
Submitted:
27 April 2023
Posted:
28 April 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background: This study investigated the biological effects on circulating monocytes after challenge with SARS-CoV-2 recombinant spike protein. Methods: Whole blood collected form seven ostensibly healthy healthcare workers was incubated for 15 min with 2 and 20 ng/mL recombinant spike protein of Ancestral, Alpha, Delta and Omicron variants. Samples were analyzed with Sysmex XN and DI-60 analyzers. Results: Cellular complexity (i.e., presence of granules, vacuoles and other cytoplasmic inclusions) increased in all samples challenged with the recombinant spike protein of Ancestral, Alpha and Delta variants, but not in those containing Omicron. The cellular content of nucleic acids was constantly decreased in most samples, achieving statistical significance in those containing 20 ng/mL of Alpha and Delta recombinant spike proteins. The heterogeneity of monocyte volumes significantly increased in all samples, achieving statistical significance in those containing 20 ng/mL of recombinant spike protein of Ancestral, Alpha and Delta variants. The monocyte morphological abnormalities after spike protein challenge included dysmorphia, granulation, intense vacuolization, platelet phagocytosis, development of aberrant nuclei and cytoplasmic extrusions. Conclusions: The SARS-CoV-2 spike protein triggers important monocyte morphological abnormalities, more evident in cells challenged with spike protein of the more clinically severe Alpha and Delta variants.
Keywords:
SARS-CoV-2
; COVID-19
; Monocyte
; Spike protein
; Morphology.
1. Introduction
The coronavirus disease 2019 (COVID-19) is the most recent life-threatening infectious disease to have reached pandemic proportions in human history [1]. More than three years after its emergence, and with over 7 million attributable deaths, SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2) infections are still raising international concern, such that the World Health Organization (WHO) has reiterated the status of public health emergency [2].
Although some aspects of the intricate pathogenesis of COVID-19 remain unraveled, there is now incontrovertible evidence that SARS-CoV-2 is capable of infecting a vast array of human cells besides type II pneumocytes. This is supported by the evidence that the virus can not only bind to its principal host cell receptor angiotensin converting enzyme 2 (ACE2), but can also associate to a variety of other proteins present at the cell surface such as heparan sulfate, proteoglycans, phosphatidylserine receptor, neuropilin-1, CD147, C-type lectins, SLC1A5, AXL tyrosine-kinase cellular receptor (AXL), kringle containing transmembrane protein 1 (KREMEN1), asialoglycoprotein receptor-1, low density lipoprotein receptor class A domain 3 (LDLRAD3), transmembrane protein 30A (TMEM30A), which may directly promote or foster the process of host cell invasion [3,4]. Additional mechanisms of cell entry have been identified, especially characterizing the latest viral variants belonging to the Omicron sub-lineages, and encompassing cathepsin-mediated endocytosis, cell-to-cell diffusion and internalization of SARS-CoV-2-bearing extracellular microparticles generated by infected cells [5].
Throughout the early history of COVID-19, one fact that clearly emerged through many experimental and clinical studies, is that the circulating monocytes play a pivotal role in the pathogenesis of this condition [6]. Succinctly, monocytes are not only recruited to the site of infection where they can further differentiate into tissue macrophages or dendritic cells, but they also directly interplay with SARS-CoV-2 in blood, becoming infected, activated and stimulated to produce a variety of cytokines and chemokines that may support sustained (either localized or systemic) inflammatory reactions [7], which has been conventionally called the “cytokine storm” [8]. Incidentally, both the number and degree of activation of classical monocytes are consistently elevated in patients with COVID-19, especially those expressing CD169 at their surface, a hallmark of monocyte activation [9], whilst their overall number tends to gradually decrease afterwards, as a result of cell injury, degeneration and death directly caused by the virus or by some of its components [10].
Besides the direct role played by the integral viral particle, several lines of evidence suggest that its spike protein may also directly interplay with human monocytes, triggering their activation and thus promoting, maintaining or even boosting the strong inflammatory reaction that characterizes the more severe cases of COVID-19 [11]. Nonetheless, since the pathogenicity of SARS-CoV-2 has considerably changed over time, generating a lower clinical burden in the infected individuals, we planned this series of experiments to investigate whether different types of recombinant SARS-CoV-2 spike protein may produce diverse alternations of circulating monocyte morphology.
2. Materials and Methods
Whole venous blood samples were drawn from seven ostensibly healthy healthcare workers (mean age 50±7 years; 5 females) in the early morning, into nine consecutive evacuated blood tubes containing 3.2 M buffered sodium citrate. All blood samples were left standing in vertical position for at least 15 min after collection, and then used for the experiments. Specifically, one blood sample was spiked with 11.6 µL saline, while in the other samples different variants and concentrations of recombinant SARS-CoV-2 trimeric spike proteins were added (ACROBiosystems, Newark, DE, USA; Ancestral spike: Cat. No. SPN-C52H3; Alpha spike: Cat. No. SPN-C52H6, Delta spike: Cat. No. SPN-C52He; Omicron spike: Cat. No. SPN-C52Hz)) (Table 1).
The material, lyophilized from 0.22 μm filtered solution in PBS with trehalose, is certified from the company, displays over >90% purity as established by Sodium Dodecyl Sulphate - PolyAcrylamide Gel Electrophoresis (SDS-PAGE) and Size-Exclusion Chromatography – Multi-Angle Light Scattering (SEC-MALS), and is stable in lyophilized form for 12 months at -20°C to -70°C. The resuspended protein is also stable for approximately 3 weeks at 4°C. The lyophilized recombinant SARS-CoV-2 trimetric spike proteins of the Ancestral, Alpha, Delta and Omicron viral variants were resuspended with saline, producing two stock solutions with concentration of 6×106 and 6×105 ng/mL, respectively. The remaining eight whole blood anticoagulated samples were then spiked with 11.6 µL of each spike protein variant and with both solutions, achieving final concentrations in the test samples of 2 and 20 ng/mL, respectively, which reflects an interval of blood values of SARS-CoV-2 spike protein that could be usually measured in patients with acute COVID-19 [12]. An interval of 15 min was observed for processing one sample from the previous, thus allowing to standardize the time of incubation with the recombinant spike protein (5 min) and for concluding each set of experiments from each single donor within 90 min, which is a guarantee of sample stability [13].
After sample spiking, the specimens were hence gently mixed and left standing for 5 min in vertical position allowing for the interaction between the recombinant spike protein and blood cells. After such period, the samples were tested on a Sysmex XN hematological analyzer (Sysmex Corporation, Kobe, Japan) connected to the DI-60 digital morphology analyzer (Sysmex Corporation, Kobe, Japan), for obtaining the monocyte parameters. The assessment of leukocytes within a flow chamber in this hematological instrumentation encompasses their illumination by means of laser light, followed by measurement of scattered light in all directions, generating accurate information about fluorescence intensity, complexity, size and width of dispersion of events scattered on the three axis of the white blood cell (WBC) differential (WDF) channel, as comprehensively described elsewhere [14]. The monocyte parameters reported on the x-axis included monocytes cells complexity (MO-X) and the relative width of dispersion of monocytes complexity (MO-WX), the parameters reported on the y-axis included monocytes fluorescence intensity (MO-Y) and the relative width of dispersion of monocytes fluorescence intensity (MO-WY), whilst those reported on the z-axis encompass monocytes cell size (MO-Z) and the relative width of dispersion of monocytes cell size (MO-WZ). These parameters and their specific clinical significance are summarized in Table 2 [14].
