
Submitted:
29 April 2023
Posted:
30 April 2023
You are already at the latest version
Abstract
Oxidative stress has been implicated in cellular senescence and aging, as well as in the onset and progression of many diverse genetic and acquired diseases and conditions. The search for biomarkers of oxidative stress has become relevant to many biomedical fields using many different methods and approaches. Fluorescence methodology is advantageous because of its simplicity and high sensitivity. Fluorescent probes are frequently used to investigate the role of reactive oxygen species in oxidative stress in experimental or clinical settings, often involving the use of flow cytometry and related single-cell based technologies. To assess the specific role of reactive oxygen species in oxidative stress studies by cytometric methodologies, it is essential to detect and characterize these species accurately. However, the detection and quantitation of individual intracellular ROS is still a challenge, but different and complementary strategies may focus on other endpoints of oxidative stress. In this review we present and discuss briefly the limitations and perspectives of such approaches. Based on our results, we also provide recommendations for proper design of cytometric studies of oxidative stress in order to prevent or minimize experimental errors..
Keywords:
Fluorescence
; Image Cytometry
; Cytomics
; Reactive Oxygen Species
; Antioxidants
; Oxydative Stress
; In Vitro Toxicology
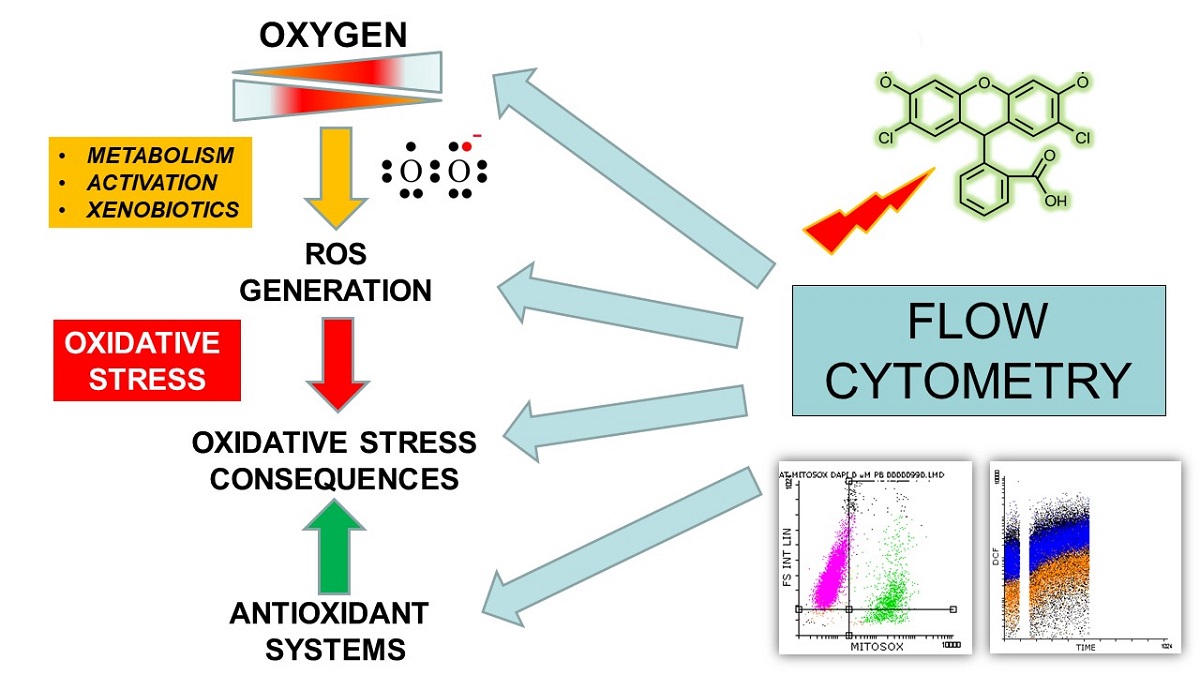
1. Introduction: Oxygen, ROS and Oxidative Stress
The evolution of Life on Earth has created organisms that need oxygen to live. Most living beings depend on oxygen to generate metabolic energy from the oxidation of biomolecules [1]. Paradoxically, the oxygen functions essential to living things depend on a chemical property dangerous to them: the structure of the oxygen molecule (O2) has two unpaired electrons, and O2 can accept individual electrons to generate unstable and highly reactive molecular forms, known as reactive oxygen species (ROS) [2].
The term ROS may be applied to a variety of molecules not only derived from O2 and includes both free radicals and species derived from free radicals [3]. O2 itself is a free radical, albeit with low reactivity [2]. More active, biologically relevant ROS derived from metabolic or abiotic reactions of O2 include singlet oxygen (1O2), superoxide anion radical (O2.-), hydrogen peroxide (H2O2) and hydroxyl radical (OH.) [1,2].
1O2 is an excited and much more reactive form of the O2 molecule, that can arise by photochemical reactions or by abiotic stress [4]. The pathway of univalent O2 reduction, as it happens along the mitochondrial respiratory chain generates O2.-, a relatively unreactive species that can interact with different molecules to generate H2O2 and other ROS through enzyme- or metal-catalysed processes [2,5]. H2O2 is not a free radical, as it lacks unpaired electrons, but diffuses easily through membranes and has relatively long half-life [5], allowing H2O2 to undergo metal-catalysed reactions that yield the OH. radical, one of the strongest free radicals that reacts readily with cellular components, including DNA, proteins, lipids and carbohydrates, thus being one of the most potentially damaging ROS [2,5].
There are also nitrogen-containing free radicals and reactive molecules, the Reactive Nitrogen Species (RNS), including nitric oxide (NO) and peroxynitrite (ONOO) [5,6]. Because RNS also contain oxygen and their generation is connected to ROS generation, ROS and RNS have been grouped together under the term RONS [7].
RONS have been implicated in cellular senescence and aging [8], as well as in the onset and progression of genetic [9,10] and acquired conditions such as inflammatory diseases [11,12,13,14], cardiovascular diseases [15,16,17,18], thrombosis [19], cancer [20,21,22,23] and anticancer chemotherapy [24], HIV-progression [25,26], neurodegenerative diseases [27,28,29] and metabolic disorders [30]. However, RONS participate in multiple intercellular and intracellular signaling pathways [31,32,33,34] and in cell-function modifying processes involved both in the destruction of invading pathogens [35] and in the fine tuning of cellular adaptation to endogenous and exogenous stress [36,37,38].
The physiological levels of ROS are maintained by a complex antioxidant machinery. However, when the capacity of these protective mechanisms is overcome by the intensity or duration of oxidative processes, a situation called oxidative stress occurs, which is defined as an alteration in the equilibrium between ROS production and antioxidant defenses, producing oxidative damage [39,40]. Oxidative stress can result from two separate, but not excluding, processes. On the one hand, the decrease in the levels or the activity of enzymes of the antioxidant defense by mutation or destruction of the active center, induced by the ROS themselves [40]. Deficiencies in the dietary supply of soluble antioxidants can also cause oxidative stress. On the other hand, increased production of ROS, exposure of cells or organisms to elevated levels of exogenous ROS or their metabolic precursors, and even excessive induction of ROS-mediated protective processes, such as phagocytosis or xenobiotic biotransformation can lead to the situation of oxidative stress [2,5].
The active search for biomarkers of oxidative stress has become relevant to many biomedical fields [41], and many different methods are applied to assess the redox state of the body or of specific tissues and cells [42,43,44].
Fluorescence methodology is advantageous because of its simplicity and high sensitivity and spatial resolution in imaging techniques [45,46]. Fluorescent probes are frequently used to investigate the role of ROS in oxidative stress in experimental or clinical settings [45,46], often involving the use of flow cytometry (FCM) [47,48] and related single-cell based technologies [49,50].
As schematically shown in Figure 1 and Figure 2, typically, the fluorescent probes are non fluorescent until being oxidised by intracellular oxidants and they are incorporated in form of fluorogenic substrates (Figure 1) which have been modified by appropriate chemical design to become both cell-permeable and susceptible to ROS-mediated oxidation (Figure 2) [51,52,53]. However, fluorescence-based detection of ROS is a complex task due to the low concentration, short half-life, and extensive interactions of ROS, as well as by intrinsic limitations of both probes and experimental conditions [51,52,53]. In addition, the efficiency and specificity of many probes for detecting ROS in vitro still need to be established [46,51,52,53]. Such limitations and potential sources of artifacts complicate quantitative measurements of intracellular generation of ROS and demand careful design of the experiments and cautious interpretation of the results [53].
2. FCM in Oxidative Stress Research
FCM can be defined basically as an analytical method which measures light scattering and emission of multiple fluorescences by single cells or microscopic particles (natural or artificial) aligned by a laminar stream, when they are presented one-by-one at high speed to suitable illumination sources [54,55]. In a conventional flow cytometer, suspensions of cells and other biological particles in an appropriate isotonic liquid medium are hydrodynamically focused across an illumination system and an optical system capable of simultaneously quantifying multiple structural and functional properties of each cell or particle. The characterization of cellular parameters at rates of thousands of events per second is achieved mostly by fluorescence measurements or by other forms of interaction between light and matter [54,55]
FCM allows rapid and simultaneous analysis of multiple parameters of individual cells in a heterogeneous mixture, transported by a liquid stream across an illumination source and light detectors. The computer-integrated data provide a comprehensive description of biological features of the sample. The increasing availabity of fluorescent reagents and the recent development of algorithms for multispectral-fluorescence unmixing allows quantifying up to 30-50 parameters at the same time [56]. The ability to identify cell subpopulations, including very rare cells, makes FCM an essential tool in Cellular and Molecular Biology, Biotechnology, Toxicology and Drug Discovery, or Environmental Studies [57]. On the other hand, FCM is a firmly established technology for clinical diagnosis and prognosis, especially in Immunology [58] and Onco-Hematology [59].
Fluorescence-based analysis of ROS and oxidative stress is a very relevant application of FCM as attested by the more than 8,000 papers that have been published between 1989 and 2023, according to PubMed Central. However, to ascertain the specific role of ROS in oxidative stress studies by cytomic methodologies, it is essential to detect and characterize these species accurately. Unfortunately, the specific analysis of individual intracellular ROS remains a challenge [51,52,53], but alternative cytometric strategies aimed to other endpoints of oxidative stress can be also be considered. In this review we present and discuss briefly the limitations and perspectives of such approaches.
2.1. Specific Features and Limitations of Functional FCM
Because of the multiparametric analysis (and physical separation) of single cells or particles at very fast rate, FCM can be considered as a particular method for biochemical analysis, with advantages over other conventional methodologies applied to the study of oxidative stress.
2.1.1. Multiparametric Data Acquisition
In general, standard biochemical procedures determine a single biomarker per assay and are not sensitive enough for single-cell analysis. FCM instruments allow routinely two morphology-related parameters (forward and side-light scatter) and up to 30-50 fluorescence signals per single particle [59,60]. In this way, in a single-tube assay, one or more parameters may be used to identify and select (“gated analysis”) cell subsets in heterogeneous populations (e.g.; live, apoptotic or necrotic cells; cells of different origin or lineage; cells in different cell cycle stage, and so on), whereas other signals may be assigned to analyze specific structures or functions in the selected populations [47,48].
2.1.2. Multivariate Data Analysis
Due to the hardware and software design of current flow cytometers, multiparametric acquisition is interfaced to multivariate data analysis. In this way, cell populations can be described by the multidimensional correlation of the individual properties measured on a single-cell basis, thus increasing the discriminating power [58,60]. Moreover, the special format in which FCM raw data and experimental details are stored as listmode files, i.e.; uncorrelated data matrices for each cell (Flow Cytometry Standard, FCS), allows to define post-hoc new parameter correlations and population-gating criteria when replaying those FCS files [58,61]. This is an invaluable tool especially when scarce- or rare samples are studied.
2.1.3. Fast Analysis of Large Number of Live Cells
FCM can be performed on a large variety of biological material in different conditions of vitality (e.g.; intact fresh cells, fixed and/or permeabilized cells) [47]. The use of live cells allows to study multiple biochemical parameters in minimally perturbed intracellular environments, as well as in near-physiological extracellular conditions. The fast rate of data acquisition makes possible to examine millions of individual cells in a reasonable time allows the detection and accurate analysis of infrequent or rare cells, down to 1 event per 108 cells [58]. This is in contrast with bulk determinations in which molecules extracted from large number of cells are analyzed, yielding single average values of gene expression, enzyme activity or metabolite concentration.
2.1.4. Real-Time Flow Cytometry
Including time as an operative parameter in FCM has led to the concept of real-time flow cytometry (RT-FCM), an experimental approach in which changes in cellular parameters within heterogeneous cell populations are followed in a kinetic fashion [47,62]. In RT-FCM, the biological process of interest happens while the sample is actually run in the flow cytometer and single cells are analyzed in sequence. RT-FCM allows to monitor functional changes in high numbers of single cells with a theoretical time-resolution of less than one second, and for experimental periods ranging from few seconds to several minutes. This type of kinetic analysis is specially relevant for following very fast or transient dynamic processes, as those typical of signal transduction [47,48,59].
2.1.5. Individual Cell Sorting
FCM can not only analyze different cell populations based on the use of fluorescent probes, but can also separate these populations using the same cytometric detection principles. Cell sorting allows the combination of the intrinsic capabilities of FCM results with information obtained by image analysis and molecular techniques and provides a preparative tool for rapid isolation of rare cells of biochemical relevance [63].
The most frequent and complex cell sorters are based on the formation and deflection of microdrops by high-frequency vibration, and charged electric plates to deflect these droplets towards specific collection tubes or supports. In general, this type of separator allows several sub-populations to be separated simultaneously and at a very high speed (thousands of cells per second). A second family of cell separators are based on microfluidics. These systems, in general, require a smaller number of starting cells, but they are of lower speed and performance than electrostatic separators. On the other hand, they present fewer biological risks for the operator and are less aggressive towards the cells in process [58].
2.1.6. Limitations of Functional FCM
While the large number of cells analyzed and the instrumental settings of current cytometers provide multiple strategies to obtain primary information, and allow a large number of general applications, there are several critical points and difficulties when performing adequate functional analysis by FCM (Table 1). The limitations depend mostly on the maintenance of adequate viability or metabolic capacity of cells along sample preparation and analysis, as well as avoiding the interference of fluorescent probes with cellular functions [47,48].
3. General Strategies in Flow Cytometric Analysis of Oxygen and Oxidative Stress
Possibly the most frequent and standardized application of FCM in the field of oxidative stress is the ex vivo analysis of the respiratory burst in phagocytes and the diagnosis or prognosis of chronic granulomatous disease and sepsis (Figure 3) [59,64]. In the areas of Cell Biology and Biotechnology, the most relevant examples of FCM applications in oxidative stress in vitro research can be found in the assessment of the involvement of ROS in physiopathological conditions [64,65,66,67,68,69], the evaluation of the mechanisms involved in xenobiotic biotransformation and toxicity (Figure 3 and Figure 4) [49,69], and the analysis of antioxidant properties of drugs and natural compounds [66,69,70].
FCM is a verastile technology that provides different approaches to study the complex processes involved in the generation of ROS, their control by antioxidant systems and the physio-pathological consequences of their action. Thus, the most common FCM strategies to the study of oxygen, ROS and oxidative stress include:
- (a)
- Performing cell-based studies in hypoxic conditions:
Hypoxia can be defined as oxygen deficiency of the cellular environment and hypoxic responses mediated by hypoxia-inducible (HIF) transcription factors are associated with several pathologies, including lipid metabolism, inflammation, cardiovascular disease, hypertension, tumor-mediated immunosuppression, and neurodegenerative disease [2,71]. Hypoxia is a key factor in primary tumors and metastasis, including cell proliferation, metabolic capacity, immune response, and drug resistance to chemotherapeutic intervention [72].
FCM is commonly applied in cell-based studies comparing experimental conditions of normoxia versus hypoxia, mostly related to stem-cell [73,74,75] and cancer research [76,77] as well as for assessing oxidative stress [78], mitochondrial function [79,80] and angiogenesis [81] associated to the hypoxia/reoxygenation transition.
- (b) Monitoring intracellular Oxygen in hypoxic conditions:
Hypoxia can be monitored in living tissues with complex methods, including positron emission tomography (PET), magnetic resonance imaging (MRI) and computerized tomography (CT) [82]. Such methods have shortcomings and cannot be applied to the single-cell level or to finely detect local heterogeneity in hypoxia. On the contrary, small-molecule fluorescent probes for hypoxia have been developed, with clear advantages of simplicity, sensitivity, and high temporal- and spatial resolution [83]. The mechanism of such probes is based on the hypoxia-induced overexpression of reductases, such as cytochrome P450 reductase and nitroreductase, that catalyze oxygen-sensitive bioreductive reactions [83,84].
- (c) Direct detection of ROS, the initiators of the oxidative stress process:
This task is complex due to the low concentration, short half-life and extensive interactions of ROS, as well as by the limitations imposed by the fluorescent probes and the experimental conditions [53].
- (d) Detection of more stable oxidized end products:
FCM can be applied to the detection of stable molecules generated by the reaction of RONS with endogenous cell components or with exogenous probes added to this purpose. This strategy includes the analysis of lipid peroxidation and oxidative damage to DNA [2,53].
- (e) Assessment of antioxidant defences, mostly GSH and SH-containing proteins:
This indirect approach to oxidative stress may be limited by issues related to the complexity of the antioxidant defense by itself and to the specificity of enzymes required to fluorescent reporting of the process [2,53].
3.1. Monitoring intracellular Oxygen in hypoxic conditions
Classical hypoxia-sensing probes, such as pimonidazole and EF5 [84,85] contain nitroimidazole groups that are reduced by reductases in hypoxia to generate covalently-bound products that require cell fixation followed by immunostaining to be detected [84]. While such probes are currently applied to FCM studies of hypoxia [86,87,88], cell permeant fluorogenic substrates have been developed that can be used in live single cells without the need of fixation/permeabilizaton procedures. Hypoxia Green ReagentTM for Flow Cytometry (λ excitation=488 nm; λ emission=530 nm) [89] and its related BioTracker 520 Green Hypoxia DyeTM [90] are suitable for the detection of hypoxia in living cells. Both probes are as sensitive as pimonidazole and can be used for live cell fluorescence imaging and flow cytometry applications, with common spectral properties (λ excitation=498 nm; λ emission=520 nm). As cellular oxygen levels decrease, these probes responds by releasing rhodamine, which results in detectable emissions in the green channel.
iT™ Green Hypoxia Reagent (λ excitation=488 nm; λ emission=520 nm) and Image-iT™ Red Hypoxia Reagent (λ excitation=490 nm; λ emission=610 nm) are live-cell permeable compounds which increase fluorescence in environments with low oxygen concentrations. Unlike pimonidazole adducts, which only respond to oxygen levels lower than 1%, Image-iT™ reagents are fluorogenic when atmospheric oxygen levels are lower than 5%, and their fluorogenic response increases as the oxygen levels decrease in the environment [91,92].
