
Submitted:
29 April 2023
Posted:
30 April 2023
You are already at the latest version
Abstract
Hybridization is recognized as a notable driver of evolution and adaptation, which closely related species may exploit in the form incomplete reproductive barriers. Three closely related species of Ceratocystis (i.e., C. fimbriata, C. manginecans and C. eucalypticola) previously have been shown to hybridize by mating naturally occurring self-sterile strains with an unusual laboratory-generated sterile isolate type, which could have impacted conclusions regarding the prevalence of hybridization and inheritance of mitochondria. In the current study we investigated whether interspecific crosses between fertile isolates of these three species are possible, and if so how mitochondria are inherited by the progeny. For this purpose, a PCR-RFLP method and a mitochondrial DNA-specific PCR technique were custom-made and applied in a novel approach of typing complete ascospore drops collected from the fruiting bodies in each cross to distinguish between self-fertilizations and potential hybridization. These markers showed hybridization between C. fimbriata and C. eucalypticola and between C. fimbriata and C. manginecans, while no hybridization was detected in the crosses involving C. manginecans and C. eucalypticola. In both sets of hybrid progeny, we detected biparental inheritance of mitochondria. This study was the first to successfully produce hybrids from a cross involving self-fertile isolates of Ceratocystis, and also provided the first direct evidence of biparental mitochondrial inheritance in the Ceratocystidaceae. This work lays the foundation for further research focused on investigating the role of hybridization in the speciation of Ceratocystis species, and if mitochondrial conflict could have influenced the process.
Keywords:
Ceratocystis
; hybridization
; PCR RFLP
1. Introduction
Hybridization is one of various forces driving the evolution of new species [1]. It is defined as the crossing of genetically distinguishable groups that leads to the production of viable offspring [2]. This process can result in genetic admixture, which may promote adaptation if the hybrid progeny possesses combinations of beneficial alleles [3]. Through selection, individuals carrying such novel and beneficial allelic combinations may evolve into separate species, having greater virulence and adaptability than their parent species. This has been seen in the evolution of some plant pathogenic fungi, where hybridization led to the emergence of novel species or genotypes responsible for widespread devastation [1,3-6].
Fungal hybrids are often encountered in studies aiming to use the biological species concept for delineating species of Ceratocystidaceae, especially in Endoconidiophora and Ceratocystis [7,8]. In these fungi, intraspecific crosses and self-fertilizations (or selfings) cause formation of ascomata (sexual fruiting bodies) with large, clear ascospore masses at the top of long ascomatal necks [7]. However, interspecific crosses between different Endoconidiophora species result in the formation of ascomata with distorted asci and ascospores, and reduced ascospore viability [7]. Indeed, single-ascospore progeny of the few successful interspecific crosses of Endoconidiophora showed very poor growth, typical of interspecific hybrids [7].
In Ceratocystis, interspecific hybridization has been used to delineate species in the C. fimbriata complex [9]. A notable example is the study of three monophyletic lineages in the complex’s Latin American clade, which are respectively host-specialised to cacao (Theobroma cacao), sweet potato (Ipomea batatas) and sycamore (Platanus spp.) [9]. Sexual mating crosses were used to assess the existence of reproductive barriers between them. Here, crosses producing unusually small numbers of ascospores were taken as evidence of species divergence between the respective lineages, and that host specialization likely mediated the process [9]. Accordingly, the cacao pathogen was described as C. cacaofunesta while the sycamore pathogen was named C. platani [9].
Species in the genus Ceratocystis include many economically important plant pathogens that affect various notable tree and crop species [8,10,11]. The genus also includes the species C. manginecans, which represents an example of a plant pathogen whose evolution has been linked to hybridization [12]. This fungus causes disease on an unusually wide range of plant hosts, e.g., Acacia mangium [13-15], Mangifera indica [16], Eucalyptus species [17], Punica granatum [18], Dalbergia sissoo and Prosopis cineraria [19]. Most other Ceratocystis species affect single plant hosts. For example, the type species of the genus, C. fimbriata [20], causes black rot only on sweet potato [21-25] and the recently described C. eucalypticola appears to infect only Eucalyptus trees [26].
All Ceratocystis species undergo unidirectional mating-type switching during sexual reproduction [27-30]. A common trait of fungi utilizing this system is the natural presence of two sexual fertility types: self-fertile and self-sterile isolate types [27,29,31-33]. At the gene level, the mating-type (MAT1) locus of self-fertile isolates are characterised by the MAT1-1 gene being split by the presence of the MAT1-2 genes [29]. The region containing the MAT1-2 genes is flanked by direct repeats [29], which support an as yet unknown recombination event that mediates permanent deletion of the MAT1-2 genes from the locus [34-36]. Isolates in which this deletion has occurred are self-sterile and contain only the MAT1-1 gene information [29]. However, the picture is somewhat more complex, as a second self-sterile type has also been described from several species in the Ceratocystidaceae, including Ceratocystis [9,37,38]. This second type of self-sterile isolate has only been generated under laboratory conditions, and the basis of its self-sterility remains unclear [9,29,37]. From what is known so far, they apparently encode an intact MAT1 locus with all the requisite MAT1-1 and MAT1-2 gene information [29]. However, as with the MAT1-2-lacking self-sterile isolates, the MAT1-2-containing self-sterile isolates also remain mating competent, and can act as a parent in sexual crosses [29].
Studies on interspecific hybridization in the Ceratocystidaceae have made use of compatible matings between MAT1-2-lacking self-sterile isolates and MAT1-2-containing self-sterile isolates [7,30,39]. This assured that the development of ascomata was due to true hybridization events and not the result of self-fertilization, as would have been the case when self-fertile isolates were used [29]. Such crosses between the two self-sterile isolate types of C. fimbriata and C. eucalypticola were investigated to determine whether reduced fertility of progeny could contribute to post-zygotic reproductive isolation [39]. Similarly, these isolate types were used in crosses of C. fimbriata and C. manginecans to study the phenotype of hybrid progeny [11,30]. In all these cases, the MAT1-2-lacking self-sterile isolates acted as the maternal parent based on the progeny’s inheritance of mitochondria [30,39]. Because none of these previous studies employed self-fertile isolates of Ceratocystis in their experiments, it is unclear how the various conclusions drawn relate to what might happen under natural conditions. Also, MAT1-2-containing self-sterile isolates are considered laboratory-based artifacts and may not be present in natural populations [7,9,30,39].
Therefore, the aim of this study was to determine whether self-fertile isolates, the most commonly encountered isolate type of most Ceratocystis species [27,29,36], are capable of hybridization. This was achieved by employing self-fertile isolates in crosses among the three closely related species C. fimbriata, C. eucalypticola and C. manginecans [8]. Based on the development of ascomata, the relative abundance of outcrossing and self-fertilization was estimated using PCR-based methods. We also used PCR to study mitochondrial inheritance in the progeny. The results of this study would contribute to the knowledge base on fungal hybrids, and more specifically sets the scene for further studies into the role of hybridization in the speciation process of these Ceratocystis species.
2. Materials and Methods
2.1. Interspecific Hybridization of Ceratocystis Isolates
Self-fertile isolates of Ceratocystis manginecans (CMW46461), C. eucalypticola (CMW9998) and C. fimbriata (CMW14799) were obtained from the culture collection (CMW) of the Forestry and Agricultural Biotechnology Institute (FABI) of the University of Pretoria, South Africa. Isolates were maintained at 25 °C MEA-TS medium, which contained 2% (w/v) Malt Extract Agar supplemented with Thiamine (100 mg/L) and Streptomycin (150 mg/L). Their self-fertile nature was evidenced by the presence of ascomata that bear ascospore drops (Figure 1).
