. Soil Biol. 2009, 45, 163–167. [Google Scholar] [CrossRef]
- Accinelli, C.; Koskinen, W.C.; Becker, J.M.; Sadowsky, M.J. . Environmental fate of two sulfonamide antimicrobial agents in soil. J. Agric. Food Chem. 2007, 55, 2677–2682. [Google Scholar] [CrossRef]
- Lancaster, S.H.; Hollister, E.B.; Senseman, S.A.; Gentry, T.J. Effects of repeated glyphosate applications on soil microbial community composition and the mineralization of glyphosate. Pest Manag. Sci. 2010, 66, 59–64. [Google Scholar] [CrossRef]
- Wilkes, T.I.; Warner, D.J.; Davies, K.G.; Edmonds-Brown, V. Tillage, glyphosate and beneficial arbuscular mycorrhizal fungi: Optimising crop management for plantfungal symbiosis. Agric. For. 2020, 10, 520. [Google Scholar] [CrossRef]
- Ch'avez-Ortiz, P.; Tapia-Torres, Y.; Larsen, J.; García-Oliva, F. Glyphosate-based herbicides alter soil carbon and phosphorus dynamics and microbial activity. Appl. Soil Ecol. 2022, 169, 104256. [Google Scholar] [CrossRef]
- Zobiole, L.; Kremer, R.; Oliveira, R.; Constantin, J. Glyphosate affects microorganisms in rhizospheres of glyphosate-resistant soybeans. J. Appl. Microbiol. 2011, 110, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Carles, L.; Artigas, J. Interaction between glyphosate and dissolved phosphorus on bacterial and eukaryotic communities from river biofilms. Sci. Total Environ. 2020, 719, 137463. [Google Scholar] [CrossRef] [PubMed]
- Annett, R.; Habibi, H.R.; Hontela, A. Impact of glyphosate and glyphosate-based her-bicides on the freshwater environment. J. Appl. Toxicol. 2014, 34, 458–479. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Glyphosate effects on soil rhizosphere-associated bacterial communities. Sci. Total Environ. 2016, 543, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Allegrini, M.; Gomez, E.; Zabaloy, M.C. Repeated glyphosate exposure induces shifts in nitrifying communities and metabolism of phenylpropanoids. Soil Biol. Biochem. 2017, 105, 206–215. [Google Scholar] [CrossRef]
- Druille, M.; Omacini, M.; Golluscio, R.A.; Cabello, M.N. Arbuscular mycorrhizal fungi are directly and indirectly affected by glyphosate application. Appl. Soil Ecol. 2013, 72, 143–149. [Google Scholar] [CrossRef]
- Soares, C.; Fernandes, B.; Paiva, C.; Nogueira, V.; Cachada, A.; Fidalgo, F.; Pereira, R. Ecotoxicological relevance of glyphosate and flazasulfuron to soil habitat and retention functions – Single vs combined exposures. J. Hazard. Mater. 2023, 442, 130128. [Google Scholar] [CrossRef]
- Zhu, C.H. The toxic effects of glyphosate and cadmium on E. coli. Master's degree thesis, Hunan University of Science and Technology, China, 16 December 2019. 16 December.
- Deng, M.C. Response of lead, cadmium and glyphosate to pollution stress and HSPs in C. elegans. Master's Thesis, Northeast Normal University, China, 16 January 2015. [Google Scholar]
- Zhou, C.F. Ecological toxicicology of heavy metals and glyphosate. Doctoral dissertation, Nanjing Forestry University, China, 16 February 2014. 16 February.
- Han, B.; Gao, M.; Wang, Z.F. , Luo, Y.J.; Wang, L.; Tian, M. Microbial ecological effects of soil contaminated with Zn, Pb and glyphosate. Journal of Southwest University (Natural Science Edition), 2010, 32, 83–87. [Google Scholar]
- Wang, Y.B.; Li, R.Q.; Deng, M.C.; Li, Z.H.; Xu, J.B. Combined toxicity of arsenic with the pesticide glyphosate, and dichlorvos against C. elegans. Journal of Ecological Toxicology, 2013, 8, 262–267. [Google Scholar]
- Chen, J.J.; Rao, C.Y.; Yuan, R.J.; Sun, D.D.; Guo, S.Q.; Li, L.L.; Yang, S.; Qian, D.D.; Lu, R.H.; Cao, X.L. Long-term exposure to polyethylene microplastics and glyphosate interferes with the behavior, intestinal microbial homeostasis, and metabolites of the common carp (Cyprinus carpio L.). Sci. Total Environ 2022, 814, 152681. [Google Scholar] [CrossRef] [PubMed]
- Millet, M. A method to assess glyphosate, glufosinate and aminomethylphosphonic acid in soil and earthworms. J. Chromatogr. A 2021, 1651, 462339. [Google Scholar] [CrossRef]
- Pizzutti, I.R.; Vela, G.M.E.; De Kok, A.; Scholten, J.M.; Dias, J.V.; Cardoso, C.D.; Vivian, R. Determination of paraquat and diquat: LC-MS method optimization and validation. Food Chemistry 2016, 209, 248–255. [Google Scholar] [CrossRef] [PubMed]
- USDA. Soil survey manual. In: Soil Survey Division Staff; Soil Conservation Service Volume Handbook 18. U.S. Department of Agriculture (chapter 3), 2017.
