
Submitted:
23 April 2023
Posted:
24 April 2023
You are already at the latest version
Abstract
Dietary iron assimilation is critical for health and essential to prevent iron deficient states and related comorbidities, such as anemia. The bioavailability of iron is limited, while its absorption and metabolism are tightly controlled to keep body iron stores within a relatively narrow range. Genetic inactivation of the iron hormone hepcidin causes hereditary hemochromatosis, an endocrine disorder of iron overload characterized by chronic hyperabsorption of dietary iron, with deleterious clinical complications if untreated. The impact of high dietary iron intake and elevated body iron stores in the general population is not well understood. Herein, we summarize epidemiological data suggesting that high intake of heme iron, which is abundant in meat products, poses a risk factor for metabolic syndrome pathologies, cardiovascular diseases, and some cancers. We discuss clinical relevance and potential limitations of data from cohort studies, as well as the need to establish causality and elucidate molecular mechanisms.
Keywords:
Iron deficiency
; iron overload
; anemia
; heme
; metabolic syndrome
; cardiovascular disease
; cancer
; microbiome
1. Nutritional Value of Iron
Iron is an essential nutrient that is required for critical biological functions such as oxygen transport and cellular respiration. The adult human body contains 3-5 g of iron, with ~70% utilized in hemoglobin of red blood cells [1]. The daily iron requirements for erythropoiesis are 25-30 mg and are mostly met by iron recycled from senescent red blood cells, which are cleared by tissue macrophages. As there is no specific mechanism for iron excretion from the body, dietary absorption of 1-2 mg iron per day is essential to compensate for non-specific iron losses. It should be noted that under physiological conditions, only a small fraction (~15-20%) of ingested luminal iron gets eventually absorbed. The limited bioavailability of iron can lead to iron deficiency anemia or non-anemic iron deficiency, which are the most common pathologies worldwide and remain leading contributors to global burden of disease [2,3,4]. Iron deficiency is associated with fatigue and may also lead to immune, growth and neurocognitive defects (Figure 1). In 2005, anemias affected roughly a quarter of the world’s population with iron deficiency anemia accounting for about half of these cases [5]. Little progress has been made since then as, in 2016, iron deficiency anemia was one of the top five causes of years lived with disability with over 1.2 billion cases reported [6].
Nutritional guidelines developed by the Food and Nutrition Board at the National Academy of Medicine in the United States recommend that infants between 7-12 months obtain 11 mg of iron from their diet daily, whereas the corresponding values for adult men, menstruating women and pregnant women are 8, 18 and 27 mg of iron, respectively [7]. These guidelines highlight the elevated iron requirements in pregnant women and infants where iron is critical for growth and development [8].
Iron deficiency and anemia are more prevalent in vulnerable populations such as indigenous peoples, refugees and immigrants from low and middle-income countries, and disadvantaged subpopulations [4,9,10]. These notions provide the rational for food iron fortification programs [11], and for the use of oral iron supplements or intravenous iron for therapeutic purposes [12]. Nevertheless, excess body iron may lead to adverse health outcomes [13], mainly due to the redox reactivity of the metal that can promote oxidative stress and tissue damage [14]. This is vividly illustrated in diseases of iron overload such as hereditary hemochromatosis or iron-loading anemias (including thalassemia), which are associated with type 1 and type 2 diabetes mellitus, arthropathy, osteoporosis, hypogonadism, liver disease (fibrosis, cirrhosis, hepatocellular carcinoma) and cardiomyopathy (Figure 1) [15,16,17,18,19,20,21,22].
The hazardous effects of excess iron are also evident from cases of acute iron poisoning, for instance following accidental ingestion of iron supplements by children [23]. The severity of iron intoxication depends on the amount of iron ingested. Symptoms include nausea, vomiting, diarrhea, gastrointestinal bleeding, coagulopathy, shock, metabolic acidosis, hepatotoxicity, and abdominal pain. Thus, balanced iron intake is critical to avoid states of iron deficiency or overload.
2. Iron Homeostasis
Systemic iron homeostasis is largely controlled by hepcidin, a peptide hormone [24]. Circulating hepcidin is primarily produced by hepatocytes in the liver and targets the iron exporter ferroportin in intestinal enterocytes, tissue macrophages and other cells. The binding of hepcidin occludes ferroportin’s iron export channel and, additionally, triggers internalization and lysosomal degradation of ferroportin, limiting iron entry into the bloodstream [25,26,27]. Hepcidin is transcriptionally induced in response to iron or inflammatory cues via BMP/SMAD and IL-6/STAT3 signaling, respectively [28,29]. Iron-dependent induction of hepcidin primarily serves to prevent excessive dietary iron absorption and systemic iron overload [24]. Inflammatory induction of hepcidin is considered as an innate immune response that causes hypoferremia to deprive extracellular pathogens of iron, which is essential for their proliferation. However, persistent hypoferremia due to chronic inflammatory induction of hepcidin results in functional iron deficiency by restricting iron availability for erythropoiesis, thereby contributing to anemia of inflammation, also known as anemia of chronic disease [30]. Defective iron-dependent regulation of hepcidin due to genetic inactivation of upstream regulators leads to hereditary hemochromatosis, while suppression of hepcidin due to ineffective erythropoiesis contributes to iron overload in thalassemia and other iron-loading anemias [31].
3. Iron Biomarkers: Applications and Limitations
The major serum biomarkers typically used to evaluate iron status are ferritin, iron, transferrin saturation, soluble transferrin receptor 1 (sTfR1; also referred to as sTfR) and hepcidin [32]. Ferritin is normally an intracellular iron storage protein that can also be found in the circulation for reasons that remain unclear. Serum ferritin typically reflects body iron stores but is also influenced by inflammation, liver disease, obesity, and malignancies and must thus be paired with other tests for accurate diagnosis [33,34,35,36].
Measurement of sTfR1 and establishing sTfR1/log ferritin ratio are currently used to assess iron deficiency states [37,38]. TfR1 is the primary cellular iron gate and mediates import of circulating transferrin-bound iron [39]. It is induced in response to iron deficiency, and its shedding from the plasma membrane gives rise to sTfR1. Thus, plasma sTfR1 concentrations reflect protein density at the cell surface and the number of cells expressing the receptor, making it a good biomarker of iron demand [40,41]. It may be used to differentiate “true” from “functional” iron deficiency under inflammatory conditions [42].
Serum hepcidin can be used for assessing iron status, diagnosing iron deficiency states and for predicting responses to iron absorption from foods and supplements [43]. However, its main limitation, much like serum ferritin, is its induction by inflammation. Despite this, hepcidin may be a useful diagnostic tool in specific conditions such as chronic renal disorders or diseases of iron overload [44]. Yet, hepcidin (and sTfR1) assays have not yet become standardized tests due to harmonization challenges [38,43]. Serum hepcidin assays can vary up to 10-fold between tests due to the absence of a main reference material, a reference technique and a commutable calibrator, making it difficult to compare data and establish a uniform reference range [45,46].
4. Dietary Iron Intake and the Risk for Disease
High iron stores are often viewed as a potential biohazard [13], indicating that excessive dietary iron intake increases the risk for disease. Dietary iron is found in forms of heme or inorganic (non-heme) iron [47]. Heme is primarily present in hemoglobin and myoglobin from animal food sources, while inorganic iron is found in food derived from both plants and animals [48]. Heme consists of ~10-15% of total dietary iron sources in meat-eating populations, but accounts for over 40% of assimilated iron due to its enhanced absorption [48]. Thus, it is estimated that the efficiency of inorganic iron absorption is ~10% and of heme ~25%, which is 2.5 times higher [48]. This may be related to the lipophilic nature of the heme molecule, and possibly also to lack of negative feedback regulation of heme absorption. In any case, the intestinal heme transporter remains elusive, and its identification and characterization are expected to increase our mechanistic understanding. Thus far, it is well established that dietary absorbed heme is catabolized within intestinal epithelial cells and liberated iron follows the fate of dietary inorganic iron assimilated via the divalent metal transporter (DMT1). DMT1 is expressed on the apical membrane of duodenal enterocytes and is subjected to negative regulation by iron [49].
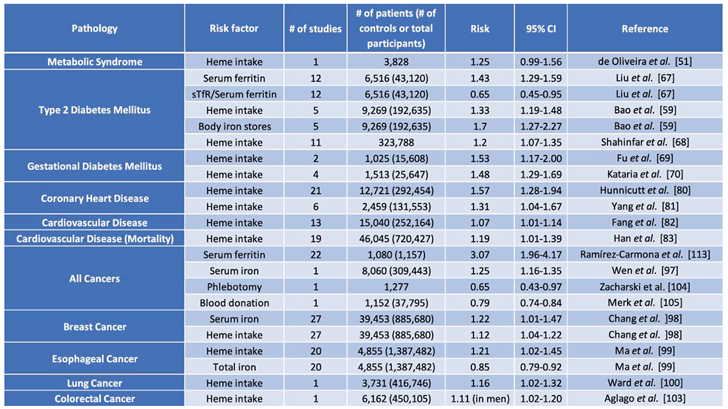
There is evidence from epidemiological studies that high dietary iron intake may predispose to diseases (Table 1). Most if not all these studies assessed dietary heme intake based on food questionnaires and data were extrapolated from meat consumption. However, it should be noted that processed red meat contains several potentially confounding substances, such as nitrate/nitrite, heterocyclic amines, polycyclic aromatic hydrocarbons etc., which may likewise affect health outcomes. Therefore, it is not clear whether the adverse effects of high dietary heme intake can always be entirely attributed to iron. This primarily applies to heme as opposed to inorganic iron as described in an umbrella review with a total of 34 meta-analyses [50]. The intake of heme but not inorganic iron was significantly associated with modestly increased risk for type 2 diabetes mellitus (T2DM), gestational diabetes mellitus, coronary heart disease, cardiovascular disease (CVD), CVD mortality, as well as colorectal, esophageal and breast cancer [50].