To produce hybrid progeny, the three self-fertile isolates were paired in all possible combinations on petri dishes containing MEA-TS. Crosses were made by using a sterile needle for collecting a single ascospore drop from an ascoma of a particular parental isolate and placing the drop approximately 1 cm away from the drop obtained from another parent. Five plates for each of the three parental pairings were prepared and incubated in containers containing silica crystals at 25 °C for a period of two weeks. A zone of interaction between the isolates was defined as the area on the medium where the mycelium of the two Ceratocystis species came in contact, and it is in this region where hybridization was expected to occur. Within the zone of interaction, the ascospore drops from five randomly selected ascomata per plate were collected for each cross (i.e., twenty-five drops were collected per cross). Single spore drops were also collected from each side of every plate, away from the interaction zone, to serve as non-hybrid controls, as the ascomata produced in this region likely resulted from self-fertilization. All the collected spore drops were plated onto fresh MEA-TS medium and incubated for 21 days at 25 °C until the mycelia covered most of the plate. The entire mating experiment was conducted in duplicate.
2.2. DNA Extraction and PCR-Based Confirmation of Parental Self-Fertility
Genomic DNA was isolated from the original parental isolates and all cultures grown from the collected ascospore masses by using an extraction protocol based on hexadecyltrimethyl ammonium bromide (CTAB) [40]. Briefly, fungal tissue was scraped from the entire surface of each plate with a sterile scalpel blade, and then vortexed in the presence of extraction buffer (0.2 M Tris, 1.4 M NaCl, 20 mM EDTA, 0.2 g/L CTAB) containing 2 or 3 glass beads. Mixtures containing the disrupted fungal tissue were incubated for 3 mins at 100 °C and then 10 mins on ice. Supernatant was collected by centrifugation for 5 mins at 18 000 rcf, after which the nucleic acids contained within the aqueous phase were purified by chloroform: isoamylalcohol (24:1) extraction. Following overnight precipitation at -20 °C in the presence of 2.5 M ammonium acetate and 2 volumes of isopropanol, nucleic acids were recovered by centrifugation at 18 000 rcf for 10 mins at 4 °C. The resulting pellets were washed with 70% ethanol, air-dried, and suspended in sterile water.
To confirm self-fertility of the parental isolates, they were screened for the presence of the MAT1-1-1 and MAT1-2-1 genes. To do this, portions with particular sizes were amplified for the respective genes using primers (Table 1) previously designed for C. fimbriata [27] and also tested on related Ceratocystis species [39]. The target fragments were amplified using 25 µl PCR mixtures that contained 1 U of KAPA Taq DNA polymerase (Kapa Biosystems, USA), 1 X KAPA Taq Buffer A, 0.4 mM of each primer, 0.25 mM of each dNTP and 90-150 ng of template DNA. An Eppendorf Thermocycler (Eppendorf AG, Mannheim, Germany) was used for amplification with the following parameters: 5 min at 95 °C followed by 35 cycles of 30 sec at 95 °C, 30 sec at 50 °C and 30 sec 72 °C; and a final elongation step of 72 °C for 7 min. Amplicon sizes were estimated using 1% agarose (LE Agarose, SeaKem, USA) by gel electrophoresis for 20 mins at 150 V, after which the DNA stained with GelRed Nucleic Acid Gel stain (Biotium, Hayward, USA) was visualised with an ultraviolet transilluminator.
2.3. Marker and Primer Design
Two sets of markers were developed in this study, one was designed using nuclear DNA regions to allow differentiation of C. fimbriata, C. eucalypticola and C. manginecans from one another, while the second targeted their mitochondrial genomes. For this purpose, the genome sequences for the respective species were obtained from the database (https://www.ncbi.nlm.nih.gov/genome/) of the National Centre for Biotechnology Information (NCBI) using the accession numbers SGIO00000000, APWK03000000 and LJOA00000000 for, respectively, C. manginecans CMW46461, C. fimbriata CMW14799 and C. eucalypticola CMW9998.
The C. fimbriata genome had been previously annotated [41] and was used to design three sets of nuclear markers to use as a diagnostic for identifying the individual species. An in-silico analysis of restriction enzyme (RE) digestion patterns was used to find intergenic regions with sequence variation that could be used in a PCR-based restriction fragment length polymorphism (PCR-RFLP) analysis. This was done by randomly taking portions of the C. fimbriata genome and doing a BLAST comparison to the C. manginecans and C. eucalypticola genomes to find regions of sequence variation. Three variable regions were selected, aligned among the three species, and then subjected to in silico digestion for identifying unique RE cut sites. Regions that were considered suitable as RFLP markers were then used as input into Primer3Plus [42,43] to design primers for PCR amplification of the respective regions (Table 1).
Mitochondrial genome sequences for the three species were compared as before to identify variable regions that could potentially be used to identify the parental origin of the mitochondria inherited by the progeny. The sequences of these variable regions together with their flanking sequences were then used as input into Primer3Plus to design primers that would allow amplicons of different sizes for the three species (Table 1). In other words, this diagnostic method included three primer sets that could, when used in combination, identify the origin of the mitochondria.
2.4. Marker Analysis
The primers designed for the RFLP analysis were used to amplify the targeted marker regions using DNA extracted from the parental isolates, as well as from the progeny. Each 25 µl reaction mixture included 1 U of KAPA Taq DNA polymerase (Kapa Biosystems, USA), 1 X KAPA Taq Buffer A, 0.4 mM of each primer, 0.25 mM of each of the dNTPs and 90-150 ng of template DNA. The PCR cycling conditions were as follows: an initial denaturation step of 3 min at 94 °C; followed by 25 cycles of 30 sec at 94 °C, 30 sec at 50 °C and 30 sec at 72 °C; and a final extension step of 7 mins at 72 °C. The amplified PCR products were stained with GelRed and visualised on 1% agarose by gel electrophoresis after 20 mins at 150 V.
RFLP analysis of the amplicons was done using the appropriate high-fidelity enzymes (New England Biolabs, Ipswich, USA) as per the manufacturer’s instructions. One unit of each RE and 1X rCutSmart Buffer was added to the PCR reaction, which was then incubated for 1 hr at 37 °C followed by an inactivation step at 80 °C for 20 mins. GelRed was added to the inactivated digestion, and the full reaction was separated on a 1% agarose gel for 60 mins at 90 V and visualised under UV light. The restriction profile of each spore drop culture was compared to the expected banding pattern for the individual parents to identify possible hybridizations. Progeny was scored as the result of hybridization if both banding patterns of the parents were present in a culture established from a single spore drop.
The primer sets that targeted mitochondrial genomes were amplified as described above, except that an annealing temperature of 52 °C was used. Following PCR, amplicons were visualised as described above, after which their sizes were used to identify the mitochondrial origin of potential hybrids from the C. manginecans X C. eucalypticola and C. manginecans X C. fimbriata crosses. However, primer set 1 produced almost the same size fragment in C. fimbriata and C. eucalypticola, which necessitated the use of sequence analysis for scoring mitochondrial origins of their progeny. To do this, the relevant amplicons were purified using Sephadex G50 columns (Sigma-Aldrich, USA), and the amplicons were sequenced using feMitSeq_F primer, the BigDye Terminator Cycle Sequencing Kit v.3.1 (Life Technologies, USA) and an ABI3500xL Genetic analyser (Applied Biosystems, ThermoFisher Scientific, Waltham, MA, USA) at the DNA Sanger Sequencing facility, Faculty of Natural and Agricultural Science at the University of Pretoria. The resulting electropherograms were visually inspected for diagnostic regions unique to the mitochondrial genome of C. eucalypticola.
3. Results
3.1. Interspecific Hybridization of Ceratocystis Isolates
On MEA-TS medium, the three parental isolates (i.e., C. manginecans, C. eucalypticola and C. fimbriata) all produced abundant ascomata, which served as an initial indication of their self-fertility. PCR amplification of the MAT1-1-1 and MAT1-2-1 genes from these cultures all produced amplicons of the expected sizes (i.e., 436 bp and 495 bp, respectively). Therefore, presence of both the MAT1-1-1 and MAT1-2-1 fragments confirmed that the isolates were indeed self-fertile and had not shifted to self-sterility through unidirectional mating-type switching [27,34,35,44].
When mated with each other, all interspecific crosses produced some ascomata within the zone of interaction (Figure 2). However, the number of fruiting bodies present in this zone was consistently less than the number present in regions outside this region where selfing reactions likely occurred. Culturing of the spore drops collected in the zones of interaction from the various MEA-TS plates again resulted in cultures that abundantly produced ascomata.