- China Pesticide Information Network. http://www.chinapesticide.org.cn/. (accessed on 28 March 2023).
- Wołejko, E.; Jabłonska-Trypuc, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil biological activity as an indicator of soil pollution with pesticides–a review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- Widenfalk, A.; Bertilsson, S.; Sundh, I.; Goedkoop, W. Effects of pesticides on community composition and activity of sediment microbes–responses at various levels of microbial community organization. Environ. Pollut. 2008, 152, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, H.; Vera, M.S.; Vinocur, A.; P´erez, G.; Ferraro, M.; Helman, R.M.; Santos, D.; Afonso, M. Glyphosate input modifies microbial community structure in clear and turbid freshwater systems. Environ. Sci. Pollut. Res. 2016, 23, 5143–5153. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; Lorenz, N.; Saxena, J.; Ramsier, C.; Dick, R.P. The effect of glyphosate on soil microbial activity, microbial community structure, and soil potassium. Pedobiol. 2012, 55, 335–342. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2008, 285, 263–269. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Liu, Y.B.; Long, M.X.; Yin, Y.J.; Si, M.R.; Zhang, L.; Lu, Z.Q.; Shen, X.H. Physiological roles of mycothiol in detoxification and tolerance to multiple poisonous chemicals in Corynebacterium glutamicum. Arch. Microbiol. 2013, 195, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.B.; Moreno, M.V.; Amodeo, M.R.; Bianchinotti, M.V. Effects of Glyphosate on soil fungal communities: A field study. Revista Argentina de Microbiología, 2021, 53, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Romdhane, S.; Devers-Lamrani, M.; Beguet, J.; Bertrand, C.; Calvayrac, C.; Salvia, M.V.; Jrad, A.B.; Dayan, F.E.; Spor, A.; Barthelmebs, L.; Martin-Laurent, F. Assessment of the ecotoxicological impact of natural and synthetic β-triketone herbicides on the diversity and activity of the soil bacterial community using omic approaches. Sci. Total Environ. 2019, 651, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Prashar, P.; Shah, S. Impact of fertilizers and pesticides on soil microflora in agriculture. In: Sustainable Agriculture Reviews. Springer, Cham 2016, 331–36.
Figure 1.
The abundance distribution of Actinobacteria (a) and Proteobacteria (b). CK, Gl, Gm, Gh, Dl, Dm, Dh, GDl, GDm, GDh: control, low concentration glyphosate, medium concentration glyphosate, high concentration glyphosate, low concentration diquat, medium concentration diquat, high concentration diquat, low concentration mixed herbicides, medium concentration mixed herbicides, and high concentration mixed herbicides. Similarly hereinafter.
Figure 1.
The abundance distribution of Actinobacteria (a) and Proteobacteria (b). CK, Gl, Gm, Gh, Dl, Dm, Dh, GDl, GDm, GDh: control, low concentration glyphosate, medium concentration glyphosate, high concentration glyphosate, low concentration diquat, medium concentration diquat, high concentration diquat, low concentration mixed herbicides, medium concentration mixed herbicides, and high concentration mixed herbicides. Similarly hereinafter.
