Submitted:
24 April 2023
Posted:
26 April 2023
You are already at the latest version
Abstract
Whole genome duplication (WGD) or polyploidization can occur at the cellular, tissue, and organismal levels. At the cellular level, tetraploidization has been proposed as a driver of aneuploidy and genome instability, and correlates strongly with cancer progression, metastasis, and development of drugs resistance[1–6]. WGD is also a key developmental strategy for regulating cell size, metabolism, and cellular function[1,7–10]. In specific tissues, WGD is involved in normal development (e.g. organogenesis), tissue homeostasis, wound healing, and regeneration[10–17]. At the organismal level, WGD propels evolutionary processes such as adaptation[18], speciation[19], and crop domestication[20]. An essential strategy to further our understanding of the mechanisms promoting WGD and its effects is to compare isogenic strains that differ only in their ploidy. Caenorhabditis elegans (C. elegans) is emerging as an animal model for these comparisons, in part because relatively stable and fertile tetraploid strains can be produced rapidly from nearly any diploid strain[7]. Here we review the use of Caenorhabditis polyploids as tools to understand important developmental processes (e.g. sex determination, dosage compensation, and allometric relationships)[21–27] and cellular processes (e.g. cell cycle regulation, chromosome dynamics during meiosis)[28–32]. We also discuss how the unique characteristics of the C. elegans WGD model will enable significant advances in our understanding of mechanisms of polyploidization and its role in development and disease.
Keywords:
Whole genome duplication
; tetraploidy
; polyploidy
; aneuploidy
; Caenorhabditis
; WGD model (List three to ten pertinent keywords specific to the article yet reasonably common within the subject discipline.)
1. Whole genome duplication in development, evolution, and disease
Whole genome duplication (WGD) or polyploidization is a key step in vital biological processes including development [1,7,8,9,10], genome instability [1,2], adaptation [18], speciation [19], and crop domestication [20] (Figure 1). In nature, WGD exists at the cellular, tissue, organ, and organismal level. Polyploid cells normally exist in most multicellular diploid organisms [33,34]. In humans, certain cells from the placenta [35], mammary glands [36], liver [37], heart [38], skin [39], and bone marrow must become polyploid to perform specialized functions [40], for tissue homeostasis [11], or as part of the wound healing process [17]. For instance, during development, megakaryocytes undergo several rounds of endomitosis until they acquire 16 copies of the genome, and thus become giant cells that are hypermetabolic. This polyploidization is a basic requirement to produce platelets in the blood, as platelets are produced by fragmenting the cytoplasm of megakaryocytes [41,42,43]. Polyploid cells can arise by the fusion of cells resulting in a larger multinucleate cell, by endoreduplication (alternating cycles between G and S phases without undergoing mitosis or cytokinesis) resulting in a mononucleated cell, or by endomitosis (the cell enters mitosis but does not complete cell division) to generate either mononucleated giant cells or multinucleated cells [44].
1.1. Causes and downstream effects for WGD
Polyploidization and its effects have been observed in different biological contexts (e.g. in cells, tissues, and organisms) [10,33,45,46] and in multiple processes (e.g. during development, evolution, and disease) [1,3,5,11,47,48,49,50,51]. For instance, environmental stresses induce WGD in tissues (e.g. wounding or viral infection) [15,52,53] and organisms (e.g. large changes in temperature or water availability and salinity) [54,55,56,57,58,59]. The effects of WGD are diverse in nature—ranging from changes in gene expression and cell size, to epigenetic remodeling, and to altered responses to environmental stress [45,60,61]. Stress-induced WGD is a platform for genomic diversity and versatility that provides the means for rapid adaptability. In whole organisms, this can elicit a competitive advantage in a changing environment [61,62]; in cancer, it can elicit an adaptative advantage that promotes cancer development, progression or resistance to chemotherapeutic drugs [61,62]. In the pathogenic yeast Candida abdicans, WGD promotes formation of drug-resistant aneuploid progeny in response to antifungal drugs [18]. Therefore, queries about mechanisms that give rise to WGD and its effects in both sub-organismal and organismal systems will likely reveal additional commonalities across systems and processes.
Polyploidy often is confused with aneuploidy, in part because in humans, both phenomena are hallmarks of cancer [1,3,50,63]. Observations from numerous studies identify WGD in about 30% of human cancers [6,64] and as a precursor for many malignancies in cancer evolution [3,65,66,67,68,69]. Although, WGD does not seem to require a preexisting cancer driver mutation, it most often emerges after oncogenic mutations, such as lesions in the TP53 genes [6,66]. WGD promotes diversification of copy number alterations (CNAs) and is permissive for other chromosomal aberrations and genome instability associated with poor cancer patient prognosis [3,68,69]. Interestingly, a recent study revealed a mechanism by which WGD enhances cancer-promoting alterations in DNA organization and gene expression [68]. Induction of WGD can reduce the levels of proteins involved in chromatin packaging resulting in a reorganization of the 3D DNA compartments and domains. Over time, loss of this domain structure predisposes the cell to additional cancer-promoting alterations and expression of oncogenic genes [68]. Polyploidization can also be protective, as in the liver where it both provides the basis for genetic adaptation to hepatotoxic stress and also protects from malignant transformations [14,42].
Organismal WGD is common in extant plants [19,61,70,71,72], and is also observed in protists [73], fungi [4,62,74], bacteria and archaebacteria [75,76], and in several animal clades [12,34,47,48,50,51,77,78]. Evidence for ancestral whole genome polyploidization events is found in the genomes of organisms of most clades, including vertebrates [48,79,80]. Polyploid metazoans arise due to errors during meiosis that result in the formation of diploid gametes that when fertilized give rise to polyploid organisms. Fertilization between diploid (2n) and haploid (n) gametes results in a triploid (3n) organism; fertilization between two diploid (2n) gametes results in a tetraploid (4n) [45,47]. When the diploid gametes of two related species fuse, the resulting organism is said to be allopolyploid, whereas fertilization of diploid gametes from the same species gives rise to autopolyploid [80]. Polyploidization from the cell to organismal level most often results in lethality [61,81]. This lethality may be caused by errors partitioning the additional chromosomes during cell division, by WGD driven increases in cell size that interfere with tissues or organ scaling, or by expression changes that affect dose-dependent processes and complexes [80]. A newly formed polyploid may very occasionally overcome these obstacles, e.g. by establishing balanced genome expression. Then, if polyploidy provides a competitive advantage—as in a changing environment—polyploid lineages may outgrow their diploid counterparts and become established [51,61,80]. The established polyploid population will continue to evolve and then may “diploidize” to become a new species that has an adaptative advantage compared to the parental diploid species [61].
1.2. Laboratory multicellular organism models for WGD
A major impediment to addressing WGD and its impacts on the physiology and function of cells, tissues and organs is the scarcity of laboratory models that permit comparison between isogenic organisms with differing ploidy [45]. Many laboratory models are sterile or embryonic lethal when polyploid [82,83,84]. Generation of autopolyploid laboratory model organisms that are not sterile (e.g. plants) frequently involves the use of chemicals such as colchicine, known to cause aneuploidy and genetic mutations. These polyploids, therefore, may not be truly isogenic with the parental diploid strains they were derived from [62,80]. The recently developed method for generating tetraploids in Caenorhabditis utilizes transient knock-down of a meiosis-specific cohesin (i.e. rec-8) by RNA interference to produce diploid gametes [7]. Therefore, the derived tetraploids are unlikely to differ significantly from the diploid strains from which they were derived (See Figure 2A to overview the method) [7].
2. Caenorhabditis elegans as an animal laboratory model for understanding WGD
Several attributes of C. elegans nematodes make it an exceptional laboratory model for querying WGD: 1) It has a small number of chromosomes (five autosomes and a sex chromosome) allowing for easy visual evaluation of ploidy and differentiation between polyploidy and aneuploidy [85] (Figures 2B,C). 2) Despite C. elegans being a self-fertilizing hermaphroditic species, males arise from meiotic nondisjunction at a 0.2% frequency [86,87,88], allowing for genetic analysis [89]. 3) Each diploid hermaphrodite produces 200-300 offspring and has a lifecycle of three to four days from egg to egg, both of which facilitate multigenerational ecological and evolutionary studies. 4) C. elegans is normally diploid but like vertebrates (including humans), it has polyploid tissues or cells [90]. These include the intestine (20 cells with a total of 30-34 nuclei and a ploidy of 32C per nucleus) and the hyp7 cell (a syncytium of 139 nuclei with an average ploidy of 10.7C) [90,91].
Critically, the quick and reliable method to generate fertile and viable tetraploid Caenorhabditis strains from nearly any diploid strain [7] allows direct comparison of identical cells in isogenic organisms with differing ploidy (Figure 2). The availability of hundreds of mutant strains that impinge on different aspects of development, aging, stress, and reproduction, will facilitate directed study of genes in all of the processes that polyploidy has been reported to affect. The transparency of the worms allows visualization and direct comparison of physiology and function in the live animal. Combined, these characteristics make the C. elegans WGD model unique and provide an unprecedented opportunity to answer long-standing questions about organismal polyploidization and its effects in a multicellular animal.
2.1. Polyploidy and aneuploidy in C. elegans
In humans, all monosomies, except of the X chromosome are lethal, whereas trisomies 13, 18, 21, X and Y are viable. In C. elegans some aneuploidies are tolerated. These include X-chromosome trisomies (2A;3X = 13 chromosomes triplo-X hermaphrodites) and monosomies (2A;1X = 11 chromosomes XO males shown in Figure 3) [92]. Compared to diploid hermaphrodites, triplo-X animals have a Dumpy body morphology, lower fertility, and slower growth rate [92]. Trisomy of chromosome 4 (LGIV) has also been reported [93]. Although animals with this autosomal trisomy are viable and have a general morphological appearance that seems normal, they produce 50% fewer progeny than normal hermaphrodites [94]. Interestingly, large chromosomal fragments, often Mb is size, have been generated and preserved as free duplications and have been found for part of each of the chromosomes of C. elegans (reviewed in [22]).
In nature, most triploids are inherently unstable, unless they reproduce asexually [95]. For instance, the parasitic nematodes of the Meloidogyne incognita group (MIG) are stable hypotriploids that reproduce by asexual parthenogenesis [96]. C. elegans triploid animals (3A;3X and 3A;2X shown in Figure 3) can be generated by crossing diploid and tetraploid animals [24]. Only 15% of the embryos sired by triploid animals’ hatch [24], and those that do hatch tend to be diploid or become diploid in the next generation [28,97,98]. Triploid progeny inviability is likely because the odd number of homologs results in abnormal meiotic pairing and recombination [28,30,31], yielding aneuploid gametes that produce embryos that are inviable.
Victor Nigon reported the first induction of polyploid C. elegans—the first in any animal—in the laboratory in 1949 [97]. These tetraploid strains were generated by treating spermatogenic animals with either heat shock (several hours at 25°C) or exposure to colchicine. Both treatments were followed by screening for ‘larger than normal’ animals and cytological assessment of chromosome numbers in diakinesis-stage oocytes, when the chromosomes are highly condensed prior to the meiotic divisions (e.g. in Figure 2C). Newly formed tetraploids sired a range of phenotypes in offspring that included: progeny with variable body sizes; infertile hermaphrodites and males; intersexuality; and progeny that had reverted to diploidy [97,98]. The frequency of these aberrant phenotypes decreased with continuous passaging, but did not disappear, even after 20 generation of selection for tetraploids with high fecundity. Two major types of tetraploids were identified [24,99]: About 66% of the induced tetraploids sired very low frequencies of males (0.6%), compared to the ~2% incidence of males in diploids; the remainder sired an average of ~42% males. Interestingly, tetraploids of both classes could interconvert and give rise to a proportion of progeny of the other type, albeit at different frequencies [98]. After 30 generations the patterns of inheritance of high and low incidence of male progeny did not change. The tetraploid strains that sired high proportions of males are now known to correspond to a 4A;3X genotype and those that produced a low incidence of males to 4A;4X genotype (see Figure 3) [24]. Even in the most stable lines, tetraploid hermaphrodites had substantially fewer self-progeny (average of ~65 progeny) than diploid (250-300 offspring in wild type, N2), and reduced hatching (decrease of 13%) [24]. Nigon’s tetraploid strains were eventually all accidentally lost after 78 generations. Several other groups have isolated fertile and relatively stable tetraploid C. elegans and C. briggsae strains [7,21,23,24,31]. Like their predecessors, each of these tetraploid strains has been reported to produce reduced numbers of self-progeny and to sporadically diploidize. Since the diploids can rapidly overtake a population due to their higher fertility, C. elegans tetraploid strains need to be maintained by continuous selection for the largest animals—at least for the first tens of generations. Studies of the early events post-tetraploidization (e.g. in generations 1-10) therefore should shed light on how organisms acclimate to tetraploidy and become established.
