
Submitted:
25 April 2023
Posted:
26 April 2023
You are already at the latest version
Abstract
In the last decades, technological advances related to RNA manipulation enabled and expanded its application in vaccine development. This approach comprises synthetic single-stranded mRNA molecules that direct the translation of the antigen responsible for activating the desired immune response. The success of RNA vaccines depends on the delivery vehicle employed. Among the systems, yeasts emerge as a new approach to a natural delivery platform. The presence of β-glucans and mannans in its wall is responsible for the adjuvant action of this system. Yeasts are already employed to deliver protein antigens, with success and efficacy demonstrated through pre-clinical and clinical trials. Yeast β-glucan capsules, microparticles, and nanoparticles are capable of modulating host immune responses and have a high capacity to carry RNA and small molecules, with bioavailability upon oral immunization and with targeting to receptors present in anti-gen-presenting cells (APCs). Besides, yeasts are interesting vehicles for the protection and specific delivery of therapeutic vaccines based on shRNA or dsRNA. In this review, we present an overview of the attributes of yeast or its derivatives for the delivery of RNA-based vaccines, discussing their current challenges and prospects for using this promising strategy.
Keywords:
therapeutic vaccines
; mRNA vaccines
; antigenic delivery
; whole yeast vaccine
; delivery vehicle
1. Introduction
Vaccination is the most effective prophylaxis measure for infectious diseases [1]. According to the WHO, the eradication of diseases, such as poliomyelitis, and the reduction of mortality of others as tetanus, whooping cough, and influenza, was only achieved thanks to vaccines. However, there are still some obstacles, such as keeping up with the changes caused by genetic variants and generating more effective responses, in addition to the slow production of some types of immunogen [2].
Within this scenario, the mRNA-based vaccine production strategy proved to be a safe, fast, and effective alternative. Primarily developed decades ago, RNA vaccines took a long time to be widely employed due to inherent problems such as low stability, excessive immunostimulation, and low efficacy in some trials [3, 4]. However, with advances in sequence modifications, such as codon optimization, chemical modifications, and the use of carrier molecules, the possibility of its use in clinical applications has grown [5].
In parallel, the interest in more efficient delivery systems has also received attention. Several methods have already been adopted, such as lipid nanoparticles used in mRNA vaccines developed for SARS-CoV-2 [6, 7]. However, the application of microorganisms as delivery systems has been gaining prominence due to their ability to generate efficient immune responses, specific capture, and physical protection for the vaccine antigen [8]. In this context, yeasts such as Saccharomyces cerevisiae, Schizosaccharomyces pombe, and Pichia pastoris have been proposed as promising delivery systems for acid nucleic-based vaccines [9, 10]. In this review, we describe how yeasts can be used as an antigen vehicle to enhance the effectiveness of RNA vaccines, besides the main applications in immunotherapy approaches.
2. RNA Vaccines and Delivery Systems
Due to limitations such as stability, intrinsic toxicity, and poor distribution, the applicability of RNA vaccines was initially restricted compared to other platforms [4]. However, the application of chemical modifications, codon optimization, and integration of carrier molecules has broadened its usefulness and sparked significant interest in the production of vaccines against infectious [5], autoimmune [11], and neoplastic diseases [12] (Figure 1).
These vaccines comprise synthetic single-stranded mRNA molecules that direct the translation of the antigen responsible for activating the immune response. Like endogenous mRNA, the in vitro assembly includes one or more antigen-encoding ORFs, untranslated regions, a 3' poly-A tail, and a 5' methylated cap that prevents unintended immune responses and degradation [5, 13]. Most RNA vaccines include modified nucleosides to avoid recognition by pattern recognition receptors (PRRs) and ensure sufficient translation of the target antigen [14]. This adjustment is necessary since unmodified mRNA molecules are highly immunogenic and recognized by Toll-like receptors (such as TLR3, TLR7, TLR8, and TLR13), RIG-1, and MDA-5, able to instigate cellular and humoral responses [15]. However, they also lead to the production of immunostimulatory molecules such as IFN I and IFNα that can block the translation of the antigenic molecule [16]. Thus, current classifications include three main types of RNA vaccines: conventional (mRNA), which have no modifications and self-amplifying capacity; base-modified non-amplifying mRNA (bmRNA) vaccines and auto-amplifying mRNA (saRNA) vaccines [17].
To be expressed, RNA vaccines must enter the cytoplasm of antigen-presenting cells, where they are translated and processed for presentation via MHC and subsequent activation of CD8+ T cells [18]. On the other hand, synthesized proteins located in the extracellular milieu can be captured, degraded, and presented via MHC-II, causing the activation of CD4+ T cells critical for the activation of defense cells and antibody production [19]. The success of mRNA vaccines also depends on delivery techniques. The delivery vehicle must protect the RNA from possible digestion by ribonucleases and allow for effective uptake by the target cell, dissociation of genetic material from the delivery vehicle, and escape from endosomes [17, 20]. Furthermore, non-toxicity and immunostimulation are imperative for an ideal delivery vehicle [21].
Despite the preferential study by delivery particles of lipid and polymeric origin, new approaches involving natural delivery systems based on microorganisms emerge as novel systems to be explored [20, 22 -23]. Due to their composition, they can signalize efficient immune responses, promoting a specific uptake, transporting nucleic acids anchored superficially or in their interior, and expanding the range of administration methods [8]. Among microorganisms, yeasts have application value as biofactories of therapeutic proteins and carriers of biological molecules [8-9]. However, despite its potential, there is still a need for a better understanding of its applicability in the delivery of mRNA vaccines, as well as a better evaluation of the mechanisms of uptake and transport.
3. Yeasts as Vaccine Carriers: Characteristics and Immunological Aspects
Besides being recognized biotechnological platforms for the obtention of immunobiological products, yeasts such as Saccharomyces cerevisiae and Pichia pastoris have been proposed as delivery systems for protein antigens or nucleic acids in the development of prophylactic and therapeutic vaccines [24]. This approach, called whole yeast vaccines, allows for better delivery and antigen presentation to the immune system through intracellular delivery or even anchoring recombinant proteins on the yeast cell surface [9 -10, 25].
An advantage of the use of yeasts as vaccine vehicles is their immunostimulatory property due to the presence of β-1,3-glucans and mannoproteins in their wall, which, when detected by Pattern Recognition Receptors (RRPs), promote the release of cytokines, chemokines, and eicosanoids that modulate the inflammatory response in addition to leading to the development of specific adaptive immune responses against the pathogen [26-28]. As with mannoproteins, the response induced by β-1,3-glucans tends towards a pro-inflammatory Th1 profile. Besides, these particles can promote the recruitment of neutrophils and specific antigenic recognition by APCs when interacting with receptors such as Dectin-1 [29]. The exposure of β-1,3-glucan can be favored by heating the yeast at 60°C for at least 1 hour, expanding the adjuvant property of this microorganism [30] (Figure 2A).
In dendritic cells (DCs), the capture and phagocytosis of yeasts are mediated by the presence of the Dectin-1 receptor that recognizes the β-1,3-glucans [31 - 32] (Figure 2B). Activation of DCs by yeast promotes the secretion of inflammatory cytokines such as TNF-α, IL-6, IL-8, IL-1β, IL-12, IL-10, IL-23, and IL-27, which polarize T cells for Th1 and Th17 [33]. This polarization is particularly interesting in the context of immunotherapies against viral infections or anticancer, reinforcing the response induced by the vaccine antigen carried by the yeast.
4. Biodelivery of mRNA Vaccines by Yeasts
The ability to use yeast for gene delivery has been explored in different studies for the therapy of different types of cancer, as immunomodulators, and for the treatment or prophylaxis of infectious diseases [34 - 36]. In this context, its use as a vehicle for RNA molecules is based on the intracellular transport of these sequences using vectors with components that allow the transport and adequate delivery of genetic material to target cells or tissues.
In general, plasmids with promoters derived from yeast are chosen to allow transcription of the gene of interest before or after phagocytosis by APCs [9]. Breining et al. (2013) tested the pPGK vector whose promoter belongs to the phosphoglycerate kinase (PGK) pathway, being recognized as strong and constitutive, mediating transcription before the yeasts are phagocytosed [37]. On the other hand, another study revealed the transcription efficiency after phagocytosis from the use of inducible promoters, such as malate synthase (MLS1) and isocitrate lyase (ICL1) [35].