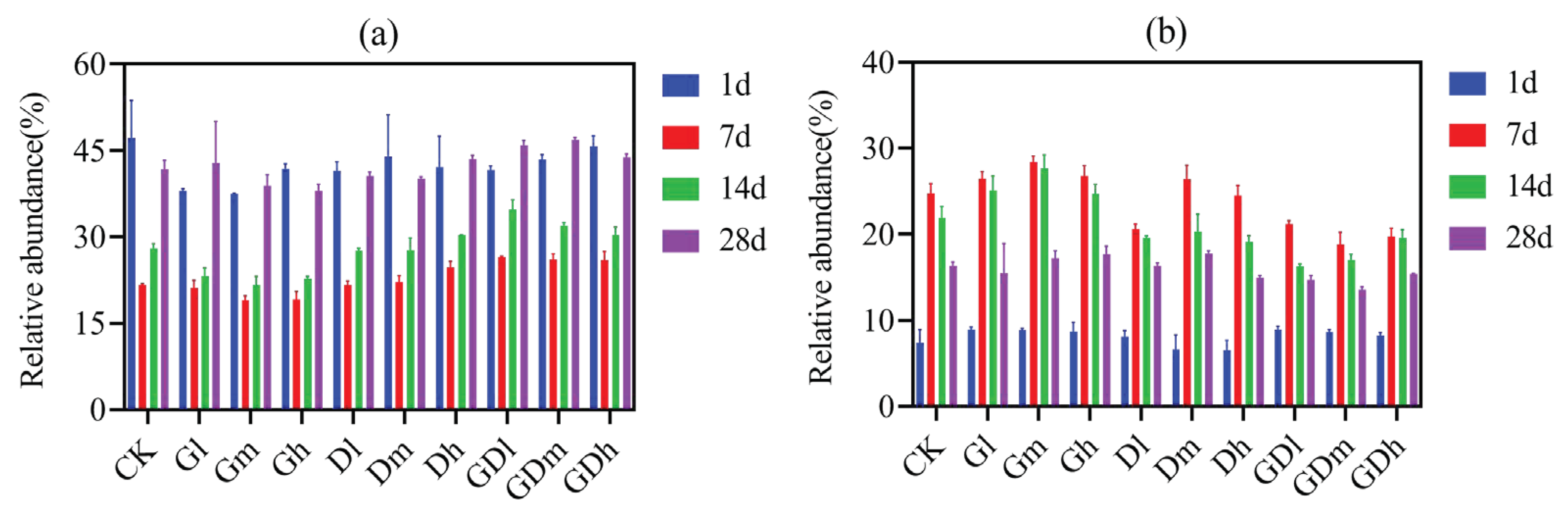
Figure 2.
Community composition of bacteria (a–d) and fungi (e–h) at genus level from different exposure time (1, 7, 14 and 28 d).
Figure 2.
Community composition of bacteria (a–d) and fungi (e–h) at genus level from different exposure time (1, 7, 14 and 28 d).
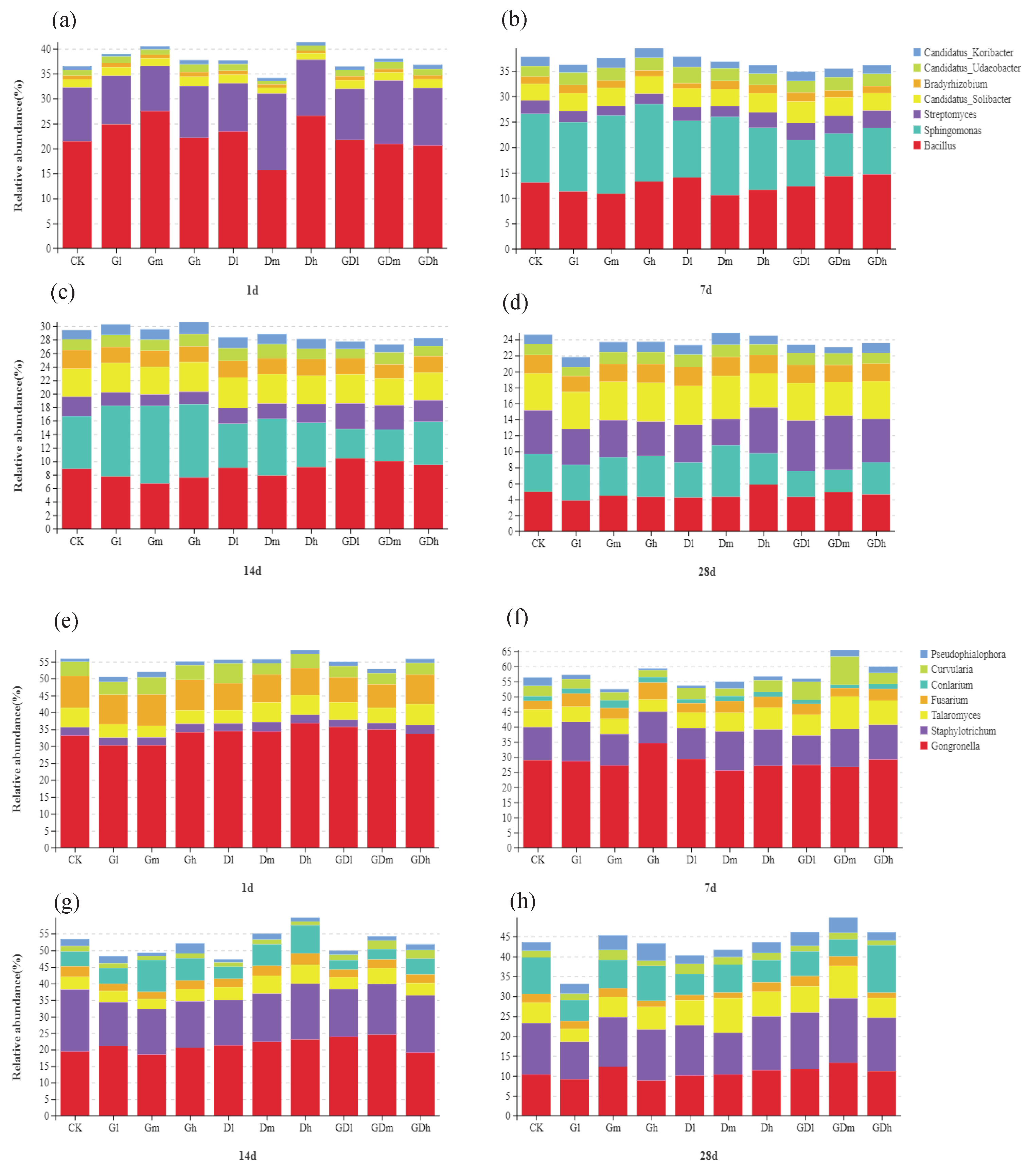
Figure 3.