3.2. Development of a PCR-RFLP Method Based on Nuclear Markers
Three sets of nuclear markers with suitable levels of polymorphism were to design a PCR-RFLP method for differentiating C. manginecans, C. eucalypticola and C. fimbriata from one another. These were based on the sequence comparisons of three genes. In the case of nuclear marker 1, our primers targeted part of the noncoding region to the one side of the endoplasmic reticulum membrane protein 65. For marker 2, the primers targeted a gene encoding endothelin-converting enzyme-like 1. The region targeted by the primers for marker 3 corresponded to the beginning of a gene encoding bifunctional purine biosynthetic protein ADE1, as well as part of the non-coding region upstream from it. Laboratory testing of these markers yielded the predicted PCR-RFLP profiles for each of the parental species. Details regarding each of these markers are provided below.
The primers for nuclear marker 1 were designed to amplify a 692 bp fragment in all three Ceratocystis species. This fragment had a single HindIII cut site in all three species, as well as two PstI cut sites in C. eucalypticola and C. fimbriata (Figure 3a). In silico digestions of C. eucalypticola and C. fimbriata with HindIII and PstI thus produced four bands of sizes (of 35, 170, 190 and 290 bp). The lack of the second PstI cut site in C. manginecans provided this fungus with a unique band pattern (35, 290 and 360 bp fragments) (Figure 3d). Therefore, this marker could differentiate C. manginecans from C. eucalypticola and C. fimbriata.
Marker region 2 was designed around an 836 bp amplicon that would be amplified in all three Ceratocystis species (Figure 3b). In silico digestion with HindIII predicted the formation of two bands (100 bp and 740 bp) for C. manginecans and C. fimbriata, with neither containing PstI cut sites (Figure 3d). In C. eucalypticola, this region had four HindIII and one PstI cut sites (producing six fragments of sizes 55 bp, 85 bp, 90 bp, 100 bp, 110 bp and 400 bp respectively. Therefore, this marker region could differentiate C. eucalypticola from C. manginecans and C. fimbriata.
The primers for nuclear marker 3 were designed to produce an amplicon of 908 bp. This marker would differentiate C. fimbriata from the other two species, as a unique PstI cut site and common HindIII cut site produces three bands sized 560, 180 and 160 bp respectively when digested in silico with these restriction enzymes (Figure 3c,d). In C. manginecans and C. eucalypticola, digestion with HindIII and PstI produces two bands of sizes 340 bp and 560 bp.
3.3. Identifying Hybrid Cultures Using PCR-RFLP Analysis of Nuclear Markers
PCR-RFLP analysis using the three polymorphic nuclear markers identified in this study allowed differentiation of C. manginecans, C. eucalypticola and C. fimbriata isolates from one another, and from cultures potentially representing hybrids of these species. If restriction profiles obtained from progeny cultures were identical to any of the three parental isolates, they were regarded as being the product of a selfing (i.e., self-fertilization of the parent isolate). If the restriction profile represented a mixture of the profiles for both parents, the progeny culture was scored as being the product of a hybridization between the parents (Figure 4).
3.3.1. C. Fimbriata X C. Eucalypticola
For the cultures grown from 25 ascospore drops collected during the first round of interspecific crosses, marker 2 was successfully amplified in only 23 of the resulting cultures, despite multiple PCR attempts. Among these, 4 cultures showed the banding pattern of C. fimbriata only (indicative of C. fimbriata selfing), 3 showed the banding pattern of only C. eucalypticola (indicative of C. eucalypticola selfing) and 16 showed a banding pattern characteristic of both C. fimbriata and C. eucalypticola, indicating hybridization. Marker 3 was amplified from all 25 spore drop cultures, with the RFLP scoring 6 isolates as C. fimbriata selfings, 7 as C. eucalypticola selfings and 12 as potential hybrids (Table S1). During the second round of crosses, marker 2 was successfully amplified from all 25 spore drop cultures, as was marker 3. However, neither marker region 2 nor 3 showed a selfing of C. fimbriata. Marker 2 showed 15 C. eucalypticola selfings and 10 hybridizations, while marker 3 showed 22 C. eucalypticola selfings and only 3 hybridizations (Table S2). The spore drops collected as controls showed the expected banding pattern of the individual parent. Taken together (Table 2), these data showed that among the 50 mating events examined during the two rounds of crosses, 26 were scored involving hybridization and 24 as selfings (20 in C. eucalypticola and 4 in C. fimbriata).
3.3.2. C. Fimbriata X C. Manginecans
For cultures grown from the 25 ascospore drops collected from the first round of crosses, both marker 1 and marker 3 were successfully amplified from 24 of the resulting cultures. Using nuclear marker 1, none of the cultures were scored as originating from hybridization, while 23 and 1 were scored as being the product selfing in, respectively, C. fimbriata and C. manginecans. With marker 3, we scored 3 of the cultures as having hybrid origins, while 1 and 20 cultures were scored as selfings of C. manginecans and C. fimbriata, respectively (Table S1). For the second round of crosses, evidence of hybridization was detected only with marker region 1. Of the selfings scored with marker 1, the majority (20) originated from C. fimbriata with only a single selfing in C. manginecans detected. With marker region 3, we only detected selfings of C. fimbriata, except for one detected as a C. manginecans selfings event (Table S2). Therefore, of the 48 mating events between C. fimbriata X C. manginecans, only 8 bore evidence of hybridization, while 40 were regarded as selfings (1 in C. manginecans and 39 in C. fimbriata) (Table 2).
3.3.3. C. Manginecans X C. Eucalypticola
For the first round of C. manginecans X C. eucalypticola crosses, 25 spore drops were collected and grown on MEA-TS medium. Marker regions 1 and 2 were successfully amplified from 24 of the resulting cultures. Based on their PCR-RFLP profiles, 23 of these were scored as being the product of selfing of C. eucalypticola and one as a selfing of C. manginecans (Table S1). In the second round of crosses, marker region 1 was successfully amplified from all 25 cultures, of which PCR-RFLP analysis suggested that 19 originate from C. eucalypticola selfings and six from C. manginecans selfings. With marker region 2, which was successfully amplified only for 22 of the spore drop cultures, 16 were scored as C. eucalypticola selfings and 6 as C. manginecans selfings (Table S2). None of the ascospore drop cultures thus appeared to have originated from hybridization (Table 2).
3.4. Mitochondrial DNA Inheritance in Hybrid Cultures
Three primer sets were designed to amplify regions of the mitochondrial genome that are diagnostic for the three Ceratocystis species examined (Figure 5). The mitochondrial genome was not annotated in the genome assembly, and therefore it was not known if these regions were within a gene. However, these markers were used to study the inheritance of mitochondria in the cultures identified as having originated from hybridization events between the crosses examined here (Table 2). All eight of the hybrid cultures obtained from the crosses between C. fimbriata and C. manginecans yielded two distinct amplicons using primer set 1. Of these, one corresponded to the 560 bp fragment diagnostic for C. manginecans and the other to the 1890 bp fragment of C. fimbriata. Similarly, for primer set 2 an amplicon was produced from all hybrid cultures, although the differences in size were too small to assign parental origin. This result was unexpected, as the in silico analysis indicated that no amplicon should be possible in the C. manginecans parent and the marker was not used to assign origin to the hybrids. With primer set 3, all of these hybrid cultures contained the 560 bp diagnostic for C. fimbriata. In other words, the mitochondria of both these parental strains were inherited by their hybrid progeny (Figure S1).
For all 26 of the ascospore drop cultures collected from the crosses between C. fimbriata and C. eucalypticola, PCR with primer set 3 yielded the 560 bp fragment expected in C. fimbriata. PCR with primer set 1 yielded an 1890 bp fragment expected for both parents. Therefore, to determine their origins, the amplicons were sequenced, which in most cultures showed the presence of C. fimbriata mitochondria (Figure S2). However, the sequence data also suggested the presence of C. eucalypticola mitochondrial DNA in at least 6 of the 26 hybrid cultures. This was apparent from the presence of two sequencing profiles in the region containing the 9 bp deletion characteristic of C. eucalypticola (Figure S2). Therefore, PCR combined with sequence information provided evidence of both parents’ mitochondrial DNA being present in some of the ascospore drops originally collected from the ascomata produced during the C. fimbriata X C. eucalypticola cross.