Another component introduced upstream of the gene in the assembly of mRNA vaccines comprises the insertion of the IRES (Internal Ribosomal Entry Site) sequence derived from EMCV (encephalomyocarditis virus). This region prevents antigen translation in yeast cells and increases translational levels in host cells, mainly in the macrophages [38]. In the studies by Walch and collaborators (2012), in vitro, assays were performed to prove that the observed expression refers to the translation of the model antigen (carrying IC21 macrophages) in mammalian cells and the absence of translational activity by yeast [39].
One of the main applications of mRNA delivery by yeast has been immunomodulation, as demonstrated by the study by Seif et al. (2017). Polarization of tumor-associated macrophages (TAMs) to an M1 expression profile has been used in several cancer immunotherapy strategies [40 - 41]. The carrying of genes encoding pro-inflammatory cytokines, such as TNF-α, by S. cerevisiae may be a useful tool for altering the profile of these TAMs within the context of the tumor microenvironment [35]. In addition to the isolated use of RNA, new studies have emerged applying yeast to carry multiple strategies in the same expression cassette. Zhang et al. (2023) showed the feasibility of combining DNA-RNA sequences with hemagglutinin as a target gene (rH9-DNA-RNA) in the same plasmid and using S. cerevisiae as a vaccine vehicle. The combined nucleic acids were effectively expressed and successfully delivered into the intestinal cells of orally immunized animals [36].
5. Capsules, Microparticles, and Nanoparticles of Yeast β-Glucans for RNA Delivery
The yeast-based biodelivery goes beyond using the whole cell as a vaccine vehicle. Yeast cell particles (YCP) obtained from the mannoprotein layer removal have also been used for this purpose [42 - 44] (Figure 3). These particles can vary in size and be classified as capsules, microparticles, or nanoparticles depending on the treatment conditions used to allow access to the β-glucans layer [45 -47]. The association with other immunostimulants such as aluminum salts [48] and formylate [49] or the introduction of chemical modifications can further improve the function of glucan particles (GPs) as a vaccine adjuvant, offering an interesting method to trigger the vaccine immune responses [50].
The capsules can be recognized as a pathogen-associated molecular pattern (PAMP), induce, and modulate immune responses, and can carry proteins, DNA, RNA, and small molecules. Antigen delivery based on these particles allows oral administration and promotes antigenic targeting to APCs due to the presence of β-1,3-D-glucan receptors in these immune cells [49, 51 - 53]. These receptors include Dectin-1, complement receptor 3 (CR3), and Toll-like receptors (TLRs). Once phagocytosed, GPs can mediate the stimulation of components of innate immunity, such as macrophages, dendritic cells, neutrophils, and NK cells, and lead to the secretion of pro-inflammatory cytokines [54 - 55] (Figure 4).
GPs are 2-4 µm hollow and porous structures that allow the encapsulation of a high load of antigens through central polyplexes and synthetic structures that help maintain the conformation and integrity of the particles for targeted delivery of antigens to APCs [50]. GPs as delivery systems for nucleic acids have demonstrated protection and ease of oral delivery targeting specific components of mucosal immunity, besides being able to promote systemic immune responses [52]. Vaccine formulations based on GP nanoparticles (NPs) offer significant potential for new vaccination strategies due to their unique physicochemical properties and the size of these structures that facilitate their uptake by dendritic cells [55]. The carriage of siRNA by GP NPs demonstrates high intracellular delivery capacity, even when compared to commercial carriers, such as Lipofectamine, and has low toxicity [56]. These NPs can stably and efficiently carry antigens, mediate the delivery of these antigens, and enhance maturation, stimulation of DCs, and the secretion of pro-inflammatory cytokines [57].
GPs lack reproduction activity and are smaller than whole yeast cells [55]. The morphology of these particles, especially nanoparticles smaller than 50 nm, facilitates their entry through lymphatic vessels, being more easily absorbed by DCs, while whole yeasts are mainly delivered to lymph nodes in a cell-mediated manner [55, 58]. Furthermore, vaccines that use whole organisms contain a wide range of antigens and, although they are relatively easy to manufacture, may present concerns about reactogenicity, autoimmunity, and possible infection in immunosuppressed populations when administered [54 - 55, 59]. The use of particles derived from the cell wall of yeasts has shown to be an option to circumvent any adverse effects arising from the administration of whole microorganisms. However, more comprehensive in vitro and in vivo evaluations are needed to address the immunotoxicity and bioavailability of these vaccine carriers.
6. Oral Yeast-Based Immunization for RNA Vaccines
When evaluating the effectiveness of a vaccine strategy, the route of administration plays an important role in immunogenicity and is a determining factor for effective antigen presentation, potency, and effective response [60]. Nucleic acid strategies, specifically RNA vaccines, need to be carried by structures that keep the antigen safe and stable until it reaches target tissues and cells, in addition to allowing recognition and robust response by the immune system [9].
On the other hand, oral administration allows, besides systemic immunostimulation, mediation on mucosal surfaces leading to an adaptive response that combines the action of IgA antibodies and resident memory T cells [61]. This last feature results in specific and efficient protection since most pathogens have mucosal areas as their entry site. In addition, the oral route is preferable due to its low cost-effectiveness of production and ease of distribution [62]. Despite this, there are some challenges due to local natural barriers as those present in the gastrointestinal tract, which hinder access and delivery of vaccine components, leading to low antigen availability. Some factors include physical structures, such as the mucus and glycocalyx layer, in addition to the low pH and presence of proteases and nucleases on the mucosal surface that cause degradation of nucleic acids and antigenic proteins present in vaccines [61, 63 - 64].
Thus, the development and use of approaches that improve the delivery of vaccines by the oral route have been the subject of studies, which generally point to the need for adjuvants and delivery systems. Alternatively, the yeast-based distribution approach, in different forms such as whole yeast, capsules, micro, and nanoparticles, is an excellent option, as seen in previous sections of this review, enabling this type of administration for RNA vaccines (Table 1). A review elaborated by Ivanova (2021) describes the effectiveness of yeast-based systems for oral gene delivery and how they have been explored in vaccine strategies and immunotherapy [62]. A relevant characteristic of this system is the specific antigen targeting through the intestinal mononuclear phagocytic system that forwards to the systemic circulation or sites of inflammation or neoplasia.
7. Delivery of RNA Interference (RNAi)
In addition to mRNA-based vaccines, different experimental models have demonstrated that it is possible to silence or modulate the expression of target genes in a specific way in eukaryotic cells through dsRNA (double-stranded RNA) or shRNA (short hairpin RNA) molecules [67 - 68]. These interference RNAs make up a regulatory pathway that leads to the silencing of genes due to sequence homology and complementarity, associated with an enzymatic complex that promotes target mRNA degradation [69 - 70].
In recent years, the understanding of RNAi has enabled its use as a gene therapy tool against infections caused by different viruses such as coronaviruses associated with Severe Acute Respiratory Syndrome, Influenza, Hepatitis B virus, and Dengue virus [711 - 74]. In addition to controlling these infections in their acute phase, this gene-silencing mechanism has also been used in therapeutic strategies aimed at cancer and chronic inflammatory diseases [75 - 76].
However, the success of this strategy involves both the effectiveness of silencing and the non-occurrence of toxic effects. Thus, some adaptations are performed, including chemical modifications and changes in size, charge, and hydrophilicity that can interfere with the cellular uptake and biodistribution of RNAi molecules [77]. The administration method and the particle size of the vehicle influence the bioavailability and biodistribution of the loaded RNAi.
In this sense, the efficiency of this approach also depends on the use of efficient delivery systems [66, 77]. Liposomes, bacteria, and viruses are examples of platforms used as vehicles for interfering RNAs, as well as already used for DNA and mRNA vaccines [78 - 80]. However, some limitations indicate the need to seek alternatives to these approaches. Among these limitations are certain restrictions on the carrying capacity, besides issues related to safety and induction of immune tolerance or neutralization of the vaccine vector [81].
In contrast, the yeasts in their various presentations (whole cell, microcapsules, capsules) are feasible vehicles for the protection and specific delivery of therapeutic vaccines based on shRNA or dsRNA [66, 81 - 82]. Antigen delivery remains efficient even if the yeasts undergo heat treatment to eliminate their reproductive activity and are freeze-dried and processed in the form of capsules or tablets [83, 84]. The therapeutic interfering RNA can be eliminated from the body in hours or days, even if it is nanoparticulate. The reduced bioavailability time may be insufficient for the immunotherapeutic product not to reach the target cells or tissues [77]. On the other hand, Zhang et al. (2014) analyzed the immune response of mice with CD40 shRNA carried by S. cerevisiae 50 days after oral administration, observing the continuity of the effects of the CD40 receptor expression silencing about the set of cytokines present in the blood and from the extraction of proteins and dendritic cells from the intestine of immunized animals [66]. Although this study was carried out in an animal model, it shows promising effects regarding the duration of the immune response by the proposed therapy, which may be evaluated in humans.
