
Submitted:
26 April 2023
Posted:
27 April 2023
You are already at the latest version
Abstract
This study aimed to select the most effective metabolites for controlling honey bee nosemosis using culture extracts from 342 entomopathogenic fungi of 24 species from 18 genera. The germination inhibitory activity of the fungal culture extract against Nosema ceranae spores was evaluated using an in vitro germination assay method. Among 89 fungal culture extracts showing germination inhibitory activity of approximately 80% or more, 44 culture extracts that maintained their inhibitory activity even at a concentration of 1% were selected. Finally, the honey bee nosemosis inhibitory activity was evaluated using the cultured extracts of 5 fungal isolates having a Nosema inhibitory activity of approximately 60% or more even when the extract was removed after treatment. As a result, the proliferation of Nosema spores was reduced by all fungal culture extract treatments. However, only the treatment of the culture extracts from Paecilomyces marquandii 364 and Pochonia bulbillosa 60 showed a reduction in honey bee mortality due to nosemosis. In particular, the extracts of these two fungal isolates also increased the survival of honey bees.
Keywords:
honey bee
; nosemosis
; Nosema ceranae
; Apis mellifera
; entomopathogenic fungi
; metabolite
1. Introduction
The honey bee (Apis mellifera L. (Hymenoptera: Apidae)) plays an important role in the pollination of flowering plants, which are essential for the production of fruits, nuts and seeds upon which animals, including humans, rely for food [1,2,3,4]. In addition to their role in agriculture, honey bees are also important for maintaining biodiversity in natural ecosystems [5]. They help pollinate wildflowers, which provide habitat and food for a variety of other species. Honey bees are also vital to the production of honey, beeswax, and other bee products that humans have used for thousands of years [6,7]. However, unfortunately, honey bee colony collapse has become frequent, with global bee populations rapidly declining in recent years due to habitat loss, pesticide use, climate change and disease [8,9]. This is a major concern for the health of both food systems and natural ecosystems. Efforts are therefore underway worldwide to protect and conserve honey bee populations, including promoting habitat restoration, reducing pesticide use, and supporting research on bee health and behavior [9].
Bee colony collapse is a complex phenomenon that can be caused by multiple environmental and human-related factors [8,9]. Habitat loss and fragmentation, climate change, pesticides, mites and other parasites, and diseases and pathogens have been highlighted as major causes of honey bee colony collapse [10,11,12,13]. It is worth noting that bee colony collapse is often the result of a combination of these factors rather than a single cause. To address honey bee colony collapse, it is important to take a multipronged approach that addresses both environmental and human-related factors. Of these factors, honey bee disease, known as nosemosis, is known to have the greatest impact on honey bees [10,13,14,15]. Nosema infection can have severe negative effects on honey bee colonies, including reduced foraging activity, decreased colony growth and productivity, increased mortality, increased susceptibility to other stressors, reduced winter survival, decreased queen bee productivity, and reduced immune function [15,16,17,18,19,20]. Nosemosis is caused by two species of microsporidia, Nosema apis and N. ceranae. Nosema ceranae is more common and more lethal than N. apis [15,21,22,23]. Between the prevention and treatment for nosemosis, preventive methods are mainly relied on. Representative preventive methods include good beekeeping practices such as keeping the hive clean and providing adequate nutrition and moisture to the bees, hygienic behaviors such as removing diseased and dead brood from the hive, and avoiding stressors such as exposure to pesticides and poor nutrition [9,24,25,26]. In contrast, for direct treatment, an antibiotic called fumagillin is used, but its use is controversial due to its potential negative effects on the environment and other organisms [27,28,29,30]. Research efforts to replace it have reported that probiotic treatment, plant essential oils, propolis, plant extracts, and royal jelly, etc., can reduce the number of Nosema spores in bees and improve the survival rate of infected bees [31,32,33,34].
Entomopathogenic fungi, which are fungi that infect and kill insects, have been widely studied and used as materials for microbial insecticides for pest control [35,36,37,38]. Metabolites of these entomopathogenic fungi have various physiological activities, such as insecticidal activity, antibacterial activity, antioxidant activity, immunomodulatory activity, and cytotoxic activity, and potential applications are being sought in various fields, such as agriculture, medicine, and biotechnology [39,40,41,42].
In our study, entomopathogenic fungal metabolites with proliferation inhibitory activity against N. ceranae were screened from various entomopathogenic fungi for the prevention and treatment of nosemosis, and the effect of improving the survival rate of bees upon Nosema infection was evaluated. The purpose of this study was to provide basic data on the possibility of using metabolites of entomopathogenic fungi for the prevention or treatment of nosemosis.
2. Materials and Methods
2.1. Honey bees and N. ceranae
A colony of the honey bee A. mellifera was reared with 50% sucrose and bee bread as the main food, and adults that emerged within 24 h were used in the experiment. Nosema ceranae spores were provided by the Sericulture and Apiculture Division of the Rural Development Administration, Republic of Korea.
2.2. Entomopathogenic fungal culture extract
In this study, 342 isolates of entomopathogenic fungi from 18 different genera were used (Table S1) [43]. Fungal isolates were initially suspended in 1 mL of Sabouraud dextrose broth containing yeast extract medium (SDYB: 10 g of Bacto peptone, 40 g of dextrose, 10 g of yeast extract in 1000 mL of distilled water, and pH 6.0). Cultures were inoculated with agar blocks (6 mm) of fungi collected from 2-week-old potato dextrose agar (PDA) and grown in the dark at 25 °C with shaking at 150 rpm. After 10 days, the samples were centrifuged at 10,000 x g for 10 minutes. After removing the pellet, the supernatant was filtered using a LaboPass™ Mini Plasmid DNA Purification Kit column (Cosmo Genetech Co. Ltd., Korea) to remove spores and mycelia. Ethyl acetate fractionation was used to separate hydrophobic substances from fungal culture filtrates. After adding the same amount of ethyl acetate as the culture medium, vortexing for 20 min and centrifugation at 4,000 rpm for 5 min, the supernatant was collected. After that, ethyl acetate was evaporated using gaseous nitrogen, and the remaining extract was dissolved in 2% acetone in the same volume as the culture filtrate and used in the next experiment. The prepared culture filtrate extract was stored at -76 °C until use. To prepare a culture filtrate extract after quantitative inoculation of entomopathogenic fungi, fungal conidia that were harvested after being cultured in PDA medium for 2 weeks were used to prepare a conidial suspension using a 0.02% Tween-80 solution. The conidia were then counted using a hemocytometer. The conidial suspension was inoculated in 30 mL PDB medium in 50 µL at a concentration of 9 x 105 conidia/mL and cultured for 10 days. After culturing, spores and mycelia were removed from the culture medium, and culture filtrate extracts were prepared using the ethyl acetate fraction in the same methods as described above.