New indolequinone-based bioreductive fluorescent probes have been synthesized that allow imaging different levels of hypoxia in 2D and 3D cell cultures [83]. These probes are modified versions of the existing resorufin and Me-Tokyo Green fluorophores. The resorufin-based probe is activated in conditions of 4% O2 and lower, while the Me-Tokyo Green-based probe is only activated in severe hypoxia, at 0.5% O2 and less [83].
Green-fluorescent protein (EGFP) constructs under the control of hypoxia-induced promoters have been used as hypoxia reporters by fluorescence imaging of single cells, allowing dye-free monitoring of hypoxic responses, despite the O2 requirement for EGFP-fluorophore formation [93,94]. More recently, a novel family of hypoxia sensors have been described, based on UnaG, a fluorescent protein from Japanese freshwater eel that does not rely on O2 to adopt the fluorescent state [94,95]. The combination of UnaG with oxygen-sensitive fluorescent proteins has led to several reporters of hypoxia and reoxygenation that allow to record the dynamics of hypoxia in living cells [96].
3.2. Direct Detection of ROS using fluorogenic substrates
Fluorescent probes and fluorogenic substrates provide a simple and easy approach for the detection and quantification of ROS production in cellular systems. However, there are many limitations and artifacts in their application that will be discussed further away.
3.2.1. 1O2 Probes
The fluorescent probes designed for 1O2 take advantage of its chemical reactivity and combine a chemical 1O2 trap and diene-containing fluorophores [45,97,98,99,100]. On this chemical motif, structural modifications of 1O2 probes result in different optical properties. Thus, 1,3 Diphenylisobenzofuran (DPBF) reacts with 1O2 to form a nonfluorescent endoperoxide [99]. On the contrary, substituted derivatives of DPBF with phenanthrene (PPBF), pyrene (PyPBF) and 4-(diphenylamino) stilbene (StPBF) can acts as ratiometric probe for 1O2 detection. These 1O2 probes exhibit significant red shift in their emission spectrum as the conjugation increases from DBPF to StPBF [99].
9-[2-(3-Carboxy-9,10-diphenyl)anthryl]-6-hydroxy-3H-xanthen-3-one (DPAX) is a sensitive and efficient fluorescent probe for the detection of 1O2 which combines a fluorescein moiety with DPA [100]. DPAX and its derivatives show very low fluorescence intensity in aqueous solution but once bound to 1O2 the corresponding endoperoxide (DPAX-EP) emits intense fluorescence [100]. DPAX and its derivatives demonstrate excellent selectivity towards 1O2 as compared with other RONS [100]. DPAXs stability of fluorescence intensity can be enhanced by incorporating electron withdrawing groups like Cl or F at the 2- and 7- positions of the xanthene moiety leading to generation of DPAX-2 (Cl derivative) and DPAX-3(F-derivative) [45,100].
9-[2-(3-Carboxy-9,10-dimethyl)anthryl]-6-hydroxy-3H-xanthen-3-one (DMAX) reacts rapidly with 1O2 with much greater sensitivity than DPAX. Both DMAX and its endoperoxide DMAX-EP have similar excitation (λ = 492 nm) and emission (λ = 515 nm) wavelengths, compatible with most FCM instruments, but DMAX-EP is highly fluorescent whereas DMAX itself is practically non-fluorescent. Further, the hydrophobicity of DMAX is less than that of DPAXs making it suitable to use for assays in biological sample [100].
More recently, dansyl-based (DanePy) [101] and Singlet Oxygen Sensor Green reagent® (SOSG) [98,102] fluorescent probes have been synthesized, both probes composed of an anthracene moiety (electron donor) that quenches the fluorescence of the fluorochrome (electron acceptor) through electron transfer [102]. When the anthracene moiety traps 1O2, the resulting adduct ceses being an electron donor and the fluorescence is recovered [103]
SOSG is currently the choice probe for detection of 1O2, due to its claimed high selectivity and specificity to 1O2 [98,102,103,104] This fluorescein-based dye probe upon reaction with 1O2, generates SOSG endoperoxide (SOSG-EP) that emit green fluorescence (λ excitation= 504-508 nm; λ emission= 525-536 nm) [103,104]. Spectral properties of SOSG are similar to those of fluorescein which made this probe suitable for FCM instruments. However, SOSG presents some drawbacks, mostly unequal penetration in cells and photosensitization resulting from 1O2 generation by SOSG itself under exposure to UV radiation (355 nm) and visible light (532 nm), respectively [104]. While SOSG does not show an appreciable response to O2•− and HO.; its fluorescence emmision increases dose-dependently when exposed to gamma-rays or X-rays, in conditions where formation of 1O2 is ruled out [105].
The modified 1O2 indicator Aarhus Sensor Green (ASG) (a tetrafluoro-substituted fluorescein derivative covalently linked to a 9,10-diphenyl anthracene moiety) has spectral properties similar to SOSG without inducing photosensitization [106].
3.2.2. 2’,7’-Dichlorodihydrofluorescein diacetate (H2DCF-DA) and related probes
Cell-permeant H2DCF-DA is one of the most popular fluorogenic substrates for ROS detection [51,107,108]. After cleavage of acetate groups by intracellular esterases, the oxidation of intracellular 2,7-dichlorodihydrofluorescein (H2DCF) yields fluorescent 2,7-dichlorofluorescein (DCF; λ excitation=498 nm; λ emission=522 nm). While widely considered a specific indicator for H2O2 [109], H2DCF is oxidized by other ROS, such as OH. and peroxyl radicals, and also by RNS like ONOO [51,107,108]. On the other hand, it seems well established that H2DCF is not oxidized by O2.-, hypochlorous acid or NO [107]. With these caveats, H2DCF has been successfully used for studies of oxidative burst in phagocytes [110,111] and to follow the generation of prooxidants in many cell models [112,113,114,115,116].
Intracellular oxidation of H2DCF in conditions of cell damage, tends to be accompanied by leakage of the product, DCF. To enhance retention of the fluorescent product, several analogs with improved retention have been designed, such as carboxylated H2DCF -DA (carboxy- H2DCF -DA), which has two negative charges at physiological pH, and its di-(acetoxymethyl ester) [117]. The halogenated derivatives 5-(and 6-)chloromethyl-2’,7’-dichlorodihydrofluorescein diacetate, acetyl ester (CM- H2DCF -DA), and 5-(and 6-) carboxy-2’,7’-difluorodihydrofluorescein diacetate (carboxy-H2DFF-DA) exhibit much better retention in live cells and have been used for monitoring oxidative burst in clinical settings and applied to different experimental studies related to oxidative stress [117].
3.2.3. Dihydrorhodamine 123 (DHR123)
DHR123 is a non-fluorescent molecule that upon oxidation oxidation generates rhodamine 123, a cell-permeable fluorescent cationic probe (λ excitation=505 nm; λ emission=529 nm) [51,107]. Upon oxidation of DHR123 to the fluorescent rhodamine 123, one of the two equivalent amino groups tautomerizes into an imino group, effectively trapping rhodamine 123 within mitochondria [118]. DHR123 is oxidized by H2O2 in the presence of peroxidases, but it can also be oxidized by other reactive oxidants, as ONOO, Fe2+, Fe3+ in the presence of ascorbate or EDTA, cytochrome c, or HOCl [62,81]. DHR123 is not directly oxidized by H2O2 alone, O2.- nor by the xanthine/xanthine oxidase system [107,118].
Although DHR123 was described initially as a fluorogenic susbstrate for H2O2 [51], currently it is the most frequently used probe for measuring ONOO [118,119,120]. However, the oxidation of DHR123 by ONOO is not induced directly by this species, but is mediated by intermediate oxidants formed from the rapid and spontaneous decomposition of ONOO [118,119,120].
3.2.4. Mitochondria peroxy yellow 1 (MitoPY1) and related arylboronate fluorescent probes
Recently, new H2O2 chemoselective probes have been developed based on the selective H2O2 -mediated transformation of arylboronates to phenols [121,122]. Arylboronates are linked to fluorogenic moieties, so that reaction with H2O2 generates a fluorescent probe [121]. Arylboronate probes include peroxyfluor-2 (PF2), peroxy yellow 1 (PY1), peroxy orange 1 (PO1), peroxyfluor-6 acetoxymethyl ester (PF6-AM) and mitochondria peroxy yellow 1 (MitoPY1) [121,122,123,124,125,126]. Adding appropriate functional groups results in organelle-specific targetting for measuring H2O2 with spatial resolution. Thus, MitoPY1 and SHP-Mito [124,125,126] target mitochondria. while Nuclear Peroxy Emerald (NucPE) is suitable for nuclear targeting [121]. In addition, Ratio Peroxyfluor 1 (RPF-1) provides a ratiometric change of two fluorescent signals upon reaction with H2O2, allowing correlating fluorescence ratio to probe concentration [125].
3.2.5. Hydroethidine and MitoSOX mitochondrial O2.- indicators
Hydroethidine (HE), also know as dihydroethidium, is widely used for detecting O2.- anion [51,107,108,117,127,128]. HE is membrane-permeant, and intracellular HE exhibits blue fluorescence, but once oxidized by O2.-, it originates 2-hydroxy-ethidium (E+), a fluorescent compound (λ excitation=520 nm; λ emission=610 nm). E+ is retained in the nucleus, intercalating with DNA, a fact that increases its fluorescence [107].
HE has been repeatedly used in studies of oxidative burst in leukocytes [92,93] and during inflammation [130,131,132,133]. HE has been used also for mitochondrial O2.- detection [117,134,135] although MitoSOX Red indicator provides more specific mitochondrial localization, as discussed later [117,136]. While HE (and also Mito-SOX Red) have been used to detect mitochondrial O2.- generation associated to the induction and execution of apoptosis [134,135], HE may have important limitations when used for analysis of intracellular O2.- as it has been shown that cytochrome c is able to oxidize HE, an aspect that might be relevant in conditions of apoptosis, where cytochrome c is released to cytosol [133]. Furthermore, HE can also be oxidized by a variety of reactive species, including ONOO. Thus, HE should be considered as an indicator of ROS and RNS production [51,107,108,133].
MitoSOX Red mitochondrial O2.- indicator (MitoSOX Red) is a cationic derivative of HE, for detection of O2.- in the mitochondria of live cells [51,117,137]. MitoSOX Red contains a triphenylphosphonium cationic substituent that selectively targets the probe to active mitochondria, as a function of mitochondrial membrane potential [117]. Oxidation of MitoSOX Red by O2.- results in hydroxylation of the ethidium moiety at the 2-position, to yield a 2-hydroxyethidium substituent. Therefore, the fluorescence spectral properties of oxidized MitoSox Red (λ excitation=488 nm; λ emission=610 nm) are comparable to those of HE. Nevertheless, MitoSOX Red shows also an absorption peak at 396, which may be used for more accurate detection of O2.- [117]. MitoSOX Red has been used for detection of mitochondrial O2.- production in a wide variety of cell types and conditions [107,117,136], including hypoxia [138]. Recently, the green-emitting version of MitoSOX Red, the probe MitoSOX Green (λ excitation=488 nm; λ emission=510 nm). has been marketed for detection of mitochondrial O2.- [117]. Since the chemical reactivity with O2.- of the MitoSOX dyes is similar to the reactivity of HE with O2.-, the limitations of HE apply also to MitoSOX Red and Green dyes [51,139].
3.2.6. CellROX® reagents
The CellROX® reagents are a series of proprietary cell-permeant dyes, weakly fluorescent in reduced state and exhibiting photostable fluorescence upon oxidation by ROS [140,141,142,143]. CellROX® Green only becomes fluorescent (λ excitation=485 nm; λ emission=520 nm) with subsequent binding to DNA, limiting its presence to the nucleus or mitochondria. This reagent can be formaldehyde-fixed and its signal survives detergent treatment, allowing it to be it multiplexed with other compatible dyes and antibodies. CellROX® Orange (λ excitation=545 nm; λ emission=565 nm) and CellROX® Deep Red (λ excitation=640 nm; λ emission=665 nm) do not require DNA binding for fluorescence and are localized in the cytoplasm [144]
3.2.7. ROS-ID® reagents
The ROS-ID® Total ROS/Superoxide detection kit is a propietary system for real-time measurement of global ROS levels and specifically O2.- in living cells [145]. The probe Oxidative Stress Green is a cell-permeable and fluorogenic probe that reacts directly with a wide range of ROS and RNS, generating a green fluorescent product [146]. The reagent Superoxide Orange is a cell permeable probe that reacts specifically with O2.- and generates an orange fluorescent product [147].
3.3. Detection of more stable products of ROS reaction
3.3.1. Detection of Lipid Peroxidation
Peroxyl radicals are formed by the decomposition of various peroxides and hydroperoxides, including lipid hydroperoxides. The hydroperoxyl radical is also the protonated form of O2.- , and approximately 0.3% of the O2.- in the cytosol is present as this protonated radical [117].
cis-Parinaric Acid
cis-Parinaric acid is a fluorescent 18-carbon polyunsaturated fatty acid, containing four conjugated double bonds in positions 9, 11, 13 and 15 [107,117]. cis-Parinaric can be metabolically integrated into phospholipids of cell membranes, where its conformation and mobility are comparable to endogenous phospholipids. Its fluorescent and peroxidative properties are combined in the conjugated system of double bonds. The fluorescence of cis-parinaric acid (λ excitation=320 nm; λ emission=432 nm) is lost upon oxidation [107,117]
cis-Parinaric has been repeatedly used to measure lipid peroxidation in a multiplicity of cell systems and conditions [148,149,150]. However, there are some limitations associated with the use of this probe in cytometry, such as its excitation by UV lasers, still absent in many flow cytometers. In addition, cis-parinaric is very sensitive to air and undergoes photodimerization under illumination, resulting in loss of fluorescence and overestimation of lipid peroxidation [117].
Lipophilic Fluorescein derivatives
The probe 5-(N-dodecanoyl) aminofluorescein (C11-Fluor) is a lipophilic derivative of fluorescein that remains associated to cell membranes in a stable and irreversible way. C11-Fluor has been used in flow cytometry for determining membrane-lipid peroxidation [43,156]. Other lipophilic derivatives of fluorescein, include 5-hexadecanoylaminofluorescein (C16-Fluor), 5-octadecanoyl-aminofluorescein (C18-Fluor) and di-hexadecanoyl-glycerophosphoethanolamine (Fluor-DHPE) [157].
3.3.2. Detection of Metabolic Derivatives of Peroxidized Lipids
Immunofluorescent detection of 4-hydroxy-2-nonenal (4-HNE)
Several aldehydes can be formed as end-products of the peroxidation process. 4-HNE is an unsaturated aldehyde arising from peroxidation of -6 unsaturated fatty acids. 4-HNE is highly reactive towards free SH- groups of proteins and is highly cytotoxic and genotoxic [158,159].
Monoclonal antibodies recognizing adducts of 4-HNE with histidine, lysine, and cysteine in proteins are now commercially available [160]. These antibodies have been conjugated with different fluorochromes and can be used with high specificity for detecting advanced stages of lipid peroxidation [161].
Immunofluorescent detection of oxidized bases in DNA
The oxidized DNA base 8-oxodeoxyguanine (8-oxoDG) is a major form of oxidative DNA damage derived from the attack by OH. radical to guanine at the C8-position, resulting in a C8-OH-adduct radical. Thus, 8-oxoDG is formed during free radical damage to DNA and is a sensitive and specific indicator of DNA oxidation [162,163].
8-oxoDG has been quantified with the OxyDNA Assay, based on the specific binding of a monoclonal antibody conjugated with FITC to the 8-oxoDG moieties in the DNA of fixed and permeabilized cells [164]. This assay has been used to detect oxidative genotoxicity in vitro [165], including environmental studies [166]. Of particular interest, the OxyDNA assay has been used in a number of fertility studies related to oxidative stress during cryopreservation of sperm cells [167] and the relation of oxidative DNA damage to fertility in humans [168,169] and animals [170].
3.4. Assessment of Antioxidant Defenses: Glutathione (GSH) and Thiols (SH)
SH groups, and especially GSH, act as nucleophiles and protect against toxicity, mutagenicity, or transformation by ionizing radiation and many carcinogens [40]. The availability of many thiol-reactive fluorescent probes since early in the 1980s has allowed FCM assays for GSH [171,172] and free SH- groups [173] in living cells, and analysis of intracellular levels of GSH and of GSH S-transferase activity (GST) are relevant applications of FCM in oxidative stress and drug resistance [174]. Cytometric assays for GSH and intracellular SH-groups have been critically reviewed in several publications [175,176,177,178].
The probes most used for cytometric analysis of GSH and GST have been the UV-excited, cell-permeant bimanes, including monobromobimane (mBrB) and the more selective monochlorobimane (mClB). Both probes are essentially nonfluorescent until conjugated to GSH [174,175,176,177]. o-Phthaldialdehyde, another UV reagent, reacts with both the thiol and the amine functions of GSH, yielding a cyclic derivative with excitation and emission maxima shifted from those of the protein adducts, improving the specificity of GSH detection [174,175,176,177]. ThiolTracker Violet (λ excitation=405 nm; λ emission=525 nm) is up to 10-fold brighter than the bimanes. This cell-permeant probe resists formaldehyde fixation and detergent extraction, allowing analyzing fixed cells [174,178].
GSH has been determined also with visible light–excitable probes, including Mercury Orange [173], 5-chloromethylfluorescein diacetate (CellTracker Green CMFDA), and chloromethyl SNARF-1 acetate. Both probes form adducts with intracellular thiols that are well retained by viable cells. CellTracker Green CMFDA is brighter than MClB, and is highly specific for GSH over free SH-groups [174]. The GSH-adduct of chloromethyl SNARF-1 emits beyond 630 nm, allowing multicolor protocols and reducing the impact of cellular autofluorescence.
4. FCM in Oxidative Stress Research
As commented above, cytometric analysis of Oxygen, ROS and Oxidative Stress is complex due to the low concentration, short half-life and extensive interactions of ROS and RNS, as well as by limitations intrinsic to the fluorescent probes or resulting from the experimental conditions. Such limitations are potential sources of artifacts that challenge quantitative analysis of intracellular ROS and require careful design of the experiments and cautious interpretation of data [53].
4.1. Variability in half-life and intracellular sources of ROS
While most ROS are short-lived molecules, there is large heterogeneity in their lifetime and the microenvironments where they are produced and consumed [51,108]. While ROS of low reactivity may accumulate with time, the more reactive ROS will reach a steady state in which the rate of their generation will be equal to the rate of disappearance by self-reaction and reaction with cellular components or with the probe.
