
Submitted:
27 April 2023
Posted:
27 April 2023
You are already at the latest version
Abstract
Mucosal vaccines offer several advantages over injectable conventional vaccines, such as adaptive immunity induction, secretory IgA production at the entry site of most pathogens, and needleless vaccinations. Despite their potential, only a few mucosal vaccines are currently used. Developing new effective mucosal vaccines strongly relies on identifying innovative anti-gens, efficient adjuvants, and delivery systems. Several approaches based on phages, bacteria, or nanoparticles have been proposed to deliver antigens to mucosal surfaces. Bacterial spores have also been considered antigen vehicles, and various antigens have been successfully exposed on their surface. Due to their peculiar structure, the spores conjugate the advantages of live microorganisms with synthetic nanoparticles. When mucosally administered, spores expressing anti-gens have been shown to induce antigen-specific, protective immune responses. This review accounts for recent progress in the formulation of spore-based mucosal vaccines, describing the spore's structure, specifically the spore surface, and the diverse approaches developed to improve its efficiency as a vehicle for heterologous antigen presentation.
Keywords:
bacterial spores
; spore display system technology
; mucosal vaccine
; spore engineering
; Bacillus subtilis
1. Introduction
The Covid-19 pandemic, with 359 M infected people and claiming 5.6 M lives, has firmly confirmed the crucial role of vaccines on human health.
Since their discovery at the end of the 18th century with Jenner and then Pasteur's studies, vaccinations represent the only way to control infectious diseases and induce herd immunity. A long list of treatments containing live, attenuated, or killed viruses or bacteria has led to the suppression or decrease of the incidence of several fatal human diseases. So, each year, more than 5 million lives are saved by implementing yellow fever, hepatitis B, Haemophilus influenza type b, diphtheria, polio, and tetanus vaccines [1].
The current pandemic has highlighted that the technologies available today enable new vaccines to be rationally designed, considering the specific disease. Still, at the same time, it has pointed out some difficulties, such as the limited supply, the storage requirements, and the high cost. Therefore, public health organizations urge the scientific community to overcome these obstacles and develop a new generation of safe, efficient, and cost-effective vaccines. For instance, the World Health Organization has attempted to end cholera by 2030 by developing lower-cost alternative oral cholera vaccines [2].
One of the biggest challenges in vaccine design is developing an efficient product that provides systemic immunity and induces protective mucosal immune responses to block pathogens at their entry site. Inducing the synthesis of secretory IgA (sIgA) at the mucosal surface, an ideal vaccine should elicit immune protection at the mucosal surfaces and provide systemic immunity.
Unfortunately, mucosal immunity is not generated when an antigen is delivered systemically as in traditional vaccines, but this is not the case when an antigen is delivered orally or nasally. Besides enhancing local immunity, vaccine administration by oral or nasal route is a non-invasive approach, with improved patient compliance, eliminates the needle-associated risk of transmission of blood-borne diseases, and bypasses hepatic first-pass drug metabolism [3]. However, despite these potential advantages, the mucosal route for vaccine administration is still limited. Currently, only nine mucosal vaccines have been approved for use in humans, and they all are live attenuated or whole-cell inactivated vaccine formulations (Figure 1).
This low number is mainly because of the greater tolerability of orally administered whole-cell killed antigens, antigens' instability and susceptibility to proteases or endonucleases, and, crucially, the lack of an efficient mucosal delivery system. Therefore, identifying safe and effective mucosal adjuvants allied to innovative antigen delivery strategies is critical to advancing mucosal vaccines [7]. The rationale and challenges for developing mucosal vaccines are presented in Figure 2.
So far, several live microorganism display systems, including activated or live-attenuated viruses, and bacteria [8,9], have been considered for developing mucosal vaccine delivery systems. However, each of these platforms has its advantages and disadvantages.
Historically, the first viral vector vaccine was developed almost forty years ago, using as platform the vaccinia virus to express the hepatitis B surface antigen and provide protective immunity to chimpanzees against the viral disease [10,11].
Since then, several viruses, including poxviruses, adenoviruses, and herpesviruses, have been proposed as antigens platforms for vaccine design [12], and some of them have successfully been used in many veterinary species to protect, for example, against wildlife rabies [13], and new Castel disease [14]. However, only a single viral carrier has been authorized for human vaccination for the Ebola virus [15].
Recently, an innovative vaccine based on adenoviral vectors expressing the Covid-19 spike-1, nucleocapsid, and RNA-dependent RNA polymerase antigens has been proposed. The nasal administration of the new multivalent vaccine showed the ability to provide protective local and mucosal immunity against both the ancestral SARS-CoV-2 and two variants in a murine model [16].
The main advantages of the viral delivery system regarding traditional vaccines are the high stability and their ability to arise a solid adaptive immune response without an adjuvant. Moreover, the viral vectors ensure protection from gastrointestinal tract conditions when orally administrated, and effective release is addressed. Nevertheless, the main drawback remains their safety: inactivated viruses are considered safe but have weak immunogenicity. At the same time, the live attenuated viral carrier shows a greater efficacy but low safety since they could revert toward virulence [17]. Besides, oral viral vaccines have led to post-vaccination complications in some cases. For example, a rotavirus vaccine has increased the risk of intussusception in children, limiting its use.
As viruses, the live recombinant bacteria are applicable for oral vaccination since they can induce both systemic and mucosal immune responses [18], have intrinsic adjuvant properties [19], protect antigens during the gastrointestinal passage [20], and finally, are easy to handle. Moreover, bacteria show the additional advantage of not having restrictions on the number or dimension of heterologous genes to be inserted into their genome, in contrast to viruses which encounter limits in the capacity to encapsulate foreign DNA.
Both attenuated pathogenic and commensal bacteria have been engineered to carry and deliver heterologous antigens to the mucosal surfaces. The attenuated pathogenic strains are particularly effective since they are well adapted to the mucosal surface environments and Mycobacterium bovis BCG [21], Listeria monocytogenes [22], Salmonellae spp [23], and Shigellae spp. [24] are capable of eliciting immune responses against important viral, bacterial, protozoan, and metazoan pathogens in animal models. However, they are unsuitable for vaccinating individuals such as infants, the elderly, or immunocompromised patients for residual virulence [23].
In this regard, probiotic bacteria have been extensively studied as safe and effective vaccine candidates. Microorganisms such as Lactic Acid Bacteria (LAB) or Bacillus spp. are generally recognized as safe (GRAS) bacteria and represent major microorganisms used for probiotic purposes. Vaccines based on LAB, especially Lactobacillus lactis, Lactobacillus plantarum, and Lactobacillus casei, have shown promising beneficial effects in prophylactic or therapeutic probiotic-based vaccines against emerging respiratory viruses, including SARS, influenza viruses [25] and more recently SARS-CoV-2 [26,27].
2. Bacterial spores as mucosal vaccine delivery system
Also, bacterial spores have been proposed for oral delivery of the Covid-19 vaccine. In particular, Bacillus subtilis recombinant spores expressing SARS-CoV-2 spike protein receptor-binding domain have been developed and shown to increase specific neutralizing antibodies when orally administrated to mice or healthy human volunteers [28]. The utilization of B. subtilis spores to display heterologous antigens has been extensively reported since its first successful application in 2001 by Isticato et al. and will be the focus of this review [29].
2.1. Bacterial spore, sporulation and intestinal life cycle
Bacterial endospores are metabolically inactive cells produced by Gram-positive bacteria belonging to the genera of aerobic or facultative anaerobic Bacilli and the strictly anaerobic Clostridia [30]. These bacteria can adapt to environmental changes through different survival mechanisms, such as competence, biofilm production, cannibalism, or sporulation [31]. This latter process is induced when the environmental conditions do not allow cell growth or survival and bring to the production of endospores (hereafter referred to as a “spore”) and have been widely investigated in Bacillus subtilis, the model system for spores [32,33,34,35]. During sporulation, the vegetative cell undergoes morphological, biochemical, and physiological changes, leading to the development of two different but genetically identical cells: the mother cell and the forespore (Figure 3). This differentiation event is possible thanks to distinct gene expression programs in the two cell types [32,33,34,35].
The mature spore released in the environment is one of the most resilient cell types in nature, able to survive extreme stresses such as extreme pH and temperatures, dehydration, ionizing radiations, mechanical abrasion, toxic chemicals, and hydrolytic enzymes [36]. Moreover, the spores can persist in this dormant state for several decades, perhaps even longer, until the environmental condition returns favorable [36].
Despite its dormancy, the spore can quickly germinate and resume vegetative growth in response to water, nutrients, and favorable environmental conditions (Figure 3).
The exceptional longevity of the spore in the environment is the main reason for the ubiquitous distribution of these organisms in nature, found in soil, air, fermented foods, and the human gut [37,39].
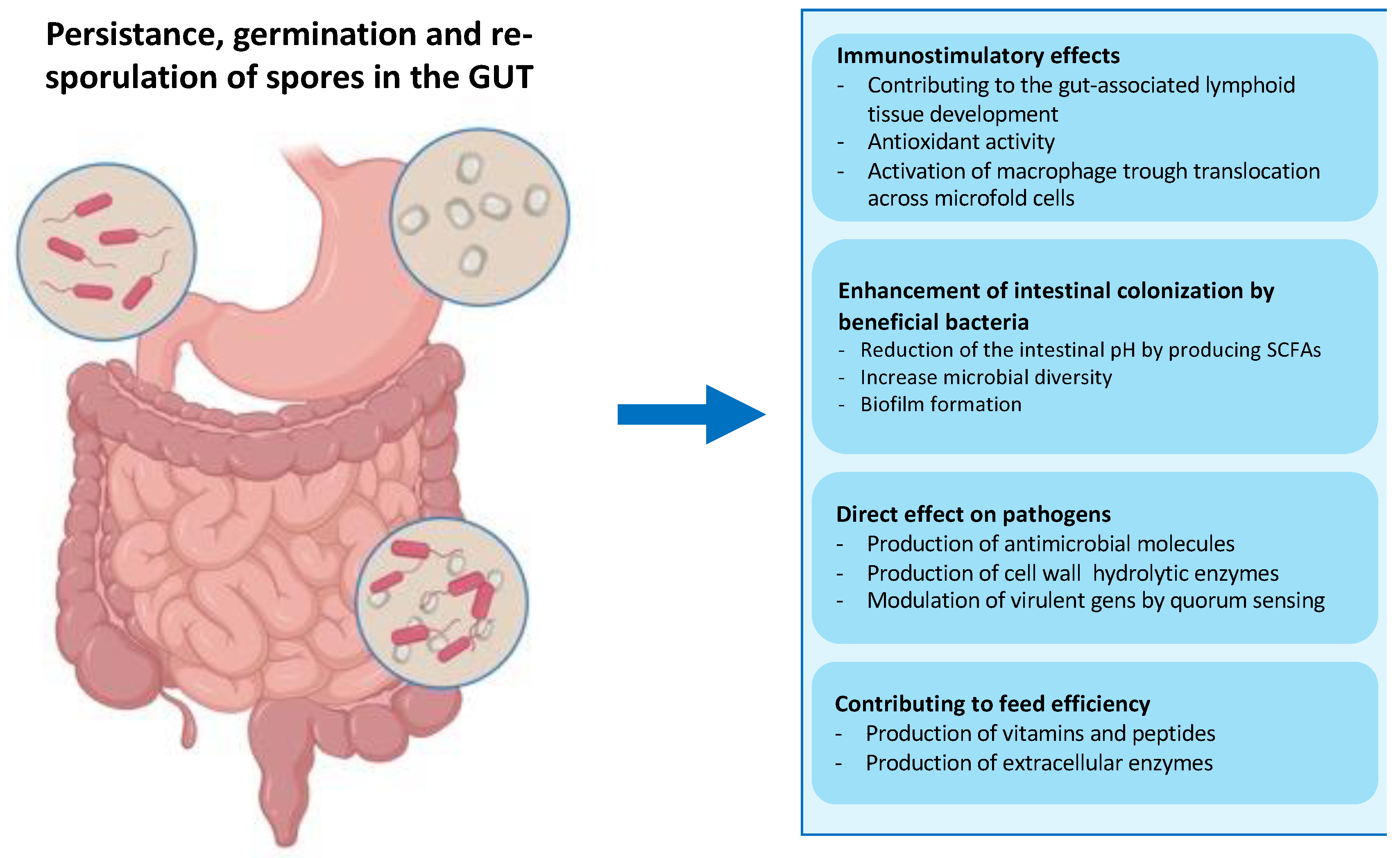
In particular, ingested spores of B. subtilis have been shown to pass safely through the stomach, germinate and proliferate in the upper intestine, triggered by the rich nutrient environment of the small intestine [38,39] (Figure 5). Then, germination-derived cells can sporulate in the lower part of the intestine, thus performing an entire life cycle in the animal GUT [39]. Several works have shown that many Bacillus species, including Bacillus clausii, Bacillus pumilus, Bacillus polyfermenticus, and B. subtilis contribute to the composition of GUT microbiota and so maintain host health by performing nutritional, immunological, and physiological functions [37,40,41,42].