Principal coordinate analysis for the bacterial communities at the phylum level at 1 day (a) and 14 day (b) exposure.
Figure 3.
Principal coordinate analysis for the bacterial communities at the phylum level at 1 day (a) and 14 day (b) exposure.
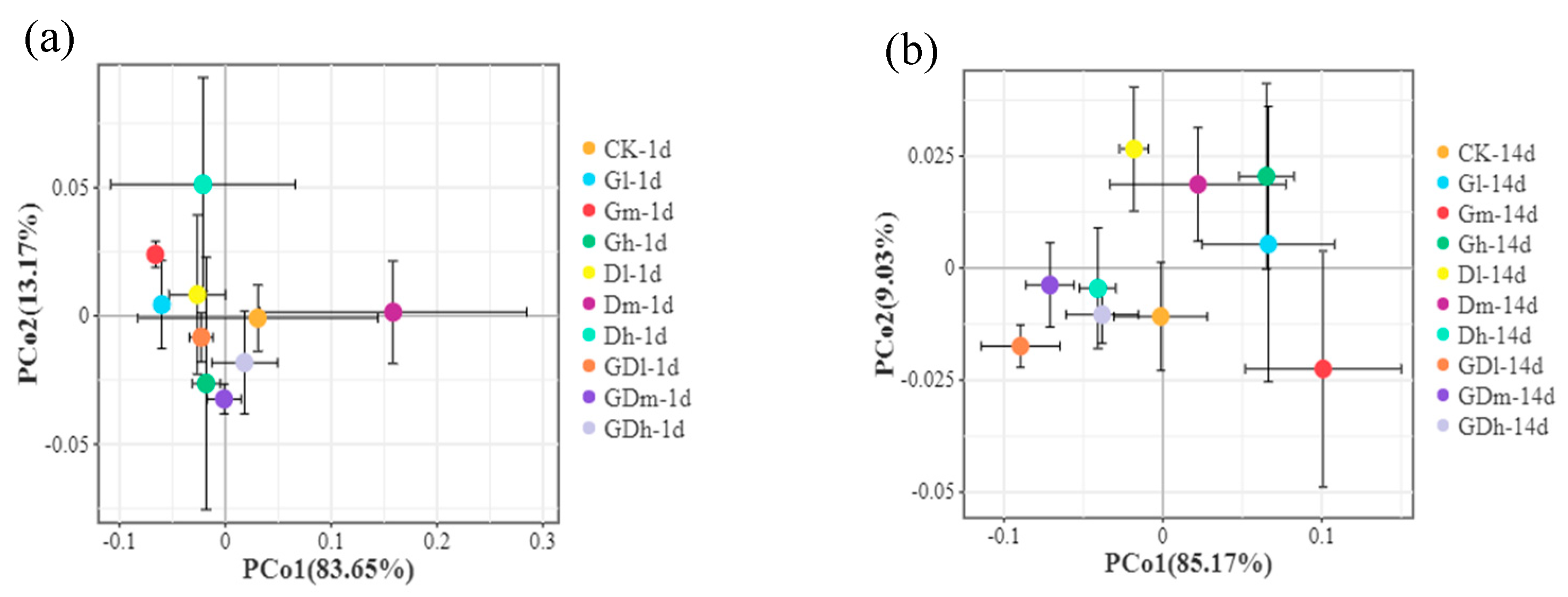
Figure 4.
Cladograms of line discriminant analysis effect size (LEfSe) analyses based on microbial community composition in different treats (The line discriminant analysis (LDA) scores above the preset value of 2.0 were considered to be significant. From the center outward, the circles represent the different taxonomic levels of the bacteria and fungi from the phylum to genus level. The yellow circles denote the taxa without significant differences among the different soil layers.) (a) bacteria, (b) fungi.
Figure 4.
Cladograms of line discriminant analysis effect size (LEfSe) analyses based on microbial community composition in different treats (The line discriminant analysis (LDA) scores above the preset value of 2.0 were considered to be significant. From the center outward, the circles represent the different taxonomic levels of the bacteria and fungi from the phylum to genus level. The yellow circles denote the taxa without significant differences among the different soil layers.) (a) bacteria, (b) fungi.