Intracellular locations of ROS and RNS can be approached through molecular design of the fluorogenic substrates [51,52,53]. Introduction of chemical modifications in their molecular scaffolds renders the probes permeable to plasma- and organelle membranes, thus allowing to target them to specific intracellular environments [51,52,107,108]. For instance, dihydrocalcein accumulates in mitochondria, in contrast to H2DCF which localizes usually in the cytoplasm [179]. However, preferential localization of H2DCF in the mitochondria of rat cardiomyocytes has been reported [180].
4.2. Interactions among and between ROS and RNS
Mitochondrial respiration is a good example of ROS interplay, as O2.- anion, H2O2 and OH. radical are sequentially produced by a series of partial reductions of O2 molecule. Incorporation of an electron into O2 gives rise to O2.- anion, which is a poorly reactive radical but which can oxidize thiols and ascorbic acid [5,10,40]. O2.- gives rise to H2O2 by spontaneous reaction or by the action of superoxide dismutase. H2O2, in turn, can react with different organic compounds to produce peroxyl radicals that will eventually release OH. radicals during their metabolism. Moreover, by way of the Fenton reaction, OH. radicals are produced when H2O2 and a transition metal, such as Fe2+, react together, yielding Fe3+ that consumes O2.- for recycling Fe2+. In the Haber-Weiss reaction, O2.- and H2O2 react together to produce OH. radicals [5,10].
On the other hand, the interaction of ROS with nitrogen derivatives can generate RNS. For instance, NO is synthesized in many cell types from L-arginine by different NO synthetase isoforms. NO is a weak reductor and reacts with O2to form NO2, but reacts much faster with O2.- to produce ONOO, a strong oxidant [33,34,181].
4.3. Influence of the probes on the experimental system
Many reagents used for flow cytometric analysis of ROS are chemically-reduced fluorogenic substrates, intrinsecally suceptible of auto-oxidation, which usually produces 1O2, O2.- and H2O2. If the rate of auto-oxidation is significant, it may result in artifactual detection of ROS and higher background, a problem especially evident for probes such as HE or MitoSOX Red [51,107,108].
The concentration of the probe is also relevant, as it may affect the stoichiometry of the process under study. For instance, the stoichiometry of the reaction between HE and O2.- depends on the ratio of O2.- flux and HE concentration. Due to HE-catalyzed O2.- dismutation, the efficiency of HE oxidation decreases at higher rates of O2.- generation and high HE concentrations might lead to fluorescence increase independent of O2.- [107].
Fluorescent probes at high concentration, may interfere with cell functions and be toxic [47,53]. For example, when irradiated with UVA, H2DCF auto-oxidizes and photo-sensitizes cells [108]. In addition, probes may affect the activity of ROS-producing enzymes. For instance, H2DCF can be a source of electrons for the oxidation of arachidonic acid by prostaglandin H synthase [108], while dihydrocalcein may inhibit mitochondrial-chain complex I [51].
4.4. Cell integrity and intracellular retention of probes
Loss of intracellular fluorogenic substrates or fluorescent probes from injured cells is a common problem in FCM analysis, most specially in pharmaco-toxicological studies. Passive leakage is always present in necrotic or apoptotic cells, due to enhanced plasma membrane permeability [47,48,53]. In oxidative-stress related studies, therefore, a certain degree of damage to plasma membrane should be expected, with leakage of intracellular probes or their oxidation products leading to artifacts or erroneous interpretation of results.
On the other hand, the expression of multidrug transporters in the plasma membrane of cells may result in probe extrusion, leading to underestimate oxidative stress [151], as multidrug-resistant cells might appear to produce less ROS. Fluorescent molecules such as rhodamine 123 and ethidium are good substrates for P glycoprotein, while fluorescein and dihydrofluorescein are substrates for MRP1 [182]. Dihydrocalcein has been preferred to H2DCF because its oxidation product calcein is believed to not leak out of cells, however, calcein is also a good substrate for MRP1 and MRP2 transporters [43].
4.5. Experimental artifacts
The photochemical reaction of diverse components of culture media may spontaneously generate ROS [43,183]. Xenobiotics and endogenous compounds such as catechols, dopamine, hydralazine and molecules with SH- groups may also produce significant ROS upon interaction with the media [43]. Interestinly, the presence of ROS has been detected even in natural environments, such as seawater [184,185].
4.6. Intrinsic limitations of fluorogenic substrates and probes
4.6.1. Probes used for detection of H2O2 and organic peroxides
H2DCF-DA is most widely used for detecting intracellular H2O2 and oxidative stress. Traditionallty, H2DCF -DA and DHR123 are assumed to be oxidized by H2O2 and organic peroxides [109,117]. However, these probes do not react directly with H2O2 in the absence of peroxidases [51,108]. Even if H2DCF oxidation also occurs by action of H2O2 or O2 in the presence of Fe2+, the OH. radical is the species responsible for such oxidation [107]. Since the oxidation of H2DCF and DHR123 by H2O2 under physiological conditions requires peroxidase-dependent systems, enzyme activity may become a limiting factor and thus measurement of probe oxidation might be rather considered a measure of peroxidase activity. However, H2DCF and DHR123 can be oxidized not only by the peroxidases, but by other related enzymes, such as xanthine oxidase, superoxide dismutase and cytochrome C [107].
H2DCF and DHR123 are not oxidized by NO or O2.- to any significant extent, but they are also very efficiently oxidized by ONOO via the radicals generated during ONOO decomposition [119,120]. DCF may undergo photoreduction by visible light or by UVA radiation [187]. This mechanism may generate a semiquinone radical from DCF which originates O2.- by reaction with O2. Sequentially, the dismutation of O2.- generates H2O2, which leads to an artificial increase of H2DCF oxidation and to amplification of DCF fluorescence.
MitoPY-1 and other aromatic boronate derivatives proposed for analysis of intramitochondrial generation of H2O2 [121,122,123] also react with ONOO a million times faster than they do with H2O2 [188]. Because of this reactivity, it is critical to perform proper controls when using a boronate-based fluorescent probe, such as expression of catalase or using a ONOO-specific probe.
4.6.2. Probes used for detection of O2.-
HE and Mito-SOX Red are widely used for detection of intracellular- and mitochondrial O2.- [51,108]. The red fluorescence of the two-electron oxidation product of HE, ethidium (E+), is usually considered the proof of intracellular O2.- formation. However, it has been demonstrated that E+ is not formed from the direct oxidation of HE by O2.- [189,190]. Instead, 2- hydroxyethidium (2-OH-E+), a different product with similar fluorescence characteristics, is formed by reaction of HE with O2.- [190]. E+ and other dimeric products, but not 2-OH-E+, are generated by the reaction between HE and oxidants such as ONOO, OH.; H2O2, and peroxidase intermediates. Thus, 2-OH-E+ is only a qualitative indicator of intracellular O2.- [51,191].
HE is oxidized directly by ferricytochrome c [127] and by other heme proteins. Oxidation of the probe by cytochromes c, c1, b562, b566 and aa3 is oxygen-independent while oxidation by met-hemoglobin and met--myoglobin is strictly oxygen-dependent, with products consisting of a mixture of species resulting from 1 to 4-electron abstraction from HE. Although they are different from the O2.- oxidation product, their excitation/emission peaks are close to those generated by O2.- [43,192].
Because of its positive charges, Mito-SOX reacts slightly faster with O2.- compared to HE [101]. Mito-SOX reacts with and forms a red fluorescent product, 2-hydroxy-mitoethidium (2-OH-Mito-E+), and not Mito-E+. 2-OH-Mito-E+, the specific product of O2.- with Mito-SOX and Mito-E+, the nonspecific product of Mito-SOX, have overlapping fluorescence spectra. Thus, the red fluorescence formed from Mito-SOX localized in mitochondria is not a reliable indicator of mitochondrial formation of O2.- , as it might arise also from an oxidation product of Mito-SOX induced by one-electron oxidants (such as cytochrome c, peroxidase, and H2O2) [51,137,189,190,191].
4.6.3. Probes used for detection of lipid peroxides
The presence of four double bonds in cis-parinaric acid makes this probe very susceptible to oxidation if not carefully protected from air [43,117], and cis-parinaric samples should be handled under inert gas and the solutions prepared with degassed buffers and solvents. cis-Parinaric acid is also photolabile and undergoes photodimerization when exposed to intense illumination, resulting in loss of fluorescence [117].
4.6.4. Probes used for the determination of GSH
In many cases, the fluorescent reagents designed to measure GSH may react with other free or protein-bound intracellular thiols [173,176,194]. An important aspect in the use of GSH reagents is the large interspecies and tissue variability of cellular GSH content and the presence of GST isozymes [194]. For instance, mClB is highly selective for GSH in rodents, but has a low affinity for human GST [176].
5. Recommendations for performing FCM analysis of ROS, RNS and Oxidative Stress
According to Woolley et al. [195], a fluorescent probe must fulfill a series of criteria in order to be an “ideal” indicator of ROS. These criteria include probe selectivity for a particular species of ROS, fast and reversible kinetics when reacting with ROS, and adequate subcellular compartmentalization. The probe should be excitable at a visible wavelength, to be resistant and not to show any toxicity in general and phototoxicity in particular. To the above criteria, it must be added that of the probe not interfering with the biological process under study [47,48].
Following these principles, we have recently performed a systematical study to detect experimental issues related to the specificity of fluorescent probes and the involvement of different ROS in E. coli [126] and eukaryotic [196] models of oxidative stress. Our results, as summarized below, may provide recommendations for proper design of cytometric studies of oxidative stress to prevent or minimize experimental errors.
5.1. Inclusion of Experimental Controls
Accordingly to the limitations and caveats presented above, including appropriate positive and negative controls is very important when performing cytometric experiments or analyses related to ROS and oxidative stress. When possible, direct visualization of intracellular of ROS generation by co-localization techniques is highly recommendable [50,53], detailed discussion of possible controls in such studies is beyond the scope of this review, as the biochemical complexity of experimental oxidants and antioxidants parallels that of their biological counterparts [40,197].
5.1.1. Positive Controls
In general, the most frequent controls are positive controls, molecules or complex systems that directly or indirectly increase the intracellular level of ROS or mimick the cellular effects of oxidative stress. Pro-oxidants are chemicals that induce oxidative stress, either by generating ROS or by inhibiting antioxidant systems [40,197].
To mimic mitochondrial H2O2 production, cells can be treated with the complex I respiratory chain inhibitor rotenone [123]. Peroxyl radicals, including alkylperoxyl and hydroperoxyl radicals can be generated from compounds such as 2,2’-azobis(2-amidinopropane) and from hydroperoxides such as tert-butyl hydroperoxide(t-BOOH) or cumene hydroperoxide (CHP) [123,126]. The OH. radical can be generated from O2.- donors (e.g.; plumbagin or menadione) [123,181] or by exogenous H2O2 in a Fenton reaction catalyzed by Fe2+ or other transition metals, as well as by the effect of ionizing radiation [123,126]. O2.- can be most effectively produced by the hypoxanthine–xanthine oxidase generating system [198].
Many xenobiotics, including anticancer agents such as anthracyclines and cis-platin [199], and natural redox active toxins, like pyocyanin, [200] generate ROS and can be used as positive controls.
When using prooxidant molecules with different physico-chemical properties, as H2O2 and organic peroxides such as t-BOOH and CHP, it is important to consider the role of their lipophilicity and their ability to penetrate or transport through membranes. Thus, at comparable oxidant concentrations, the oxidative stress induced in membrane is always much higher for t-BOOH and CHP while intracellular oxidative effects are observed with H2O2 [3,126]. Typically, the intracellular action of t-BOOH and CHP is gradual, generating ROS that produce membrane-initiated oxidative stress, which increases over time to a maximum and then decreases. On the contrary, H2O2 reacts rapidly generating ROS highly reactive towards intracellular components. H2O2 -induced oxidative stress usually is maximal immediately after addition and rapidly disappears.
Intracellular levels of ROS can be also increased by attenuating or inhibiting antioxidant defense. A convenient strategy involves depletion of intracellular GSH stores by inhibiting GSH biosynthesis or by accelerating GSH oxidation [201]. Also, inhibitors of antioxidant enzymes, such as superoxide dismutase [202] and catalase [203] have been used to increase intracellular ROS and induce oxidative stress.
5.1.2. Negative Controls
Negative controls are designed to reduce the levels of ROS or to atenuate their biological effects. If possible, controls should be specific about which particular reactive species is (are) or enzyme system is involved, but in most cases, controls do not attempt to that degree of specificity [51,52,108]. Antioxidants can be categorized as enzymatic and nonenzymatic [197].
Enzymatic antioxidants work by transforming oxidative products to H2O2 and then to H2O, in a sequential process. Cell-permeable forms of antioxidant enzymes, such as polyethylenglycol-superoxide dismutase [204] can be also used to decrease specifically intracelllular ROS.
Non-enzymatic antioxidants work by disrupting free-radical initiated chain reactions. Such antioxidants can be classified depending on whether they are hydrophilic (e.g. ascorbic acid, N-acetyl cysteine, GSH-esters) or lipophilic (e.g. -tocopherol and Trolox)[40]. In general, water-soluble antioxidants react with oxidants in the cytosol while lipid-soluble antioxidants protect cell membranes from lipid peroxidation [40]. In addition, chelators of transition metals exert also antioxidant effects, based upon the attenuation of Fenton-type reactions [40,197].
Regarding the use of chemical antioxidants as negative controls, it should be kept in mind that reducing agents may become pro-oxidants. For instance, ascorbate has antioxidant activity when it reduces oxidizing substances such as H2O2, but it can also reduce metal ions, generating free radicals by the Fenton reaction [40,197].
Regarding the specificity of antioxidants, all organic compounds react with OH. radicals with rate constants approaching the diffusion limit. Thus, in solution, no compound really has anymore significant OH. radical scavenging activity than other molecules (proteins, lipids, nucleic acids, amino acids, numerous metabolites, etc.) already present in the biological system [40]. On the contrary, α-tocopherol, due to its specific uptake into membranes and relatively rapid kinetics of reaction with lipid hydroperoxyl radicals compared with their propagation reaction is an effective chain breaker of lipid peroxidation [40].
5.1.3. Genetically-Modified organisms as controls
Genetically-modified organisms (GMO) are alternative approaches to standard positive and negative controls in studies of oxidative stress. For instance, Guo et al. [124] used an enzymatic method to generate cytoplasmic H2O2 in astrocytes. Primary astrocytes were transduced with adenoviruses containing the cDNA for cytoplasmic D-Amino acid oxidase (DAAO). DAAO oxidatively deaminates d-amino acids using FAD as electron acceptor. At the same time, DAAO uses O2 to oxidize FAD, thus generating H2O2 in a dose-dependent manner relative to the concentration of D-alanine.
Bacterial strains, mostly E. coli, with genetical modifications in genes related to the antioxidant defense have been used repeatedly as models for studying ROS mechanisms and oxidative stress [70,126,205,206]. However, functional FCM assays in live bacteria are limited by the impaired penetration of vital dyes across the cell wall, thus imposing permeabilization procedures that are time-consuming and affect cell physiology, leading to aggregation or cell lysis [206].
E. coli B is extensively used for mutagenic assays [207,208,209,210], while E. coli K12 strain is applied mostly to genetic and biochemical studies. E. coli B WP2 strains express constitutively a different cell-wall and outer-membrane lipopolysaccharide composition that results in increased membrane permeability and renders this strain more favorable for both protein secretion and uptake of exogenous chemicals, as recently confirmed by multi-omic analysis comparison of E.coli B and E. coli K12 strains [211].
In accordance with these findings, we have previously demonstrated the utility of E. coli B as a naturally-permeable strain in straight-forward functional FCM assays [205]. The E. coli B strain IC188 exhibited more efficient staining with vital fluorochromes than the E. coli K-12 strain AB1157, while maintaining similar membrane potential. In addition, we found IC188 strain to be more sensitive than AB1157 for revealing oxidative stress when challenged with prooxidants, supporting its suitability as a biosensor of oxidative stress [205]. On this background, we have developed a collection of genetically modified strains of E. coli B WP2 [209] based on the inactivation of the oxyR operon, a main sensor of oxidative stress [212]. Such oxyR-deficient bacterial strains show enhanced sensitivity to oxidative stress and increased accumulation of intracellular ROS when examined by FCM using fluorogenic susbstrates [206]. More recently [126], we have characterized two novel biosensors derived from the E. coli B parental wild type (strain IC188) which are deficient either in the OxyR function (strain IC203) or simultaneously in OxyR, SodA and SodB functions (strain IC5233). On these models, we have quantified the intracellular levels of ROS by FCM using ROS-sensitive fluorescent probes after exposure to relevant peroxidative xenobiotics differing in solubility and prooxidant mechanisms.
5.2. Choice of Fluorescent Probes
Our results are consistent with a higher sensitivity and specificity of the green-emitting MitoPY1 probe for the detection of H2O2, especially in the bacterial model [126] and in RT-FCM experiments with hepatoma cells [196]. On the contrary, our results showed that MitoPY1 is not be the fluorescent probe of choice for oxidative stress induced by organic peroxides or redox cycle compounds. In fact, MitoPY1 is a chemoselective fluorescent reporter of the arylboronate family, with improved selectivity for H2O2 over other ROS [220]. In addition, MitoPY1 is a probe designed for its mitochondrial localization, which is why it probably detects H2O2 present in the mitochondria [221], which would explain the low response of MitoPY1 to exogenous organic hydroperoxides and redox cycle compounds, which initially generate H2O2 in membrane environments and initiate chain reactions that lead to the diffusion of ROS and the generation of the highly reactive OH. radical [126].
Our results with E. coli [126] and cancer cell lines [196] also showed that the green-emitting probes H2DCF -DA and DHR123 are more sensitive to organic peroxides, including t-BOOH and CHP, than to H2O2, confirming their lack of selectivity of H2DCF for H2O2. These results confirm the caveats regarding H2DCF and DHR123 [213], possibly the most widely used probes to detect intracellular oxidative stress [53]. Accordingly, both probes should be used rather than MitoPY1 for studies involving organic peroxides acting through H2O2 -indepent mechanisms.
The orange-red emission fluorochromes HE and its mitochondrial localization derivative MitoSOX Red are widely used in CMF as indicators of high specificity towards O2.- [127,137]. In line with this idea, a greater response has been shown in the MitoSox Red probe when inducing with menadione and greater HE specificity with plumbagin, both O2.- donor xenobiotics. Based on these observations, these probes could be recommended for use in oxidative stress studies in which it is desired to quantify O2.- generation. A study in HT22 cells exposed to menadione showed an accumulation of O2.- in mitochondria [214]. This is related to what was observed in Jurkat cells, with higher ratios of MitoSOX Red, a mitochondrial probe, after incubation with menadione.