Figure 4.
Fate of ingested bacterial spores in the gut. Spores administered by the intragastric route can transit through the stomach thanks to their extreme resistance. The low pH of the stomach activates the spores to germinate before they reach the small intestine, which is rich in nutrients. In the GUT, the vegetative cells help modulate the immune response and secrete hydrolytic enzymes, antioxidants, vitamins, peptides, and antimicrobial compounds, contributing to the GUT microbiota balance and facilitating digestion [39]. The particularly toxic environment of the GUT, including anoxia, low pH, bile salts, and an extremely high concentration of commensal bacteria, can induce the Bacillus cells to re-sporulate [37].
Figure 4.
Fate of ingested bacterial spores in the gut. Spores administered by the intragastric route can transit through the stomach thanks to their extreme resistance. The low pH of the stomach activates the spores to germinate before they reach the small intestine, which is rich in nutrients. In the GUT, the vegetative cells help modulate the immune response and secrete hydrolytic enzymes, antioxidants, vitamins, peptides, and antimicrobial compounds, contributing to the GUT microbiota balance and facilitating digestion [39]. The particularly toxic environment of the GUT, including anoxia, low pH, bile salts, and an extremely high concentration of commensal bacteria, can induce the Bacillus cells to re-sporulate [37].
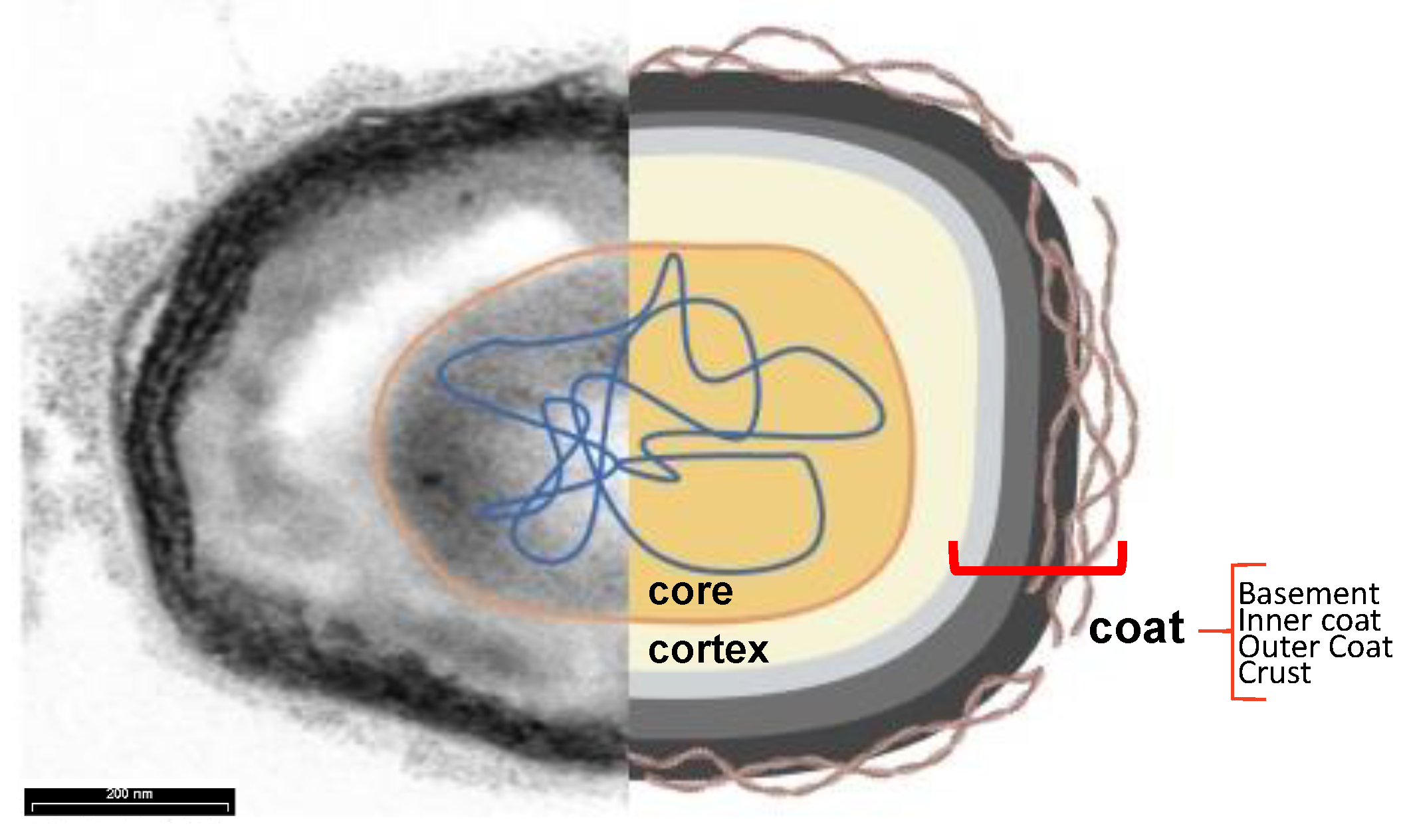
At an Electron microscope, the spore of B. subtilis appears to be formed by several layers surrounding an innermost part, the core. The core contains a partially dehydrated cytoplasm, a copy of the chromosome, RNAs, and inactive [32,35]. The highly dehydrated state of the core is due to the replacement of water with Ca2+-dipicolinic acid and is responsible for the heat resistance of the spores. The tolerance to damage from UA radiation is due to small acid-soluble proteins (SAPS) that maintain the DNA in a compact state. The core is protected by the cortex, a thick peptidoglycan-like structure, and the coat, a multilayered proteinaceous layer [43]. In turn, the coat is formed by four concentric sublayers: the basement layer, inner coat, outer coat, and crust, made of glycoproteins [44].
Figure 5.
Subcellular Spore Structure. Transmission electron microscopy analysis (left) and cartoon (right) of a typical B. subtilis spore. The core (orange)contains the dehydrated cytoplasm and the nucleic acids (blue) and is protected by the cortex (yellow) and the four sublayers of the coat: the basement layer (light grey), the inner coat (grey), the outer coat (black) and the crust (brown filamentous), visible only after ruthenium red staining.
Figure 5.
Subcellular Spore Structure. Transmission electron microscopy analysis (left) and cartoon (right) of a typical B. subtilis spore. The core (orange)contains the dehydrated cytoplasm and the nucleic acids (blue) and is protected by the cortex (yellow) and the four sublayers of the coat: the basement layer (light grey), the inner coat (grey), the outer coat (black) and the crust (brown filamentous), visible only after ruthenium red staining.
2.2. Spore surface
In B. subtilis, the coat represents the spore surface and comprises over 80 proteins, representing ∼25% of total spore proteins. All the genes coding for the Cot proteins are expressed in the mother cell under the control of two sporulation-specific sigma factors of the RNA polymerase, SigE and SigK, and at least three transcription regulators, SpoIIID, GerE, and GerR [45]. This complex transcriptional regulation ensures the synthesis of each spore surface component at the correct time during sporulation [32,33,34,35]. Post-translational modifications, such as phosphorylation, cross-linking, or glycosylation, occur after their synthesis by enzymes specific to sporulation. While the cross-linking, including disulfide cross-links and dityrosine cross-links, contributes to the strength of the coat, glycosylation of coat proteins determines the hydrophobicity of the spore and, therefore, its adherence properties [46,47].
Finally, the assembly of the coat components involves various coat proteins with morphogenetic activity, i.e. proteins that do not affect the synthesis of other coat proteins but are essential to organize the coat into the multi-layered final structure [32,48,49,50]. In particular, some coat proteins are glycosylated in the late-stage spore development and assembled on the surface of the pre-spores forming the crust layer [37,44,51].
Several studies have highlighted the ability of some coat components to self-assemble and form remarkably robust structures characterized by hexagonally symmetric semipermeable arrays [52]. The final result of the coat development is a complex layer arranged into two major layers: an outer, darker layer with coarse striations and a lighter inner layer with a lamellar appearance [53]. The two other sublayers, the basement layer, and the crust, respectively under the inner coat and external to the outer coat, complete the proteinaceous spore surface. The physical-chemical properties of proteins and carbohydrates composing the coat of the mature spore give a negative charge and a relative hydrophilicity to the spore surface [54], as suggested by the altered electric charge and hydrophilicity of mutant spores lacking some coat components [44,51,55].
Spores of Bacillus megaterium or in Bacillus cereus and its close relatives Bacillus anthracis and Bacillus thuringensis are surrounded by a further layer, the exosporium [56,57]. A basal layer of glycoproteins forms this structure with hair-like protrusions [32]. Among Bacillus species, the spore's surface differs not only by the presence or absence of the exosporium but also by the diversity of proteins that component the coat. Genome sequence analysis of the Bacillus species has highlighted that less than one-half of the known B. subtilis coat protein genes have recognizable orthologues in its close relatives. In the genomes of Clostridium spp., the B. subtilis coat genes are even less conserved [56]. Despite the spore surface's physical-chemical properties and structure varying from specie to species, the coat of the bacterial spores, by itself or in combination with the exosporium, plays a crucial role in spore protection and the initiation of germination [53,57].
2.3. Bacterial spore as a recombinant vaccine platform
The rigidity and compactness of the coat, along with spore stability and resistance, have suggested using spores as a display system for formulating mucosal vaccines. A genetic system to engineer the surface of the B. subtilis spore was developed in a pioneering study in 2001, and model antigens efficiently displayed on the spore surface using the coat components as anchoring motifs [29]. Since then, various antigens have been successfully exposed on the surface of recombinant spores of B. subtilis and other spore formers and shown to induce antigen-specific, protective immune responses in mucosally immunized mice [58,59,60,61].