Figure 3.
Sexual fate in C. elegans is determined by the X-chromosome/autosome ratio. C. elegans has six chromosomes, 5 autosomes and one X chromosome. Diploid males and hermaphrodites have 2 sets of autosomes and one or two X chromosomes, respectively. Triploid males and hermaphrodites have three sets of autosomes and one or two X chromosomes making 17 or 18 total chromosomes, respectively. Tetraploid males have 22 chromosomes: four sets of autosomes and two X chromosomes. By contrast, tetraploid hermaphrodites have four sets of autosomes and either three or four X chromosomes. .
Figure 3.
Sexual fate in C. elegans is determined by the X-chromosome/autosome ratio. C. elegans has six chromosomes, 5 autosomes and one X chromosome. Diploid males and hermaphrodites have 2 sets of autosomes and one or two X chromosomes, respectively. Triploid males and hermaphrodites have three sets of autosomes and one or two X chromosomes making 17 or 18 total chromosomes, respectively. Tetraploid males have 22 chromosomes: four sets of autosomes and two X chromosomes. By contrast, tetraploid hermaphrodites have four sets of autosomes and either three or four X chromosomes. .

2.2. Polyploid and aneuploid animals uncover C. elegans modes of sex determination and dosage compensation
One of the most critical developmental decisions that a heterogametic organism must make early in life is whether to develop as male or female and, concomitantly, whether to activate dosage compensation. From studies in C. elegans with different ploidies, it became apparent that sex is assessed by the ratio of sex chromosomes to autosomes (X/A) and that induction of hermaphrodite fate activates dosage compensation, which is achieved by reducing expression from the pair of X chromosomes [87,100]. Male phenotypes arose from animals with 2A;1X, 3A:2X, and 4A;2X, whereas 2A;2X, 3A;3X, 4A;4X, and 4A;3X gave rise to hermaphrodite phenotypes [24] (Table 1 and Figure 3).
Together, these studies led to the conclusion that an X:A ratio > 0.74 induces female/hermaphrodite identity and < 0.67 promotes male development. Further support for these conclusions came from manipulation of the X/A ratio using X-linked gene duplications in the 2X;3A triploids to show at least 3 loci were involved in determining sex [23]. We now know the molecular details about the genes and processes involved in both assessing the X/A ratio and in executing male and hermaphrodite fates and dosage compensation [101]. Nevertheless, questions remain about how allotetraploids generated by hybridization of different tetraploid Caenorhabditis species would assess the X/A ratio, and whether manipulation or modification of this system could be used to stabilize polyploid organisms.
3. Utilizing polyploid C. elegans as tools to investigate developmental processes
In the last ~50 years, comparisons between C. elegans strains of differing ploidy have been utilized as tools to query developmental processes including meiosis, embryonic divisions, in addition to sex determination and dosage compensation.
3.1. Understanding early events of meiotic prophase I
During meiosis, recombination between the duplicated maternal and paternal homologous chromosomes transiently connects the homologs to orderly partition them in the first division, while maintaining sister chromatids together until the second division. To recombine, homologous chromosomes must first find one another in the nucleus (i.e. pair), become aligned along their lengths, and then stabilize their association by assembling the synaptonemal complex (i.e. synapse) [102]. Observations of the effects of having additional chromosomes in C. elegans triploid and tetraploid animals have helped to uncover rules underpinning homologous chromosome dynamics during meiosis [30,31]. Additional chromosomes did not significantly alter the initial steps of pairing into groups of three or four homologs in triploids and tetraploids, respectively. However, full alignment was often perturbed and synapsis was frequently incomplete, resulting in lengthening of the period in which synapsis normally takes place [30]. The imperfectly synapsed chromosomes became marked by di-methylation (H3K9me2), which has been proposed to prevent the synapsis defects from being detected by the quality control system [30]. Further analysis revealed that two pairs of synapsed homologous chromosomes form in tetraploids [31]. This pairwise synapsis also takes place in germ lines of mutants (i.e. spo-11) that are deficient in early steps of recombination, as in diploids. In triploid germ lines, one pair of homologs synapse and the remaining homolog eventually folds onto itself, in a process known as self-synapse. By contrast to diploids, the pairwise synapsis is disrupted in the presence of the recombination-deficient mutations, suggesting that in C. elegans, as in other organisms, homologous recombination plays a role in promoting synapsis [31]. Polyploid animals heterozygous for translocation chromosomes (e.g. nT1(IV;V)) were utilized to test whether pairwise synapsis is dictated by full sequence homology along the chromosome length. Synapsis was normal in diploid translocation heterozygotes (i.e. nT1/+ or mln1/+), as long as they contained the pairing center (PC) sequence that is required for homologous chromosomes to initially find each other in the nucleus [103]. In tetraploids (nT1/nT1/+/+), there was no preference for a synapsis partner as translocation chromosomes paired with normal chromosomes with the same frequency as with themselves. In contrast, associations and synapsis between pairs of homologous chromosomes was biased towards homologs with identical sequences in the triploids [31]. These findings support the proposed model in which the initiation of synapsis depends on PC-mediated associations of homologs but completion of synapsis along the homologous pair is promoted by recombination [31].
3.2. Understanding meiotic and early embryonic cell divisions
Female meiotic divisions are asymmetric. Each division gives rise to a large oocyte and a tiny polar body. The resulting egg contains a haploid set of chromosomes, as half of the chromosomes in each division are discarded into the polar bodies. Therefore, C. elegans hermaphrodites trisomic for the X chromosome (2A;3X) would be expected to produce equal proportions of eggs that carry either one or two X chromosomes (one sister from the paired homologues +/- one sister from the self-synapsed chromosome). However, the observed X:XX ratio in these eggs is 2:1 [92]. Live and high-resolution visualization of the oocyte cell division in these trisomic worms showed that the self-synapsed chromosomes were preferentially partitioned into the polar body during oogenesis [28,32]. In addition, in triploid animals that have one set of homologous chromosome pairs (bivalents) and a set of unpaired chromosomes (univalents); univalents were most often discarded into the polar body of the first oocyte division [32]. In these triploids, preferential segregation of the single sister chromatids into the polar body could also be observed in the second division. These results provide mechanistic insights into why triploid animals rapidly diploidize.
Several seminal studies elucidating mechanism of embryonic cell division have utilized C. elegans tetraploid and haploid (1A;1X) embryos [26,104,105,106,107]. One of the first questions this work addressed was how the paternally-inherited centrosomes duplicate and become associated with the nucleus. In the zygote, the migration of the maternal and paternal pronucleus towards one another and their subsequent fusion is important to ensure the fidelity of the first division of the embryo. In addition to contributing a haploid genome, the sperm also contributes a pair of centrioles to the zygote. These centrioles remain in the proximity of the male pronucleus and, as they gather pericentriolar proteins (PCM) from the maternal cytosol, they duplicate to form the first pair of centrosomes of the zygote [108]. Concurrently, the male pronucleus grows as DNA decondenses and the centrosomes then become attached to its surface [109]. Comparison of zygotes with different ploidies suggests that centrosome attachment to the nuclear membrane depends on the size of the male pronucleus [104]. Live imaging showed that diploid embryos with mutations in nuclear envelope genes have aberrantly small male pronuclei with only one attached centrosome; the second centrosome only became attached as the male pronucleus surface increases [104]. The detached centrosome phenotype was suppressed in zygotes from tetraploid animals that had larger pronuclei. Conversely, the detached centrosome phenotype was enhanced in haploid zygotes with a smaller pronuclei. It has been speculated that dynein motor accumulation on the pronuclear surface area is the limiting factor for centrosome attachment with a threshold of dynein required for both centrosomes to attach [106,110]. Of note, the haploid embryos in this study were generated utilizing conditional mutant sperm that lack nuclear DNA (i.e. emb-27(g48ts)) [106].
A second major question that was addressed used animals of differing ploidy was whether nuclear to cytosol ratio regulates cell cycle timing which had been hypothesized from work in other organisms [111,112,113]. The roles of nuclear size, its ratio to cytosolic volume, DNA content, and overall cell size in cell cycle timing of the C. elegans embryo were evaluated by comparing diploid (N2 Bristol), tetraploid (strain SP343), and haploid embryos (generated by extracting the pronucleus from the fertilized egg before the zygote formed) [105]. In these embryos, different nuclear/cytosolic volume ratios were examined by extruding nuclei with little to no cytosol, some cytosol, or increased cytosolic volume, the latter created by laser-induced cell fusion followed by extrusion of one nucleus. Live imaging revealed that cell cycle timing was neither affected significantly by altering cell size, nor by nuclear to cytosol ratio, nor by DNA content [105]. Instead, cell fusion experiments revealed that cell cycle periods could be altered by the cytosol of another cell in a different cell cycle period. Together with other findings, these early results are consistent with studies that posited models for the existence of “cytoplasmic factor(s)” that controls cell cycle timing [114,115,116]. The cytoplasmic factors that regulate many aspects of the cell cycle and its timing have now been identified [117].
Alterations of ploidy in embryos was also utilized to assess scaling of the microtubule-based spindle [26]. Accurate chromosome partitioning relies on the assembly of a properly shaped spindle during cell division. The metaphase spindle forms a diamond shape, with two opposing corners at each centrosome and two at the ends of the metaphase plate. The ratio between the spindle length (from centrosome to centrosome) and width (of the metaphase plate at the equator) was constant in in vitro Xenopus extract studies that either molecularly or mechanically perturbed the dimensions of the metaphase spindles [118,119]. In vivo studies in C. elegans have been instrumental to our understanding of spindle shape. Initial studies showed that spindle length is affected by cell size as spindle length and width decreased as embryogenesis progressed [26,114]. Interestingly, reduction of the spindle width was observed when the spindle length was decreased (e.g. by RNAi depletion of tpxl-1 (the C. elegans homolog of Xenopus TPX2 involved in spindle assembly) or spd-2 (involved on multiple aspects of centrosome biogenesis and thus spindle formation)) [120]. Conversely, the opposite was not the case: changing the width of the spindle did not affect the spindle length. In haploid embryos the spindle width is reduced and in polyploid embryos [26] increased; yet in neither case was embryo size or spindle length affected [121,122,123,124,125]. Correlation analysis between spindle length and width in haploid, diploid and polyploid embryos revealed a simple equation that can predict spindle width based on the spindle length and ploidy of the embryo [26].
Another important correlation that was observed with the reduction in cell size during embryogenesis is related to chromosome condensation, a chromosome feature that is important for accurate chromosome inheritance [120,126,127,128,129]. Haploid and tetraploid embryos produced by diploid klp-18(RNAi) mothers show differential condensation of chromosomes compared to diploid control embryos [120]. Chromosome condensation decreased in haploid embryos in comparison to the diploid embryos, whereas condensation increased in tetraploid embryos. Differential condensation was independent of cell size, the speed of spindle elongation, and spindle length [26,120]. Therefore, an inverse correlation between nuclear DNA density and chromosome condensation was proposed and tested by observing chromosome condensation in dividing diploid and klp-18(RNAi) haploid embryos, during divisions as the size of the nucleus decreases during development [120]. The relationship between the DNA density in the nucleus and chromosome length (a measure of chromosome condensation) fit a regression line for the dividing diploid embryo. Importantly, this regression line also fitted well the measurements of the dividing haploid embryo, as well as, when the nuclear size was manipulated by reducing the transcripts by RNAi of ima-3 (nuclear pore import protein), ran-3 (ortholog of the human RCC1 regulator of chromosome condensation) and C27D9.1 [120], suspected or known to affect nucleus size [130]. These analyses revealed an allometric relationship between chromosome length/condensation and the relative ratio of DNA amount per nucleus size that is consistent with the hypothesis that chromosome condensation is regulated to adapt to fit optimally the amount of DNA in the physical space within the nucleus.