Consistent with the specificity of HE and MitoSOX Red, the results with Jurkat and N13 cells [196] show the very low response of both probes to treatment with H2O2 but dose-dependent weak responses to all agents prooxidants used in the Jurkat cell model. On the other hand, in the bacterial strain model [126], although we do not have data for MitoSox Red, the HE responded to all the oxidizing agents used, with different levels of intensity, which casts doubts to the direct applicability of this probe as a specific sensor. of O2.- and requires a careful review of the chosen experimental model.
When fine-tuning multiparametric kinetic and end-point assays of oxidative stress [196] we have found striking discrepancies in the responses of the ROS-sensitive probes analyzed individually or in spectrally compatible pairs. Since in such experiments fluorescence compensation was performed between the pairs of fluorochromes, the data obtained seemed to suggest the existence of biochemical interferences between the probes of each pair and/or of the probes with ROS metabolism and oxidative processes induced by xenobiotics [57]. Thus, while the different fluorescent probes responded individually to the addition of pro-oxidant compounds as expected, the probes in spectrally-compatible combination with other probes showed important increases or reductions in the fluorescence generation rate induced by the same prooxidant. The interpretation is complicated and requires further experiments, which may include more specific control systems for generating ROS as well as novel, single-cell based imaging technologies, such as multispectral-imaging flow cytometry [50].
5.3. Fluorescent Probe titration
A very important procedure when setting up a florescence-based determination in flow cytometry is to titrate properly the fluorescent probes, in order to define their optimal staining concentration for minimizing saturation and suboptimal detection of the relevant biological parameters [58].
The very first point to address in functional cytometry is to avoid or quantify the in-fluence of the probes on the experimental system. In the case of oxidative stress, all re-duced fluorogenic substrates are subject to auto-oxidation, which usually produces 1O2, O2.- and by its dismutation, H2O2. If the auto-oxidation rate is significant, it may result in artifactual detection of ROS and higher background, a problem especially important for probes such as HE [46]. The concentration of the probe is also relevant, as it may affect the stoichiometry of the process under study. In fact, the probes themselves may affect the activity of ROS-producing enzymes [53]. Finally, fluorescent probes at high concentration may be toxic to the cells [47].
To minimize artifacts derived from excessive probe concentration, fluorochrome titration is an essential procedure when setting up a florescence-based determination in flow cytometry, to define their optimal concentration for staining [58]. By defining the minimal concentration of a given fluorescent probe required for sensitive detection of a given ROS, the non-specific detection of ROS can be minimized, as demonstrated by the low level of intracellular fluorescence observed in cells not exposed to exogenous peroxides (Figure 4)
5.4. Range-finding experiments and Exclusion of dead/injured cells
A significant number of oxidative stress studies by FCM involve detection of ROS generation by in vitro exposure to xenobiotics [48]. In the Pharmaco-Toxicological context, it is important to quantify the toxic potency of the drug [215] and still the most used method for quantifying viable cells is the classical colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) [216]. However, this is a bulk method that provides only an average estimation of the viable-cell number based on the dehydrogenase activity ability of metabolically-competent cells. On the contrary, setting up a FCM analysis of cell function requires clear identification of single living cells in order to exclude cell aggregates and dead or dying cells [55,58]. Moreover, identifying live cells by FCM provides an alternative approach to assess the toxic potency of xenobiotics [48,217]. While the FCM procedures for single live-cell identification are relatively easy when analyzing human or mammalian cells [48,55,58], they become complicated for bacteria, mostly due to their smaller size and different permeability to viability dyes than in eukaryotic cells [126,218].
According to these aspects, performing a range-finding experiment for determining the toxicity (e.g. the IC50 value) of a test- or positive-control chemical is essential to establish toxic-exposure conditions that allow the presence of identificable and selectable live cells in the sample (Figure 4). This will require only including viability markers that are spectrally compatible with the other fluorescent probes that define the experiment [48,55,58,126,218]. In this way, the loss of intracellular fluorogenic substrates or fluorescent probes due to plasma membrane lesion may be detected and minimized, as commented in previous sections of this review [47,48,53].
5.6. Data generation, presentation, data analysis and publication
As for all the laboratory technologies, data generation in FCM is highly dependent on the quality issues that define the pre-analytical phase (i.e.; sample obtention, storage, handling and preparation) and the analytical phase (i.e.; Instrument control and optimization) should be carefully kept [47,48,53,56,58]. In addition, the special way in which data are generated from specific cell subpopulations (i.e.; single cells versus cell aggregates; live cells versus non-viable cells; cells with a particular phenotype or biological properties) requires that the procedures followed for such identification are clearly presented and justified in the post-analytical phase [47,48,58] in order to assess the reproducibility of the results and their interpretation. These considerations are at the basis of the publication policy of cytometry-specialized journals, which require that data suitable for independent reproduction of the results be made available prior to publication, and recommend that authors submit their data files to a repository [219] prior to submission. In addition, such journals require submitting a file with Minimum Information about a Flow Cytometry Experiment (MIFlowCyt Standard), that has been set by the International Society for the Advancement of Cytometry (ISAC). MIFlowCyt states the minimum information required to report FCM experiments to allow for future independent validation and interpretation of experimental data [220].
The issues related to the presentation of FCM data in the context of studies of ROS generation and oxidative stress are exemplified in Figure 4.
Figure 4A shows the strategy followed by us to identify single live cells, separated from debris and cell doublets (or aggregates). The scatter dotplot (FS Integral vs SS Integral) shows two populations of events, likely including cells and debris. A second scatter dotplot (FS Peak vs FS Integral) 2 shows the gate allowing to discriminate single cells from doublets/aggregates. The identification of live cells is confirmed with the vital probe DAPI, which stains live cells (panel 3) and with FS characteristics, by identifying dead or dying cells. Accordingly, all the analyses related to the use of ROS-sensitive fluorescent probes in FCM studies, including range-finding and titration experiments should be performed on events that were included simultaneously in the gate of “live cells” and “single cells”.
Finally, and based upon our own experience and the experimental evidences discussed in this review, in Table 2 we propose some recommendations for performing in vitro studies of Oxygen and Oxygen-related stress by FCM. Recommendations include the selection of the more appropriate fluorescent reagents and of suitable positive biological control,s as well as for the inclusion of viability markers in the experiments. Such recommendations are aimed to help to standardize and to improve the specificity and sensitivity of RONS analysis by FCM.
Author Contributions
Conceptualization, JEO; Investigation, BJ, GH, AMR, JEO; Writing – Original Draft Preparation, BJ, GH, AMR, JEO; Writing – Review & Editing, X BJ, GH, AMR, JEO; Supervision, JEO.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Taverne, Y.J.; Merkus, D.; Bogers, A.J.; Halliwell, B.; Duncker, D.J.; Lyons, T.W. Reactive Oxygen Species: Radical Factors in the Evolution of Animal Life. BioEssays. 2018, 40, 1700158. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Fasciolo, G.; Venditti, P. The Ambiguos Aspects of Oxygen. Oxygen. 2022, 2, 382–409. [Google Scholar] [CrossRef]
- Forman, H.J.; Augusto, O.; Brigelius-Flohe, R.; Dennery, P.A.; Kalyanaraman, B.; Ischiropoulos, H.; Mann, G.E.; Radi, R.; Roberts, L.J., 2nd; Vina, J.; et al. Even free radicals should follow some rules: A guide to free radical research terminology and methodology. Free Radic Biol Med. 2015, 78, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Sedlářová, M.; Pospíšil, P. Singlet oxygen imaging using fluorescent probe Singlet Oxygen Sensor Green in photosynthetic organisms. Sci Rep. 2018, 8, 13685. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid Med Cell Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Speckmann, B.; Steinbrenner, H.; Grune, T.; Klotz, L.O. Peroxynitrite: From interception to signaling. Arch Biochem Biophys. 2016, 595, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and Oxidative Stress: An Overview of Basic Concepts. Oxygen. 2022, 2, 437–478. [Google Scholar] [CrossRef]
- Clancy, D.; Birdsall, J. Flies, worms and the Free Radical Theory of ageing. Ageing Res Rev. 2013, 12, 404–412. [Google Scholar] [CrossRef]
- Hayashi, G.; Cortopassi, G. Oxidative stress in inherited mitochondrial diseases. Free Radic Biol Med. 2015, 88, 10–17. [Google Scholar] [CrossRef]
- Moulin, M.; Ferreiro, A. Muscle redox disturbances and oxidative stress as pathomechanisms and therapeutic targets in early-onset myopathies. Semin Cell Dev Biol. 2017, 64, 213–223. [Google Scholar] [CrossRef]
- Reula, A.; Pellicer, D.; Castillo, S.; Magallón, M.; Armengot, M.; Herrera, G.; O’Connor, J.E.; Bañuls, L.; Navarro-García, M.M.; Escribano, A.; et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. J Clin Med. 2021, 10, 1172. [Google Scholar] [CrossRef] [PubMed]
- Beltrán, B.; Nos, P.; Dasí, F.; Iborra, M.; Bastida, G.; Martínez, M.; O’Connor, J.E.; Sáez, G.; Moret, I.; Ponce, J. Mitochondrial dysfunction, persistent oxidative damage, and catalase inhibition in immune cells of naïve and treated Crohn’s disease. Inflamm Bowel Dis. 2010, 16, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int J Mol Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vascul Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Santilli, F.; D’Ardes, D.; Davì, G. Oxidative stress in chronic vascular disease: From prediction to prevention. Vascul Pharmacol. 2015, 74, 23–37. [Google Scholar] [CrossRef]
- Li, H.; Horke, S.; Förstermann, U. Vascular oxidative stress, nitric oxide and atherosclerosis. Atherosclerosis. 2014, 237, 208–219. [Google Scholar] [CrossRef]
- Montezano, A.C.; Dulak-Lis, M.; Tsiropoulou, S.; Harvey, A.; Briones, A.M.; Touyz, R.M. Oxidative stress and human hypertension: vascular mechanisms, biomarkers, and novel therapies. Can J Cardiol. 2015, 31, 631–641. [Google Scholar] [CrossRef]
- Fuentes, E.; Palomo, I. Role of oxidative stress on platelet hyperreactivity during aging. Life Sci. 2016, 148, 17–23. [Google Scholar] [CrossRef]
- Collado, R.; Ivars, D.; Oliver, I.; Tormos, C.; Egea, M.; Miguel, A.; Sáez, G.T.; Carbonell, F. Increased oxidative damage associated with unfavorable cytogenetic subgroups in chronic lymphocytic leukemia. Biomed Res Int. 2014, 2014, 686392. [Google Scholar] [CrossRef]
- Oh, B.; Figtree, G.; Costa, D.; Eade, T.; Hruby, G.; Lim, S.; Elfiky, A.; Martine, N.; Rosenthal, D.; Clarke, S.; et al. Oxidative stress in prostate cancer patients: A systematic review of case control studies. Prostate Int. 2016, 4, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wen, J.; Huang, Z.; Nice, E.C.; Huang, C.; Zhang, H.; Li, Q. Redox proteomics screening cellular factors associated with oxidative stress in hepatocarcinogenesis. Proteomics Clin Appl. 2017, 11, 1600089. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Nitti, M.; Furfaro, A.L.; Colla, R.; Ciucis, C.D.; Marinari, U.M.; Pronzato, M.A.; Traverso, N.; Domenicotti, C. Redox Homeostasis and Cellular Antioxidant Systems: Crucial Players in Cancer Growth and Therapy. Oxid Med Cell Longev. 2016, 2016, 6235641. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, D.; Zhelev, Z.; Aoki, I.; Bakalova, R.; Higashi, T. Overproduction of reactive oxygen species - obligatory or not for induction of apoptosis by anticancer drugs. Chin J Cancer Res. 2016, 28, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid Med Cell Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef] [PubMed]
- Elbim, C.; Pillet, S.; Prevost, M.H.; Preira, A.; Girard, P.M.; Rogine, N.; Hakim, J.; Israel, N.; Gougerot-Pocidalo, M.A. The role of phagocytes in HIV-related oxidative stress. J Clin Virol. 2001, 20, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Henchcliffe, C.; Beal, M. Mitochondrial biology and oxidative stress in Parkinson disease pathogenesis. Nat Clin Pract Neurol. 2008, 4, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.; Masters, C.; Busch, A.J. Neurodegenerative diseases and oxidative stress. Nat Rev Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef]
- Kamat, P.K.; Kalani, A.; Rai, S.; Swarnkar, S.; Tota, S.; Nath, C.; Tyagi, N. Mechanism of Oxidative Stress and Synapse Dysfunction in the Pathogenesis of Alzheimer’s Disease: Understanding the Therapeutics Strategies. Mol Neurobiol. 2016, 53, 648–661. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic Biol Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Dugas, B.; Debré, P.; Moncada, S. Nitric oxide, a vital poison inside the immune and inflammatory network. Res Immunol. 1995, 146, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Erusalimsky, J.D.; Moncada, S. Nitric oxide and mitochondrial signaling: from physiology to pathophysiology. Arterioscler Thromb Vasc Biol. 2007, 27, 2524–2531. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nature reviews. Molecular cell biology 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M. Priming of the neutrophil respiratory burst: role in host defense and inflammation. Immunol Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Burhans, W.C.; Heintz, N.H. The cell cycle is a redox cycle: linking phase-specific targets to cell fate. Free Radic Biol Med. 2009, 47, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Lionaki, E.; Markaki, M.; Tavernarakis, N. Autophagy and ageing: insights from invertebrate model organisms. Ageing Res Rev. 2013, 12, 413–428. [Google Scholar] [CrossRef]
- Gibellini, L.; De Biasi, S.; Pinti, M.; Nasi, M.; Riccio, M.; Carnevale, G.; Cavallini, G.M.; Sala de Oyanguren, F.J.; O’Connor, J.E.; Mussini, C.; et al. The protease inhibitor atazanavir triggers autophagy and mitophagy in human preadipocytes. AIDS. 2012, 26, 2017–2026. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat Chem Biol. 2011, 7, 504–511. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed Res Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Measuring reactive species and oxidative damage in vivo and in cell cutures: how should you do it and what do the results mean? Br J Pharmacol. 2004, 142, 231–252. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Use of spectroscopic probes for detection of reactive oxygen species. Clin Chim Acta. 2006, 368, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Sung, G.; Lin, J.M. Reactive oxygen species and their chemiluminescence-detection methods. Trends Anal Chem. 2006, 25, 985–995. [Google Scholar] [CrossRef]
- Maity, A. Photophysical Detection of Singlet Oxygen. In Reactive Oxygen Species; Ahmad, R., Ed.; Intechopen Limited: London, United Kingdom, 2022. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Singh, S.P.; Häder, D.-P.; Sinha, R.P. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2′,7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem Biophys Res Commun. 2010, 397, 603–607. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.E.; Callaghan, R.C.; Escudero, M.; Herrera, G.; Martínez, A.; Monteiro, M.C.; Montolíu, H. The relevance of flow cytometry for biochemical analysis. IUBMB Life 2001, 51, 231–239. [Google Scholar] [CrossRef]
- Herrera, G.; Diaz, L.; Martinez, A.; Gomes, A.; Villamón, E.; Callaghan, R.C.; O’Connor, J.E. Cytomics: A multiparametric, dynamic approach to cell research. Toxicol In Vitro. 2007, 21, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Barrientos, A.; O’Connor, J.E.; Nieto-Castillo, R.; Moreno-Moreno, A.B.; Prieto, P. Use of flow cytometry and confocal microscopy techniques to investigate early CdCl2-induced nephrotoxicity in vitro. Toxicol In Vitro. 2001, 15, 407–412. [Google Scholar] [CrossRef]
- Ploppa, A.; George, T.C.; Unertl, K.E.; Nohe, B.; Durieux, M.E. ImageStream cytometry extends the analysis of phagocytosis and oxidative burst. Scand J Clin Lab Invest. 2011, 71, 362–369. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J.; 2nd, Ischiropoulos, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free Radic Biol Med. 2012, 52, 1–6. [Google Scholar] [CrossRef]
- Debowska, K.; Debski, D.; Hardy, M.; Jakubowska, M.; Kalyanaraman, B.; Marcinek, A.; Michalski, R.; Michalowski, B.; Ouari, O.; Sikora, A.; et al. Toward selective detection of reactive oxygen and nitrogen species with the use of fluorogenic probes--Limitations, progress, and perspectives. Pharmacol Rep. 2015, 67, 756–764. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.E.; Herrera, G.; Sala-de-Oyanguren, F.; Jávega, B.; Martínez-Romero, A. Cytomics of oxidative stress: Probes and problems. In Single Cell Analysis: Contemporary Research and Clinical Applications, Robinson, J.; Cossarizza, A., Ed.; Springer: Singapore, 2017; pp. 83–118. [Google Scholar]
- Shapiro, H. Practical Flow Cytometry, 4th ed, John Wiley and Sons: Hoboken, NJ, USA, 2003.