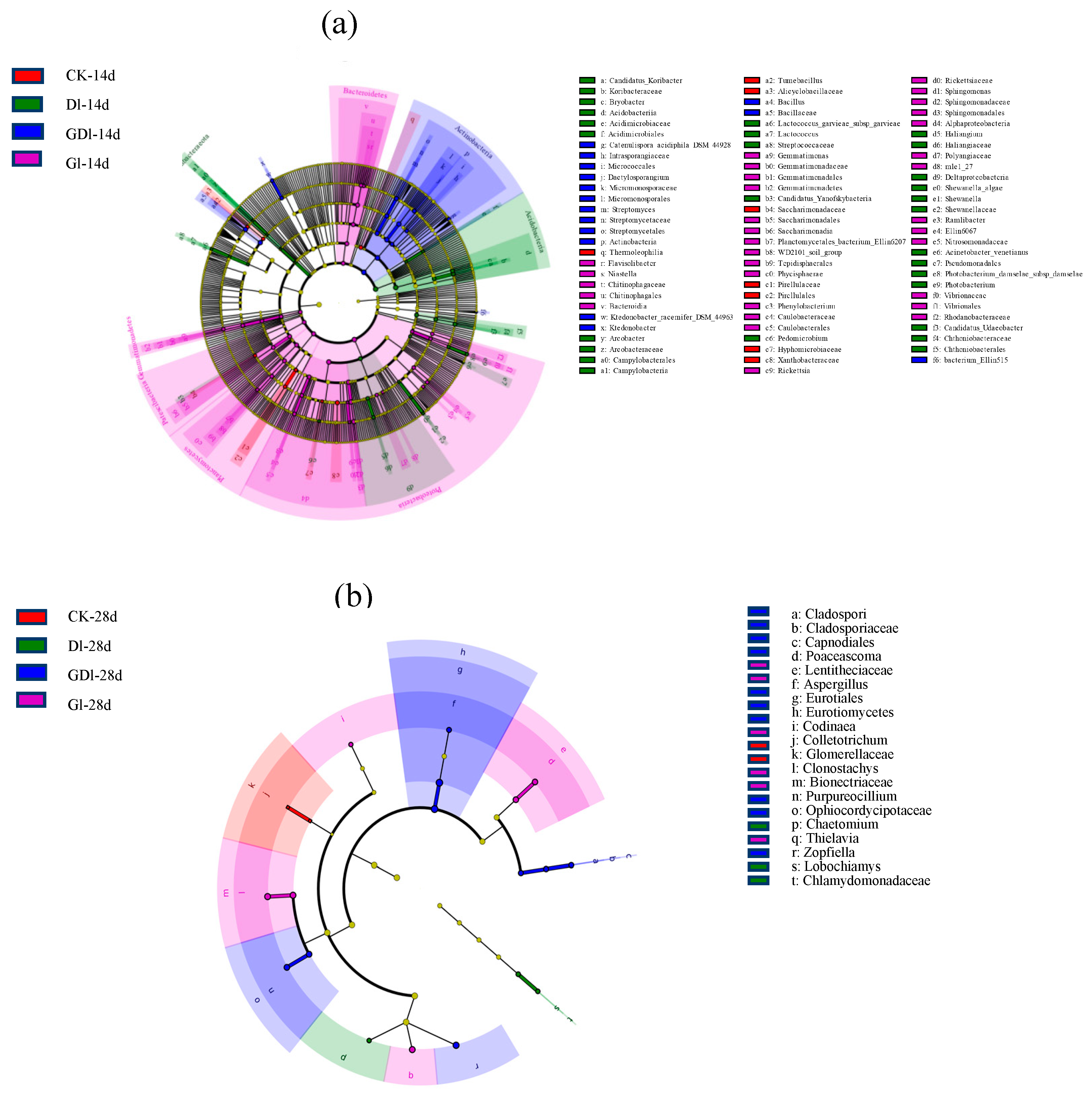
Figure 5.
Community composition of fungi at phylum level from different exposure time (1, 7, 14 and 28 d).
Figure 5.
Community composition of fungi at phylum level from different exposure time (1, 7, 14 and 28 d).