Results of all these experiments were finally reported as mean ± SD (standard deviation), whilst the significance of variation from the paired control specimen (i.e., only saline spiked) were assessed with Mann-Whitney paired test (Analyse-it; Analyse-it Software Ltd, Leeds, UK). All subjects provided an informed consent for being included in this study, which was performed in accordance with the Helsinki Declaration and was approved by the Ethics Committee of the Provinces of Verona and Rovigo (2622CESC).
3. Results
The results of this study are summarized in Figure 1, Figure 2 and Figure 3. The values of MO-X, reflecting cellular complexity for the presence of granules, vacuoles and other cytoplasmic inclusions, increased as compared to control in all samples where SARS-CoV-2 recombinant spike protein of Ancestral, Alpha and Delta variants were added, but not in those containing the Omicron variant, with values being typically higher in samples spiked with the higher 20 ng/mL final concentration compared to those challenged with 2 ng/mL.
The heterogeneity of monocytes complexity was also found to be consistently increased in all samples spiked with the two concentrations of the four SARS-CoV-2 recombinant spike protein variants, achieving statistical significance in samples containing Alpha and Delta 20 ng/mL recombinant spike protein (Figure 1).
The values of MO-Y, reflecting the cellular content of DNA and RNA, were found to be consistently decreased in most samples spiked with SARS-CoV-2 recombinant spike proteins, achieving statistical significance in those containing Alpha and Delta recombinant spike proteins at 20 ng/mL final concentration (Figure 2).
Unlike MO-Y, MO-WY which reflects the heterogeneity of monocyte content of DNA and RNA did not vary significantly between the two concentrations of the four different SARS-CoV-2 recombinant spike protein tested (Figure 2).
As shown in Figure 3, the values of MO-Z, mirroring the presence of monocytes of abnormal size, were found to be significantly decreased in all samples spiked with SARS-CoV-2 recombinant spike proteins (except in that spiked with 2 ng/mL ancestral variant).
The values of MO-WZ, reflecting the heterogeneity of monocyte volumes were found to be increased in all samples, achieving statistical significance in those where the SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha and Delta variants were added at the higher 20 ng/mL final concentration (Figure 3).
The revision of the blood smear performed with the DI-60 digital morphology analyzer confirmed the presence of a broad spectrum of morphological monocyte abnormalities, as shown in Figure 4, which represents some monocytes in the whole sample where the SARS-CoV-2 recombinant spike protein of the Delta variant was added at the final concentration of 20 ng/mL.
Briefly, the cells were found to be largely dysmorphic, granulated, intensely vacuolated, with aberrant nucleus and clear alterations of chromatin structure, as well as presenting cytoplasmic extrusions. Platelet phagocytosis could also be clearly observed.
4. Discussion
The results of our experimental study, where human monocytes where challenged with different types (Ancestral, Alpha, Delta and Omicron) and concentrations (2 and 20 ng/mL) of SARS-CoV-2 recombinant spike protein reveal that the morphology of these cells is dramatically perturbed upon incubation. More precisely, we found an increased monocyte complexity (i.e., MO-X), which reflects the presence of granules, vacuoles and other cytoplasmic inclusions, whilst the monocyte content of DNA and RNA (MO-Y) as well as their size (MO-Z) appeared to be consistently reduced. These changes were also mirrored by a trend towards increased heterogeneity of monocyte complexity and size (MO-WX and MO-WZ, respectively), whilst the heterogeneity of DNA and RNA varied less markedly. Notably, these changes appeared to be consistently dose-dependent, in that the changes become more prominent in samples where the higher (20 ng/mL) concentration of SARS-CoV-2 recombinant spike protein was added, and in specimens spiked especially with the Alpha and Delta variants. Importantly, most of these abnormalities found a reliable reflection in the digital images, where we could see that samples with most evident variations of monocyte parameters displayed clear alterations of monocyte morphology, namely the presence of dysmorphia, increased content of granules and vacuoles, nucleus abnormalities reflecting a considerable alteration of chromatin structure and even platelet phagocytosis. All these findings provide convincing support to different biological and epidemiological information previously published and reviewed in the following parts of this article.
The first important aspect that emerged from our study is that the recombinant spike protein of SARS-CoV-2 is capable of strongly disrupting monocyte structure, thus supporting earlier evidence of such intriguing interaction between the cell and the virus. Resting monocytes harbor an abundant concentration of ACE2 and transmembrane serine protease 2 (TMPRSS2) in their cytoplasm, which upon stimulation, can be rapidly transported to the cell surface, where these two proteins are used by SARS-CoV-2 for binding to and fusing with the cell through its spike protein [15]. Virus-cell interaction may also be mediated by the myeloid cell specific I-type lectin CD169, to which the spike protein can efficiently bind [16].
In the seminal study published by Ait-Belkacem et al. [11], the authors found that incubation of SARS-CoV-2 spike protein with human monocytes was capable of triggering a substantial release of inflammatory biomarkers. Important evidence of monocyte-macrophage lineage disruption has been reported in the interesting study published by Cao et al. [17], where the authors found that spike protein-pseudotyped lentiviruses were capable to bind to these cell lines, triggering a sustained pro-inflammatory cytokines production in the lung, which could be abolished by deletion of the sequence of the receptor binding domain (RBD) from the spike protein. In a subsequent study, other authors found that the spike protein could per se promote monocyte activation to an extent similar to that seen using other potent monocyte activators such as lipopolysaccharide [18]. Interestingly, an enhanced content on lysosomal ionized calcium (Ca2+) was also observed, which is an important marker of cell death processes including apoptosis and autophagy [19,20]. A similar effect (i.e., monocyte apoptosis) was observed in a separate experiment by Chandrasekar et al., who challenged human monocytes isolated from the blood of ostensibly healthy individuals with SARS-CoV-2 recombinant spike protein [21], and reported that such incubation with spike protein triggered Caspase 3 activation and subsequent cell death.