- Ortolani, C. Flow Cytometry Today: Everything You Need to Know about Flow Cytometry; Springer Nature Switzerland: Cham, Switzerland, 2022. [Google Scholar]
- Robinson, J.P. Spectral flow cytometry—Quo vadimus? Cytometry 2019, 95, 823–824. [Google Scholar] [CrossRef] [PubMed]
- Schmid, I. (Ed.) Flow Cytometry: Recent Perspectives; Intechopen Limited: London, United Kingdom, 2012. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur J Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed]
- Ortolani, C. Flow Cytometry of Hematological Malignancies, 2nd ed.; Wiley Blackwell: Hoboken, NJ, USA, 2021. [Google Scholar]
- Robinson, J.P. Flow cytometry: past and future. Biotechniques. 2022, 72, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Spidlen, J.; Moore, W.; Parks, D.; Goldberg, M.; Bray, C.; Bierre, P.; Gorombey, P.; Hyun, B.; Hubbard, M.; Lange, S.; et al. Data File Standard for Flow Cytometry, version FCS 3.1. Cytometry A. 2010, 77, 97–100. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.E.; Herrera, G.; Corrochano, V. Flow versus Flux: Functional Assays by Flow Cytometry. In Fluorescence and Fluorescent Probes II, Slavík, J.; ed.; Plenum Press: New York, USA, 1998; pp. 47–54. [Google Scholar]
- Sun, G.; Qu, L.; Azi, F.; Liu, Y.; Li, J.; Lv, X.; Du, G.; Chen, J.; Chen, C.H.; Liu, L. Recent progress in high-throughput droplet screening and sorting for bioanalysis. Biosens Bioelectron. 2023, 225, 115107. [Google Scholar] [CrossRef] [PubMed]
- Elbim, C.; Lizard, G. Flow cytometric investigation of neutrophil oxidative burst and apoptosis in physiological and pathological situations. Cytometry A. 2009, 75, 475–481. [Google Scholar] [CrossRef]
- De Gaetano, A.; Gibellini, L.; Zanini, G.; Nasi, M.; Cossarizza, A.; Pinti, M. Mitophagy and Oxidative Stress: The Role of Aging. Antioxidants 2021, 10, 794. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cossarizza, A. Interfering with ROS Metabolism in Cancer Cells: The Potential Role of Quercetin. Cancers (Basel). 2010, 2, 1288–1311. [Google Scholar] [CrossRef]
- Petit, P.X.; Ardilla-Osorio, H.; Penalvia, L.; Rainey, N.E. Tafazzin Mutation Affecting Cardiolipin Leads to Increased Mitochondrial Superoxide Anions and Mitophagy Inhibition in Barth Syndrome. Cells. 2020, 9, 2333. [Google Scholar] [CrossRef]
- Ivars, D.; Orero, M.T.; Javier, K.; Díaz-Vico, L.; García-Giménez, J.L.; Mena, S.; Tormos, C.; Egea, M.; Pérez, P.L.; Arrizabalaga, B.; et al. Oxidative imbalance in low/intermediate-1-risk myelodysplastic syndrome patients: The influence of iron overload. Clin Biochem. 2017, 50, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, V.; Jávega, B.; Gasperini, S.; O’Connor, J.E.; Lenzi, M.; Hrelia, P. 6-(Methylsulfonyl) Hexyl Isothiocyanate: A Chemopreventive Agent Inducing Autophagy in Leukemia Cell Lines. Biomolecules. 2022, 12, 1485. [Google Scholar] [CrossRef] [PubMed]
- Urios, A.; López-Gresa, M.P.; González, M.C.; Primo, J.; Martínez, A.; Herrera, G.; Escudero, J.C.; O’Connor, J.E.; Blanco, M. Nitric oxide promotes strong cytotoxicity of phenolic compounds against Escherichia coli: the influence of antioxidant defenses. Free Radic Biol Med. 2003, 35, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K. Basic Biology of Hypoxic Responses Mediated by the Transcription Factor HIFs and its Implication for Medicine. Biomedicines 2020, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Godet, I.; Doctorman, S.; Wu, F.; Gilkes, D.M. Detection of Hypoxia in Cancer Models: Significance, Challenges, and Advances. Cells 2022, 11, 686. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Shrivastava, V.; Joshi, D.; Singal, C.M.S.; Tyagi, S.; Bhat, M.A.; Jaiswal, P.; Sharma, J.B.; Palanichamy, J.K.; Sinha, S.; et al. Hypoxia Induces Early Neurogenesis in Human Fetal Neural Stem Cells by Activating the WNT Pathway. Mol Neurobiol. 2023, 60, 2910–2921. [Google Scholar] [CrossRef] [PubMed]
- Pachón-Peña, G.; Serena, C.; Ejarque, M.; Petriz, J.; Duran, X.; Oliva-Olivera, W.; Simó, R.; Tinahones, F.J.; Fernández-Veledo, S.; Vendrell, J. Obesity Determines the Immunophenotypic Profile and Functional Characteristics of Human Mesenchymal Stem Cells From Adipose Tissue. Stem Cells Transl Med. 2016, 5, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Espinosa, C.; Ferreira, I.; Ríos-Kristjánsson, J.G.; Rizo-Roca, D.; García Godoy, M.D.; Rico, L.G.; Rubi-Sans, G.; Torrella, J.R.; Pagès, T.; Petriz, J.; et al. Effects of intermittent hypoxia and light aerobic exercise on circulating stem cells and side population, after strenuous eccentric exercise in trained rats. Curr Stem Cell Res Ther. 2015, 10, 132–139. [Google Scholar] [CrossRef]
- Synowiec, A.; Brodaczewska, K.; Wcisło, G.; Majewska, A.; Borkowska, A.; Filipiak-Duliban, A.; Gawrylak, A.; Wilkus, K.; Piwocka, K.; Kominek, A.; et al. Hypoxia, but Not Normoxia, Reduces Effects of Resveratrol on Cisplatin Treatment in A2780 Ovarian Cancer Cells: A Challenge for Resveratrol Use in Anticancer Adjuvant Cisplatin Therapy. Int J Mol Sci. 2023, 24, 5715. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Jin, Y.; Lin, J.; Gong, L.; Xu, Y. Hypoxia upregulates the expression of lncRNA H19 in non-small cell lung cancer cells and induces drug resistance. Transl Cancer Res. 2022, 11, 2876–2886. [Google Scholar] [CrossRef]
- Such, L.; O’Connor, J.E.; Sáez, G.T.; Gil, F.; Beltrán, J.F.; Moya, A.; Alberola, A. Flow cytometric analysis of peroxidative activity in granulocytes from coronary and peripheral blood in acute myocardial ischemia and reperfusion in dogs: protective effect of methionine. Cytometry. 1999, 37, 140–146. [Google Scholar] [CrossRef]
- Chen, G.; Yang, Y.; Xu, C.; Gao, S. A Flow Cytometry-based Assay for Measuring Mitochondrial Membrane Potential in Cardiac Myocytes After Hypoxia/Reoxygenation. J Vis Exp. 2018, 137, 57725. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.; Hao, Y.; Hou, D.; Yang, R. Lycium barbarum polysaccharide protects cardiomyocytes from hypoxia/reoxygenation injury via activation of SIRT3/CypD signaling. Ann Transl Med. 2023, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; He, W.; Xia, J.; Huang, Q.; Yang, J.; Gu, W.P.; Zhang, N.; Liu, Y.H. Human umbilical cord mesenchymal stem cells-derived exosomal circDLGAP4 promotes angiogenesis after cerebral ischemia-reperfusion injury by regulating miR-320/KLF5 axis. FASEB J. 2023, 37, e22733. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Wang, M.; Cui, M.; Fang, Y.; Li, H.; Zheng, C.; Li, Z.; Xu, Y.; Hua, H.; Li, D. Small-molecule probes for fluorescent detection of cellular hypoxia-related nitroreductase. J Pharm Biomed Anal. 2021, 203, 114199. [Google Scholar] [CrossRef] [PubMed]
- Wallabregue, A.L.D.; Bolland, H.; Faulkner, S.; Hammond, E.M.; Conway, S.J. Two Color Imaging of Different Hypoxia Levels in Cancer Cells. J Am Chem Soc. 2023, 145, 2572–2583. [Google Scholar] [CrossRef] [PubMed]
- Nordsmark, M.; Loncaster, J.; Aquino-Parsons, C.; Chou, S.C.; Ladekarl, M.; Havsteen, H.; Lindegaard, J.C.; Davidson, S.E.; Varia, M.; West, C.; et al. Measurements of hypoxia using pimonidazole and polarographic oxygen-sensitive electrodes in human cervix carcinomas. Radiother Oncol. 2003, 67, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.J. Measurement of absolute oxygen levels in cells and tissues using oxygen sensors and 2-nitroimidazole EF5. Methods Enzymol. 2002, 352, 3–31. [Google Scholar] [CrossRef]
- Nakajima, K.; Homma, M.; Suzuki, M.; Yokouchi, Y.; Matsuda, T.; Takakura, H.; Hirata, K.; Kuge, Y.; Ogawa, M. Reduction of tumor hypoxia by anti-PD-1 therapy assessed using pimonidazole and [18F]FMISO. Nucl Med Biol. 2022, 108-109, 85–92. [Google Scholar] [CrossRef]
- Gravelle, P.; Jean, C.; Familiades, J.; Decaup, E.; Blanc, A.; Bezombes-Cagnac, C.; Laurent, C.; Savina, A.; Fournié, J.J.; Laurent, G. Cell growth in aggregates determines gene expression, proliferation, survival, chemoresistance, and sensitivity to immune effectors in follicular lymphoma. Am J Pathol. 2014, 184, 282–295. [Google Scholar] [CrossRef]
- Chang, Q.; Ornatsky, O.I.; Koch, C.J.; Chaudary, N.; Marie-Egyptienne, D.T.; Hill, R.P.; Tanner, S.D.; Hedley, D.W. Single-cell measurement of the uptake, intratumoral distribution and cell cycle effects of cisplatin using mass cytometry. Int J Cancer. 2015, 136, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Document Connect (thermofisher.com). Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0017632_Hypoxia_Green_for_Flow_Cytometry_UG.pdf (accessed on 27 April 2023).
- BioTracker 520 Green Hypoxia Dye Live Cell Imaging Millipore (sigmaaldrich.com). Available online: https://www.sigmaaldrich.com/ES/es/product/mm/sct033 (accessed on 27 April 2023).
- Document Connect (thermofisher.com). Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0013497_Image_iT_Hypoxia_Reagents_UG.pdf (accessed on 27 April 2023).
- Zhang, S.; Hosaka, M.; Yoshihara, T.; Negishi, K.; Iida, Y.; Tobita, S.; Takeuchi, T. Phosphorescent light–emitting iridium complexes serve as a hypoxia-sensing probe for tumor imaging in living animals. Cancer Res. 2010, 70, 4490–4498. [Google Scholar] [CrossRef] [PubMed]
- Vordermark, D.; Shibata, T.; Brown, J.M. Green fluorescent protein is a suitable reporter of tumor hypoxia despite an oxygen requirement for chromophore formation. Neoplasia. 2001, 3, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Gately, D.P.; Hom, D.; Mishima, M.; Los, G.; Howell, S.B. Quantification of tumor cell injury in vitro and in vivo using expression of green fluorescent protein under the control of the GADD153 promoter. Int J Cancer. 2001, 91, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Erapaneedi, R.; Belousov, V.V.; Schäfers, M.; Kiefer, F. A novel family of fluorescent hypoxia sensors reveal strong heterogeneity in tumor hypoxia at the cellular level. EMBO J. 2016, 35, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, A.; Huang, L.; Zou, Y.; Gu, Y.; Chen, X.; Zhao, Y.; Yang, Y. Monitoring cellular redox state under hypoxia using a fluorescent sensor based on eel fluorescent protein. Free Radic Biol Med. 2018, 120, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Miura, T.; Umezawa, N.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T.; Rational design of fluorescein-based fluorescence probes. Mechanism-based design of a maximum fluorescence probe for singlet oxygen. J Am Chem Soc. 2001, 123, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Ragas, X.; Jimenez-Banzo, A.; Sanchez-Garcia, D.; Batllori, X.; Nonell, S. Singlet oxygen photosensitisation by the fluorescent probe Singlet Oxygen Sensor Green. Chem Commun (Camb). 2009, 20, 2920–2922. [Google Scholar] [CrossRef]
- Song, D.; Cho, S.; Han, Y.; You, Y.; Nam, W. Ratiometric Fluorescent Probes for Detection of Intracellular Singlet Oxygen. Org Lett. 2013, 15, 3582–3585. [Google Scholar] [CrossRef]
- Umezawa, N.; Tanaka, K.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Novel Fluorescent Probes for Singlet Oxygen. Angew Chem Int Ed Engl 1999, 38, 2899–2901. [Google Scholar] [CrossRef]
- Hideg, E.; Kalai, T.; Kos, P.B.; Asada, K.; Hideg, K. Singlet oxygen in plants- Its significance and possible detection with double (fluorescent and spin) indicator reagents. Photochem Photobiol. 2006, 82, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Gollmer, A.; Arnbjerg, J.; Blaikie, F.H.; Pedersen, B.W.; Breitenbach, T.; Daasbjerg, K.; Glasius, M.; Ogilby, P.R. Singlet Oxygen Sensor Green®: photochemical behavior in solution and in a mammalian cell. Photochem Photobiol. 2011, 87, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Fujitsuka, M.; Majima, T. Photochemistry of Singlet Oxygen Sensor Green. J Phys Chem B. 2013, 117, 13985–13992. [Google Scholar] [CrossRef] [PubMed]
- Flors, C.; Fryer, M.J.; Waring, J.; Reeder, B.; Bechtold, U.; Mullineaux, P.M.; Nonell, S.; Wilson, M.T.; Baker, N.R. Imaging the production of singlet oxygen in vivo using a new fluorescent sensor, Singlet Oxygen Sensor Green®. J Exp Bot. 2006, 57, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Carter, P.J.H.; Laan, A.C.; Eelkema, R.; Denkova, A.G. Singlet Oxygen Sensor Green is not a Suitable Probe for 1O2 in the Presence of Ionizing Radiation. Sci Rep. 2019, 9, 8393. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.K.; Holmehave, J.; Blaikie, F.H.; Gollmer, A.; Breitenbach, T.; Jensen, H.H.; Ogilby, P.R. Aarhus Sensor Green: A Fluorescent Probe for Singlet Oxygen. J Org Chem. 2014, 79, 3079–3087. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Fernandes, E.; Lima, J.L. Fluorescence probes used for detection of reactive oxygen species. J Biochem Biophys Methods. 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Wardman, P. Fluorescent and luminescent probes for measurement of oxidative and nitrosative species in cells and tissues: progress, pitfalls, and prospects. Free Radic Biol Med. 2007, 43, 995–1022. [Google Scholar] [CrossRef]
- Keston, A.S.; Brandt, R. The fluorometric analysis of ultramicro quantities of hydrogen peroxide. Anal Biochem. 1965, 11, 1–5. [Google Scholar] [CrossRef]
- van Eeden, S.F.; Klut, M.E.; Walker, B.A.; Hogg, J.C. The use of flow cytometry to measure neutrophil function. J Immunol Methods. 1999, 232, 23–43. [Google Scholar] [CrossRef]
- Caldefie-Chézet, F.; Walrand, S.; Moinard, C.; Tridon, A.; Chassagne, J.; Vasson, M.P. Is the neutrophil reactive oxygen species production measured by luminol and lucigenin chemiluminescence intra or extracellular? Comparison with DCFH-DA flow cytometry and cytochrome c reduction. Clin Chim Acta 2002, 319, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Bourré, L.; Thibaut, S.; Briffaud, A.; Rousset, N.; Eléouet, S.; Lajat, Y.; Patrice, T. Indirect detection of photosensitizer ex vivo. J Photochem Photobiol, B Biol 2002, 67, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Silveira, L.R.; Pereira-Da-Silva, L.; Juel, C.; Hellsten, Y. Formation of hydrogen peroxide and nitric oxide in rat skeletal muscle cells during contractions. Free Radic Biol Med. 2003, 35, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Tampo, Y.; Kotamraju, S.; Chitambar, C.R.; Kalivendi, S.V.; Keszler, A.; Joseph, J.; Kalyanaraman, B. Oxidative stress-induced iron signaling is responsible for peroxide-dependent oxidation of dichlorodihydrofluorescein in endothelial cells: role of transferrin receptor-dependent iron uptake in apoptosis. Circ Res. 2003, 92, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kotamraju, S.; Tampo, Y.; Keszler, A.; Chitambar, C.R.; Joseph, J.; Haas, A.L.; Kalyanaraman, B. Nitric oxide inhibits H2O2-induced transferrin receptor-dependent apoptosis in endothelial cells: role of ubiquitin–proteasome pathway. Proc Natl Acad Sci U S A. 2003, 100, 10653–10658. [Google Scholar] [CrossRef] [PubMed]
- Kotamraju, S.; Kalivendi, S.V.; Konorev, E.; Chitambar, C.R.; Joseph, J.; Kalyanaraman, B. Oxidant induced iron signaling in doxorubicin-mediated apoptosis. Methods Enzymol. 2004, 378, 362–382. [Google Scholar] [CrossRef] [PubMed]
- Probes for Reactive Oxygen Species, Including Nitric Oxide—Chapter 18 | Thermo Fisher Scientific – ES. Available online: https://www.thermofisher.com/es/es/home/references/molecular-probes-the-handbook/probes-for-reactive-oxygen-species-including-nitric-oxide.html (accessed on 27 April 2023).
- Crow, J.P. Dichlorodihydrofluorescein and dihydrorhodamine 123 are sensitive indicators of peroxynitrite in vitro: implications for intracellular measurement of reactive nitrogen and oxygen species. Nitric Oxide. 1997, 1, 145–157. [Google Scholar] [CrossRef]
- Kooy, N.W.; Royall, J.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite mediated oxidation of dihydrorhodamine 123. Free Radic Biol Med. 1994, 16, 149–156. [Google Scholar] [CrossRef]
- Kooy, N.W.; Royall, J.A.; Ischiropoulos, H. Oxidation of 2,7-dichlorofluorescin by peroxynitrite. Free Radic Res. 1997, 27, 245–254. [Google Scholar] [CrossRef]
- Miller, E.W.; Albers, A.E.; Pralle, A.; Isacoff, E.Y.; Chang, C.J. Boronate-based fluorescent probes for imaging cellular hydrogen peroxide. J Am Chem Soc. 2005, 127, 16652–16659. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Huynh, C.; Chang, C.J. A palette of fluorescent probes with varying emission colors for imaging hydrogen peroxide signaling in living cells. J Am Chem Soc. 2010, 132, 5906–5915. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Aleyasin, H.; Dickinson, B.C.; Haskew-Layton, R.E.; Ratan, R.R. Recent advances in hydrogen peroxide imaging for biological applications. Cell Biosci. 2014, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Aleyasin, H.; Howard, S.S.; Dickinson, B.C.; Lin, V.S.; Haskew-Layton, R.E.; Xu, C.; Chen, Y.; Ratan, R.R. Two-photon fluorescence imaging of intracellular hydrogen peroxide with chemoselective fluorescent probes. J Biomed Opt. 2013, 18, 106002. [Google Scholar] [CrossRef] [PubMed]
- Albers, A.E.; Okreglak, V.S.; Chang, C.J. A FRET-based approach to ratiometric fluorescence detection of hydrogen peroxide. J Am Chem Soc. 2006, 128, 9640–9641. [Google Scholar] [CrossRef] [PubMed]
- Jávega, B.; Herrera, G.; O’Connor, J.E. Flow Cytometric Analysis of Oxidative Stress in Escherichia coli B Strains Deficient in Genes of the Antioxidant Defence. Int J Mol Sci. 2022, 10, 6537. [Google Scholar] [CrossRef] [PubMed]
- Benov, L.; Sztejnberg, L.; Fridovich, I. Critical evaluation of the use of hydroethidine as a measure of superoxide anion radical. Free Radic Biol Med 1998, 25, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Rothe, G.; Valet, G. Flow cytometric analysis of respiratory burst activity in phagocytes with hydroethidine and 2,7-dichlorofluorescin. J Leukoc Biol. 47:440-448.