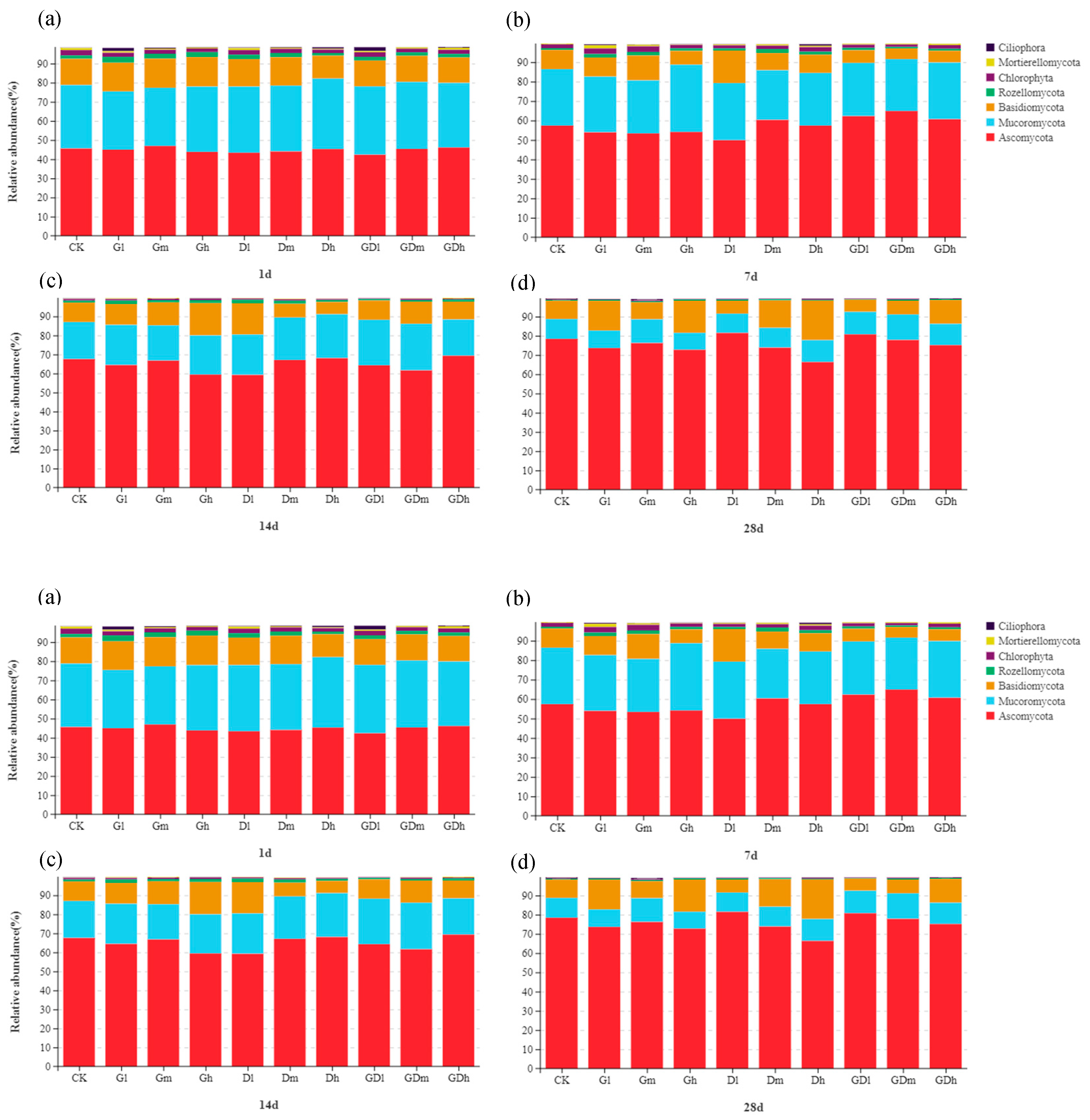
Figure 6.
Principal coordinate analysis (a–d) for fungus communities at the genus level at 1, 7, 14 and 28 day exposure.
Figure 6.
Principal coordinate analysis (a–d) for fungus communities at the genus level at 1, 7, 14 and 28 day exposure.
