
Submitted:
29 April 2023
Posted:
30 April 2023
You are already at the latest version
Abstract
Transient Receptor Potential Vanilloid 4 (TRPV4) channel is a non-selective cation channel, mostly permeable to calcium (Ca2+), which participates to intracellular Ca2+ handling in cardiac cells. It is widely expressed through the body and is activated by a large spectrum of physico-chemical stimuli, conferring it a role in a variety of sensorial and physiological functions. With-in the cardiovascular system, TRPV4 expression was reported in cardiomyocytes, endothelial cells (ECs) and smooth muscle cells (SMCs) where it modulates mitochondrial activity, Ca2+ ho-meostasis, cardiomyocytes electrical activity and contractility, cardiac embryonic development, fibroblast proliferation but also vascular permeability, dilatation and constriction. On the other hand, TRPV4 channels participate in several cardiac pathological processes such as development of cardiac fibrosis, hypertrophy, ischemia-reperfusion injuries, heart failure, myocardial infarc-tion, and arrhythmia. In this manuscript, we provide an overview of TRPV4 channel implica-tions in cardiac physiology and discuss the potential of TRPV4 channel as therapeutic target against cardiovascular diseases.
Keywords:
TRPV4 channels
; TRPV4-KO mice
; heart
; myocytes
; fibroblasts
; action potential
; calcium homeo-stasis
1. Introduction
The mammalian TRP channels are widely expressed in the heart and can be considered as real ’’cellular switches’’ able to respond to numerous physical and chemical stimuli related to sensory physiology [1], [2], [3]. In mammals, TRP family is composed of 28 members and classified according to amino acid sequence homology into six families: TRPA (ankyrin; TRPA1), TRPC (canonical; TRPC1–TRPC7), TRPM (melastatin; TRPM1–TRPM8), TRPML (mucolipin; TRPML1–TRPML3), TRPP (polycystin; TRPP1–TRPP3) and TRPV (vanilloid; TRPV1–TRPV6) [3], [4]. With some exceptions, most of the TRP channels are able to conduct Ca2+ which plays an important role in stimulus-response reactions of cardiac cells. Interestingly, altered expression, localization and regulation of TRP channels have already been related to cardiovascular disorders due to Ca2+ handling dysregulation [4]. Among the TRPV channels family [5], TRPV4 channel emerged as a key modulator in cardiac cell structure and activity [6], [7]. Recent identification of pharmacological modulators and construction of TRPV4 knockout (KO) mice unmasked several physiopathological roles of this channel [5], [8], [9]. However, the mechanisms that regulate cardiac TRPV4 channels still remain not well apprehended. In this context, this review will discuss the latest findings on the pathophysiological role of TRPV4 channel in the cardiovascular system and why understanding of TRPV4 regulation may lead to novel therapeutical strategies related to cardiac diseases.
2. Gene, structure, function and electrical properties
TRPV4 channel have received a particular attention according to its large expression in the cardiovascular system [5], [8], [10], [11]. TRPV4 protein is encoded by the trpv4 gene, present on the long (q) arm of chromosome 12 at position 24.1 in human [4]. The corresponding locus is found between the bases 109,783,087 & 109,783,406 of the genome and is composed of 15 exons. Alternative exon-splicing leads to 5 channel isoforms (TRPV4-A to E). Only TRPV4-A and TRPV4-D isoforms are localized at the plasma membrane and display identical biophysical properties [12], while the other splice variants (N-terminal deletions) remain in the endoplasmic reticulum (ER) [4] and do not form functional ion channels [12]. In the following sections, we will refer to TRPV4 as the TRPV4-A and TRPV4-D isoforms without distinction. TRPV4 has tetrameric structure, each subunit being composed of six transmembrane segments (S1 to S6), a pattern shared with other TRP channels and voltage-gated ion channel subunits (VGIC). The S1 to S4 segments constitute the peripheral structure while the central S5-S6 loops border the pore of the channel [13], [14], [15], [16]. The N and C-terminal extremities are intracellular and contain a variety of functional domains [17]. A recent crystallographic study combined to X-ray approach insights more details about the structure of Xenopus tropicalis TRPV4 with a resolution of 3.8 Å [16].
The N-terminal region represents more than half of the protein and plays a critical role in channel assembly, trafficking and regulation [16], [18], [19], [20], [21], [22]. It harbours six Ankyrin repeated domains (ARD1–6) that participate in channel oligomerization, protein-protein interaction and trafficking [4]. Note that the absence of these specific domains blocks the TRPV4 trafficking at the ER level [12]. This region also presents a Ca2+-calmodulin kinase type II (CaMKII) regulation site conferring channel sensitivity to intracellular Ca2+. In mammalian cells, a proline-rich sequence in TRPV4 N-terminal can interact with the cytoskeleton protein PACSIN 3 (protein kinase C and casein kinase substrate in neurons 3) thereby regulating channel trafficking, sensitivity to membrane stretch [23] and preventing/reducing TRPV4 activation by heat [24]. In this context, PACSIN3 can be considered as a TRPV4 channel regulatory protein that regulates both the TRPV4 subcellular localization and its function. Obviously, further studies on cardiomyocytes are needed to better understand the mechanism and the role of this interaction. In addition, the TRPV4 N-terminal region is able to interact with OS-9, an ER-resident lectin, in order to prevent channel trafficking to the plasma membrane [25] suggesting that OS-9 is an important auxiliary protein for TRPV4 maturation.
TRPV4 C-terminal extremity contains different domains: i) “TRPbox” (carrying the consensus sequence WKFQR) [26], ii) a CaM binding domain which modulates tetramerization, gate opening [27] and Ca2+ influx [28], iii) a PDZ domain which interacts with numerous cell auxiliary proteins [4], [13]. Furthermore, the microtubule-associated protein 7 (MAP7) interaction with the C-terminal extremity of TRPV4 increases both expression and functional activity at the plasma membrane in Chinese hamster ovary cells [29]. Because MAP7 is directly related to the cytoskeletal filaments, this interaction may underline the TRPV4 mechanotransduction mechanism [29] nevertheless this hypothesis requires further attention in myocytes.
TRPV4 principally assemble as homotetramers [13], [30] to form a functional channel but can also form heterotetramers with few other members of the TRP channels family : TRPC1 [31], [32] and TRPP2 [30], [32]. It can also interact with the α-subunit of few ion channels and aquaporins [33], [34], [35], [36], [37], [38]. This heteromerization was shown to modify TRPV4 channel biophysical properties in heterologous expression systems [32], [39], [40]. TRPV4 channel shows a lower voltage sensitivity compared to other TRP channels but still harbors an outward rectification current when expressed in heterologous expression system, constituting the signature of most TRP channels. To date, this weak voltage dependence of TRPV4 is not yet clearly elucidated. Nevertheless, the low density of positive charges in the S4 voltage sensor domain [41] combined with a better understanding of its crystal structure [16], allows to get a plausible mechanism of its gating. Indeed, TRPV4 adopts a domain-swapped arrangement between the S1-S4 domain and the pore domain S5-S6, similarly to TRPV1, TRPV2 and numerous VGICs whereas the connector between these last two domains adopts an ordered loop structure rather than an α-helix segment present within TRPV1 and TRPV2. This last structural aspect is important since VGICs connector acts as a mechanical lever to couple the pore opening and the voltage sensor activation [42], [43] and its absence can change the gating behavior [44] [45]. Taken together, the interface behavior between the S1–S4 and pore domains in TRPV4 is unique among TRP channels and not closely related to the VGICs.
The TRPV4 single channel conductances are ~ 30-60 pS and ~ 80-100 pS for inward and outward currents respectively [46]. Like most of TRP channels, TRPV4 is mainly permeable to Ca2+ over other ions and its permeability sequence was established to be as follows: Ca2+ >> Mg2+ > K+ > Na+ (PCa/PNa ~ 10 and PMg/PNa ~ 2-3) [31], [47], [48] . Surprisingly, the macroscopic TRPV4 current in cardiomyocytes has not been unmasked, yet. Recent structural characterization performed by Yuan group has reported a larger selectivity filter (10.6-12.6 Å) within the upper gate of TRPV4 compared to other TRPV channels (TRPV1, TRPV2 & TRPV6 ranging from 4.2 to 7.4 Å) (Figure 1- [16]).
Note that the presence of this specificity does not significantly increases the degree of Ca2+ permeability within this ion channel family but should potentially be considered as an important structural feature when TRPV4 has to deal with mechanical forces associated with increased membrane tension.
3. Available tools to investigate TRPV4 roles
Intracellular Ca2+ concentration plays a pivotal role in living organisms because it is involved in several physiological regulations but also in response to various pathological states [4], [49]. In the myocardium, cellular Ca2+ homeostasis maintains normal heart function which requires various specialized proteins such as ion channels and exchangers [50]. The central role Ca2+ in cardiac excitation-contraction is well established where Ca2+ is at the interface of electrical membrane activity (Action potential (AP)) and cell contraction [51]. Any disturbance in intracellular Ca2+ homeostasis may thus lead to contractile and/or electrical defects in the myocardium. In this context, TRPV4 function related to Ca2+ permeability is evaluated with the greatest attention [52], [53]. Consequently, synthetic specific antagonists of TRPV4 have been designed and evaluated to treat human diseases including cardiac pathologies [49], [53], [54].
3.1. Pharmacological modulators
To date, several synthetic molecules that modulate TRPV4 channels were developed to aim at treating diseases such as osteoarthritis, respiratory diseases, cancers, gastrointestinal disorders, and pain [53]. Recently, some of these compounds entered safety trials to treat heart failure [53]. Progress in the development of these agents led to a last generation of more potent and specific modulators that have been used to unmask TRPV4 channels pathophysiological roles in heart.
3.1.1. TRPV4 agonists
Four generations of TRPV4 agonists (see Table 1) have been used on different tissues and cell types to determine its physiological and pathological functions in the myocardium. The 4α-phorbol 12,13-didecanoate (4α-PDD, a semisynthetic derivatives of the diterpenoid phorbol), 5,6 epoxyeicosatrienoicacid (5,6-EET, an oxidative metabolite of arachidonic acid), the RN-1747 (a benzenesulfonamide), and the GSK1016790A agonist (fully synthetic) which has been extensively used in cardiovascular investigations [8], [11], [55], [56], [57]. In addition, Atobe and al., have recently reported the new TRPV4 agonist quinazolin-4(3 H)-one derivative intended to treat osteoarthritis [58]. (see table 1)
3.1.2. TRPV4 antagonists
Over the last decade, several TRPV4 antagonists were used to highlight TRPV4’s roles in the cardiovascular system. Initial studies used several non-selective TRPV4 antagonists such as ruthenium red (RuR) and gadolinium [59]. Subsequent studies have evaluated the effect of more selective antagonists including: RN-1734 which completely blocks TRPV4 channels without affecting other TRP channels [60] or the widely-used HC067047 antagonist which was used in cardiac investigations in the context of ischemia/reperfusion [56], [61]. Recently, a promising clinical antagonist GSK2193874 which possesses high TRPV4 affinity, has been shown to efficiently prevent and treat lung edema in heart failure models and congestive heart failure [62] (Table 2). Several optimized antagonists exhibiting better in vivo availability (RN-9893) [63], solubility (GSK3527497) [64], efficacy (novel 2′,4′-dimethyl-[4,5′-bithiazol]-2-yl amino derivatives [65], [66] and GSK2798745 [53]) or with a decreased toxicity (Sulfone Pyrrolidine Sulfonamide) [54], [67] have been developed since then. Knowing that TRPV4 antagonist GSK2798745 [53] is the first inhibitor tested in humans (phase 2), more homologs are likely to appear in the near future (see below Table 2). Indeed, the recently unveiled Cryo-EM structure [16] of TRPV4 channels combined with the pharmaceutical industry efforts should accelerate knowledge related to the mechanism of action of current modulators and may help to identify the next generation of TRPV4 drugs.