- Walrand, S.; Valeix, S.; Rodriguez, C.; Ligot, P.; Chassagne, J.; Vasson, M.P. Flow cytometry study of polymorphonuclear neutrophil oxidative burst: a comparison of three fluorescent probes. Clin Chim Acta. 2003, 331, 103–110. [Google Scholar] [CrossRef]
- Carter, W.O.; Narayanan, P.K.; Robinson, J.P. Intracellular hydrogen peroxide and superoxide anion detection in endothelial cells. J Leukoc Biol. 1994, 55, 253–258. [Google Scholar] [CrossRef]
- Barbacanne, M.A.; Souchard, J.P.; Darblade, B.; Iliou, J.P.; Nepveu, F.; Pipy, B.; Bayard, F.; Arnal, J.F. Detection of superoxide anion released extracellularly by endothelial cells using cytochrome c reduction, ESR, fluorescence and lucigenin-enhanced chemiluminescence techniques. Free Radic Biol Med. 2000, 29, 388–396. [Google Scholar] [CrossRef]
- Münzel, T.; Afanas’ev, I.B.; Kleschyov, A.L.; Harrison, D.G. Detection of superoxide in vascular tissue. Arterioscler Thromb Vasc Biol. 2002, 22, 1761–1768. [Google Scholar] [CrossRef]
- Tarpey, M.M.; Wink, D.A.; Grisham, M.B. Methods for detection of reactive metabolites of oxygen and nitrogen: in vitro and in vivo considerations. Am J Physiol Regul Integr Comp Physiol. 2004, 286, R431–R444. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.L.; Miller, M.A.; Shapiro, I.M.; Shenker, B.J. Mercuric chloride induces apoptosis in human T lymphocytes: evidence of mitochondrial dysfunction. Toxicol Appl Pharmacol. 1998, 153, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Le, S.B.; Hailer, M.K.; Buhrow, S.; Wang, Q.; Flatten, K.; Pediaditakis, P.; Bible, K.C.; Lewis, L.D.; Sausville, E.A.; Pang, Y.P.; et al. Inhibition of mitochondrial respiration as a source of adaphostin-induced reactive oxygen species and cytotoxicity. J Biol Chem. 2007, 282, 8860–8872. [Google Scholar] [CrossRef] [PubMed]
- De Biasi, S.; Gibellini, L.; Bianchini, E.; Nasi, M.; Pinti, M.; Salvioli, S.; Cossarizza, A. Quantification of mitochondrial reactive oxygen species in living cells by using multi-laser polychromatic flow cytometry. Cytometry A. 2016, 89, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.M.; Janes, M.S.; Beckman, J.S. The selective detection of mitochondrial superoxide by live cell imaging. Nat Protoc. 2008, 3, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Kalyanaraman, B. Hydroethidine- and MitoSOX-derived red fluorescence is not a reliable indicator of intracellular superoxide formation: another inconvenient truth. Free Radic Biol Med. 2010, 48, 983–1001. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, G.; Yang, T.; Gan, J.; Xu, L.; Yang, H. A flow-cytometry-based protocol for detection of mitochondrial ROS production under hypoxia. STAR Protoc. 2021, 2, 100466. [Google Scholar] [CrossRef]
- Ahn, H.Y.; Fairfull-Smith, K.E.; Morrow, B.J.; Lussini, V.; Kim, B.; Bondar, M.V.; Bottle, S.E.; Belfield, K.D. Two-photon fluorescence microscopy imaging of cellular oxidative stress using profluorescent nitroxides. J Am Chem Soc. 2012, 134, 4721–4730. [Google Scholar] [CrossRef]
- Davison, C.A.; Durbin, S.M.; Thau, M.R.; Zellmer, V.R.; Chapman, S.E.; Diener, J.; Wathen, C.; Leevy, W.M.; Schafer, Z.T. Antioxidant Enzymes Mediate Survival of Breast Cancer Cells Deprived of Extracellular Matrix. Cancer Res. 2013, 73, 3704–3715. [Google Scholar] [CrossRef]
- DeLoughery, Z.; Luczak, M.W.; Zhitkovich, A. Monitoring Cr intermediates and reactive oxygen species with fluorescent probes during chromate reduction. Chem Res Toxicol. 2014, 27, 843–851. [Google Scholar] [CrossRef]
- Plaza Davila, M.; Martin Muñoz, P.; Tapia, J.A.; Ortega Ferrusola, C.; Balao da Silva, C.C.; Peña, F.J. Inhibition of Mitochondrial Complex I Leads to Decreased Motility and Membrane Integrity Related to Increased Hydrogen Peroxide and Reduced ATP Production, while the Inhibition of Glycolysis Has Less Impact on Sperm Motility. PLoS One 2015, 10, e0138777. [Google Scholar] [CrossRef] [PubMed]
- mp10422.pdf (fishersci.com). Available online: https://assets.fishersci.com/TFS-Assets/LSG/manuals/mp10422.pdf?_ga=2.231040351.1726096505.1681672212-1827320652.1681672212 (accessed on 27 April 2023).
- ROS-ID® Total ROS/Superoxide detection kit - ENZ-51010 - Enzo Life Sciences. Available online: https://www.enzolifesciences.com/ENZ-51010/ros-id-total-ros-superoxide-detection-kit/ (accessed on 27 April 2023).
- Egawa, N.; Tanaka, T.; Matsufuji, S.; Yamada, K.; Ito, K.; Kitagawa, H.; Okuyama, K.; Kitajima, Y.; Noshiro, H. Antitumor effects of low-dose tipifarnib on the mTOR signaling pathway and reactive oxygen species production in HIF-1α-expressing gastric cancer cells. FEBS Open Bio. 2021, 11, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Liu, S.; Wang, X.; Deng, X.; Fan, Y.; Sun, C.; Wang, Y.; Mehta, J.L. Hemodynamic Shear Stress via ROS Modulates PCSK9 Expression in Human Vascular Endothelial and Smooth Muscle Cells and Along the Mouse Aorta. Antioxid Redox Signal. 2015, 22, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, F.A.; van den Berg, J.J.; Schalkwijk, C.; Roelofsen, B.; Op den Kamp, J.A. Parinaric acid as a sensitive fluorescent probe for the determination of lipid peroxidation. Biochim Biophys Acta. 1987, 921, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Hedley, D.; Chow, S. Flow cytometric measurement of lipid peroxidation in vital cells using parinaric acid. Cytometry. 1992, 13, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Drummen, G.P.; Makkinje, M.; Verkleij, A.J.; Op den Kamp, J.A.; Post, J.A. Attenuation of lipid peroxidation by antioxidants in rat-1 fibroblasts: comparison of the lipid peroxidation reporter molecules cis-parinaric acid and C11-BODIPY(581/591) in a biological setting. Biochim Biophys Acta. 2004, 1636, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Shimakawa, S.; Itoh, N.; Niki, E. Action of DCFH and BODIPY as a probe for radical oxidation in hydrophilic and lipophilic domain. Free Radic Res. 2003, 37, 861–872. [Google Scholar] [CrossRef]
- Brouwers, J.F.; Gadella, B. M In situ detection and localization of lipid peroxidation in individual bovine sperm cells. Free Radic Biol Med. 2003, 35, 1382–1391. [Google Scholar] [CrossRef]
- Cheloni, G.; Slaveykova, V.I. Optimization of the C11-BODIPY(581/591) dye for the determination of lipid oxidation in Chlamydomonas reinhardtii by flow cytometry. Cytometry A. 2013, 83, 952–961. [Google Scholar] [CrossRef]
- Peluso, I.; Adorno, G.; Raguzzini, A.; Urban, L.; Ghiselli, A.; Serafini, M. A new flow cytometry method to measure oxidative status: the Peroxidation of Leukocytes Index Ratio (PLIR). J Immunol Methods. 2013, 390, 113–120. [Google Scholar] [CrossRef]
- Donato, M.T.; Martínez-Romero, A.; Jiménez, N.; Negro, A.; Herrera, G.; Castell, J.V.; O’Connor, J.E.; Gómez-Lechón, M.J. Cytometric analysis for drug-induced steatosis in HepG2 cells. Chem Biol Interact. 2009, 181, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Makrigiorgos, G.M.; Kassis, A.I.; Mahmood, A.; Bump, E.A.; Savvides, P. Novel fluorescein-based flow–cytometric method for detection of lipid peroxidation. Free Radic Biol Med. 1997, 22, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Maulik, G.; Kassis, A.I.; Savvides, P.; Makrigiorgos, G.M. Fluoresceinated phosphoethanolamine for flow–cytometric measurement of lipid peroxidation. Free Radic Biol Med. 1998, 26, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Blair, I.A. Characterization of 4-oxo-2-nonenal as a novel product of lipid peroxidation. Chem Res Toxicol. 2000, 13, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Csala, M.; Kardon, T.; Legeza, B.; Lizák, B.; Mandl, J.; Margittai, É.; Puskás, F.; Száraz, P.; Szelényi, P.; Bánhegyi, G. On the role of 4-hydroxynonenal in health and disease. Biochim Biophys Acta. 2015, 1852, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Miyake, N.; Hiai, H.; Hagiwara, M.; Kawakishi, S.; Osawa, T.; Uchida, K. The monoclonal antibody specific for the 4-hydroxy-2-nonenal histidine adduct. FEBS Lett. 1995, 359, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Martin Muñoz, P.; Ortega Ferrusola, C.; Vizuete, G.; Plaza Dávila, M.; Rodriguez Martinez, H.; Peña, F.J. Depletion of Intracellular Thiols and Increased Production of 4-Hydroxynonenal that Occur During Cryopreservation of Stallion Spermatozoa Lead to Caspase Activation, Loss of Motility, and Cell Death. Biol Reprod. 2015, 93, 143. [Google Scholar] [CrossRef]
- Fortini, P.; Pascucci, B.; Parlanti, E.; D’Errico, M.; Simonelli, V.; Dogliotti, E. 8-Oxoguanine DNA damage: at the crossroad of alternative repair pathways. Mutat Res. 2003, 531, 127–139. [Google Scholar] [CrossRef]
- Barregard, L.; Møller, P.; Henriksen, T.; Mistry, V.; Koppen, G.; Rossner, P. ; Jr, Sram, R. J.; Weimann, A.; Poulsen, H.E.; Nataf, R.; et al. Human and methodological sources of variability in the measurement of urinary 8-oxo-7,8-dihydro-2’-deoxyguanosine. Antioxid Redox Signal. 2013, 18, 2377–2391. [Google Scholar] [CrossRef]
- Oxy dna assay kit | Sigma-Aldrich (sigmaaldrich.com). Available online: https://www.sigmaaldrich.com/ES/en/search/oxy-dna-assay-kit?focus=products&page=1&perpage=30&sort=relevance&term=oxy%20dna%20assay%20kit&type=product (accessed on 27 April 2023).
- Nagy, S.; Kakasi, B.; Bercsényi, M. Flow cytometric detection of oxidative DNA damage in fish spermatozoa exposed to cadmium—short communication. Acta Vet Hung. 2016, 64, 120–124. [Google Scholar] [CrossRef]
- Esperanza, M.; Cid, Á.; Herrero, C.; Rioboo, C. Acute effects of a prooxidant herbicide on the microalga Chlamydomonas reinhardtii: Screening cytotoxicity and genotoxicity endpoints. Aquat Toxicol. 2015, 165, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Cambi, M.; Tamburrino, L.; Marchiani, S.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E.; Muratori, M. Development of a specific method to evaluate 8-hydroxy, 2-deoxyguanosine in sperm nuclei: relationship with semen quality in a cohort of 94 subjects. Reproduction. 2013, 145, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Meseguer, M.; García-Herrero, S.; Gil-Salom, M.; O’Connor, J.E.; Garrido, N. Relevance of testicular sperm DNA oxidation for the outcome of ovum donation cycles. Fertil Steril. 2010, 94, 979–988. [Google Scholar] [CrossRef]
- Meseguer, M.; Martínez-Conejero, J.A.; O’Connor, J.E.; Pellicer, A.; Remohí, J.; Garrido, N. The significance of sperm DNA oxidation in embryo development and reproductive outcome in an oocyte donation program: a new model to study a male infertility prognostic factor. Fertil Steril. 2008, 89, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Balao da Silva, C.M.; Ortega-Ferrusola, C.; Morrell, J.M.; Rodriguez Martínez, H.; Peña, F.J. Flow Cytometric Chromosomal Sex Sorting of Stallion Spermatozoa Induces Oxidative Stress on Mitochondria and Genomic DNA. Reprod Domest Anim. 2016, 51, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Durand, R.E.; Olive, P.L. Flow cytometry techniques for studying cellular thiols. Radiat Res 1983, 95, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Treumer, J.; Valet, G. Flow-cytometric determination of glutathione alterations in vital cells by o-phthaldialdehyde (OPT) staining. Exp Cell Res. 1986, 163, 518–524. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.E.; Kimler, B.F.; Morgan, M.C.; Tempas, K.J. A flow cytometric assay for intracellular nonprotein thiols using mercury orange. Cytometry. 1988, 9, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Cell Viability Assays | Thermo Fisher Scientific – ES. Availble online: https://www.thermofisher.com/es/es/home/life-science/cell-analysis/cell-viability-and-regulation/cell-viability. (accessed on 27 April 2023).
- Nair, S.; Singh, S.V.; Krishan, A. Flow cytometric monitoring of glutathione content and anthracycline retention in tumor cells. Cytometry 1991, 12, 336–342. [Google Scholar] [CrossRef]
- Hedley, D.W.; Chow, S. Evaluation of methods for measuring cellular glutathione content using flow cytometry. Cytometry. 1994, 15, 349–358. [Google Scholar] [CrossRef]
- Chow, S.; Hedley, D. Flow cytometric determination of glutathione in clinical samples. Cytometry. 1995, 21, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Skindersoe, M.E.; Kjaerulff, S. Comparison of three thiol probes for determination of apoptosis-related changes in cellular redox status. Cytometry A. 2014, 85, 179–187. [Google Scholar] [CrossRef]
- Keller, A.; Mohamed, A.; Drose, S.; Brandt, U.; Fleming, I.; Brandes, R.P. Analysis of dichlorodihydrofluorescein and dihydrocalcein as probes for the detection of intracellular reactive oxygen species. Free Radic Res. 2004, 38, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Swift, L.M.; Sarvazyan, N. Localization of dichlorofluorescin in cardiac myocytes: implications for assessment of oxidative stress. Am J Physiol Heart Circ Physiol. [CrossRef]
- Balaguer, S.; Diaz, L.; Gomes, A.; Herrera, G.; O’Connor, J.E.; Urios, A.; Felipo, V.; Montoliu, C. Real-time cytometric assay of nitric oxide and superoxide interaction in peripheral blood monocytes: A no-wash, no-lyse kinetic method. Cytometry B Clin Cytom. 2017, 92, 211–217. [Google Scholar] [CrossRef]
- Saengkhae, C.; Loetchutinat, C.; Garnier-Suillerot, A. Kinetic analysis of fluorescein and dihydrofluorescein effluxes in tumour cells expressing the multidrug resistance protein, MRP1. Biochem Pharmacol. 2003, 65, 969–977. [Google Scholar] [CrossRef]
- Grzelak, A.; Rychlik, B.; Bartosz, G. Reactive oxygen species are formed in cell culture media. Acta Biochim Pol. 2000, 47, 47,1197–1198. [Google Scholar] [PubMed]
- Petasne, R.G.; Zika, R.G. Fate of superoxide in coastal sea water. Nature. 1987, 325, 516–518. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Rapp, I.; Schlosser, C.; Achterberg, E.P. Hydrogen peroxide in deep waters from the Mediterranean Sea, South Atlantic and South Pacific Oceans. Sci Rep. 2017, 7, 43436. [Google Scholar] [CrossRef]
- Subramaniam, R.; Fan, X.J.; Scivittaro, V.; Yang, J.; Ha, C.E.; Petersen, C.E.; Surewicz, W.K.; Bhagavan, N.V.; Weiss, M.F.; Monnier, V.M. Cellular oxidant stress and advanced glycation endproducts of albumin: caveats of the dichlorofluorescein assay. Arch Biochem Biophys. 2002, 400, 15–25. [Google Scholar] [CrossRef]
- Chignell, C.F.; Sik, R.H. A photochemical study of cells loaded with 2,7-dichlorofluorescin: implications for the detection of reactive oxygen species generated during UVA irradiation. Free Radic Biol Med. 2003, 34, 1029–1034. [Google Scholar] [CrossRef]
- Sikora, A.; Zielonka, J.; Lopez, M.; Joseph, J.; Kalyanaraman, B. Direct oxidation of boronates by peroxynitrite: mechanism and implications in fluorescence imaging of peroxynitrite. Free Radic Biol Med. 2009, 47, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Kalivendi, S.; Zhang, H.; Joseph, J.; Nithipatikom, K.; Vásquez-Vivar, J.; Kalyanaraman, B. Superoxide reacts with hydroethidine but forms a fluorescent product that is distinctly different from ethidium: potential implications in intracellular fluorescence detection of superoxide. Free Radic Biol Med. 2003, 34, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Joseph, J.; Fales, H.M.; Sokoloski, E.A.; Levine, R.L.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc Natl Acad Sci USA. 2005, 102, 5727–5732. [Google Scholar] [CrossRef] [PubMed]
- Papapostolou, I.; Patsoukis, N.; Georgiou, C.D. The fluorescence detection of superoxide radical using hydroethidine could be complicated by the presence of heme proteins. Anal Biochem. 2004, 332, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Roychowdhury, S.; Luthe, A.; Keilhoff, G.; Wolf, G.; Horn, T.F. Oxidative stress in glial cultures: detection by DAF-2 fluorescence used as a tool to measure peroxynitrite rather than nitric oxide. Glia. 2002, 38, 103–114. [Google Scholar] [CrossRef] [PubMed]
- van der Ven, A.J.; Mier, P.; Peters, W.H.; Dolstra, H.; van Erp, P.E.; Koopmans, P.P.; van der Meer, J.W. Monochlorobimane does not selectively label glutathione in peripheral blood mononuclear cells. Anal Biochem. 1994, 217, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Palb, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Woolley, J.F.; Stanicka, J.; Cotter, T.G. Recent advances in reactive oxygen species measurement in biological systems. Trends Biochem Sci. 2013, 38, 556–565. [Google Scholar] [CrossRef]
- Jávega, B. Aplicación de la citometría de flujo al estudio en tiempo real de las interacciones entre especies reactivas de oxígeno y nitrógeno en el estrés oxidativo inducido por xenobióticos. PhD Thesis, Universitat de València, Valencia, Spain.