The concept that the SARS-CoV-2 spike protein alone could be capable to bind and alter monocyte biology has been convincingly confirmed in a series of experiments conducted by Schroeder et al. [22]. In brief, the authors immobilized different spike protein components within microtiter wells, showing that its S1 subunit effectively bound and activated human monocytes, generating a proinflammatory state characterized by a pattern of cytokine release virtually identical to that seen in patients with COVID-19-induced cytokine storm. In a subsequent study, Barhoumi et al. challenged THP-1 cells (i.e., derived from a human monocytic cell line) with SARS-CoV-2 recombinant spike protein [23], and found significant induction of apoptosis, with cell activity considerably dysregulated towards production of reactive oxygen species (ROS). A similar effect was noted using peripheral blood mononuclear cells. Notably, all such apoptotic effects triggered by the SARS-CoV-2 spike protein in other studies are thoughtfully mirrored by our experimental findings. In fact, we also found that adding SARS-CoV-2 recombinant spike protein to human blood samples caused a significant reduction of monocytes fluorescence intensity (a reduced MO-Y indicates a lower cellular amount of DNA and RNA, characterizing apoptotic cells) (Figure 3) and the appearance of dysmorphic and intensely vacuolated monocytes in the peripheral blood smear (Figure 4). Both these events reflect a degenerative process, which is a predictable anticipation of cell death. Accordingly, a recent meta-analysis has concluded that the monocyte count was significantly lower in COVID-19 patients with severe illness compared to those with milder symptoms (Standardized Mean Difference: -0.56; 95%CI, from -0.8 to -0.32; p<0.001) [24], which is in keeping with our findings that monocytes may be seriously injured after being infected by SARS-CoV-2 or even by interacting with its spike protein alone.
As concerns the morphological abnormalities triggered by monocyte-spike protein interaction that were observed in our study, these changes seem to have similar counterparts ex vivo. An interesting study published by Utrero-Rico et al. assayed circulating monocytes upon emergency department admission in 131 patients with acute SARS-CoV-2 infection [25], reporting the presence of characteristic dysmorphia and intense granularity, with enhanced propensity to release pro-inflammatory cytokines. Volumetric monocyte alternations have been clearly seen ex vivo in the study of Kubánková et al. [26], who reported a significant change in monocyte size of patients with active COVID-19 compared to a healthy control population. In keeping with our observations, the authors also found an increased SD of the cross-sectional monocyte area in patients with COVID-19, which is reflected by the enhanced MO-WZ value that we found in samples where SARS-CoV-2 recombinant spike proteins (except that of the Omicron variant) was added at the final concentration of 20 ng/mL, clearly reflecting a broader heterogeneity of monocyte volumes. In another study, Zhang et al. used flow cytometry for studying peripheral blood samples collected from 34 patients with COVID-19 [27]. These authors observed substantial morphologic and functional differences in monocytes of COVID-19 patients, which appeared even more evident in those with severe SARS-CoV-2 infection needing prolonged hospitalization and/or intensive care. These authors also reported a remarkably increased number of atypical and vacuolated monocytes in COVID-19 patients, which could not be seen in the blood of healthy individuals. Exceptionally, the ratio between these atypical to typical monocytes in blood was comprised between 60-100% in COVID-19 patients compared to 0% in healthy control blood, respectively. It is hence not surprising that we found a high degree of heterogeneity of monocyte volumes in our experiments (i.e., the higher MO-WZ) which could be reflected by an increased value of the monocyte distribution width (MDW; i.e., the relative change in the volume of circulating monocytes). Recent studies and meta-analyses concluded that the MDW is not only increased in patients with SARS-CoV-2 infection compared to those without [28], but is also a significant predictor of unfavorable clinical progression in those with COVID-19 [29,30,31,32]. Importantly, platelet phagocytosis could also be observed especially in specimens challenged with 20 ng/mL of Alpha and Delta recombinant spike protein (Figure 4), a pathological process that has been previously described in the blood smears of patients with SARS-CoV-2 infection and reflecting intense monocyte activation and haemophagocytic lymphohistiocytosis [33], a frequent condition seen in patients with COVID-19 [34].
Another important aspect emerged from our experimental study is that the alteration of monocyte structure seen in human blood samples challenged with different SARS-CoV-2 recombinant spike proteins were substantially dependent upon concentration and type of the recombinant protein. Succinctly, we typically found larger alterations in samples challenged with 20 ng/mL of the recombinant protein as compared to those where the lower 2 ng/mL concentration was added, as well as in those especially challenged with SARS-CoV-2 recombinant spike protein of the Alpha and Delta variants. Thus, it is conceivable that the sequence and structure of these two spike protein variants may display better affinity for some receptors expressed at monocyte surface and/or may be capable to more intensely trigger monocyte activation and injury compared to the Omicron spike protein, as was earlier conjectured [35]. The lower chemical stability of the Omicron spike protein compared to the spike protein of former variants the has also been suggested as an important characteristic that may explain the lower pathogenetic burden of this strain [36].
Notably, the more accentuated reduction of monocyte fluorescence (i.e., the more markedly reduced MO-Y) observed after challenging human whole blood with the higher concentration of the recombinant spike protein of the Alpha and Delta variants is theoretically consistent with a more intense disruption of cell integrity. On the other hand, the fact that all monocyte parameters were less affected in whole blood challenged with the recombinant spike protein of the Omicron variant seemingly reflect the clinical evidence that the clinical spectrum of disease after infection with the sub-lineages belonging to the Omicron family is substantially milder compared to that caused by the previous strains. Several epidemiological studies, recently meta-analyzed by Hu et al. (33 studies, including over 6 million patients with COVID-19) [37] have convincingly shown that Omicron infection was associated with a 2.9-fold lower risk of hospitalization, 3.6-fold lower risk of ICU admission and a cumulatively 3.0-fold lower risk of death compared to the Delta variant in the general population. Esper and colleagues correlated COVID-19 disease severity and outcomes throughout the periods of major variants dominance [38], reporting that the clinical burden (as defined in terms of hospitalization, ICU admission and mortality) was similar between the Alpha and Delta waves, but in both cases was higher than that seen after the emergence of the Omicron variant. A comparative analysis of clinical severity of COVID-19 cases sustained by the Alpha, Delta and Omicron SARS-CoV-2 variants has also been conducted by Varea-Jiménez et al. [39], who concluded that infections sustained by the Alpha and especially Delta variants were associated with higher clinical severity compared to Omicron infections, irrespective of the vaccination status. Even more interestingly, Korobova et al. assayed the concentration of multiple cytokines in COVID-19 patients infected by the four major strains (i.e., Ancestral, Alpha, Delta and Omicron) [40] and reported that only four of these mediators displayed were constantly increased irrespective of the genetic variant. These authors also reported that the so-called cytokine storm and the general “hypercytokinemia” exhibited a marked attenuation after emergence of the new Omicron variant. Interestingly, these findings were confirmed in a subsequent study published by Barh et al. [41], who reported that pro-inflammatory cytokines production (especially interleukin 6) is decreased in patients infected with the Omicron compared to those infected by Delta variant. In keeping with these findings, Park et al. studied nearly 30000 veterans with Alpha, Delta and Omicron infections [42] and observed that the risk of having enhanced levels of C Reactive protein was higher in those infected by Delta (Odds Ratio [OR], 2.30; 95%CI, 2.18-2.4) or Alpha (OR, 2.29; 95%CI, 2.14-2.45) compared to Omicron. A similar trend was seen by measuring ferritin (i.e., another well-known inflammatory biomarker), in that the likelihood of having increased concentrations of this protein was higher in COVID-19 patients infected by Delta (OR, 1.91; 95%CI, 1.81-2.02) or Alpha (OR, 1.54; 95%CI, 1.45-1.65) compared to Omicron. Finally, International Kawasaki Disease Registry reported that the clinical burden of the multisystem inflammatory syndrome in children (MIS-C), i.e., a delayed hyperinflammatory response to SARS-CoV-2 infection in children, was found to be considerably lower during the omicron period compared to the previous Ancestral, Alpha and Delta waves [43].