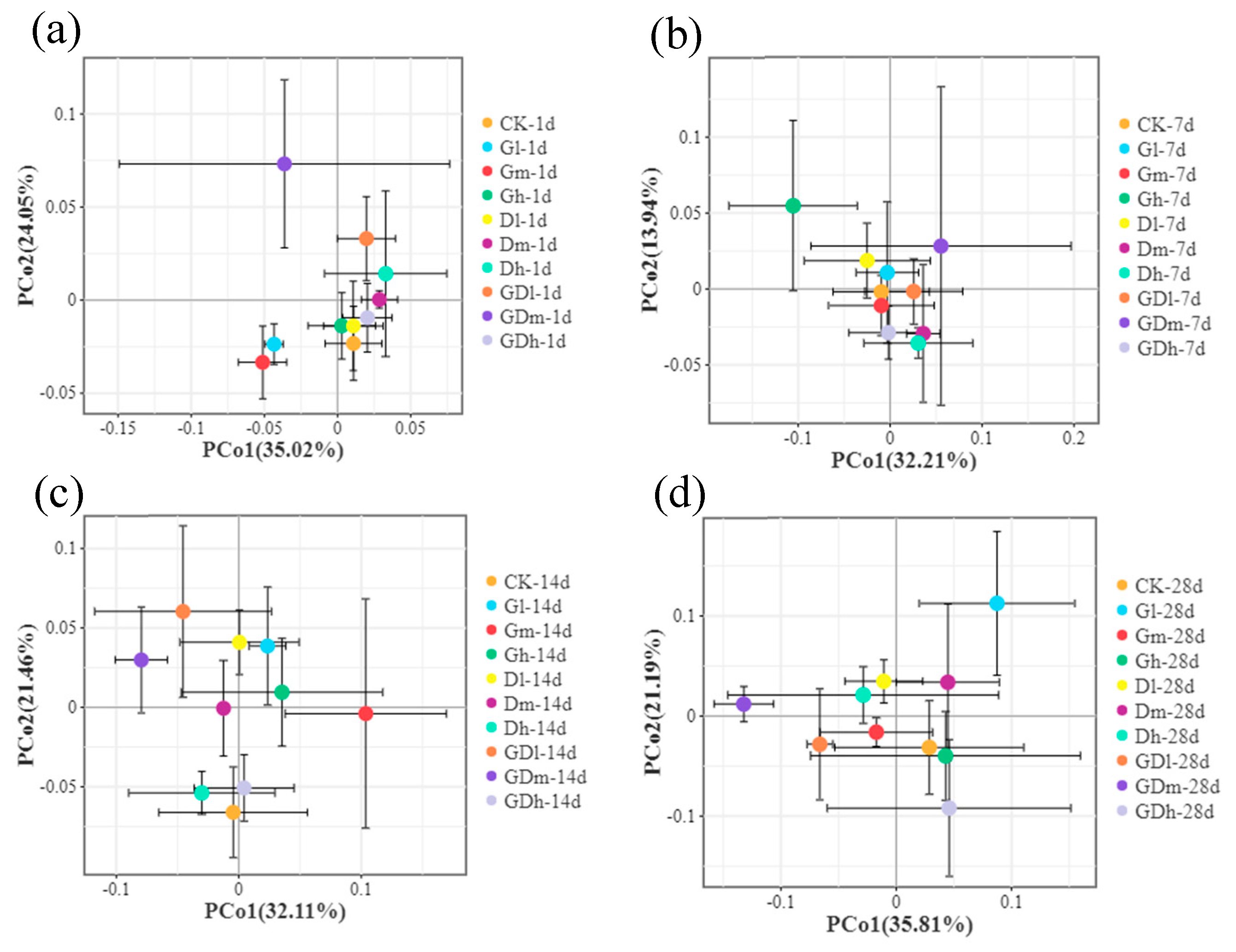

Table 1.
Trial treatment setting. The low concentration (L) was 1 times the recommended dosage, the medium concentration (M) was 10 times the recommended dosage, and the high concentration (H) was 100 times the recommended dosage. The recommended dosage were set 0.6 and 0.4 mg kg−1 for glyphosate (GP) and diquat (DQ).
Table 1.
Trial treatment setting. The low concentration (L) was 1 times the recommended dosage, the medium concentration (M) was 10 times the recommended dosage, and the high concentration (H) was 100 times the recommended dosage. The recommended dosage were set 0.6 and 0.4 mg kg−1 for glyphosate (GP) and diquat (DQ).
| Treatment | Concentration (mg kg−1) | ||
|---|---|---|---|
| Low (l) | Middle (m) | High (h) | |
| Blank (CK) | 0 | ||
| Glyphosate (G) | 0.6 (Gl) | 6 (Gm) | 60 (Gh) |
| Diquat (D) | 0.4 (Dl) | 4 (Dm) | 40 (Dh) |
| Glyphosate+diquat (GD) | 0.6+0.4 (GDl) | 6+4 (GDm) | 60+40 (GDh) |
Table 2.
Alpha diversity of bacterial communities under different treatments. Different letters indicate significant differences at the p <0.05 level between different treatments at the same time. Mean values (n = 3)±S.E.
Table 2.
Alpha diversity of bacterial communities under different treatments. Different letters indicate significant differences at the p <0.05 level between different treatments at the same time. Mean values (n = 3)±S.E.