2.3. Purification of Nosema spores
To produce infective spores, honey bees were placed in plastic cages and inoculated with 1 x 106 spores of N. ceranae in sucrose solution (50% v/v in water). To obtain purified Nosema spores, after 10 days the midgut tissues from heavily infected honey bees were individually separated using forceps and washed with phosphate buffered saline [44]. The isolated midgut was ground in 200 μL of sterile distilled water in a Bullet Blender® Homogenizer (Scientific Instrument Services Inc., Ringoes, USA) set to speed 8 with 2 mm diameter tungsten carbide beads (Sigma‒Aldrich, St. Louis, USA) for 2 min. The homogenate increased the volume up to 1 mL, and the mixture was filtered through Qualitative No. 2 filter paper (Advantec MFS Inc., Dublin, USA) with an 8-11 μm pore size to remove tissue debris [45]. The filtered suspension was overlaid very gently on discontinuous 25%, 50%, 75% and 90% Percoll® (Sigma‒Aldrich, St. Louis, USA) gradient and centrifuged twice at 15,000 × g for 30 min at 20 °C [46]. A small but dense band just above the bottom of the tube was collected and resuspended in sterile distilled water. After a final centrifugation at 15,000 × g for 10 min at 20 °C, the spore pellet was resuspended in sterile distilled water and stored at room temperature until use. The spore concentration was measured by counting with a hemocytometer [47], and the spore suspension was freshly prepared before use.
2.4. In vitro germination assay
Aliquots of purified Nosema spores (10 μL; 1 x 103 spores) were spotted onto glass slide reaction cells (12 wells; Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany) and air-dried for 2 h at room temperature. Germination was triggered by adding 1.5 μL of 0.1 M sucrose in distilled water to the air-dried spores, a procedure that mimics natural conditions for the germination of environmental spores [48]. After maintaining the covered glass slide at room temperature for 6 h, the germinated spores were observed under a light microscope (magnification, 400 x) (Nikon Instech Co., Ltd., Tokyo, Japan). The germination rate was calculated as the percentage of total observed spores that had germinated.
2.5. Safety test of fungal culture extract
Twenty honey bees that emerged within 24 hours were transferred to one cage and allowed to adapt for 3 days before being used in the experiment. To evaluate the safety of the fungal culture extract, 50% sucrose mixed with culture extract was fed once to the honey bees, and for comparison, 50% sucrose containing the same concentration of acetone as the treated solvent was fed once. The survival rate of honey bees was observed and recorded every day for 14 days after treatment, and the experiment was repeated three times for each treatment group.
2.6. Nosema inoculation and culture extract feeding assay
To evaluate the effect of fungal culture extract on Nosema infection in honey bees, 25 honey bees that emerged within 24 h were used for each experimental group. Honey bees were orally infected with purified Nosema spores by making a 50% sucrose suspension at 1 × 10⁶ spores/mL and administering 1 mL to each experimental group. Treatment of fungal culture filtrate extract was carried out before and after Nosema inoculation. For the control group, honey bees were infected with Nosema and no treatment of culture extract was used. To evaluate the inhibitory effect of Nosema infection, the survival rate of honey bees was observed and recorded every day for 14 days. The proliferation of Nosema spores was examined by isolating and counting spores from the midgut of honey bees on the 14th day after infection to evaluate the effect of inhibition of Nosema growth. The experiment was repeated three times for each treatment group.
2.7. Statistical analysis
Spore germination and honey bee survival rate results were analyzed with SPSS statistical software v12.0 (SPSS, Inc., Chicago, IL, USA). Data were subjected to one-way analysis of variance (ANOVA), and comparisons among groups were performed with the SNK test. Data are expressed as the means ± standard errors (SEs), and statistical significance was set at the conventional α < 0.05 level.
3. Results
3.1. Inhibitory activity of entomopathogenic fungal culture extracts on Nosema germination
To evaluate the inhibitory activity of the entomopathogenic fungal culture extract on Nosema spore germination, the effect of acetone, a solvent used in preparing the culture extract, on Nosema spore germination was first evaluated. After treating Nosema spores with various concentrations of acetone from 0.25% to 20%, no significant effect of acetone on the germination of Nosema spores was observed at all concentrations (Figure S1). Therefore, it was confirmed that the 0.2% concentration of acetone used in this experiment had no effect on the germination of Nosema spores. All fungal isolates used in the experiment were mostly only expressed with numbers for convenience, and the identification information for the fungal isolates is shown in Table S1. As a result of evaluating the germination inhibitory activity of the culture extracts of 342 fungal isolates on Nosema spores, the germination of spores was inhibited in a variety of ways from 0% to a maximum of 96.5% (Table S2). Most of the spores did not germinate when the germination inhibitory activity was greater than approximately 80%, some spores germinated when the germination inhibitory activity was greater than 60%, and many spores germinated when the activity was less than 60% (Figure 1). Among the 342 fungal isolates, culture extracts from 89 isolates showed spore germination inhibitory activity of more than 80%, and 20 isolates showed activity of more than 90% (Table S2). By classification of entomopathogenic fungi, approximately 80% or more of the germination inhibitory activity against Nosema was observed in 15 species of 10 genera among 24 species from 18 genera (Table 1). Fifty of 126 isolates of Beauveria spp., 18 of 81 isolates of Metarhizium spp., 11 of 40 isolates of Cordyceps spp., 3 of 17 isolates of Pochonia spp. and 2 of 12 isolates of Paecilomyces spp. showed spore germination activity against Nosema. As various activities were confirmed in various fungi, culture extracts of 89 isolates showing spore germination inhibitory activity of 80% or more were prepared again by quantitative inoculation of fungi, and the germination inhibitory activity of Nosema spores was re-evaluated.
3.2. Nosema spore germination inhibitory activity according to the concentration of culture extract
For 89 selected entomopathogenic fungi, culture extracts were prepared by inoculation with the same conidia concentration, and then the extracts were diluted in distilled water at concentrations of 100, 10, and 1% to evaluate the germination inhibitory activity on Nosema spores (Figure 2). As a result, 44 of the 89 isolates showed a high spore germination inhibitory activity of 80% or more even at a diluted concentration of 1% as well as the extract stock solution. In particular, the decrease in activity according to the concentration of the extract of these isolates did not exceed 20% of the activity difference between the original solution and the diluted concentration of 1%. The activity of the remaining 45 isolates decreased significantly depending on the concentration of the extract, showing activity from approximately 50% to 5% at 1% extract concentration. Twenty-five isolates of Beauveria spp., 6 isolates of Met. anisopliae, 6 isolates of Cordyceps spp., 2 isolates of Paecilomyces spp., 2 isolates of Pochonia spp., and 3 other fungal isolates showed high activity similar to the stock solution even in the 100-fold diluted extract (Table 2). The following experiment was conducted using these 44 fungal isolates showing high spore germination inhibitory activity even at 100-fold dilution concentrations.