4. Utilizing Caenorhabditis to understand the effects of polyploidization
In addition to being utilized as tools to study development, C. elegans polyploids have also been used to study the process of polyploidization itself and its immediate after-effects.
4.1. Polyploid tissues in C. elegans
At least two tissues are polyploid in C. elegans: the intestine and hypodermis (part of the nematode epidermis) [90]. The intestine is a tube responsible for digestion and makes up nearly a third of the animal’s body. All intestinal cells are derived from a single blastomere (E) starting at the eight-cell embryo stage [131,132]. The intestine of a newly hatched embryo has twenty diploid cells. When the L1 larvae hatch, they have 20 intestinal cells that have 20 diploid nuclei [90]. Of these nuclei six do not divide nor replicate their DNA, four nuclei may or may not replicate their DNA or divide, and 10 nuclei replicate and divide by endomitosis cells with two diploid nuclei each. Therefore, by the end of the L1 stage the 20 cells of the intestine have 30-34 diploid nuclei. Prior to the transition molt to the L2 stage all nuclei of the intestine endoreduplicate their DNA (4C each nucleus). Nuclei continue to endoreduplicate prior to the molts into the L2 (8C each nucleus), the L3 (16C), and finally the L4 (32C) larval stages. Thus, the adult intestine is composed of 20 cells with a total of 30-34 nuclei, each with 32C ploidy [90]. The hypodermis has many roles in development including establishment of the basic body plan in the embryo such as body shape and size, regulation of cell fate specification, and guidance of migrating axons and cells [133,134,135]. The hypodermal cell hyp7 is a multi-nucleated syncytium that encases most of the adult animal body (the tail and head of the animal are covered by smaller hypodermal cells) [90]. The hyp7 syncytium contains 139 nuclei in the adult and arises from many cell fusions (during embryogenesis 23 cells fuse to hyp7 and post-embryonically 116 cells also fuse to it) [90,136]. The embryonically-derived nuclei in hyp7 remain diploid, whereas the ~98 cells generated post-embryonically endoreduplicate to become tetraploid [90,136]. Most of these nuclei undergo up to 2 more endoreduplication cycles that result in the hypodermal nuclei having an average 10.7C ploidy by the fully grown adult stage [91]. The ploidy of the hyp7 cell is key for adult animal size regulation, (see section on allometric studies, below) [137].
In diploid cells, coordination of the centrosome and cell cycle ensures a single duplication of the centrosome per mitotic cycle, which is crucial to prevent supernumerary centrosomes accumulation that can lead to genomic instability [138,139]. Thus, the regulation of the centrosome in polyploid tissues is equally important for maintaining genome stability [140]. The endopolyploidy of the intestine and hypodermis of developing C. elegans allowed for comparative studies of how tissues respond to this challenge. In both the polyploid intestine and the hypodermis V1 cell nuclear lineages of the hyp7 syncytium, loss of PCM correlates with an apparent lack of centriole duplication resulting from uncoupling of the centrosome cycle in the first endo-cycle followed by progressive centriole elimination [141]. The SPD-2 protein with a normal dynamic localization to the PCM and the centrioles is key in for centriole duplication. An spd-2 phosphomimetic mutant, SPD-2S545E that can be phosphorylated in vitro by CDK1 and CDK2 [141] leads to supernumerary centrosomes in the intestinal cells [141]. This supports the hypothesis that the putative CDK phosphorylation site at S545 is a part of the molecular mechanism preventing centriole duplication during the endo-S phases of the intestine [141]. Since the SPD-2S545E mutation does not affect centriole number in the hypodermal cells, this regulation appears to be specific to intestinal cells. A different putative phosphomimetic SPD-2S375E mutation, affects a Polo Like Kinase-1 (PLK-1 in C. elegans) consensus phosphorylation site, and results in the persistence of SPD-2 on centrioles at the L2 stage that agrees with the observed delay in centriole elimination in some of the intestinal nuclei of this mutant. Surprisingly however, PLK-1 does not seem to regulate SPD-2 stabilization nor centriole elimination [141]. Instead, knockdown of the proteasome b-subunit (pbs-3) results in both SPD-2 persistence indicative of a lack of centriole elimination in many of the intestinal cells. This finding implicates ubiquitin-mediated proteolysis in the uncoupling of the endocycle and centriole cycle in the intestine [141].
One question that arose when looking at polyploid tissues is whether there is a biologically-significant functional difference between mononucleated polyploidy (restricted to a single nucleus) and multinucleate polyploidy (in more than one nucleus) [12,142]. The developing intestine in C. elegans was used to address this question by converting the normally binucleated intestinal cells into mononucleated cells of identical ploidy using auxin- induced degradation [143] of either a regulator of mitotic entry (e.i. CDK-1) or a kinetochore protein required for chromosome partitioning in mitosis (i.e. KNL-1) to block endomitosis [49]. Whereas absence of CDK-1 prevents entry into mitosis and essentially converts the endomitosis into an endocycle, degradation of KNL-1 allows for entry into mitosis but prevents chromosome partitioning into two nuclei [49]. Neither knock-down affects cell size or morphology, but the mononucleated intestinal cells have reduced nuclear to cytoplasmic ratio given that their nuclei more than double in size [49]. Transcriptome analysis revealed that all of the vitellogenin genes (vit-1 to vit-6) are downregulated in mononucleated intestines. Vitellogenins are yolk proteins that are required for lipid transport and lipid loading onto oocytes and embryos and promote progeny fitness. Accordingly, the embryos from mothers with mononucleated intestines have reduced survival. Similarly, decreasing vitellogenin expression in binucleated gut cells phenocopies the impact on progeny fitness and increasing vitellogenin expression, by overexpressing their transcriptional activators, results in mothers siring progeny with similar fitness regardless of whether they had mono- or bi-nucleated intestines [49]. Importantly, many of the genes that are downregulated in animals with mononucleated intestines normally increase in expression during the L4 to adult transiition, implicating a major role for binucleation at this developmental stage [49].
4.2. Biological size and scaling (allometry)
Regulation of cell size and scaling of tissues and organs are critical for development and influence homeostasis, metabolism, and function [45]. Polyploidization has been generally associated with increases in cell and organism size. For example, genome size and ploidy correlate linearly with cell volume in yeast [144,145,146]. However, effects of polyploidy are cell-type specific in many multicellular organisms, with different cell types having a nonlinear relationship between ploidy and cell size [147]. For instance, in plants there is a strong correlation between ploidy and cell volume in epidermal pavement cells, but the volume of palisade mesophyll cells remains at a constant size despite changes in ploidy [148]. These and other observations [45,149] make it apparent that the DNA mass does not fully explain the effect of ploidy on cell (and nuclear) size [45,149,150,151,152]. In most polyploidization models, including plants, interpreting the direct effect of ploidy is complex because the increase in cell size in tissues and organs is balanced by a reduction in cell number [45,150,153,154,155]. The invariant cell lineage in C. elegans simplifies the identification of mechanisms underpinning correlations between ploidy and scaling of cells, tissues and organs.
Allometry studies related to the effects of ploidy in C. elegans have been performed for cells and animals [25,26,91,98,120,156,157,158,159]. Nigon (1949) [99] reported that, compared to the 1300 µm long diploid hermaphrodites, 4A;3X hermaphrodites were on average 1360 µm and the 4A;4X hermaphrodites were 1560 µm long. The more profound increase in 4A;4X hermaphrodites containing 24 chromosomes (See Figure 3) compared to the 4A;3X hermaphrodites (23 chromosomes) is surprising and suggests that gene expression from the X chromosome may contribute to animal size significantly more than gene expression from autosomes.
The hypodermis secretes the cuticle forming the exoskeleton that houses the nematode’s internal organs and nervous system. There is a proportional relationship between the size of the C. elegans body and the size of the hypodermal cells (seam cells and the syncytium hyp7) [156,160,161]. Comparison of 12 nematode species from the order Rhabditida revealed a weak correlation between the number of nuclei and hypodermal volume (hyp7 in C. elegans), suggesting that body size was at least partly independent of cell number [91]. A significant correlation between animal size and the product of the number of nuclei and the ploidy was observed across species. Therefore, unlike most organisms where the evolution of body size relies in changes in cell number, in nematodes body size evolution was likely driven by the size of the hypodermal syncytium [91]. Comparison of a tetraploid strain and its diploid revertant revealed a 39% increase in the adult body size of tetraploids [25]. The unexpected 1.4-fold increase in volume (instead of the expected 2-fold) in tetraploids is at least in part explained by the lower-than-expected ploidy in the hyp7 cell (16.7C instead of 22C) and the small reduction in the number of cells (by less than two nuclei) [25]. However, one should be careful in making conclusions from this comparison because it is likely that the reverted diploid differed from the original, but these were not examined. Multiple independent tetraploid strains from nearly any diploid strain provides the possibility of conducting this comparison more rigorously. Levels of endoreduplication in the hyp of C. elegans was found to be important in regulation of the adult animal size. When C. elegans young adults were exposed to the DNA replication inhibitor, hydroxyurea (HU), endoreduplication of the hyp7 nuclei was inhibited and a reduction in body size was observed [25]. Cyclin E is required for endoreduplication in mice and Drosophila [162,163]. Null mutants carrying mutations the C. elegans homolog of Cyclin E, cye-1, have an ~1.8-fold reduction in size of the hyp7 syncytium and are 35-54% the size of wild-type worms [25].
Many signaling pathways and genes affecting diploid animal size have been characterized in C. elegans. These include pathways that only regulate embryo size, that regulate multiple developmental stages, and that influence adult body size. The TGF/BMP signaling pathway has a major role in the response to ploidy as it regulates both adult size and hyp7 ploidy [25,137,160,161,164,165]. The TGF signaling pathway that regulates body size in C. elegans includes the DAF-4 type two receptor, the DBL-1 ligand, and the downstream SMADs (SMA-2, SMA-3, SMA-4, and SMA-6) [166]. DBL-1, the human ortholog of human BMP7, is required for post-embryonic growth animal [91,159,167]. Whereas DBL-1 overexpression results in longer-than-normal animals (Lon phenotype) [164], mutations in the dbl-1 gene cause a dwarf phenotype (Sma) and reduced hypodermal ploidy [25,164]. Interestingly, as with the cye-1 mutant, exposure of dbl-1 mutants to hydroxyurea neither further reduced hyp7 ploidy nor affected its body size, suggesting that the DBL-1 normally promotes hyp7 endoreduplication [25]. Together these and other findings [25,161] suggest that DBL-1 regulates adult growth in a dose dependent manner [158,164] by promoting endoreduplication in the adult hypodermis (hyp7) [161], likely via the CYE-1 cyclin [25].
The regulation of body size by DBL-1 pathway is complex. Many of the molecular components in the DBL-1 pathway and other pathways affecting endoreduplication in the hypodermis have been identified, and their localization and molecular or genetic interactions described. For instance, overexpression of LON-1, a member of the conserved PR-protein superfamily [157,168], causes hypo-endoreduplication and absence of LON-1 protein causes hyper-endoreduplication in the hypodermis [157]. Specifically, the levels of lon-1 transcript depend on the dosage of the dbl-1 gene and TGF signaling. Other genes and processes affecting diploid animal size are also likely to play important roles in this process. These include cuticle collagens, ß-H Spectrin, TOR kinase, MAPK signaling, Hippo-Warts signaling, and the insulin signaling pathway [159,169,170,171,172,173,174,175,176,177,178]. These genes are expected to show altered expression in polyploid animals [45,51,179,180]. Global metabolic genes, such as rRNA or tRNA genes, are likely also involved in regulating biological scaling in polyploid animals [45,181]. Whether these pathways contribute equally to growth regulation in polyploids remains an open question.
5. Potential future queries utilizing C. elegans polyploids
The ease of creating synthetic autotetraploids from any strain utilizing RNAi is shepherding in a new era. We can now expand the use of C. elegans polyploids to study cellular and developmental processes (e.g. allometric relationships of different cell types, cell division dynamics) and disease (e.g. genome instability, pathological cell divisions, cancer) by comparing equivalent organisms, tissues, organs and cells from isogenic diploid, triploid, and tetraploid animals. This method allows for the first time in C. elegans the ability to query the direct impact of organismal polyploidization on individual tissues across time. In addition, this polyploid model's unique features will be of particular benefit for advancing our understanding of WGD and its effect on genome structure, transcriptome size (total RNA per unit of DNA mass) [182], specific-gene expression, regulation of gene expression by chromatin modification, and importantly how these changes affect the physiology, metabolism and developmental character of the organism.