In summary, the previous described clinical evidence are strongly congruent with our findings, since disruption of classical monocyte biology seems highest after challenge with recombinant spike protein of Alpha and Delta variants, intermediate with that of the Ancestral strain, and the lowest with the recombinant spike protein of the new Omicron sublineages. Nonetheless, the still abnormal value of some monocyte parameters (especially MO-Z, i.e., reflecting an increased presence of monocytes of abnormal size) observed when we added the recombinant spike protein of the Omicron variant would suggest that the pandemic may not be over, at least clinically, as monocyte biology and structure may still be disrupted by the descendants of this most recently emerged variant.
5. Conclusions
In this study, we demonstrated that the spike protein of SARS-CoV-2 is capable to trigger per se morphological abnormalities in monocytes. For the first time, we demonstrated that disruption of monocyte structure becomes more evident when these cells are challenged with the recombinant spike proteins of the more clinically severe Alpha and Delta variants, which may hence explain the attenuated pathogenetic and clinical burden seen after emergence of the Omicron lineages.
Author Contributions
Conceptualization, G.L.S. and G.L.; methodology, G.L.; formal analysis, G.L.; investigation, M.V., F.D., G.C.. M.G. and G.C.; data curation, G.L.; writing—original draft preparation, G.L.; writing—review and editing, B.M.H.; funding acquisition, G.L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of the Provinces of Verona and Rovigo (2622CESC).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Complete data can be available upon reasonable request to the corresponding author.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sampath, S.; Khedr, A.; Qamar, S.; Tekin, A.; Singh, R.; Green, R.; Kashyap, R. Pandemics Throughout the History. Cureus 2021, 13, e18136. [Google Scholar] [CrossRef] [PubMed]
- McVernon, J.; Liberman, J. WHO keeps covid-19 a public health emergency of international concern. BMJ 2023, 380, 504. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.P.; Liu, S.L. Role of host factors in SARS-CoV-2 entry. J Biol Chem. 2021, 297, 100847. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Zhang, M.; Chang, T.L. ACE2-Independent Alternative Receptors for SARS-CoV-2. Viruses 2022, 14, 2535. [Google Scholar] [CrossRef]
- Nocini, R.; Henry, B.M.; Mattiuzzi, C.; Lippi, G. Improving Nasal Protection for Preventing SARS-CoV-2 Infection. Biomedicines 2022, 10, 2966. [Google Scholar] [CrossRef]
- Ligi, D.; Lo Sasso, B.; Henry, B.M.; Ciaccio, M.; Lippi, G.; Plebani, M.; Mannello, F. Deciphering the role of monocyte and monocyte distribution width (MDW) in COVID-19: An updated systematic review and meta-analysis. Clin Chem Lab Med. 2023, 61, 960–973. [Google Scholar] [CrossRef]
- Grant, R.A.; Morales-Nebreda, L.; Markov, N.S.; Swaminathan, S.; Querrey, M.; Guzman, E.R.; Abbott, D.A.; Donnelly, H.K.; Donayre, A.; Goldberg, I.A.; et al. Circuits between infected macrophages and T cells in SARS-CoV-2 pneumonia. Nature 2021, 590, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Plebani, M. Cytokine "storm", cytokine "breeze", or both in COVID-19? Clin Chem Lab Med. 2020, 59, 637–639. [Google Scholar] [CrossRef]
- Park, J.; Dean, L.S.; Jiyarom, B.; Gangcuangco, L.M.; Shah, P.; Awamura, T.; Ching, L.L.; Nerurkar, V.R.; Chow, D.C.; Igno, F.; et al. Elevated circulating monocytes and monocyte activation in COVID-19 convalescent individuals. Front Immunol. 2023, 14, 1151780. [Google Scholar] [CrossRef]
- Kilercik, M.; Demirelce, Ö.; Serdar, M.A.; Mikailova, P.; Serteser, M. A new haematocytometric index: Predicting severity and mortality risk value in COVID-19 patients. PLoS ONE 2021, 16, e0254073. [Google Scholar] [CrossRef]
- Ait-Belkacem, I.; Cartagena García, C.; Millet-Wallisky, E.; Izquierdo, N.; Loosveld, M.; Arnoux, I.; Morange, P.E.; Galland, F.; Lambert, N.; Malergue, F.; et al. SARS-CoV-2 spike protein induces a differential monocyte activation that may contribute to age bias in COVID-19 severity. Sci Rep. 2022, 12, 20824. [Google Scholar] [CrossRef] [PubMed]
- Craddock, V.; Mahajan, A.; Spikes, L.; Krishnamachary, B.; Ram, A.K.; Kumar, A.; Chen, L.; Chalise, P.; Dhillon, N.K. Persistent circulation of soluble and extracellular vesicle-linked Spike protein in individuals with postacute sequelae of COVID-19. J Med Virol. 2023, 95, e28568. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Salvagno, G.L.; Montagnana, M.; Lima-Oliveira, G.; Guidi, G.C.; Favaloro, E.J. Quality standards for sample collection in coagulation testing. Semin Thromb Hemost. 2012, 38, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Buoro, S.; Seghezzi, M.; Vavassori, M.; Dominoni, P.; Apassiti Esposito, S.; Manenti, B.; Mecca, T.; Marchesi, G.; Castellucci, E.; Azzarà, G.; et al. Clinical significance of cell population data (CPD) on Sysmex XN-9000 in septic patients with our without liver impairment. Ann Transl Med. 2016, 4, 418. [Google Scholar] [CrossRef]