With respect to systems based on microbial cells or phages described above, the spore-based approach provides at least three main advantages:
- (1)
- the extreme stability due to the well-documented resistance of the bacterial spore to high temperatures, acidic pH, and the presence of chemicals and enzymes [29,30,31,32]. Guaranteeing the high stability of the vaccine carrier system from production to administration to the patient is a crucial requirement of vaccine development. Bacillus spores are more stable than vegetative cells during the processing and storage of commercial preparations, making them a suitable candidate for vaccine formulations [62]. Moreover, stability at extreme temperatures is strongly required in the development of mucosal vaccines, mainly for those intended for use in developing countries, where poor distribution and storage conditions are the main limitations [3,7,60,61,62];
- (2)
- the exceptional safety record of several spore-former species used worldwide in probiotic preparations for human and animal use as dietary supplements and growth promoters [41,42,63]. As mentioned above, several spore former species are part of the animal gut microbiota, which have a role in the development of the immune system, protection against intestinal pathogens, induction of cytoprotective responses, and of anti-oxidative stress responses in epithelial cells (Figure 4) [38,39,40,41,42]. The safety record is an essential requirement if the display system is intended for the delivery of antigens molecules to human mucosal surfaces;
- (3)
- All known coat proteins are synthesized in the mother compartment during sporulation [31,32,43]. Consequently, coat components and the antigens fused to them do not need to undergo a cell wall translocation step to expose the recombinant proteins externally, thus overcoming the size limitation often encountered with cell-based display systems [59,64];
- (4)
2.4. Spore-based vaccine design strategy
In recombinant vaccines, the immunogenicity of an antigen depends on several factors, such as its surface exposition and the fold stability of the protein chimer.
The conformational stability of the antigen, for example, affects its intracellular fate during processing in antigen-presenting cell APC (from the uptake to the final presentation of antigenic peptides) and the downstream events, such as the stimulation of B and T cells or the differentiation of T cells into effector cells [65].
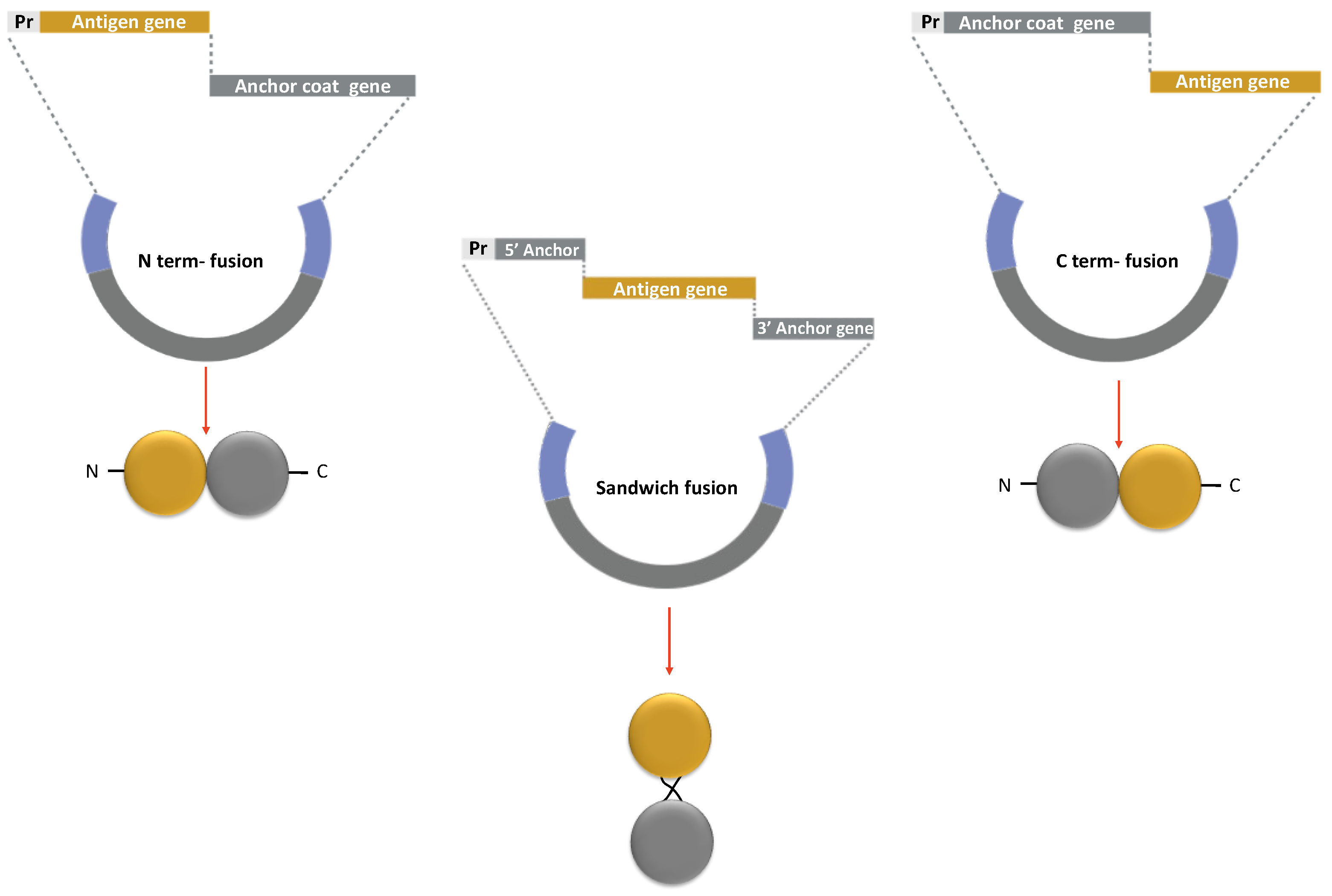
The strategy to display antigens on the spore surface and the choice of anchor protein are crucial for the success of the recombinant vaccine. Efficient coat-carrier proteins have to meet specific criteria, such as having a solid anchoring motif to ensure the attachment of the fusion protein on the spore surface and a domain to correctly displayed the epitopes. Moreover, the coat component should ensure the conformational stability of the recombinant protein to natural stresses, i.e., pH, temperature, or redox environment. Maintaining the antigen folding is crucial to avoid the peptide backbone being accessible to the proteases, which are abundant on mucosal surfaces and in the extracellular matrix [66]. Finally, the fusion of the antigen to the spore component should not affect any functions or proprieties of the first one [59,60,61]. Therefore, fusions at the N-terminus, C-terminus, or interior of the Cot protein (sandwich fusions) are often constructed with the antigen to obtain an efficient display (Figure 6). The N-terminus fusion is preferred when the Cot protein has the anchoring domain in its C-terminus part or when the epitope is present at the N- terminal of the antigen. On the contrary, the C-terminus fusion could opt if the anchoring domain is in the N-terminal of the coat protein [59].
2.4.1. Anchor Proteins
Most antigens exposed on the spore surface have been anchored to the crust-associated components CotB and CotG [29,67,68,69] or the outer coat protein CotC [49,70]. The surface localization of the three spore coat proteins and their abundance have ensured the exposure of many heterologous proteins (Table 1).
In particular, CotB was used in the first attempt at spore surface display technology, fusing the non-toxic C-terminal fragment of the tetanus toxin (TTFC), a highly immunogenic and well-characterized peptide encoded by the tetC gene of Clostridium tetani [29]. The correct timing of expression of the CotB-TTFC during sporulation was guaranteed by placing the chimera protein under the cotB promoter, while the chromosomal integration of the cotB-tetC fusion ensured the genetic stability of the construct (Figure 6)[29].
Since the anchor motif of CotB was not yet identified, the tetC gene was cloned at the 3' and 5' of the cotB gene and inserted in an internal region, obtaining a sandwich fusion. Western, dot blot and fluorescent-activated cell sorting analyses showed that similar amounts of TTFC were expressed on the surface of the three recombinant spores, suggesting that the fusion position did not affect the efficiency of the display system. Moreover, it was estimated that a single spore exposed more than 1.5 x 103 TTFC molecules on its surface [29]. When orally or nasally administrated to mice, the recombinant spores induced the production of TTFC -specific Faecal sIgA and serum IgG and protected mice in a challenging experiment with purified tetanus toxin [29,71].
After these first studies, other antigens have been successfully displayed on B. subtilis spores using CotB as anchor protein, such as the domains 1b-3 and 4 of the Protective Antigen (PA) of B. anthracis [72,73], the C-terminal part of the alpha toxin of Clostridium perfringens [74], the UreA protein of Helicobacter acinonychis [67], the carboxy-terminal repeat domains of toxins A and B (TcdA-TcdB) of Clostridium difficile [75], and the VP28 protein of the white spot syndrome virus [76] (Table 1).
The strategy initially developed to use CotB as an antigen-displayed protein has been followed for other coat components. For example, TTFC and the subunit B of Escherichia coli labile toxin (LTB) [70] and TcdA-TcdB of C. difficile [75] have been exposed on spores using the 12 kDa CotC as anchor motif (Table 1). In all cases, the coat protein coding gene and its promoter have been cloned inframe with the antigen coding sequence to construct a translational fusion followed by the chromosomal integration of the gene fusions [77]. Contrary to what was observed for CotB, the antigen display efficiency of the CotC-based chimera depends on the position of insertion of the heterologous part, and a 5-fold increase in the efficiency of the display was observed when TTFC was at the N-terminal end of CotC rather than at its C-terminal end [70,77]. In recent years, mucosal spore-based vaccines against Clonorchis sinensis were proposed using CotC as anchoring protein since the increase of food-borne infectious diseases caused by the ingestion of contaminated raw or undercooked fish. Several liver fluke antigens, such as CsTP22.3, TP20.8, CsPmy, and CsLAP2, were successfully displayed on spore surfaces [64,79,80,81,82,83]. When the recombinant spores were intragastrically administrated to mice, an immunological response was observed with specific IgGs in sera and sIgA increase [64,79,81].
CotG also was successfully used as a carrier to display proteins such as UreA of H. acinonychis [67] and the fragment of C. difficile flagellar cap FliD protein [84]. Interestingly, when the biotin-binding streptavidin was fused to CotG, the recombinant proteins could still form the native tetrameric form on the spore surface [85,86]. Several antigens, i.e. viral antigens, are multimer and can only induce an immune response in this complex form. Therefore, the spore's ability to display the antigen in a multimeric form that mimics epitopes' natural presentation could ensure protective responses.
In recent work, the transmissible gastroenteritis virus spike (TGEV-S) protein was anchored on the spores by CotG [87]. When vaccinated with the recombinant spores, suckling piglets showed an elevated specific SIgA titer in feces and IgG titers and neutralizing antibodies in serum. Moreover, the animals were protected in TGEV challenge experiments. Altogether these results highlight the considerable potentiality of CotG as carrier proteins for the formulation of the mucosal vaccine [87].
Other two coat proteins have been used as anchor motifs to expose antigens, CotZ and CgeA, both crust components [44,52]. CotZ was proposed as an anchor motif for the first time by Negri et al. to expose the fragment of C. difficile FliD protein [84]. Despite the CotZ localization in the crust, the outermost layer of the spore, the surface display efficiency of recombinant FliD protein was similar to that measured when the heterologous protein was fused to CotB, CotG, and CotC [84]. These results suggested that the exposition of the chimera protein on the spore surface does not necessarily depend on the localization of the coat protein used as carrier protein but also the passenger protein and the properties of the obtained fusion.