After oral administration, the yeasts used in vaccine preparations are phagocytosed by macrophages and dendritic cells in the gut-associated lymphatic tissues (GALT). These phagocytic cells that act as professional antigen presenters can migrate and infiltrate other lymphatic tissues and induce specific responses even in sites distant from the point of administration [47]. This mechanism was observed by Zhang et al. (2020), that evaluated the delivery of an shRNA for silencing the IL-1β cytokine as a therapeutic strategy for post-traumatic osteoarthritis. In this study, to access the eventual tissue-specific migration of phagocytosed yeasts, yeast cells were labeled with near-infrared fluorescence, which allows the in vivo evaluation of the delivery of biomolecules to verify the specificity of this delivery and the induction of immunological response at the target sites. Such marking allowed observing the presence of macrophages carrying the recombinant yeasts in the joint and knee regions of the studied mice [47, 85].
As already mentioned, the size of the yeast and the binding to specific receptors such as Dectin-1 and CD206 favor the process of phagocytosis by antigen-presenting cells [31]. These immune cells are appropriate targets for RNAi-based therapies as they mediate the chronic inflammatory response caused by certain clinical conditions and autoimmune diseases [86]. This type of specificity is also essential to minimize toxic effects that could result from silencing in inappropriate cells or tissues.
In therapies based on RNAi transport by yeast, the main targets explored for silencing and immunomodulation are cytokines that play an effector role in the differentiation or maturation of APCs (e.g. IL-1β and IL-21), cellular receptors (e.g. CD40), repressor molecules (e.g. indoleamine 2, 3-dioxygenase - IDO), or even specific antigens of infectious agents [47, 66, 53, 87]. Zakria et al. (2019) developed an interesting study using S. cerevisiae as a vehicle for a vector designed for a DNA vaccine encoding the myostatin gene and for delivery of an IL-21 shRNA vaccine as therapeutic targets. Studies like this open the way for improving vaccines with more than one type of vaccine antigen that can complement or amplify the desired immune response [87].
In addition to selecting therapeutic targets, it is necessary to consider how to construct the expression cassettes to be inserted into the yeast. Species like S. cerevisiae have a vast set of expression vectors that can be adapted for this type of application. The main components of these cassettes are the promoter, the selection mark, and eventual integration sites; they are liable to optimization and vary according to the strategy (Figure 5). Regarding transcriptional control, promoters functional in mammalian cells as hU6 [66] or active only in yeast as GDP1 (constitutive) and Gal1 (inducible) can be used (Duman-Scheel, 2019). When using a promoter such as hU6, transcription must occur only after yeast phagocytosis and the release of its intracellular content where are the recombinant plasmids.
On the other hand, when are adopted native yeast promoters, it is expected the transcription and accumulation of RNAi in the yeast cell. This is feasible due to the absence of RNAi processing machinery in yeast (they do not express Dicer and Argonauta), allowing the accumulation of bioengineered RNAi molecules without inducing degradation [81, 83]. It is also important to point out that the type of promoter chosen (inducible versus constitutive) can also impact the scaling of cultures to prepare vaccine doses. Induction of gene expression by adding certain compounds can be an unfeasible process to produce yeast-based vaccines on an industrial scale [24, 81].
Concerning the construction of vectors used for RNAi insertion into yeast cells, it is preferable to use auxotrophic selection marks such as URA3/TRP1 that minimize the use of antibiotics and the possible horizontal transfer of resistance genes. Both episomal and integrative vectors have been used, and integration has been seen as a way to ensure more stable delivery. However, how this stability and the number of RNAi copies per cell can influence the effect or duration of silencing remains open [81, 83].
8. Conclusions
The development of RNA vaccines have gained a greater dimension due to the addition of optimizations in its structure and the discovery of new carriers capable of protecting the molecule, besides allowing specific uptake of the antigen to the target cells, in addition to acting as adjuvants or immunomodulators. Yeast cells or their derivatives (β-glucan particles) for RNA delivery present the potential for immunomodulation within therapeutic strategies. However, it is important not to disregard the immunostimulatory properties inherent to yeasts due to their cell wall components, which can induce or modulate immune responses. There are some points of study and investigation, like the yeast species that can be used (beyond S. cerevisiae), technical aspects, and the transposition of the results obtained in vitro and an animal model to the clinic. Such improvements are essential for vaccines, prophylactic or therapeutic, using yeasts to reach the pharmaceutical market.
Author Contributions
Conceptualization; data curation; writing—review and editing, Anna Jéssica Duarte Silva. Writing—original draft preparation, Mylenna Máyra Gois de Sousa, Larissa Silva de Macêdo, Pedro Luiz de França Neto, Ingrid Andressa de Moura, Benigno Cristofer Flores Espinoza, Maria Da Conceição Viana Invenção, Samara Sousa de Pinho, Marco Antonio Turiah Machado da Gama. Supervision; project administration; funding acquisition, Antonio Carlos de Freitas. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by FACEPE/PPSUS2020, grant number APQ-0260-2.02/20; CNPq Bolsa Produtividade, grant number 308489/2019-5; CAPES – Epidemias, grant number 88887.507421/ 2020-00; CNPq/MCTI/CT-BIOTEC Nº 31/2022, grant number 440315/2022-0 and Programa de Apoio a Fixação de Jovens Doutores em Pernambuco APQ-1482-2.02/22.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.B.; Ovsyannikova, I.G.; Palese, P.; Poland, G.A. Current Challenges in Vaccinology. Front. Immunol. 2020, 11, 1181. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Lagniton, P.N.P.; Liu, Y.; Xu, R.-H. mRNA vaccines for COVID-19: What, why and how. Int. J. Biol. Sci. 2021, 17, 1446–1460. [Google Scholar] [CrossRef] [PubMed]