In the course of evolution, parts of the genomes of polyploids are lost [183,184,185], a process known as fractionation [186]. This can occur because the purifying selection – the drive to remove mutated genes – is relaxed on duplicated genes, leading to both gene and regulatory element loss [186,187]. In allotetraploids, allele dominance can facilitate gene retention/fractionation, but genes involved in multi-subunit complexes (ribosomes, polymerases, etc.) tend to be retained [186]. Previous observations in plants showed that they undergo loss of repetitive sequences, mostly transposable elements, by recombination [188,189,190]. This can reduce the genome by up to 25% within the first few generations of newly formed autopolyploid [188]. Synthetic auto- and allo-tetrapolyploid C. elegans will allow both short term and longitudinal studies of the process of genome downsizing given the nematodes’ short life cycle and the ability to manipulate its environment. Additionally, these animals would facilitate study of tissue-specific differences and mitotic stability of ploidy during development. Of particular interest will be comparative analyses of the endopolyploid hyp7 and intestinal cells to determine how their DNA size/mass scales with ploidy.
Gene expression studies have been done to assess differential gene expression between in allopolyploid plants, often comparing diploids to an ancestral hybrid species [45]. An important mystery remaining is the stoichiometry of the changes in whole transcriptomes of similar cells from animals with differing ploidy [182,189]. Two competing hypotheses regarding the effect of organism polyploidization on global gene expression have been proposed; 1) each duplicated genome retains diploid transcription levels, or 2) all gene loci are affected by dosage compensation to maintain gene dose similar to that of diploid. Although very few studies have been done to compare the effect of differing ploidy on transcriptome size, neither global suppression nor doubling of expression is uniquely likely since allopolyploids studies showed an intermediate transcriptome size. Importantly, single-cell transcriptomic studies showed a high variability or noise in gene expression within a cell type and between different cell types. Specific regulatory genes (e.g. RNA polymerase 2 and 5.8S rRNA) have been found to differentially increase in expression with increasing ploidy [190]. Global and specific gene expression can also be affected by chromatin remodeling (e.g. chromatin and histone modifications) and specific histone modification, particularly histone H3K56 acetylation, are thought to buffer increases in mRNA synthesis due to increased number of gene copies [191].
The ability to create tetraploids with any strain facilitates longitudinal studies of the tetraploid. For example, one can now create recombinant inbred strains of auto- and allotetraploids to address how the genome changes over 10s -100s of generations. Combining these studies with exposures to environmental stressors, such as disease, temperature, infection and DNA damage, has the potential to shed light on genomic changes that provide flexibility and drive adaptation.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NIGMS, grant number R01GN104007 to JLY; PSC-CUNY award, 65508-00 53 and the CUNY-Research Foundation, FFPP award to MS; Tow mentoring and research program to MT and MS; Brooklyn College’s, Samuel and Rose Goldstein award to MT.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Davoli, T.; Lange, T. de The Causes and Consequences of Polyploidy in Normal Development and Cancer. 10.1146/annurev-cellbio-092910-154234 2011. [CrossRef]
- Quinton, R.J.; DiDomizio, A.; Vittoria, M.A.; Kotýnková, K.; Ticas, C.J.; Patel, S.; Koga, Y.; Vakhshoorzadeh, J.; Hermance, N.; Kuroda, T.S.; et al. Whole-Genome Doubling Confers Unique Genetic Vulnerabilities on Tumour Cells. Nature 2021, 590, 492–497. [CrossRef]
- Storchova, Z.; Pellman, D. From Polyploidy to Aneuploidy, Genome Instability and Cancer. Nat Rev Mol Cell Bio 2004, 5, 45–54. [CrossRef]
- Storchová, Z.; Breneman, A.; Cande, J.; Dunn, J.; Burbank, K.; O’Toole, E.; Pellman, D. Genome-Wide Genetic Analysis of Polyploidy in Yeast. Nature 2006, 443, 541–547. [CrossRef]
- Nguyen, H.G.; Makitalo, M.; Yang, D.; Chinnappan, D.; St.Hilaire, C.; Ravid, K. Deregulated Aurora-B Induced Tetraploidy Promotes Tumorigenesis. Faseb J 2009, 23, 2741–2748. [CrossRef]
- Bielski, C.M.; Zehir, A.; Penson, A.V.; Donoghue, M.T.A.; Chatila, W.; Armenia, J.; Chang, M.T.; Schram, A.M.; Jonsson, P.; Bandlamudi, C.; et al. Genome Doubling Shapes the Evolution and Prognosis of Advanced Cancers. Nat Genet 2018, 50, 1189–1195. [CrossRef]
- Clarke, E.K.; Gomez, K.A.R.; Mustachi, Z.; Murph, M.C.; Schvarzstein, M. Manipulation of Ploidy in <em>Caenorhabditis Elegans</Em>. J Vis Exp 2018, 57296. [CrossRef]
- Kawade, K.; Horiguchi, G.; Tsukaya, H. Non-Cell-Autonomously Coordinated Organ Size Regulation in Leaf Development. Development 2010, 137, 4221–4227. [CrossRef]
- Horiguchi, G.; Tsukaya, H. Organ Size Regulation in Plants: Insights from Compensation. Front Plant Sci 2011, 2, 24. [CrossRef]
- Bailey, E.C.; Kobielski, S.; Park, J.; Losick, V.P. Polyploidy in Tissue Repair and Regeneration. Csh Perspect Biol 2021, 13. [CrossRef]
- Donne, R.; Saroul-Aïnama, M.; Cordier, P.; Celton-Morizur, S.; Desdouets, C. Polyploidy in Liver Development, Homeostasis and Disease. Nat Rev Gastroentero 2020, 17, 391–405. [CrossRef]
- Øvrebø, J.I.; Edgar, B.A. Polyploidy in Tissue Homeostasis and Regeneration. Development 2018, 145, dev156034. [CrossRef]
- Besen-McNally, R.; Gjelsvik, K.J.; Losick, V.P. Wound-Induced Polyploidization Is Dependent on Integrin-Yki Signaling. Biol Open 2021, 10, bio055996. [CrossRef]
- Lin, Y.-H.; Zhang, S.; Zhu, M.; Lu, T.; Chen, K.; Wen, Z.; Wang, S.; Xiao, G.; Luo, D.; Jia, Y.; et al. Mice With Increased Numbers of Polyploid Hepatocytes Maintain Regenerative Capacity But Develop Fewer Hepatocellular Carcinomas Following Chronic Liver Injury. Gastroenterology 2020, 158, 1698-1712.e14. [CrossRef]
- Losick, V.P.; Fox, D.T.; Spradling, A.C. Polyploidization and Cell Fusion Contribute to Wound Healing in the Adult Drosophila Epithelium. Curr Biol 2013, 23, 2224–2232. [CrossRef]
- irsten, H.W. Wound Healing Is a First Response in a Cancerous Pathway: Hyperplasia Developments to 4n Cell Cycling in Dysplasia Linked to Rb-Inactivation. J Cancer Ther 2015, 06, 906–916. [CrossRef]
- Losick, V.P. Wound-Induced Polyploidy Is Required for Tissue Repair. Adv Wound Care 2016, 5, 271–278. [CrossRef]
- Berman, J. Ploidy Plasticity: A Rapid and Reversible Strategy for Adaptation to Stress. FEMS Yeast Res 2016, 16. [CrossRef]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The Frequency of Polyploid Speciation in Vascular Plants. Proc National Acad Sci 2009, 106, 13875–13879. [CrossRef]
- Akagi, T.; Jung, K.; Masuda, K.; Shimizu, K.K. Polyploidy before and after Domestication of Crop Species. Curr Opin Plant Biol 2022, 69, 102255. [CrossRef]
- Hodgkin, J. Exploring the Envelope. Systematic Alteration in the Sex-Determination System of the Nematode Caenorhabditis Elegans. Genetics 2002, 162, 767–780.
- Hodgkin, J. Karyotype, Ploidy, and Gene Dosage. WormBook 2005.
- Meneely, P.M. Sex Determination in Polyploids of Caenorhabditis Elegans. Genetics 1994, 137, 467–481.
- Madl, J.E.; Herman, R.K. POLYPLOIDS AND SEX DETERMINATION IN CAENORHABDITIS ELEGANS. Genetics 1979, 93, 393–402.
- Lozano, E.; Sáez, A.G.; Flemming, A.J.; Cunha, A.; Leroi, A.M. Regulation of Growth by Ploidy in Caenorhabditis Elegans. Current Biology 2006, 16, 493–498. [CrossRef]
- Hara, Y.; Kimura, A. An Allometric Relationship between Mitotic Spindle Width, Spindle Length, and Ploidy in Caenorhabditis Elegans Embryos. Mol. Biol. Cell 2013, 24, 1411–1419. [CrossRef]
- Woodruff, G.C.; Eke, O.; Baird, S.E.; Félix, M.-A.; Haag, E.S. Insights Into Species Divergence and the Evolution of Hermaphroditism From Fertile Interspecies Hybrids of Caenorhabditis Nematodes. Genetics 2010, 186, 997–1012. [CrossRef]
- Vargas, E.; McNally, K.; Friedman, J.A.; Cortes, D.B.; Wang, D.Y.; Korf, I.F.; McNally, F.J. Autosomal Trisomy and Triploidy Are Corrected During Female Meiosis in Caenorhabditis Elegans. Genetics 2017, genetics.300259.2017. [CrossRef]
- Ozugergin, I.; Mastronardi, K.; Law, C.; Piekny, A. Diverse Mechanisms Regulate Contractile Ring Assembly for Cytokinesis in the Two-Cell Caenorhabditis Elegans Embryo. J Cell Sci 2021, 135. [CrossRef]
- Mlynarczyk-Evans, S.; Roelens, B.; Villeneuve, A.M. Evidence That Masking of Synapsis Imperfections Counterbalances Quality Control to Promote Efficient Meiosis. PLoS Genet 2013, 9, e1003963. [CrossRef]
- Roelens, B.; Schvarzstein, M.; Villeneuve, A.M. Manipulation of Karyotype in Caenorhabditis Elegans Reveals Multiple Inputs Driving Pairwise Chromosome Synapsis During Meiosis. Genetics 2015, 201, 1363–1379. [CrossRef]
- Cortes, D.B.; McNally, K.L.; Mains, P.E.; McNally, F.J. The Asymmetry of Female Meiosis Reduces the Frequency of Inheritance of Unpaired Chromosomes. Elife 2015.