3.3. Mechanism of spore germination inhibitory activity of culture extracts
To investigate the active mechanism of inhibition of Nosema spore germination by 44 fungal culture extracts, the extracts of each fungal isolate were treated on Nosema spores for 2 h, and after removal of the extracts, spore germination was observed. As a result, the isolate showing a germination inhibition rate of approximately 60% or more even after removal of the culture extract were M. anisopliae 296, Pae. marquandii 364, Poc. bulbillosa 60, Bea. brongniartii 183, and Bea. bassiana 35, 161, and 59 (Figure 3). Among the remaining isolates, 5 isolates showed a germination inhibition rate of approximately 30% or more even after removing the extract, and the extracts of the other isolates showed no anti-germination activity or very low inhibition activity of approximately 5% or 20%. If the fungal culture extract showed high spore germination inhibitory activity even after removal, it was assumed that Nosema spores were inactivated by the culture extract, and the Nosema spore germination inhibitory activity of the culture extract was determined to be fungicidal activity. The inhibitory effect of Nosema infection on honey bees was continuously evaluated using the culture extracts of 6 fungal isolates, except for Bea. bassiana 161, which showed the lowest fungicidal activity.
3.4. Effects of culture extracts on honey bees
Prior to the evaluation of the inhibitory activity of the culture extract against Nosema infection in honey bees, the effect of the extracts on the survival of honey bees was evaluated. Acetone, used as a solvent for the culture extract, showed a similar survival rate for the untreated group up to 2% concentration, but at a concentration of 4%, the survival rate decreased from 4 days after treatment, and the final survival rate showed a difference of approximately 5% from the untreated group (Figure S2). In the Nosema infection experiment of the culture extract for honey bees, the extract that was diluted more than 10 times was used, so the actual concentration of acetone did not exceed 0.2%, confirming that the possibility of acetone affecting the lifespan of honey bees was very low. The effects of culture extracts on 6 fung6al isolates in honey bees were evaluated by dividing the concentrations of the extracts into 1% and 10%. As a result of treatment with 1% diluted extract, 5 fungal isolates except Bea. brongniartii 183 had no significant effect on the lifespan of honey bees (Figure 4A). Even at 10% dilution, only Bea. brongniartii 183 partially reduced the survival rate of honey bees, so it was excluded from further experiments (Figure 4B).
3.5. Inhibitory effect of culture extract on honey bee nosemosis
To evaluate the effect of inhibiting honey bee nosemosis by fungal culture extracts, honey bees were treated with each extract at a concentration of 5%, and the survival rate of honey bees and Nosema spore production were evaluated. The effect of the culture extract was evaluated in two ways: by treating the extract before Nosema infection of honey bees and by treating the extract after Nosema infection. After treating each culture extract before infection with Nosema, Pae. marquandii 364 culture extract only improved the survival rate of honey bees by approximately 13% compared to Nosema-infected honey bees (Figure 5A). Other fungal culture extracts showed similar or lower survival rates of honey bees compared to Nosema-infected honey bees. Treatment of culture extracts of M. anisopliae 296, Pae. marquandii 364, Bea. bassiana 59 and Poc. bulbillosa 60 reduced spore production by approximately 65%, 32%, 80% and 33%, respectively, compared to that of honey bees infected with Nosema alone (Figure 6A).
When honey bees were infected with Nosema and then treated with culture extracts, the extracts of Poc. bulbillosa 60 and Pae. marquandii 364 showed approximately 11% and 8% increases in honey bee survival rates, respectively, while the survival rates of the others decreased slightly (Figure 5B). Nosema spore production was reduced compared to Nosema-infected honey bees in all fungal culture extract treatments (Figure 6B). These results suggested that the culture extracts of Pae. marquandii 364 and Poc. bulbillosa 60 were effective in inhibiting honey bee nosemosis.
4. Discussion
This study was conducted to screen and select metabolites of entomopathogenic fungal cultures that are effective for the control of honey bee nosemosis. Among the 342 entomopathogenic fungal isolates of 24 species from 18 genera tested, the germination inhibitory effect of Nosema was shown in the culture extracts of more than 95% of the isolates (Tables 1 and S2). These results suggested that the germination inhibitors of Nosema were contained in the fungal culture extract and were consistent with previous studies showing that entomopathogenic fungal metabolites had various biological activities. It has been reported that entomopathogenic fungal metabolites have antimicrobial activity against various bacteria and fungi, so such antifungal activity could be sufficiently expected for fungi such as Nosema in our study [39,40,41,42]. To the best of our knowledge, this is the first report that various diverse types of fungal metabolites have antifungal activity against Nosema.
To determine the most effective culture extract among fungal isolates with antifungal activity against Nosema, 6 isolates showing high anti-germination effects at a 100-fold diluted concentration and even when removed after treatment with the extract were selected and used for the evaluation of honey bee nosemosis inhibitory activity (Figure 2 and Figure 3). In our study, the inhibitory activity of the culture extract against Nosema spores was evaluated by dividing the fungistatic activity, which is inhibited only when the extract is present, and the fungicidal activity, which inactivates the spores by the extract. It was judged that the fungicidal active culture extract was more effective in controlling nosemosis, and such isolates were selected. In addition, the nosemosis inhibitory activity of the culture extract showed a preventive effect by the treatment before infection with Nosema in one fungal isolate and a therapeutic effect after infection in two fungal isolates (Figure 5). However, in the experimental groups treated with the culture extract, lower honey bee survival rates were observed compared to the control group in which the culture extract was not treated, and these results were presumed to be due to the toxicity of the culture extract. Although the honey bee survival rate was not significantly reduced compared to that in the control under treatment with 10% of each culture extract, it is presumed that the low toxicity of the culture extract increased the susceptibility of honey bees to Nosema, thereby lowering the survival rate. However, it was confirmed that the production of Nosema spores was reduced by treatment with the culture extract, and it was confirmed that all the culture extracts could inhibit the growth of Nosema (Figure 6). As a particularly noteworthy result, treatment with the culture extracts of Poc. bulbillosa 60 and Pae. marquandii 364 showed a higher honey bee survival rate than uninfected honey bees, confirming that these culture extracts not only inhibit nosemosis but also increase the lifespan of honey bees. However, it has been shown that these effects may vary depending on the treatment concentration of these culture extracts. When only the culture extract was treated at a 10% concentration, the survival rate of honey bees was not higher than that of the control group, but at a 1% concentration, the culture extracts of Poc. bulbillosa 60 and Bea. bassiana 59 partially increased the survival rate of honey bees (Figure 4). To date, there has been no report on substances showing positive activity on the lifespan of bees among entomopathogenic fungal metabolites. In our results, further research is needed to determine whether the metabolite that shows Nosema inhibitory activity and the metabolite that increases the lifespan of honey bees are the same.