4. Discussion
Here we used a novel approach to demonstrate that self-fertile isolates of different Ceratocystis species can hybridize and that their progeny often bear the mitochondria of both parental species. Although the importance of hybridization during adaptation and evolution is widely recognized [45,46], fungal hybridization is not completely understood and few studies have explored the level to which sexual reproductive barriers and species boundaries are permeable [47,48]. By employing cultures grown from single ascospore drops obtained from crosses between C. fimbriata and C. manginecans and between C. fimbriata and C. eucalypticola we showed for the first time that self-fertile isolates of closely related Ceratocystis species can sexually reproduce, forming viable progeny with mitochondrial inheritance from both parents. All previous attempts at this “forced” outcrossing by mating self-sterile isolates lacking MAT1-2 with self-sterile isolates containing MAT1-2 [30,39]. Contrast to MAT1-2-lacking self-sterile isolates that are present among the progeny of self-fertilizations [7,29,36,39], MAT1-2-containing self-sterile strains are not encountered in nature [7,9,30,39]. The use of these two types of self-sterile isolates might thus skew the data obtained from sexual mating experiments. Therefore, our experiments with self-fertile isolates provided more conclusive support to the leaky reproductive barriers previously reported between some of these species [39,49,50]. Additionally, these findings provide a strong basis from which to explore the roles of decreased fertility and fitness as post-zygotic reproductive barriers in these fungi [39].
The fact that crosses between C. manginecans and C. eucalypticola did not produce hybrid progeny was unexpected. Both of these species were interfertile with C. fimbriata and based on rigorous phylogenetic analyses the three species are very closely related [8]. However, neither C. manginecans nor C. eucalypticola have been widely studied, and we likely have an incomplete picture of their ecology and distribution. C. eucalypticola has only been reported from Eucalyptus species in South Africa [26,51], while C. manginecans has been reported on various hosts and from various localities in the Middle East, Malaysia, India, Brazil, and South China [13,14,17,19,20,28]. Therefore, meaningful explorations of the basis of the apparent reproductive barrier between C. manginecans and C. eucalypticola require analyses of broad collections of isolates that are representative of both their geographic distribution and hosts range [52,53].
The presence of PCR markers from both parents in the mitochondria of the examined hybrid progeny is interesting, as these organelles typically show a uniparental inheritance pattern in fungi [54-56] that is distinct to that of nuclear genes [57,58]. In plants and animals, uniparental inheritance of mitochondria is mostly facilitated by pre-zygotic processes related to gamete anisogamy where the maternal gamete is larger and contains many more mitochondria compared to the paternal gamete [54-56,59]. Not much is known about the regulation of mitochondrial inheritance in fungi, but various possible mechanisms might be involved, ranging from the physical position of mitochondria within dividing cells through to enzymatic degradation of additional types of mitochondria or mitochondrial DNAs [60]. Nevertheless, the paradigm of uniparental mitochondrial inheritance was previously reported for interspecific [30,39] and intraspecific [30,39] crosses of Ceratocystis. For interactions between conspecifics, one Ceratocystis parent’s mitochondria or mitochondrial DNAs are purged from the progeny [60-62]. In the case of hybrid crosses, however, these results of these initial studies were likely impacted by the use of MAT1-2-containing self-sterile strains, especially when evaluating reproductive barriers. In other words, the reduced fitness associated with these types of sterile isolates likely extends to their mitochondria, as our data clearly showed that mitochondria of both parental species can end up in the hybrid progeny.
Biparental inheritance of mitochondria is often associated with hybridization, where the condition of harbouring more than one type of mitochondrion is referred to as heteroplasmy [58,63]. This condition is due to a breakdown in the coordinated set of mechanisms governing the typical uniparental inheritance of these organelles [58]. Biparental inheritance has been reported before in crosses between the plant pathogens Verticillium dahlia and V. nonalfalfae [52,64], as well as among various species yeast and human pathogens (e.g., [65,66]. In the current study, the crosses between C. fimbriata and C. manginecans and the crosses between C. fimbriata and C. eucalypticola all produced heteroplasmic progeny, and their analysis using genetics and comparative genomics methodologies would be invaluable, not only for understanding species boundaries, but also for developing a holistic knowledge framework for why and how these fungi emerged as pathogens.
Another intriguing explanation for the presence of mitochondria from both parents in the hybrid progeny could stem from the sexual strategy employed by Ceratocystis. During unidirectional mating-type switching, apparently self-fertile isolates produce ascospores that are either self-fertile or self-sterile [27,34,35,44]. In Chromocrea spinulosa, another species capable of unidirectional mating-type switching, there is evidence that nuclei of opposite mating-type co-exist within the a single cytoplasm, and karyogamy followed by meiosis would produce a mixture of self-fertile and self-sterile ascospores [34]. Whether this mechanism is at play in Ceratocystis is currently not known. However, it could be speculated that in the Ceratocystis hybrid crosses presented here, each parent both contributed two nuclei to the fertilization event that produced an ascoma – one self-fertile nucleus and one self-sterile nucleus, in a single cell with its own mitochondria. The parent that acted as the maternal parent (i.e., the cell that donated the self-sterile nucleus [30,39]) would then dictate which cell donates cytoplasm and mitochondria during karyogamy. This would result in the mitochondria of a single ascospore originating from one parent only. However, among the many ascospores in a spore drop, there could be variations in which cell acted as maternal parent and therefore acted as mitochondrial donor. Genetic typing of complete spore drops (as done here) would then produce a profile associated with biparental mitochondrial inheritance. Our future work will seek to explore this hypothesis further by examining the mitochondria present in single isolates derived from these self-fertile Ceratocystis crosses.
Our novel use of cultures established from single ascospore drops obtained from distinct ascomata for detecting hybridization and to track mitochondrial inheritance is reminiscent of a technique previously used to score self-fertile and MAT1-2-lacking self-sterile isolates in progeny of Ceratocystis drops [67]. Because PCR may be associated amplification bias [68], it is conceivable that an ascospore drop produced from a hybrid interaction is characterized by only one parent profile. Similarly, any segregation bias in the marker regions or differences in the viability of ascospores contained within a spore drop could have caused an underestimation of the possible number of hybridization events examined. Furthermore, the use of only two marker regions may further impact this underestimation. Additionally, we only used one individual from each of the three species, but fertility within populations of Ceratocystis have been shown to vary widely [30]. However, neither PCR bias, nor the choice of isolates would have impacted our conclusions regarding hybridization and mitochondrial inheritance in the three fungi examined. Our methodology therefore introduces a feasible strategy with which to identify hybrids and trace mitochondrial inheritance across Ceratocystis and the broader Ceratocystidaceae.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Amplicons produced by the mitochondrial primer sets 1;Figure S2: Electropherograms from sequencing the amplicons using the mitochondrial sequencing primer; Table S1: RFLP results showing the parental origin of individual spore drops for three self-fertile interspecific crosses generated during the first round of crosses; Table S2: RFLP results showing the parental origin of individual spore drops for three self-fertile interspecific crosses generated during the second round of crosses.
Author Contributions
Conceptualization, D.v.d.W., E.T.S., B.D.W. and P.M.W.; methodology, D.v.d.W., E.T.S., B.D.W. and P.M.W.; formal analysis, D.v.d.W.; investigation, D.v.d.W.; data curation, D.v.d.W.; writing—original draft preparation, D.v.d.W.; writing—review and editing, E.T.S., B.D.W. and P.M.W.; visualization, D.v.d.W. and P.M.W.; supervision, E.T.S., B.D.W. and P.M.W.; project administration, D.v.d.W. and P.M.W.; funding acquisition, B.D.W. All authors have read and agreed to the published version of the manuscript.