- Aljabali, A.A.A.; Bashatwah, R.M.; Obeid, M.A.; Mishra, V.; Mishra, Y.; Serrano-Aroca, Á.; Lundstrom, K.; Tambuwala, M.M. Current state of, prospects for, and obstacles to mRNA vaccine development. Drug Discovery Today. 2023, 28, 103458. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-Assembled MRNA Vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112. [Google Scholar] [CrossRef]
- Kauffman, K.J.; Webber, M.J.; Anderson, D.G. Materials for Non-Viral Intracellular Delivery of Messenger RNA Therapeutics. J. Control. Release 2016, 240, 227–234. [Google Scholar] [CrossRef]
- de Moura, I.A.; Silva, A.J.D.; de Macêdo, L.S.; Invenção M da, C.V.; de Sousa, M.M.G.; de Freitas, A.C. Enhancing the Effect of Nucleic Acid Vaccines in the Treatment of HPV-Related Cancers: An Overview of Delivery Systems. Pathogens 2022, 11, 1444. [Google Scholar] [CrossRef]
- Silva, A.J.D.; de Macêdo, L.S.; Leal, L.R.S.; de Jesus, A.L.S.; Freitas, A.C. Yeasts as a promising delivery platform for DNA and RNA vaccines. FEMS Yeast Res. 2021. 21, foab018. [CrossRef]
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. 2019, 19, foz007. [Google Scholar] [CrossRef]
- Krienke, C.; Kolb, L.; Diken, E.; Streuber, M.; Kirchhoff, S.; Bukur, T.; Akilli-Öztürk, Ö.; Kranz, L.M.; Berger, H.; Petschenka, J.; Diken, M.; et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science 2021, 371, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Zhang, Y.; Huang, L. MRNA vaccine for cancer immunotherapy. Mol. Cancer 2021, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- Schlake, T.; Thess, A.; Fotin-Mleczek, M.; Kallen, K.-J. Developing mRNA-vaccine technologies. RNA Biol. 2012, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. MRNA Vaccine Era-Mechanisms, Drug Platform and Clinical Prospection. Int. J. Mol. Sci. 2020, 21, 6582. [Google Scholar] [CrossRef] [PubMed]
- Freund, I.; Eigenbrod, T.; Helm, M.; Dalpke, A. RNA Modifications Modulate Activation of Innate Toll-Like Receptors. Genes 2019, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, S.; Azizian, K.T.; Haque, A.K.M.A.; Henderson, J.M.; Hendel, A.; Shore, S.; Antony, J.S.; Hogrefe, R.I.; Kormann, M.S.D.; Porteus, M.H.; et al. Uridine Depletion and Chemical Modification Increase Cas9 mRNA Activity and Reduce Immunogenicity without HPLC Purification. Mol. Ther. Nucleic Acids 2018. 12, 530–542. [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in mRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Rijkers, G.T.; Weterings, N.; Obregon-Henao, A.; Lepolder, M.; Dutt, T.S.; van Overveld, F.J.; Henao-Tamayo, M. Antigen Presentation of mRNA-Based and Virus-Vectored SARS-CoV-2 Vaccines. Vaccines 2021, 9, 848. [Google Scholar] [CrossRef]
- Matsumura, T.; Takano, T.; Takahashi, Y. Immune responses related to the immunogenicity and reactogenicity of COVID-19 mRNA vaccines. Int. Immunol. 2022, dxac064. [Google Scholar] [CrossRef]
- Ramachandran, S.; Satapathy, S.R.; Dutta, T. Delivery Strategies for mRNA Vaccines. Pharm. Med. 2022, 36, 11–20. [Google Scholar] [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef]
- Eygeris, Y.; Gupta, M.; Kim, J.; Sahay, G. of Lipid Nanoparticles for RNA Delivery. Acc. Chem. Res. 2022, 55, 2–12. [Google Scholar] [CrossRef]
- Shende, P.; Basarkar, V. Recent trends and advances in microbe-based drug delivery systems. DARU J. Pharm. Sci. 2019, 27, 799–809. [Google Scholar] [CrossRef]
- Silva, A.J.D.; Rocha CK da, S.; de Freitas, A.C. Standardization and Key Aspects of the Development of Whole Yeast Cell Vaccines. Pharmaceutics 2022, 14, 2792. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Heterologous Protein Expression in Pichia pastoris : Latest Research Progress and Applications. ChemBioChem 2018, 19, 7–21. [Google Scholar] [CrossRef]
- Bal, J.; Luong, N.N.; Park, J.; Song, K.D.; Jang, Y.S.; Kim, D.H.l. Comparative immunogenicity of preparations of yeast-derived dengue oral vaccine candidate. Microb. Cell Fact. 2018, 17, 1–14. [Google Scholar] [CrossRef]
- Salazar, F.; Brown, G.D. Antifungal Innate Immunity: A Perspective from the Last 10 Years. J. Innate Immun. 2018, 10, 373–397. [Google Scholar] [CrossRef]
- Brown, G.D. Innate antifungal immunity: The key role of phagocytes. Annu. Rev. Immunol. 2011, 29, 1–21. [Google Scholar] [CrossRef]
- Erwig, L.P.; Gow, N.A. Interactions of fungal pathogens with phagocytes. Nat. Rev. Microbiol. 2016, 14, 163–176. [Google Scholar] [CrossRef]
- Bazan, S.B. , Breinig, T., Schmitt, M.J., Breinig, F. Heat treatment improves antigen-specific T cell activation after protein delivery by several but not all yeast genera. Vaccine 2014, 32, 2591–2598. [Google Scholar] [CrossRef]
- Bazan, S.B.; Geginat, G.; Breinig, T.; Schmitt, M.J.; Breinig, F. Uptake of various yeast genera by antigen-presenting cells and influence of subcellular antigen localization on the activation of ovalbumin-specific CD8 T lymphocytes. Vaccine 2011, 29, 8165–8173. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Bazan, S.B.; Walch-Rückheim, B.; Schmitt, M.J.; Breinig, F. Maturation and cytokine pattern of human dendritic cells in response to different yeasts. Med. Microbiol. Immunol. 2018, 207, 75–81. [Google Scholar] [CrossRef]
- Bilusic, M.; Heery, C.R.; Arlen, P.M.; Rauckhorst, M.; Apelian, D.; Tsang, K.Y.; Tucker, J.A.; Jochems, C.; Schlom, J.; Gulley, J.L.; et al. Phase I trial of a recombinant yeast-CEA vaccine (GI-6207) in adults with metastatic CEA-expressing carcinoma. Cancer Immunol. Immunother. 2014, 63, 225–234. [Google Scholar] [CrossRef]
- Seif, M.; Hoppstädter, J.; Breinig, F.; Kiemer, A.K. Yeast-mediated mRNA delivery polarizes immuno-suppressive macrophages towards an immuno-stimulatory phenotype. Eur. J. Pharm. Biopharm. 2017, 117, 1–13. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, R.; Zhang, H.; Sun, R.; Li, S.; Xia, C.; Li, Z.; Zhang, L.; Guo, Y.; Huang, J. Recombinant hemagglutinin protein and DNA-RNA-combined nucleic acid vaccines harbored by yeast elicit protective immunity against H9N2 avian influenza infection. Poultry Sci. 2023, 102, 102662. [Google Scholar] [CrossRef]
- Breinig, F.; Breinig, T.; Schmitt, M.J. mRNA Delivery to Human Dendritic Cells by Recombinant Yeast and Activation of Antigen-Specific Memory T Cells, in: Rabinovich, P.M. (Ed.), Synthetic Messenger RNA and Cell Metabolism Modulation, Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; pp. 163–184. [Google Scholar] [CrossRef]
- Evstafieva, A.G.; Beletsky, A.V.; Borovjagin, A.V.; Bogdanov, A.A. Internal Ribosome Entry Site Ofencephalomyocarditis Virus RNA Is Unable to Direct Translation in Saccharomyces Cerevisiae. FEBS Lett. 1993, 335, 273–276. [Google Scholar] [CrossRef]