3.2. TRPV4 knockout mice
To our knowledge, two transgenic TRPV4 knock-out mice strains were developed from two different groups. Suzuki et al, used the 129/SvJ strain via a cassette insertion mutagenesis of exon 5 [96] whereas Liedtke and al, used the C57bl/6J strain with a Cre-lox-mediated excision of exon 12 [97]. Both mice strains were viable and fertile [96], [97] (see Table 3).
4. Physiological roles in cardiovascular system
4.1. TRPV4 expression profile under physiological condition
In mammals, TRPV4 channel expression is distributed in various organs and tissues including blood vessels and heart. RT-PCR, immunostainings, and functional recordings demonstrated that TRPV4 channels are expressed in cardiac cell types such as atrial and ventricular cardiomyocytes [8], [10] [11], [52], [56], [61], [113], [114], human embryonic stem cell-derived cardiomyocytes [115], cardiofibroblasts (CFs) [52], [81], [90], [116], [117], [118], [119], endothelial cells (ECs) [84], [101], [120], [121], [122], [123] and smooth muscle cells [71], [84], [87], [122], [124], [125], [126], [127], [128], [129] (See Table 4).
Taken together, TRPV4 channel is present and functional at the plasma membrane of cardiac cells where it plays important physiological roles.
4.2. Modulation of ventricular electrical activity
In healthy mammalian hearts, the action potential (AP) waveform initiates and modulates cardiac contractility. It occurs suddenly and transitorily when the resting membrane potential depolarizes and repolarizes according to the successive opening and closing of several ion channels. A lot of attention is paid to the ventricular repolarization phase in pre-clinical investigations, as AP prolongation is often associated with an increased risk of potentially lethal arrhythmias. Recent evidence has shown that TRP channels opening produces a depolarizing current, since the net flux of cations is inward under physiological conditions [3]. Therefore, these channels are particularly important in excitable cells, such as cardiomyocytes, where they can both trigger and modulate AP shape. To date, some TRP channels, such as TRPC3 [131], [132], TRPM4 [133], [134, 135] and TRPM7 [136] were shown to modulate both sinus node and cardiac AP. Interestingly, deletion or mutations of TRPM4 were reported to be associated with several cardiac electrical disorders (Brugada, long QT, progressive cardiac conduction disorder) [137], [138], [139], [140], [141] and hypertrophy [142]. Since other TRP channels are expressed in cardiac myocytes they may participate in cardiac electrical activity and their mutations could lead to inherited cardiac electrical disorders. Thus, it appears necessary to specifically describe the effects of each of these channels, including TRPV4, and evaluate their physiological roles in the cardiovascular system.
TRPV4 protein expression was detected in mouse [8], [56], [61] and rat [56] ventricular myocytes. It shows a membrane specific expression and location (plasma membrane and t-tubules) which is age-dependent [8], [11]. In young mice [8], TRPV4 agonist GSK1016790A induced a dose-dependent and transient increase in AP duration in trpv4+/+ ventricular myocytes. In this study, GSK1016790A was proposed to promote the trafficking TRPV4 channel to the membrane. This phenomenon was transient, because of subsequent rapid channel endocytosis [8], [57], [143]. A similar effect of GSK1016790A was also observed on rat atrial myocytes [52]. According to its biophysical properties, TRPV4 potentiation in a physiological or pathological context would prolong AP. Conversely, the TRPV4 inhibitor GSK2193874 (100 nM) significantly shortened AP duration in trpv4+/+ ventricular myocytes. The effect of these pharmacological modulators on the ventricular AP were not observed in trpv4-/- mice, highlighting the effects observed on WT cardiomyocytes were exclusively due to TRPV4 modulation. These data suggest that TRPV4 channels carry an inward current, which remains to be characterized, during the ventricular AP in basal conditions. Computational modelling was used to predict the shape of TRPV4 current during AP [8]. This modelling considered channel permeation and open probability depending on the voltage and subsarcolemmal [Ca2+]i that will be sensed by the channel. Since TRPV4 channel open probability increases with depolarization and decreases with [Ca2+]i elevation, the model predicted a transient inward current, that develops rapidly after the AP upstroke and declines with the rise in [Ca2+]i and membrane repolarization [8]. The consequence of trpv4 deletion on cardiac electrical activity was also evaluated. In vivo electrocardiogram recordings revealed significantly shortened QT interval in trpv4-/- mice compared to their wild type littermates (C57bl/6J strain). In accordance with these data, a 19% reduction in AP duration was found in ventricular myocytes from trpv4-/- mice compared to trpv4+/+ mice. No change in the properties of the main VGICs participating in AP was observed between trpv4+/+ and trpv4-/- myocytes, suggesting that QT and AP duration shortening in trpv4-/- mice are exclusively attributable to trpv4 deletion [8] . Finally, a recent investigation has shown that TRPV4 expression level is increased in cardiomyocytes of aged (24–27 months) mice compared with young (3–6 months) mice [11] but TRPV4 contribution to the prolonged repolarization commonly observed with aging remains to be studied.
Together, these studies suggest that TRPV4 modulates ventricular electrical activity under basal conditions and may thus be involved in cardiac arrhythmias in aging and/or under pathological conditions. [11] (see section 4 for details).
4.3. Modulation of cardiac contractility
Muscular contractility is onset by an increase in cytoplasmic Ca2+. For so, Ca2+ handling results from a tight balance between both the Ca2+ influx, intracellular store release and uptake and cell extrusion [51]. In a ventricular myocyte, Ca2+ influx is mainly mediated by voltage-gated calcium channels (Cav1.2) whereas the Ca2+ extrusion is ensured by Na/Ca exchanger (NCX1) and plasma membrane Ca2+ pump (PMCA) [144]. Several TRP channels are present and functional and can constitute an alternative source of Ca2+ entry. Furthermore, it is well established that impaired Ca2+ handling leads to abnormal contractility [145], [146], [142]. Therefore, a better understanding of TRP channels, involved in Ca2+ handling and contractility constitutes an important step to treat deleterious cardiac diseases involving cytosolic Ca2+ overload in myocytes [147]. Regarding TRPV4, several studies reported its implication in these processes while others did not observe obvious contribution. Indeed, its contribution to intracellular Ca2+ influx and myocardial contractility was examined by Li et al., using the TRPV4 agonist 4-αPDD on isolated rat papillary muscle. The authors revealed that the 4-αPDD had no effect on contractility [148]. In the same line, another study revealed in isolated perfused rat hearts, TRPV4 channels potentiation by the agonist GSK1016790A also had no effect on beating rate and contractility but induced, at the in vivo level, circulatory collapse which was most likely due to vascular leakage and tissue haemorrhage in the lung [80]. On the other side, our group has recently shown that the TRPV4 agonist GSK101679A perfusion induced a significant increase of the transient Ca2+ current influx in trpv4+/+ but not in trpv4-/- mouse myocytes [8] confirming TRPV4 involvement in modulating ventricular intracellular Ca2+ concentration. Recently, Veteto et al., explored the relationship between the TRPV4 function and Ca2+ handling after mechanical stimulation in aged hearts [114]. To this end, the authors explored the effect of left ventricular preload elevation in working-heart perfused aged mice hearts and found that following the initial Frank-Starling response, these hearts exhibited a secondary increase in left ventricular maximal pressure which was absent when perfusing TRPV4 antagonist HC067047. Interestingly, when preload elevation was maintained for a longer period of time (20 min), maximal left ventricular pressure declined less in HC067047-treated than in untreated aged hearts. The authors then studied Ca2+ handling in ventricular myocytes following uniaxial stretch and found a delayed increase in intracellular Ca2+ in myocytes from aged mice ultimately leading to a contractured state which were both prevented by HC067047 treatment. These results suggest that TRPV4 is responsible for Ca2+ entry leading to a hypercontractile response secondary to myocardial stretch in the aged heart but when stretched was maintained, TRPV4-mediated Ca2+ influx was detrimental to cardiac contractility. Further investigations have reported that TRPV4 expression increases in mouse cardiomyocytes with advancing age [11] and after in vivo ischemia-reperfusion [61]. Note that TRPV4 upregulation in aged mice was associated to increased hypoosmotic stress-mediated contractility, enhanced cell death and increased ischemia-reperfusion injuries [11]. Finally, inhibition of TRPV4 by HC067047 exerts cardioprotective effect during ischemia-reperfusion, attesting the key role of TRPV4 in Ca2+ handling and contractility [149]. It is important to underline that TRPV4 channel can also dock with other TRP channels and little is known about the contribution of these types of channels in intracellular Ca2+ dynamics and their involvement in the myocyte contractility. In this context, detailed interplay between TRPV4 channels and various α-subunits requires additional research in cardiomyocytes.
Collectively, these data suggest that TRPV4 channel represents an attractive ion channel target to prevent Ca2+ overload and cardiac contractility dysfunction [56], [61], [10], [150]. Indeed, TRPV4 seems to contribute to Ca2+ homeostasis regulation under physiological condition without impacting the contractility, whereas its implication may to be more deleterious during aging or under pathological conditions and lead to Ca2+ overload and altered contractility. Further investigations are undoubtedly needed to fully understand TRPV4 role in cardiac contractility in cardiovascular diseases.