Funding
This project was supported by the University of Pretoria, the Department of Science and Technology (DST)/National Research Foundation (NRF) Centre of Excellence in Tree Health Biotechnology (CTHB). The project was additionally supported by B. D. Wingfield’s DST/NRF SARChI chair in Fungal Genomics (grant number 98353). The grant holders acknowledge that opinions, findings and conclusions, or recommendations expressed in this piece of work are those of the researchers and that the funding bodies accept no liability whatsoever in this regard.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analysed in this study. These data can be found at https://www.ncbi.nlm.nih.gov/using the information provided in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Callaghan, S.; Guest, D. Globalisation, the founder effect, hybrid Phytophthora species and rapid evolution: New headaches for biosecurity. Australas. Plant Pathol. 2015, 44, 255–262. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hessenauer, P.; Fijarczyk, A.; Martin, H.; Prunier, J.; Charron, G.; Chapuis, J.; Bernier, L.; Tanguay, P.; Hamelin, R.C.; Landry, C.R. Hybridization and introgression drive genome evolution of Dutch elm disease pathogens. Nat. Ecol. Evol. 2020, 4, 626–638. [Google Scholar] [CrossRef]
- Samarasinghe, H.; You, M.; Jenkinson, T.S.; Xu, J.; James, T.Y. Hybridization facilitates adaptive evolution in two major fungal pathogens. Genes 2020, 11, 101. [Google Scholar] [CrossRef] [PubMed]
- Feurtey, A.; Stukenbrock, E.H. Interspecific gene exchange as a driver of adaptive evolution in fungi. Annu. Rev. Microbiol. 2018, 72, 377–398. [Google Scholar] [CrossRef]
- Stukenbrock, E.H. The role of hybridization in the evolution and emergence of new fungal plant pathogens. Phytopathology 2016, 106, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C.; McNew, D.L. Partial interfertility among the Ceratocystis species on conifers. Fungal Genet. Biol. 1998, 25, 44–53. [Google Scholar] [CrossRef]
- De Beer, Z.W.; Duong, T.A.; Barnes, I.; Wingfield, B.D.; Wingfield, M.J. Redefining Ceratocystis and allied genera. Stud. Mycol. 2014, 79, 187–219. [Google Scholar] [CrossRef]
- Engelbrecht, C.J.B.; Harrington, T.C. Intersterility, morphology and taxonomy of Ceratocystis fimbriata on sweet potato, cacao and sycamore. Mycologia 2005, 97, 57–69. [Google Scholar] [CrossRef]
- van der Nest, M.A.; Steenkamp, E.T.; Roodt, D.; Soal, N.C.; Palmer, M.; Chan, W.-Y.; Wilken, P.M.; Duong, T.A.; Naidoo, K.; Santana, Q.C.; et al. Genomic analysis of the aggressive tree pathogen Ceratocystis albifundus. Fungal Biol. 2019, 123, 351–363. [Google Scholar] [CrossRef]
- Fourie, A.; van der Nest, M.A.; de Vos, L.; Wingfield, M.J.; Wingfield, B.D.; Barnes, I. QTL mapping of mycelial growth and aggressiveness to distinct hosts in Ceratocystis pathogens. Fungal Genet. Biol. 2019, 131, 103242. [Google Scholar] [CrossRef]
- Naidoo, K.; Steenkamp, E.T.; Coetzee, M.P.; Wingfield, M.J.; Wingfield, B.D. Concerted evolution in the ribosomal RNA cistron. PLoS ONE 2013, 8, e59355. [Google Scholar] [CrossRef] [PubMed]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; Thu, P.Q.; Barnes, I. A possible centre of diversity in South East Asia for the tree pathogen, Ceratocystis manginecans. Infect. Genet. Evol. 2016, 41, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Brawner, J.; Japarudin, Y.; Lapammu, M.; Rauf, R.; Boden, D.; Wingfield, M., J. . Evaluating the inheritance of Ceratocystis acaciivora symptom expression in a diverse Acacia mangium breeding population. South. For. A J. For. Sci. 2015, 77, 83–90. [Google Scholar] [CrossRef]
- Tarigan, M.; Roux, J.; Van Wyk, M.; Tjahjono, B.; Wingfield, M.J. A new wilt and die-back disease of Acacia mangium associated with Ceratocystis manginecans and C. acaciivora sp. nov. in Indonesia. South Afr. J. Bot. 2011, 77, 292–304. [Google Scholar] [CrossRef]
- Al Adawi, A.; Deadman, M.; Al Rawahi, A.; Al Maqbali, Y.; Al Jahwari, A.; Al Saadi, B.; Al Amri, I.; Wingfield, M. Aetiology and causal agents of mango sudden decline disease in the Sultanate of Oman. Eur. J. Plant Pathol. 2006, 116, 247–254. [Google Scholar] [CrossRef]
- Chen, S.; Van Wyk, M.; Roux, J.; Wingfield, M.J.; Xie, Y.; Zhou, X. Taxonomy and pathogenicity of Ceratocystis species on Eucalyptus trees in South China, including C. chinaeucensis sp. nov. Fungal Divers. 2013, 58, 267–279. [Google Scholar] [CrossRef]
- Harrington, T.C.; Huang, Q.; Ferreira, M.A.; Alfenas, A.C. Genetic analyses trace the Yunnan, China population of Ceratocystis fimbriata on pomegranate and taro to populations on Eucalyptus in Brazil. Plant Dis. 2015, 99, 106–111. [Google Scholar] [CrossRef]
- Al Adawi, A.O.; Barnes, I.; Khan, I.A.; Al Subhi, A.M.; Al Jahwari, A.A.; Deadman, M.L.; Wingfield, B.D.; Wingfield, M.J. Ceratocystis manginecans associated with a serious wilt disease of two native legume trees in Oman and Pakistan. Australas. Plant Pathol. 2013, 42, 179–193. [Google Scholar] [CrossRef]
- Van Wyk, M.; Al Adawi, A.O.; Khan, I.A.; Deadman, M.L.; Al Jahwari, A.A.; Wingfield, B.D.; Ploetz, R.; Wingfield, M.J. Ceratocystis manginecans sp. nov., causal agent of a destructive mango wilt disease in Oman and Pakistan. Fungal Divers. 2007, 27, 213–230. [Google Scholar]
- Halsted, B.D.; Fairchild, D.G. Sweet-potato black rot. J. Mycol. 1891, 1–11. [Google Scholar] [CrossRef]
- Halsted, B.D. Some fungus diseases of the sweet potato; New Jersey Agricultural College Experiment Station: 1890; pp. 25–27.
- Li, Q.; Harrington, T.C.; McNew, D.; Li, J.; Huang, Q.; Somasekhara, Y.M.; Alfenas, A.C. Genetic bottlenecks for two populations of Ceratocystis fimbriata on sweet potato and pomegranate in China. Plant Dis. 2016, 100, 2266–2274. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.C.; Nam, S.-S.; Kachroo, A.; Kim, Y.-H.; Yang, J.-W. Characterization and pathogenicity of sweet potato (Ipomoea batatas) black rot caused by Ceratocystis fimbriata in Korea. Eur. J. Plant Pathol. 2018, 152, 833–840. [Google Scholar] [CrossRef]
- Harter, L.L.; Weimer, J. A monographic study of sweet-potato diseases and their control; Washington, D.C., 29; pp. 1–118. 19 April.