Various entomopathogenic fungal metabolites have been reported, and the most representative substance is beauvericin from Bea. bassiana and destruxins from Metarhizium spp., which are the most actively studied and utilized [40,49,50,51,52]. These substances have both antifungal activity and insecticidal activity. Other substances known to have antifungal activity include muscodorin, oosporein, patulin, enniatins, pradimicin, flavoglaucin, and terpenoids [40,51,53,54,55]. However, among these substances, inhibitory activity against Nosema has not been reported. Furthermore, since various fungal metabolites show various antifungal activities, the possibility that anti-Nosema active substances exist among fungal metabolites is considered sufficient. In a previous report on Poc. bulbillosa and Pae. marquandii, which showed the highest Nosema inhibitory activity in our study, there was no result for Nosema inhibitory activity. However, the antimicrobial activity, including antifungal activity and insecticidal activity, of the metabolites in Poc. chlamydosporia, Pae. variotii, and Pae. lilacinus have been reported [56,57,58]. Therefore, it is considered sufficient that the 2 fungal isolates in our study also have antifungal activity against Nosema. Through further research on the metabolites of Poc. bulbillosa 60 and Pae. marquandii 364, it may be possible to develop an effective control agent for honey bee nosemosis.
5. Conclusions
Metabolites of 342 entomopathogenic fungal isolates of 24 species from 18 genera were used to evaluate the germination inhibitory activity of Nosema to search for fungal metabolites with effective inhibitory activity against Nosema. As a result, inhibitory activity was observed in all cultured metabolites except for 2 genera and 1 species of fungus, and high inhibitory activity of approximately 80% or more was shown in 15 species and 10 genera. Among them, the culture extracts of Poc. bulbillosa 60 and Pae. marquandii 364 had fungicidal activity at low concentrations and effectively inhibited honey bee nosemosis.
Supplementary Materials
Table S1. The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Entomopathogenic fungal isolates used in the experiment. Table S2. Germination inhibition activity of entomopathogenic fungal culture extracts against Nosema ceranae spores. Figure S1. Germination of Nosema ceranae spores by acetone solvent treatment. After treating the spores with acetone at each concentration, the germination rate was determined by an in vitro germination assay. Figure S2. Honey bee survival by acetone treatment at different concentrations. A mixture of acetone and 50% sucrose solution was fed to honey bees. After that, the survival rate of honey bees was determined for 14 days. The control group was fed only a 50% sucrose solution. Data show the mean ± SE.
Author Contributions
Conceptualization: S.W., and D.K.; Design of experiment and methodology: S.W., and D.K.; Laboratory analysis: D.K., R.W., and K.K.; Data curation: D.K., R.W., and K.K.; Writing, original draft, review and editing: S.W., and D.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Agricultural Machinery/Equipment Localization Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (No.321054-05)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [CrossRef]
- Breeze, T.D.; Bailey, A.P.; Balcombe, K.G.; Potts, S.G. Pollination services in the UK: How important are honeybees? Agric. Ecosyst. Environ. 2011, 142, 137-143. [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321-326. [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [CrossRef]
- Papa, G.; Maier, R.; Durazzo, A.; Lucarini, M.; Karabagias, I.K.; Plutino, M.; Bianchetto, E.; Aromolo, R.; Pignatti, G.; Ambrogio, A.; Pellecchia, M.; Negri, I. The honey bee Apis mellifera: An insect at the interface between human and ecosystem health. Biology 2022, 11, 233. [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. Honey and health: A review of recent clinical research. Pharmacogn. Res. 2017, 9, 121-127.
- Luo, X.; Dong, Y.; Gu, C.; Zhang, X.; Ma, H. Processing technologies for bee products: An overview of recent developments and perspectives. Front. Nutr. 2021, 8, 727181. [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.P.; Vanengelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142-148. [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors associated with honey bee colony losses: A mini-review. Vet. Sci. 2020, 7, 166. [CrossRef]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; et al. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110-113. [CrossRef]
- Vanengelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [CrossRef]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can Varroa explain part of the colony losses? Apidologie 2010, 41, 353-363.
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; et al. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [CrossRef]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of Eukaryotes. J. Eukaryotic Microbiol. 2012, 59, 429-514. [CrossRef]
- Galajda, R.; Valenčáková, A.; Sučik, M.; Kandráčová, P. Nosema disease of European honey bees. J. Fungi 2021, 7, 714. [CrossRef]
- Fries, I. Nosema in European honey bees (Apis mellifera): An emerging threat to bee health. Insect Sci. 2010, 17, 418-424.
- Mayack, C.; Naug, D. Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 2010, 105, 20-27. [CrossRef]
- Goblirsch, M.J.; Spivak, M.; Kurtti, T.J. Nosema spp. infections in honey bees (Apis mellifera): Literature review and a survey of prevalence in North America. Insect Sci. 2013, 20, 1-22.
- Huang, W.F.; Solter, L.F.; Yau, P.M. Nosema ceranae causes asynchronous emergence of honey bee workers and reduces colony productivity. Apidologie 2018, 49, 11-22.
- Higes, M.; Martín-Hernández, R.; Meana, A. Nosema ceranae in European honey bees (Apis mellifera): An emerging infectious disease threatening apiaries and beekeeping. Insects 2020, 11, 886.
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chin, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Wide spread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1-10. [CrossRef]
- Chen, Y.P.; Evans, J.D.; Simith, I.B.; Pettis, J.S. Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 2008, 97, 186-188. [CrossRef]
- Emsen, B.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Eccles, L.; Lacey, B.; Ruiz-Perez, R.A.; Nasr, M. Higher prevalence and levels of Nosema ceranae than Nosema apis infections in Canadian honey bee colonies. Parasitol. Res. 2016, 115, 175-181. [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [CrossRef]
- Vanengelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J.; Pettis, J.S. Entombed pollen: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009. 101, 147-149. [CrossRef]
- Requier, F.; Garnery, L.; Kohl, P.L.; Njovu, H.K.; Pirk, C.W.W.; Crewe, R.M.; Steffan-Dewenter, I. The conservation of native honey bees is crucial. Trends Ecol. Evol. 2019, 34, 789-798. [CrossRef]
- Stanimirovic, Z.; Stevanovic, J.; Bajic, V.; Radovic, I. Evaluation of genotoxic effects of fumagillin (dicyclohexylamine) by citogenetic tests in vivo. Mutat. Res. 2006, 628, 1-10. [CrossRef]
- Pajuelo, A.G.; Torres, C.; Bermejo, F.J.O. Colony losses: A double blind trial on the influence of supplementary protein nutrition and preventative treatment with fumagillin against Nosema ceranae. J. Apic. Res. 2008, 47, 84-86. [CrossRef]
- Williams, G.R.; Shutler, D.; Little, C.M.; Burgher-MacLellan, K.L.; Rogers, R.E.L. The microsporidian Nosema ceranae, the antibiotic Fumagilin-B®, and western honey bee (Apis mellifera) colony strength. Apidologie 2011, 42, 15-22. [CrossRef]
- Williams, G.R.; Sampson, M.A.; Shutler, D.; Rogers, R.E. Does fumagillin control the recently detected invasive parasite Nosema ceranae in western honey bees (Apis mellifera)? J. Apic. Res. 2018, 57, 714-721. [CrossRef]
- Sabaté, D.C.; Cruz, M.S.; Benítez-Ahrendts, M.R.; Audisio, M.C. Beneficial effects of Bacillus subtilis subsp. subtilis Mori2, a honey-associated strain, on honeybee colony performance. Probiot. Antimicrob. Proteins 2012, 4, 39-46. [CrossRef]
- Glavinic, U.; Blagojevic, J.; Ristanic, M.; Stevanovic, J.; Lakic, N.; Mirilovic, M.; Stanimirovic, Z. Use of thymol in Nosema ceranae control and health improvement of infected honey bees. Insects 2022, 13, 574. [CrossRef]
- Klassen, S.S.; VanBlyderveen, W.; Eccles, L.; Kelly, P.G.; Borges, D.; Goodwin, P.H.; Petukhova, T.; Wang, Q.; Guzman-Novoa, E. Nosema ceranae infections in honey bees (Apis mellifera) treated with pre/probiotics and impacts on colonies in the field. Vet. Sci. 2021, 8, 107. [CrossRef]
- Kunat-Budzyńska, M.; Budzyński, M.; Schulz, M.; Strachecka, A.; Gancarz, M.; Rusinek, R.; Ptaszyńska, A.A. Natural substances, probiotics, and synthetic agents in the treatment and prevention of honeybee nosemosis. Pathogens 2022, 11, 1269. [CrossRef]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230-248. [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879-920. [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzon, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149-159. [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307-364. [CrossRef]
- Isaka, M.; Kittakoop, P.; Kirtikara, K.; Hywel-Jones, N.L.; Thebtaranonth, Y. Bioactive substances from insect pathogenic fungi. Acc. Chem. Res. 2005, 38, 813-823. [CrossRef]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367-2377. [CrossRef]
- Shin, T.Y.; Bae, S.M.; Kim, D.J.; Yun, H.G.; Woo, S.D. Evaluation of virulence, tolerance to environmental factors and antimicrobial activities of entomopathogenic fungi against two-spotted spider mite, Tetranychus urticae. Mycoscience 2017, 58, 204-212. [CrossRef]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181-1206. [CrossRef]
- Shin, T.Y.; Lee, W.W.; Ko, S.H.; Choi, J.B.; Bae, S.M.; Choi, J.Y.; Lee, K.S.; Je, Y.H.; Jin, B.R.; Woo, S.D. Distribution and characterisation of entomopathogenic fungi from Korean soils. Biocontrol Sci. Technol. 2013, 23, 288-304. [CrossRef]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032-3038. [CrossRef]
- Kim, D.J.; Yun, H.G.; Kim, I.H.; Gwak, W.S.; Woo, S.D. Efficient method for the rapid purification of Nosema ceranae spores. Mycobiology 2017, 45, 204-208. [CrossRef]
- Chen, Y.P.; Evans, J.D.; Murphy, C.; Gutell, R.; Zuker, M.; Gundensen-Rindal, D.; Pettis, J.S. Morphological, molecular, and phylogenetic characterization of Nosema ceranae, a microsporidian parasite isolated from the European honey bee, Apis mellifera. J. Eukaryotic Microbiol. 2009, 56, 142-147. [CrossRef]
- Shimanuki, H.; Knox, D.A. Diagnosis of honey bee diseases. U.S. Department of Agriculture, Agriculture Handbook No. AH-690. Washington, DC, USA, 2000; p.61.
- Olsen, P.E.; Rice, W.A.; Liu, T.P. In vitro germination of Nosema apis spores under conditions favorable for the generation and maintenance of sporoplasms. J. Invertebr. Pathol. 1986, 47, 65-73. [CrossRef]
- Klarić, M.S.; Pepeljnjak, S. Bovericin: Kemizam, bioloski aspekti i rasirenost [Beauvericin: Chemical and biological aspects and occurrence]. Arh. Hig. Rada Toksikol. 2005, 56, 343-350.
- Liu, B.L.; Tzeng, Y.M. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242-1254. [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Fron. Microbiol. 2021, 12, 705343. [CrossRef]
- Houard, J.; Aumelas, A.; Noël, T.; Pages, S.; Givaudan, A.; Fitton-Ouhabi, V.; Villain-Guillot, P.; Gualtieri, M. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J. Antibiot. 2013, 66, 617-620. [CrossRef]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for health and wellbeing: A potent bioactive metabolite of an entomopathogenic Cordyceps medicinal fungus and Its nutraceutical and therapeutic potential. Molecules 2020, 25, 2735. [CrossRef]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181-1206. [CrossRef]
- Lacatena, F.; Marra, R.; Mazzei, P.; Piccolo, A.; Digilio, M.C.; Giorgini, M.; Woo, S.L.; Cavallo, P.; Lorito, M.; Vinale, F. Chlamyphilone, a novel Pochonia chlamydosporia metabolite with insecticidal activity. Molecules 2019, 24, 750. [CrossRef]
- Qin, F.; Li, Y.; Lin, R.; Zhang, X.; Mao, Z.; Ling, J.; Yang, Y.; Zhuang, X.; Du, S.; Cheng, X.; Xie, B. Antibacterial radicicol analogues from Pochonia chlamydosporia and their biosynthetic gene cluster. J. Agric. Food Chem. 2019, 67, 7266-7273. [CrossRef]
- Dai, Z.B.; Wang, X.; Li, G.H. Secondary metabolites and their bioactivities produced by Paecilomyces. Molecules 2020, 25, 5077. [CrossRef]
Figure 1.
Spore germination inhibitory activity of fungal culture extract against Nosema ceranae. A, more than 80% inhibition; B, more than 60% inhibition; C, less than 60% inhibition. The scale bar is 20 μm. Arrows indicate mycelia that have grown after germination.
Figure 1.
Spore germination inhibitory activity of fungal culture extract against Nosema ceranae. A, more than 80% inhibition; B, more than 60% inhibition; C, less than 60% inhibition. The scale bar is 20 μm. Arrows indicate mycelia that have grown after germination.
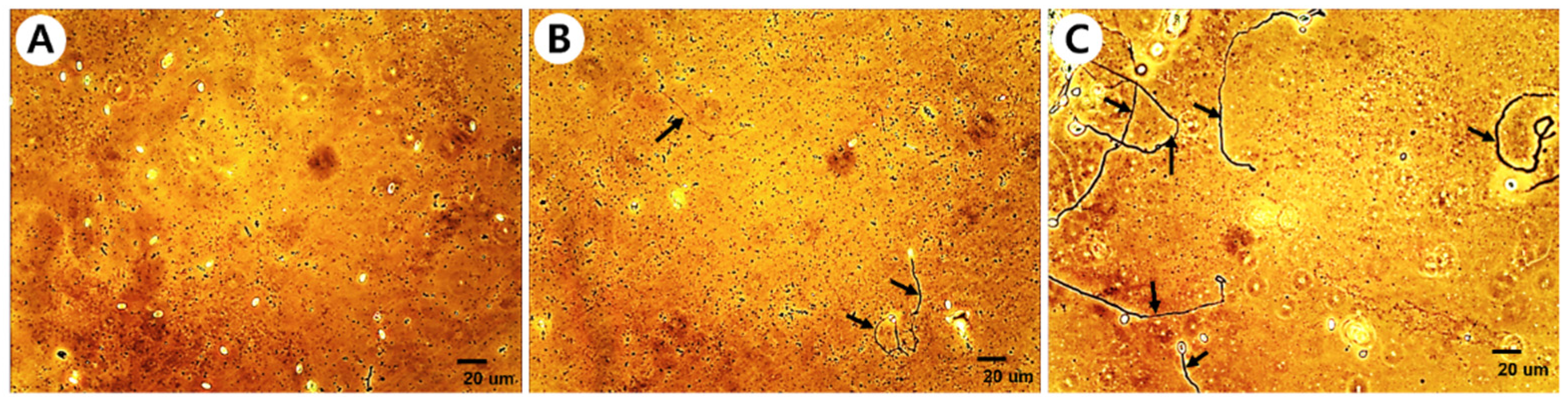
Figure 2.