Finally, CgeA was used to express on the spore surface the H. pylori NCTC 11637 cytotoxic-associated gene A protein (CagA) [88].
| Anchor [Refs] | Target pathogen | Antigen | Fusion method | linker | Application |
| CotB | |||||
| [29] | Clostridium tetani | C-term fragment of the tetanus toxin, TTFC | C-term, N-term, sandwich | - | Oral vaccination for tetanus |
| [84] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEA; AAKGGG | C. difficile oral vaccine |
| [75] | C-term repeat domains of toxins A and B, TcdA-TcdB | C-terminal | - | ||
| [74] | Clostridium perfringens | C-term of alpha toxin gene, Cpa247-370 fused to the GST gene | C-terminal | - | Oral and Nasal Vaccine against necrotic enteritis |
| [72,73] | Bacillus anthracis | anthrax protective antigen ,PA | C-terminal | - | Anthrax vaccine |
| [67] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | GGGEAA; AKGGG | Anti-Helicobacter vaccine |
| [88] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N- terminal | GGGGS | Anti-Helicobacter vaccine |
| [90] | Mycobacterium tuberculosis | immunodominant secretory antigen, MPT64 | C-terminal | - | Nasal Vaccine against tuberculosis |
| [91] | Streptococcus mutans | truncated P1 protein | N-terminal | NR | S. mutans vaccine |
| [76] | White spot syndrome virus | major envelope proteins, VP28 | C-terminal | - | Oral vaccine for shrimps |
| [92] | Influenza virus | ectodomain of influenza virus M2 protein | C-terminal | - | Oral Influenza vaccine |
| [93] | Adjuvant | human IL-2 | C-terminal | adiuvant to Helicobacter pylori vaccine | |
| CotC | |||||
| [70] | Clostridium tetani | C-term fragment of the tetanus toxin, TTFC | C-terminal N- terminal | - | Oral vaccination for tetanus |
| [75] | Clostrium difficile | C- term repeat domains of toxins A and B TcdA-TcdB | C-terminal | - | C. dìfficile oral vaccine |
| [70] | Escherichia coli | heat-labile enterotoxin B, LTB | C-terminal N- terminal | - | E. coli vaccine |
| [94] | Salmonella serovar pullorum | outer membrane protein (porin), OmpC | C-terminal | - | Salmonella vaccine |
| [72,73] | Bacillus anthracis | anthrax protective antigen, PA | C-terminal | - | Anthrax vaccine |
| [67] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | - | Anti-Helicobacter vaccine |
| [89] | Helicobacter pylori | urease subunit beta, UreB | C-terminal | - | Oral vaccine for H. pylori |
| [88] | vacuolating cytotoxin A, CagA | C-terminal N- terminal | GGGGS | ||
| [95] | cholera toxin B subunit, CTB and UreB | C-terminal | - | ||
| [78] | Clonorchis sinensis | tegumental protein 20.8 kD,TP20.8 | C-terminal | - | Liver flukes vaccine |
| [78] | tegumental protein 22.3 kDa, CsTP22.3 | C-terminal | - | ||
| [79] | cysteine proteases, CsCP | C-terminal | - | ||
| [80] | leucine aminopeptidase 2, CsLAP2 | C-terminal | - | ||
| [81] | enolase | C-terminal | - | ||
| [82] | paramyosin antigen, CsPmy | C-terminal | |||
| [83] | serpin, CsSer-3 | C-terminal | |||
| [96] | Schistosoma japonicum | 26 kDa full- length GST protein, SjGST | C-terminal | - | Liver flukes oral vaccine |
| [97] | grass carp reovirus | major outer capsid protein , VP4 | C-terminal | - | grass carp reovirus vaccine |
| [98] | White spot syndrome virus | major envelope proteins, VP28 and VP62 | C-terminal | - | Oral vaccine for shrimps |
| [99] | Bombyx mori | nucleopolyhedrovirus, GP64 | C-terminal | - | Bombyx mori vaccine |
| CotG | |||||
| [67] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | - | Anti-Helicobacter vaccine |
| [84] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEAAAKGGG | Oral vaccine against C. difficile |
| [88] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N- terminal | GGGGS | Anti-Helicobacter vaccine |
| [87] | Transmissible gastroenteritis virus | transmissible gastroenteritis virus spike , TGEV-S | C-terminal | Transmissible gastroenteritis vaccine | |
| [85] | - | Streptavidin | C-terminal | GGGGS | - |
| CotZ | |||||
| [84] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEAAAKGGG | Oral vaccine against C. difficile |
| [67] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | GGGGS | Anti-Helicobacter vaccine |
| [88] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N- terminal | GGGEAAAKGGG | Anti-Helicobacter vaccine |
| CgeA | |||||
| [88] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N- terminal | GGGEAAAKGGG | Anti-Helicobacter vaccine |
2.5. Optimizing antigen exposure on spore surface strategies.
Several strategies have been developed to augment the efficiency of spore-based mucosal vaccines since spores were genetically manipulated in 2001.
2.5.1. Linker peptides to increase stability and flexibility of recombinant protein
To ensure the folding structure of the exposed antigens, different linker peptides of 5-11 residues were introduced between the antigen and the Cot protein, working as a carrier [100,101]. The linker can affect the exposed antigen's immunogenicity by increasing the chimeric proteins' stability or flexibility. For example, the display efficiency of CgeA-CagA and CotB-UreA was remarkably increased when a linker was added between the antigen and the anchor proteins [67,88,101].
Several linker libraries with different proprieties have been utilized, and promising results have been efforted by inserting flexible linker peptides. For example, the insertion of GGGEAAAKGGG between the C-terminus of the anchor protein and the N-terminus of the exogenous protein UreA is more efficient compared with using GGGGS as the linker. However, when the latter linker was inserted between CotB and UreA, the recombinant proteins failed to be displayed on the spore surface, probably because the short linker did not allow the formation of any secondary structure, making the chimera unstable. Similar results were obtained with FliD when expressed using the same approach [84]. Therefore, the authors have speculated that an α-helix could be formed in the GGGEAAAKGGG peptide chain but not in a shorter linker as GGGGS, favoring the antigen display on the spore surface [67,101].
2.5.2. Multi antigens spore-based mucosal vaccine
For vaccine development, the crucial point is to activate multiple immune cell types to be effective against aggressive pathogens. Using spores as a vaccine delivery system can expose different molecules, multiple antigens or adjuvants, on the same spore. The deep knowledge of the genetics of B. subtilis has allowed the identification of different integration loci, such as thrC, pyrD, and gltA loci, to insert the chimeric proteins in its chromosome [102,103]. The integration of the heterologous constructs into the target genes does not perturb the sporulation rate and the spore structure and resistance [102,103]. Using alternative integration loci gives strength and flexibility to design different fusions on the spore surface simultaneously. For instance, a higher number of exposed molecules can be achieved by the co-expression of the same antigen anchored to different coat proteins on the spore surface. A collection of 16 integrative plasmids has been created for ectopic integration using the five mentioned cot proteins as anchor motifs. In particular, plasmids for obtaining N-terminal and C-terminal fusions have been constructed for CotB, CotC, and CotG, while integrator vectors have been designed to produce only C-terminal fusions for CotZ and CgeA [88].
Another advantage of the extreme versatility of spores as a vaccine-display system is the possibility of administering for immunization a unique spore presenting different antigens on the same surface or using a mixture of spores singly-expressing antigens. The immunization by different types of recombinant spores ensures a perfectly dosed amount of each antigen or adjuvant. Hinc and collaborators showed that recombinant spores expressing a fragment of the H. acinonychis UreB protein elicit a more robust cellular immune response in orally immunized mice when co-administered with spores expressing IL-2 [93].
2.5.3. Non-recombinant display of antigens on the spore surface.
The release of genetically modified microorganisms into nature and their clearance can sometimes be a drawback. To overcome this issue, a non-recombinant to display antigens on the spore surface was proposed by Huang and collaborators in 2010 [104], and various model antigens have been efficiently displayed [106,107,108,109]. This approach uses spores' negatively charged and hydrophobic surface layer to adsorb foreign proteins and is more efficient at acidic pH [110,111,112].
Antigen-adsorbed spores have been used to immunize mice mucosally, and protective humoral and cellular immune responses have been measured [109,110]. For example, a report by Isticato et al. has shown that LTB of E. coli was present on the spore surface in its pentameric native form when adsorbed on the spore surface, improving the antigen recognition by its natural receptor and activation of a proper immune response [106]. Remarkably, killed or inactivated spores, unable to germinate, appeared equally effective as live spores in stably displaying the adsorbed proteins. Although the molecular details of spore adsorption are still not totally understood, it is clear that: 1) spore-adsorbed molecules are more stable than unbound, free molecules at both high temperatures and low pH values, suggesting that the interaction with the spore stabilizes and protects the heterologous protein [113]; 2) some mutant spores, either lacking or strongly altered in their outer surface structures, are more efficient than their isogenic wild type strain in adsorbing antigens [114]; 3) adsorbed molecules are stably attached to the spore surface and can be detached only by drastic treatments [114].
Since the remarkable progress achieved in spore physiology, recombinant and non-recombinant spore-based vaccines have been improved to ensure higher immune responses. For instance, the observation that B. megaterium, B. cereus [115], and B. subtilis [116] spores of several Bacillus species produced under different laboratory conditions have structurally different coat layers has suggested an innovative approach to modulating the number of exposed antigens. In particular, in B. subtilis, CotB, CotG, and CotZ are more abundantly represented when spores are produced at low temperatures (25°C) than higher ones (37°C- 42°C). Others, such as CotC and CotU, show an opposite trend. These findings have been exploited to control the display of heterologous proteins on spores [117].
3. Conclusions
Mucosal infections are a major worldwide health concern, and it is generally accepted that mucosal vaccination strategies that can block infection at the point of entry would be preferable to other prevention approaches. However, relatively few mucosal vaccines are still available, mainly because of the need for efficient delivery systems. Recombinant bacterial spores displaying a heterologous antigen have been shown to induce protective immune responses and proposed as a mucosal delivery system. Several reasons support the use of spores as a vaccine delivery system: the remarkable and well-documented resistance of spores, which ensures high stability of the display system, the safety record of many spore-forming species, which makes spores of these species ideal candidates as vehicles to transport molecules to the surface of human mucous membranes.
Over the past two decades, significant progress in bacterial spores' morphology and physiology insights have been achieved, and the mechanisms of their beneficial effect on humans and animals have been. Advances in the understanding of spore structure have allowed at the same time, the improvement of the spore-based display system, developing different efficient approaches. Several researchers in this field have proposed combining the recombinant and non-recombinant approaches to produce single spores displaying a protein fused to a spore coat protein and a second molecule adsorbed. Potocki et al. (2017) have increased the immune response against C. difficile in mice by intranasal administration of recombinant IL-2-expressing spores previously adsorbed with the antigen FliD. Such dual display technology can be useful both to elicit a higher immune response exposing simultaneity antigen-adjuvant on a single spore and for the specific delivery of heterologous molecules. Nguyen et al. (2013) expressed CotB-streptavidin on the spore surface and bound it to a biotinylated antibody recognizing specific tumor cells. The authors showed that the recombinant spores adsorbed with a mitotic inhibitor used in cancer therapy were able to bind HT 29 colon cancer cells and thereby deliver the drug to the cells.
In conclusion, the huge flexibility of the spore delivery system allows for selecting the best-performing display approach, the number and the kind of molecules to display for the formulation of safe, economic, and targeted mucosal vaccines
Author Contributions
not applicable
Funding
No specific funds were received to carry on this work
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mojgani, N.; Shahali, Y.; Dadar, M. Immune modulatory capacity of probiotic lactic acid bacteria and applications in vaccine development. Benef. Microbes 2020, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global task force on cholera control: overview of ending cholera — a global roadmap to 2030. 2018.