5. Iron and the Risk for Metabolic Syndrome
Metabolic syndrome is a pathologic state defined by combined manifestation of abdominal obesity, hyperglycemia due to insulin resistance, dyslipidemia and/or hypertension. Consequently, patients with metabolic syndrome are at greater risk of developing type 2 diabetes mellitus (T2DM), CVD, non-alcoholic fatty liver disease (NAFLD), and some types of cancer. Interestingly, in a multi-ethnic, population-based cohort of 3,828 participants, metabolic syndrome itself was associated with intake of heme iron [hazard ratio (HR) = 1.25 (95% CI = 0.99 to 1.56)] and zinc [HR = 1.29 (95% CI = 1.03 to 1.61)] from red meat [51]. Several clinical studies established a strong association between high body iron stores and insulin resistance [52,53,54,55], and the combination of iron overload with metabolic defects has been labeled as “dysmetabolic iron overload syndrome” (DIOS) [56]. Importantly, the alterations in iron metabolism appear to be multifactorial and dynamic due to unhealthy diet combined with environmental and genetic cofactors [56]. Despite evidence that dietary iron-depletion can attenuate NAFLD progression in mice [57], the benefits of phlebotomy on patients with DIOS remain controversial with a meta-analysis of 9 studies not finding significant improvement [58]. Nevertheless, the strong correlation of high dietary heme intake with T2DM and metabolic syndrome is consistent with iron-dependent effects [59,60]. In fact, there are several molecular links between iron and glucose metabolism [61]. For instance, insulin stimulates ferritin synthesis thereby increasing tissue iron stores; in addition, it promotes redistribution of transferrin receptors to the cell surface, thereby stimulating iron uptake [62].
Insulin was positively associated with serum ferritin levels in the 1988-1994 cross-sectional study from the Third National Health and Nutrition Examination Survey suggesting a possible link between T2DM and iron [63]. This association has since been corroborated in smaller cohort studies in Norwegian men, subjects with excessive body weight, and adolescent girls [64,65,66]. A meta-analysis found a correlation between high serum ferritin and T2DM in 11 out of 12 studies analyzed, with an odds ratio of 1.43 (95% CI 1.29 to 1.59) [67]. The ratio of sTfR1 to serum ferritin was also inversely related to risk of T2DM with an odds ratio of 0.65 (95% CI 0.45 to 0.95) in participants with high serum ferritin [67].
A 2012 meta-analysis of 5 primary prospective cohort studies suggested a relative risk of 1.33 (95% CI 1.19 to 1.48) for T2DM in individuals with highest vs lowest heme intake [59]. High body iron stores were likewise linked to T2DM with a relative risk of 1.70 (95% CI 1.27 to 2.27); however total dietary or supplemental iron intake did not show any significant association [59]. Another systematic review performed in 2021 came to the same conclusion [68]. This review included data from 323,788 participants over 11 studies and found that higher heme intake was associated with a 20% increased risk for developing T2DM (95% CI 1.07 to 1.35) [68].
Similar results were reported in two meta-analyses on iron and gestational diabetes mellitus (GDM). Fu et al. analyzed 1,025 GDM patients against 15,608 controls across 2 studies to compare the lowest and highest consumers of heme iron, and found a relative risk of 1.53 (95% CI 1.17 to 2.00) for development of GDM [69]. Kataria et al. analyzed the association between heme iron intake and GDM across 4 studies including the two studies from the previous meta-analysis and found an adjusted odds ratio of 1.48 (95% CI 1.29 to 1.69) [70]. Both studies confirmed a strong association between GDM and high body iron stores, but not intake of total dietary iron or iron supplements [69,70]. Due to the limited number of studies examining heme iron intake and GDM compared to T2DM, further investigation should be performed to provide a reliable correlation.
Molecular studies in cell and mouse models have linked iron overload to insulin resistance through mechanisms involving autophagy in skeletal muscles and cardiomyocytes [71,72]. Iron loading in cell and mouse models exacerbated palmitate-induced insulin resistance, impaired glucose-stimulated insulin secretion from pancreatic β cells, and increased gluconeogenesis in the liver [73,74,75]. Conversely, iron deficiency has been associated with insulin sensitivity in rats [76,77]. In young women, correction of iron deficiency anemia decreased insulin levels [78]. Taken together, these data provide a basis to investigate molecular mechanisms underlying the increased risk for metabolic syndrome pathologies in individuals with high dietary iron intake and elevated body iron stores.
6. Iron and the Risk for Cardiovascular Disease
The association between iron and CVD was first proposed in 1981 from observations that the incidence of CVD was elevated in men and post-menopausal women [79]. A meta-analysis of 21 cohort studies with 292,454 participants revealed a significant association between heme iron intake and coronary heart disease incidence [80], with relative risk of 1.57 (95% CI 1.28 to 1.94). These findings were corroborated in another meta-analysis from 6 different studies including 131,553 participants [81]. Interestingly, total iron intake, serum iron levels and transferrin saturation were inversely correlated with coronary heart disease incidence [80]. A meta-analysis of 13 primary studies with 252,164 participants reported a relative risk of 1.07 (95% CI 1.01 to 1.14) for CVD in individuals with high dietary heme intake [82]. Another meta-analysis of 19 studies with 720,427 participants reported an association between high dietary heme intake and CVD mortality, with a relative risk of 1.19 (95% CI 1.01 to 1.39) [83]. Iron status has also been positively associated with carotid atherosclerosis in the absence of inflammation [84]. Additionally, abdominal walls from patients having suffered abdominal aortic aneurysms displayed iron accumulation compared to healthy controls with elevated expression of TfR1 [85]. On the other hand, iron deficiency is a known comorbidity in patients with heart failure [86,87] and its correction with intravenous iron administration has been shown to reduce hospitalizations [88]. Taken together, these data highlight epidemiological links between iron status and CVD risk.
Animal studies have provided supportive evidence. Thus, early experiments in rabbits injected with iron dextran and fed a 0.5% cholesterol diet demonstrated greater atherosclerotic lesion development compared to diet alone [89]. Recent work in mice suggested that non-transferrin bound iron (NTBI), a highly redox active form of iron that appears in the circulation primarily during conditions of iron overload, aggravates atherosclerosis [90]. Crossing apolipoprotein E knockout (apoE–/–) mice, an established model of atherosclerosis, with mice that express a hepcidin-resistant ferroportin mutant (Fpnwt/C326S) aggravated atherosclerosis via increased levels of NTBI and oxidative stress [91]. Iron loading of the heart appears to be critically important when it is in cardiomyocytes as demonstrated by the reduced survival of mice lacking ferroportin in this cell type [92]. Thus, despite having elevated cardiac iron content, mouse models of hemochromatosis exhibit only minor cardiac dysfunction and develop cardiomyopathy only in response to chronic dietary or parenteral iron loading [93,94]. The functional importance of cardiomyocyte iron load is also emphasized by the lethal cardiomyopathy documented in mice with iron-deficient cardiomyocytes due to ablation of TfR1 [95] or expression of hepcidin-resistant FpnC326S [96]. Notably, local production of hepcidin is necessary for proper iron homeostasis in the heart [96], and a better understanding of its function and regulation is needed.
7. Iron and Cancer Risk
An observational study in a cohort of 309,443 adults in Taiwan identified an increased incidence of all cancers in individuals with high serum iron with a hazard ratio of 1.25 (95% CI 1.16 to 1.35), and a hazard ratio for mortality from all cancers of 1.39 (95% CI 1.23 to 1.57) [97]. Similar results were reported in a meta-analysis of 27 studies, where high serum iron correlated with higher relative risk for breast cancer (1.22; 95% CI 1.01 to 1.47) [98]. While no association between breast cancer and total dietary iron intake or inorganic iron supplementation was documented, dietary heme intake was associated with significant relative risk for breast cancer of 1.12 (95% CI 1.04 to 1.22) [97]. Dietary heme intake was also significantly associated with increased relative risk for esophageal cancer (1.21; 95% CI 1.02 to 1.45) [99], and lung cancer (1.16; 95% CI 1.02 to 1.32) after adjustment for smoking history [100].
The meta-analysis on esophageal cancer involved 20 studies with 1,387,482 study participants; interestingly total iron intake was found to be protective against esophageal cancer development (relative risk 0.85; 95% CI 0.79 to 0.92) [99]. A similar trend was also observed in the lung cancer study that involved 416,746 individuals from European countries [100]. Comparable data were also obtained from meta-analyses of studies focusing on colorectal cancer [101,102]. A European prospective cohort study of 450,105 participants followed for 14.2 ± 4.0 years found a stronger positive association of colorectal cancer in the proximal vs distal colon in men with high dietary heme intake (1.11; 95% CI:1.02 to 1.20) [103]. Yet, this trend was not observed in women, and other sources of iron were not associated with colorectal cancer [103]. Conversely, phlebotomy was associated with less new visceral malignancy in a prospective multicenter randomized clinical trial of 1,277 patients with peripheral arterial disease (0.65; 95% CI 0.43 to 0.97) [104]. Moreover, an analysis of 37,795 blood donors found an association with decreased risk of cancer (0.79; 95% CI 0.74 to 0.84) [105].