| Index | Treatment | Days after application | |||
|---|---|---|---|---|---|
| 1 | 7 | 14 | 28 | ||
| Shannon | CK | 5.918±0.311ab | 7.264±0.027bc | 7.556±0.023cd | 7.315±0.093abc |
| Gl | 6.390±0.071a | 7.458±0.009a | 7.650±0.017abc | 7.254±0.320bc | |
| Gm | 6.249±0.063a | 7.405±0.032ab | 7.736±0.022a | 7.497±0.090ab | |
| Gh | 6.266±0.161a | 7.145±0.051c | 7.671±0.018ab | 7.453±0.067abc | |
| Dl | 6.131±0.140a | 7.407±0.033ab | 7.583±0.015bcd | 7.416±0.027abc | |
| Dm | 5.382±0.379b | 7.485±0.044a | 7.669±0.030ab | 7.629±0.006a | |
| Dh | 5.770±0.256ab | 7.416±0.038ab | 7.404±0.011e | 7.152±0.014bc | |
| GDl | 6.298±0.046a | 7.381±0.029ab | 7.284±0.078f | 7.100±0.034c | |
| GDm | 6.409±0.001a | 7.414±0.090ab | 7.505±0.015d | 7.090±0.023c | |
| GDh | 6.092±0.104a | 7.370±0.106ab | 7.499±0.038de | 7.238±0.039bc | |
| Chao1 | CK | 2188.649±150.191bcd | 2263.584±36.558ab | 2369.947±39.031a | 2303.254±45.996ab |
| Gl | 2536.324±75.475ab | 2377.208±3.108a | 2352.648±36.608a | 2219.027±135.281b | |
| Gm | 2602.062±60.436a | 2338.966±56.674a | 2386.601±49.563a | 2450.166±44.036aA | |
| Gh | 2508.073±91.128abc | 2184.617±30.267b | 2417.438±28.661a | 2285.848±30.396ab | |
| Dl | 2229.911±153.341bcd | 2406.547±28.774a | 2334.005±44.040a | 2352.122±40.203ab | |
| Dm | 1993.626±203.891d | 2417.230±8.741a | 2351.966±34.642a | 2254.606±47.180b | |
| Dh | 2154.460±98.706cd | 2329.326±48.987ab | 2249.354±70.180a | 2297.709±26.360ab | |
| GDl | 2372.901±44.448abc | 2332.078±103.682ab | 2305.535±69.564a | 2309.014±8.035ab | |
| GDm | 2488.896±9.119abc | 2363.224±52.350a | 2358.002±63.922a | 2366.971±23.944ab | |
| GDh | 2250.869±64.077abcd | 2380.824±14.352a | 2332.714±39.074a | 2250.471±9.346b | |
Table 3.
Alpha diversity of fungal communities under different treatments. Different letters indicate significant differences at the p < 0.05 level between different treatments at the same time. Mean values (n = 3)±S.E.
Table 3.
Alpha diversity of fungal communities under different treatments. Different letters indicate significant differences at the p < 0.05 level between different treatments at the same time. Mean values (n = 3)±S.E.
| Index | Treatment | Days after Application | |||
|---|---|---|---|---|---|
| 1 | 7 | 14 | 28 | ||
| Shannon | CK | 4.780±0.083abc | 4.368±0.046cd | 4.614±0.015a | 4.727±0.044abc |
| Gl | 4.882±0.054a | 4.634±0.023ab | 4.363±0.029ab | 4.783±0.039ab | |
| Gm | 4.765±0.011ab | 4.635±0.086bc | 4.259±0.135ab | 4.980±0.057a | |
| Gh | 4.544±0.129d | 4.231±0.043d | 4.388±0.121ab | 4.684±0.023cd | |
| Dl | 4.686±0.072bcd | 4.371±0.020cd | 4.259±0.076ab | 4.708±0.007bcd | |
| Dm | 4.683±0.018abc | 4.652±0.044ab | 4.516±0.052ab | 4.661±0.028bcd | |
| Dh | 4.509±0.014cd | 4.800±0.053a | 4.488±0.043ab | 4.598±0.053cd | |
| GDl | 4.688±0.033abc | 4.405±0.049cd | 4.128±0.111b | 4.867±0.076bcd | |
| GDm | 4.452±0.014d | 4.325±0.028cd | 4.376±0.097ab | 4.821±0.134abcd | |
| GDh | 4.763±0.060abc | 4.490±0.069cd | 4.369±0.010ab | 4.547±0.053d | |
| Chao1 | CK | 673.214±21.296ab | 543.470±18.721bc | 462.809±3.810b | 455.377±60.937a |
| Gl | 773.434±39.275ab | 579.153±19.726abc | 464.205±2.574b | 479.181±56.792a | |
| Gm | 772.912±28.255a | 626.580±29.634a | 465.700±4.486b | 469.934±53.990a | |
| Gh | 717.335±44.500ab | 510.983±12.810c | 452.857±1.069b | 438.914±55.892a | |
| Dl | 733.530±38.877ab | 546.209±15.359bc | 461.212±6.469b | 438.962±53.068a | |
| Dm | 706.838±26.245ab | 595.042±19.484ab | 477.764±9.637b | 425.494±44.871a | |
| Dh | 641.645±17.447b | 578.217±19.415ab | 468.624±8.868b | 437.094±48.999a | |
| GDl | 697.382±28.165ab | 540.641±15.796bc | 452.803±0.858b | 455.715±39.099a | |
| GDm | 666.482±30.530b | 500.521±2.086bc | 496.728±3.212a | 491.149±35.763a | |
| GDh | 679.040±27.961ab | 541.952±8.107abc | 499.915±16.673b | 426.687±6.304a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