- Yao, Y.; Subedi, K.; Liu, T.; Khalasawi, N.; Pretto-Kernahan, C.D.; Wotring, J.W.; Wang, J.; Yin, C.; Jiang, A.; Fu, C.; et al. Surface translocation of ACE2 and TMPRSS2 upon TLR4/7/8 activation is required for SARS-CoV-2 infection in circulating monocytes. Cell Discov. 2022, 8, 89. [Google Scholar] [CrossRef]
- Jalloh, S.; Olejnik, J.; Berrigan, J.; Nisa, A.; Suder, E.L.; Akiyama, H.; Lei, M.; Ramaswamy, S.; Tyagi, S.; Bushkin, Y.; et al. CD169-mediated restrictive SARS-CoV-2 infection of macrophages induces pro-inflammatory responses. PLoS Pathog. 2022, 18, e1010479. [Google Scholar] [CrossRef]
- Cao, X.; Tian, Y.; Nguyen, V.; Zhang, Y.; Gao, C.; Yin, R.; Carver, W.; Fan, D.; Albrecht, H.; Cui, T.; et al. Spike protein of SARS-CoV-2 activates macrophages and contributes to induction of acute lung inflammation in male mice. FASEB J. 2021, 35, e21801. [Google Scholar] [CrossRef]
- Palestra, F.; Poto, R.; Ciardi, R.; Opromolla, G.; Secondo, A.; Tedeschi, V.; Ferrara, A.L.; Di Crescenzo, R.M.; Galdiero, M.R.; Cristinziano, L.; et al. SARS-CoV-2 Spike Protein Activates Human Lung Macrophages. Int J Mol Sci. 2023, 24, 3036. [Google Scholar] [CrossRef]
- Sukumaran, P.; Nascimento Da Conceicao, V.; Sun, Y.; Ahamad, N.; Saraiva, L.R.; Selvaraj, S.; Singh, B.B. Calcium Signaling Regulates Autophagy and Apoptosis. Cells 2021, 10, 2125. [Google Scholar] [CrossRef]
- Medina, D.L.; Di Paola, S.; Peluso, I.; Armani, A.; De Stefani, D.; Venditti, R.; Montefusco, S.; Scotto-Rosato, A.; Prezioso, C.; Forrester, A.; et al. Lysosomal calcium signalling regulates autophagy through calcineurin and TFEB. Nat Cell Biol. 2015, 17, 288–299. [Google Scholar] [CrossRef]
- Chandrasekar, A.P.; Maynes, M.; Natesampillai, S.; Shweta, F.N.U.; Badley, A.D.; Cummins, N.W. SARS-CoV-2 spike protein induces monocyte apoptosis and interleukin-8 production. Top Antivir Med 2021, 29, 61. [Google Scholar]
- Schroeder, J.T.; Bieneman, A.P. The S1 Subunit of the SARS-CoV-2 Spike Protein Activates Human Monocytes to Produce Cytokines Linked to COVID-19: Relevance to Galectin-3. Front Immunol. 2022, 13, 831763. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, T.; Alghanem, B.; Shaibah, H.; Mansour, F.A.; Alamri, H.S.; Akiel, M.A.; Alroqi, F.; Boudjelal, M. SARS-CoV-2 Coronavirus Spike Protein-Induced Apoptosis, Inflammatory, and Oxidative Stress Responses in THP-1-Like-Macrophages: Potential Role of Angiotensin-Converting Enzyme Inhibitor (Perindopril). Front Immunol. 2021, 12, 728896. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Jing, X.; Liu, Y.; Wen, R.; Wang, C. Comparison of laboratory parameters in mild vs. severe cases and died vs. survived patients with COVID-19: Systematic review and meta-analysis. J Thorac Dis. 2022, 14, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Utrero-Rico, A.; González-Cuadrado, C.; Chivite-Lacaba, M.; Cabrera-Marante, O.; Laguna-Goya, R.; Almendro-Vazquez, P.; Díaz-Pedroche, C.; Ruiz-Ruigómez, M.; Lalueza, A.; Folgueira, M.D.; et al. Alterations in Circulating Monocytes Predict COVID-19 Severity and Include Chromatin Modifications Still Detectable Six Months after Recovery. Biomedicines 2021, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Kubánková, M.; Hohberger, B.; Hoffmanns, J.; Fürst, J.; Herrmann, M.; Guck, J.; Kräter, M. Physical phenotype of blood cells is altered in COVID-19. Biophys J. 2021, 120, 2838–2847. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Guo, R.; Lei, L.; Liu, H.; Wang, Y.; Wang, Y.; Qian, H.; Dai, T.; Zhang, T.; Lai, Y.; et al. Frontline Science: COVID-19 infection induces readily detectable morphologic and inflammation-related phenotypic changes in peripheral blood monocytes. J Leukoc Biol. 2021, 109, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Malinovska, A.; Hernried, B.; Lin, A.; Badaki-Makun, O.; Fenstermacher, K.; Ervin, A.M.; Ehrhardt, S.; Levin, S.; Hinson, J.S. Monocyte Distribution Width as a Diagnostic Marker for Infection: A Systematic Review and Meta-analysis. Chest, 2023; Epub ahead of print. [Google Scholar] [CrossRef]
- Bajić, D.; Matijašević, J.; Andrijević, L.; Zarić, B.; Lalić-Popović, M.; Andrijević, I.; Todorović, N.; Mihajlović, A.; Tapavički, B.; Ostojić, J. Prognostic Role of Monocyte Distribution Width, CRP, Procalcitonin and Lactate as Sepsis Biomarkers in Critically Ill COVID-19 Patients. J Clin Med. 2023, 12, 1197. [Google Scholar] [CrossRef]