Heme may have diverse and often opposing functions in carcinogenesis [106]. In essence, above average levels of heme intake may lead to cellular cytotoxicity, lipid peroxidation, DNA and protein oxidation, and genetic mutations promoting carcinogenesis; however, excessive heme may also protect from carcinogenesis by shifting cell metabolism towards oxidative phosphorylation and eventually inducing cell death through ferroptosis, an iron-regulated programmed cell death [106]. It should be noted that highly proliferative cancer cells have increased needs for iron and undergo reprogramming for efficient iron acquisition and retention [107]. Studies in breast, ovarian, and prostate cancer have demonstrated reduced expression of ferroportin in the tumor to increase iron retention [108,109,110]. On the other hand, TfR1 is upregulated to increase iron uptake [111,112]. In line with these data, a meta-analysis of 22 studies revealed that serum ferritin levels were significantly higher in cancer patients with a standardized mean difference of 3.07 (CI 1.96 to 4.17) [113].
Mouse studies have validated clinical and epidemiological data on the role of iron as driver of carcinogenesis and, furthermore, have provided mechanistic insights. For instance, deletion of the Apc gene, an important precursor in colorectal cancer development, made mice susceptible to tumorigenesis from luminal iron [114]. Other studies demonstrated that dietary heme promotes epithelial hyperplasia in mice due to oxidative stress, hyperproliferation and reduced apoptosis of intestinal epithelial cells [115,116]. Thus, it is likely that toxicity of excess dietary heme is directly relevant to colorectal cancer. Nevertheless, since dietary heme promotes gut dysbiosis [117], which in turn may affect metabolic syndrome and other pathologies [118,119,120], it is possible that several of its adverse effects are indirect.
Mutations predisposing to hereditary hemochromatosis have been associated with increased risk of colorectal and breast cancer providing an association between iron and these malignancies [121]. Yet the most common type of cancer associated with hereditary hemochromatosis is hepatocellular carcinoma [122], which is also a common complication of transfusion-dependent thalassemias [123]. Interestingly, hemojuvelin knockout (Hjv–/–) mice, a model of juvenile hereditary hemochromatosis, are predisposed to hepatocarcinogenesis by a mechanism linked to mitochondrial hyperactivity [124].
8. Iron and the Intestinal Microbiome
Iron is also essential for microorganisms, and excessive dietary iron intake may affect the composition of bacterial communities in the gut. Iron deprivation has been shown to cause irreversible community alterations in the human intestinal microbiome, whereas iron supplementation resulted in small person-specific shifts [125]. Changes in microbial composition may be more relevant to specific populations or patients with increased sensitivity to such alterations. For example, the shifts in the gut microbiota from iron intake and supplementation has been proposed to influence progression of NAFLD in obese individuals [126].
Recent studies revealed adverse effects of iron fortification or supplementation in the intestinal microbiome of children in tropical countries that are vulnerable to infectious diseases [127,128] or of inflammatory bowel disease (IBD) patients [129]. In the latter, oral iron caused a decrease in Faecalibacterium prausnitzii and Ruminococcus bromi which are important anti-inflammatory bacteria by producing butyrate [129]. IBD poses an extra challenge as it is often associated with iron-deficiency anemia in part due to gastrointestinal bleeding [130], which provides an additional source of heme that may alter microbiome composition. Experiments in mice showed that dietary heme exacerbated dextran sodium sulphate (DSS)-induced colitis and promoted formation of adenomas [131]. Similar results were obtained in further studies involving the use of iron fortified chow in mouse models of colitis [132,133].
The gut microbiota has also been shown to play a role in colorectal cancer progression in mice, by sensitizing the mucus barrier in the presence of heme [134]. Heme may favor the growth of Akkermansia muciniphila, a mucin-degrading bacterium, which may lead to disruption of the mucus barrier [135]. Nevertheless, there is evidence suggesting that iron supplementation may be beneficial for the microbiome depending on formulation [136,137]. Additional studies would be needed to evaluate the impact of dietary or various forms of supplementary iron on the intestinal microbiome in the general population and in specific patient groups.
9. Conclusions
There is strong evidence from epidemiological studies that high dietary heme intake via meat consumption increases the risk for adverse health effects (Figure 2), particularly T2DM, GDM, coronary heart disease, CVD, and some cancers (such as colorectal, esophageal and breast cancer). These effects can be at least partially attributed to excessive iron accumulation due to the increased bioavailability of heme iron. However, a contribution of confounding factors present in meat products is also possible. It is reasonable to assume that iron-dependent adverse effects of heme are more evident in conditions which are also associated with high body iron stores, such as metabolic syndrome pathologies, GDM and some cancers, especially colorectal. Nevertheless, a clear causal relationship has not been established, and further experimental studies are needed to corroborate epidemiological data and explore underlying mechanisms. There is no evidence that dietary intake of inorganic iron, even from iron fortified foods or supplements, is unsafe for the general population. Regardless, it will be important to understand the impact of dietary iron on the intestinal microbiome, especially in the context of inflammatory and infectious diseases.
Author Contributions
Conceptualization, K.P.; writing—original draft preparation, E.C.; writing—review and editing, K.P.; funding acquisition, K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Canadian Institutes of Health Research (CIHR; PJT-486651). EC was funded by fellowships from the Natural Sciences and Engineering Research Council of Canada (NSERC) and from the Fonds de recherche du Québec – Santé (FRQS).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Katsarou, A.; Pantopoulos, K. Basics and principles of cellular and systemic iron homeostasis. Mol Aspects Med 2020, 75, 100866. https://doi.org/10.1016/j.mam.2020.100866. [CrossRef]
- Camaschella, C. Iron-deficiency anemia. N Engl J Med 2015, 372, 1832–1843. https://doi.org/10.1056/NEJMra1401038. [CrossRef]
- Al-Naseem, A.; Sallam, A.; Choudhury, S.; Thachil, J. Iron deficiency without anaemia: A diagnosis that matters. Clin Med (Lond) 2021, 21, 107–113. https://doi.org/10.7861/clinmed.2020-0582. [CrossRef]
- Pasricha, S.-R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. The Lancet 2021, 397, 233–248. https://doi.org/10.1016/S0140-6736(20)32594-0. [CrossRef]
- De Benoist, B.; Cogswell, M.; Egli, I.; McLean, E. Worldwide prevalence of anaemia 1993-2005; WHO global database of anaemia. 2008.
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. The Lancet 2017, 390, 1211–1259. https://doi.org/10.1016/S0140-6736(17)32154-2. [CrossRef]
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 294–301. https://doi.org/10.1016/S0002-8223(01)00078-5. [CrossRef]
- Pivina, L.; Semenova, Y.; Doşa, M.D.; Dauletyarova, M.; Bjørklund, G. Iron Deficiency, Cognitive Functions, and Neurobehavioral Disorders in Children. J. Mol. Neurosci. 2019, 68, 1–10. https://doi.org/10.1007/s12031-019-01276-1. [CrossRef]
- Pottie, K.; Greenaway, C.; Feightner, J.; Welch, V.; Swinkels, H.; Rashid, M.; Narasiah, L.; Kirmayer, L.J.; Ueffing, E.; MacDonald, N.E.; et al. Evidence-based clinical guidelines for immigrants and refugees. Can. Med. Assoc. J. 2011, 183, E824–E925. https://doi.org/10.1503/cmaj.090313. [CrossRef]
- Khambalia, A.Z.; Aimone, A.M.; Zlotkin, S.H. Burden of anemia among indigenous populations. Nutr. Rev. 2011, 69, 693–719. https://doi.org/10.1111/j.1753-4887.2011.00437.x. [CrossRef]
- Uauy, R.; Hertrampf, E.; Reddy, M. Iron fortification of foods: Overcoming technical and practical barriers. J Nutr 2002, 132, 849S–852S. https://doi.org/10.1093/jn/132.4.849S. [CrossRef]
- Auerbach, M.; Adamson, J.W. How we diagnose and treat iron deficiency anemia. Am J Hematol 2016, 91, 31–38. https://doi.org/10.1002/ajh.24201. [CrossRef]
- Weinberg, E.D. The hazards of iron loading. Metallomics 2010, 2, 732–740. https://doi.org/10.1039/c0mt00023j. [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim Biophys Acta Mol Cell Res 2019, 1866, 118535. https://doi.org/10.1016/j.bbamcr.2019.118535. [CrossRef]
- Deugnier, Y.; Turlin, B. Pathology of hepatic iron overload. Semin Liver Dis 2011, 31, 260–271. https://doi.org/10.1055/s-0031-1286057. [CrossRef]
- Kowdley, K.V. Iron, hemochromatosis, and hepatocellular carcinoma. Gastroenterology 2004, 127, S79–S86.