- Orr-Weaver, T.L. When Bigger Is Better: The Role of Polyploidy in Organogenesis. Trends in Genetics 2015, 31, 307–315. [CrossRef]
- Frawley, L.E.; Orr-Weaver, T.L. Polyploidy. Current Biology 2015, 25, R353–R358. [CrossRef]
- Chen, H.-Z.; Ouseph, M.M.; Li, J.; Pécot, T.; Chokshi, V.; Kent, L.; Bae, S.; Byrne, M.; Duran, C.; Comstock, G.; et al. Canonical and Atypical E2Fs Regulate the Mammalian Endocycle. Nat Cell Biol 2012, 14, 1192–1202. [CrossRef]
- Rios, A.C.; Fu, N.Y.; Jamieson, P.R.; Pal, B.; Whitehead, L.; Nicholas, K.R.; Lindeman, G.J.; Visvader, J.E. Essential Role for a Novel Population of Binucleated Mammary Epithelial Cells in Lactation. Nat Commun 2016, 7, 11400. [CrossRef]
- Pandit, S.K.; Westendorp, B.; Nantasanti, S.; Liere, E. van; Tooten, P.C.J.; Cornelissen, P.W.A.; Toussaint, M.J.M.; Lamers, W.H.; Bruin, A. de E2F8 Is Essential for Polyploidization in Mammalian Cells. Nat Cell Biol 2012, 14, 1181–1191. [CrossRef]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.-D.; Guerquin-Kern, J.-L.; Lechene, C.P.; Lee, R.T. Mammalian Heart Renewal by Pre-Existing Cardiomyocytes. Nature 2013, 493, 433–436. [CrossRef]
- Zanet, J.; Freije, A.; Ruiz, M.; Coulon, V.; Sanz, J.R.; Chiesa, J.; Gandarillas, A. A Mitosis Block Links Active Cell Cycle with Human Epidermal Differentiation and Results in Endoreplication. Plos One 2010, 5, e15701. [CrossRef]
- Mattia, G.; Vulcano, F.; Milazzo, L.; Barca, A.; Macioce, G.; Giampaolo, A.; Hassan, H.J. Different Ploidy Levels of Megakaryocytes Generated from Peripheral or Cord Blood CD34+ Cells Are Correlated with Different Levels of Platelet Release. Blood 2002, 99, 888–897. [CrossRef]
- Zimmet, J.; Ravid, K. Polyploidy Occurrence in Nature, Mechanisms, and Significance for the Megakaryocyte-Platelet System. Exp Hematol 2000, 28, 3–16. [CrossRef]
- Sladky, V.C.; Eichin, F.; Reiberger, T.; Villunger, A. Polyploidy Control in Hepatic Health and Disease. J Hepatol 2021, 75, 1177–1191. [CrossRef]
- Mazzi, S.; Lordier, L.; Debili, N.; Raslova, H.; Vainchenker, W. Megakaryocyte and Polyploidization. Exp Hematol 2018, 57, 1–13. [CrossRef]
- Edgar, B.A.; Zielke, N.; Gutierrez, C. Endocycles: A Recurrent Evolutionary Innovation for Post-Mitotic Cell Growth. Nat Rev Mol Cell Bio 2014, 15, 197–210. [CrossRef]
- Doyle, J.J.; Coate, J.E. Polyploidy, the Nucleotype, and Novelty: The Impact of Genome Doubling on the Biology of the Cell. Int J Plant Sci 2019, 180, 1–52. [CrossRef]
- Ren, R.; Wang, H.; Guo, C.; Zhang, N.; Zeng, L.; Chen, Y.; Ma, H.; Qi, J. Widespread Whole Genome Duplications Contribute to Genome Complexity and Species Diversity in Angiosperms. Mol Plant 2018, 11, 414–428. [CrossRef]
- Fox, D.T.; Soltis, D.E.; Soltis, P.S.; Ashman, T.-L.; Peer, Y.V. de Polyploidy: A Biological Force From Cells to Ecosystems. Trends Cell Biol 2020, 30, 688–694. [CrossRef]
- Maciver, S.K. Ancestral Eukaryotes Reproduced Asexually, Facilitated by Polyploidy: A Hypothesis. Bioessays 2019, 41, e1900152. [CrossRef]
- Rijnberk, L.M. van; Barrull-Mascaró, R.; Palen, R.L. van der; Schild, E.S.; Korswagen, H.C.; Galli, M. Endomitosis Controls Tissue-Specific Gene Expression during Development. Plos Biol 2022, 20, e3001597. [CrossRef]
- Zhang, J.; Qiao, Q.; Xu, H.; Zhou, R.; Liu, X. Human Cell Polyploidization: The Good and the Evil. Semin Cancer Biol 2022, 81, 54–63. [CrossRef]
- Otto, S.P. The Evolutionary Consequences of Polyploidy. Cell 2007, 131, 452–462. [CrossRef]
- Lazzeri, E.; Angelotti, M.L.; Peired, A.; Conte, C.; Marschner, J.A.; Maggi, L.; Mazzinghi, B.; Lombardi, D.; Melica, M.E.; Nardi, S.; et al. Endocycle-Related Tubular Cell Hypertrophy and Progenitor Proliferation Recover Renal Function after Acute Kidney Injury. Nat Commun 2018, 9, 1344. [CrossRef]
- Li, T.; Wu, Y.; Tsai, H.; Sun, C.; Wu, Y.; Wu, H.; Pei, Y.; Lu, K.; Yen, T.T.; Chang, C.; et al. Intrahepatic Hepatitis B Virus Large Surface Antigen Induces Hepatocyte Hyperploidy via Failure of Cytokinesis. J. Pathol. 2018, 245, 502–513. [CrossRef]
- David, K.T.; Halanych, K.M. Spatial Proximity between Polyploids across South American Frog Genera. J Biogeogr 2021, 48, 991–1000. [CrossRef]
- Glennon, K.L.; Ritchie, M.E.; Segraves, K.A. Evidence for Shared Broad-scale Climatic Niches of Diploid and Polyploid Plants. Ecol Lett 2014, 17, 574–582. [CrossRef]
- Arnold, B.; Kim, S.-T.; Bomblies, K. Single Geographic Origin of a Widespread Autotetraploid Arabidopsis Arenosa Lineage Followed by Interploidy Admixture. Mol. Biol. Evol. 2015, 32, 1382–1395. [CrossRef]
- Molina-Henao, Y.F.; Hopkins, R. Autopolyploid Lineage Shows Climatic Niche Expansion but Not Divergence in Arabidopsis Arenosa. Am J Bot 2019, 106, 61–70. [CrossRef]
- Ramsey, J. Polyploidy and Ecological Adaptation in Wild Yarrow. Proc National Acad Sci 2011, 108, 7096–7101. [CrossRef]
- Martin, S.L.; Husband, B.C. ADAPTATION OF DIPLOID AND TETRAPLOID CHAMERION ANGUSTIFOLIUM TO ELEVATION BUT NOT LOCAL ENVIRONMENT: ADAPTATION OF CHAMERION ANGUSTIFOLIUM PLOIDIES. Evolution 2013, 67, 1780–1791. [CrossRef]
- Schoenfelder, K.P.; Fox, D.T. The Expanding Implications of Polyploidy. J Cell Biol 2015, 209, 485–491. [CrossRef]
- Bomblies, K. When Everything Changes at Once: Finding a New Normal after Genome Duplication. Proc Royal Soc B Biological Sci 2020, 287, 20202154. [CrossRef]
- Selmecki, A.M.; Maruvka, Y.E.; Richmond, P.A.; Guillet, M.; Shoresh, N.; Sorenson, A.L.; De, S.; Kishony, R.; Michor, F.; Dowell, R.; et al. Polyploidy Can Drive Rapid Adaptation in Yeast. Nature 2015, 519, 349–352. [CrossRef]
- Peer, Y.V. de; Mizrachi, E.; Marchal, K. The Evolutionary Significance of Polyploidy. Nat Rev Genet 2017, 18, 411–424. [CrossRef]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhang, C.-Z.; Wala, J.; Mermel, C.H.; et al. Pan-Cancer Patterns of Somatic Copy Number Alteration. Nat Genet 2013, 45, 1134–1140. [CrossRef]
- Sansregret, L.; Vanhaesebroeck, B.; Swanton, C. Determinants and Clinical Implications of Chromosomal Instability in Cancer. Nat Rev Clin Oncol 2018, 15, 139–150. [CrossRef]
- Fujiwara, T.; Bandi, M.; Nitta, M.; Ivanova, E.V.; Bronson, R.T.; Pellman, D. Cytokinesis Failure Generating Tetraploids Promotes Tumorigenesis in P53-Null Cells. Nature 2005, 437, 1043–1047. [CrossRef]
- Matsumoto, T.; Wakefield, L.; Peters, A.; Peto, M.; Spellman, P.; Grompe, M. Proliferative Polyploid Cells Give Rise to Tumors via Ploidy Reduction. Nat Commun 2021, 12, 646. [CrossRef]
- Lambuta, R.A.; Nanni, L.; Liu, Y.; Diaz-Miyar, J.; Iyer, A.; Tavernari, D.; Katanayeva, N.; Ciriello, G.; Oricchio, E. Whole-Genome Doubling Drives Oncogenic Loss of Chromatin Segregation. Nature 2023, 1–9. [CrossRef]
- Dewhurst, S.M.; McGranahan, N.; Burrell, R.A.; Rowan, A.J.; Grönroos, E.; Endesfelder, D.; Joshi, T.; Mouradov, D.; Gibbs, P.; Ward, R.L.; et al. Tolerance of Whole-Genome Doubling Propagates Chromosomal Instability and Accelerates Cancer Genome Evolution. Cancer Discov 2014, 4, 175–185. [CrossRef]
- Adams, K.L.; Wendel, J.F. Polyploidy and Genome Evolution in Plants. Current Opinion in Plant Biology 2005, 8, 135–141. [CrossRef]
- Cheng, A.; Hanafiah, N.M.; Harikrishna, J.A.; Eem, L.P.; Baisakh, N.; Mispan, M.S. A Reappraisal of Polyploidy Events in Grasses (Poaceae) in a Rapidly Changing World. Biology 2022, 11, 636. [CrossRef]
- Park, C.-W.; Bhandari, G.S.; Won, H.; Park, J.H.; Park, D.S. Polyploidy and Introgression in Invasive Giant Knotweed (Fallopia Sachalinensis) during the Colonization of Remote Volcanic Islands. Sci Rep-uk 2018, 8, 16021. [CrossRef]
- Demin, S.Yu.; Berdieva, M.A.; Goodkov, A.V. Cyclic Polyploidy in Obligate Agamic Amoebae. Cell Tissue Biology 2019, 13, 242–246. [CrossRef]
- Albertin, W.; Marullo, P. Polyploidy in Fungi: Evolution after Whole-Genome Duplication. Proc Royal Soc B Biological Sci 2012, 279, 2497–2509. [CrossRef]
- Soppa, J. Polyploidy in Archaea and Bacteria: About Desiccation Resistance, Giant Cell Size, Long-Term Survival, Enforcement by a Eukaryotic Host and Additional Aspects. J Mol Microb Biotech 2015, 24, 409–419. [CrossRef]
- Markov, A.V.; Kaznacheev, I.S. Evolutionary Consequences of Polyploidy in Prokaryotes and the Origin of Mitosis and Meiosis. Biol Direct 2016, 11, 28. [CrossRef]
- Wertheim, B.; Beukeboom, L.W.; Zande, L. van de Polyploidy in Animals: Effects of Gene Expression on Sex Determination, Evolution and Ecology. Cytogenet Genome Res 2013, 140, 256–269. [CrossRef]
- Mable, B.K.; Alexandrou, M.A.; Taylor, M.I. Genome Duplication in Amphibians and Fish: An Extended Synthesis. J Zool 2011, 284, 151–182. [CrossRef]
- Sacerdot, C.; Louis, A.; Bon, C.; Berthelot, C.; Crollius, H.R. Chromosome Evolution at the Origin of the Ancestral Vertebrate Genome. Genome Biol 2018, 19, 166. [CrossRef]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat Rev Genet 2005, 6, 836–846. [CrossRef]
- Monnahan, P.; Kolář, F.; Baduel, P.; Sailer, C.; Koch, J.; Horvath, R.; Laenen, B.; Schmickl, R.; Paajanen, P.; Šrámková, G.; et al. Pervasive Population Genomic Consequences of Genome Duplication in Arabidopsis Arenosa. Nat Ecol Evol 2019, 3, 457–468. [CrossRef]
- Yamazaki, W.; Takahashi, M.; Kawahara, M. Restricted Development of Mouse Triploid Fetuses with Disorganized Expression of Imprinted Genes. Zygote 2015, 23, 874–884. [CrossRef]
- Menon, T.; Nair, S. Experimental Manipulation of Ploidy in Zebrafish Embryos and Its Application in Genetic Screens. Methods Mol Biology Clifton N J 2019, 1920, 111–128. [CrossRef]
- Eakin, G.S.; Hadjantonakis, A.-K.; Papaioannou, V.E.; Behringer, R.R. Developmental Potential and Behavior of Tetraploid Cells in the Mouse Embryo. Dev Biol 2005, 288, 150–159. [CrossRef]
- Nigon; V. Les Modalites de La Reproduction et Le Determinisme Du Sexe Chez Quelques Nematodes Libres. Annales de Sciences Naturelles - Zool. Biol. Anim. 1949, 11, 1–132.
- Hodgkin, J. MALE PHENOTYPES AND MATING EFFICIENCY IN CAENORHABDITIS ELEGANS. Genetics 1983, 103, 43–64. [CrossRef]
- Hodgkin, J. Primary Sex Determination in the Nematode C. Elegans. Development 1987, 101, 5–16.