- Lorubbio, M.; Tacconi, D.; Iannelli, G.; Feri, M.; Scala, R.; Montemerani, S.; Mandò, M.; Ognibene, A. The role of Monocyte Distribution Width (MDW) in the prognosis and monitoring of COVID-19 patients. Clin Biochem. 2022, 103, 29–31. [Google Scholar] [CrossRef]
- Riva, G.; Castellano, S.; Nasillo, V.; Ottomano, A.M.; Bergonzini, G.; Paolini, A.; Lusenti, B.; Milić, J.; De Biasi, S.; Gibellini, L.; et al. Monocyte Distribution Width (MDW) as novel inflammatory marker with prognostic significance in COVID-19 patients. Sci Rep. 2021, 11, 12716. [Google Scholar] [CrossRef]
- Lippi, G.; Sanchis-Gomar, F.; Henry, B.M. Pooled analysis of monocyte distribution width in subjects with SARS-CoV-2 infection. Int J Lab Hematol. 2021, 43, O161–O163. [Google Scholar] [CrossRef] [PubMed]
- Khedmati, M.E.; Sharifi, M.J. Platelet phagocytosis by monocytes. Clin Chem Lab Med. 2022, 60, e204–e206. [Google Scholar] [CrossRef] [PubMed]
- Fadlallah, M.M.; Salman, S.M.; Fadlallah, M.M.; Rahal, H. Hemophagocytic Syndrome and COVID-19: A Comprehensive Review. Cureus 2023, 15, e36140. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, W.; Liu, Z.; Kang, Y.; Zhang, X.; Xu, Z.; Gao, Y.; Qi, Y. A comparative study of spike protein of SARS-CoV-2 and its variant Omicron (B.1.1.529) on some immune characteristics. Sci Rep. 2022, 12, 17058. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Geng, B.; Marcon, E.; Pu, S.; Tang, H.; Merluza, J.; Bello, A.; Snider, J.; Lu, P.; Wood, H.; et al. Omicron Spike Protein Is Vulnerable to Reduction. J Mol Biol. 2023; Epub ahead of print. [Google Scholar] [CrossRef]
- Hu, F.H.; Jia, Y.J.; Zhao, D.Y.; Fu, X.L.; Zhang, W.Q.; Tang, W.; Hu, S.Q.; Wu, H.; Ge, M.W.; Du, W.; et al. Clinical outcomes of the severe acute respiratory syndrome coronavirus 2 Omicron and Delta variant: Systematic review and meta-analysis of 33 studies covering 6 037 144 coronavirus disease 2019-positive patients. Clin Microbiol Infect. 2023; Epub ahead of print. [Google Scholar] [CrossRef]
- Esper, F.P.; Adhikari, T.M.; Tu, Z.J.; Cheng, Y.W.; El-Haddad, K.; Farkas, D.H.; Bosler, D.; Rhoads, D.; Procop, G.W.; Ko, J.S.; et al. Alpha to Omicron: Disease Severity and Clinical Outcomes of Major SARS-CoV-2 Variants. J Infect Dis. 2023, 227, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Varea-Jiménez, E.; Aznar Cano, E.; Vega-Piris, L.; Martínez Sánchez, E.V.; Mazagatos, C.; García San Miguel Rodríguez-Alarcón, L.; Casas, I.; Sierra Moros, M.J.; Iglesias-Caballero, M.; Vazquez-Morón, S.; et al. Comparative severity of COVID-19 cases caused by Alpha, Delta or Omicron SARS-CoV-2 variants and its association with vaccination. Enferm Infecc Microbiol Clin (Engl Ed). 2023; Epub ahead of print. [Google Scholar] [CrossRef]
- Korobova, Z.R.; Arsentieva, N.A.; Liubimova, N.E.; Batsunov, O.K.; Dedkov, V.G.; Gladkikh, A.S.; Sharova, A.A.; Adish, Z.; Chernykh, E.I.; Kaschenko, V.A.; et al. Cytokine Profiling in Different SARS-CoV-2 Genetic Variants. Int J Mol Sci. 2022, 23, 14146. [Google Scholar] [CrossRef]
- Barh, D.; Tiwari, S.; Rodrigues Gomes, L.G.; Ramalho Pinto, C.H.; Andrade, B.S.; Ahmad, S.; Aljabali, A.A.A.; Alzahrani, K.J.; Banjer, H.J.; Hassan, S.S.; et al. SARS-CoV-2 Variants Show a Gradual Declining Pathogenicity and Pro-Inflammatory Cytokine Stimulation, an Increasing Antigenic and Anti-Inflammatory Cytokine Induction, and Rising Structural Protein Instability: A Minimal Number Genome-Based Approach. Inflammation 2023, 46, 297–312. [Google Scholar] [CrossRef]
- Park, C.; Tavakoli-Tabasi, S.; Sharafkhaneh, A.; Seligman, B.J.; Hicken, B.; Amos, C.I.; Chou, A.; Razjouyan, J. Inflammatory Biomarkers Differ among Hospitalized Veterans Infected with Alpha, Delta, and Omicron SARS-CoV-2 Variants. Int J Environ Res Public Health 2023, 20, 2987. [Google Scholar] [CrossRef]
- McCrindle, B.W.; Harahsheh, A.S.; Handoko, R.; Raghuveer, G.; Portman, M.A.; Khoury, M.; Newburger, J.W.; Lee, S.; Jain, S.S.; Khare, M.; et al. SARS-CoV-2 Variants and Multisystem Inflammatory Syndrome in Children. N Engl J Med. 2023, 388, 1624–1626. [Google Scholar] [CrossRef]
Figure 1.
Values distribution of MO-X (monocyte complexity) and MO-WX (heterogeneity of monocyte complexity) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
Figure 1.
Values distribution of MO-X (monocyte complexity) and MO-WX (heterogeneity of monocyte complexity) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
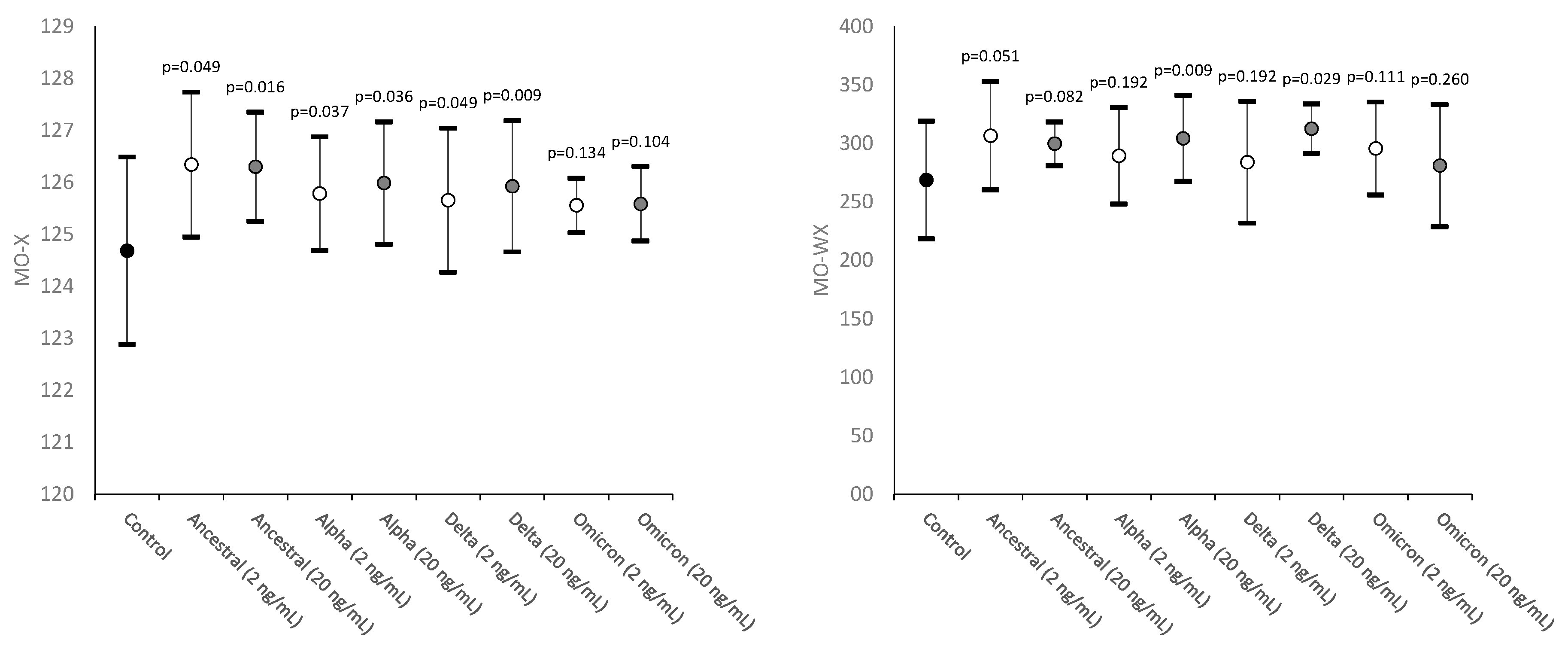
Figure 2.
Values distribution of MO-Y (monocytes fluorescence intensity) and MO-WY (heterogeneity of monocytes fluorescence intensity) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
Figure 2.