- Utzschneider, K.M.; Kowdley, K.V. Hereditary hemochromatosis and diabetes mellitus: Implications for clinical practice. Nat Rev Endocrinol 2010, 6, 26–33, doi:nrendo.2009.241 [pii] https://doi.org/10.1038/nrendo.2009.241. [CrossRef]
- Husar-Memmer, E.; Stadlmayr, A.; Datz, C.; Zwerina, J. HFE-related hemochromatosis: An update for the rheumatologist. Curr Rheumatol Rep 2014, 16, 393. https://doi.org/10.1007/s11926-013-0393-4. [CrossRef]
- Jeney, V. Clinical Impact and Cellular Mechanisms of Iron Overload-Associated Bone Loss. Front Pharmacol 2017, 8, 77. https://doi.org/10.3389/fphar.2017.00077. [CrossRef]
- Kremastinos, D.T.; Farmakis, D. Iron overload cardiomyopathy in clinical practice. Circulation 2011, 124, 2253–2263, doi:124/20/2253 [pii] https://doi.org/10.1161/CIRCULATIONAHA.111.050773. [CrossRef]
- Pelusi, C.; Gasparini, D.I.; Bianchi, N.; Pasquali, R. Endocrine dysfunction in hereditary hemochromatosis. J Endocrinol Invest 2016, 39, 837–847. https://doi.org/10.1007/s40618-016-0451-7. [CrossRef]
- De Sanctis, V.; Soliman, A.T.; Elsedfy, H.; Pepe, A.; Kattamis, C.; El Kholy, M.; Yassin, M. Diabetes and Glucose Metabolism in Thalassemia Major: An Update. Expert Rev Hematol 2016, 9, 401–408. https://doi.org/10.1586/17474086.2016.1136209. [CrossRef]
- Chang, T.P.; Rangan, C. Iron poisoning: A literature-based review of epidemiology, diagnosis, and management. Pediatr Emerg Care 2011, 27, 978–985. https://doi.org/10.1097/PEC.0b013e3182302604. [CrossRef]
- Nemeth, E.; Ganz, T. Hepcidin and Iron in Health and Disease. Annu Rev Med 2023, 74, 261–277. https://doi.org/10.1146/annurev-med-043021-032816. [CrossRef]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Ruwe, T.A.; Vieth, K.R.; Jung, G.; Casu, C.; Rivella, S.; et al. Structure-function analysis of ferroportin defines the binding site and an alternative mechanism of action of hepcidin. Blood 2018, 131, 899–910. https://doi.org/10.1182/blood-2017-05-786590. [CrossRef]
- Billesbølle, C.B.; Azumaya, C.M.; Kretsch, R.C.; Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of hepcidin-bound ferroportin reveals iron homeostatic mechanisms. Nature 2020, 10.1038/s41586-020-2668-z. https://doi.org/10.1038/s41586-020-2668-z. [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [CrossRef]
- Nemeth, E.; Rivera, S.; Gabayan, V.; Keller, C.; Taudorf, S.; Pedersen, B.K.; Ganz, T. IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. J. Clin. Invest. 2004, 113, 1271–1276. https://doi.org/10.1172/jci20945. [CrossRef]
- Babitt, J.L.; Huang, F.W.; Wrighting, D.M.; Xia, Y.; Sidis, Y.; Samad, T.A.; Campagna, J.A.; Chung, R.T.; Schneyer, A.L.; Woolf, C.J.; et al. Bone morphogenetic protein signaling by hemojuvelin regulates hepcidin expression. Nat. Genet. 2006, 38, 531–539. https://doi.org/10.1038/ng1777. [CrossRef]
- Weiss, G.; Ganz, T.; Goodnough, L.T. Anemia of inflammation. Blood 2019, 133, 40–50. https://doi.org/10.1182/blood-2018-06-856500. [CrossRef]
- Pantopoulos, K. Inherited Disorders of Iron Overload. Front Nutr 2018, 5, 103. https://doi.org/10.3389/fnut.2018.00103. [CrossRef]
- Lynch, S.; Pfeiffer, C.M.; Georgieff, M.K.; Brittenham, G.; Fairweather-Tait, S.; Hurrell, R.F.; McArdle, H.J.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Iron Review. J Nutr 2018, 148, 1001S–1067S. https://doi.org/10.1093/jn/nxx036. [CrossRef]
- Walters, G.O.; Miller, F.M.; Worwood, M. Serum ferritin concentration and iron stores in normal subjects. J. Clin. Pathol. 1973, 26, 770–772. https://doi.org/10.1136/jcp.26.10.770. [CrossRef]
- Lipschitz, D.A.; Cook, J.D.; Finch, C.A. A Clinical Evaluation of Serum Ferritin as an Index of Iron Stores. New England Journal of Medicine 1974, 290, 1213–1216. https://doi.org/10.1056/nejm197405302902201. [CrossRef]
- Marcus, D.M.; Zinberg, N. Measurement of Serum Ferritin by Radioimmunoassay: Results in Normal Individuals and Patients With Breast Cancer2. JNCI: Journal of the National Cancer Institute 1975, 55, 791–795. https://doi.org/10.1093/jnci/55.4.791. [CrossRef]
- McKinnon, E.J.; Rossi, E.; Beilby, J.P.; Trinder, D.; Olynyk, J.K. Factors That Affect Serum Levels of Ferritin in Australian Adults and Implications for Follow-up. Clin. Gastroenterol. Hepatol. 2014, 12, 101–108.e104. https://doi.org/10.1016/j.cgh.2013.07.019. [CrossRef]
- Infusino, I.; Braga, F.; Dolci, A.; Panteghini, M. Soluble Transferrin Receptor (sTfR) and sTfR/log Ferritin Index for the Diagnosis of Iron-Deficiency Anemia A Meta-Analysis. Am. J. Clin. Pathol. 2012, 138, 642–649. https://doi.org/10.1309/ajcp16ntxzlzfaib. [CrossRef]
- Shin, D.H.; Kim, H.S.; Park, M.J.; Suh, I.B.; Shin, K.S. Utility of access soluble transferrin receptor (sTfR) and sTfR/log ferritin index in diagnosing iron deficiency anemia. Ann. Clin. Lab. Sci. 2015, 45, 396–402.
- Gkouvatsos, K.; Papanikolaou, G.; Pantopoulos, K. Regulation of iron transport and the role of transferrin. Biochim Biophys Acta 2012, 1820, 188–202. https://doi.org/10.1016/j.bbagen.2011.10.013. [CrossRef]
- Speeckaert, M.M.; Speeckaert, R.; Delanghe, J.R. Biological and clinical aspects of soluble transferrin receptor. Crit. Rev. Clin. Lab. Sci. 2010, 47, 213–228. https://doi.org/10.3109/10408363.2010.550461. [CrossRef]
- Skikne, B.S. Serum transferrin receptor. Am. J. Hematol. 2008, 83, 872–875. https://doi.org/10.1002/ajh.21279. [CrossRef]
- Berlin, T.; Meyer, A.; Rotman-Pikielny, P.; Natur, A.; Levy, Y. Soluble transferrin receptor as a diagnostic laboratory test for detection of iron deficiency anemia in acute illness of hospitalized patients. The Israel Medical Association journal : IMAJ 2011, 13, 96–98.
- Girelli, D.; Nemeth, E.; Swinkels, D.W. Hepcidin in the diagnosis of iron disorders. Blood 2016, 127, 2809–2813. https://doi.org/10.1182/blood-2015-12-639112. [CrossRef]
- Fathi, Z.H.; Mohammad, J.A.; Younus, Z.M.; Mahmood, S.M. Hepcidin as a Potential Biomarker for the Diagnosis of Anemia. Turk J Pharm Sci 2022, 19, 603–609. https://doi.org/10.4274/tjps.galenos.2021.29488. [CrossRef]
- Kroot, J.J.; van Herwaarden, A.E.; Tjalsma, H.; Jansen, R.T.; Hendriks, J.C.; Swinkels, D.W. Second round robin for plasma hepcidin methods: First steps toward harmonization. Am. J. Hematol. 2012, 87, 977–983. [CrossRef]
- van der Vorm, L.N.; Hendriks, J.C.; Laarakkers, C.M.; Klaver, S.; Armitage, A.E.; Bamberg, A.; Geurts-Moespot, A.J.; Girelli, D.; Herkert, M.; Itkonen, O. Toward worldwide hepcidin assay harmonization: Identification of a commutable secondary reference material. Clin. Chem. 2016, 62, 993–1001. [CrossRef]
- Carpenter, C.E.; Mahoney, A.W. Contributions of heme and nonheme iron to human nutrition. Critical Reviews in Food Science and Nutrition 1992, 31, 333–367. https://doi.org/10.1080/10408399209527576. [CrossRef]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. The American Journal of Clinical Nutrition 2010, 91, 1461S–1467S. https://doi.org/10.3945/ajcn.2010.28674F. [CrossRef]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am J Physiol Gastrointest Liver Physiol 2014, 307, G397-G409, doi:ajpgi.00348.2013 [pii] https://doi.org/10.1152/ajpgi.00348.2013. [CrossRef]
- Huang, Y.; Cao, D.; Chen, Z.; Chen, B.; Li, J.; Wang, R.; Guo, J.; Dong, Q.; Liu, C.; Wei, Q.; et al. Iron intake and multiple health outcomes: Umbrella review. Critical Reviews in Food Science and Nutrition 2021, 10.1080/10408398.2021.1982861, 1-18. https://doi.org/10.1080/10408398.2021.1982861. [CrossRef]
- de Oliveira Otto, M.C.; Alonso, A.; Lee, D.-H.; Delclos, G.L.; Bertoni, A.G.; Jiang, R.; Lima, J.A.; Symanski, E.; Jacobs, D.R., Jr; Nettleton, J.A. Dietary Intakes of Zinc and Heme Iron from Red Meat, but Not from Other Sources, Are Associated with Greater Risk of Metabolic Syndrome and Cardiovascular Disease. The Journal of Nutrition 2012, 142, 526–533. https://doi.org/10.3945/jn.111.149781. [CrossRef]