- Aitken, R.J.; Buckingham, D.; Harkiss, D. Use of a xanthine oxidase free radical generating system to investigate the cytotoxic effects of reactive oxygen species on human spermatozoa. J Reprod Fertil. 1993, 97, 441–450. [Google Scholar] [CrossRef]
- Alexandre, J.; Nicco, C.; Chéreau, C.; Laurent, A.; Weill, B.; Goldwasser, F.; Batteux, F. Improvement of the therapeutic index of anticancer drugs by the superoxide dismutase mimic mangafodipir. J Natl Cancer Inst 2006, 98, 236–244. [Google Scholar] [CrossRef]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.; Hansen, J.M. Oxidative stress, thiols, and redox profiles. Methods Mol Biol. 2012, 889, 325–346. [Google Scholar] [CrossRef] [PubMed]
- Siwik, D.A.; Tzortzis, J.D.; Pimental, D.R.; Chang, D.L.; Pagano, P.J.; Singh, K.; Sawyer, D.B.; Colucci, W.S. Inhibition of copper-zinc superoxide dismutase induces cell growth, hypertrophic phenotype, and apoptosis in neonatal rat cardiac myocytes in vitro. Circ Res. 1999, 85, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Titov, V.Y.; Osipov, A.N. Nitrite and nitroso compounds can serve as specific catalase inhibitors. Redox Rep. 2017, 22, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Serafim, R.A.; Primi, M.C.; Trossini, G.H.; Ferreira, E.I. Nitric oxide: state of the art in drug design. Curr Med Chem. 2012, 19, 386–405. [Google Scholar] [CrossRef] [PubMed]
- Hrušková, K.; Potůčková, E.; Hergeselová, T.; Liptáková, L.; Hašková, P.; Mingas, P.; Kovaříková, P.; Šimůnek, T.; Vávrová, K. Aroylhydrazone iron chelators: Tuning antioxidant and antiproliferative properties by hydrazide modifications. Eur J Med Chem. 2016, 120, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.; Martinez, A.; Blanco, M.; O’Connor, J.E. Assessment of Escherichia coli B with enhanced permeability to fluorochromes for flow cytometric assays of bacterial cell function. Cytometry. 2002, 49, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.; Martinez, A.; O’Connor, J.E.; Blanco, M. Functional Assays of Oxidative Stress Using Genetically Engineered Escherichia coli Strains. Curr Protoc Cytom. 2003, 24, 11–16. [Google Scholar] [CrossRef]
- Yoon, S.H.; Jeong, H.; Kwon, S.K.; Kim, J.F. Genomics, biological features, and biotechnological applications of Escherichia coli B: “Is B for better?!”. In Systems Biology and Biotechnology of Escherichia coli, Lee, S.Y.; Ed.; Springer, Dordrecht, 2009, pp. 1–17. doi.org/10.1007/978-1-4020-9394-4_1.
- Blanco, M.; Martı́nez, A.; Urios, A.; Herrera, G.; O’Connor, J.E. Detection of oxidative mutagenesis by isoniazid and other hydrazine derivatives in Escherichia coli WP2 tester strain IC203, deficient in OxyR: strong protective effects of rat liver S9. Mutat Res Genet Toxicol Environ Mutagen. 1998, 417, 39–46. [Google Scholar] [CrossRef]
- Blanco, M.; Urios, A.; Martı́nez, A. New Escherichia coli WP2 tester strains highly sensitive to reversion by oxidative mutagens. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1998, 413, 95–101. [Google Scholar] [CrossRef]
- Martı́nez, A.; Urios, A.; Blanco, M. Mutagenicity of 80 chemicals in Escherichia coli tester strains IC203, deficient in OxyR, and its oxyR+ parent WP2 uvrA/pKM101: detection of 31 oxidative mutagens. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 467, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Han, M.J.; Jeong, H.; Lee, C.H.; Xia, X.X.; Lee, D.H.; Shim, J.H.; Lee, S.Y.; Oh, T.K.; Kim, J.F. Comparative multi-omics systems analysis of Escherichia coli strains B and K-12. Genome Biol. 2012, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kim, S.; Mukhopadhyay, P.; Cho, S.; Woo, J.; Storz, G.; Ryu, S.E. Structural basis of redox switch in the OxyR transcription factor. Cell, 2001; 105, 103–113. [Google Scholar] [CrossRef]
- Wardman, P. Methods to measure the reactivity of peroxynitrite-derived oxidants toward reduced fluoresceins and rhodamines. Methods Enzymol. 2008, 441, 261–282. [Google Scholar] [CrossRef] [PubMed]
- Fukui, M.; Choi, H.J.; Zhu, B.T. Rapid generation of mitochondrial superoxide induces mitochondrion-dependent but caspase-independent cell death in hippocampal neuronal cells that morphologically resembles necroptosis. Toxicol Appl Pharmacol. 2012, 262, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Clothier, R.; Gómez-Lechón, M.J.; Kinsner-Ovaskainen, A.; Kopp-Schneider, A.; O’Connor, J.E.; Prieto, P.; Stanzel, S. Comparative analysis of eight cytotoxicity assays evaluated within the ACuteTox Project. Toxicol In Vitro. 2013, 27, 1347–1356. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Rooprai, H.K.; Lawrence, P.; Keshavarz, S.; Yashod, P.; Gullan, R.W.; Selway, R.P.; Davies, D. DRAQ7 as an Alternative to MTT Assay for Measuring Viability of Glioma Cells Treated With Polyphenols. Anticancer Res. 2020, 40, 5427–5436. [Google Scholar] [CrossRef]
- McBee, M.E.; Chionh, Y.H.; Sharaf, M.L.; Ho, P.; Cai, M.W.L.; Dedon, P.C. Production of Superoxide in Bacteria Is Stress- and Cell State-Dependent: A Gating-Optimized Flow Cytometry Method that Minimizes ROS Measurement Artifacts with Fluorescent Dyes. Front Microbiol. 2017, 8, 459. [Google Scholar] [CrossRef]
- Spidlen, J.; Brinkman, R.R. Use FlowRepository to share your clinical data upon study publication. Cytometry B Clin Cytom. 2018, 94, 196–198. [Google Scholar] [CrossRef]
- Spidlen, J.; Breuer, K.; Brinkman, R. Preparing a Minimum Information about a Flow Cytometry Experiment (MIFlowCyt) compliant manuscript using the International Society for Advancement of Cytometry (ISAC) FCS file repository (FlowRepository. org). Curr Protoc Cytom. 2012; 61, 10–18. [Google Scholar] [CrossRef]
Figure 1.
Example of biochemical mechanisms involved in the localization and generation of intracellular ROS-sensitive fluorescent probes from extracellular fluorogenic substrates. (A) The incorporation of two acetyl-ester groups renders the chemically reduced, non-fluorescent H2DCF-DA molecule totally non-polar and, thus, able to cross freely plasma membrane. Intracellular esterases cleave acetyl groups and expose two negative charges, thus retaining this substrate in the cytosolic compartment. Intracellular ROS oxidize the molecule and generate a fluorescent probe. This general mechanism is extensible to other esterified fluorogenic substrates. (B) Because of positively charged imino groups (=NH2), DHRH123 is an amphipathic, non-fluorescent fluorogenic substrate permeable to the plasma membrane. Intracellular DHRH123 is transported into mitochondria driven by the mitochondrial membrane potential that reflects the proton-motive force (m) generated by the respiratory electron transfer chain. Once in the mitochondrial matrix, DHRH123 is oxidized by intramitochondrial ROS to the fluorescent probe RH123. A similar mechanism applies for other cationic fluorogenic substrates.
Figure 1.
Example of biochemical mechanisms involved in the localization and generation of intracellular ROS-sensitive fluorescent probes from extracellular fluorogenic substrates. (A) The incorporation of two acetyl-ester groups renders the chemically reduced, non-fluorescent H2DCF-DA molecule totally non-polar and, thus, able to cross freely plasma membrane. Intracellular esterases cleave acetyl groups and expose two negative charges, thus retaining this substrate in the cytosolic compartment. Intracellular ROS oxidize the molecule and generate a fluorescent probe. This general mechanism is extensible to other esterified fluorogenic substrates. (B) Because of positively charged imino groups (=NH2), DHRH123 is an amphipathic, non-fluorescent fluorogenic substrate permeable to the plasma membrane. Intracellular DHRH123 is transported into mitochondria driven by the mitochondrial membrane potential that reflects the proton-motive force (m) generated by the respiratory electron transfer chain. Once in the mitochondrial matrix, DHRH123 is oxidized by intramitochondrial ROS to the fluorescent probe RH123. A similar mechanism applies for other cationic fluorogenic substrates.
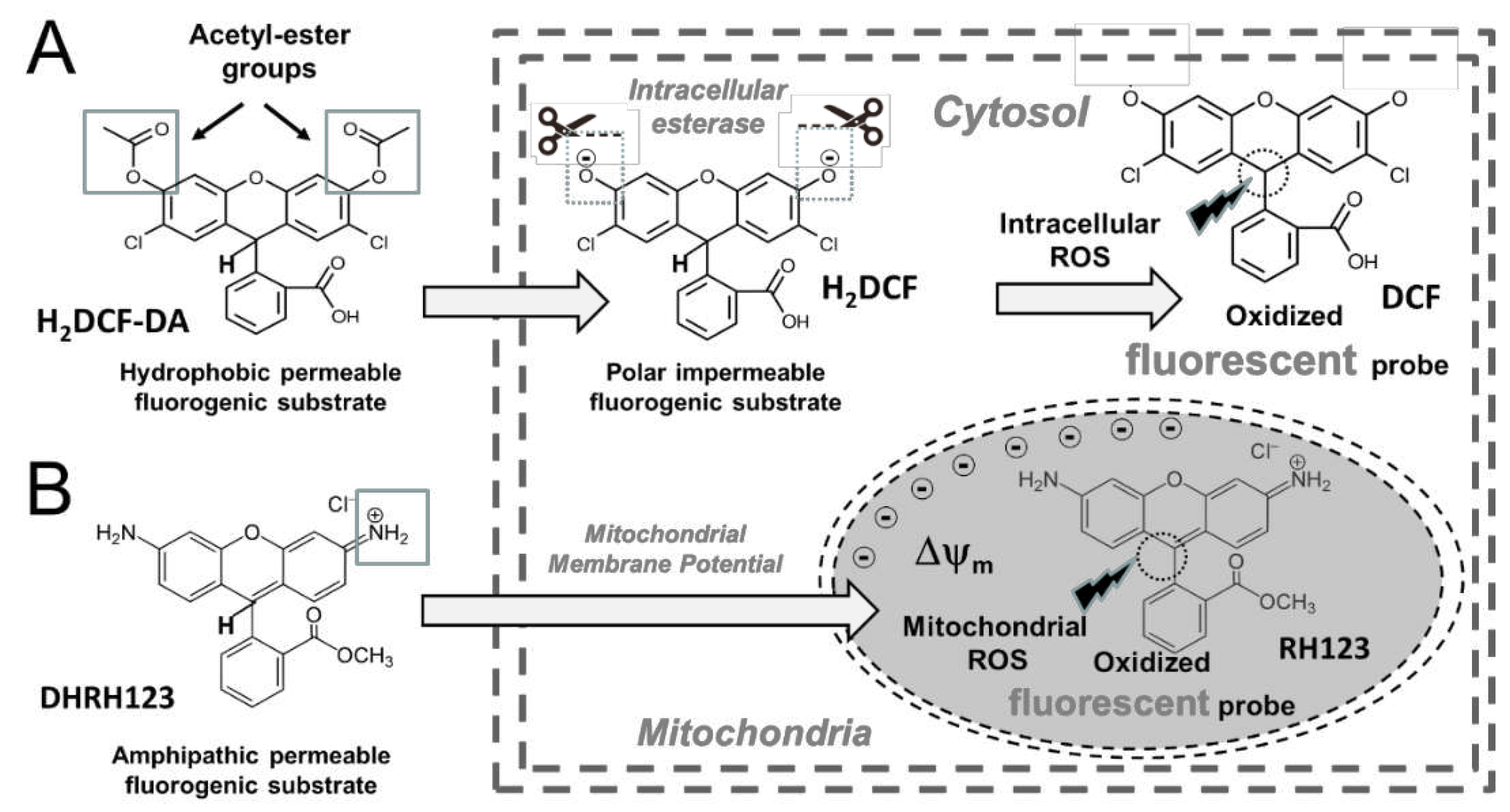
Figure 2.
Examples of specificity issues in common fluorescent probes used for detecting ROS in mitochondria (A, B) and for assessing Glutathione (GSH)-related antioxidant activity. DHRH123 (A) and Mito-PY1 (B) are both used for detection of intramitochondrial ROS. Both probes share the same electrochemical mechanism for intramitochondrial localization, as described in Figure 1 for DHRH123. However, several ROS and RNS can react with the indicated H- atom in DHRH123, while the boronate group in Mito-PY1 undergoes a highly chemospecific reaction only with H2O2. Chloromethyl SNARF (C) and monochloro-bimane (D) are commonly used for GSH detection. However, because of its strong reactivity, the chloromethyl group in chloromethyl SNARF forms adducts with most thiol (-SH) groups. On the contrary, monochloro-bimane is a substrate of the enzyme GSH-transferase (GST) and forms adducts only with GSH.
Figure 2.
Examples of specificity issues in common fluorescent probes used for detecting ROS in mitochondria (A, B) and for assessing Glutathione (GSH)-related antioxidant activity. DHRH123 (A) and Mito-PY1 (B) are both used for detection of intramitochondrial ROS. Both probes share the same electrochemical mechanism for intramitochondrial localization, as described in Figure 1 for DHRH123. However, several ROS and RNS can react with the indicated H- atom in DHRH123, while the boronate group in Mito-PY1 undergoes a highly chemospecific reaction only with H2O2. Chloromethyl SNARF (C) and monochloro-bimane (D) are commonly used for GSH detection. However, because of its strong reactivity, the chloromethyl group in chloromethyl SNARF forms adducts with most thiol (-SH) groups. On the contrary, monochloro-bimane is a substrate of the enzyme GSH-transferase (GST) and forms adducts only with GSH.
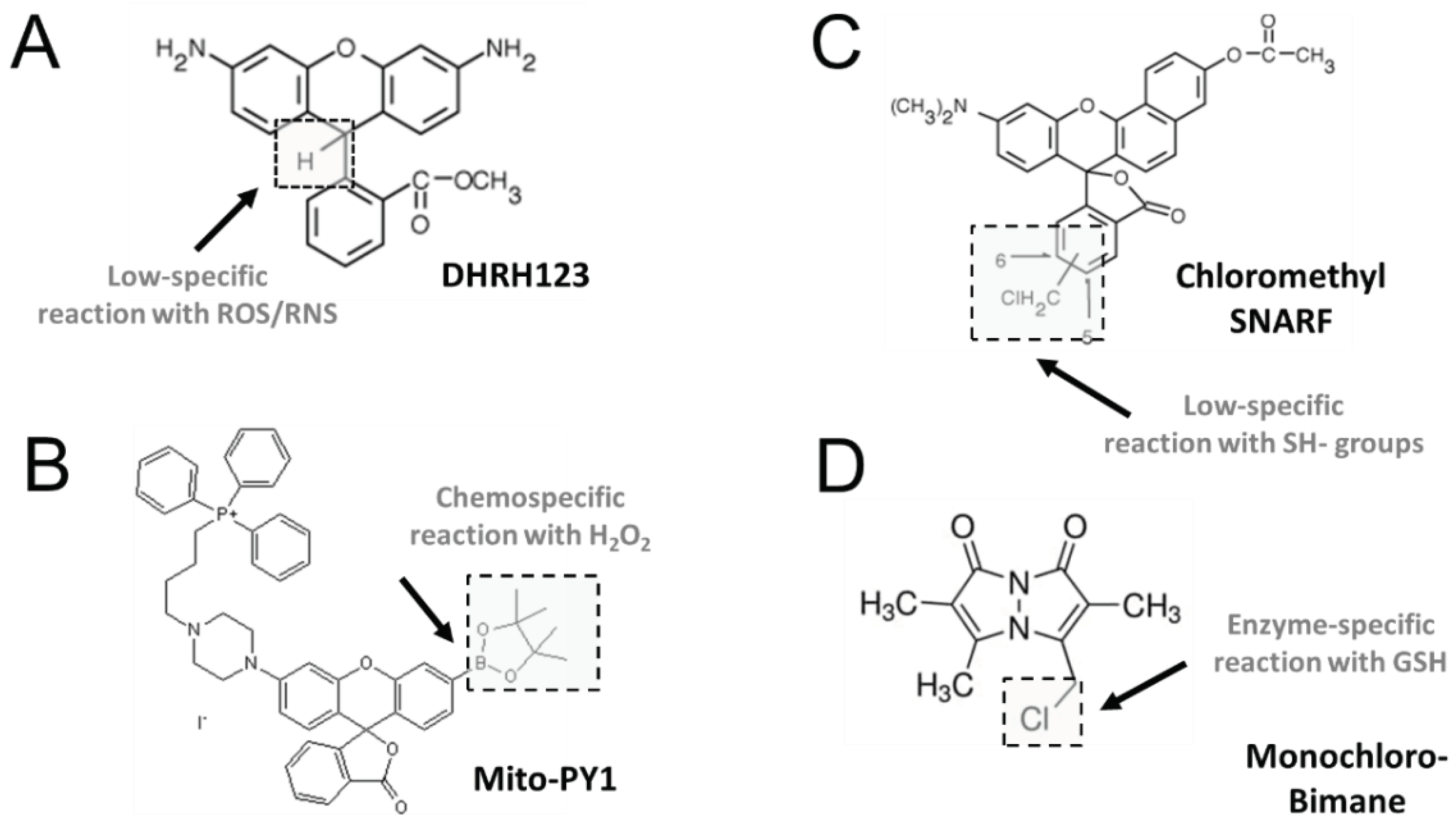
Figure 3.