- Pavot, V.; Rochereau, N.; Genin, C.; Verrier, B.; Paul, S. New insights in mucosal vaccine development. Vaccine 2012, 30, 142–154. [Google Scholar] [CrossRef]
- Kim, S.-H.; Jang, Y.-S. Antigen targeting to M cells for enhancing the efficacy of mucosal vaccines. Exp. Mol. Med. 2014, 46, e85–e85. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.J.; Curran, M.P. Live Attenuated Influenza Vaccine (FluMist®; Fluenz™). Drugs 2011, 71, 1591–1622. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.D.; Adhikari, A.; Yang, Y.; Kuschner, R.A.; Karasavvas, N.; Binn, L.N.; Walls, S.D.; Graf, P.C.F.; Myers, C.A.; Jarman, R.G.; et al. Live Oral Adenovirus Type 4 and Type 7 Vaccine Induces Durable Antibody Response. Vaccines 2020, 8, 411. [Google Scholar] [CrossRef]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines — fortifying the frontiers. Nat. Rev. Immunol. 2021, 22, 236–250. [Google Scholar] [CrossRef]
- Tsakiri, M.; Naziris, N.; Demetzos, C. Innovative vaccine platforms against infectious diseases: Under the scope of the COVID-19 pandemic. Int. J. Pharm. 2021, 610, 121212–121212. [Google Scholar] [CrossRef]
- da Silva, A.J.; Zangirolami, T.C.; Novo-Mansur, M.T.M.; Giordano, R.d.C.; Martins, E.A.L. Live bacterial vaccine vectors: An overview. Braz. J. Microbiol. 2014, 45, 1117–1129. [Google Scholar] [CrossRef]
- Smith, G.L.; Mackett, M.; Moss, B. Infectious vaccinia virus recombinants that express hepatitis B virus surface antigen. Nature 1983, 302, 490–495. [Google Scholar] [CrossRef]
- Moss, B.; Smith, G.L.; Gerin, J.L.; Purcell, R.H. Live recombinant vaccinia virus protects chimpanzees against hepatitis B. Nature 1984, 311, 67–69. [Google Scholar] [CrossRef]
- Draper, S.J.; Heeney, J.L. Viruses as vaccine vectors for infectious diseases and cancer. Nat. Rev. Microbiol. 2009, 8, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Jas, D.; Coupier, C.; Toulemonde, C.E.; Guigal, P.-M.; Poulet, H. Three-year duration of immunity in cats vaccinated with a canarypox-vectored recombinant rabies virus vaccine. Vaccine 2012, 30, 6991–6996. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and Antibody Response Caused by Turkey Herpesvirus Vector Newcastle Disease Vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef]
- FDA. First FDA-approved vaccine for the prevention of Ebola virus disease, marking a critical milestone in public health preparedness and response. 2020 Available at: https://www.fda.gov/news-events/press-announcements/first- fda-approved-vaccine-prevention.
- Afkhami, S.; D’agostino, M.R.; Zhang, A.; Stacey, H.D.; Marzok, A.; Kang, A.; Singh, R.; Bavananthasivam, J.; Ye, G.; Luo, X.; et al. Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 2022, 185, 896–915. [Google Scholar] [CrossRef]
- Peng, B.; Wang, L.R.; Gómez-Román, V.R.; Davis-Warren, A.; Montefiori, D.C.; Kalyanaraman, V.S.; Venzon, D.; Zhao, J.; Kan, E.; Rowell, T.J.; et al. Replicating Rather than Nonreplicating Adenovirus-Human Immunodeficiency Virus Recombinant Vaccines Are Better at Eliciting Potent Cellular Immunity and Priming High-Titer Antibodies. J. Virol. 2005, 79, 10200–10209. [Google Scholar] [CrossRef]
- Mielcarek, N.; Alonso, S.; Locht, C. Nasal vaccination using live bacterial vectors. Adv. Drug Deliv. Rev. 2001, 51, 55–69. [Google Scholar] [CrossRef]
- Detmer, A.; Glenting, J. Live bacterial vaccines – a review and identification of potential hazards. Microb. Cell Factories 2006, 5, 23–23. [Google Scholar] [CrossRef]
- Zhu, Q.; Berzofsky, J.A. Oral vaccines. Gut Microbes 2013, 4, 246–252. [Google Scholar] [CrossRef]
- Bastos, R.G.; Borsuk, S.; Seixas, F.K.; Dellagostin, O.A. 2009. Recom- binant Mycobacterium bovis BCG. Vaccine, 2009, 27, 6495–6503. [Google Scholar]
- Bruhn, K.W.; Craft, N.; Miller, J.F. Listeria as a vaccine vector. Microbes Infect. 2007, 9, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Curtiss, R.; Xin, W.; Li, Y. ; Kong, ; Wanda, S.Y.; Gunn, B.; et al. New technologies in using recombinant attenuated Salmonella vaccine vectors. Crit Rev Immunol, 2010, 30, 255–270. [Google Scholar]
- Roland, K.L.; A Tinge, S.; Killeen, K.P.; Kochi, S.K. Recent advances in the development of live, attenuated bacterial vectors. . 2005, 7, 62–72. [Google Scholar] [PubMed]
- Sundararaman, A. M.; Ray, P.V.; Ravindra, J.; Halami, P.M. Role of probiotics to combat viral infections with emphasison COVID-19. Appl Microbiol Biotechnol, 2020, 104, 8089–8104. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fu, T.; Hao, J.; Li, L.; Tian, M.; Jin, N.; Ren, L.; Li, C. A recombinant Lactobacillus plantarum strain expressing the spike protein of SARS-CoV-2. Int. J. Biol. Macromol. 2020, 160, 736–740. [Google Scholar] [CrossRef]
- Xu, J.; Ren, Z.; Cao, K.; Li, X.; Yang, J.; Luo, X.; Zhu, L.; Wang, X.; Ding, L.; Liang, J.; et al. Boosting Vaccine-Elicited Respiratory Mucosal and Systemic COVID-19 Immunity in Mice With the Oral Lactobacillus plantarum. Front. Nutr. 2021, 8, 789242. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.C.-C.; Liu, Y.; Wu, K.-C.; Choi, M.-C.; Ma, C.H.-Y.; Lin, J.; He, E.I.C.; Leung, D.Y.-M.; Sze, E.T.-P.; Hamied, Y.K.; et al. Retraction: Sung et al. Expression of SARS-CoV-2 Spike Protein Receptor Binding Domain on Recombinant B. subtilis on Spore Surface: A Potential COVID-19 Oral Vaccine Candidate. Vaccines 2022, 10, 2. Vaccines 2022, 10, 1852. [Google Scholar] [CrossRef]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface Display of Recombinant Proteins on Bacillus subtilis Spores. J. Bacteriol. 2001, 183, 6294–6301. [Google Scholar] [CrossRef]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ. Microbiol. Rep. 2013, 6, 212–225. [Google Scholar] [CrossRef]
- Chastanet, A.; Vitkup, D.; Yuan, G.-C.; Norman, T.M.; Liu, J.S.; Losick, R.M. Broadly heterogeneous activation of the master regulator for sporulation in Bacillus subtilis. Proc. Natl. Acad. Sci. 2010, 107, 8486–8491. [Google Scholar] [CrossRef]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2012, 11, 33–44. [Google Scholar] [CrossRef]
- Setlow, P. Germination of Spores of Bacillus Species: What We Know and Do Not Know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P.; Christie, G. Bacterial Spore mRNA – What’s Up With That? Front. Microbiol. 2020, 11, 596092. [Google Scholar] [CrossRef] [PubMed]
- Swarge, B.; Abhyankar, W.; Jonker, M.; Hoefsloot, H.; Kramer, G.; Setlow, P.; Brul, S.; de Koning, L.J. Integrative Analysis of Proteome and Transcriptome Dynamics during Bacillus subtilis Spore Revival. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed]
- Koopman, N.; Remijas, L.; Seppen, J.; Setlow, P.; Brul, S. Mechanisms and Applications of Bacterial Sporulation and Germination in the Intestine. Int. J. Mol. Sci. 2022, 23, 3405. [Google Scholar] [CrossRef] [PubMed]
- Casula, G.; Cutting, S.M. Bacillus Probiotics: Spore Germination in the Gastrointestinal Tract. Appl. Environ. Microbiol. 2002, 68, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Duc, L.H.; Hong, A.H.; Nguyen, Q.U.; Cutting, S.M. Intracellular fate and immunogenicity of B. subtilis spores. Vaccine, 2004, 22, 1873–1885. [Google Scholar] [CrossRef]
- Tam, N.K.M.; Uyen, N.Q.; Hong, H.A.; Duc, L.H.; Hoa, T.T.; Serra, C.R.; Henriques, A.O.; Cutting, S.M. The Intestinal Life Cycle of Bacillus subtilis and Close Relatives. J. Bacteriol. 2006, 188, 2692–2700. [Google Scholar] [CrossRef]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the gastrointestinal life cycle of Bacillus for probiotic functionality. J. Food Sci. Technol. 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- Elshaghabee, E.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef]
- McKenney, P.T.; Eichenberger, P. Dynamics of spore coat morphogenesis in Bacillus subtilis. Mol. Microbiol. 2011, 83, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Shuster, B.; Khemmani, M.; Abe, K.; Huang, X.; Nakaya, Y.; Maryn, N.; Buttar, S.; Gonzalez, A.N.; Driks, A.; Sato, T.; et al. Contributions of crust proteins to spore surface properties in Bacillus subtilis. Mol. Microbiol. 2018, 111, 825–843. [Google Scholar] [CrossRef]
- Cangiano, G.; Mazzone, A.; Baccigalupi, L.; Isticato, R.; Eichenberger, P.; De Felice, M.; Ricca, E. Direct and Indirect Control of Late Sporulation Genes by GerR of Bacillus subtilis. J. Bacteriol. 2010, 192, 3406–3413. [Google Scholar] [CrossRef] [PubMed]
- Ursem, R.; Swarge, B.; Abhyankar, W.R.; Buncherd, H.; de Koning, L.J.; Setlow, P.; Brul, S.; Kramer, G. Identification of Native Cross-Links in Bacillus subtilis Spore Coat Proteins. J. Proteome Res. 2021, 20, 1809–1816. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Pelosi, A.; Zilhão, R.; Baccigalupi, L.; Henriques, A.O.; De Felice, M.; Ricca, E. CotC-CotU Heterodimerization during Assembly of the Bacillus subtilis Spore Coat. J. Bacteriol. 2008, 190, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Sirec, T.; Vecchione, S.; Crispino, A.; Saggese, A.; Baccigalupi, L.; Notomista, E.; Driks, A.; Ricca, E. The Direct Interaction between Two Morphogenetic Proteins Is Essential for Spore Coat Formation in Bacillus subtilis. PLOS ONE 2015, 10, e0141040. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Pelosi, A.; De Felice, M.; Ricca, E. CotE Binds to CotC and CotU and Mediates Their Interaction during Spore Coat Formation in Bacillus subtilis. J. Bacteriol. 2010, 192, 949–954. [Google Scholar] [CrossRef]
- Isticato, R.; Sirec, T.; Giglio, R.; Baccigalupi, L.; Rusciano, G.; Pesce, G.; Zito, G.; Sasso, A.; De Felice, M.; Ricca, E. Flexibility of the Prograamme of Spore Coat Formation in Bacillus subtilis: Bypass of CotE Requirement by Over-Production of CotH. PLOS ONE 2013, 8, e74949. [Google Scholar] [CrossRef]
- Cangiano, G.; Sirec, T.; Panarella, C.; Isticato, R.; Baccigalupi, L.; De Felice, M.; Ricca, E. The sps gene products affect germination, hydrophobicity and protein adsorption of Bacillus subtilis spores. Appl Environ Microbiol, 2014, 80, 7293–7302. [Google Scholar] [CrossRef]
- Ricca, E.; Baccigalupi, L.; Isticato, R. Spore-adsorption: Mechanism and applications of a non-recombinant display system. Biotechnol. Adv. 2020, 47, 107693. [Google Scholar] [CrossRef]
- Driks, A.; Eichenberger, P. The Spore Coat. Microbiol Spectr, 2016, 4(2).