- Bozzini, C.; Girelli, D.; Olivieri, O.; Martinelli, N.; Bassi, A.; De Matteis, G.; Tenuti, I.; Lotto, V.; Friso, S.; Pizzolo, F.; et al. Prevalence of body iron excess in the metabolic syndrome. Diabetes Care 2005, 28, 2061–2063. [CrossRef]
- Fernandez-Real, J.M.; Ricart-Engel, W.; Arroyo, E.; Balanca, R.; Casamitjana-Abella, R.; Cabrero, D.; Fernandez-Castaner, M.; Soler, J. Serum ferritin as a component of the insulin resistance syndrome. Diabetes Care 1998, 21, 62–68. [CrossRef]
- Lee, D.H.; Liu, D.Y.; Jacobs, D.R., Jr.; Shin, H.R.; Song, K.; Lee, I.K.; Kim, B.; Hider, R.C. Common presence of non-transferrin-bound iron among patients with type 2 diabetes. Diabetes Care 2006, 29, 1090–1095. https://doi.org/10.2337/diacare.2951090. [CrossRef]
- Mendler, M.H.; Turlin, B.; Moirand, R.; Jouanolle, A.M.; Sapey, T.; Guyader, D.; Le Gall, J.Y.; Brissot, P.; David, V.; Deugnier, Y. Insulin resistance-associated hepatic iron overload. Gastroenterology 1999, 117, 1155–1163, doi:S0016508599006150 [pii]. [CrossRef]
- Sachinidis, A.; Doumas, M.; Imprialos, K.; Stavropoulos, K.; Katsimardou, A.; Athyros, V.G. Dysmetabolic Iron Overload in Metabolic Syndrome. Curr Pharm Des 2020, 26, 1019–1024. https://doi.org/10.2174/1381612826666200130090703. [CrossRef]
- Crawford, D.H.G.; Ross, D.G.F.; Jaskowski, L.A.; Burke, L.J.; Britton, L.J.; Musgrave, N.; Briskey, D.; Rishi, G.; Bridle, K.R.; Subramaniam, V.N. Iron depletion attenuates steatosis in a mouse model of non-alcoholic fatty liver disease: Role of iron-dependent pathways. Biochim Biophys Acta Mol Basis Dis 2021, 1867, 166142. https://doi.org/10.1016/j.bbadis.2021.166142. [CrossRef]
- Murali, A.R.; Gupta, A.; Brown, K. Systematic review and meta-analysis to determine the impact of iron depletion in dysmetabolic iron overload syndrome and non-alcoholic fatty liver disease. Hepatol Res 2018, 48, E30–E41. https://doi.org/10.1111/hepr.12921. [CrossRef]
- Bao, W.; Rong, Y.; Rong, S.; Liu, L. Dietary iron intake, body iron stores, and the risk of type 2 diabetes: A systematic review and meta-analysis. BMC Med 2012, 10, 119. https://doi.org/10.1186/1741-7015-10-119. [CrossRef]
- Dos Santos Vieira, D.A.; Hermes Sales, C.; Galvão Cesar, C.L.; Marchioni, D.M.; Fisberg, R.M. Influence of Haem, Non-Haem, and Total Iron Intake on Metabolic Syndrome and Its Components: A Population-Based Study. Nutrients 2018, 10, 314. [CrossRef]
- Fillebeen, C.; Lam, N.H.; Chow, S.; Botta, A.; Sweeney, G.; Pantopoulos, K. Regulatory Connections between Iron and Glucose Metabolism. Int J Mol Sci 2020, 21. https://doi.org/10.3390/ijms21207773. [CrossRef]
- Davis, R.J.; Corvera, S.; Czech, M.P. Insulin stimulates cellular iron uptake and causes the redistribution of intracellular transferrin receptors to the plasma membrane. J. Biol. Chem. 1986, 261, 8708–8711. https://doi.org/10.1016/S0021-9258(19)84438-1. [CrossRef]
- Ford, E.S.; Cogswell, M.E. Diabetes and serum ferritin concentration among U.S. adults. Diabetes Care 1999, 22, 1978–1983. https://doi.org/10.2337/diacare.22.12.1978. [CrossRef]
- Oshaug, A.; Bugge, K.H.; Bjønnes, C.H.; Borch-Iohnsen, B.; Neslein, I.L. Associations between serum ferritin and cardiovascular risk factors in healthy young men. A cross sectional study. Eur. J. Clin. Nutr. 1995, 49, 430–438.
- Vaquero, M.P.; Martínez-Maqueda, D.; Gallego-Narbón, A.; Zapatera, B.; Pérez-Jiménez, J. Relationship between iron status markers and insulin resistance: An exploratory study in subjects with excess body weight. PeerJ 2020, 8, e9528. https://doi.org/10.7717/peerj.9528. [CrossRef]
- Klisic, A.; Kavaric, N.; Kotur, J.; Ninic, A. Serum soluble transferrin receptor levels are independently associated with homeostasis model assessment of insulin resistance in adolescent girls. Arch. Med. Sci. 2021, 10.5114/aoms/132757. https://doi.org/10.5114/aoms/132757. [CrossRef]
- Liu, J.; Li, Q.; Yang, Y.; Ma, L. Iron metabolism and type 2 diabetes mellitus: A meta-analysis and systematic review. Journal of Diabetes Investigation 2020, 11, 946–955. https://doi.org/10.1111/jdi.13216. [CrossRef]
- Shahinfar, H.; Jayedi, A.; Shab-Bidar, S. Dietary iron intake and the risk of type 2 diabetes: A systematic review and dose–response meta-analysis of prospective cohort studies. Eur. J. Nutr. 2022, 61, 2279–2296. https://doi.org/10.1007/s00394-022-02813-2. [CrossRef]
- Fu, S.; Li, F.; Zhou, J.; Liu, Z. The Relationship Between Body Iron Status, Iron Intake And Gestational Diabetes: A Systematic Review and Meta-Analysis. Medicine (Baltimore) 2016, 95, e2383. https://doi.org/10.1097/MD.0000000000002383. [CrossRef]
- Kataria, Y.; Wu, Y.; Horskjaer, P.H.; Mandrup-Poulsen, T.; Ellervik, C. Iron Status and Gestational Diabetes-A Meta-Analysis. Nutrients 2018, 10. https://doi.org/10.3390/nu10050621. [CrossRef]
- Jahng, J.W.S.; Alsaadi, R.M.; Palanivel, R.; Song, E.; Hipolito, V.E.B.; Sung, H.K.; Botelho, R.J.; Russell, R.C.; Sweeney, G. Iron overload inhibits late stage autophagic flux leading to insulin resistance. EMBO reports 2019, 20, e47911. https://doi.org/10.15252/embr.201947911. [CrossRef]
- Sung, H.K.; Song, E.; Jahng, J.W.S.; Pantopoulos, K.; Sweeney, G. Iron induces insulin resistance in cardiomyocytes via regulation of oxidative stress. Sci. Rep. 2019, 9, 4668. https://doi.org/10.1038/s41598-019-41111-6. [CrossRef]
- Cui, R.; Choi, S.-E.; Kim, T.H.; Lee, H.J.; Lee, S.J.; Kang, Y.; Jeon, J.Y.; Kim, H.J.; Lee, K.-W. Iron overload by transferrin receptor protein 1 regulation plays an important role in palmitate-induced insulin resistance in human skeletal muscle cells. The FASEB Journal 2019, 33, 1771–1786. https://doi.org/10.1096/fj.201800448R. [CrossRef]
- Hansen, Jakob B.; Tonnesen, Morten F.; Madsen, Andreas N.; Hagedorn, Peter H.; Friberg, J.; Grunnet, Lars G.; Heller, R.S.; Nielsen, Anja Ø.; Størling, J.; Baeyens, L.; et al. Divalent Metal Transporter 1 Regulates Iron-Mediated ROS and Pancreatic β Cell Fate in Response to Cytokines. Cell Metab. 2012, 16, 449–461. https://doi.org/10.1016/j.cmet.2012.09.001. [CrossRef]
- Huang, J.; Jones, D.; Luo, B.; Sanderson, M.; Soto, J.; Abel, E.D.; Cooksey, R.C.; McClain, D.A. Iron Overload and Diabetes Risk: A Shift From Glucose to Fatty Acid Oxidation and Increased Hepatic Glucose Production in a Mouse Model of Hereditary Hemochromatosis. Diabetes 2010, 60, 80–87. https://doi.org/10.2337/db10-0593. [CrossRef]
- Borel, M.J.; Beard, J.L.; Farrell, P.A. Hepatic glucose production and insulin sensitivity and responsiveness in iron-deficient anemic rats. American Journal of Physiology-Endocrinology and Metabolism 1993, 264, E380–E390. https://doi.org/10.1152/ajpendo.1993.264.3.E380. [CrossRef]
- Farrell, P.A.; Beard, J.L.; Druckenmiller, M. Increased Insulin Sensitivity in Iron-Deficient Rats. The Journal of Nutrition 1988, 118, 1104–1109. https://doi.org/10.1093/jn/118.9.1104. [CrossRef]
- Ozdemir, A.; Sevinç, C.; Selamet, U.; Kamaci, B.; Atalay, S. Age- and body mass index-dependent relationship between correction of iron deficiency anemia and insulin resistance in non-diabetic premenopausal women. Ann. Saudi Med. 2007, 27, 356–361. https://doi.org/10.5144/0256-4947.2007.356. [CrossRef]
- Sullivan, J. Iron and the sex difference in heart disease risk. The Lancet 1981, 317, 1293–1294. [CrossRef]
- Hunnicutt, J.; He, K.; Xun, P. Dietary iron intake and body iron stores are associated with risk of coronary heart disease in a meta-analysis of prospective cohort studies. J Nutr 2014, 144, 359–366. https://doi.org/10.3945/jn.113.185124. [CrossRef]
- Yang, W.; Li, B.; Dong, X.; Zhang, X.Q.; Zeng, Y.; Zhou, J.L.; Tang, Y.H.; Xu, J.J. Is heme iron intake associated with risk of coronary heart disease? A meta-analysis of prospective studies. Eur J Nutr 2014, 53, 395–400. https://doi.org/10.1007/s00394-013-0535-5. [CrossRef]