4.4. Modulation of vascular tone
TRPV4 is expressed in ECs where its activation results in vasodilatation [151], [152]. TRPV4 is sensitive to shear stress [153] and is thus able to be activated by blood flow and induce Ca2+-entry when expressed in endothelium. In ECs, Ca2+-entry activates endothelial nitric oxide synthase (eNOS) pathway and the production of nitric oxide (NO), which can, in turn, diffuse to smooth muscle cells (SMC). In SMCs, NO activates cyclic GMP / PKG signaling inducing vasodilatation [152], leading to endothelium-derived factors release, causing SMC hyperpolarization and vasodilatation [152]. Furthermore, Ca2+-entry activates in parallel intermediate (IK) and small (SK) conductance Ca2+-sensitive K+ channels in ECs, leading to hyperpolarization of ECs, and then, hyperpolarization of SMC via myoendothelial gap junctions [152]. Genetic deletion or pharmacological blockade of TRPV4 channels inhibits NO- and endothelium-derived hyperpolarizing factor (EDHF)-dependent relaxation of mouse small mesenteric arteries in response to flow [101]. Accordingly, transfection of TRPV4 channels in HEK293 cells confers them sensitivity to flow and induces shear stress-dependent Ca2+-entry [101] confirming the important role of TRPV4 in sensing shear stress. TRPV4 transfected HEK293T cells are also sensitive to cell confluence, modulating TRPV4 response to hypoxia at high cell density suggesting another important feature of TRPV4 activation i.e. hypoxia[154]. Indeed, in vessels, hypoxia induces vasodilatation in systemic circulation, whereas it induces vasoconstriction in pulmonary circulation. TRPV4 activation with its agonist GSK1016790A causes endothelium-dependent relaxation of pre-contracted rat pulmonary artery rings, and this relaxation is inhibited in de-endothelized vessels. The authors of this study suggested that both NO and EDHF contribute to GSK1016790A-induced relaxation [122]. Ottolini et al. found that TRPV4 channels in mouse ECs co-localize with IK/SK channels in mesenteric arteries but not in pulmonary arteries, which explains that TRPV4 sparklets can activate IK/SK channels in mesenteric arteries but not in pulmonary arteries, where ECs TRPV4 activates preferentially eNOS [130]. Additionally, flow-induced vasorelaxation in human coronary micro vessels requires endothelial TRPV4 activation which increases mitochondrial ROS production in ECs and induces ROS-dependent vasodilation [123], [155]. TRPV4 is also expressed in SMC where it participates in vascular contraction, cell migration and proliferation[124], [127]. TRPV4 activation in pulmonary artery SMC results in a Ca2+ entry and Ca2+ release from the SR through the activation of ryanodine receptors[71] . This TRPV4-mediated Ca2+ elevation leading to SMC contraction [71], [87] can be triggered by serotonin [87], [128] or mechanical stimuli such as flow shear stress [129]. Dysregulation of TRPV4 expression between ECs and SMC could lead to an impaired balance between vasorelaxation and vasoconstriction and result in an altered myogenic tone [71], [156], [125].
5. Pathological implications of TRPV4 channels
5.1. Expression remodelling under pathological condition
TRPV4 expression increases under certain pathophysiological conditions, such as aging [11] pressure overload [157], [158], ischemia-reperfusion [56], [61], increased membrane tension [114], [115], pericarditis in rats and patients with atrial fibrillation [52]. In this context, it seems to be important to investigate the TRPV4 expression profile during life but also during the development of cardiac diseases.
5.2. Arrhythmias
Atrial fibrillation is the most common cardiac arrhythmia and is related, in part, to Ca2+ handling defects [159]. Since TRPV4 channels are expressed in native cardiac cells including atrial cardiomyocytes their implication in atrial fibrillation was evaluated [52]. In a model of pericarditis-related atrial fibrillation phenotype, TRPV4 channel inhibition by GSK2193874 limited abnormal electrophysiological changes, cardiac fibrosis and inflammation, resulting in a reduced vulnerability to atrial fibrillation [52]. Accordingly, TRPV4 may constitute an interesting therapeutic target to treat human atrial fibrillation. Another recent investigation pointed the importance to consider aging and his potential negative effect on Ca2+ handling, the resting membrane potential and the risk to develop ventricular arrhythmia after ischemia-reperfusion. In this context and because TRPV4 channel is up-regulated in cardiomyocytes of aged mice [11], the perfusion of HC067047 (TRPV4 antagonist) after ischemia-reperfusion, reduces incidence of pro-arrhythmic diastolic Ca2+ signaling, maintains the resting membrane potential and decreases the ventricular arrhythmia score [160]. Thereby, TRPV4 blockade may also constitute an interesting strategy to limit the occurrence of arrhythmogenic events for aged populations following ischemia-reperfusion and MI.
5.3. Cardiac Remodelling and fibrosis
There are some evidence that TRPV4 may be involved in cardiac pathological remodelling and could thus be an interesting therapeutic target in these contexts. In a recent study, TRPV4 channel expression was found to be significantly increased in the ventricles of mice with left ventricular pressure-overload induced hypertrophy and in human failing ventricles [158]. Interestingly, pressure-overload resulted in reduced cardiac hypertrophy, cardiac dysfunction, fibrosis, inflammation in TRPV4 knockout mice compared to wild-type animals. Moreover, treatment with GSK2193874 (TRPV4 antagonist) prevented the pressure-overload induced remodelling and dysfunction further confirming TRPV4 involvement. In the same study, the authors showed that the increase in CaMKII phosphorylation found in the pressure-overloaded left ventricle in wild-type mice was absent in TRPV4 knockout mice. They suggested that this TRPV4-related increase in CaMKII phosphorylation, leads to NF-κB phosphorylation and NLRP3 activation, which both contribute to the pro-inflammatory remodelling found in these pressure-overloaded hearts. Therefore, inhibiting TRPV4 appears promising to limit CaMKII phosphorylation and its multiple consequences and inflammation in cardiac hypertrophy. The role of TRPV4 in cardiac remodelling has also been studied in the context of myocardial infarction and subsequent fibrosis through channels expressed in CFs.CFs originate from mesenchymal stem cells, and represent 2/3 of the cardiac cell population [161]. They contribute to maintain the structural, biochemical, mechanical and electrical properties of the healthy myocardium and constitute the main source of extracellular matrix (ECM) protein production (collagen types I and III and metalloproteases) [162]. Fibrosis is known to be related to an expansion of the cardiac interstitial space due to an accumulation of ECM proteins. During this phenomenon, CFs proliferate, migrate and differentiate into myofibroblasts [163] under the effect of a variety of factors including transforming growth factor β1 (TGF-β1), tumor necrosis factor-α (TNF-α), Angiotensin II (Ang II), Platelet derived growth factor (PDGF), Endothelin 1 (ET-1)) and mechanical (stretch and stiffness) factors [116], [164]. Interestingly, these changes in the heart eventually lead to an increased matrix stiffness, structural damaging effect, electrical disorders [6] and predispose it to diastolic dysfunction [165], [166]. Moreover, intracellular Ca2+ signaling, involved in fibroblast proliferation and differentiation, is mainly activated in CFs through TRP channels, including TRPV4 channels [7], [52], [90], [118] [167].Indeed, accumulating evidence hints that TRPV4 mechanosensor is an important factor in the progression of fibrosis and/or prevention of fibroproliferative disorders in several organs such as skin, lung [168], liver, kidney, brain, blood vessel and heart [7]. The role of TRPV4 mechanotransduction in cardiac fibrosis was demonstrated using trpv4-/- mice model [81]. The results suggest that the TRPV4 deletion preserves cardiac function and reduces fibrosis in trpv4-/- mice but not in WT mice after transverse aortic constriction and MI [167]. Interestingly, trpv4-/- mice displayed a marked decrease in fibrosis, as well as a decrease in the pro-fibrotic gene expression level including col1A2, α-SMA, NFAT, TGF-β1, and MRTF, a mechanosensitive transcription factor. Conversely, the WT mice subjected to MI or congestive heart failure had an up-regulation of TRPV4 channels and profibrotic genes compared to the untreated WT mice which lead, in fine, to increase the intracellular Ca2+ level and amplify the pathological fibrosis process. These findings demonstrated that TRPV4 is functional in mouse CFs and directly related to fibrosis process insofar as its genetic deletion preserves cardiac function and protects the heart against the adverse fibrosis effects [167]. In line with these results, several in vitro investigations of rat or human CFs have shown a rapid but sustained increase in Ca2+ influx in response to TRPV4 agonist perfusion (4αPDD [118] or GSK1016790A [82]) which promotes the fibroblasts differentiation into myofibroblasts. Consistently, the blockade of TRPV4 by different antagonists (AB159908 [81] or RuR [118] or GSK2193874 [52]) or TRPV4 short-interference RNA (TRPV4-siRNA) [118] inhibits TGF-β-induced fibroblast differentiation [81] or abolishes Ca2+ conductance [118] or decreases the Ca2+ influx level [52], [116]. Recently, and in agreement with the previous investigations, treatment with the pro-fibrotic cytokine, TGF-β, increases TRPV4 channel expression in human ventricular CFs [117]. Simultaneously with this change, the authors noted both an increase in a fibrosis biomarker (the Plasminogen activator inhibitor-1) and an increase in a fibroblast differentiation biomarker (the α-smooth muscle actin : α-SMA) [117]. In addition, and via the MAPK/ERK pathway, TRPV4 channel activation by its specific agonist (GSK1016790A) triggers the CFs transformation into myofibroblasts while its antagonist (RN-9893) combined with a Ca2+ chelating agent (BAPTA-AM) reduces it [117]. These different points are important because they show that the Ca2+ permeability of TRPV4 channels is an essential component of human ventricular CFs differentiation. A recent investigation has demonstrated that TRPV4 channel expression level was higher in CFs isolated from Sprague-Dawley neonatal diabetic rats and from Sprague-Dawley adult rats, cultured in glucose-rich medium. Interestingly, the TRPV4 channel agonist HC067047 perfusion abolished significantly these CF changes. It has also been reported that HC067047 decreases the collagen Ι synthesis and suppresses the presence of the growth factor TGF-β necessary for the differentiation of CFs into myofibroblasts. In this context, different approaches could be used directly or indirectly to manage TRPV4 channel function and imagine new therapeutic strategies to limit fibrosis within cardiac diabetic patients [90] or in various cardiac pathologies involving many different patient populations. Finally, the link between fibrosis and the increased risk for atrial [52], [169], [170], [171] or ventricular [116] arrhythmias has been reported in several cardiac diseases [166], [172]. In this context, the anti-fibrosis approach via the TRPV4 inhibition constitutes a promising therapeutic way to limit adverse outcomes and prevent arrhythmias in the heart.
Taken together, these findings have clinical significance by identifying TRPV4 as an interesting therapeutic molecular target to attenuate cardiac fibrosis but still require further attention especially during arrhythmogenesis and pathological cardiac development.
6. Conclusions and perspectives
Evidence for a crucial role of TRPV4 channel has emerged from a large range of experiments in which its inhibition preserves the physiological intracellular Ca2+ dynamics and protects the heart form several dysfunctions including pathological cardiac disorders (Figure 2), HF and arrhythmia. Unfortunately, the molecular mechanism involving the transition from a healthy heart towards a cardiac pathology is poorly understood. The use of specific TRPV4 modulators combined to transgenic animal models has given valuable information about its involvement in physiological or pathological cardiovascular remodelling. Note that the recent availability of the crystal structure of TRPV4 channel will facilitate drug development to counteract diseases including potentially pathological cardiac phenotypes. Furthermore, various genetic diseases involving abnormal TRPV4 channel function have highlighted the key role of this ion channel in a broad spectrum of cellular processes. Surprisingly, no trpv4 mutation was found in patients with cardiac diseases so far. It would be relevant to further investigate the role of TRPV4 in the elderly population given the increase in its expression and the risk of cardiac diseases (ischemia-reperfusion lesion, MI, pressure-overload, arrhythmia) found in this population. Future work in this promising field may help to better understand the function and regulation of this channel and identify its interactions with other channels and the implication in cardiac physiological and pathological processes.