Representative examples of (A) end-point and (B) real-time kinetic flow cytometric analysis of ROS generation. Panel A summarizes graphically the performance of the oxidative burst in whole-blood phagocytes of a healthy donor, using a commercial kit for clinical diagnosis of the Chronic Granulomatous Disease (CGD) (FagoFlowex) under manufacturer’s instructions. Samples of human peripheral whole blood are incubated for 30 min with the fluorogenic substrate DHRH123 in the presence of vehicle (column Control) or Escherichia coli (column +E. coli). For positive control (column +PMA), blood samples are incubated with the phorbol ester PMA, an activator of Protein kinase C (PKC). Row A1 shows the morphological identification and gating of phagocytic cells in a FS INT Lin vs SS INT Lin dotplot. Row A2 displays the ROS generation (DHRH123 vs SS INT Lin plots) during the oxidative burst induced by phagocytosis (+E. coli) or PKC activation (+PMA), as compared with the spontaneous ROS generation induced by the experimental conditions (Control). Row 3 compares the monoparametric histograms of fluorescence intensity of ROS-generated RH123 in phagocytes of the three experimental conditions. Such histograms are clinically used to diagnose CGD or other oxidative-burst related genetic defects [58]. Panel B exemplifies graphically the performance of a flow cytometry kinetic analysis (RT-FCM or In Fluxo analysis [62]) of the ROS generation in Jurkat cells treated with exogenous H2O2 or with the mitochondrial uncoupler FCCP [196]. Graph B1 shows the identification and gating of live (blue) and dead (orange) cells using FS signal and the viability stain DAPI [196] and graph B2 displays the gate used for single-cell selection in a FS Peak vs FS INT dotplot. For In Fluxo experiments, Jurkat cells are incubated with the appropiate fluorogenic substrates and the viabiity marker. Dotplots display real time on X-axis and the fluorescence intensity of each probe in the Y-axis (log scale). Cells are run in the cytometer for some seconds to establish baseline fluorescence, the run is halted for addition of the exogenous challenge (H2O2 or FCCP) and immediately continued until Graph B3 shows that H2O2 addition to the cells results in a fast generation of intracellular ROS (measured with ROS-generated DCF from the substrate H2DCF-DA), especially in the living cells (blue). Graph B4 shows that mitochondrial membrane potential (MMP) is slightly decreased in live cells (blue), but not in dead cells (orange) by H2O2 addition, as indicated by the fluorescence of TMRM mitochondrial probe, the product of the fluorogenic substrate TMRM-ME. The effect of FCCP is shown in graphs B5 and B6. Mitochondrial uncoupling by FCCP induces a slight generation of ROS in live cells, but not in dead ones (graph B5), while MMP is strongly decreased by FCCP in live cells (graph B6). As seen in this plot, dead cells are characterized by an already very low MMP, which is not modified by FCCP.
Figure 3.
Representative examples of (A) end-point and (B) real-time kinetic flow cytometric analysis of ROS generation. Panel A summarizes graphically the performance of the oxidative burst in whole-blood phagocytes of a healthy donor, using a commercial kit for clinical diagnosis of the Chronic Granulomatous Disease (CGD) (FagoFlowex) under manufacturer’s instructions. Samples of human peripheral whole blood are incubated for 30 min with the fluorogenic substrate DHRH123 in the presence of vehicle (column Control) or Escherichia coli (column +E. coli). For positive control (column +PMA), blood samples are incubated with the phorbol ester PMA, an activator of Protein kinase C (PKC). Row A1 shows the morphological identification and gating of phagocytic cells in a FS INT Lin vs SS INT Lin dotplot. Row A2 displays the ROS generation (DHRH123 vs SS INT Lin plots) during the oxidative burst induced by phagocytosis (+E. coli) or PKC activation (+PMA), as compared with the spontaneous ROS generation induced by the experimental conditions (Control). Row 3 compares the monoparametric histograms of fluorescence intensity of ROS-generated RH123 in phagocytes of the three experimental conditions. Such histograms are clinically used to diagnose CGD or other oxidative-burst related genetic defects [58]. Panel B exemplifies graphically the performance of a flow cytometry kinetic analysis (RT-FCM or In Fluxo analysis [62]) of the ROS generation in Jurkat cells treated with exogenous H2O2 or with the mitochondrial uncoupler FCCP [196]. Graph B1 shows the identification and gating of live (blue) and dead (orange) cells using FS signal and the viability stain DAPI [196] and graph B2 displays the gate used for single-cell selection in a FS Peak vs FS INT dotplot. For In Fluxo experiments, Jurkat cells are incubated with the appropiate fluorogenic substrates and the viabiity marker. Dotplots display real time on X-axis and the fluorescence intensity of each probe in the Y-axis (log scale). Cells are run in the cytometer for some seconds to establish baseline fluorescence, the run is halted for addition of the exogenous challenge (H2O2 or FCCP) and immediately continued until Graph B3 shows that H2O2 addition to the cells results in a fast generation of intracellular ROS (measured with ROS-generated DCF from the substrate H2DCF-DA), especially in the living cells (blue). Graph B4 shows that mitochondrial membrane potential (MMP) is slightly decreased in live cells (blue), but not in dead cells (orange) by H2O2 addition, as indicated by the fluorescence of TMRM mitochondrial probe, the product of the fluorogenic substrate TMRM-ME. The effect of FCCP is shown in graphs B5 and B6. Mitochondrial uncoupling by FCCP induces a slight generation of ROS in live cells, but not in dead ones (graph B5), while MMP is strongly decreased by FCCP in live cells (graph B6). As seen in this plot, dead cells are characterized by an already very low MMP, which is not modified by FCCP.
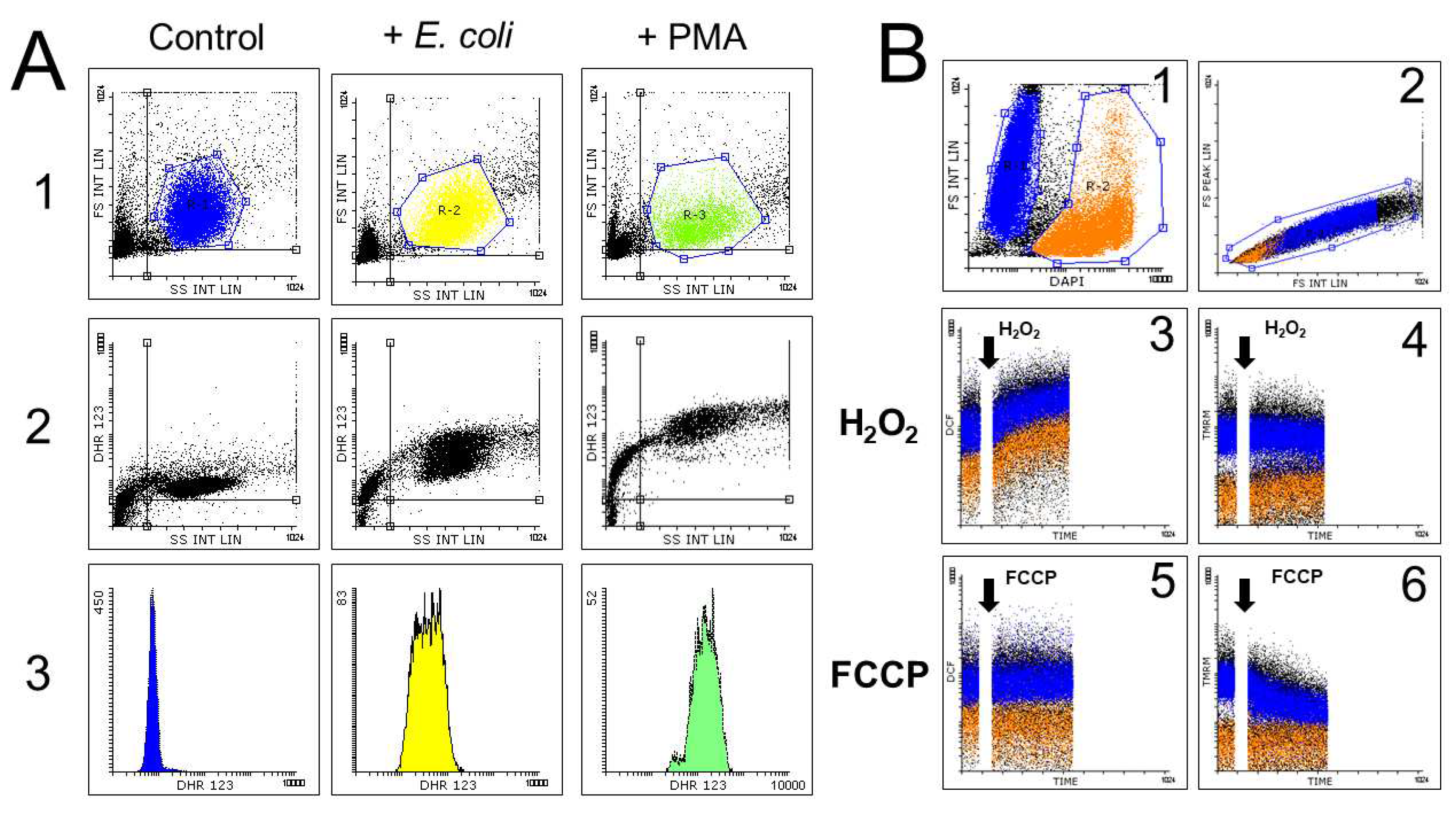
Figure 4.
Scheme of the most relevant aspects of the experimental design and performance of a typical flow cytometric assay aimed to detecting intracellular ROS generation induced by an exogenous prooxidant chemical, such as plumbagin, a redox cycling compound generating superoxide anion radical. Graphs in row A show the strategy for observation of live and dead Jurkat cells by their scatter properties (Graph A1, FS INT Lin vs SS INT Lin) and for selection of single cells (Graph A2 FS Peak vs FS INT dotplot). In graph A3, single cells either live (fuchsia) or dead (green) are confirmed and selected by using the viability stain DAPI. On the basis of live cells, panels B and C show the performance of range-finding experiments to define plumbagin cytotoxicity (panel B) and for titrating the fluorescent probe MitoSOX Red (panel C), selected for assessing O2.- generation by plumbagin. Plumbagin is relatively low-toxic for Jurkat cells (panel B), and the concentration 80 M is chosen for titrating MitoSOX Red (panel C). By incubating Jurkat cells at a fixed cell density (100,000 cells/mL) with a series of Mitosox Red concentrations in the presence of 80 M plumbagin, it is shown that over 500 nm Mitosox Red no significan increase of fluorescence intensity is observed, and thus this dye concentration (and dye-to-cell ratio) is selected for the toxicity examples shown in panels D and E. A similar titration procedure is followed for the probes Hydroethidine (HE) and H2DCF -DA used in these examples. Graphs in row D (control experiment) show that the mitochondrial probe MitoSOX Red (D4) allows a much better discrimination of intracellular ROS levels between live (fuchsia) and dead (green) cells than its related probe HE (D5). Both orange-emitting probes have preferential sensitivity towards O2.- anion. Cytosolic ROS-sensitive probe H2DCF -DA (D6) has an intermediate efficiency in separating ROS levels in live or dead cells in control conditions, as reflected by the green fluorescence emission of DCF. Graphs in row E, however, show a similar efficiency between MitoSOX Red (E4) and HE (E5) for detecting plumbagin-induced ROS generation. Also, H2DCF -DA is able to respond, albeit at lower efficiency, to the incubation with the redox cycling compound plumbagin, supporting the relative lack of specificity of this probe usually considered as a sensor of H2O2 or peroxidative activity.
Figure 4.
Scheme of the most relevant aspects of the experimental design and performance of a typical flow cytometric assay aimed to detecting intracellular ROS generation induced by an exogenous prooxidant chemical, such as plumbagin, a redox cycling compound generating superoxide anion radical. Graphs in row A show the strategy for observation of live and dead Jurkat cells by their scatter properties (Graph A1, FS INT Lin vs SS INT Lin) and for selection of single cells (Graph A2 FS Peak vs FS INT dotplot). In graph A3, single cells either live (fuchsia) or dead (green) are confirmed and selected by using the viability stain DAPI. On the basis of live cells, panels B and C show the performance of range-finding experiments to define plumbagin cytotoxicity (panel B) and for titrating the fluorescent probe MitoSOX Red (panel C), selected for assessing O2.- generation by plumbagin. Plumbagin is relatively low-toxic for Jurkat cells (panel B), and the concentration 80 M is chosen for titrating MitoSOX Red (panel C). By incubating Jurkat cells at a fixed cell density (100,000 cells/mL) with a series of Mitosox Red concentrations in the presence of 80 M plumbagin, it is shown that over 500 nm Mitosox Red no significan increase of fluorescence intensity is observed, and thus this dye concentration (and dye-to-cell ratio) is selected for the toxicity examples shown in panels D and E. A similar titration procedure is followed for the probes Hydroethidine (HE) and H2DCF -DA used in these examples. Graphs in row D (control experiment) show that the mitochondrial probe MitoSOX Red (D4) allows a much better discrimination of intracellular ROS levels between live (fuchsia) and dead (green) cells than its related probe HE (D5). Both orange-emitting probes have preferential sensitivity towards O2.- anion. Cytosolic ROS-sensitive probe H2DCF -DA (D6) has an intermediate efficiency in separating ROS levels in live or dead cells in control conditions, as reflected by the green fluorescence emission of DCF. Graphs in row E, however, show a similar efficiency between MitoSOX Red (E4) and HE (E5) for detecting plumbagin-induced ROS generation. Also, H2DCF -DA is able to respond, albeit at lower efficiency, to the incubation with the redox cycling compound plumbagin, supporting the relative lack of specificity of this probe usually considered as a sensor of H2O2 or peroxidative activity.
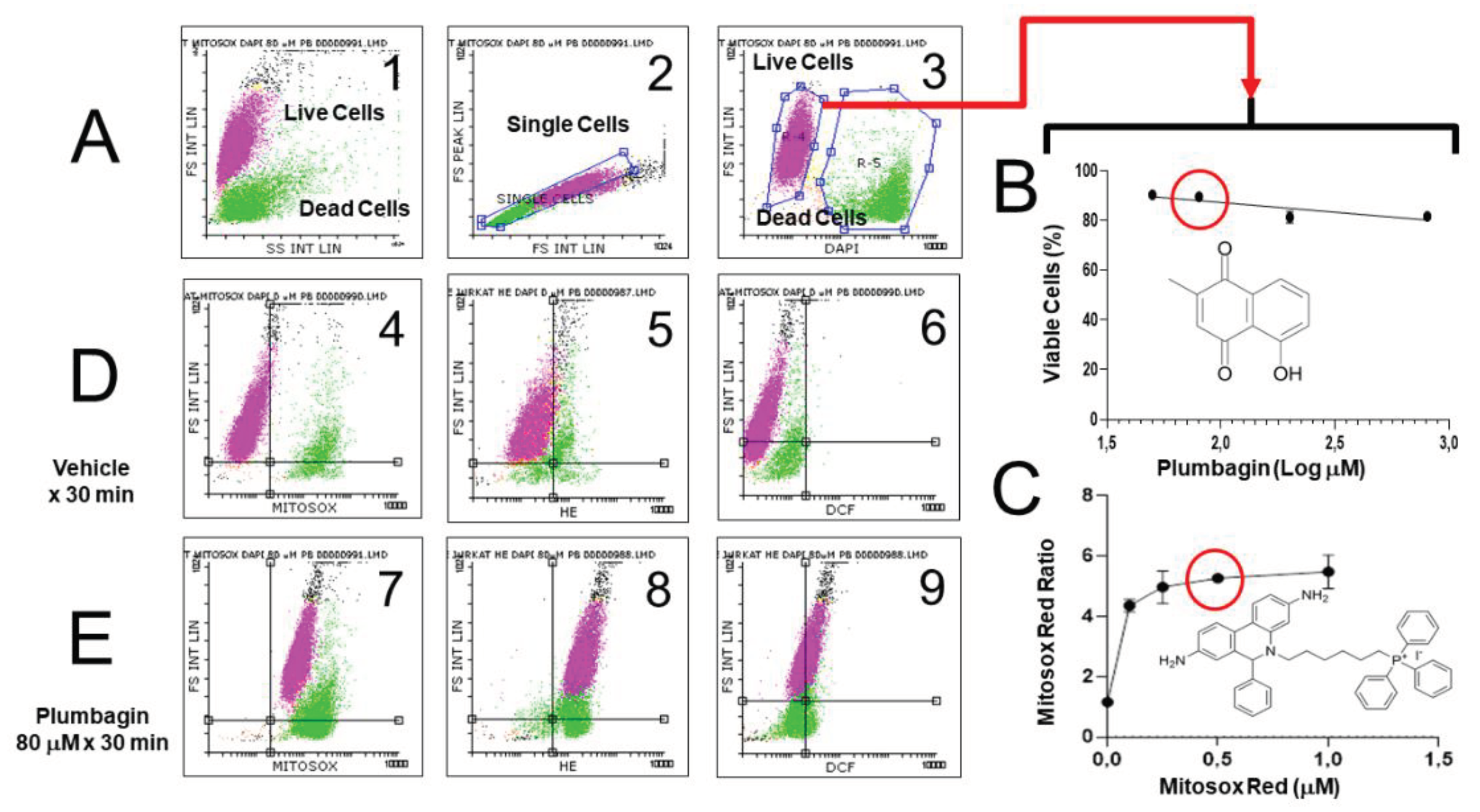

Table 1.
Summary of main critical points and limitations of flow cytometry in functional assays of Oxygen and Oxygen-related stress.
Table 1.
Summary of main critical points and limitations of flow cytometry in functional assays of Oxygen and Oxygen-related stress.
| CRITICAL POINTS AND LIMITATIONS |
|---|
| Identification of blood cells in whole blood samples without lysis of erythrocytes |
| Preparation of single-cell suspensions from adherent cell models |
| Maintenance of viability and functional competence of the cells along sample preparation and experiment performance |
| Identification of the optimal incubation time and concentration for staining |
| Access of fluorogenic substrates to intracellular sites or intracellular processes |
| Retention of fluorogenic substrates and oxidized fluorescent probes |
| Preparation of single-cell suspensions from adherent cell models |
| Lack of absolute specificity of fluorogenic substrates for specific RONS |
| Interference of the probes with ROS biology or ROS-relevant cell functions |
| Interference of the probes with ROS biology or ROS-relevant cell functions |
| Interference of the probes with ROS biology or ROS-relevant cell functions |
| Interference of the probes with ROS biology or ROS-relevant cell functions |
| Selection of the time window for kinetic assays |
| Assay calibration for data expression in biochemical units |
| Adapted from ref. [47] |
Table 2.
Some recommendations for performing in vitro functional studies of Oxygen and Oxygen-related stress by Flow Cytometry.
Table 2.
Some recommendations for performing in vitro functional studies of Oxygen and Oxygen-related stress by Flow Cytometry.
| Biological Process of Interest and Experimental In Vitro Setting | ||||||
|---|---|---|---|---|---|---|
| Peroxidative Activity | Redox Cycling | Antioxidant Defense | ||||
| Probe | MitoPY1 | H2DCF-DA | DHRH123 | MitoSOX Red | HE | Monochlorobimane |
| Viability Stain | DAPI or PI |
DAPI or PI |
DAPI or PI |
DAPI | DAPI | PI |
| Positive control | H2O2 | t-BOOH | CHP | Menadione | Plumbagin | N-Acetyl cysteine GSH-ester |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