- Fang, X.; An, P.; Wang, H.; Wang, X.; Shen, X.; Li, X.; Min, J.; Liu, S.; Wang, F. Dietary intake of heme iron and risk of cardiovascular disease: A dose-response meta-analysis of prospective cohort studies. Nutr Metab Cardiovasc Dis 2015, 25, 24–35. https://doi.org/10.1016/j.numecd.2014.09.002. [CrossRef]
- Han, M.; Guan, L.; Ren, Y.; Zhao, Y.; Liu, D.; Zhang, D.; Liu, L.; Liu, F.; Chen, X.; Cheng, C.; et al. Dietary iron intake and risk of death due to cardiovascular diseases: A systematic review and dose-response meta-analysis of prospective cohort studies. Asia Pac J Clin Nutr 2020, 29, 309–321. https://doi.org/10.6133/apjcn.202007_29(2).0014. [CrossRef]
- Ahluwalia, N.; Genoux, A.; Ferrieres, J.; Perret, B.; Carayol, M.; Drouet, L.; Ruidavets, J.-B. Iron Status Is Associated with Carotid Atherosclerotic Plaques in Middle-Aged Adults. The Journal of Nutrition 2010, 140, 812–816. https://doi.org/10.3945/jn.109.110353. [CrossRef]
- Sawada, H.; Hao, H.; Naito, Y.; Oboshi, M.; Hirotani, S.; Mitsuno, M.; Miyamoto, Y.; Hirota, S.; Masuyama, T. Aortic Iron Overload with Oxidative Stress and Inflammation in Human and Murine Abdominal Aortic Aneurysm. Arteriosclerosis, Thrombosis, and Vascular Biology 2015, 35, 1507–1514. https://doi.org/10.1161/ATVBAHA.115.305586. [CrossRef]
- Alnuwaysir, R.I.S.; Hoes, M.F.; van Veldhuisen, D.J.; van der Meer, P.; Grote Beverborg, N. Iron Deficiency in Heart Failure: Mechanisms and Pathophysiology. J Clin Med 2021, 11. https://doi.org/10.3390/jcm11010125. [CrossRef]
- Anand, I.S.; Gupta, P. Anemia and Iron Deficiency in Heart Failure: Current Concepts and Emerging Therapies. Circulation 2018, 138, 80–98. https://doi.org/10.1161/CIRCULATIONAHA.118.030099. [CrossRef]
- Salah, H.M.; Savarese, G.; Rosano, G.M.C.; Ambrosy, A.P.; Mentz, R.J.; Fudim, M. Intravenous iron infusion in patients with heart failure: A systematic review and study-level meta-analysis. ESC Heart Fail 2023, 10, 1473–1480. https://doi.org/10.1002/ehf2.14310. [CrossRef]
- Araujo, J.A.; Romano, E.L.; Brito, B.E.; Parthé, V.; Romano, M.; Bracho, M.; Montaño, R.F.; Cardier, J. Iron Overload Augments the Development of Atherosclerotic Lesions in Rabbits. Arteriosclerosis, Thrombosis, and Vascular Biology 1995, 15, 1172–1180, https://doi.org/10.1161/01.ATV.15.8.1172. [CrossRef]
- Vinchi, F.; Porto, G.; Simmelbauer, A.; Altamura, S.; Passos, S.T.; Garbowski, M.; Silva, A.M.N.; Spaich, S.; Seide, S.E.; Sparla, R.; et al. Atherosclerosis is aggravated by iron overload and ameliorated by dietary and pharmacological iron restriction. Eur. Heart J. 2019, 41, 2681–2695. https://doi.org/10.1093/eurheartj/ehz112. [CrossRef]
- Vinchi, F.; Porto, G.; Simmelbauer, A.; Altamura, S.; Passos, S.T.; Garbowski, M.; Silva, A.M.N.; Spaich, S.; Seide, S.E.; Sparla, R.; et al. Atherosclerosis is aggravated by iron overload and ameliorated by dietary and pharmacological iron restriction. Eur Heart J 2020, 41, 2681–2695. https://doi.org/10.1093/eurheartj/ehz112. [CrossRef]
- Lakhal-Littleton, S.; Wolna, M.; Carr, C.A.; Miller, J.J.J.; Christian, H.C.; Ball, V.; Santos, A.; Diaz, R.; Biggs, D.; Stillion, R.; et al. Cardiac ferroportin regulates cellular iron homeostasis and is important for cardiac function. Proceedings of the National Academy of Sciences 2015, 112, 3164–3169, https://doi.org/10.1073/pnas.1422373112. [CrossRef]
- Bigorra Mir, M.; Charlebois, E.; Tsyplenkova, S.; Fillebeen, C.; Pantopoulos, K. Cardiac Hamp mRNA Is Predominantly Expressed in the Right Atrium and Does Not Respond to Iron. Int. J. Mol. Sci. 2023, 24, 5163. [CrossRef]
- Zhabyeyev, P.; Oudit, G.Y. Unravelling the molecular basis for cardiac iron metabolism and deficiency in heart failure. Eur Heart J 2017, 38, 373–375. https://doi.org/10.1093/eurheartj/ehw386. [CrossRef]
- Xu, W.; Barrientos, T.; Mao, L.; Rockman, H.A.; Sauve, A.A.; Andrews, N.C. Lethal Cardiomyopathy in Mice Lacking Transferrin Receptor in the Heart. Cell Rep 2015, 13, 533–545. https://doi.org/10.1016/j.celrep.2015.09.023. [CrossRef]
- Lakhal-Littleton, S.; Wolna, M.; Chung, Y.J.; Christian, H.C.; Heather, L.C.; Brescia, M.; Ball, V.; Diaz, R.; Santos, A.; Biggs, D.; et al. An essential cell-autonomous role for hepcidin in cardiac iron homeostasis. Elife 2016, 5. https://doi.org/10.7554/eLife.19804. [CrossRef]
- Wen, C.P.; Lee, J.H.; Tai, Y.P.; Wen, C.; Wu, S.B.; Tsai, M.K.; Hsieh, D.P.; Chiang, H.C.; Hsiung, C.A.; Hsu, C.Y.; et al. High serum iron is associated with increased cancer risk. Cancer Res 2014, 74, 6589–6597. https://doi.org/10.1158/0008-5472.CAN-14-0360. [CrossRef]
- Chang, V.C.; Cotterchio, M.; Khoo, E. Iron intake, body iron status, and risk of breast cancer: A systematic review and meta-analysis. BMC Cancer 2019, 19, 543. https://doi.org/10.1186/s12885-019-5642-0. [CrossRef]
- Ma, J.; Li, Q.; Fang, X.; Chen, L.; Qiang, Y.; Wang, J.; Wang, Q.; Min, J.; Zhang, S.; Wang, F. Increased total iron and zinc intake and lower heme iron intake reduce the risk of esophageal cancer: A dose-response meta-analysis. Nutr Res 2018, 59, 16–28. https://doi.org/10.1016/j.nutres.2018.07.007. [CrossRef]
- Ward, H.A.; Whitman, J.; Muller, D.C.; Johansson, M.; Jakszyn, P.; Weiderpass, E.; Palli, D.; Fanidi, A.; Vermeulen, R.; Tjonneland, A.; et al. Haem iron intake and risk of lung cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Eur J Clin Nutr 2019, 73, 1122–1132. https://doi.org/10.1038/s41430-018-0271-2. [CrossRef]
- Cao, H.; Wang, C.; Chai, R.; Dong, Q.; Tu, S. Iron intake, serum iron indices and risk of colorectal adenomas: A meta-analysis of observational studies. Eur J Cancer Care (Engl) 2017, 26. https://doi.org/10.1111/ecc.12486. [CrossRef]
- Qiao, L.; Feng, Y. Intakes of heme iron and zinc and colorectal cancer incidence: A meta-analysis of prospective studies. Cancer Causes Control 2013, 24, 1175–1183. https://doi.org/10.1007/s10552-013-0197-x. [CrossRef]
- Aglago, E.K.; Cross, A.J.; Riboli, E.; Fedirko, V.; Hughes, D.J.; Fournier, A.; Jakszyn, P.; Freisling, H.; Gunter, M.J.; Dahm, C.C.; et al. Dietary intake of total, heme and non-heme iron and the risk of colorectal cancer in a European prospective cohort study. Br. J. Cancer 2023, 10.1038/s41416-023-02164-7. https://doi.org/10.1038/s41416-023-02164-7. [CrossRef]
- Zacharski, L.R.; Chow, B.K.; Howes, P.S.; Shamayeva, G.; Baron, J.A.; Dalman, R.L.; Malenka, D.J.; Ozaki, C.K.; Lavori, P.W. Decreased cancer risk after iron reduction in patients with peripheral arterial disease: Results from a randomized trial. J Natl Cancer Inst 2008, 100, 996–1002. https://doi.org/10.1093/jnci/djn209. [CrossRef]
- Merk, K.; Mattsson, B.; Mattsson, A.; Holm, G.; Gullbring, B.; Bjorkholm, M. The Incidence of Cancer among Blood Donors. Int. J. Epidemiol. 1990, 19, 505–509. https://doi.org/10.1093/ije/19.3.505. [CrossRef]
- Gamage, S.M.K.; Lee, K.T.W.; Dissabandara, D.L.O.; Lam, A.K.-Y.; Gopalan, V. Dual role of heme iron in cancer; promotor of carcinogenesis and an inducer of tumour suppression. Exp. Mol. Pathol. 2021, 120, 104642. https://doi.org/10.1016/j.yexmp.2021.104642. [CrossRef]
- Torti, S.V.; Manz, D.H.; Paul, B.T.; Blanchette-Farra, N.; Torti, F.M. Iron and Cancer. Annu Rev Nutr 2018, 38, 97–125. https://doi.org/10.1146/annurev-nutr-082117-051732. [CrossRef]
- Pinnix, Z.K.; Miller, L.D.; Wang, W.; D’Agostino, R.; Kute, T.; Willingham, M.C.; Hatcher, H.; Tesfay, L.; Sui, G.; Di, X.; et al. Ferroportin and Iron Regulation in Breast Cancer Progression and Prognosis. Sci. Transl. Med. 2010, 2, 43ra56-43ra56, https://doi.org/10.1126/scitranslmed.3001127. [CrossRef]
- Basuli, D.; Tesfay, L.; Deng, Z.; Paul, B.; Yamamoto, Y.; Ning, G.; Xian, W.; McKeon, F.; Lynch, M.; Crum, C.P. Iron addiction: A novel therapeutic target in ovarian cancer. Oncogene 2017, 36, 4089–4099. [CrossRef]