References
- Moran, M.M., H. Xu, and D.E. Clapham, TRP ion channels in the nervous system. Curr Opin Neurobiol, 2004. 14(3): p. 362-9. [CrossRef]
- Moran, M.M., TRP Channels as Potential Drug Targets. Annu Rev Pharmacol Toxicol, 2018. 58: p. 309-330. [CrossRef]
- Hof, T., et al., Transient receptor potential channels in cardiac health and disease. Nat Rev Cardiol, 2019. 16(6): p. 344-360. [CrossRef]
- White, J.P., et al., TRPV4: Molecular Conductor of a Diverse Orchestra. Physiol Rev, 2016. 96(3): p. 911-73. [CrossRef]
- Miller, M., et al., Role of Known Transient Receptor Potential Vanilloid Channels in Modulating Cardiac Mechanobiology. Front Physiol, 2021. 12: p. 734113. [CrossRef]
- Randhawa, P.K. and A.S. Jaggi, TRPV4 channels: physiological and pathological role in cardiovascular system. Basic Res Cardiol, 2015. 110(6): p. 54. [CrossRef]
- Inoue, R., L.H. Kurahara, and K. Hiraishi, TRP channels in cardiac and intestinal fibrosis. Semin Cell Dev Biol, 2019. 94: p. 40-49. [CrossRef]
- Chaigne, S., et al., Transient receptor potential vanilloid 4 channel participates in mouse ventricular electrical activity. Am J Physiol Heart Circ Physiol, 2021. 320(3): p. H1156-H1169. [CrossRef]
- Gorbunov, A.S., et al., Physiological and Pathological Role of TRPV1, TRPV2 and TRPV4 Channels in Heart. Curr Cardiol Rev, 2019. 15(4): p. 244-251. [CrossRef]
- Zhao, Y., et al., Unusual localization and translocation of TRPV4 protein in cultured ventricular myocytes of the neonatal rat. Eur J Histochem, 2012. 56(3): p. e32. [CrossRef]
- Jones, J.L., et al., TRPV4 increases cardiomyocyte calcium cycling and contractility yet contributes to damage in the aged heart following hypoosmotic stress. Cardiovasc Res, 2019. 115(1): p. 46-56. [CrossRef]
- Arniges, M., et al., Human TRPV4 channel splice variants revealed a key role of ankyrin domains in multimerization and trafficking. J Biol Chem, 2006. 281(3): p. 1580-6. [CrossRef]
- Shigematsu, H., et al., A 3.5-nm structure of rat TRPV4 cation channel revealed by Zernike phase-contrast cryoelectron microscopy. J Biol Chem, 2010. 285(15): p. 11210-8. [CrossRef]
- Inada, H., et al., Structural and biochemical consequences of disease-causing mutations in the ankyrin repeat domain of the human TRPV4 channel. Biochemistry, 2012. 51(31): p. 6195-206. [CrossRef]
- Takahashi, N., et al., TRPV4 channel activity is modulated by direct interaction of the ankyrin domain to PI(4,5)P(2). Nat Commun, 2014. 5: p. 4994. [CrossRef]
- Deng, Z., et al., Cryo-EM and X-ray structures of TRPV4 reveal insight into ion permeation and gating mechanisms. Nat Struct Mol Biol, 2018. 25(3): p. 252-260. [CrossRef]
- Nilius, B. and T. Voets, The puzzle of TRPV4 channelopathies. EMBO Rep, 2013. 14(2): p. 152-63. [CrossRef]
- Hellwig, N., et al., Homo- and heteromeric assembly of TRPV channel subunits. J Cell Sci, 2005. 118(Pt 5): p. 917-28. [CrossRef]
- Jin, X., J. Touhey, and R. Gaudet, Structure of the N-terminal ankyrin repeat domain of the TRPV2 ion channel. J Biol Chem, 2006. 281(35): p. 25006-10. [CrossRef]
- Liao, M., et al., Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature, 2013. 504(7478): p. 107-12. [CrossRef]
- Lishko, P.V., et al., The ankyrin repeats of TRPV1 bind multiple ligands and modulate channel sensitivity. Neuron, 2007. 54(6): p. 905-18. [CrossRef]
- McCleverty, C.J., et al., Crystal structure of the human TRPV2 channel ankyrin repeat domain. Protein Sci, 2006. 15(9): p. 2201-6. [CrossRef]
- Michalick, L. and W.M. Kuebler, TRPV4-A Missing Link Between Mechanosensation and Immunity. Front Immunol, 2020. 11: p. 413. [CrossRef]
- D'Hoedt, D., et al., Stimulus-specific modulation of the cation channel TRPV4 by PACSIN 3. J Biol Chem, 2008. 283(10): p. 6272-80. [CrossRef]
- Wang, Y., et al., OS-9 regulates the transit and polyubiquitination of TRPV4 in the endoplasmic reticulum. J Biol Chem, 2007. 282(50): p. 36561-70. [CrossRef]
- Toft-Bertelsen, T.L., B.R. Larsen, and N. MacAulay, Sensing and regulation of cell volume - we know so much and yet understand so little: TRPV4 as a sensor of volume changes but possibly without a volume-regulatory role? Channels (Austin), 2018. 12(1): p. 100-108. [CrossRef]
- Strotmann, R., G. Schultz, and T.D. Plant, Ca2+-dependent potentiation of the nonselective cation channel TRPV4 is mediated by a C-terminal calmodulin binding site. J Biol Chem, 2003. 278(29): p. 26541-9. [CrossRef]
- Loukin, S.H., J. Teng, and C. Kung, A channelopathy mechanism revealed by direct calmodulin activation of TrpV4. Proc Natl Acad Sci U S A, 2015. 112(30): p. 9400-5. [CrossRef]
- Suzuki, M., A. Hirao, and A. Mizuno, Microtubule-associated [corrected] protein 7 increases the membrane expression of transient receptor potential vanilloid 4 (TRPV4). J Biol Chem, 2003. 278(51): p. 51448-53. [CrossRef]
- Stewart, A.P., et al., Atomic force microscopy reveals the alternating subunit arrangement of the TRPP2-TRPV4 heterotetramer. Biophys J, 2010. 99(3): p. 790-7. [CrossRef]
- Ma, X., et al., Heteromeric TRPV4-C1 channels contribute to store-operated Ca(2+) entry in vascular endothelial cells. Cell Calcium, 2011. 50(6): p. 502-9. [CrossRef]
- Du, J., et al., TRPV4, TRPC1, and TRPP2 assemble to form a flow-sensitive heteromeric channel. FASEB J, 2014. 28(11): p. 4677-85. [CrossRef]
- Benfenati, V., et al., An aquaporin-4/transient receptor potential vanilloid 4 (AQP4/TRPV4) complex is essential for cell-volume control in astrocytes. Proc Natl Acad Sci U S A, 2011. 108(6): p. 2563-8. [CrossRef]
- Liu, X., et al., A role for AQP5 in activation of TRPV4 by hypotonicity: concerted involvement of AQP5 and TRPV4 in regulation of cell volume recovery. J Biol Chem, 2006. 281(22): p. 15485-95. [CrossRef]
- Galizia, L., et al., Functional interaction between AQP2 and TRPV4 in renal cells. J Cell Biochem, 2012. 113(2): p. 580-9. [CrossRef]
- Verkerk, A.O., E.M. Lodder, and R. Wilders, Aquaporin Channels in the Heart-Physiology and Pathophysiology. Int J Mol Sci, 2019. 20(8). [CrossRef]
- Shin, S.H., et al., Phosphorylation on TRPV4 Serine 824 Regulates Interaction with STIM1. Open Biochem J, 2015. 9: p. 24-33. [CrossRef]
- Shukla, A.K., et al., Arresting a transient receptor potential (TRP) channel: beta-arrestin 1 mediates ubiquitination and functional down-regulation of TRPV4. J Biol Chem, 2010. 285(39): p. 30115-25. [CrossRef]
- Ma, X., et al., Electrophysiological properties of heteromeric TRPV4-C1 channels. Biochim Biophys Acta, 2011. 1808(12): p. 2789-97. [CrossRef]
- Kottgen, M., et al., TRPP2 and TRPV4 form a polymodal sensory channel complex. J Cell Biol, 2008. 182(3): p. 437-47. [CrossRef]
- Brauchi, S. and P. Orio, Voltage sensing in thermo-TRP channels. Adv Exp Med Biol, 2011. 704: p. 517-30. [CrossRef]
- Long, S.B., E.B. Campbell, and R. Mackinnon, Voltage sensor of Kv1.2: structural basis of electromechanical coupling. Science, 2005. 309(5736): p. 903-8. [CrossRef]
- Long, S.B., E.B. Campbell, and R. Mackinnon, Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science, 2005. 309(5736): p. 897-903. [CrossRef]
- Whicher, J.R. and R. MacKinnon, Structure of the voltage-gated K(+) channel Eag1 reveals an alternative voltage sensing mechanism. Science, 2016. 353(6300): p. 664-9. [CrossRef]
- Lee, C.H. and R. MacKinnon, Structures of the Human HCN1 Hyperpolarization-Activated Channel. Cell, 2017. 168(1-2): p. 111-120 e11. [CrossRef]
- Watanabe, H., et al., Heat-evoked activation of TRPV4 channels in a HEK293 cell expression system and in native mouse aorta endothelial cells. J Biol Chem, 2002. 277(49): p. 47044-51. [CrossRef]
- Voets, T., et al., Molecular determinants of permeation through the cation channel TRPV4. J Biol Chem, 2002. 277(37): p. 33704-10. [CrossRef]
- Nilius, B., et al., TRPV4 calcium entry channel: a paradigm for gating diversity. Am J Physiol Cell Physiol, 2004. 286(2): p. C195-205. [CrossRef]
- Lawhorn, B.G., E.J. Brnardic, and D.J. Behm, TRPV4 antagonists: a patent review (2015-2020). Expert Opin Ther Pat, 2021: p. 1-12. [CrossRef]
- Coetzee, W.A., Channel-mediated calcium current in the heart. Cardiovasc Drugs Ther, 1988. 1(5): p. 447-59. [CrossRef]
- Bers, D.M., Cardiac excitation-contraction coupling. Nature, 2002. 415(6868): p. 198-205. [CrossRef]
- Liao, J., et al., TRPV4 blockade suppresses atrial fibrillation in sterile pericarditis rats. JCI Insight, 2020. 5(23). [CrossRef]
- Goyal, N., et al., Clinical Pharmacokinetics, Safety, and Tolerability of a Novel, First-in-Class TRPV4 Ion Channel Inhibitor, GSK2798745, in Healthy and Heart Failure Subjects. Am J Cardiovasc Drugs, 2019. 19(3): p. 335-342. [CrossRef]
- Lawhorn, B.G., E.J. Brnardic, and D.J. Behm, Recent advances in TRPV4 agonists and antagonists. Bioorg Med Chem Lett, 2020. 30(8): p. 127022. [CrossRef]
- Thorneloe, K.S., et al., PROPERTIES OF THE TRPV4 AGONIST GSK1016790A AND the TRPV4 ANTAGONIST GSK2193874. Physiol Rev, 2017. 97(4): p. 1231-1232. [CrossRef]
- Wu, Q.F., et al., Activation of transient receptor potential vanilloid 4 involves in hypoxia/reoxygenation injury in cardiomyocytes. Cell Death Dis, 2017. 8(5): p. e2828. [CrossRef]
- Baratchi, S., et al., The TRPV4 Agonist GSK1016790A Regulates the Membrane Expression of TRPV4 Channels. Front Pharmacol, 2019. 10: p. 6. [CrossRef]
- Atobe, M., et al., Discovery of Novel Transient Receptor Potential Vanilloid 4 (TRPV4) Agonists as Regulators of Chondrogenic Differentiation: Identification of Quinazolin-4(3 H)-ones and in Vivo Studies on a Surgically Induced Rat Model of Osteoarthritis. J Med Chem, 2019. 62(3): p. 1468-1483. [CrossRef]
- Grace, M.S., et al., Modulation of the TRPV4 ion channel as a therapeutic target for disease. Pharmacol Ther, 2017. 177: p. 9-22. [CrossRef]
- Vincent, F., et al., Identification and characterization of novel TRPV4 modulators. Biochem Biophys Res Commun, 2009. 389(3): p. 490-4. [CrossRef]
- Dong, Q., et al., Blockage of transient receptor potential vanilloid 4 alleviates myocardial ischemia/reperfusion injury in mice. Sci Rep, 2017. 7: p. 42678. [CrossRef]
- Hilfiker, M.A., et al., Optimization of a Novel Series of TRPV4 Antagonists with In Vivo Activity in a Model of Pulmonary Edema. ACS Med Chem Lett, 2013. 4(2): p. 293-6. [CrossRef]
- Wei, Z.L., et al., Identification of orally-bioavailable antagonists of the TRPV4 ion-channel. Bioorg Med Chem Lett, 2015. 25(18): p. 4011-5. [CrossRef]
- Brooks, C.A., et al., Discovery of GSK3527497: A Candidate for the Inhibition of Transient Receptor Potential Vanilloid-4 (TRPV4). J Med Chem, 2019. 62(20): p. 9270-9280. [CrossRef]
- Tsuno, N., et al., Discovery of novel 2',4'-dimethyl-[4,5'-bithiazol]-2-yl amino derivatives as orally bioavailable TRPV4 antagonists for the treatment of pain: Part 1. Bioorg Med Chem Lett, 2016. 26(20): p. 4930-4935. [CrossRef]
- Tsuno, N., et al., Discovery of novel 2',4'-dimethyl-[4,5'-bithiazol]-2-yl amino derivatives as orally bioavailable TRPV4 antagonists for the treatment of pain: Part 2. Bioorg Med Chem Lett, 2016. 26(20): p. 4936-4941. [CrossRef]
- Pero, J.E., et al., Design and Optimization of Sulfone Pyrrolidine Sulfonamide Antagonists of Transient Receptor Potential Vanilloid-4 with in Vivo Activity in a Pulmonary Edema Model. J Med Chem, 2018. 61(24): p. 11209-11220. [CrossRef]
- Xu, F., E. Satoh, and T. Iijima, Protein kinase C-mediated Ca2+ entry in HEK 293 cells transiently expressing human TRPV4. Br J Pharmacol, 2003. 140(2): p. 413-21. [CrossRef]
- A.J.Duncton, M., Chapter 12 - Small Molecule Agonists and Antagonists of TRPV4. TRP Channels as Therapeutic Targets From Basic Science to Clinical Use 2015: p. 205-219.