- Nigon, V.M.; Félix, M.-A. History of Research on C. Elegans and Other Free-Living Nematodes as Model Organisms. Wormbook Online Rev C Elegans Biology 2017, 2017, 1–84. [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94.
- Hedgecock, E.M.; White, J.G. Polyploid Tissues in the Nematode Caenorhabditis Elegans. Developmental Biology 1985, 107, 128–133. [CrossRef]
- Flemming, A.J.; Shen, Z.-Z.; Cunha, A.; Emmons, S.W.; Leroi, A.M. Somatic Polyploidization and Cellular Proliferation Drive Body Size Evolution in Nematodes. Proceedings of the National Academy of Sciences 2000, 97, 5285–5290. [CrossRef]
- Hodgkin, J.; Horvitz, H.R.; Brenner, S. NONDISJUNCTION MUTANTS OF THE NEMATODE CAENORHABDITIS ELEGANS. Genetics 1979, 91, 67–94. [CrossRef]
- Sigurdson, D.C.; Spanier, G.J.; Herman, R.K. CAENORHABDITIS ELEGANS DEFICIENCY MAPPING. Genetics 1984, 108, 331–345. [CrossRef]
- Haack, H.; Hodgkin, J. Tests for Parental Imprinting in the Nematode Caenorhabditis Elegans. Molec. Gen. Genet. 1991, 228, 482–485. [CrossRef]
- Larkin, K.; Tucci, C.; Neiman, M. Effects of Polyploidy and Reproductive Mode on Life History Trait Expression. Ecol Evol 2016, 6, 765–778. [CrossRef]
- Schoonmaker, A.; Hao, Y.; Bird, D.McK.; Conant, G.C. A Single, Shared Triploidy in Three Species of Parasitic Nematodes. G3 Genes Genomes Genetics 2019, 10, 225–233. [CrossRef]
- Nigon; V. Effets de La Polyploïdie Chez Un Nématode Libre. C. R. Acad. Sc. Paris 1949, D 228, 1161-1162.
- Nigon, V. Polyploidie Experimentale Chez Un Nematode Libre, Rhabditis Elegans. Maupas. Bull. Biol. Fr. Belg. 1951, 85, 187–225. [CrossRef]
- V, N. Effets de La Polyploïdie Chez Un Nématode Libre. C. R. Acad. Sc. 1949, 228, 1161-1162.
- Meyer, B.J. X-Chromosome Dosage Compensation. Wormbook 2005, 1–14. [CrossRef]
- Meyer, B.J. The X Chromosome in C. Elegans Sex Determination and Dosage Compensation. Curr Opin Genet Dev 2022, 74, 101912. [CrossRef]
- Schvarzstein, M.; Wignall, S.M.; Villeneuve, A.M. Coordinating Cohesion, Co-Orientation, and Congression during Meiosis: Lessons from Holocentric Chromosomes. Genes & Development 2010, 24, 219–228. [CrossRef]
- MacQueen, A.J.; Phillips, C.M.; Bhalla, N.; Weiser, P.; Villeneuve, A.M.; Dernburg, A.F. Chromosome Sites Play Dual Roles to Establish Homologous Synapsis during Meiosis in C. Elegans. Cell 2005, 123, 1037–1050. [CrossRef]
- Meyerzon, M.; Gao, Z.; Liu, J.; Wu, J.-C.; Malone, C.J.; Starr, D.A. Centrosome Attachment to the C. Elegans Male Pronucleus Is Dependent on the Surface Area of the Nuclear Envelope. Developmental Biology 2009, 327, 433–446. [CrossRef]
- Schierenberg, E.; Wood, W.B. Control of Cell-Cycle Timing in Early Embryos of Caenorhabditis Elegans. Dev Biol 1985, 107, 337–354. [CrossRef]
- Sadler, P.L.; Shakes, D.C. Anucleate Caenorhabditis Elegans Sperm Can Crawl, Fertilize Oocytes and Direct Anterior-Posterior Polarization of the 1-Cell Embryo. Development 2000, 127, 355–366. [CrossRef]
- Schierenberg, E. Cell Fusion. 1987, 409–418. [CrossRef]
- Oegema, K.; Hyman, A.A. Cell Division. Wormbook 2006, 1–40. [CrossRef]
- Malone, C.J.; Misner, L.; Bot, N.L.; Tsai, M.-C.; Campbell, J.M.; Ahringer, J.; White, J.G. The C. Elegans Hook Protein, ZYG-12, Mediates the Essential Attachment between the Centrosome and Nucleus. Cell 2003, 115, 825–836. [CrossRef]
- Gönczy, P.; Pichler, S.; Kirkham, M.; Hyman, A.A. Cytoplasmic Dynein Is Required for Distinct Aspects of Mtoc Positioning, Including Centrosome Separation, in the One Cell Stage Caenorhabditis Elegans Embryo. J Cell Biology 1999, 147, 135–150. [CrossRef]
- Newport, J.; Kirschner, M. A Major Developmental Transition in Early Xenopus Embryos: I. Characterization and Timing of Cellular Changes at the Midblastula Stage. Cell 1982, 30, 675–686. [CrossRef]
- Sachsenmaier, W.; Remy, U.; Plattner-Schobel, R. Initiation of Synchronous Mitosis in Physarum Polycephalum A Model of the Control of Cell Division in Eukariots. Exp Cell Res 1972, 73, 41–48. [CrossRef]
- Fantes, P.A. Isolation of Cell Size Mutants of a Fission Yeast by a New Selective Method: Characterization of Mutants and Implications for Division Control Mechanisms. J Bacteriol 1981, 146, 746–754. [CrossRef]
- Matson, J.P.; Cook, J.G. Cell Cycle Proliferation Decisions: The Impact of Single Cell Analyses. Febs J 2017, 284, 362–375. [CrossRef]
- Arata, Y.; Takagi, H. Quantitative Studies for Cell-Division Cycle Control. Front Physiol 2019, 10, 1022. [CrossRef]
- Koreth, J.; Heuvel, S. van den Cell-Cycle Control in Caenorhabditis Elegans: How the Worm Moves from G1 to S. Oncogene 2005, 24, 2756–2764. [CrossRef]
- Wang, Z. Regulation of Cell Cycle Progression by Growth Factor-Induced Cell Signaling. Cells 2021, 10, 3327. [CrossRef]
- Dumont, S.; Mitchison, T.J. Compression Regulates Mitotic Spindle Length by a Mechanochemical Switch at the Poles. Curr Biol 2009, 19, 1086–1095. [CrossRef]
- Itabashi, T.; Takagi, J.; Shimamoto, Y.; Onoe, H.; Kuwana, K.; Shimoyama, I.; Gaetz, J.; Kapoor, T.M.; Ishiwata, S. Probing the Mechanical Architecture of the Vertebrate Meiotic Spindle. Nat Methods 2009, 6, 167–172. [CrossRef]
- Hara, Y.; Iwabuchi, M.; Ohsumi, K.; Kimura, A. Intranuclear DNA Density Affects Chromosome Condensation in Metazoans. Mol Biol Cell 2013, 24, 2442–2453. [CrossRef]
- Mains, P.E.; Kemphues, K.J.; Sprunger, S.A.; Sulston, I.A.; Wood, W.B. Mutations Affecting the Meiotic and Mitotic Divisions of the Early Caenorhabditis Elegans Embryo. Genetics 1990, 126, 593–605. [CrossRef]
- Quintin, S.; Mains, P.E.; Zinke, A.; Hyman, A.A. The Mbk-2 Kinase Is Required for Inactivation of MEI-1/Katanin in the One-cell Caenorhabditis Elegans Embryo. Embo Rep 2003, 4, 1175–1181. [CrossRef]
- Segbert, C.; Barkus, R.; Powers, J.; Strome, S.; Saxton, W.M.; Bossinger, O. KLP-18, a Klp2 Kinesin, Is Required for Assembly of Acentrosomal Meiotic Spindles in Caenorhabditis Elegans. Mol Biol Cell 2003, 14, 4458–4469. [CrossRef]
- Maddox, A.S.; Habermann, B.; Desai, A.; Oegema, K. Distinct Roles for Two C. Elegans Anillins in the Gonad and Early Embryo. Development 2005, 132, 2837–2848. [CrossRef]
- Dorn, J.F.; Zhang, L.; Paradis, V.; Edoh-Bedi, D.; Jusu, S.; Maddox, P.S.; Maddox, A.S. Actomyosin Tube Formation in Polar Body Cytokinesis Requires Anillin in C. Elegans. Curr Biol 2010, 20, 2046–2051. [CrossRef]
- Fankhauser, G. Cytological Studies on Egg Fragments of the Salamander Triton. V. Chromosome Number and Chromosome Individuality in the Cleavage Mitoses of Merogonic Fragments. J. Exp. Zool. 1934, 68, 1–57. [CrossRef]
- Conklin, E.G. Cell Size and Nuclear Size. J. Exp. Zool. 1912, 12, 1–98. [CrossRef]
- Berardino, M.A.D. The Karyotype of Rana Pipiens and Investigation of Its Stability during Embryonic Differentiation. Dev Biol 1962, 5, 101–126. [CrossRef]
- Belmont, A.S.; Sedat, J.W.; Agard, D.A. A Three-Dimensional Approach to Mitotic Chromosome Structure: Evidence for a Complex Hierarchical Organization. J Cell Biology 1987, 105, 77–92. [CrossRef]
- Sönnichsen, B.; Koski, L.B.; Walsh, A.; Marschall, P.; Neumann, B.; Brehm, M.; Alleaume, A.-M.; Artelt, J.; Bettencourt, P.; Cassin, E.; et al. Full-Genome RNAi Profiling of Early Embryogenesis in Caenorhabditis Elegans. Nature 2005, 434, 462–469. [CrossRef]
- Deppe, U.; Schierenberg, E.; Cole, T.; Krieg, C.; Schmitt, D.; Yoder, B.; Ehrenstein, G. von Cell Lineages of the Embryo of the Nematode Caenorhabditis Elegans. Proc National Acad Sci 1978, 75, 376–380. [CrossRef]
- Sulston, J.E.; Schierenberg, E.; White, J.G.; Thomson, J.N. The Embryonic Cell Lineage of the Nematode Caenorhabditis Elegans. Dev Biol 1983, 100, 64–119. [CrossRef]
- I., G. Development of the Vulva. In C. elegans II; R., R.L., T., B., J., M.B., R., P.J., Eds.; Cold Spring Harbor Laboratory Press, Cold Spring Harbor: New York, 1997; Vol. Chapter 19, pp. 519–541.