- van Wyk, M.; Roux, J.; Nkuekam, G.K.; Wingfield, B.D.; Wingfield, M.J. Ceratocystis eucalypticola sp. nov. from Eucalyptus in South Africa and comparison to global isolates from this tree. IMA Fungus 2012, 3, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; De Beer, Z.W.; Wingfield, B.D. DNA loss at the Ceratocystis fimbriata mating locus results in self-sterility. PLoS ONE 2014, 9, e92180. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C.; Kazmi, M.R.; Al-Sadi, A.M.; Ismail, S.I. Intraspecific and intragenomic variability of ITS rDNA sequences reveals taxonomic problems in Ceratocystis fimbriata sensu stricto. Mycologia 2014, 106, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.S.; Harrington, T.C.; Ferreira, M.A.; Damacena, M.B.; Al-Sadi, A.M.; Al-Mahmooli, I.H.; Alfenas, A.C. Species or genotypes? Reassessment of four recently described species of the Ceratocystis wilt pathogen, Ceratocystis fimbriata, on Mangifera indica. Phytopathology 2015, 105, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; van der Nest, M.A.; Loots, M.T.; Barnes, I. Inheritance of phenotypic traits in the progeny of a Ceratocystis interspecific cross. Fungal Biol. 2018, 122, 717–729. [Google Scholar] [CrossRef]
- Harrington, T.C.; McNew, D.L. Self-fertility and uni-directional mating-type switching in Ceratocystis coerulescens, a filamentous ascomycete. Curr. Genet. 1997, 32, 52–59. [Google Scholar] [CrossRef]
- Harrington, T.C.; Steimel, J.; Kile, G. Genetic variation in three Ceratocystis species with outcrossing, selfing and asexual reproductive strategies. Eur. J. For. Pathol. 1998, 28, 217–226. [Google Scholar] [CrossRef]
- Lee, D.H.; Roux, J.; Wingfield, B.D.; Wingfield, M.J. Variation in growth rates and aggressiveness of naturally occurring self-fertile and self-sterile isolates of the wilt pathogen Ceratocystis albifundus. Plant Pathol. 2015, 64, 1103–1109. [Google Scholar] [CrossRef]
- Yun, S.H.; Kim, H.K.; Lee, T.; Turgeon, B.G. Self-fertility in Chromocrea spinulosa is a consequence of direct repeat-mediated loss of MAT1-2, subsequent imbalance of nuclei differing in mating type, and recognition between unlike nuclei in a common cytoplasm. PLoS Genet. 2017, 13, e1006981. [Google Scholar] [CrossRef]
- Xu, L.; Jardini, T.M.; Chen, W. Direct repeat-mediated DNA deletion of the mating type MAT1-2 genes results in unidirectional mating type switching in Sclerotinia trifoliorum. Sci. Rep. 2016, 6, 27083. [Google Scholar] [CrossRef]
- Witthuhn, R.C.; Harrington, T.C.; Wingfield, B.D.; Steimel, J.P.; Wingfield, M.J. Deletion of the MAT- 2 mating-type gene during uni-directional mating-type switching in Ceratocystis. Curr. Genet. 2000, 38, 48–52. [Google Scholar] [CrossRef]
- Johnson, J.; Harrington, T.; Engelbrecht, C.J.B. Phylogeny and taxonomy of the North American clade of the Ceratocystis fimbriata complex. Mycologia 2005, 97, 1067–1092. [Google Scholar] [CrossRef]
- Harrington, T.C.; Pashenova, N.V.; McNew, D.L.; Steimel, J.; Konstantinov, M.Y. Species delimitation and host specialization of Ceratocystis laricicola and C. polonica to larch and spruce. Plant Dis. 2002, 86, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Wilken, P.M. Mating in Ceratocystis. University of Pretoria, 2015.
- Damm, U.; Mostert, L.; Crous, P.W.; Fourie, P.H. Novel Phaeoacremonium species associated with necrotic wood of Prunus trees. Persoonia 2008, 20, 87–102. [Google Scholar] [CrossRef]
- Wingfield, B.; Fourie, A.; Simpson, M.; Bushula-Njah, V.; Aylward, J.; Barnes, I.; Coetzee, M.; Dreyer, L.; Duong, T.; Geiser, D.; et al. IMA Genome-F 11: Draft genome sequences of Fusarium xylarioides, Teratosphaeria gauchensis and T. zuluensis and genome annotation for Ceratocystis fimbriata. IMA Fungus 2019, 10. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 1999, 132, 365–386. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, W71–W74. [Google Scholar] [CrossRef]
- Kramer, D.; Lane, F.A.; Steenkamp, E.T.; Wingfield, B.D.; Wilken, P.M. Unidirectional mating-type switching confers self-fertility to Thielaviopsis cerberus, the only homothallic species in the genus. Fungal Biol. 2021, 125, 427–434. [Google Scholar] [CrossRef]
- Morard, M.; Ibanez, C.; Adam, A.C.; Querol, A.; Barrio, E.; Toft, C. Genomic instability in an interspecific hybrid of the genus Saccharomyces: A matter of adaptability. Microb. Genom. 2020, 6, mgen000448. [Google Scholar] [CrossRef] [PubMed]
- Feurtey, A.; Stevens, D.M.; Stephan, W.; Stukenbrock, E.H. Interspecific gene exchange introduces high genetic variability in crop pathogen. Genome Biol. Evol. 2019, 11, 3095–3105. [Google Scholar] [CrossRef] [PubMed]
- Steenkamp, E.T.; Wingfield, M.J.; McTaggart, A.R.; Wingfield, B.D. Fungal species and their boundaries matter—Definitions, mechanisms and practical implications. Fungal Biol. Rev. 2018, 32, 104–116. [Google Scholar] [CrossRef]
- Harrison, R.G.; Larson, E.L. Hybridization, introgression, and the nature of species boundaries. J. Hered. 2014, 105, 795–809. [Google Scholar] [CrossRef]
- Ferreira, E.M.; Harrington, T.C.; Thorpe, D.J.; Alfenas, A.C. Genetic diversity and interfertility among highly differentiated populations of Ceratocystis fimbriata in Brazil. Plant Pathol. 2010, 59, 721–735. [Google Scholar] [CrossRef]
- Harrington, T.C.; Thorpe, D.J.; Alfenas, A.C. Genetic variation and variation in aggressiveness to native and exotic hosts among Brazilian populations of Ceratocystis fimbriata. Phytopathology 2011, 101, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Nkuekam, G.K.; Wingfield, M.J.; Roux, J. Ceratocystis species, including two new taxa, from Eucalyptus trees in South Africa. Australas. Plant Pathol. 2013, 42, 283–311. [Google Scholar] [CrossRef]
- Depotter, J.R.L.; Seidl, M.F.; Wood, T.A.; Thomma, B.P.H.J. Interspecific hybridization impacts host range and pathogenicity of filamentous microbes. Curr. Opin. Microbiol. 2016, 32, 7–13. [Google Scholar] [CrossRef]
- Brasier, C. The rise of the hybrid fungi. Nature 2000, 405, 134–135. [Google Scholar] [CrossRef]
- Xu, J. The inheritance of organelle genes and genomes: Patterns and mechanisms. Genome 2005, 48, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Birky, C.W. The inheritance of genes in mitochondria and chloroplasts: Laws, mechanisms, and models. Annu. Rev. Genet. 2001, 35, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Birky, C.W. Uniparental inheritance of mitochondrial and chloroplast genes: Mechanisms and evolution. Proc. Natl. Acad. Sci. 1995, 92, 11331–11338. [Google Scholar] [CrossRef]
- Xu, J.; Li, H. Current perspectives on mitochondrial inheritance in fungi. Cell Health Cytoskelet. 2015, 7, 143. [Google Scholar] [CrossRef]
- Wilson, A.J.; Xu, J. Mitochondrial inheritance: Diverse patterns and mechanisms with an emphasis on fungi. Mycology 2012, 3, 158–166. [Google Scholar] [CrossRef]
- Reboud, X.; Zeyl, C. Organelle inheritance in plants. Heredity 1994, 72, 132–140. [Google Scholar] [CrossRef]
- Mendoza, H.; Perlin, M.H.; Schirawski, J. Mitochondrial inheritance in phytopathogenic fungi - everything is known, or is it? Int. J. Mol. Sci. 2020, 21, 3883. [Google Scholar] [CrossRef]
- Sun, S.; Fu, C.; Ianiri, G.; Heitman, J. The pheromone and pheromone receptor mating-type locus is involved in controlling uniparental mitochondrial inheritance in Cryptococcus. Genetics 2020, 214, 703–717. [Google Scholar] [CrossRef]
- Barr, C.M.; Neiman, M.; Taylor, D.R. Inheritance and recombination of mitochondrial genomes in plants, fungi and animals. New Phytol. 2005, 168, 39–50. [Google Scholar] [CrossRef]
- Xu, J.; Yan, Z.; Guo, H. Divergence, hybridization, and recombination in the mitochondrial genome of the human pathogenic yeast Cryptococcus gattii. Mol. Ecol. 2009, 18, 2628–2642. [Google Scholar] [CrossRef]
- Depotter, J.R.L.; Beveren, v.F.; Moreno, L.; Kramer, H.M.; Chavarro Carrero, E.A.; Fiorin, G.L.; Berg, v.d.G.C.M.; Wood, T.A.; Thomma, B.P.H.J.; Seidl, M.F. The interspecific fungal hybrid Verticillium longisporum displays subgenome-specific gene expression. mBio 2021, 12, e0149621. [Google Scholar] [CrossRef]
- Hénault, M.; Marsit, S.; Charron, G.; Landry, C.R. Hybridization drives mitochondrial DNA degeneration and metabolic shift in a species with biparental mitochondrial inheritance. Genome Res. 2022, 32, 2043–2056. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, J. Mitochondrial genome polymorphisms in the human pathogenic fungus Cryptococcus neoformans. Front. Microbiol. 2020, 11, 706. [Google Scholar] [CrossRef]
- Lee, D.-H.; Roux, J.; Wingfield, B.D.; Wingfield, M.J. Non-Mendelian segregation influences the infection biology and genetic structure of the African tree pathogen Ceratocystis albifundus. Fungal Biol. 2018, 122, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.D.; Bloom, R.J.; Jiang, S.; Durand, H.K.; Dallow, E.; Mukherjee, S.; David, L.A. Measuring and mitigating PCR bias in microbiota datasets. PLoS Comput. Biol. 2021, 17, e1009113. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Ceratocystis growing on MEA media, showing the sexual fruiting bodies (ascomata). Ascomata have round, dark bases with long necks, at the top out of which slimy masses of ascospores exude.