- Walch-Rückheim, B.; Kiefer, R.; Geginat, G.; Schmitt, M.J.; Breinig, F. Coexpression of Human Perforin Improves Yeast-Mediated Delivery of DNA and MRNA to Mammalian Antigen-Presenting Cells. Gene Ther. 2016, 23, 103–107. [Google Scholar] [CrossRef]
- He, L.; Bai, Y.; Xia, L.; Pan, J.; Sun, X.; Zhu, Z.; Ding, J.; Qi, C.; Tang, C. Oral Administration of a Whole Glucan Particle (WGP)-Based Therapeutic Cancer Vaccine Targeting Macrophages Inhibits Tumor Growth. Cancer Immunol. Immunother. 2022, 71, 2007–2028. [Google Scholar] [CrossRef]
- Xu, T.; Yu, S.; Zhang, J.; Wu, S. Dysregulated Tumor-Associated Macrophages in Carcinogenesis, Progression and Targeted Therapy of Gynecological and Breast Cancers. J. Hematol. Oncol. 2021, 14, 181. [Google Scholar] [CrossRef]
- Berner, V.K.; Sura, M.E.; Hunter, K.W. Conjugation of Protein Antigen to Microparticulate β-Glucan from Saccharomyces Cerevisiae: A New Adjuvant for Intradermal and Oral Immunizations. Appl. Microbiol. Biotechnol. 2008, 80, 1053–1061. [Google Scholar] [CrossRef]
- De Smet, R.; Allais, L.; Cuvelier, C.A. Recent Advances in Oral Vaccine Development: Yeast-Derived β-Glucan Particles. Human. Vaccines Immunother. 2014, 10, 1309–1318. [Google Scholar] [CrossRef]
- Tipper, D.J.; Szomolanyi-Tsuda, E. Scaffolded Antigens in Yeast Cell Particle Vaccines Provide Protection against Systemic Polyoma Virus Infection. J. Immunol. Res. 2016, 2016, 1–15. [Google Scholar] [CrossRef]
- Dinarvand, R.; Cesar De Morais, P.; D’Emanuele, A. Nanoparticles for Targeted Delivery of Active Agents against Tumor Cells. J. Drug Deliv. 2012, 2012, 1–2. [Google Scholar] [CrossRef]
- Zhou, X.; Ling, K.; Liu, M.; Zhang, X.; Ding, J.; Dong, Y.; Liang, Z.; Li, J.; Zhang, J. Targeted Delivery of Cisplatin-Derived Nanoprecursors via a Biomimetic Yeast Microcapsule for Tumor Therapy by the Oral Route. Theranostics 2019, 9, 6568–6586. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, H.; Feng, M.; Zhang, W.; Li, Y. Yeast Microcapsule-Mediated Oral Delivery of IL-1β ShRNA for Post-Traumatic Osteoarthritis Therapy. Mol. Ther. - Nucleic Acids 2021, 23, 336–346. [Google Scholar] [CrossRef]
- Liu, H.; Meng, Z.; Wang, H.; Zhang, S.; Huang, Z.; Geng, X.; Guo, R.; Wu, Z.; Hong, Z. Robust Immune Responses Elicited by a Hybrid Adjuvant Based on β-Glucan Particles from Yeast for the Hepatitis B Vaccine. ACS Appl. Bio Mater. 2021, 4, 3614–3622. [Google Scholar] [CrossRef]
- Xu, Y.; Liang, M.; Huang, J.; Fan, Y.; Long, H.; Chen, Q.; Ren, Z.; Wu, C.; Wang, Y. Single-Helical Formyl β-Glucan Effectively Deliver CpG DNA with Poly(DA) to Macrophages for Enhanced Vaccine Effects. Int. J. Biol. Macromol. 2022, 223, 67–76. [Google Scholar] [CrossRef]
- Soto, E.R.; Caras, A.C.; Kut, L.C.; Castle, M.K.; Ostroff, G.R. Glucan Particles for Macrophage Targeted Delivery of Nanoparticles. J. Drug Deliv. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Muta, T. Molecular Basis for Invertebrate Innate Immune Recognition of (1→3)-β- D-Glucan as A Pathogen-Associated Molecular Pattern. CPD 2006, 12, 4155–4161. [Google Scholar] [CrossRef]
- Soto, E.R.; Ostroff, G.R. Characterization of Multilayered Nanoparticles Encapsulated in Yeast Cell Wall Particles for DNA Delivery. Bioconjugate Chem. 2008, 19, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Lu, S.; Zhang, L.; Ji, M.; Liu, S.; Wang, S.; Liu, R. An Indoleamine 2, 3-Dioxygenase SiRNA Nanoparticle-Coated and Trp2-Displayed Recombinant Yeast Vaccine Inhibits Melanoma Tumor Growth in Mice. J. Control. Release 2018, 273, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Specht, C.A.; Levitz, S.M. Robust Stimulation of Humoral and Cellular Immune Responses Following Vaccination with Antigen-Loaded β-Glucan Particles. mBio 2010, 1, e00164–10. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ma, Q.; Zhang, Y.; Fei, Z.; Sun, Y.; Fan, Q.; Liu, B.; Bai, J.; Yu, Y.; Chu, J.; et al. Yeast-Derived Nanoparticles Remodel the Immunosuppressive Microenvironment in Tumor and Tumor-Draining Lymph Nodes to Suppress Tumor Growth. Nat. Commun. 2022, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Min, D.; Choi, Y.; Yoon, S.; Jang, J.; Hwang, J.; Jeon, H.; Cho, Y.W.; Choi, J. Self-Assembling β-Glucan Nanomedicine for the Delivery of SiRNA. Biomedicines 2020, 8, 497. [Google Scholar] [CrossRef]
- Hwang, J.; Lee, K.; Gilad, Assaf. A.; Choi, J. Synthesis of Beta-Glucan Nanoparticles for the Delivery of Single Strand DNA. Biotechnol. Bioproc E 2018, 23, 144–149. [Google Scholar] [CrossRef]
- Zinkhan, S.; Ogrina, A.; Balke, I.; Reseviča, G.; Zeltins, A.; De Brot, S.; Lipp, C.; Chang, X.; Zha, L.; Vogel, M.; et al. The Impact of Size on Particle Drainage Dynamics and Antibody Response. J. Control. Release 2021, 331, 296–308. [Google Scholar] [CrossRef]
- Guimarães, L.E.; Baker, B.; Perricone, C.; Shoenfeld, Y. Vaccines, Adjuvants and Autoimmunity. Pharmacol. Res. 2015, 100, 190–209. [Google Scholar] [CrossRef]
- Zhang, P.; Andorko, J.I.; Jewell, C.M. Impact of Dose, Route, and Composition on the Immunogenicity of Immune Polyelectrolyte Multilayers Delivered on Gold Templates. Biotechnol. Bioeng. 2017, 114, 423–431. [Google Scholar] [CrossRef]
- Alu, A.; Chen, L.; Lei, H.; Wei, Y.; Tian, X.; Wei, X. Intranasal COVID-19 Vaccines: From Bench to Bed. eBioMedicine 2022, 76, 103841. [Google Scholar] [CrossRef]
- Alexander, E. Yeasts in Nanotechnology-Enabled Oral Vaccine and Gene Delivery. Bioengineered 2021, 12, 8325–8335. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.R.; Farazuddin, M.; Wong, P.T.; O’Konek, J.J. The Unfulfilled Potential of Mucosal Immunization. J. Allergy Clin. Immunol. 2022, 150, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kour, P.; Rath, G.; Sharma, G.; Goyal, A.K. Recent Advancement in Nanocarriers for Oral Vaccination. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Aouadi, M.; Tesz, G.J.; Nicoloro, S.M.; Wang, M.; Chouinard, M.; Soto, E.; Ostroff, G.R.; Czech, M.P. Orally Delivered SiRNA Targeting Macrophage Map4k4 Suppresses Systemic Inflammation. Nature 2009, 458, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, T.; Wang, L.; Shao, S.; Chen, Z.; Zhang, Z. In Vivo Targeted Delivery of CD40 ShRNA to Mouse Intestinal Dendritic Cells by Oral Administration of Recombinant Sacchromyces Cerevisiae. Gene Ther. 2014, 21, 709–714. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, H.; Zhang, W.; Li, Y.; Liu, L.; Leng, T. Yeast Cell Wall Particle Mediated Nanotube-RNA Delivery System Loaded with MiR365 Antagomir for Post-Traumatic Osteoarthritis Therapy via Oral Route. Theranostics 2020, 10, 8479–8493. [Google Scholar] [CrossRef]