- Alexander, R., et al., 4alpha-phorbol 12,13-didecanoate activates cultured mouse dorsal root ganglia neurons independently of TRPV4. Br J Pharmacol, 2013. 168(3): p. 761-72. [CrossRef]
- Dahan, D., et al., Implication of the ryanodine receptor in TRPV4-induced calcium response in pulmonary arterial smooth muscle cells from normoxic and chronically hypoxic rats. Am J Physiol Lung Cell Mol Physiol, 2012. 303(9): p. L824-33. [CrossRef]
- Watanabe, H., et al., Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature, 2003. 424(6947): p. 434-8. [CrossRef]
- Katragadda, D., et al., Epoxyeicosatrienoic acids limit damage to mitochondrial function following stress in cardiac cells. J Mol Cell Cardiol, 2009. 46(6): p. 867-75. [CrossRef]
- Lai, J. and C. Chen, The Role of Epoxyeicosatrienoic Acids in Cardiac Remodeling. Front Physiol, 2021. 12: p. 642470. [CrossRef]
- Yang, L., et al., The role of epoxyeicosatrienoic acids in the cardiovascular system. Br J Clin Pharmacol, 2015. 80(1): p. 28-44. [CrossRef]
- Thorneloe, K.S., et al., An orally active TRPV4 channel blocker prevents and resolves pulmonary edema induced by heart failure. Sci Transl Med, 2012. 4(159): p. 159ra148. [CrossRef]
- Xu, S., et al., A novel TRPV4-specific agonist inhibits monocyte adhesion and atherosclerosis. Oncotarget, 2016. 7(25): p. 37622-37635. [CrossRef]
- Pankey, E.A., et al., Analysis of responses to the TRPV4 agonist GSK1016790A in the pulmonary vascular bed of the intact-chest rat. Am J Physiol Heart Circ Physiol, 2014. 306(1): p. H33-40. [CrossRef]
- Zhang, S., et al., Activation of transient receptor potential vanilloid 4 exacerbates myocardial ischemia-reperfusion injury via JNK-CaMKII phosphorylation pathway in isolated mice hearts. Cell Calcium, 2021. 100: p. 102483. [CrossRef]
- Willette, R.N., et al., Systemic activation of the transient receptor potential vanilloid subtype 4 channel causes endothelial failure and circulatory collapse: Part 2. J Pharmacol Exp Ther, 2008. 326(2): p. 443-52. [CrossRef]
- Adapala, R.K., et al., TRPV4 channels mediate cardiac fibroblast differentiation by integrating mechanical and soluble signals. J Mol Cell Cardiol, 2013. 54: p. 45-52. [CrossRef]
- Ahn, M.S., et al., Transient receptor potential channel TRPV4 mediates TGF-beta1-induced differentiation of human ventricular fibroblasts. Cardiol J, 2020. [CrossRef]
- Donate-Macian, P., et al., Structural determinants of TRPV4 inhibition and identification of new antagonists with antiviral activity. Br J Pharmacol, 2022. 179(14): p. 3576-3591. [CrossRef]
- Filosa, J.A., X. Yao, and G. Rath, TRPV4 and the regulation of vascular tone. J Cardiovasc Pharmacol, 2013. 61(2): p. 113-9. [CrossRef]
- Greenberg, H.Z.E., et al., Heteromeric TRPV4/TRPC1 channels mediate calcium-sensing receptor-induced nitric oxide production and vasorelaxation in rabbit mesenteric arteries. Vascul Pharmacol, 2017. 96-98: p. 53-62. [CrossRef]
- Lu, J., et al., An abnormal TRPV4-related cytosolic Ca(2+) rise in response to uniaxial stretch in induced pluripotent stem cells-derived cardiomyocytes from dilated cardiomyopathy patients. Biochim Biophys Acta Mol Basis Dis, 2017. 1863(11): p. 2964-2972. [CrossRef]
- Xia, Y., et al., TRPV4 channel contributes to serotonin-induced pulmonary vasoconstriction and the enhanced vascular reactivity in chronic hypoxic pulmonary hypertension. Am J Physiol Cell Physiol, 2013. 305(7): p. C704-15. [CrossRef]
- Gevaert, T., et al., Deletion of the transient receptor potential cation channel TRPV4 impairs murine bladder voiding. J Clin Invest, 2007. 117(11): p. 3453-62. [CrossRef]
- Vizin, R.C., et al., TRPV4 activates autonomic and behavioural warmth-defence responses in Wistar rats. Acta Physiol (Oxf), 2015. 214(2): p. 275-89. [CrossRef]
- Jia, X., et al., TRPV4 Mediates Cardiac Fibrosis via the TGF-beta1/Smad3 Signaling Pathway in Diabetic Rats. Cardiovasc Toxicol, 2020. [CrossRef]
- Veteto, A.B., et al., TRPV4 Contributes to Stretch-Induced Hypercontractility and Time-Dependent Dysfunction in the Aged Heart. Cardiovasc Res, 2019. [CrossRef]
- Cheung, M., et al., Discovery of GSK2193874: An Orally Active, Potent, and Selective Blocker of Transient Receptor Potential Vanilloid 4. ACS Med Chem Lett, 2017. 8(5): p. 549-554. [CrossRef]
- O'Brien, F., C.A. Staunton, and R. Barrett-Jolley, Systemic application of the transient receptor potential vanilloid-type 4 antagonist GSK2193874 induces tail vasodilation in a mouse model of thermoregulation. Biol Lett, 2022. 18(6): p. 20220129. [CrossRef]
- Pero, J.E., et al., Identification, Synthesis, and Characterization of a Major Circulating Human Metabolite of TRPV4 Antagonist GSK2798745. ACS Med Chem Lett, 2021. 12(9): p. 1498-1502. [CrossRef]
- Brnardic, E.J., et al., Discovery of Pyrrolidine Sulfonamides as Selective and Orally Bioavailable Antagonists of Transient Receptor Potential Vanilloid-4 (TRPV4). J Med Chem, 2018. 61(21): p. 9738-9755. [CrossRef]
- Suzuki, M., et al., Impaired pressure sensation in mice lacking TRPV4. J Biol Chem, 2003. 278(25): p. 22664-8. [CrossRef]
- Liedtke, W. and J.M. Friedman, Abnormal osmotic regulation in trpv4-/- mice. Proc Natl Acad Sci U S A, 2003. 100(23): p. 13698-703. [CrossRef]
- Liu, L., et al., Role of Transient Receptor Potential Vanilloid 4 in Vascular Function. Front Mol Biosci, 2021. 8: p. 677661. [CrossRef]
- Zhang, D.X., et al., Transient receptor potential vanilloid type 4-deficient mice exhibit impaired endothelium-dependent relaxation induced by acetylcholine in vitro and in vivo. Hypertension, 2009. 53(3): p. 532-8. [CrossRef]
- Vriens, J., et al., Modulation of the Ca2 permeable cation channel TRPV4 by cytochrome P450 epoxygenases in vascular endothelium. Circ Res, 2005. 97(9): p. 908-15. [CrossRef]
- Mendoza, S.A., et al., TRPV4-mediated endothelial Ca2+ influx and vasodilation in response to shear stress. Am J Physiol Heart Circ Physiol, 2010. 298(2): p. H466-76. [CrossRef]
- Sonkusare, S.K., et al., Elementary Ca2+ signals through endothelial TRPV4 channels regulate vascular function. Science, 2012. 336(6081): p. 597-601. [CrossRef]
- Earley, S., et al., TRPV4-dependent dilation of peripheral resistance arteries influences arterial pressure. Am J Physiol Heart Circ Physiol, 2009. 297(3): p. H1096-102. [CrossRef]
- Loot, A.E., et al., Role of cytochrome P450-dependent transient receptor potential V4 activation in flow-induced vasodilatation. Cardiovasc Res, 2008. 80(3): p. 445-52. [CrossRef]
- Hartmannsgruber, V., et al., Arterial response to shear stress critically depends on endothelial TRPV4 expression. PLoS One, 2007. 2(9): p. e827. [CrossRef]
- Mizuno, A., et al., Impaired osmotic sensation in mice lacking TRPV4. Am J Physiol Cell Physiol, 2003. 285(1): p. C96-101. [CrossRef]
- Tabuchi, K., et al., Hearing impairment in TRPV4 knockout mice. Neurosci Lett, 2005. 382(3): p. 304-8. [CrossRef]
- Alvarez, D.F., et al., Transient receptor potential vanilloid 4-mediated disruption of the alveolar septal barrier: a novel mechanism of acute lung injury. Circ Res, 2006. 99(9): p. 988-95. [CrossRef]
- Taniguchi, J., et al., TRPV4 as a flow sensor in flow-dependent K+ secretion from the cortical collecting duct. Am J Physiol Renal Physiol, 2007. 292(2): p. F667-73. [CrossRef]