Inhibition rate of Nosema ceranae spore germination in response to 89 entomopathogenic fungal culture extracts. Each extract was used to treat Nosema ceranae spores at concentrations of 100%, 10% and 1%, and then the germination rate was examined. Fungal isolates exhibiting an inhibition rate of 80% or more are indicated by black bars. Data show the mean ± SE.
Figure 2.
Inhibition rate of Nosema ceranae spore germination in response to 89 entomopathogenic fungal culture extracts. Each extract was used to treat Nosema ceranae spores at concentrations of 100%, 10% and 1%, and then the germination rate was examined. Fungal isolates exhibiting an inhibition rate of 80% or more are indicated by black bars. Data show the mean ± SE.
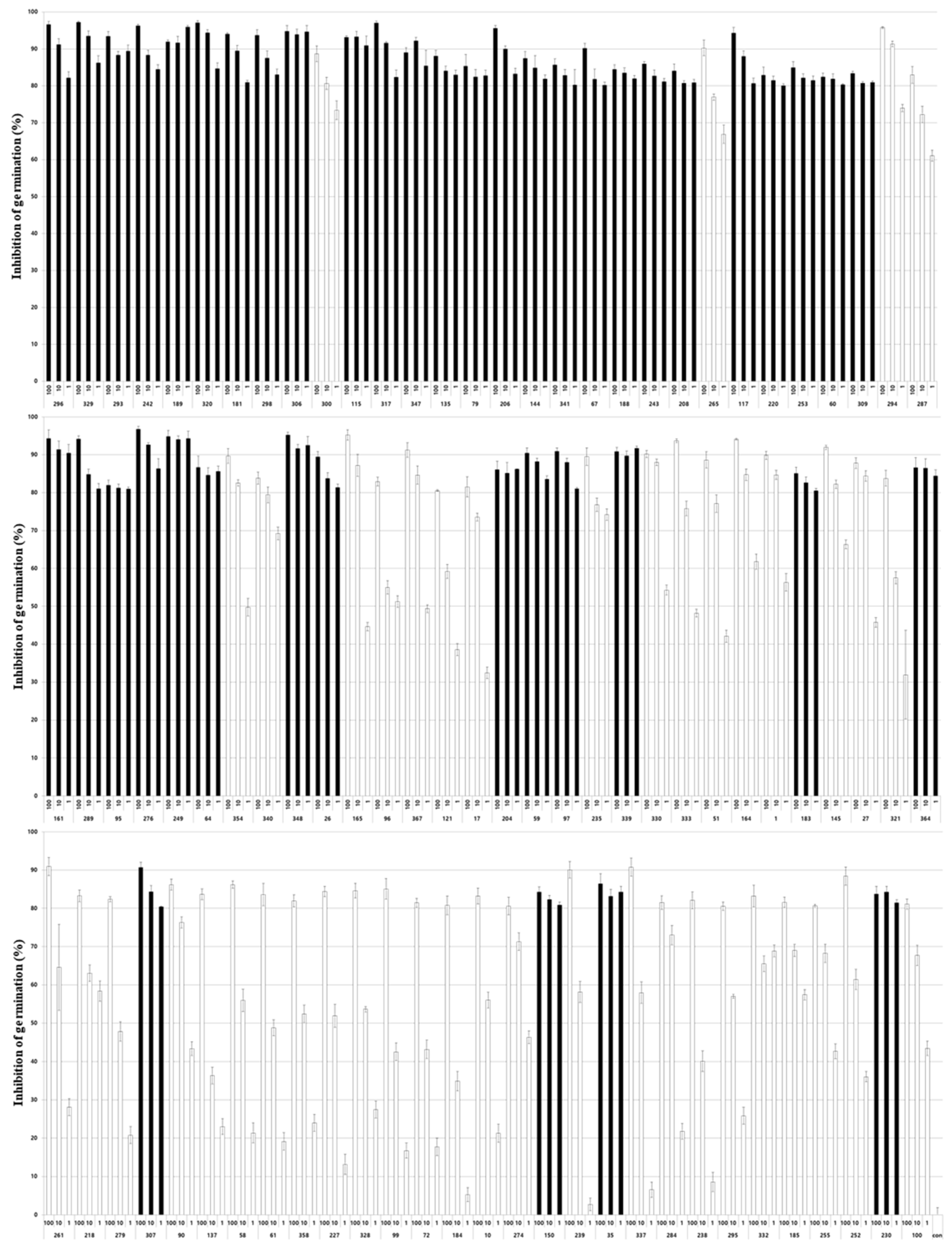
Figure 3.
Evaluation of the fungistatic and fungicidal activities of entomopathogenic fungal culture extracts against Nosema ceranae spores. After treatment with the fungal culture extract on Nosema ceranae spores, germination was observed without removing the extract (white bar) or after removing the extract (black bar). Data show the mean ± SE.
Figure 3.
Evaluation of the fungistatic and fungicidal activities of entomopathogenic fungal culture extracts against Nosema ceranae spores. After treatment with the fungal culture extract on Nosema ceranae spores, germination was observed without removing the extract (white bar) or after removing the extract (black bar). Data show the mean ± SE.

Figure 4.
Honey bee survival under 1% (A) and 10% (B) fungal culture extract treatments. A mixture of fungal culture extract and 50% sucrose solution was fed to honey bees. After that, the survival rate of honey bees was determined for 14 days. The control group was fed only a 50% sucrose solution. Data show the mean ± SE.
Figure 4.
Honey bee survival under 1% (A) and 10% (B) fungal culture extract treatments. A mixture of fungal culture extract and 50% sucrose solution was fed to honey bees. After that, the survival rate of honey bees was determined for 14 days. The control group was fed only a 50% sucrose solution. Data show the mean ± SE.