- Tesfay, L.; Clausen, K.A.; Kim, J.W.; Hegde, P.; Wang, X.; Miller, L.D.; Deng, Z.; Blanchette, N.; Arvedson, T.; Miranti, C.K.; et al. Hepcidin Regulation in Prostate and Its Disruption in Prostate Cancer. Cancer Res. 2015, 75, 2254–2263. https://doi.org/10.1158/0008-5472.Can-14-2465. [CrossRef]
- Schonberg, David L.; Miller, Tyler E.; Wu, Q.; Flavahan, William A.; Das, Nupur K.; Hale, James S.; Hubert, Christopher G.; Mack, Stephen C.; Jarrar, Awad M.; Karl, Robert T.; et al. Preferential Iron Trafficking Characterizes Glioblastoma Stem-like Cells. Cancer Cell 2015, 28, 441–455. https://doi.org/10.1016/j.ccell.2015.09.002. [CrossRef]
- Jeong, S.M.; Hwang, S.; Seong, R.H. Transferrin receptor regulates pancreatic cancer growth by modulating mitochondrial respiration and ROS generation. Biochemical and Biophysical Research Communications 2016, 471, 373–379. https://doi.org/10.1016/j.bbrc.2016.02.023. [CrossRef]
- Ramírez-Carmona, W.; Díaz-Fabregat, B.; Yuri Yoshigae, A.; Musa de Aquino, A.; Scarano, W.R.; de Souza Castilho, A.C.; Avansini Marsicano, J.; Leal do Prado, R.; Pessan, J.P.; de Oliveira Mendes, L. Are Serum Ferritin Levels a Reliable Cancer Biomarker? A Systematic Review and Meta-Analysis. Nutrition and Cancer 2022, 74, 1917–1926. https://doi.org/10.1080/01635581.2021.1982996. [CrossRef]
- Radulescu, S.; Brookes, M.J.; Salgueiro, P.; Ridgway, R.A.; McGhee, E.; Anderson, K.; Ford, S.J.; Stones, D.H.; Iqbal, T.H.; Tselepis, C.; et al. Luminal iron levels govern intestinal tumorigenesis after apc loss in vivo. Cell Rep 2012, 2, 270–282, doi:S2211-1247(12)00199-4 [pii] https://doi.org/10.1016/j.celrep.2012.07.003. [CrossRef]
- Ijssennagger, N.; Rijnierse, A.; de Wit, N.; Jonker-Termont, D.; Dekker, J.; Muller, M.; van der Meer, R. Dietary haem stimulates epithelial cell turnover by downregulating feedback inhibitors of proliferation in murine colon. Gut 2012, 61, 1041–1049. https://doi.org/10.1136/gutjnl-2011-300239. gutjnl-2011-300239 [pii]. [CrossRef]
- Ijssennagger, N.; Rijnierse, A.; de Wit, N.J.; Boekschoten, M.V.; Dekker, J.; Schonewille, A.; Muller, M.; van der Meer, R. Dietary heme induces acute oxidative stress, but delayed cytotoxicity and compensatory hyperproliferation in mouse colon. Carcinogenesis 2013, 34, 1628–1635. https://doi.org/10.1093/carcin/bgt084. [CrossRef]
- Constante, M.; Fragoso, G.; Calve, A.; Samba-Mondonga, M.; Santos, M.M. Dietary Heme Induces Gut Dysbiosis, Aggravates Colitis, and Potentiates the Development of Adenomas in Mice. Front Microbiol 2017, 8, 1809. https://doi.org/10.3389/fmicb.2017.01809. [CrossRef]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in NAFLD. Nat Rev Gastroenterol Hepatol 2016, 13, 412–425. https://doi.org/10.1038/nrgastro.2016.85. [CrossRef]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Pedersen, H.K.; et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature 2015, 528, 262–266. https://doi.org/10.1038/nature15766. [CrossRef]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. https://doi.org/10.1016/j.immuni.2017.04.008. [CrossRef]
- Osborne, N.J.; Gurrin, L.C.; Allen, K.J.; Constantine, C.C.; Delatycki, M.B.; McLaren, C.E.; Gertig, D.M.; Anderson, G.J.; Southey, M.C.; Olynyk, J.K.; et al. HFE C282Y homozygotes are at increased risk of breast and colorectal cancer. Hepatology 2010, 51, 1311–1318. https://doi.org/10.1002/hep.23448. [CrossRef]
- Fargion, S.; Valenti, L.; Fracanzani, A.L. Role of iron in hepatocellular carcinoma. Clin Liver Dis (Hoboken) 2014, 3, 108–110. https://doi.org/10.1002/cld.350. [CrossRef]
- Finianos, A.; Matar, C.F.; Taher, A. Hepatocellular Carcinoma in beta-Thalassemia Patients: Review of the Literature with Molecular Insight into Liver Carcinogenesis. Int J Mol Sci 2018, 19. https://doi.org/10.3390/ijms19124070. [CrossRef]
- Allameh, A.; Hüttmann, N.; Charlebois, E.; Katsarou, A.; Gu, W.; Gkouvatsos, K.; Pasini, E.; Bhat, M.; Minic, Z.; Berezovski, M.; et al. Hemojuvelin deficiency promotes liver mitochondrial dysfunction and predisposes mice to hepatocellular carcinoma. Commun Biol 2022, 5, 153. https://doi.org/10.1038/s42003-022-03108-2. [CrossRef]
- Celis, A.I.; Relman, D.A.; Huang, K.C. The impact of iron and heme availability on the healthy human gut microbiome in vivo and in vitro. Cell Chemical Biology 2023, 30, 110–126.e113. https://doi.org/10.1016/j.chembiol.2022.12.001. [CrossRef]
- Mayneris-Perxachs, J.; Cardellini, M.; Hoyles, L.; Latorre, J.; Davato, F.; Moreno-Navarrete, J.M.; Arnoriaga-Rodríguez, M.; Serino, M.; Abbott, J.; Barton, R.H.; et al. Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome. Microbiome 2021, 9, 104. https://doi.org/10.1186/s40168-021-01052-7. [CrossRef]
- Jaeggi, T.; Kortman, G.A.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. https://doi.org/10.1136/gutjnl-2014-307720. [CrossRef]
- Gwamaka, M.; Kurtis, J.D.; Sorensen, B.E.; Holte, S.; Morrison, R.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. Iron deficiency protects against severe Plasmodium falciparum malaria and death in young children. Clin Infect Dis 2012, 54, 1137–1144. https://doi.org/10.1093/cid/cis010. [CrossRef]
- Lee, T.; Clavel, T.; Smirnov, K.; Schmidt, A.; Lagkouvardos, I.; Walker, A.; Lucio, M.; Michalke, B.; Schmitt-Kopplin, P.; Fedorak, R.; et al. Oral versus intravenous iron replacement therapy distinctly alters the gut microbiota and metabolome in patients with IBD. Gut 2017, 66, 863–871. https://doi.org/10.1136/gutjnl-2015-309940. [CrossRef]
- Nielsen, O.H.; Soendergaard, C.; Vikner, M.E.; Weiss, G. Rational Management of Iron-Deficiency Anaemia in Inflammatory Bowel Disease. Nutrients 2018, 10. https://doi.org/10.3390/nu10010082. [CrossRef]
- Constante, M.; Fragoso, G.; Calvé, A.; Samba-Mondonga, M.; Santos, M.M. Dietary Heme Induces Gut Dysbiosis, Aggravates Colitis, and Potentiates the Development of Adenomas in Mice. Front. Microbiol. 2017, 8. https://doi.org/10.3389/fmicb.2017.01809. [CrossRef]
- Mahalhal, A.; Williams, J.M.; Johnson, S.; Ellaby, N.; Duckworth, C.A.; Burkitt, M.D.; Liu, X.; Hold, G.L.; Campbell, B.J.; Pritchard, D.M.; et al. Oral iron exacerbates colitis and influences the intestinal microbiome. PLoS ONE 2018, 13, e0202460. https://doi.org/10.1371/journal.pone.0202460. [CrossRef]
- Mahalhal, A.; Burkitt, M.D.; Duckworth, C.A.; Hold, G.L.; Campbell, B.J.; Pritchard, D.M.; Probert, C.S. Long-Term Iron Deficiency and Dietary Iron Excess Exacerbate Acute Dextran Sodium Sulphate-Induced Colitis and Are Associated with Significant Dysbiosis. Int J Mol Sci 2021, 22. https://doi.org/10.3390/ijms22073646. [CrossRef]
- Ijssennagger, N.; Belzer, C.; Hooiveld, G.J.; Dekker, J.; van Mil, S.W.; Muller, M.; Kleerebezem, M.; van der Meer, R. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc Natl Acad Sci U S A 2015, 112, 10038–10043. https://doi.org/10.1073/pnas.1507645112. [CrossRef]
- Song, M.; Chan, A.T. Environmental Factors, Gut Microbiota, and Colorectal Cancer Prevention. Clin. Gastroenterol. Hepatol. 2019, 17, 275–289. https://doi.org/10.1016/j.cgh.2018.07.012. [CrossRef]
- Zakrzewski, M.; Wilkins, S.J.; Helman, S.L.; Brilli, E.; Tarantino, G.; Anderson, G.J.; Frazer, D.M. Supplementation with Sucrosomial(R) iron leads to favourable changes in the intestinal microbiome when compared to ferrous sulfate in mice. Biometals 2021, 10.1007/s10534-021-00348-3. https://doi.org/10.1007/s10534-021-00348-3. [CrossRef]
- Constante, M.; Fragoso, G.; Lupien-Meilleur, J.; Calve, A.; Santos, M.M. Iron Supplements Modulate Colon Microbiota Composition and Potentiate the Protective Effects of Probiotics in Dextran Sodium Sulfate-induced Colitis. Inflamm Bowel Dis 2017, 23, 753–766. https://doi.org/10.1097/MIB.0000000000001089. [CrossRef]
Figure 1.