- Gualdani, R., et al., Mechanical activation of TRPV4 channels controls albumin reabsorption by proximal tubule cells. Sci Signal, 2020. 13(653). [CrossRef]
- Masuyama, R., et al., TRPV4-mediated calcium influx regulates terminal differentiation of osteoclasts. Cell Metab, 2008. 8(3): p. 257-65. [CrossRef]
- Seghers, F., et al., TRPV4 participates in pressure-induced inhibition of renin secretion by juxtaglomerular cells. J Physiol, 2016. 594(24): p. 7327-7340. [CrossRef]
- Kunert-Keil, C., et al., Tissue-specific expression of TRP channel genes in the mouse and its variation in three different mouse strains. BMC Genomics, 2006. 7: p. 159. [CrossRef]
- Veteto, A.B., et al., Transient receptor potential vanilloid-4 contributes to stretch-induced hypercontractility and time-dependent dysfunction in the aged heart. Cardiovasc Res, 2020. 116(11): p. 1887-1896. [CrossRef]
- Qi, Y., et al., Uniaxial cyclic stretch stimulates TRPV4 to induce realignment of human embryonic stem cell-derived cardiomyocytes. J Mol Cell Cardiol, 2015. 87: p. 65-73. [CrossRef]
- Thodeti, C.K., S. Paruchuri, and J.G. Meszaros, A TRP to cardiac fibroblast differentiation. Channels (Austin), 2013. 7(3): p. 211-4. [CrossRef]
- Ahn, M.S., et al., Transient receptor potential channel TRPV4 mediates TGF-beta1-induced differentiation of human ventricular fibroblasts. Cardiol J, 2020. 27(2): p. 162-170. [CrossRef]
- Hatano, N., Y. Itoh, and K. Muraki, Cardiac fibroblasts have functional TRPV4 activated by 4alpha-phorbol 12,13-didecanoate. Life Sci, 2009. 85(23-26): p. 808-14. [CrossRef]
- Du, J., et al., TRPM7-mediated Ca2+ signals confer fibrogenesis in human atrial fibrillation. Circ Res, 2010. 106(5): p. 992-1003. [CrossRef]
- Kohler, R., et al., Evidence for a functional role of endothelial transient receptor potential V4 in shear stress-induced vasodilatation. Arterioscler Thromb Vasc Biol, 2006. 26(7): p. 1495-502. [CrossRef]
- Sullivan, M.N., et al., Optical recording reveals novel properties of GSK1016790A-induced vanilloid transient receptor potential channel TRPV4 activity in primary human endothelial cells. Mol Pharmacol, 2012. 82(3): p. 464-72. [CrossRef]
- Sukumaran, S.V., et al., TRPV4 channel activation leads to endothelium-dependent relaxation mediated by nitric oxide and endothelium-derived hyperpolarizing factor in rat pulmonary artery. Pharmacol Res, 2013. 78: p. 18-27. [CrossRef]
- Bubolz, A.H., et al., Activation of endothelial TRPV4 channels mediates flow-induced dilation in human coronary arterioles: role of Ca2+ entry and mitochondrial ROS signaling. Am J Physiol Heart Circ Physiol, 2012. 302(3): p. H634-42. [CrossRef]
- Barbeau, S., et al., Mechanosensitivity in Pulmonary Circulation: Pathophysiological Relevance of Stretch-Activated Channels in Pulmonary Hypertension. Biomolecules, 2021. 11(9). [CrossRef]
- Yang, X.R., et al., Upregulation of osmo-mechanosensitive TRPV4 channel facilitates chronic hypoxia-induced myogenic tone and pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol, 2012. 302(6): p. L555-68. [CrossRef]
- Parpaite, T., et al., Effect of hypoxia on TRPV1 and TRPV4 channels in rat pulmonary arterial smooth muscle cells. Pflugers Arch, 2016. 468(1): p. 111-130. [CrossRef]
- Baylie, R.L. and J.E. Brayden, TRPV channels and vascular function. Acta Physiol (Oxf), 2011. 203(1): p. 99-116. [CrossRef]
- Ducret, T., et al., Serotonin-induced activation of TRPV4-like current in rat intrapulmonary arterial smooth muscle cells. Cell Calcium, 2008. 43(4): p. 315-23. [CrossRef]
- Song, S., et al., Flow shear stress enhances intracellular Ca2+ signaling in pulmonary artery smooth muscle cells from patients with pulmonary arterial hypertension. Am J Physiol Cell Physiol, 2014. 307(4): p. C373-83. [CrossRef]
- Ottolini, M., et al., Mechanisms underlying selective coupling of endothelial Ca(2+) signals with eNOS vs. IK/SK channels in systemic and pulmonary arteries. J Physiol, 2020. 598(17): p. 3577-3596. [CrossRef]
- Doleschal, B., et al., TRPC3 contributes to regulation of cardiac contractility and arrhythmogenesis by dynamic interaction with NCX1. Cardiovasc Res, 2015. 106(1): p. 163-73. [CrossRef]
- Qi, Z., et al., TRPC3 regulates the automaticity of embryonic stem cell-derived cardiomyocytes. Int J Cardiol, 2016. 203: p. 169-81. [CrossRef]
- Guinamard, R., et al., TRPM4 in cardiac electrical activity. Cardiovasc Res, 2015. 108(1): p. 21-30. [CrossRef]
- Hof, T., et al., TRPM4 non-selective cation channels influence action potentials in rabbit Purkinje fibres. J Physiol, 2016. 594(2): p. 295-306. [CrossRef]
- Hof, T., et al., Implication of the TRPM4 nonselective cation channel in mammalian sinus rhythm. Heart Rhythm, 2013. 10(11): p. 1683-9. [CrossRef]
- Sah, R., et al., Ion channel-kinase TRPM7 is required for maintaining cardiac automaticity. Proc Natl Acad Sci U S A, 2013. 110(32): p. E3037-46. [CrossRef]
- Saito, Y., et al., TRPM4 Mutation in Patients With Ventricular Noncompaction and Cardiac Conduction Disease. Circ Genom Precis Med, 2018. 11(5): p. e002103. [CrossRef]
- Bianchi, B., et al., Four TRPM4 Cation Channel Mutations Found in Cardiac Conduction Diseases Lead to Altered Protein Stability. Front Physiol, 2018. 9: p. 177. [CrossRef]
- Liu, H., et al., Gain-of-function mutations in TRPM4 cause autosomal dominant isolated cardiac conduction disease. Circ Cardiovasc Genet, 2010. 3(4): p. 374-85. [CrossRef]
- Stallmeyer, B., et al., Mutational spectrum in the Ca(2+)--activated cation channel gene TRPM4 in patients with cardiac conductance disturbances. Hum Mutat, 2012. 33(1): p. 109-17. [CrossRef]
- Palladino, A., et al., The Role of TRPM4 Gene Mutations in Causing Familial Progressive Cardiac Conduction Disease: A Further Contribution. Genes (Basel), 2022. 13(2). [CrossRef]
- Demion, M., et al., Trpm4 gene invalidation leads to cardiac hypertrophy and electrophysiological alterations. PLoS One, 2014. 9(12): p. e115256. [CrossRef]
- Jin, M., et al., Determinants of TRPV4 activity following selective activation by small molecule agonist GSK1016790A. PLoS One, 2011. 6(2): p. e16713. [CrossRef]
- Eisner, D.A., et al., Calcium and Excitation-Contraction Coupling in the Heart. Circ Res, 2017. 121(2): p. 181-195. [CrossRef]
- Abi-Samra, F. and D. Gutterman, Cardiac contractility modulation: a novel approach for the treatment of heart failure. Heart Fail Rev, 2016. 21(6): p. 645-660. [CrossRef]
- Ezeani, M., TRP Channels Mediated Pathological Ca2+-Handling and Spontaneous Ectopy. Front Cardiovasc Med. 2019; 6: 83. , 2020. [CrossRef]
- Freichel, M., et al., TRP Channels in the Heart, in Neurobiology of TRP Channels, T.L.R. Emir, Editor. 2017: Boca Raton (FL). p. 149-185.
- Li, J., et al., Role of transient receptor potential vanilloid 4 in the effect of osmotic pressure on myocardial contractility in rat. Sheng Li Xue Bao, 2008. 60(2): p. 181-8.
- Wu, Q., et al., Blockade of Transient Receptor Potential Vanilloid 4 Enhances Antioxidation after Myocardial Ischemia/Reperfusion. Oxid Med Cell Longev, 2019. 2019: p. 7283683. [CrossRef]
- Gregory H Turner1, W.B., Beat M Jucker2, John J Lepore2, Robert N Willette2, and Kevin S Thorneloe2, Preservation of Cardiac Function and Attenuation of Remodelling in Transient Receptor Potential Vanilloid 4 Knockout Mice Following Myocardial Infarction. Clinical & Experimental Cardiology, 2015. 6(366).