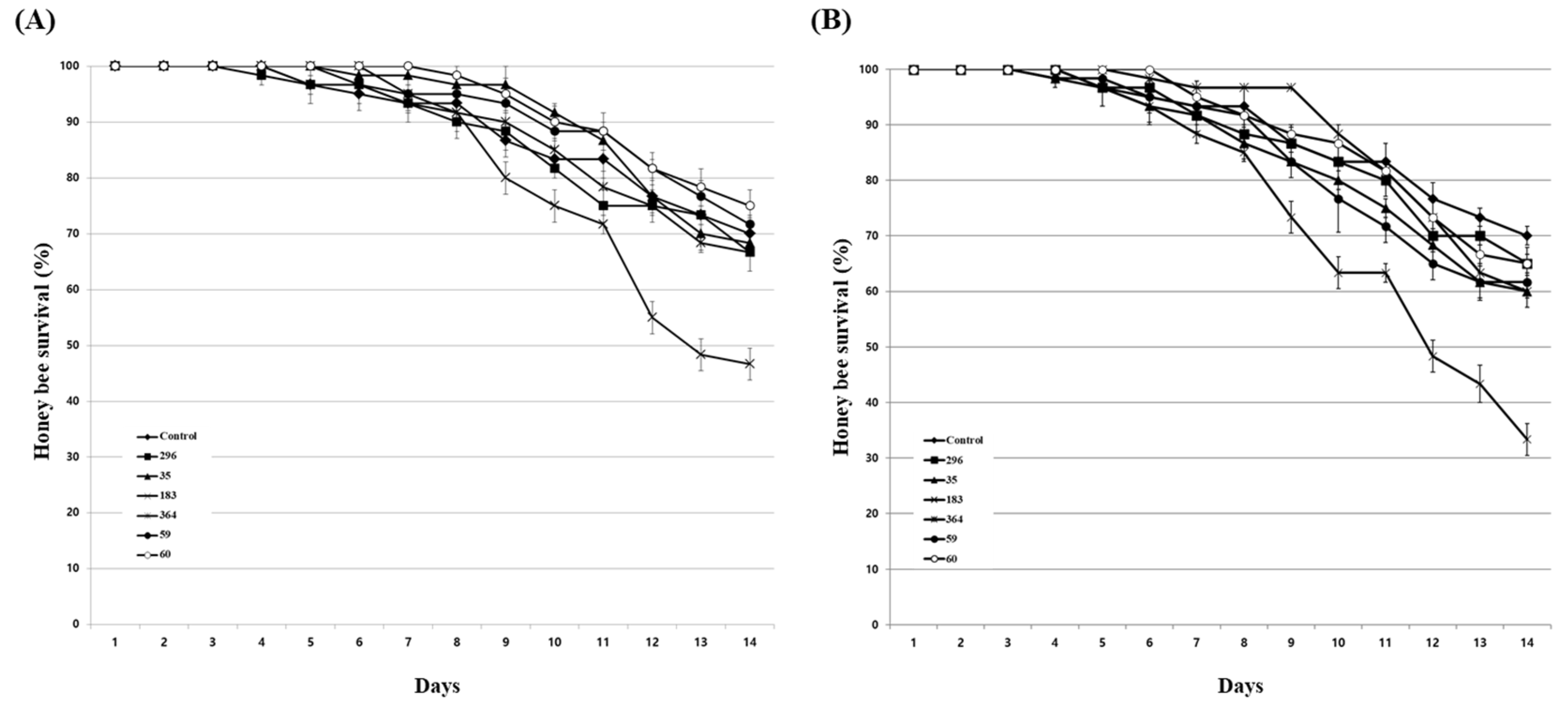
Figure 5.
Honey bee survival against Nosema ceranae infection after treatment with 5% fungal culture extract before (A) or after (B) infection. Nosema only is a control inoculated with a mixture of 50% sucrose solution and spores to honey bees. The control group was fed only a 50% sucrose solution. Data show the mean ± SE.
Figure 5.
Honey bee survival against Nosema ceranae infection after treatment with 5% fungal culture extract before (A) or after (B) infection. Nosema only is a control inoculated with a mixture of 50% sucrose solution and spores to honey bees. The control group was fed only a 50% sucrose solution. Data show the mean ± SE.
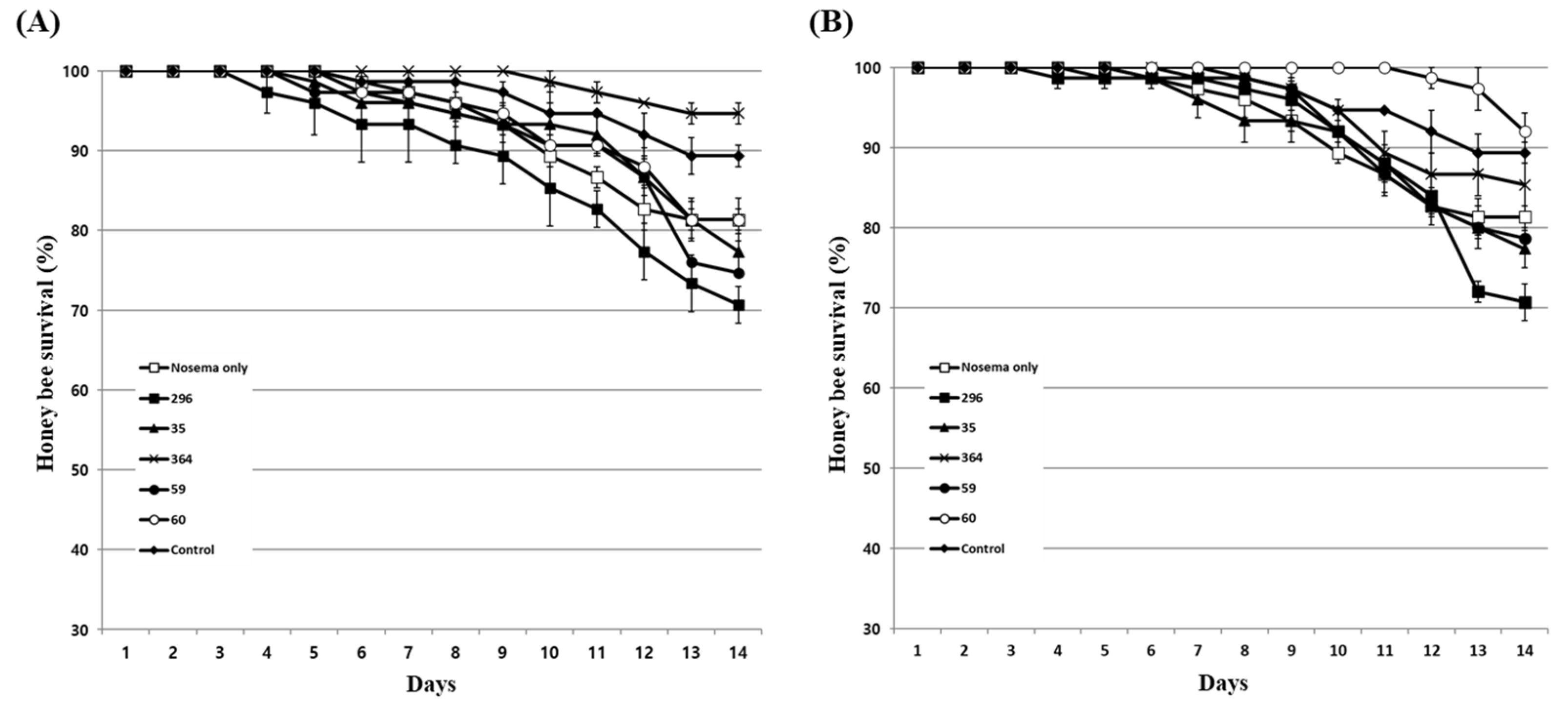
Figure 6.