- Bumcrot, D.; Manoharan, M.; Koteliansky, V.; Sah, D.W.Y. RNAi Therapeutics: A Potential New Class of Pharmaceutical Drugs. Nat. Chem. Biol. 2006, 2, 711–719. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.-J. Therapeutic SiRNA: State of the Art. Sig Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef]
- Karagiannis, T.C.; El-Osta, A. RNA Interference and Potential Therapeutic Applications of Short Interfering RNAs. Cancer Gene Ther. 2005, 12, 787–795. [Google Scholar] [CrossRef]
- Mahmoodi Chalbatani, G.; Dana, H.; Gharagouzloo, E.; Grijalvo, S.; Eritja, R.; Logsdon, C.D.; Memari, F.; Miri, S.R.; Rezvani Rad, M.; Marmari, V. Small Interfering RNAs (SiRNAs) in Cancer Therapy: A Nano-Based Approach. IJN 2019, 14, 3111–3128. [Google Scholar] [CrossRef]
- Huang, D.T.-N.; Lu, C.-Y.; Shao, P.-L.; Chang, L.-Y.; Wang, J.-Y.; Chang, Y.-H.; Lai, M.-J.; Chi, Y.-H.; Huang, L.-M. In Vivo Inhibition of Influenza A Virus Replication by RNA Interference Targeting the PB2 Subunit via Intratracheal Delivery. PLoS ONE 2017, 12, e0174523. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, F.; Limani, S.W.; Mnyandu, N.; Maepa, M.B.; Ely, A.; Arbuthnot, P. Advances with RNAi-Based Therapy for Hepatitis B Virus Infection. Viruses 2020, 12, 851. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Min, D.; Choi, Y.; Yoon, S.; Jang, J.; Hwang, J.; Jeon, H.; Cho, Y.W.; Choi, J. Self-Assembling β-Glucan Nanomedicine for the Delivery of SiRNA. Biomedicines 2020, 8, 497. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O.; Khan, D.N.; Singh, T.; Shukla, S.; Prakash, S.; Amita, J. Effect of SiRNA Targeting Dengue Virus Genes on Replication of Dengue Virus: An in Vitro Experimental Study. VirusDis. 2021, 32, 518–525. [Google Scholar] [CrossRef]
- Fujita, Y.; Takeshita, F.; Kuwano, K.; Ochiya, T. RNAi Therapeutic Platforms for Lung Diseases. Pharmaceuticals 2013, 6, 223–250. [Google Scholar] [CrossRef]
- Wu, S.; Liu, C.; Bai, S.; Lu, Z.; Liu, G. Broadening the Horizons of RNA Delivery Strategies in Cancer Therapy. Bioengineering 2022, 9, 576. [Google Scholar] [CrossRef] [PubMed]
- Setten, R.L.; Rossi, J.J.; Han, S. The Current State and Future Directions of RNAi-Based Therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef]
- Xiang, S.; Fruehauf, J.; Li, C.J. Short Hairpin RNA–Expressing Bacteria Elicit RNA Interference in Mammals. Nat. Biotechnol. 2006, 24, 697–702. [Google Scholar] [CrossRef]
- Bochicchio, S.; Dalmoro, A.; Barba, A.; Grassi, G.; Lamberti, G. Liposomes as SiRNA Delivery Vectors. CDM 2015, 15, 882–892. [Google Scholar] [CrossRef]
- Lundstrom, K. Viral Vectors Applied for RNAi-Based Antiviral Therapy. Viruses 2020, 12, 924. [Google Scholar] [CrossRef]
- Duman-Scheel, M. Saccharomyces Cerevisiae (Baker’s Yeast) as an Interfering RNA Expression and Delivery System. CDT 2019, 20, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Barreby, E.; Sulen, A.; Aouadi, M. Glucan-Encapsulated SiRNA Particles (GeRPs) for Specific Gene Silencing in Adipose Tissue Macrophages. In Lipid-Activated Nuclear Receptors; Gage, M.C., Pineda-Torra, I., Eds.; Methods in Molecular Biology; Springer New York: New York, NY, 2019; ISBN 978-1-4939-9129-7. [Google Scholar]
- Hapairai, L.K.; Mysore, K.; Chen, Y.; Harper, E.I.; Scheel, M.P.; Lesnik, A.M.; Sun, L.; Severson, D.W.; Wei, N.; Duman-Scheel, M. Lure-and-Kill Yeast Interfering RNA Larvicides Targeting Neural Genes in the Human Disease Vector Mosquito Aedes Aegypti. Sci. Rep. 2017, 7, 13223. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.; Hapairai, L.K.; Sun, L.; Harper, E.I.; Chen, Y.; Eggleson, K.K.; Realey, J.S.; Scheel, N.D.; Severson, D.W.; Wei, N.; et al. Yeast Interfering RNA Larvicides Targeting Neural Genes Induce High Rates of Anopheles Larval Mortality. Malar. J. 2017, 16, 461. [Google Scholar] [CrossRef] [PubMed]
- Hilderbrand, S.A.; Weissleder, R. Near-Infrared Fluorescence: Application to in Vivo Molecular Imaging. Curr. Opin. Chem. Biol. 2010, 14, 71–79. [Google Scholar] [CrossRef]
- Tesz, G.J.; Aouadi, M.; Prot, M.; Nicoloro, S.M.; Boutet, E.; Amano, S.U.; Goller, A.; Wang, M.; Guo, C.-A.; Salomon, W.E.; et al. Glucan Particles for Selective Delivery of SiRNA to Phagocytic Cells in Mice. Biochem. J. 2011, 436, 351–362. [Google Scholar] [CrossRef]
- Zakria, H.M.; Han, B.; Yue, F.; Mu, L.; Fang, Y.; Li, X.; Xu, K.; Zhang, Z. Significant Body Mass Increase by Oral Administration of a Cascade of ShIL21-MSTN Yeast-Based DNA Vaccine in Mice. Biomed. Pharmacother. 2019, 118, 109147. [Google Scholar] [CrossRef]
Figure 1.
A general summary of composition, features, and applications of mRNA vaccines.
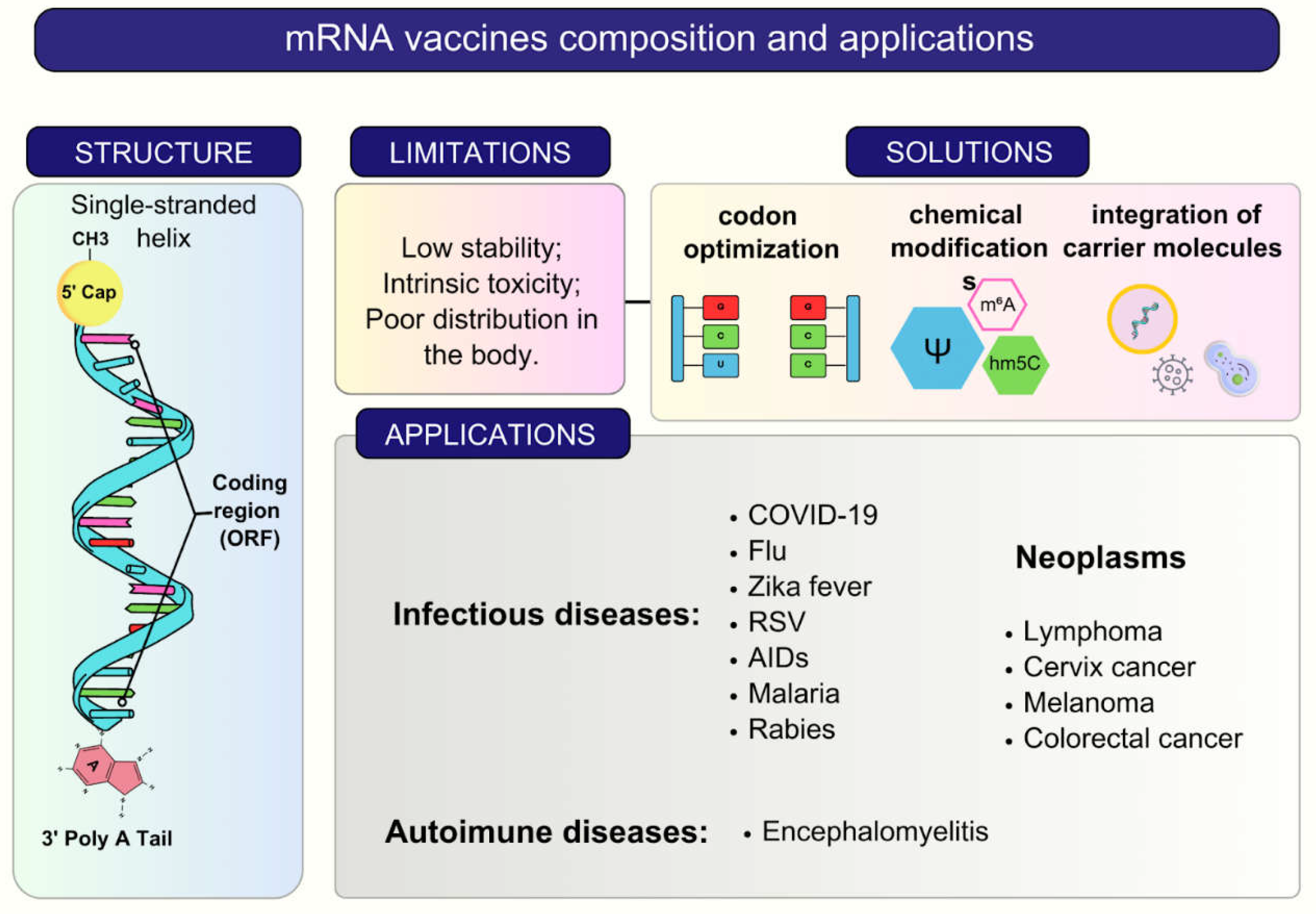
Figure 2.
A) Heat treatment promotes an increase in the exposure of β-1,3-glucans on the yeast surface (heating at 60°C for at least 1 hour), enhancing the adjuvant capacity of the yeast, favoring its recognition and binding to receptors present in antigen-presenting cells. B) Dendritic cells recognize yeast β-1,3-glucans through the Dectin-1 receptor, which promotes and facilitates phagocytosis (1). After the uptake, is formed the phagosome (2), where the yeasts are processed (3); the formed fragments and released antigens can be presented by MHC-I leading to the activation of cytotoxic TCD8+ lymphocytes, or by MHC-II inducing the activation of TCD4+Helper lymphocytes (4). Inflammatory cytokines such as TNF-α, IL-6, IL-8, and IL-1β or IL-12, IL-10, IL-23, and IL-27 released by activated dendritic cells polarize T cells to Th1 and Th17 profiles.
Figure 2.
A) Heat treatment promotes an increase in the exposure of β-1,3-glucans on the yeast surface (heating at 60°C for at least 1 hour), enhancing the adjuvant capacity of the yeast, favoring its recognition and binding to receptors present in antigen-presenting cells. B) Dendritic cells recognize yeast β-1,3-glucans through the Dectin-1 receptor, which promotes and facilitates phagocytosis (1). After the uptake, is formed the phagosome (2), where the yeasts are processed (3); the formed fragments and released antigens can be presented by MHC-I leading to the activation of cytotoxic TCD8+ lymphocytes, or by MHC-II inducing the activation of TCD4+Helper lymphocytes (4). Inflammatory cytokines such as TNF-α, IL-6, IL-8, and IL-1β or IL-12, IL-10, IL-23, and IL-27 released by activated dendritic cells polarize T cells to Th1 and Th17 profiles.