- Simske, J.S.; Hardin, J. Getting into Shape: Epidermal Morphogenesis in Caenorhabditis Elegans Embryos. Bioessays 2001, 23, 12–23. [CrossRef]
- Wong, M.; Schwarzbauer, J.E. Gonad Morphogenesis and Distal Tip Cell Migration in the Caenorhabditis Elegans Hermaphrodite. Wiley Interdiscip Rev Dev Biology 2012, 1, 519–531. [CrossRef]
- Shemer, G.; Podbilewicz, B. Fusomorphogenesis: Cell Fusion in Organ Formation. Dev. Dyn. 2000, 218, 30–51. [CrossRef]
- Tuck, S. The Control of Cell Growth and Body Size in Caenorhabditis Elegans. Exp Cell Res 2014, 321, 71–76. [CrossRef]
- Tsou, M.-F.B.; Stearns, T. Mechanism Limiting Centrosome Duplication to Once per Cell Cycle. Nature 2006, 442, 947–951. [CrossRef]
- Tsou, M.-F.B.; Stearns, T. Controlling Centrosome Number: Licenses and Blocks. Current Opinion in Cell Biology 2006, 18, 74–78. [CrossRef]
- Duensing, A.; Duensing, S. Polyploidization and Cancer. Adv Exp Med Biol 2010, 676, 93–103. [CrossRef]
- Lu, Y.; Roy, R. Centrosome/Cell Cycle Uncoupling and Elimination in the Endoreduplicating Intestinal Cells of C. Elegans. PLoS ONE 2014, 9, e110958-17. [CrossRef]
- Lee, H.O.; Davidson, J.M.; Duronio, R.J. Endoreplication: Polyploidy with Purpose. Gene Dev 2009, 23, 2461–2477. [CrossRef]
- Zhang, L.; Ward, J.D.; Cheng, Z.; Dernburg, A.F. The Auxin-Inducible Degradation (AID) System Enables Versatile Conditional Protein Depletion in C. Elegans. Development 2015, 142, 4374–4384. [CrossRef]
- Jorgensen, P.; Edgington, N.P.; Schneider, B.L.; Rupeš, I.; Tyers, M.; Futcher, B. The Size of the Nucleus Increases as Yeast Cells Grow. Mol Biol Cell 2007, 18, 3523–3532. [CrossRef]
- Yahya, G.; Menges, P.; Amponsah, P.S.; Ngandiri, D.A.; Schulz, D.; Wallek, A.; Kulak, N.; Mann, M.; Cramer, P.; Savage, V.; et al. Sublinear Scaling of the Cellular Proteome with Ploidy. Nat Commun 2022, 13, 6182. [CrossRef]
- Neumann, F.R.; Nurse, P. Nuclear Size Control in Fission Yeast. J Cell Biology 2007, 179, 593–600. [CrossRef]
- Gillooly, J.F.; Hein, A.; Damiani, R. Nuclear DNA Content Varies with Cell Size across Human Cell Types. Csh Perspect Biol 2015, 7, a019091. [CrossRef]
- Katagiri, Y.; Hasegawa, J.; Fujikura, U.; Hoshino, R.; Matsunaga, S.; Tsukaya, H. The Coordination of Ploidy and Cell Size Differs between Cell Layers in Leaves. Development 2016, 143, 1120–1125. [CrossRef]
- Cadart, C.; Heald, R. Scaling of Biosynthesis and Metabolism with Cell Size. Mol Biol Cell 2022, 33, pe5. [CrossRef]
- Robinson, D.O.; Coate, J.E.; Singh, A.; Hong, L.; Bush, M.; Doyle, J.J.; Roeder, A.H.K. Ploidy and Size at Multiple Scales in the Arabidopsis Sepal. Plant Cell 2018, 30, 2308–2329. [CrossRef]
- Cantwell, H.; Dey, G. Nuclear Size and Shape Control. Semin Cell Dev Biol 2022, 130, 90–97. [CrossRef]
- Weber, G.M.; Wiens, G.D.; Welch, T.J.; Hostuttler, M.A.; Leeds, T.D. Comparison of Disease Resistance between Diploid, Induced-Triploid, and Intercross-Triploid Rainbow Trout Including Trout Selected for Resistance to Flavobacterium Psychrophilum. Aquaculture 2013, 410–411, 66–71. [CrossRef]
- Horiguchi, G.; Tsukaya, H. Organ Size Regulation in Plants: Insights from Compensation. Front Plant Sci 2011, 2, 24. [CrossRef]
- Kawade, K.; Horiguchi, G.; Tsukaya, H. Non-Cell-Autonomously Coordinated Organ Size Regulation in Leaf Development. Development 2010, 137, 4221–4227. [CrossRef]
- Tsukaya, H. Re-Examination of the Role of Endoreduplication on Cell-Size Control in Leaves. J Plant Res 2019, 132, 571–580. [CrossRef]
- Nyström, J.; Shen, Z.-Z.; Aili, M.; Flemming, A.J.; Leroi, A.; Tuck, S. Increased or Decreased Levels of Caenorhabditis Elegans Lon-3, a Gene Encoding a Collagen, Cause Reciprocal Changes in Body Length. Genetics 2002, 161, 83–97.
- Morita, K.; Flemming, A.J.; Sugihara, Y.; Mochii, M.; Suzuki, Y.; Yoshida, S.; Wood, W.B.; Kohara, Y.; Leroi, A.M.; Ueno, N. A Caenorhabditis Elegans TGF-β, DBL-1, Controls the Expression of LON-1, a PR-related Protein, That Regulates Polyploidization and Body Length. The EMBO Journal 2002, 21, 1063–1073. [CrossRef]
- Morita, K.; Chow, K.L.; Ueno, N. Regulation of Body Length and Male Tail Ray Pattern Formation of Caenorhabditis Elegans by a Member of TGF-Beta Family. Dev Camb Engl 1999, 126, 1337–1347. [CrossRef]
- Hirose, T.; Nakano, Y.; Nagamatsu, Y.; Misumi, T.; Ohta, H.; Ohshima, Y. Cyclic GMP-Dependent Protein Kinase EGL-4 Controls Body Size and Lifespan in C. Elegans. Development 2003, 130, 1089–1099. [CrossRef]
- Nagamatsu, Y.; Ohshima, Y. Mechanisms for the Control of Body Size by a G-kinase and a Downstream TGFβ Signal Pathway in Caenorhabditis Elegans. Genes Cells 2004, 9, 39–47. [CrossRef]
- Wang, J.; Tokarz, R.; Savage-Dunn, C. The Expression of TGFβ Signal Transducers in the Hypodermis Regulates Body Size in C. Elegans. Development 2002, 129, 4989–4998. [CrossRef]
- Parisi, T.; Beck, A.R.; Rougier, N.; McNeil, T.; Lucian, L.; Werb, Z.; Amati, B. Cyclins E1 and E2 Are Required for Endoreplication in Placental Trophoblast Giant Cells. Embo J 2003, 22, 4794–4803. [CrossRef]
- Geng, Y.; Yu, Q.; Sicinska, E.; Das, M.; Schneider, J.E.; Bhattacharya, S.; Rideout, W.M.; Bronson, R.T.; Gardner, H.; Sicinski, P. Cyclin E Ablation in the Mouse. Cell 2003, 114, 431–443. [CrossRef]
- Suzuki, Y.; Yandell, M.D.; Roy, P.J.; Krishna, S.; Savage-Dunn, C.; Ross, R.M.; Padgett, R.W.; Wood, W.B. A BMP Homolog Acts as a Dose-Dependent Regulator of Body Size and Male Tail Patterning in Caenorhabditis Elegans. Development 1999, 126, 241–250. [CrossRef]
- SAVAGE-DUNN, C.; YU, L.; GILL, K.; AWAN, M.; FERNANDO, T. Non-Stringent Tissue-Source Requirements for BMP Ligand Expression in Regulation of Body Size in Caenorhabditis Elegans. Genet Res 2011, 93, 427–432. [CrossRef]
- Savage-Dunn, C. Targets of TGFβ-Related Signaling in Caenorhabditis Elegans. Cytokine Growth F R 2001, 12, 305–312. [CrossRef]
- Savage-Dunn, C.; Tokarz, R.; Wang, H.; Cohen, S.; Giannikas, C.; Padgett, R.W. SMA-3 Smad Has Specific and Critical Functions in DBL-1/SMA-6 TGFβ-Related Signaling. Dev Biol 2000, 223, 70–76. [CrossRef]
- Szyperski, T.; Fernández, C.; Mumenthaler, C.; Wüthrich, K. Structure Comparison of Human Glioma Pathogenesis-Related Protein GliPR and the Plant Pathogenesis-Related Protein P14a Indicates a Functional Link between the Human Immune System and a Plant Defense System. Proc National Acad Sci 1998, 95, 2262–2266. [CrossRef]
- Cai, Q.; Wang, W.; Gao, Y.; Yang, Y.; Zhu, Z.; Fan, Q. Ce-wts-1 Plays Important Roles in Caenorhabditis Elegans Development. Febs Lett 2009, 583, 3158–3164. [CrossRef]
- Jones, K.T.; Greer, E.R.; Pearce, D.; Ashrafi, K. Rictor/TORC2 Regulates Caenorhabditis Elegans Fat Storage, Body Size, and Development through Sgk-1. Plos Biol 2009, 7, e1000060. [CrossRef]
- Oldham, S.; Böhni, R.; Stocker, H.; Brogiolo, W.; Hafen, E. Genetic Control of Size in Drosophila. Philosophical Transactions Royal Soc Lond Ser B Biological Sci 2000, 355, 945–952. [CrossRef]
- Schmelzle, T.; Hall, M.N. TOR, a Central Controller of Cell Growth. Cell 2000, 103, 253–262. [CrossRef]
- ZHANG, W.; LIU, H.T. MAPK Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res 2002, 12, 9–18. [CrossRef]
- Watanabe, N.; Nagamatsu, Y.; Gengyo-Ando, K.; Mitani, S.; Ohshima, Y. Control of Body Size by SMA-5, a Homolog of MAP Kinase BMK1/ERK5, in C. Elegans. Development 2005, 132, 3175–3184. [CrossRef]
- Hyun, S. Body Size Regulation and Insulin-like Growth Factor Signaling. Cell Mol Life Sci 2013, 70, 2351–2365. [CrossRef]
- McKeown, C.; Praitis, V.; Austin, J. Sma-1 Encodes a ΒH-Spectrin Homolog Required for Caenorhabditis Elegans Morphogenesis. Development 1998, 125, 2087–2098. [CrossRef]
- Hayashi, S.; Yokoyama, H.; Tamura, K. Roles of Hippo Signaling Pathway in Size Control of Organ Regeneration. Dev Growth Differ 2015, 57, 341–351. [CrossRef]
- Goodman, M.B.; Savage-Dunn, C. Reciprocal Interactions between Transforming Growth Factor Beta Signaling and Collagens: Insights from Caenorhabditis Elegans. Dev. Dyn. 2022, 251, 47–60. [CrossRef]
- Song, Q.; Ando, A.; Jiang, N.; Ikeda, Y.; Chen, Z.J. Single-Cell RNA-Seq Analysis Reveals Ploidy-Dependent and Cell-Specific Transcriptome Changes in Arabidopsis Female Gametophytes. Genome Biol 2020, 21, 178. [CrossRef]
- Wu, C.-Y.; Rolfe, P.A.; Gifford, D.K.; Fink, G.R. Control of Transcription by Cell Size. Plos Biol 2010, 8, e1000523. [CrossRef]
- Marguerat, S.; Bähler, J. Coordinating Genome Expression with Cell Size. Trends Genet 2012, 28, 560–565. [CrossRef]
- Coate, J.E.; Doyle, J.J. Variation in Transcriptome Size: Are We Getting the Message? Chromosoma 2015, 124, 27–43. [CrossRef]
- LEITCH, I.J.; BENNETT, M.D. Genome Downsizing in Polyploid Plants. Biol J Linn Soc 2004, 82, 651–663. [CrossRef]
- Zenil-Ferguson, R.; Ponciano, J.M.; Burleigh, J.G. Evaluating the Role of Genome Downsizing and Size Thresholds from Genome Size Distributions in Angiosperms. Am J Bot 2016, 103, 1175–1186. [CrossRef]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I.J. Genome Size Diversity and Its Impact on the Evolution of Land Plants. Genes-basel 2018, 9, 88. [CrossRef]
- Cheng, F.; Wu, J.; Cai, X.; Liang, J.; Freeling, M.; Wang, X. Gene Retention, Fractionation and Subgenome Differences in Polyploid Plants. Nat Plants 2018, 4, 258–268. [CrossRef]
- Baduel, P.; Quadrana, L.; Hunter, B.; Bomblies, K.; Colot, V. Relaxed Purifying Selection in Autopolyploids Drives Transposable Element Over-Accumulation Which Provides Variants for Local Adaptation. Nat Commun 2019, 10, 5818. [CrossRef]
- Raina, S.N.; Parida, A.; Koul, K.K.; Salimath, S.S.; Bisht, M.S.; Raja, V.; Khoshoo, T.N. Associated Chromosomal DNA Changes in Polyploids. Genome 1994, 37, 560–564. [CrossRef]
- Lovén, J.; Orlando, D.A.; Sigova, A.A.; Lin, C.Y.; Rahl, P.B.; Burge, C.B.; Levens, D.L.; Lee, T.I.; Young, R.A. Revisiting Global Gene Expression Analysis. Cell 2012, 151, 476–482. [CrossRef]
- Bourdon, M.; Pirrello, J.; Cheniclet, C.; Coriton, O.; Bourge, M.; Brown, S.; Moïse, A.; Peypelut, M.; Rouyère, V.; Renaudin, J.-P.; et al. Evidence for Karyoplasmic Homeostasis during Endoreduplication and a Ploidy-Dependent Increase in Gene Transcription during Tomato Fruit Growth. Development 2012, 139, 3817–3826. [CrossRef]
- Voichek, Y.; Bar-Ziv, R.; Barkai, N. Expression Homeostasis during DNA Replication. Science 2016, 351, 1087–1090. [CrossRef]
Figure 1.