- Pesce, G.; Rusciano, G.; Sasso, A.; Isticato, R.; Sirec, T.; Ricca, E. Surface charge and hydrodynamic coefficient measurements of Bacillus subtilis spore by optical tweezers. Colloids Surfaces B: Biointerfaces 2014, 116, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Dubois, T.; Krzewinski, F.; Yamakawa, N.; Lemy, C.; Hamiot, A.; Brunet, L.; Lacoste, A.-S.; Knirel, Y.; Guerardel, Y.; Faille, C. The sps Genes Encode an Original Legionaminic Acid Pathway Required for Crust Assembly in Bacillus subtilis. Mbio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.C. The Exosporium Layer of Bacterial Spores: a Connection to the Environment and the Infected Host. Microbiol. Mol. Biol. Rev. 2015, 79, 437–457. [Google Scholar] [CrossRef] [PubMed]
- Vittoria, M.; Saggese, A.; Barletta, G.D.G.; Castaldi, S.; Isticato, R.; Baccigalupi, L.; Ricca, E. Sporulation efficiency and spore quality in a human intestinal isolate of Bacillus cereus. Res. Microbiol. 2023, 174, 104030. [Google Scholar] [CrossRef] [PubMed]
- Sirec, T.; Cangiano, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The spore surface of intestinal isolates ofBacillus subtilis. FEMS Microbiol. Lett. 2014, 358, 194–201. [Google Scholar] [CrossRef]
- Isticato, R.; Ricca, E. Spore Surface Display. Microbiol. Spectr. 2014, 2, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Yuan, H.; Du, J.; Liu, K.; Liu, H.; Wang, T. Progress in research and application development of surface display technology using Bacillus subtilis spores. Appl. Microbiol. Biotechnol. 2020, 104, 2319–2331. [Google Scholar] [CrossRef]
- Chen, H.; Ullah, J.; Jia, J. Progress in Bacillus subtilis Spore Surface Display Technology towards Environment, Vaccine Development, and Biocatalysis. Microb. Physiol. 2017, 27, 159–167. [Google Scholar] [CrossRef]
- Tang, Z.; Wu, Z.; Sun, H.; Zhao, L.; Shang, M.; Shi, M.; Jiang, H.; Lin, Z.; Zhou, X.; Li, X.; et al. The storage stability of Bacillus subtilis spore displaying cysteine protease of Clonorchis sinensis and its effect on improving the gut microbiota of mice. Appl. Microbiol. Biotechnol. 2021, 105, 2513–2526. [Google Scholar] [CrossRef]
- Lee, N.-K.; Kim, W.-S.; Paik, H.-D. Bacillus strains as human probiotics: characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Knecht, L.D.; Pasini, P.; Daunert, S. Bacterial spores as platforms for bioanalytical and biomedical applications. Anal. Bioanal. Chem. 2011, 400, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Scheiblhofer, S.; Laimer, J.; Machado, Y.; Weiss, R.; Thalhamer, J. Influence of protein fold stability on immunogenicity and its implications for vaccine design. Expert Rev. Vaccines 2017, 16, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Turk, B. Targeting proteases: successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef]
- Hinc, K.; Isticato, R.; Dembek, M.; Karczewska, J.; Iwanicki, A.; Peszyńska-Sularz, G.; De Felice, M.; Obuchowski, M.; Ricca, E. Expression and display of UreA of Helicobacter acinonychis on the surface of Bacillus subtilis spores. Microb. Cell Factories 2010, 9, 2–2. [Google Scholar] [CrossRef] [PubMed]
- Zilhão, R.; Serrano, M.; Isticato, R.; Ricca, E.; Moran, C.P.; Henriques, A.O. Interactions among CotB, CotG, and CotH during Assembly of the Bacillus subtilis Spore Coat. J. Bacteriol. 2004, 186, 1110–1119. [Google Scholar] [CrossRef]
- Giglio, R.; Fani, R.; Isticato, R.; De Felice, M.; Ricca, E.; Baccigalupi, L. Organization and Evolution of the cotG and cotH Genes of Bacillus subtilis. J. Bacteriol. 2011, 193, 6664–6673. [Google Scholar] [CrossRef]
- Mauriello, E.M.; Duc, L.H.; Isticato, R.; Cangiano, G.; Hong, H.A.; De Felice, M.; Ricca, E.; Cutting, S.M. Display of heterologous antigens on the Bacillus subtilis spore coat using CotC as a fusion partner. Vaccine 2004, 22, 1177–1187. [Google Scholar] [CrossRef]
- Ciabattini, A.; Parigi, R.; Isticato, R.; Oggioni, M.R.; Pozzi, G. Oral priming of mice by recombinant spores of Bacillus subtilis. Vaccine 2004, 22, 4139–4143. [Google Scholar] [CrossRef]
- Duc, L.H.; Hong, H.A.; Atkins, H.S.; Flick-Smith, H.C.; Durrani, Z.; Rijpkema, S.; Titball, R.W.; Cutting, S.M. Immunization against anthrax using Bacillus subtilis spores expressing the anthrax protective antigen. Vaccine 2007, 25, 346–355. [Google Scholar] [CrossRef]
- Oh, Y.; Kim, J.A.; Kim, C.-H.; Choi, S.-K.; Pan, J.-G. Bacillus subtilis spore vaccines displaying protective antigen induce functional antibodies and protective potency. BMC Veter- Res. 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Hoang, T.H.; Hong, H.A.; Clark, G.C.; Titball, R.W.; Cutting, S.M. RecombinantBacillus subtilisExpressing theClostridium perfringensAlpha Toxoid Is a Candidate Orally Delivered Vaccine against Necrotic Enteritis. Infect. Immun. 2008, 76, 5257–5265. [Google Scholar] [CrossRef]
- Hong, H.A.; Hitri, K.; Hosseini, S.; Kotowicz, N.; Bryan, D.; Mawas, F.; Wilkinson, A.J.; van Broekhoven, A.; Kearsey, J.; Cutting, S.M. Mucosal Antibodies to the C Terminus of Toxin A Prevent Colonization of Clostridium difficile. Infect. Immun. 2017, 85, e01060–16. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Pham, C.K.; Pham, H.T.; Pham, H.L.; Dang, L.T.; Huynh, H.A.; Cutting, S.M.; Phan, T.-N. Bacillus subtilisspores expressing the VP28 antigen: a potential oral treatment to protectLitopenaeus vannameiagainst white spot syndrome. FEMS Microbiol. Lett. 2014, 358, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Di Mase, D.S.; Mauriello, E.M.; De Felice, M.; Ricca, E. Amino terminal fusion of heterologous proteins to CotC increases display efficiencies in the Bacillus subtilis spore system. BioTechniques 2007, 42, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xia, H.; Hu, X.; Huang, Y.; Li, Y.; Li, L.; Ma, C.; Chen, X.; Hu, F.; Xu, J.; et al. Oral administration of a Bacillus subtilis spore-based vaccine expressing Clonorchis sinensis tegumental protein 22.3kDa confers protection against Clonorchis sinensis. Vaccine 2008, 26, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Shang, M.; Chen, T.; Ren, P.; Sun, H.; Qu, H.; Lin, Z.; Zhou, L.; Yu, J.; Jiang, H.; et al. The immunological characteristics and probiotic function of recombinant Bacillus subtilis spore expressing Clonorchis sinensis cysteine protease. Parasites Vectors 2016, 9, 1–14. [Google Scholar] [CrossRef]
- Qu, H.; Xu, Y.; Sun, H.; Lin, J.; Yu, J.; Tang, Z.; Shen, J.; Liang, C.; Li, S.; Chen, W.; et al. Systemic and local mucosal immune responses induced by orally delivered Bacillus subtilis spore expressing leucine aminopeptidase 2 of Clonorchis sinensis. Parasitol. Res. 2014, 113, 3095–3103. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, T.; Xie, Z.; Liang, P.; Qu, H.; Shang, M.; Mao, Q.; Ning, D.; Tang, Z.; Shi, M.; et al. Oral delivery of Bacillus subtilis spore expressing enolase of Clonorchis sinensis in rat model: induce systemic and local mucosal immune responses and has no side effect on liver function. Parasitol. Res. 2015, 114, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lin, Z.; Zhao, L.; Chen, T.; Shang, M.; Jiang, H.; Tang, Z.; Zhou, X.; Shi, M.; Zhou, L.; et al. Bacillus subtilis spore with surface display of paramyosin from Clonorchis sinensis potentializes a promising oral vaccine candidate. Parasites Vectors 2018, 11, 1–15. [Google Scholar] [CrossRef]
- Lin, Z.; Sun, H.; Ma, Y.; Zhou, X.; Jiang, H.; Wang, X.; Song, J.; Tang, Z.; Bian, Q.; Zhang, Z.; et al. Evaluation of immune response toBacillus subtilisspores expressingClonorchis sinensisserpin3. Parasitology 2020, 147, 1080–1087. [Google Scholar] [CrossRef]
- Negri, A.; Potocki, W.; Iwanicki, A.; Obuchowski, M.; Hinc, K. Expression and display of Clostridium difficile protein FliD on the surface of Bacillus subtilis spores. J. Med Microbiol. 2013, 62, 1379–1385. [Google Scholar] [CrossRef]
- Kim, J.-H.; Lee, C.-S.; Kim, B.-G. Spore-displayed streptavidin: A live diagnostic tool in biotechnology. Biochem. Biophys. Res. Commun. 2005, 331, 210–214. [Google Scholar] [CrossRef]
- Kim., J.; Schumann, W. Kim. J.; Schumann, W. Display of proteins on Bacillus subtilis endospores. Cell Mol Life Sci, 2009, 66(19),3127–313.