- Rosenbaum, T., et al., TRPV4: A Physio and Pathophysiologically Significant Ion Channel. Int J Mol Sci, 2020. 21(11). [CrossRef]
- Chen, Y.L. and S.K. Sonkusare, Endothelial TRPV4 channels and vasodilator reactivity. Curr Top Membr, 2020. 85: p. 89-117. [CrossRef]
- Geng, L., et al., Physiological levels of fluid shear stress modulate vascular function through TRPV4 sparklets. Acta Biochim Biophys Sin (Shanghai), 2022. 54(9): p. 1268-1277. [CrossRef]
- Barbeau, S., et al., Cell Confluence Modulates TRPV4 Channel Activity in Response to Hypoxia. Biomolecules, 2022. 12(7). [CrossRef]
- Earley, S., et al., TRPV4 forms a novel Ca2+ signaling complex with ryanodine receptors and BKCa channels. Circ Res, 2005. 97(12): p. 1270-9. [CrossRef]
- Suresh, K., et al., Reactive oxygen species induced Ca(2+) influx via TRPV4 and microvascular endothelial dysfunction in the SU5416/hypoxia model of pulmonary arterial hypertension. Am J Physiol Lung Cell Mol Physiol, 2018. 314(5): p. L893-L907. [CrossRef]
- Morine, K.J., et al., Endoglin selectively modulates transient receptor potential channel expression in left and right heart failure. Cardiovasc Pathol, 2016. 25(6): p. 478-482. [CrossRef]
- Zou, Y., et al., Activation of transient receptor potential vanilloid 4 is involved in pressure overload-induced cardiac hypertrophy. Elife, 2022. 11. [CrossRef]
- Connell, P., T.A. Word, and X.H.T. Wehrens, Targeting pathological leak of ryanodine receptors: preclinical progress and the potential impact on treatments for cardiac arrhythmias and heart failure. Expert Opin Ther Targets, 2020. 24(1): p. 25-36. [CrossRef]
- Peana, D., L. Polo-Parada, and T.L. Domeier, Arrhythmogenesis in the aged heart following ischaemia-reperfusion: role of transient receptor potential vanilloid 4. Cardiovasc Res, 2022. 118(4): p. 1126-1137. [CrossRef]
- van Nieuwenhoven, F.A. and N.A. Turner, The role of cardiac fibroblasts in the transition from inflammation to fibrosis following myocardial infarction. Vascul Pharmacol, 2013. 58(3): p. 182-8. [CrossRef]
- Banerjee, I., et al., Dynamic interactions between myocytes, fibroblasts, and extracellular matrix. Ann N Y Acad Sci, 2006. 1080: p. 76-84. [CrossRef]
- Disertori, M., M. Mase, and F. Ravelli, Myocardial fibrosis predicts ventricular tachyarrhythmias. Trends Cardiovasc Med, 2017. 27(5): p. 363-372. [CrossRef]
- Adapala, R.K., et al., TRPV4 Mechanotransduction in Fibrosis. Cells, 2021. 10(11). [CrossRef]
- Biernacka, A. and N.G. Frangogiannis, Aging and Cardiac Fibrosis. Aging Dis, 2011. 2(2): p. 158-173.
- Frangogiannis, N.G., Cardiac fibrosis. Cardiovasc Res, 2021. 117(6): p. 1450-1488.
- Adapala, R.K., et al., TRPV4 deletion protects heart from myocardial infarction-induced adverse remodeling via modulation of cardiac fibroblast differentiation. Basic Res Cardiol, 2020. 115(2): p. 14. [CrossRef]
- Cussac, L.A., et al., TRPV4 channel mediates adventitial fibroblast activation and adventitial remodeling in pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol, 2020. 318(1): p. L135-L146. [CrossRef]
- Verheule, S., et al., Increased vulnerability to atrial fibrillation in transgenic mice with selective atrial fibrosis caused by overexpression of TGF-beta1. Circ Res, 2004. 94(11): p. 1458-65. [CrossRef]
- Verheule, S., et al., Fibrillatory conduction in the atrial free walls of goats in persistent and permanent atrial fibrillation. Circ Arrhythm Electrophysiol, 2010. 3(6): p. 590-9. [CrossRef]
- Choi, E.K., et al., Triggered firing and atrial fibrillation in transgenic mice with selective atrial fibrosis induced by overexpression of TGF-beta1. Circ J, 2012. 76(6): p. 1354-62. [CrossRef]
- Verheule, S. and U. Schotten, Electrophysiological Consequences of Cardiac Fibrosis. Cells, 2021. 10(11). [CrossRef]
Figure 1.
Schematic view of TRPV channel structures and associated ion permeation pathway. (A). Comparison of TRPV pore structures. For clarity, only two opposing subunits are shown (segment S6). Interestingly the TRPV4 upper gate is larger compared to the other members of TRPV family (TRPV1, TRPV2 and TRPV5). (B) Homotetrameric structure of TRPV4. (C) Functional characteristics of TRPV channels (permeability ratio PCa/PNa).
Figure 1.
Schematic view of TRPV channel structures and associated ion permeation pathway. (A). Comparison of TRPV pore structures. For clarity, only two opposing subunits are shown (segment S6). Interestingly the TRPV4 upper gate is larger compared to the other members of TRPV family (TRPV1, TRPV2 and TRPV5). (B) Homotetrameric structure of TRPV4. (C) Functional characteristics of TRPV channels (permeability ratio PCa/PNa).
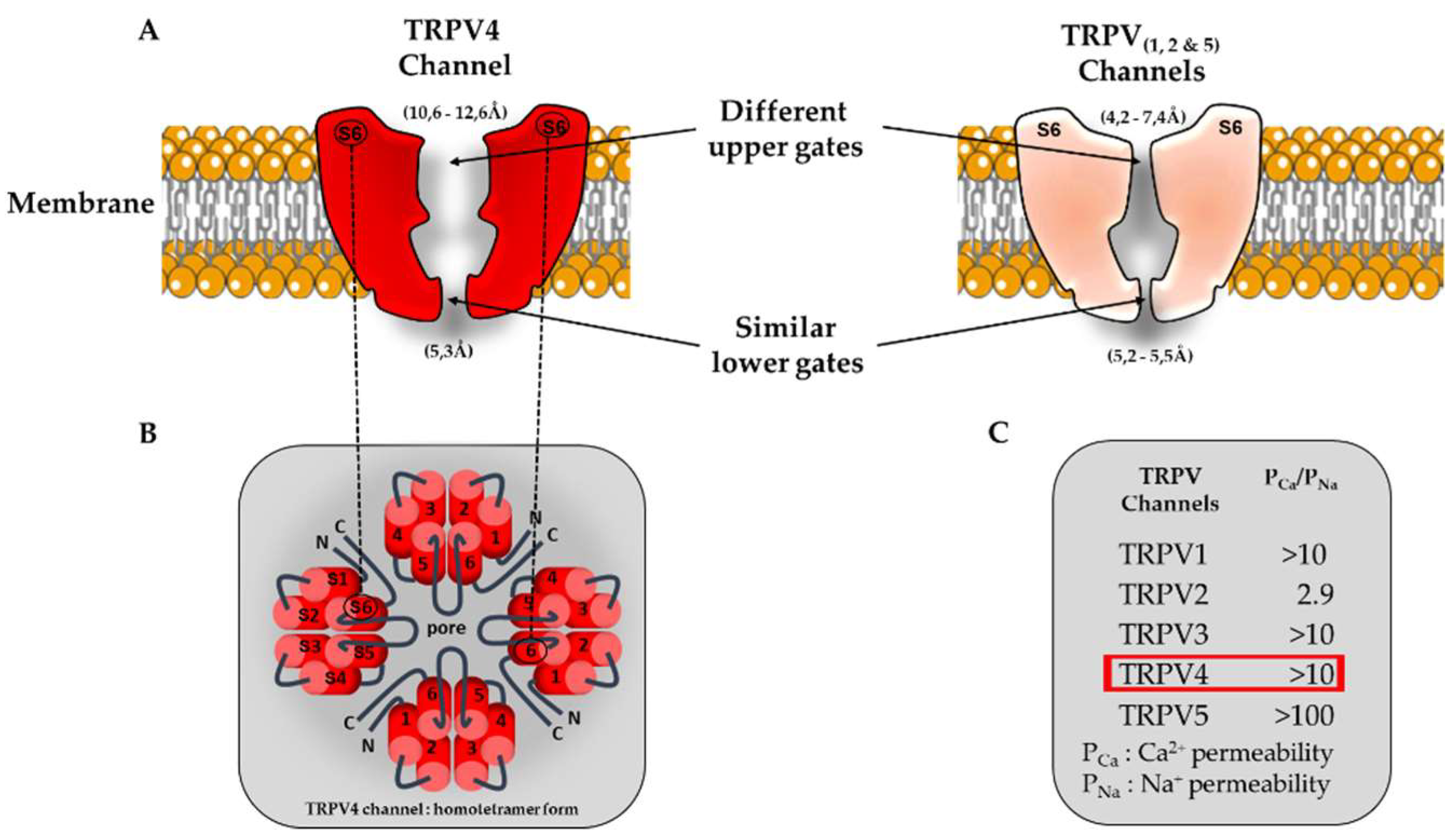
Figure 2.
TRPV4 expression and function model in healthy (left) and pathologic (right) cardiac cells. Under physiological condition, TRPV4 channels (yellow channels) present a low expression at the plasma membrane of cardiac cells. With aging, pressure overload, ischemia/reperfusion and others deleterious factors, TRPV4 exhibits both an increase in expression and activity at the plasma membrane of cardiac cells which tend progression towards of cardiovascular diseases.
Figure 2.
TRPV4 expression and function model in healthy (left) and pathologic (right) cardiac cells. Under physiological condition, TRPV4 channels (yellow channels) present a low expression at the plasma membrane of cardiac cells. With aging, pressure overload, ischemia/reperfusion and others deleterious factors, TRPV4 exhibits both an increase in expression and activity at the plasma membrane of cardiac cells which tend progression towards of cardiovascular diseases.
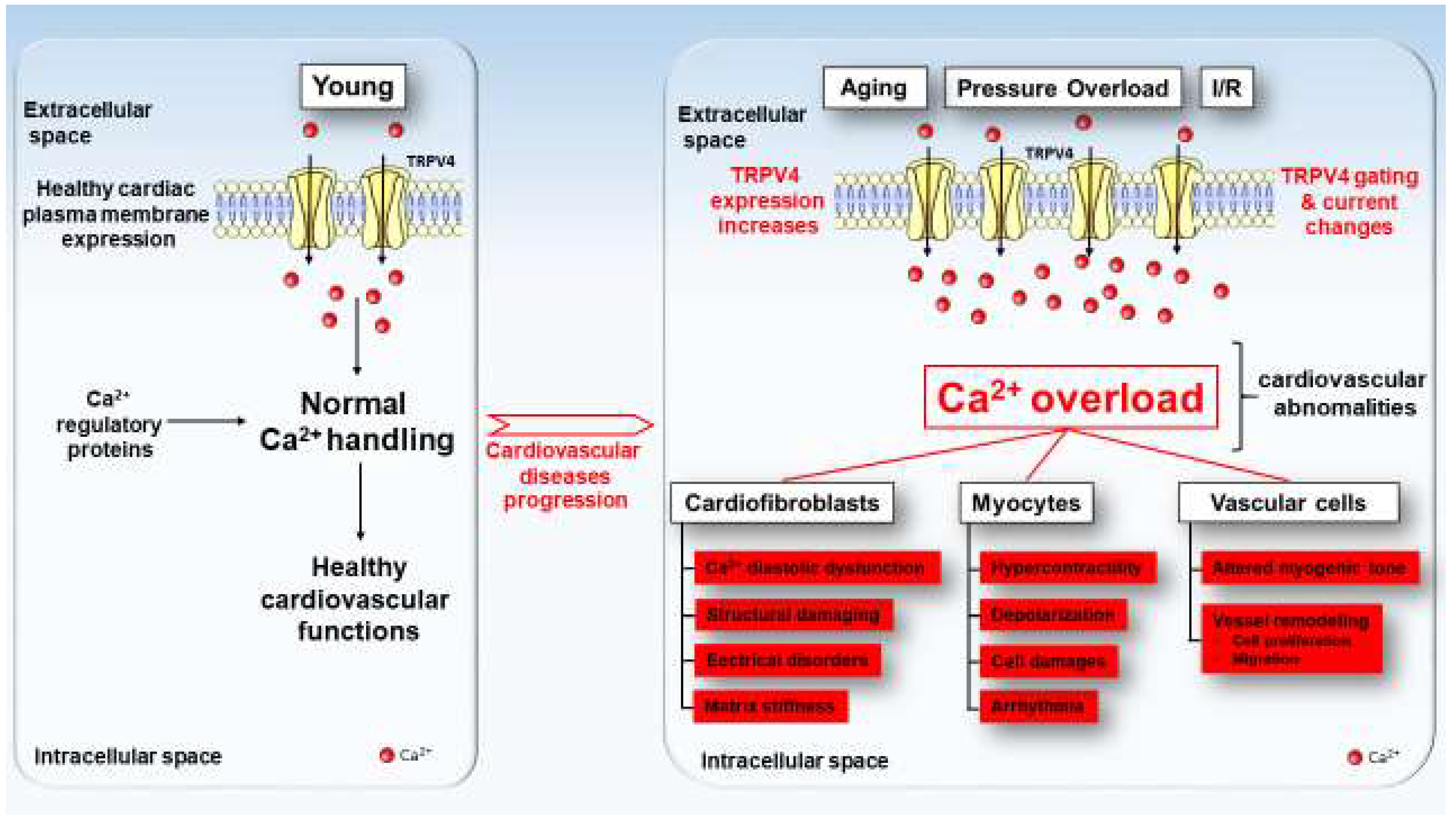

Table 1.