Figure 1.
Ceratocystis growing on MEA media, showing the sexual fruiting bodies (ascomata). Ascomata have round, dark bases with long necks, at the top out of which slimy masses of ascospores exude.

Figure 2.
A cross of C. fimbriata and C. eucalypticola following incubation on MEA-TS medium. a) Parallel blue lines show the zone of interaction. b) Close-up of the interaction zone.
Figure 2.
A cross of C. fimbriata and C. eucalypticola following incubation on MEA-TS medium. a) Parallel blue lines show the zone of interaction. b) Close-up of the interaction zone.
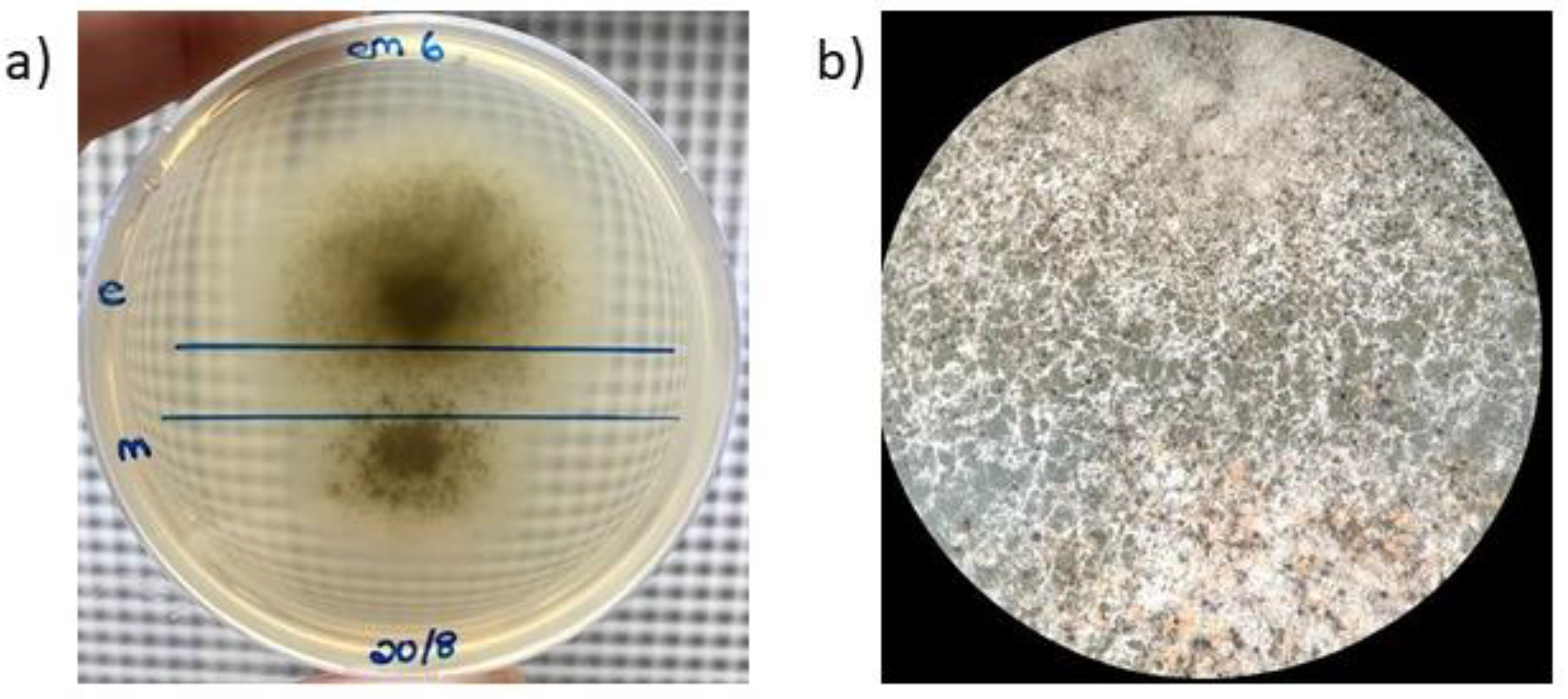
Figure 3.
In silico PCR-RFLP analysis of the three nuclear genomic regions containing HindIII and PstI restriction enzyme cut site variation in the examined Ceratocystis species. a) Marker region 1 contains a PstI cut site that is absent only in C. manginecans. b) Marker region 2 contains a PstI cut site present only in C. eucalypticola. c) Marker region 3 has PstI cut site unique to C. fimbriata. d) Predicted profiles following digestion of the three marker regions with HindIII and PstI (molecular marker in base pairs).
Figure 3.
In silico PCR-RFLP analysis of the three nuclear genomic regions containing HindIII and PstI restriction enzyme cut site variation in the examined Ceratocystis species. a) Marker region 1 contains a PstI cut site that is absent only in C. manginecans. b) Marker region 2 contains a PstI cut site present only in C. eucalypticola. c) Marker region 3 has PstI cut site unique to C. fimbriata. d) Predicted profiles following digestion of the three marker regions with HindIII and PstI (molecular marker in base pairs).