Values distribution of MO-Y (monocytes fluorescence intensity) and MO-WY (heterogeneity of monocytes fluorescence intensity) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
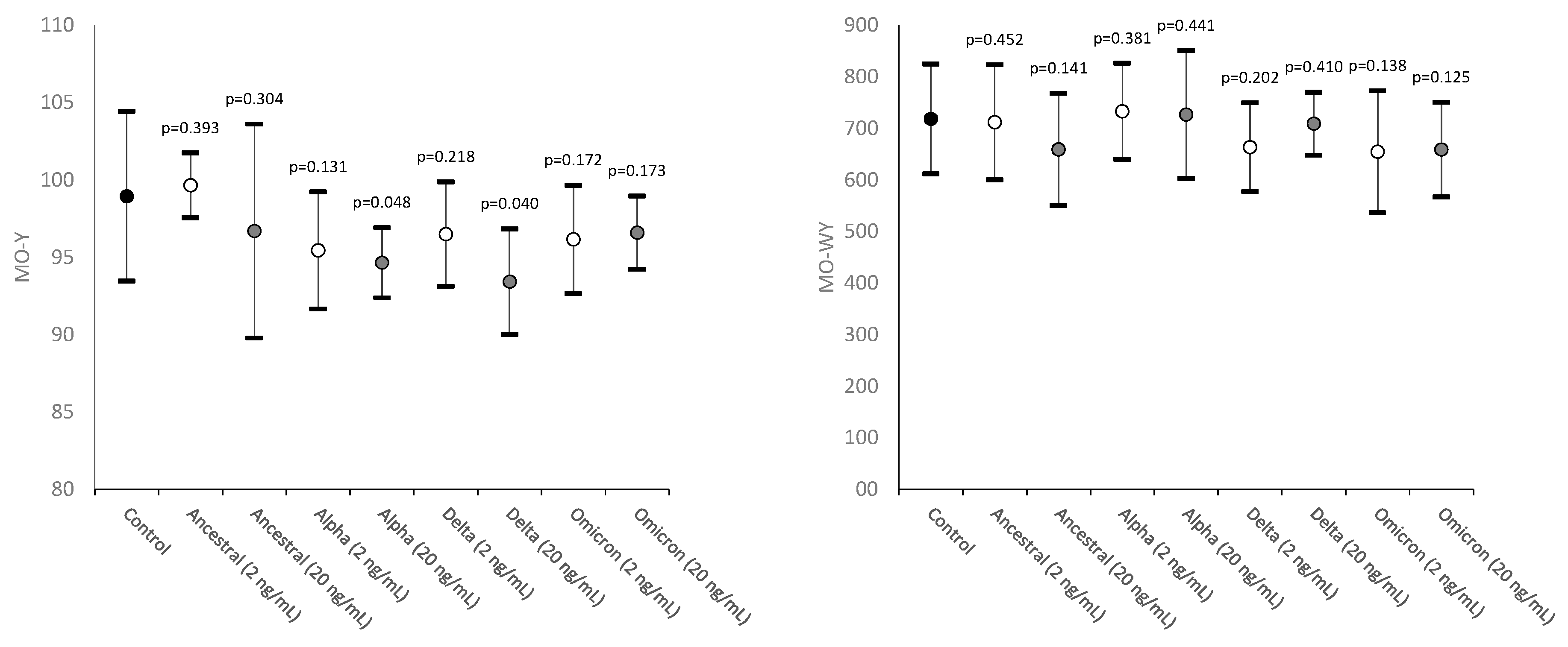
Figure 3.
Values distribution of MO-Z (monocytes cell size) and MO-WZ (heterogeneity of monocytes cell size) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
Figure 3.
Values distribution of MO-Z (monocytes cell size) and MO-WZ (heterogeneity of monocytes cell size) in human whole blood samples where saline (i.e., control sample) or SARS-CoV-2 recombinant spike proteins of Ancestral, Alpha, Delta and Omicron variants were added at a final concentration of 2 and 20 ng/mL. Values are given as mean and standard deviation (SD); the “p” value above the upper limit of the SD indicates the value of statistical significance compared to the control sample where only saline was added.
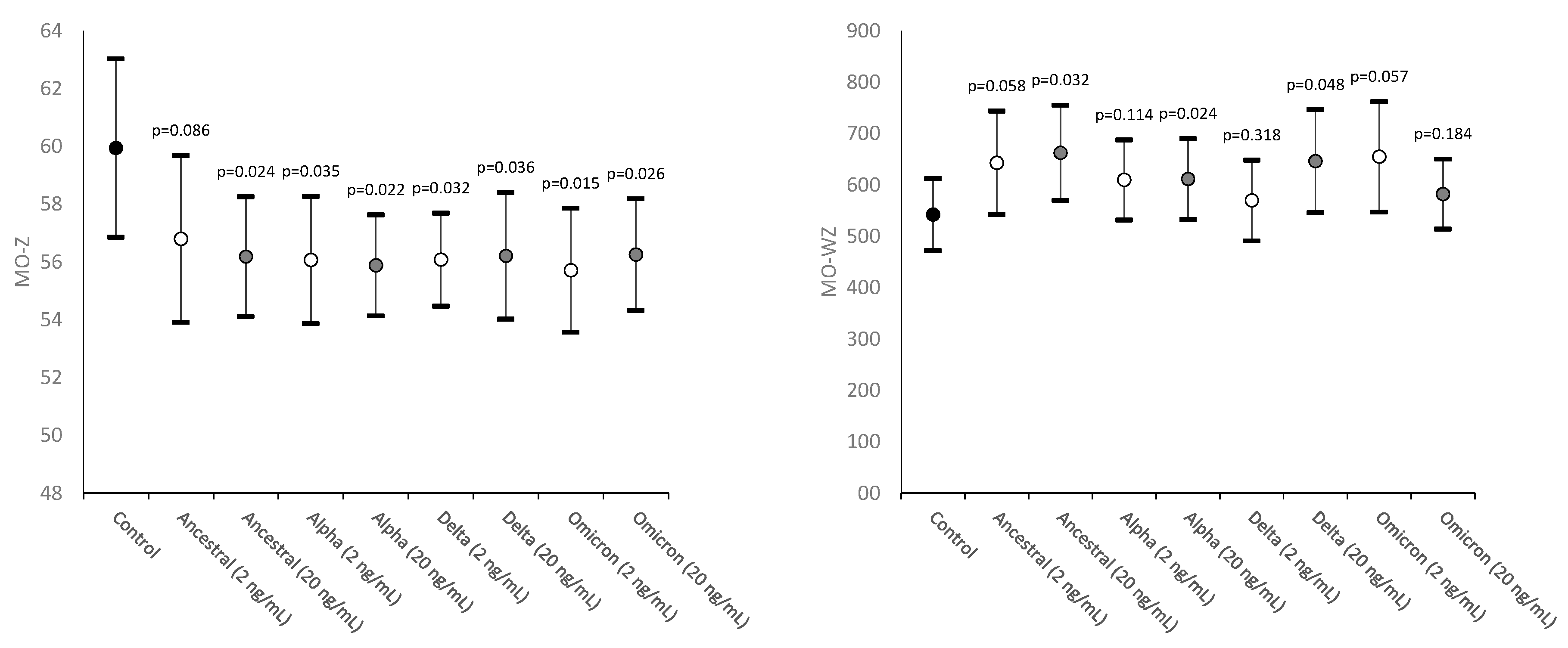
Figure 4.