Common causes and clinical complications of iron deficiency and iron overload.
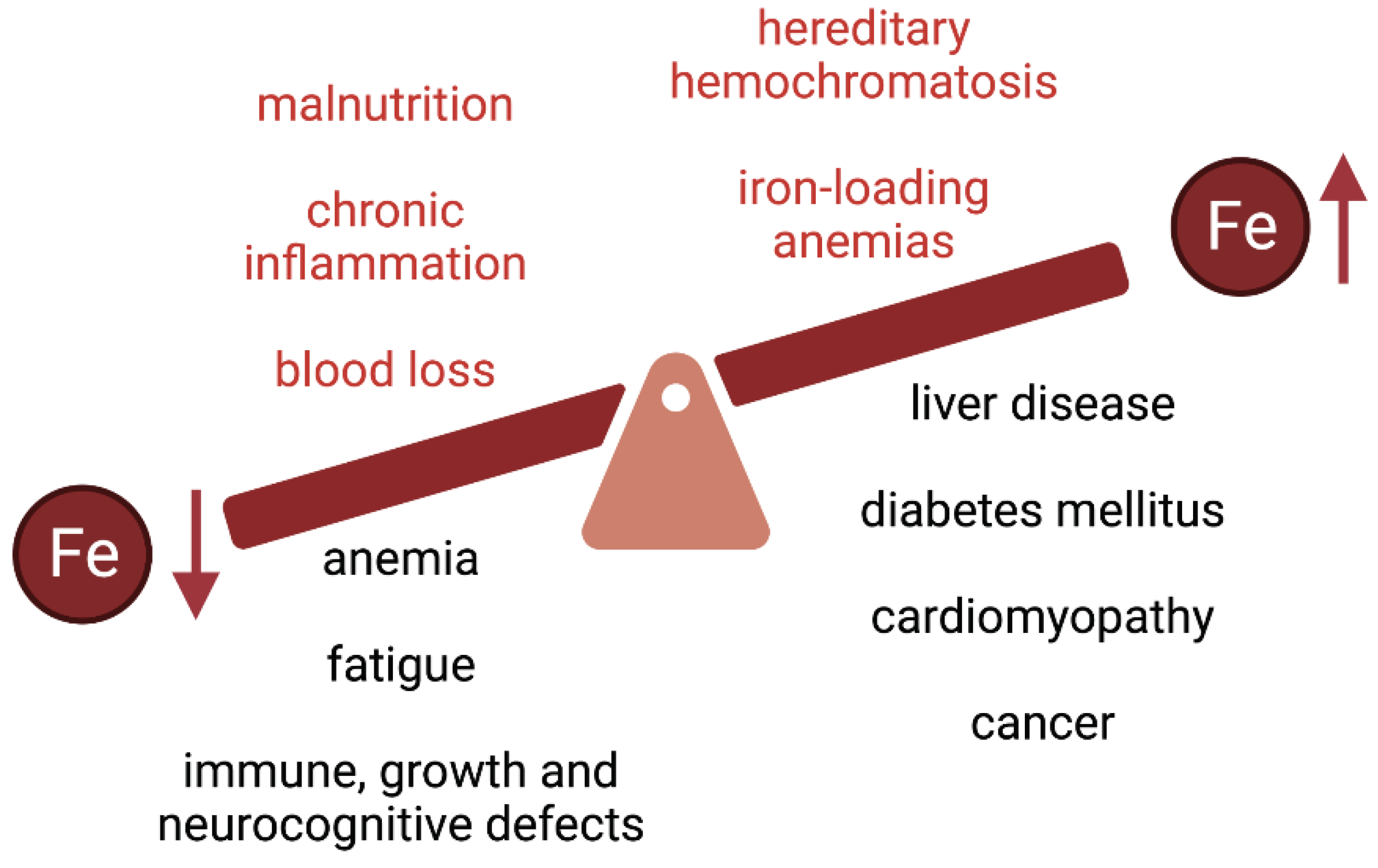
Figure 2.
Disease risks associated with enhanced dietary heme intake.
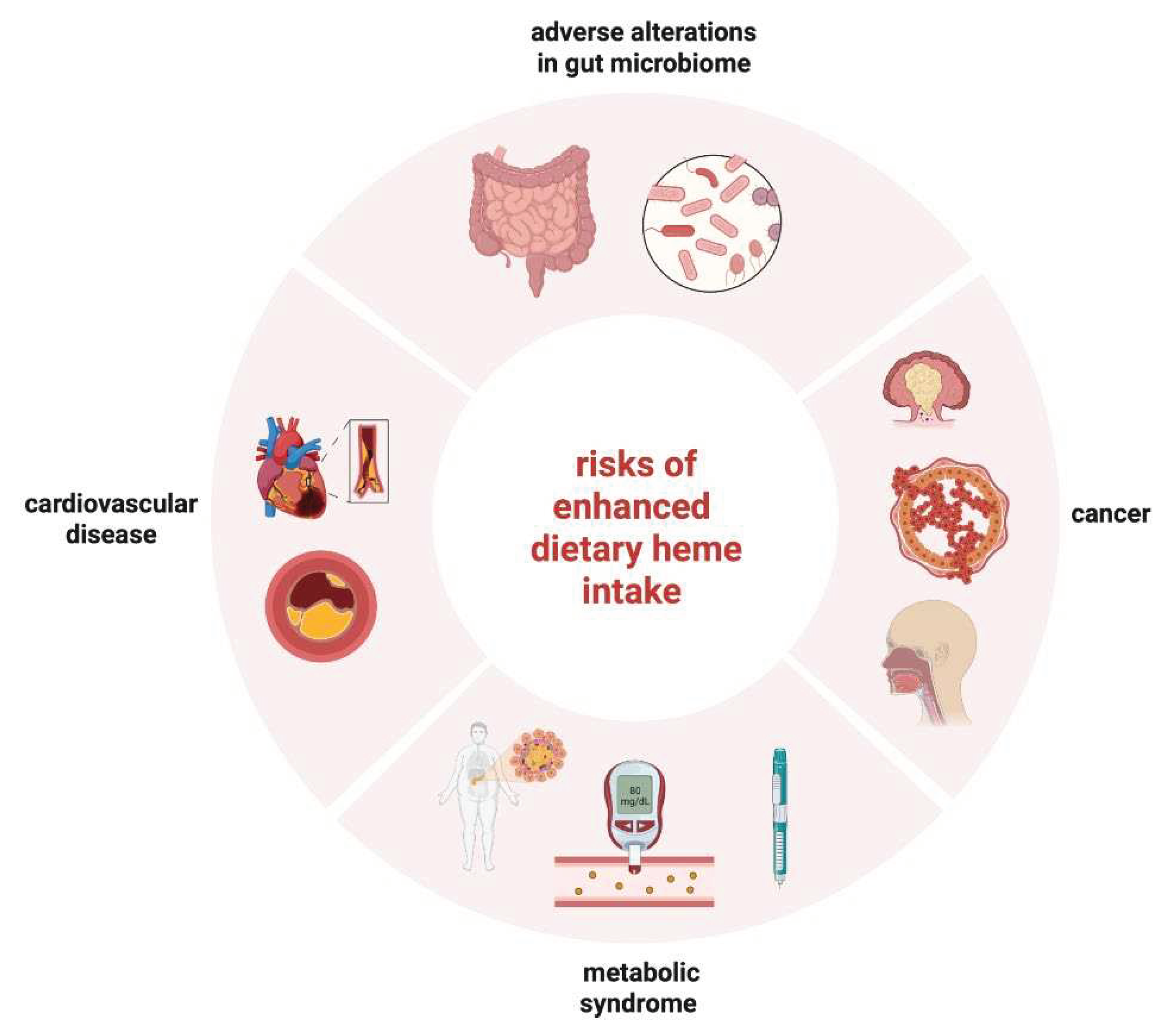

Table 1.
Pathologies with significant iron-related associations.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