Spore proliferation in Nosema-infected honey bees treated with 5% fungal culture extract before (A) or after (B) Nosema ceranae infection. Data show the mean ± SE. Values with different letters are significantly different (p < 0.05, SNK test in one-way ANOVA).
Figure 6.
Spore proliferation in Nosema-infected honey bees treated with 5% fungal culture extract before (A) or after (B) Nosema ceranae infection. Data show the mean ± SE. Values with different letters are significantly different (p < 0.05, SNK test in one-way ANOVA).
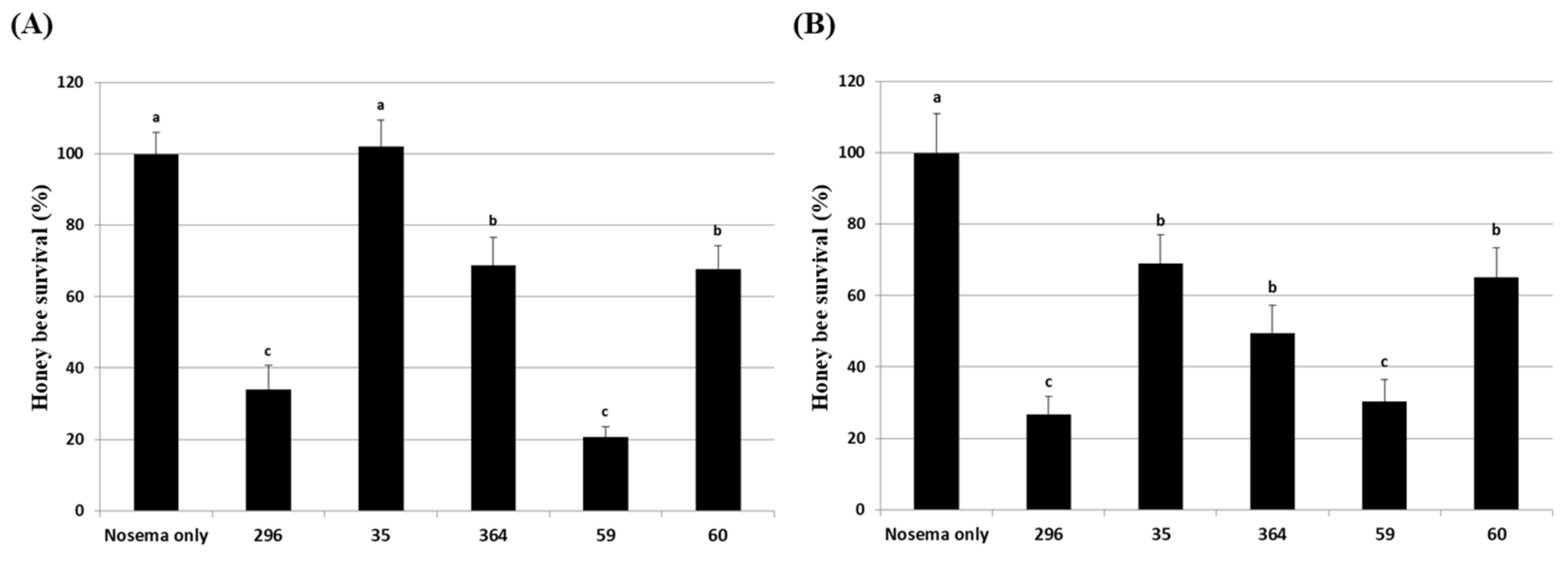

Table 1.
Entomopathogenic fungi showing more than 80% Nosema ceranae germination inhibition activity.
Table 1.
Entomopathogenic fungi showing more than 80% Nosema ceranae germination inhibition activity.
| Fungus | No. of tested isolates | No. of isolates showing the inhibition of spore germination |
| All fungal isolates | 342 | 89 (26%)* |
| Acremonium strictum | 1 | 0 (0%) |
| Aspergilluslentulus. | 5 | 0 (0%) |
| Aspergillus versicolor | 3 | 1 (33.3%) |
| Beauveria bassiana | 110 | 48 (43.6%) |
| Beauveria brongniartii | 8 | 1 (12.5%) |
| Beauveria pseudobassiana | 8 | 1 (12.5%) |
| Bionectria ochroleuca | 7 | 0 (0%) |
| Clonostachys rosea | 1 | 0 (0%) |
| Cordyceps farinosa | 12 | 5 (41.7%) |
| Cordyceps fumosorosea | 6 | 1 (16.7%) |
| Cordyceps javanica | 22 | 5 (22.7%) |
| Fusarium oxysporum | 2 | 0 (0%) |
| Lecanicillium spp. | 8 | 1 (12.5%) |
| Metarhizium anisopliae | 64 | 15 (23.4%) |
| Metarhizium lepidiotae | 1 | 0 (0%) |
| Metarhizium pemphigus | 16 | 3 (18.8%) |
| Mucoromycotina spp. | 1 | 0 (0%) |
| Myrothecium spp. | 5 | 0 (0%) |
| Paecilomyces lilacinus | 7 | 1 (14.3%) |
| Paecilomyces marquandii | 5 | 1 (20%) |
| Paraconiothyrium sporulosum | 2 | 1 (50%) |
| Phialocephala spp. | 1 | 0 (0%) |
| Pochonia bulbillosa | 16 | 3 (18.8%) |
| Pochonia rubescens | 1 | 0 (0%) |
| Simplicillium aogashimaense | 1 | 1 (100%) |
| Simplicillium sp. | 2 | 0 (0%) |
| Tolypocladium album | 23 | 1 (4.3%) |
| Tolypocladium cylindrosporum | 3 | 0 (0%) |
| Verticillium insectorum | 1 | 0 (0%) |
* Relative % to number of isolates (species or genus).
Table 2.
Entomopathogenic fungi showing the inhibition of Nosema ceranae spore germination by more than 80% in 1% concentration culture extract.
Table 2.
Entomopathogenic fungi showing the inhibition of Nosema ceranae spore germination by more than 80% in 1% concentration culture extract.
| Fungus | No. of isolate (%) |
| Aspergillus versicolor | 1 (2.3) * |
| Beauveria bassiana | 24 (54.5) |
| Beauveria brongniartii | 1 (2.3) |
| Cordyceps farinosa | 4 (9.1) |
| Cordyceps fumosorosea | 1 (2.3) |
| Cordyceps javanica | 1 (2.3) |
| Lecanicillium spp. | 1 (2.3) |
| Metarhizium anisopliae | 6 (13.6) |
| Paecilomyces lilacinus | 1 (2.3) |
| Paecilomyces marquandii | 1 (2.3) |
| Pochonia bulbillosa | 2 (4.5) |
| Tolypocladium album | 1 (2.3) |
* Relative % to number of isolates (species or genus).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