Alteration of monocyte morphology in human whole blood sample incubated for 15 min with 20 ng/mL SARS-CoV-2 spike protein of the Delta variant. Typical distinguishing features encompass dysmorphia, granulation, intense vacuolization, presence of an aberrant nucleus and development of cytoplasmic extrusions. Platelet phagocytosis can also be clearly seen.
Figure 4.
Alteration of monocyte morphology in human whole blood sample incubated for 15 min with 20 ng/mL SARS-CoV-2 spike protein of the Delta variant. Typical distinguishing features encompass dysmorphia, granulation, intense vacuolization, presence of an aberrant nucleus and development of cytoplasmic extrusions. Platelet phagocytosis can also be clearly seen.
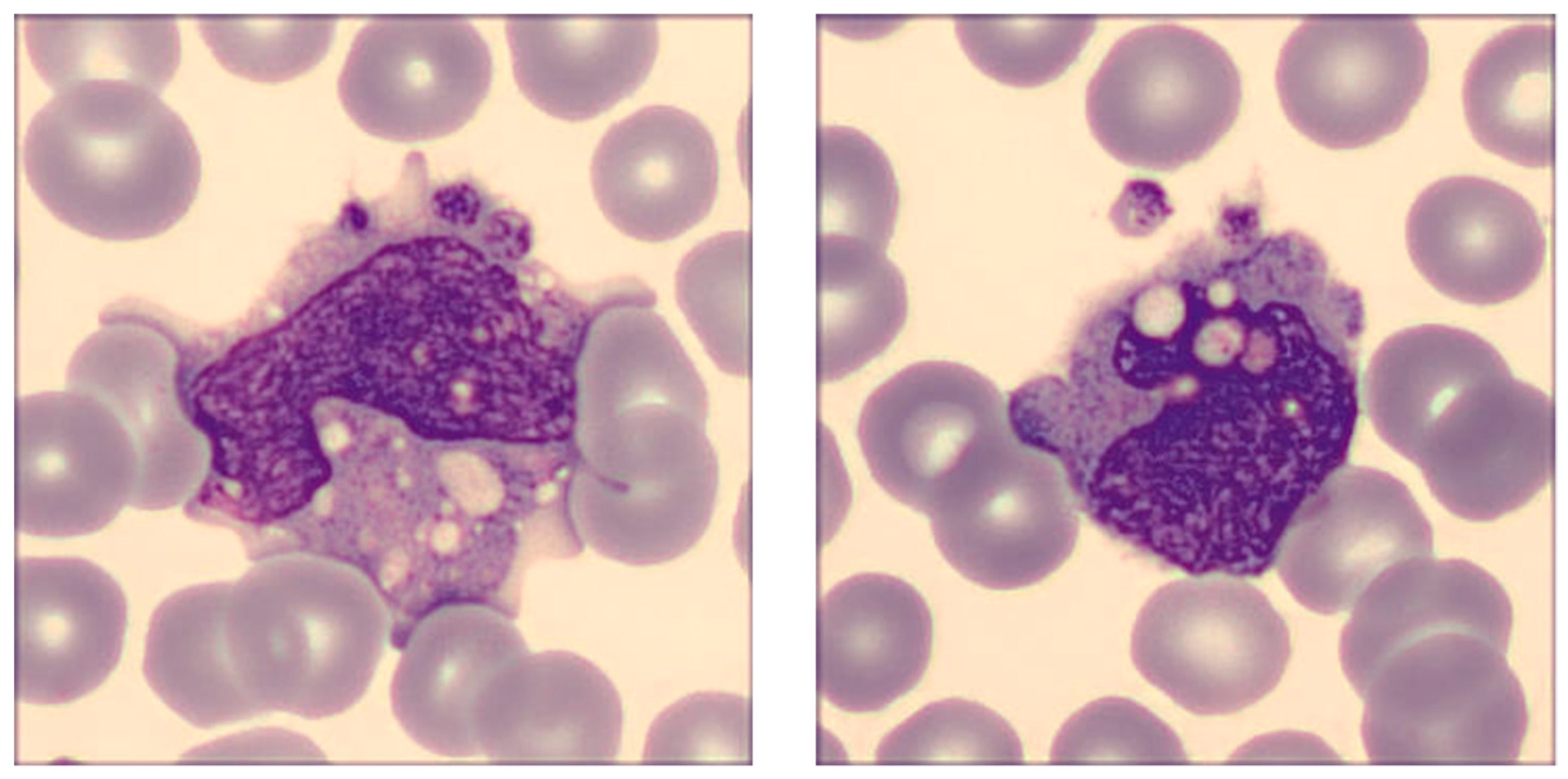

Table 1.
SARS-CoV-2 recombinant spike proteins used for the experiments, as purchased in lyophilized stable and certified form from ACROBiosystems (Newark, DE, USA).
Table 1.
SARS-CoV-2 recombinant spike proteins used for the experiments, as purchased in lyophilized stable and certified form from ACROBiosystems (Newark, DE, USA).
| Spike protein variants | Additional mutations |
|---|---|
| Ancestral | D614G |
| Alpha (B.1.1.7) | AAVal16, Pro1213, HV69-70del, Y144del, N501Y, A570D, D614G, P681H, T716I, S982A, D1118H |
| Delta (B.1.617.2) | AAVal16, Pro 1213, T19R, G142D, EF156-157del, R158G, L452R, T478K, D614G, P681R, D950N |
| Omicron (B.1.1.529) | AAVal16, Pro 1213, A67V, HV69-70del, T95I, G142D, VYY143-145del, N211del, L212I, ins214EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F |
Table 2.
Description of the monocyte parameters obtained using the Sysmex XN hematological analyzer (Sysmex Corporation, Kobe, Japan).
Table 2.
Description of the monocyte parameters obtained using the Sysmex XN hematological analyzer (Sysmex Corporation, Kobe, Japan).
| Parameter | Abbreviation | Clinical significance |
|---|---|---|
| Monocytes complexity | MO-X | Presence of granules, vacuoles and other cytoplasmic inclusions |
| Width of dispersion of monocytes complexity | MO-WX | Heterogeneity of monocytes complexity |
| Monocytes fluorescence intensity | MO-Y | Quantity of cellular DNA and RNA |
| Width of dispersion of monocytes fluorescence intensity | MO-WY | Heterogeneity of monocytes fluorescence intensity |
| Monocytes cell size | MO-Z | Abnormal sized cells |
| Width of dispersion of monocytes cell size | MO-WZ | Heterogeneity of monocyte cell size |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