- Mou, C.; Zhu, L.; Xing, X.; Lin, J.; Yang, Q. Immune responses induced by recombinant Bacillus subtilis expressing the spike protein of transmissible gastroenteritis virus in pigs. Antivir. Res. 2016, 131, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Iwanicki, A.; Piątek, I.; Stasiłojć, M.; Grela, A.; Łęga, T.; Obuchowski, M.; Hinc, K. A system of vectors for Bacillus subtilis spore surface display. Microb. Cell Factories 2014, 13, 30–30. [Google Scholar] [CrossRef] [PubMed]
- Stasiłojć, M.; Hinc, K.; Peszyńska-Sularz, G.; Obuchowski, M.; Iwanicki, A. Recombinant Bacillus subtilis Spores Elicit Th1/Th17-Polarized Immune Response in a Murine Model of Helicobacter pylori Vaccination. Mol. Biotechnol. 2015, 57, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Sibley, L.; Reljic, R.; Radford, D.S.; Huang, J.M.; Hong, H.A.; Cranenburgh, R.M.; et al. 2014. Recombinant Bacillus subtilis spores expressing MPT64 evaluated as a vaccine against tuberculosis in the murine model. FEMS Microbiol. Lett, 2014, 358, 170–179. [Google Scholar] [PubMed]
- Tavares, M.B.; Silva, B.M.; Cavalcante, R.C.; Souza, R.D.; Luiz, W.B.; Paccez, J.D.; Crowley, P.J.; Brady, L.J.; Ferreira, L.C.; Ferreira, R.C. Induction of neutralizing antibodies in mice immunized with an amino-terminal polypeptide ofStreptococcus mutansP1 protein produced by a recombinantBacillus subtilisstrain. FEMS Immunol. Med Microbiol. 2010, 59, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G. ; Miao,Y. ; Guo,Y.; Qiu, H.; Sun, S.; Kou, Z.; et al. Development of a heat-stable and orally delivered recombinant M2e-expressing B. subtilis spore-based influenza vaccine. Hum Vaccin Immunother, 2014, 10, 3649–3658. [Google Scholar]
- Hinc, K.; Stasiłojć, M.; Piątek, I.; Peszyńska-Sularz, G.; Isticato, R.; Ricca, E.; Obuchowski, M.; Iwanicki, A. Mucosal Adjuvant Activity of IL-2 Presenting Spores of Bacillus subtilis in a Murine Model of Helicobacter pylori Vaccination. PLOS ONE 2014, 9, e95187. [Google Scholar] [CrossRef]
- Dai, X.; Liu, M.; Pan, K.; Yang, J. Surface display of OmpC of Salmonella serovar Pullorum on Bacillus subtilis spores. PLOS ONE 2018, 13, e0191627. [Google Scholar] [CrossRef]
- Zhou, Z.; Gong, S.; Li, X.-M.; Yang, Y.; Guan, R.; Zhou, S.; Yao, S.; Xie, Y.; Ou, Z.; Zhao, J.; et al. Expression of Helicobacter pylori urease B on the surface of Bacillus subtilis spores. J. Med Microbiol. 2015, 64, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hu, X.; Wu, Z.; Xiong, S.; Zhou, Z.; Wang, X.; Xu, J.; Lu, F.; Yu, X. Immunogenicity of self-adjuvanticity oral vaccine candidate based on use of Bacillus subtilis spore displaying Schistosoma japonicum 26 KDa GST protein. Parasitol. Res. 2009, 105, 1643–1651. [Google Scholar] [CrossRef]
- Jiang, H.; Bian, Q.; Zeng, W.; Ren, P.; Sun, H.; Lin, Z.; Tang, Z.; Zhou, X.; Wang, Q.; Wang, Y.; et al. Oral delivery of Bacillus subtilis spores expressing grass carp reovirus VP4 protein produces protection against grass carp reovirus infection. Fish Shellfish. Immunol. 2018, 84, 768–780. [Google Scholar] [CrossRef]
- Valdez, A.; Yepiz-Plascencia, G.; Ricca, E.; Olmos, J. First Litopenaeus vannamei WSSV 100% oral vaccination protection using CotC::Vp26 fusion protein displayed on Bacillus subtilis spores surface. J. Appl. Microbiol. 2014, 117, 347–357. [Google Scholar] [CrossRef]
- Li, G.; Tang, Q.; Chen, H.; Yao, Q.; Ning, D.; Chen, K. Display of Bombyx mori Nucleopolyhedrovirus GP64 on the Bacillus subtilis Spore Coat. Curr. Microbiol. 2011, 62, 1368–1373. [Google Scholar] [CrossRef]
- Hinc, K.; Iwanicki, A.; Obuchowski, M. New stable anchor protein and peptide linker suitable for successful spore surface display in B. subtilis. Microb. Cell Factories 2013, 12, 22–22. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Li, G.; Zhang, C.; Xing, X.-H. A study on the effects of linker flexibility on acid phosphatase PhoC-GFP fusion protein using a novel linker library. Enzym. Microb. Technol. 2016, 83, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guérout-Fleury, A.-M.; Frandsen, N.; Stragier, P. Plasmids for ectopic integration in Bacillus subtilis. Gene 1996, 180, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Middleton, R.; Hofmeister, A. New shuttle vectors for ectopic insertion of genes into Bacillus subtilis. Plasmid 2004, 51, 238–245. [Google Scholar] [CrossRef]
- Huang, J.-M.; Hong, H.A.; Van Tong, H.; Hoang, T.H.; Brisson, A.; Cutting, S.M. Mucosal delivery of antigens using adsorption to bacterial spores. Vaccine 2010, 28, 1021–1030. [Google Scholar] [CrossRef]
- Bonavita, R.; Isticato, R.; Maurano, F.; Ricca, E.; Rossi, M. Mucosal immunity induced by gliadin-presenting spores of Bacillus subtilis in HLA-DQ8-transgenic mice. Immunol. Lett. 2015, 165, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Sirec, T.; Treppiccione, L.; Maurano, F.; De Felice, M.; Rossi, M.; Ricca, E. Non-recombinant display of the B subunit of the heat labile toxin of Escherichia coli on wild type and mutant spores of Bacillus subtilis. Microb. Cell Factories 2013, 12, 98–98. [Google Scholar] [CrossRef]
- Maia, A.R.; Reyes-Ramírez, R.; Pizarro-Guajardo, M.; Saggese, A.; Castro-Córdova, P.; Isticato, R.; Ricca, E.; Paredes-Sabja, D.; Baccigalupi, L. Induction of a Specific Humoral Immune Response by Nasal Delivery of Bcla2ctd of Clostridioides difficile. Int. J. Mol. Sci. 2020, 21, 1277. [Google Scholar] [CrossRef]
- Santos, F.D.S.; Mazzoli, A.; Maia, A.R.; Saggese, A.; Isticato, R.; Leite, F.; Iossa, S.; Ricca, E.; Baccigalupi, L. A probiotic treatment increases the immune response induced by the nasal delivery of spore-adsorbed TTFC. Microb. Cell Factories 2020, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, M.E.M.; Alves, K.C.S.; de Vasconcelos, M.G.S.; Pinto, T.S.; Glória, J.C.; Chaves, Y.O.; Neves, W.L.L.; Tarragô, A.M.; Neto, J.N.d.S.; Astolfi-Filho, S.; et al. Bacillus subtilis spores as delivery system for nasal Plasmodium falciparum circumsporozoite surface protein immunization in a murine model. Sci. Rep. 2022, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ricca, E.; Baccigalupi, L.; Cangiano, G.; De Felice, M.; Isticato, R. Mucosal vaccine delivery by non-recombinant spores of Bacillus subtilis. Microb. Cell Factories 2014, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pesce, G.; Rusciano, G.; Sasso, A.; Isticato, R.; Sirec, T.; Ricca, E. Surface charge and hydrodynamic coefficient measurements of Bacillus subtilis spore by optical tweezers. Colloids Surfaces B: Biointerfaces 2014, 116, 568–575. [Google Scholar] [CrossRef]
- Sirec, T.; Cangiano, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The spore surface of intestinal isolates ofBacillus subtilis. FEMS Microbiol. Lett. 2014, 358, 194–201. [Google Scholar] [CrossRef]
- Donadio, G.; Lanzilli, M.; Sirec, T.; Ricca, E.; Isticato, R. Localization of a red fluorescence protein adsorbed on wild type and mutant spores of Bacillus subtilis. Microb. Cell Factories 2016, 15, 1–12. [Google Scholar] [CrossRef]
- Lanzilli, M.; Donadio, G.; Fusco, F.A.; Sarcinelli, C.; Limauro, D.; Ricca, E.; Isticato, R. Display of the peroxiredoxin Bcp1 of Sulfolobus solfataricus on probiotic spores of Bacillus megaterium. New Biotechnol. 2018, 46, 38–44. [Google Scholar] [CrossRef]
- Bressuire-Isoard, C.; Bornard, I.; Henriques, A.O.; Carlin, F.; Broussolle, V. Sporulation Temperature Reveals a Requirement for CotE in the Assembly of both the Coat and Exosporium Layers of Bacillus cereus Spores. Appl. Environ. Microbiol. 2016, 82, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Lanzilli, M.; Petrillo, C.; Donadio, G.; Baccigalupi, L.; Ricca, E. Bacillus subtilis builds structurally and functionally different spores in response to the temperature of growth. Environ. Microbiol. 2019, 22, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, C.; Castaldi, S.; Lanzilli, M.; Saggese, A.; Donadio, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The temperature of growth and sporulation modulates the efficiency of spore-display in Bacillus subtilis. Microb. Cell Factories 2020, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Licensed mucosal vaccine formulations for human use. Nine mucosal vaccines are currently being used against cholera, salmonella, acute gastroenteritis, influenza, poliovirus, rotavirus, and adenovirus. To date, live attenuated and inactive vaccines have proven to be the most effective platforms for developing mucosal vaccines. Excluding the influenza vaccines that use the intranasal route, the remaining eight vaccines are all orally administrated. CTB: cholera toxin B subunit; LPS, lipopolysaccharide.
Figure 1.
Licensed mucosal vaccine formulations for human use. Nine mucosal vaccines are currently being used against cholera, salmonella, acute gastroenteritis, influenza, poliovirus, rotavirus, and adenovirus. To date, live attenuated and inactive vaccines have proven to be the most effective platforms for developing mucosal vaccines. Excluding the influenza vaccines that use the intranasal route, the remaining eight vaccines are all orally administrated. CTB: cholera toxin B subunit; LPS, lipopolysaccharide.
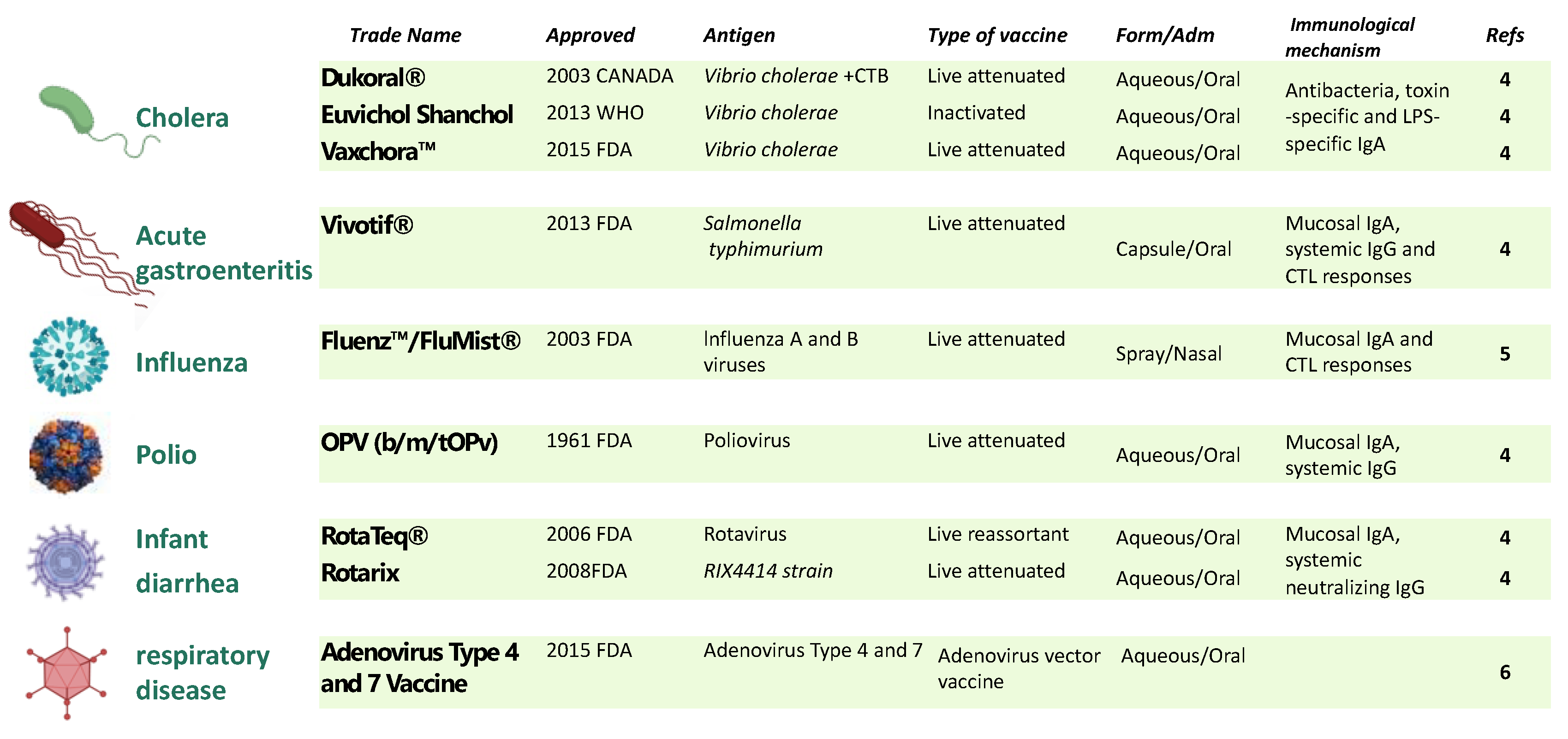
Figure 2.