Overview of the evolution of TRPV4 agonist, the associated physiological processes and their clinical application. 4α-PDD: 4alpha-Phorbol 12,13-didecanoate; CF: Cardiofibroblasts; PKC: Protein kinase C; 5,6-EET: 5,6 epoxyeicosatrienoicacid; mPTP: Mitochondrial Permeability Transition Pore; iv: intravenous injection; TNF-α: Tumor Necrosis Factor;.
Table 1.
Overview of the evolution of TRPV4 agonist, the associated physiological processes and their clinical application. 4α-PDD: 4alpha-Phorbol 12,13-didecanoate; CF: Cardiofibroblasts; PKC: Protein kinase C; 5,6-EET: 5,6 epoxyeicosatrienoicacid; mPTP: Mitochondrial Permeability Transition Pore; iv: intravenous injection; TNF-α: Tumor Necrosis Factor;.
| Molecules | Year of identification | EC50/IC50 | Other targets | Features | Cardiovascular effects | Clinical trials / uses | References |
|---|---|---|---|---|---|---|---|
| Agonists | |||||||
| 4α-PDD | 2003 | 50 µM | ● Dorsal root ganglia neurons independently of TRPV4 | ● Negative control for phorbol esters (PKC inhibitors) | ● Ca2+ influx in CF & myocytes ● Ca2+ entry in pulmonary artery smooth muscle cells and increased isometric tension in artery rings |
● None | [68], [69], [70] & [71] |
| 5,6-EET | 2003 | 0.13 µM | ● mPTP | ● Metabolite of arachidonic acid by cytochrome P450 | ● Reduction in: vascular tone, inflammatory response, pathological cardiac remodeling (fibrosis, hypertrophy) & apoptosis ● Improvement in cardiomyocytes function ● Cardioprotection ● Promotes angiogenesis |
● None because of poor solubility and short half life | [72], [73], [74] & [75] |
| RN-1747 | 2009 | 5.9 - 7.7 µM | ● TRPM8 antagonist (IC50 = 4 µM) | ● Benzenesulfonamide derivative | ● None reported | ● None | [60] |
| GSK1016790A | 2008 | 1-18 nM | ● Unknown | ● Oral administration ● iv |
● Endothelial failure and circulatory collapse ● Reduction of TNF-α induced monocyte adhesion to human endothelial cells and atherosclerosis ● Cationic non-selective current activation in rat atrial fibroblasts ● Ca2+ influx in CF and differentiation into myofibroblasts and cardiomyocytes ● Worsening of ishemia-reperfusion injuries in isolated mice hearts and in H9c2 cell line and neonatal rat myocytes ● Decrease in systemic arterial pressure, small decrease in pulmonary arterial pressure, and small increase in cardiac output |
● None | [8], [52], [56], [76], [77], [78], [79], [80], [81] & [82] |
| Quinazolin-4(3H) | 2019 | 280 nM | ● Unknown | ● Orally bioactive ? | None reported | ● None in cardiovascular diseases |
[58] |
Table 2.
Overview of the evolution of TRPV4 antagonists, the associated physiological processes and their clinical application. mPTP: Mitochondrial Permeability Transition Pore; iv: intravenous injection; RISK: Reperfusion injury salvage kinase; DCM-hiPSC-CMs: Dilated cardiomyopathy induced pluripotent stem cell.
Table 2.
Overview of the evolution of TRPV4 antagonists, the associated physiological processes and their clinical application. mPTP: Mitochondrial Permeability Transition Pore; iv: intravenous injection; RISK: Reperfusion injury salvage kinase; DCM-hiPSC-CMs: Dilated cardiomyopathy induced pluripotent stem cell.
| Molecules | Year of identification | EC50/IC50 | Other targets | Features | Cardiovascular effects | Clinical trials / uses | References |
|---|---|---|---|---|---|---|---|
| Antagonists | |||||||
| RN-1734 | 2009 | 2 to 6 µM | ● Poor pharmacokinetics and toxicity | ● Highly selective | ● Prevention of Ca2+ entry mediated vasorelaxation of mesenteric arteries ● Abolition of stretch-activated Ca2+-entry in human induced pluripotent stem cells-derived cardiomyocytes ● Inhibition of the phenylephrine-induced contraction in pulmonary artery smooth muscle cells but nonspecific off-target effects |
● None | [60], [83], [84], [85], [86] & [87] |
| HC-067047 | 2010 | 17 to 133 nM | ● ROS production, depolarization of mitochondrial membrane potential (Δψm) and mPTP opening during H/R | ● iv administration ● Intraperitonealy injected ● Potent ● Selective? |
● Cardioprotection (significantly reduced infarct size, decreased troponin T levels and improved cardiac function in murine model myocardial I/R) [57] ● Anti-apoptotic effects via the activation of RISK pathway ● Reduced TRPV4-related mechanosensitive Ca2+ signaling in DCM-hiPSC-CMs ● Prevent entry of divalent cation in response to myocyte-stretch & hypoosmotic stress-induced cardiomyocyte death and ischemia/reperfusion-induced cardiac damage ● Reduced significantly diabetes-induced cardiac fibrosis ● Inhibition of the PE-induced contraction in pulmonary artery smooth muscle cells |
● None | [11], [56] , [62], [86], [87], [88], [89] & [90] |
| RN-9893 | 2015 | 320 to 660 nM | ● Exhibits >15-fold selectivity for TRPV4 over TRPV1&V3 and TRPM8 | ● Moderate oral bioavailability ● Potent ● Selective |
● Cardioprotection (blocked collagen production following stretch in human valve interstitial cells) ● Reduced cardiac fibrosis |
● None | [82], [63], [89], [88], [89] & [91] |
| GSK2193874 | 2017 | 2 to 50 nM | ● Unknown | ● Orally active ● Potent ● Selective |
● Abolition of pulmonary edema associated with heart failure and enhanced arterial oxygenation ● Increased tail blood flow |
● None | [76], [87], [92] & [93] |
| GSK3527497 | 2019 | 12 nM | ● Unknown | ● Suitable for oral and iv administration ● Reduced bioavailability ● Poor pharmacokinetics and low solubility |
● Unknown | ● None | [88] & [64] |
| GSK2798745 | 2019 | 2 to 16 nM | ● Without any clinically significant safety concerns | ● Highly potent ● Selective ● Orally active |
● Resolve pulmonary edema in heart failure models and attenuate lung damage induced by chemical agents | ● Cardiac heart failure and respiratory diseases ● Diabetic macular edema and cough: https://clinicaltrials.gov/ |
[49], [53], [61], [76], [87] [88], [89] & [94] |
| GSK3395879 | 2018 | 1 nM | ● IC50 > 10µM for TRPA1, TRPV1, TRPM2, TRPM4, TRPM8, TRPC3, TRPC4, TRPC5, TRPC6 | ● Orally bioactive ● Highly potent | ● Abolition of pulmonary edema associated with heart failure | ● None | [11], [91], [67] & [95] |
Table 3.
Comparison of 129/SvJ trpv4-/- and C57bl/6J trpv4-/-. Ref. : References.
| Ref. | Ref. | |||
|---|---|---|---|---|
| Outcomes | 129/SvJ trpv4-/- | [96] | C57bl/6J trpv4-/- | [97] |
| Generation method | 129/SvJ strain via a cassette insertion mutagenesis of exon 5 | C57bl/6J strain with a Cre-lox-mediated excision of exon 12 | ||
| Cardiovacular phenotypes | Impaired vasorelaxation, endothelial calcium response, systemic tonicity | [98] [99] [100] [101] | Impaired vasorelaxation | [98] [102] [103] |
| Altered flow-induced vasodilatation | [104] | Loss of shear stress-induced vasodilation | [105] | |
| - | Cardiac electrophysiological changes | [8] | ||
| - | Absence of VGIC remodelling (Na+, Ca2+ and K+ VGIC) in the left ventricle | [8] | ||
| Extracardiac phenotypes | Viable and fertile | Viable and fertile | [97] | |
| Normal appearance, growth, size, and temperature and no obvious behavioral (including drinking) abnormalities | [106] | Tendency to a lower body weight | [8] | |
| Reduced response to harmful stimuli caused by pressure | [96] | Reduction in water intake and serum osmolality changes | [97] | |
| Intact heat detection but abnormal sensory phenotype | [96] | Reduced response to noxious mechanical stimuli and impaired response to mechanical stimulation | [97] | |
| Altered hearing | [107] | Intact thermal sensing | [97] | |
| Inability to thermoregulate | [100] | Loss of the permeability response in lungs, alveolar barrier | [108] | |
| Deficits in renal tubular K+ secretion | [109] | Proximal tubule defect | [110] | |
| Increased bone mass, decreased osteoclast differentiation | [111] | Blood metabolite changes | [112] | |
| - | Increased bladder capacity | [88] |
Table 4.
TRPV4 channel expression and demonstrated function in cardiovascular system.
| TRPV4 | Atrium | Ventricles | Fibroblasts | Endothelial Cells | Smooth Muscle Cells |
|---|---|---|---|---|---|
| • mRNA | - | Mouse [61], neo rat [10], [56], rat [56] | Mouse [119] , rat [118], human [119], [82] | Mouse [101], rat [120], [122], human [121], [123] | Rat [122], [125], [126], [127] |
| • Protein | Rat [52] | Mouse [8], [11], [61], neo rat [56], [10], rat [56] | Rat [81], neo rat [90], human [82], [119] | Mouse [101], [130] , rat [120], [122], human [121], [123] | Rat [122], [71], [125], [127], [128], human [128], [129] |
| • Function | Rat [52] | Mouse [8], [11], [52], [61], [117], neo rat [10], rat [52] | Rat [52], [81], [90], human [82], [119] | Mouse [101], [130], rat [120], [122], human [121], [123] | Mouse [83], rat [122], [71], [124], [125], [128], human [128], [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