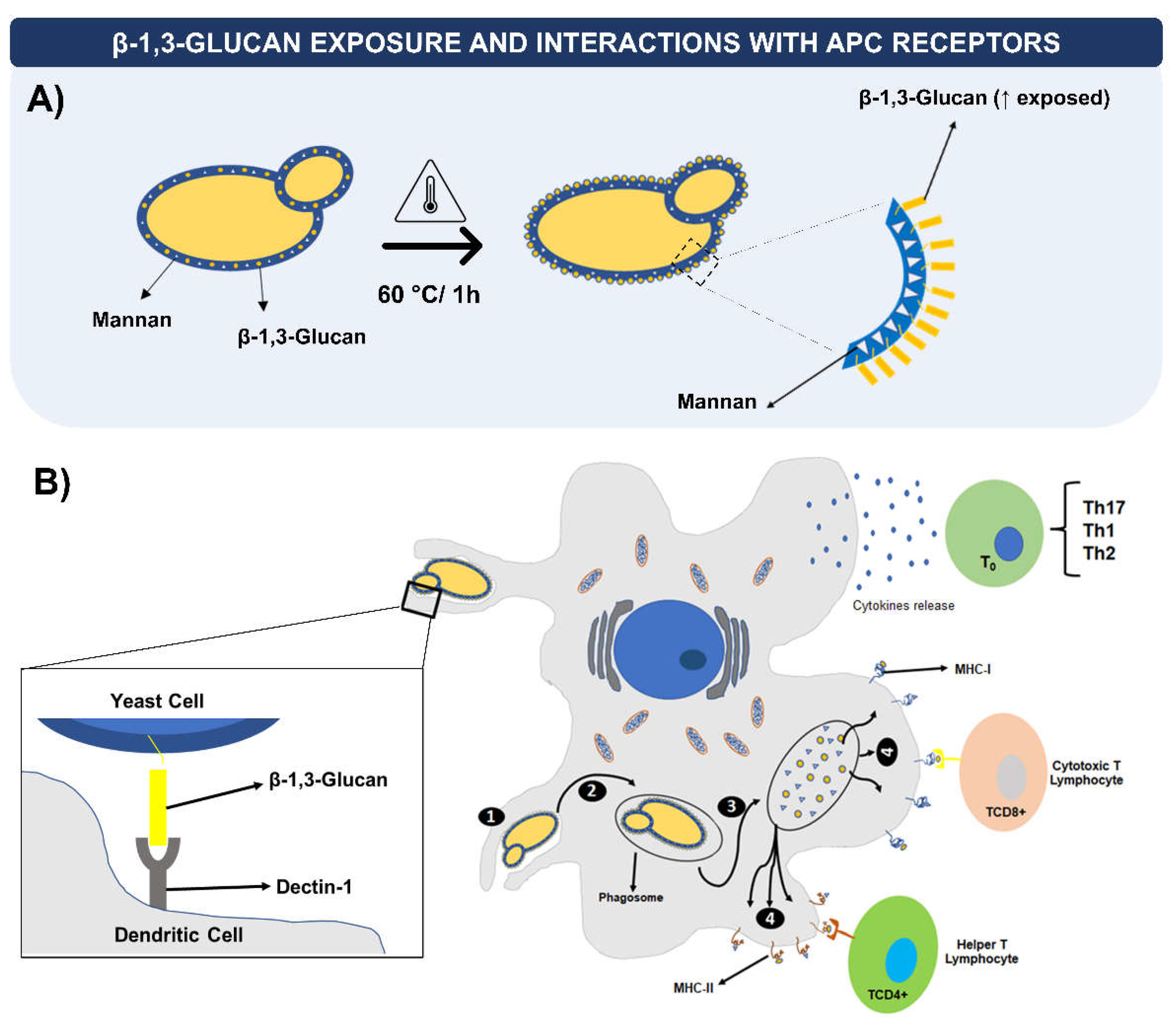
Figure 3.
Yeast-based vaccine modalities. A) Yeast, as a complete organism, can carry plasmids with the desired antigen gene inside them. The immunostimulatory effects result from the interaction between the yeast cell wall components and macrophage and dendritic cell receptors. B) β-glucan microcapsules or nanocapsules are particles obtained after physical-chemical treatment to remove the mannoprotein layer. Differences in size between the whole cell and the β-glucan particles can influence the phagocytosis process and the delivery and dissemination of antigens through lymphatic vessels.
Figure 3.
Yeast-based vaccine modalities. A) Yeast, as a complete organism, can carry plasmids with the desired antigen gene inside them. The immunostimulatory effects result from the interaction between the yeast cell wall components and macrophage and dendritic cell receptors. B) β-glucan microcapsules or nanocapsules are particles obtained after physical-chemical treatment to remove the mannoprotein layer. Differences in size between the whole cell and the β-glucan particles can influence the phagocytosis process and the delivery and dissemination of antigens through lymphatic vessels.
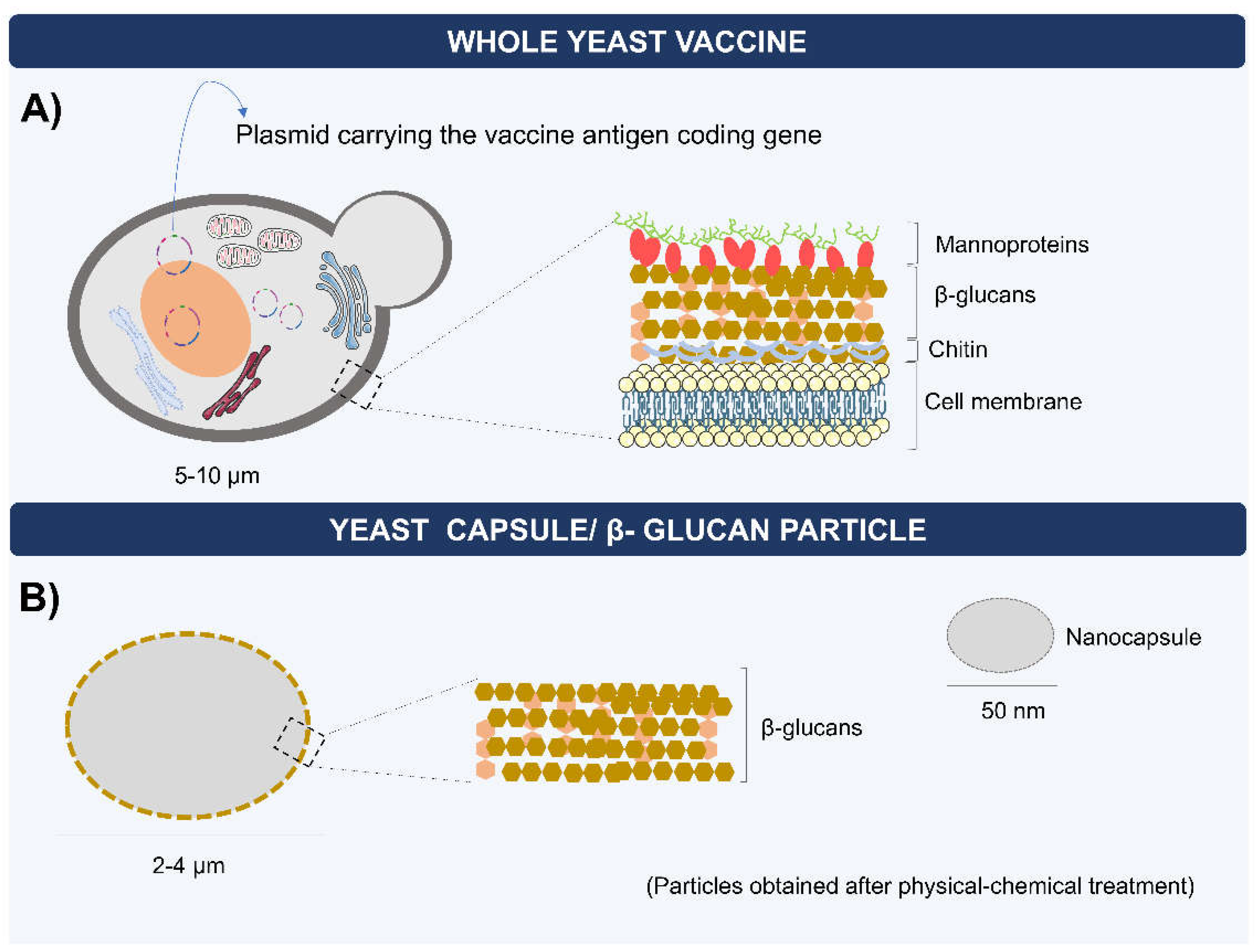
Figure 4.