Advantages and disadvantages of mucosal vaccine development.
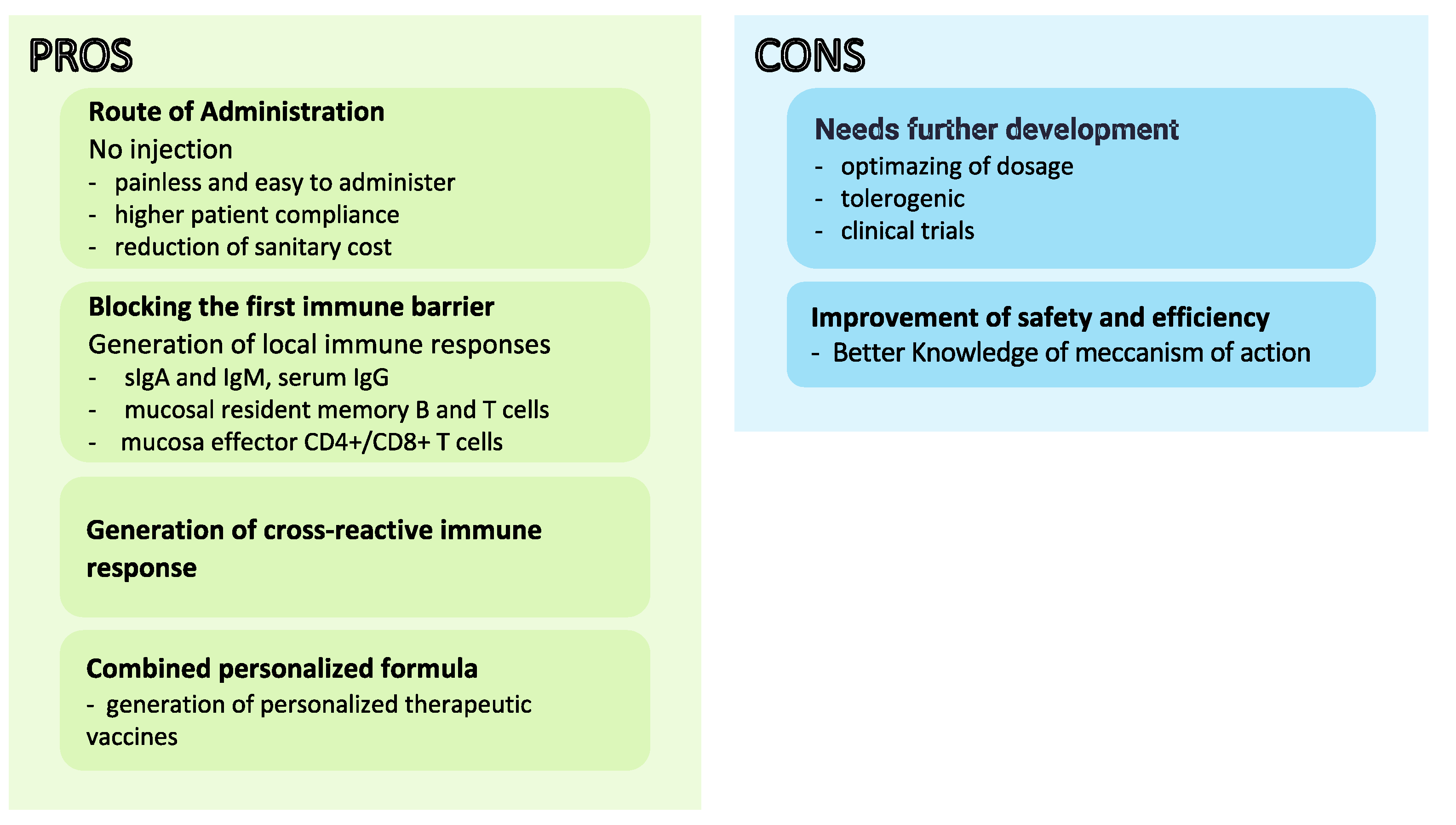
Figure 3.
Schematic representation of sporulation and germination in B. subtilis. The sporulation process starts with forming an asymmetrical septum that produces two different compartments: a bigger one called the mother cell and the forespore, the smaller one. Next, the mother cell engulfs the forespore, and following membrane fission at the opposite pole of the sporangium, a double-membrane bound forespore is formed. After the engulfment and throughout sporulation, several layers are placed around the forespore: the cortex, modified peptidoglycan, is deposited between the inner and outer forespore membranes, while the proteins of the coat and then the glycoproteins of the crust are synthesized in the mother cell and assembled on the forespore surface. In the final step, the mother cell lyses to release a mature spore into the environment. If the environmental conditions ameliorate, the spore can quickly germinate and resume vegetative growth.
Figure 3.
Schematic representation of sporulation and germination in B. subtilis. The sporulation process starts with forming an asymmetrical septum that produces two different compartments: a bigger one called the mother cell and the forespore, the smaller one. Next, the mother cell engulfs the forespore, and following membrane fission at the opposite pole of the sporangium, a double-membrane bound forespore is formed. After the engulfment and throughout sporulation, several layers are placed around the forespore: the cortex, modified peptidoglycan, is deposited between the inner and outer forespore membranes, while the proteins of the coat and then the glycoproteins of the crust are synthesized in the mother cell and assembled on the forespore surface. In the final step, the mother cell lyses to release a mature spore into the environment. If the environmental conditions ameliorate, the spore can quickly germinate and resume vegetative growth.
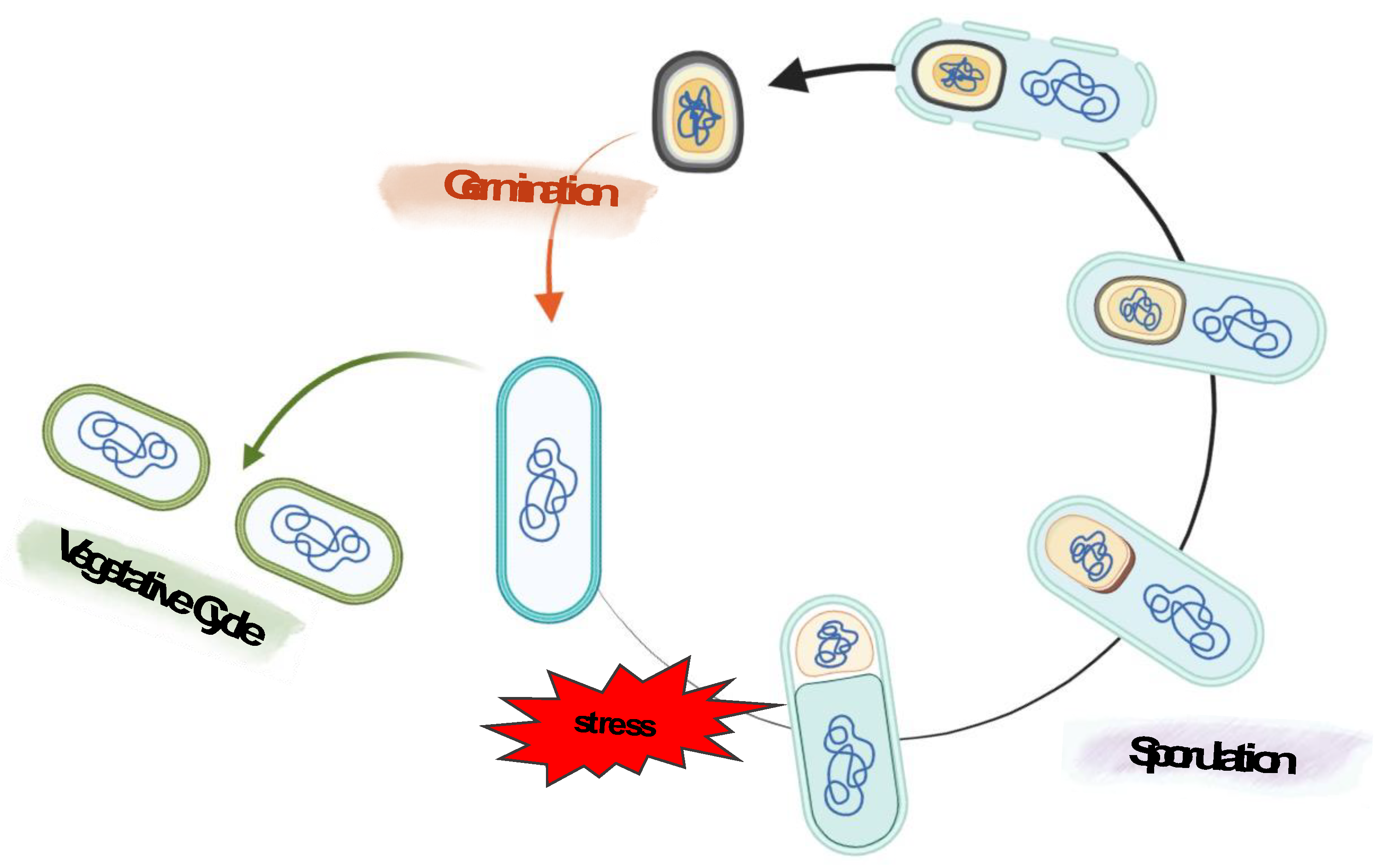
Figure 6.
Strategy for a recombinant display of antigen on the spore surface. The protein chimera is obtained by fusing the DNA coding for Cot protein (Anchor coat gene in grey) to the gene of the selected antigen (Antigen gene in yellow). The gene fusion is then cloned into an integrative vector between two regions of a non-essential gene of B. subtilis (in blue). The fusion is under the transcriptional and translational signals of the Cot protein chosen as the anchor (Pr in light grey). Similar strategies are used to obtain N-terminus, C-terminus, and sandwich fusions.
Figure 6.
Strategy for a recombinant display of antigen on the spore surface. The protein chimera is obtained by fusing the DNA coding for Cot protein (Anchor coat gene in grey) to the gene of the selected antigen (Antigen gene in yellow). The gene fusion is then cloned into an integrative vector between two regions of a non-essential gene of B. subtilis (in blue). The fusion is under the transcriptional and translational signals of the Cot protein chosen as the anchor (Pr in light grey). Similar strategies are used to obtain N-terminus, C-terminus, and sandwich fusions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