Roles of Whole Genome Duplication (WGD). Diagram depicting key examples of processes affected by polyploidization/WGD. Polyploidy can occur at the cellular, tissue or organismal level. It is a key step in important biological processes affecting cellular metabolism and function, normal development, tissue homeostasis, regeneration, wound healing, organismal adaptation to the environment, and speciation. In addition, polyploidization is can cause and/or prevent disease, repair injury, and promote tissue regeneration during aging, damage, and wound healing.
Figure 1.
Roles of Whole Genome Duplication (WGD). Diagram depicting key examples of processes affected by polyploidization/WGD. Polyploidy can occur at the cellular, tissue or organismal level. It is a key step in important biological processes affecting cellular metabolism and function, normal development, tissue homeostasis, regeneration, wound healing, organismal adaptation to the environment, and speciation. In addition, polyploidization is can cause and/or prevent disease, repair injury, and promote tissue regeneration during aging, damage, and wound healing.
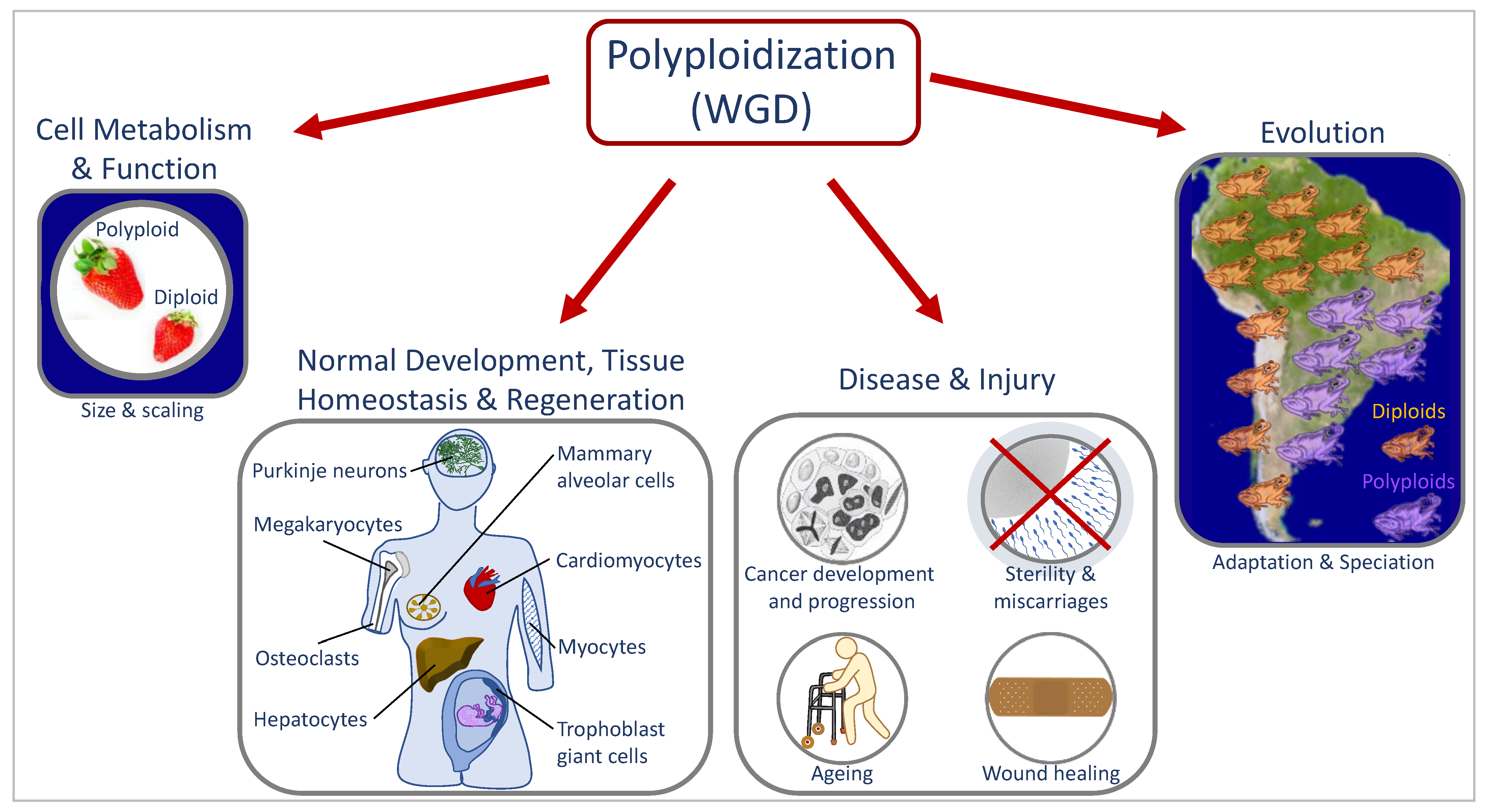
Figure 2.
Method for rapid isolation of C. elegans tetraploids. A). Diagram of the published method for generating tetraploid strains from diploids. This protocol relies on the transient knockdown of the meiosis-specific cohesin rec-8 which results in the production of diploid instead of haploid gametes. Thus upon fertilization, tetraploid progeny (which are longer and bigger) are produced. The method works for both during selfing of the hermaphrodites (left) and during outcrossing (right). Crossing allows for the generation of tetraploids with two different genetic backgrounds. After two to three generations of exposure to rec-8 dsRNA, the larger hermaphrodites are individually plated on normal growth media. Strains that consistently sire large (Lon) hermaphrodites are then screened to confirm these the strains are tetraploid. B). Diploid hermaphrodites (left) produce haploid gametes. Haploid sperm are stored in the spermatheca. Diploid oocytes have 6 pairs of connected homologous chromosomes that can be visualized by DAPI in the diakinesis-stage oocytes (C, below) just adjacent to the spermatheca (-1, -2, -3 oocytes based on position). The oocyte moves through the spermatheca and is fertilized and completes the meiotic divisions to produce a haploid pronucleus. The tetraploid worms derived from exposure to rec-8 dsRNA, (right) produce diploid gametes with 12 DAPI bodies. C) Examples of fixed hermaphrodites stained with DAPI to screen for ploidy by counting chromosome pairs in the -1, -2 and -3 diakinesis oocytes.
Figure 2.
Method for rapid isolation of C. elegans tetraploids. A). Diagram of the published method for generating tetraploid strains from diploids. This protocol relies on the transient knockdown of the meiosis-specific cohesin rec-8 which results in the production of diploid instead of haploid gametes. Thus upon fertilization, tetraploid progeny (which are longer and bigger) are produced. The method works for both during selfing of the hermaphrodites (left) and during outcrossing (right). Crossing allows for the generation of tetraploids with two different genetic backgrounds. After two to three generations of exposure to rec-8 dsRNA, the larger hermaphrodites are individually plated on normal growth media. Strains that consistently sire large (Lon) hermaphrodites are then screened to confirm these the strains are tetraploid. B). Diploid hermaphrodites (left) produce haploid gametes. Haploid sperm are stored in the spermatheca. Diploid oocytes have 6 pairs of connected homologous chromosomes that can be visualized by DAPI in the diakinesis-stage oocytes (C, below) just adjacent to the spermatheca (-1, -2, -3 oocytes based on position). The oocyte moves through the spermatheca and is fertilized and completes the meiotic divisions to produce a haploid pronucleus. The tetraploid worms derived from exposure to rec-8 dsRNA, (right) produce diploid gametes with 12 DAPI bodies. C) Examples of fixed hermaphrodites stained with DAPI to screen for ploidy by counting chromosome pairs in the -1, -2 and -3 diakinesis oocytes.
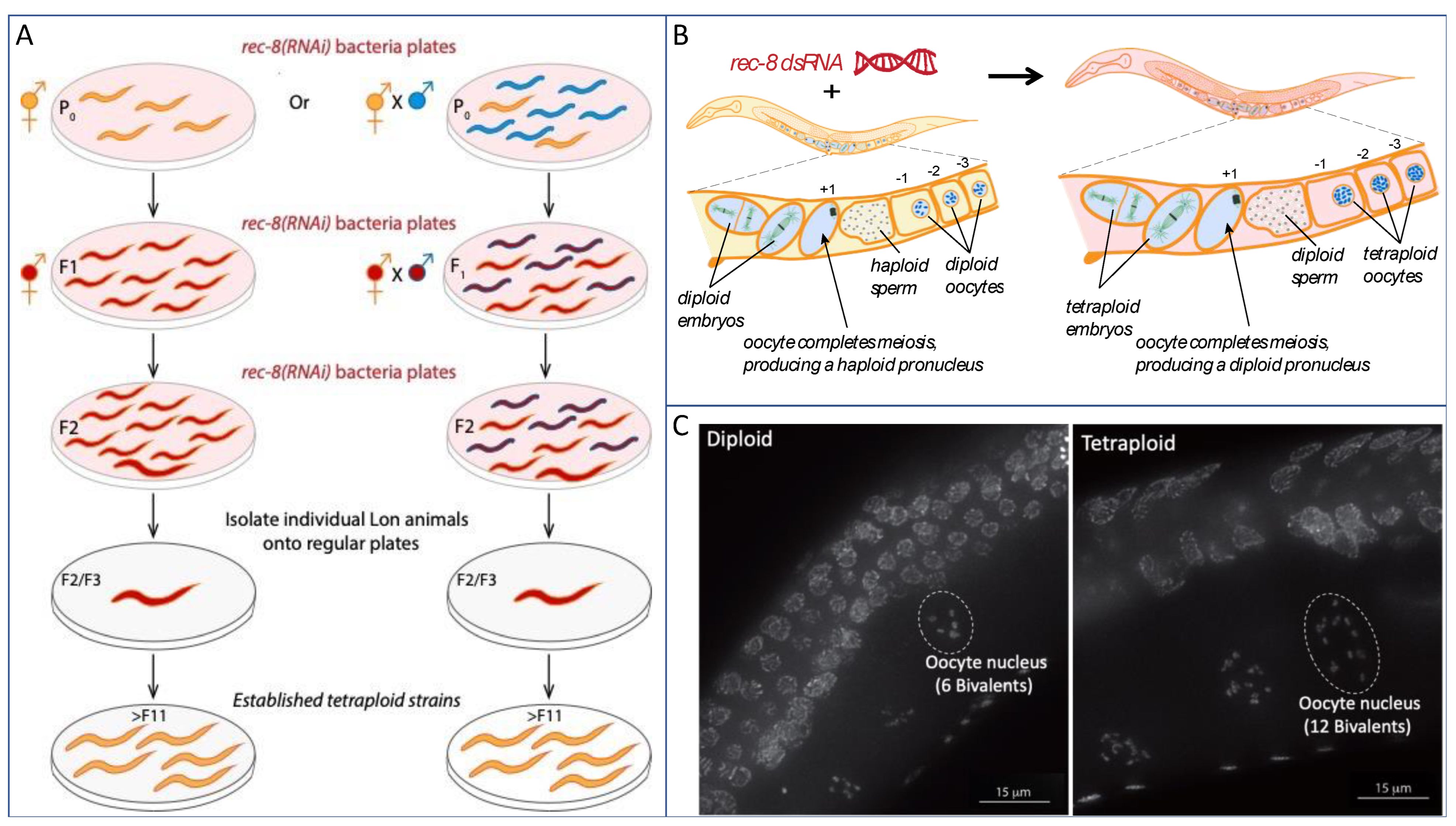

Table 1.
X/A ratio determines sex.
| Karyotype | X/A ratio | Sexual Fate |
|---|---|---|
| 2A;2X | 1 | female |
| 2A;1X | 0.5 | male |
| 3A;3X | 1 | female |
| 3A;2X | 0.67* | male |
| 4A;4X | 1 | female |
| 4A;3X | 0.75* | female |
| 4A;2X | 0.5 | male |
*X/A limits for female and male development. An X/A ratio ≥ 0.75 yields female fates and a X/A ratio ≤0.67 promotes male fates.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