Scheme demonstrating the GPs capture mechanism by APCs via Dectin-1 and TLR-2/6 receptor. In the Dectin-1 pathway, through the phosphorylation of phospholipase C (PLC)γ2, occurs the activation of protein kinase C (PKC) and syk activates the CARD9-Bcl10-MALT1 complex to induce the activation of the nuclear transcription factor (NF-κB). The GP recognition via TLR2/6 receptors, the MyD88 protein is activated by recruiting interleukin-1 receptor-associated kinases (IRAK-1 and IRAK-4) activating factor 6 associated with tumor necrosis factor receptor (TRAF6). This process activates growth factor β-associated kinase 1 (TAK1), triggering the activation of the IKK complex. This complex allows the phosphorylation of IκB, and its degradation results in the activation of NF-κB, which will be translocated to the nucleus to induce the expression of pro-inflammatory cytokines. .
Figure 4.
Scheme demonstrating the GPs capture mechanism by APCs via Dectin-1 and TLR-2/6 receptor. In the Dectin-1 pathway, through the phosphorylation of phospholipase C (PLC)γ2, occurs the activation of protein kinase C (PKC) and syk activates the CARD9-Bcl10-MALT1 complex to induce the activation of the nuclear transcription factor (NF-κB). The GP recognition via TLR2/6 receptors, the MyD88 protein is activated by recruiting interleukin-1 receptor-associated kinases (IRAK-1 and IRAK-4) activating factor 6 associated with tumor necrosis factor receptor (TRAF6). This process activates growth factor β-associated kinase 1 (TAK1), triggering the activation of the IKK complex. This complex allows the phosphorylation of IκB, and its degradation results in the activation of NF-κB, which will be translocated to the nucleus to induce the expression of pro-inflammatory cytokines. .

Figure 5.
Schematic representation of the main components present in the vectors used for yeast transformation. A) Vector with promoter recognized by mammalian cells. The cassette should also contain a leader sequence just after the promoter, and it is recommended to flank the target gene shRNA by endogenous miRNA such as miR30. Uracil is the selection marker usually employed. B) Cassette for expression with yeast promoters (constitutive or inducible). Besides auxotrophic marker genes, URA3/TRP1 sequences are loci for cassette integration.
Figure 5.
Schematic representation of the main components present in the vectors used for yeast transformation. A) Vector with promoter recognized by mammalian cells. The cassette should also contain a leader sequence just after the promoter, and it is recommended to flank the target gene shRNA by endogenous miRNA such as miR30. Uracil is the selection marker usually employed. B) Cassette for expression with yeast promoters (constitutive or inducible). Besides auxotrophic marker genes, URA3/TRP1 sequences are loci for cassette integration.
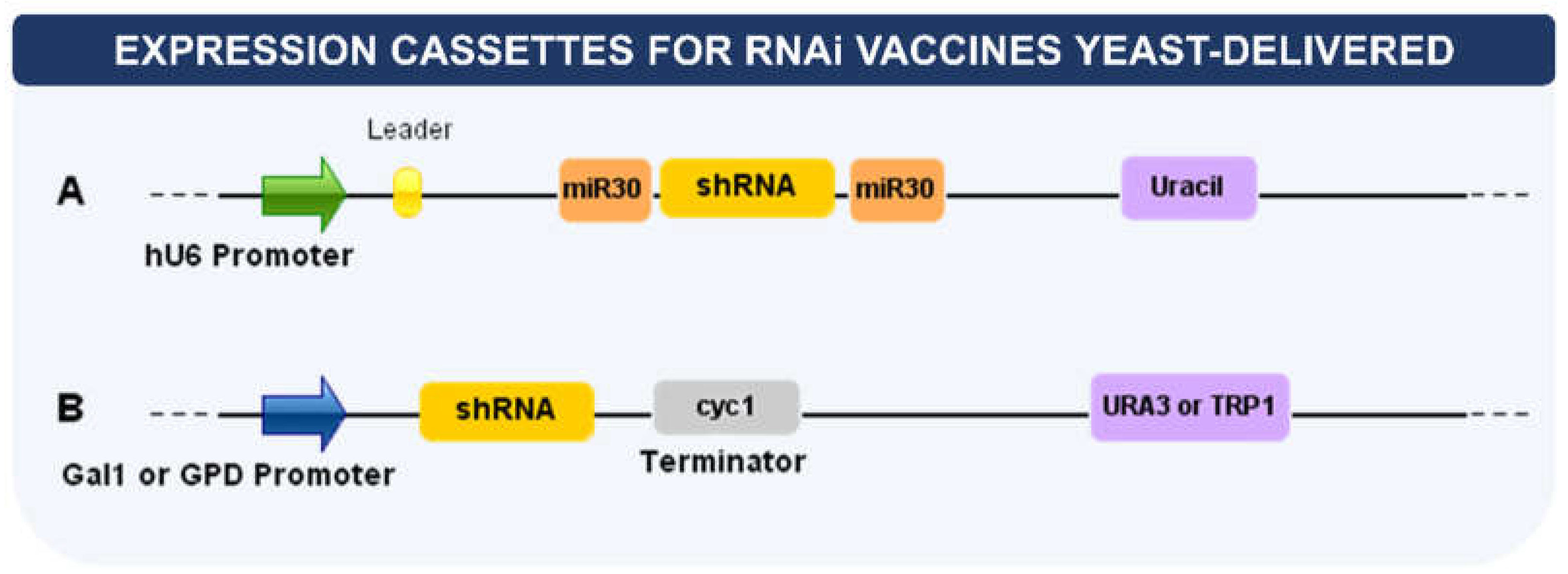

Table 1.
Main studies about the yeast-based oral route for RNA delivery.
| Type of RNA | Antigen | Vehicle type | Main findings | Ref. |
|---|---|---|---|---|
| siRNA | Tnf-α and Map4k4 | Yeast capsule | - Macrophages in the GALT internalize orally absorbed GeRPs, undergo siRNA-mediated gene silencing, and migrate into tissues throughout the body.- Oral gavage with Map4k4-siRNA-containing GeRPs significantly protects mice from LPS/d-GalN-induced lethality through inhibition of TNF-α and Il-1β production in macrophages. | [65] |
| mRNA | human pp65 | Yeast-derived microparticles | - Using CMV pp65 as a model antigen, showed that recombinant yeast cells can deliver functional mRNA very efficiently to murine macrophages and, importantly, to human DCs. | [39] |
| shRNA | CD40 | Whole yeast | - A shRNA driven by the U6 promoter along with a U6 snRNA leader sequence was delivered into intestinal DCs via orally administered yeast and effectively repressed the target gene (CD40) in vivo.- Compared with the control group, all shRNAs had a significant effect on IL-6, IL-10, IL-12, and TNF-α (P<0.01). And, the inhibition of the CD40 gene by CD40 shRNA1656-induced INF-γ increased expression (P<0.05). Thus, the inhibition of CD40 via orally administered recombinant yeast may be used in immunotherapy approaches. | [66] |
| shRNA | IL-1β | Yeast microcapsule | - The expression of IL-1β in small intestinal macrophage was downregulated with the treatment of recombinant IL-1β shRNA yeast;- The results showed that yeast microcapsule-mediated targeted delivery of IL-1β shRNA successfully, downregulating the intestinal inflammatory response in PTOA mice; | [47] |
| miRNA | miR365 antagomir | Yeast cell wall particle (YCWP) | - The results showed that most of YCWP can effectively resist the corrosion of SGF;- Evidence suggests that yeast was engulfed by gut-associated lymphatic tissue macrophages. These macrophages may traffic away from the gut and infiltrate other reticuloendothelial and mononuclear phagocyte tissues; | [67] |
| mRNA | rH9-DNA-RNA | Whole yeast | - Yeast vaccines elicited humoral and cellular immune responses;- The expression of the eGFP and mCherry in phagocytes of the duodenum was detected after the oral administration. The results suggested that both DNA and RNA cassettes were successfully delivered. The vaccine antigen, HA protein, was also expressed. | [36] |
shRNA (short hairpin RNA); siRNA (short interfering RNA); GeRPs (β1,3-d-glucan-encapsulated siRNA particles); Tnf-α (tumour necrosis factor α); IL-1β (interleukin-1β); miRNA (microRNA); SGF (Simulated gastric fluid).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