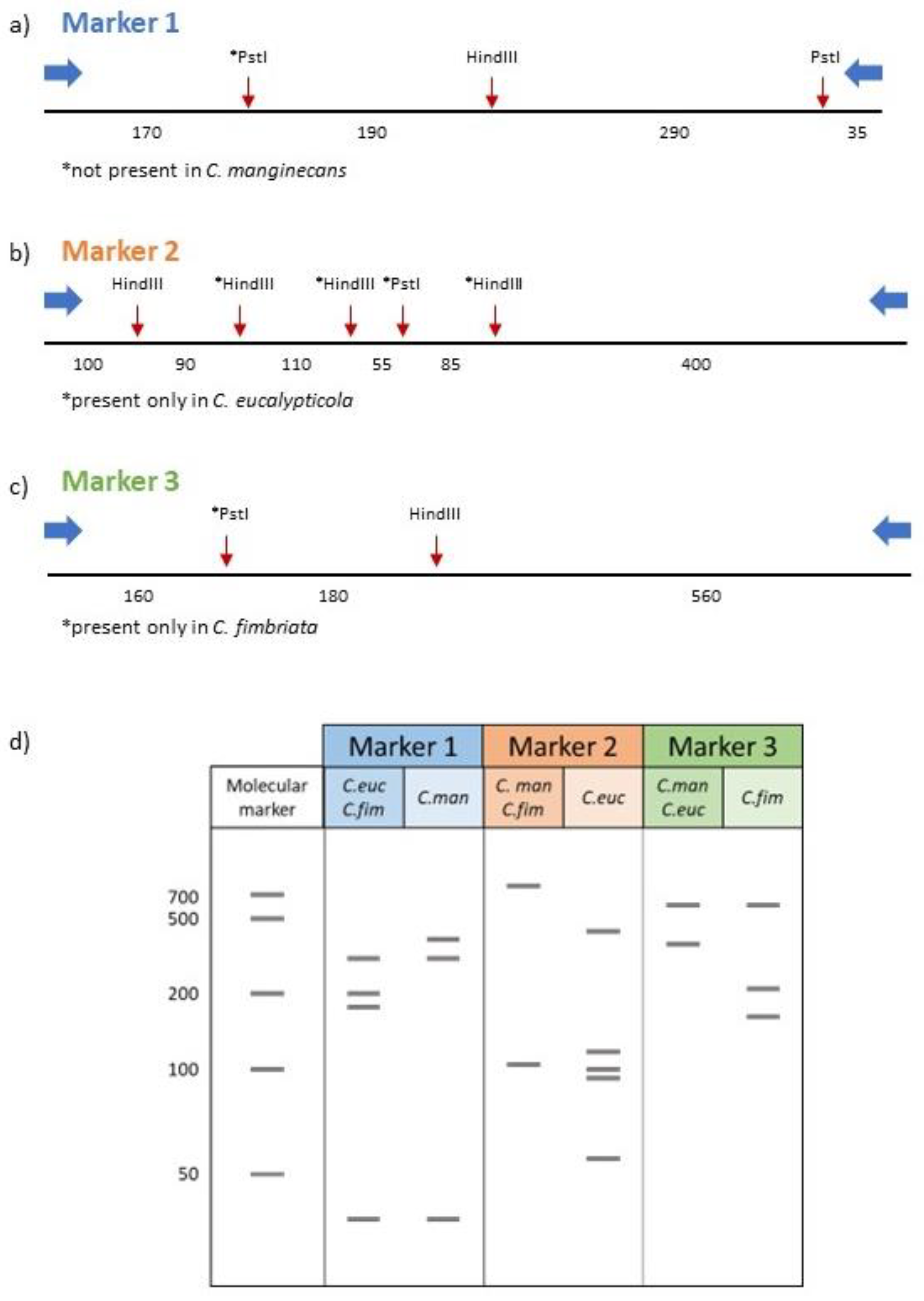
Figure 4.
Agarose gel showing the RFLP profiles of the diagnostic nuclear markers for 23 spore masses of the C. fimbriata x C. eucalypticola mating. Lanes without numbers indicate the profile for the respective parent isolate. Also shown are 23 collected spore drops showing either the profile of a parent isolate, or a mix of profiles indicating hybridization. Sizes for the reference markers are given, as well as fragment sizes for the parents and spore mass 2.
Figure 4.
Agarose gel showing the RFLP profiles of the diagnostic nuclear markers for 23 spore masses of the C. fimbriata x C. eucalypticola mating. Lanes without numbers indicate the profile for the respective parent isolate. Also shown are 23 collected spore drops showing either the profile of a parent isolate, or a mix of profiles indicating hybridization. Sizes for the reference markers are given, as well as fragment sizes for the parents and spore mass 2.
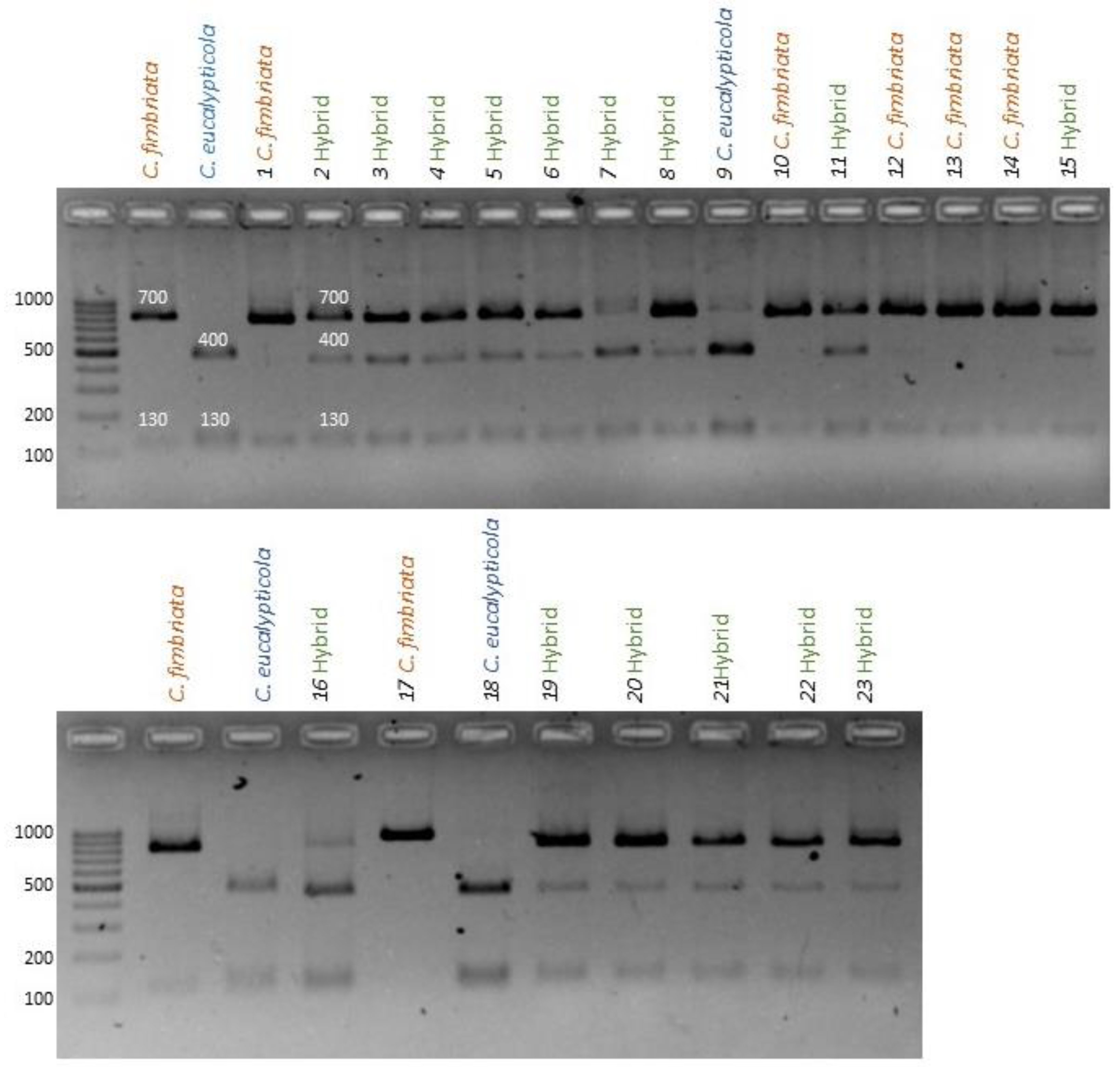
Figure 5.
Agarose gel showing amplification products of the diagnostic regions of the mitochondrial genomes of C. fimbriata, C. manginecans and C. eucalypticola. Primer set 1 produces an amplicon of around 1890 bp in C. fimbriata and C. eucalypticola, and an amplicon of around 560 bp in C. manginecans. Primer set 2 produces an amplicon of nearly 400 bp in both C. fimbriata and C. eucalypticola. An approximately 500 bp fragment was produced in C. manginecans although this fragment was not predicted in the initial in silico analysis. Primer set 3 only produces an amplicon in isolates containing mitochondrial DNA of C. fimbriata.
Figure 5.
Agarose gel showing amplification products of the diagnostic regions of the mitochondrial genomes of C. fimbriata, C. manginecans and C. eucalypticola. Primer set 1 produces an amplicon of around 1890 bp in C. fimbriata and C. eucalypticola, and an amplicon of around 560 bp in C. manginecans. Primer set 2 produces an amplicon of nearly 400 bp in both C. fimbriata and C. eucalypticola. An approximately 500 bp fragment was produced in C. manginecans although this fragment was not predicted in the initial in silico analysis. Primer set 3 only produces an amplicon in isolates containing mitochondrial DNA of C. fimbriata.
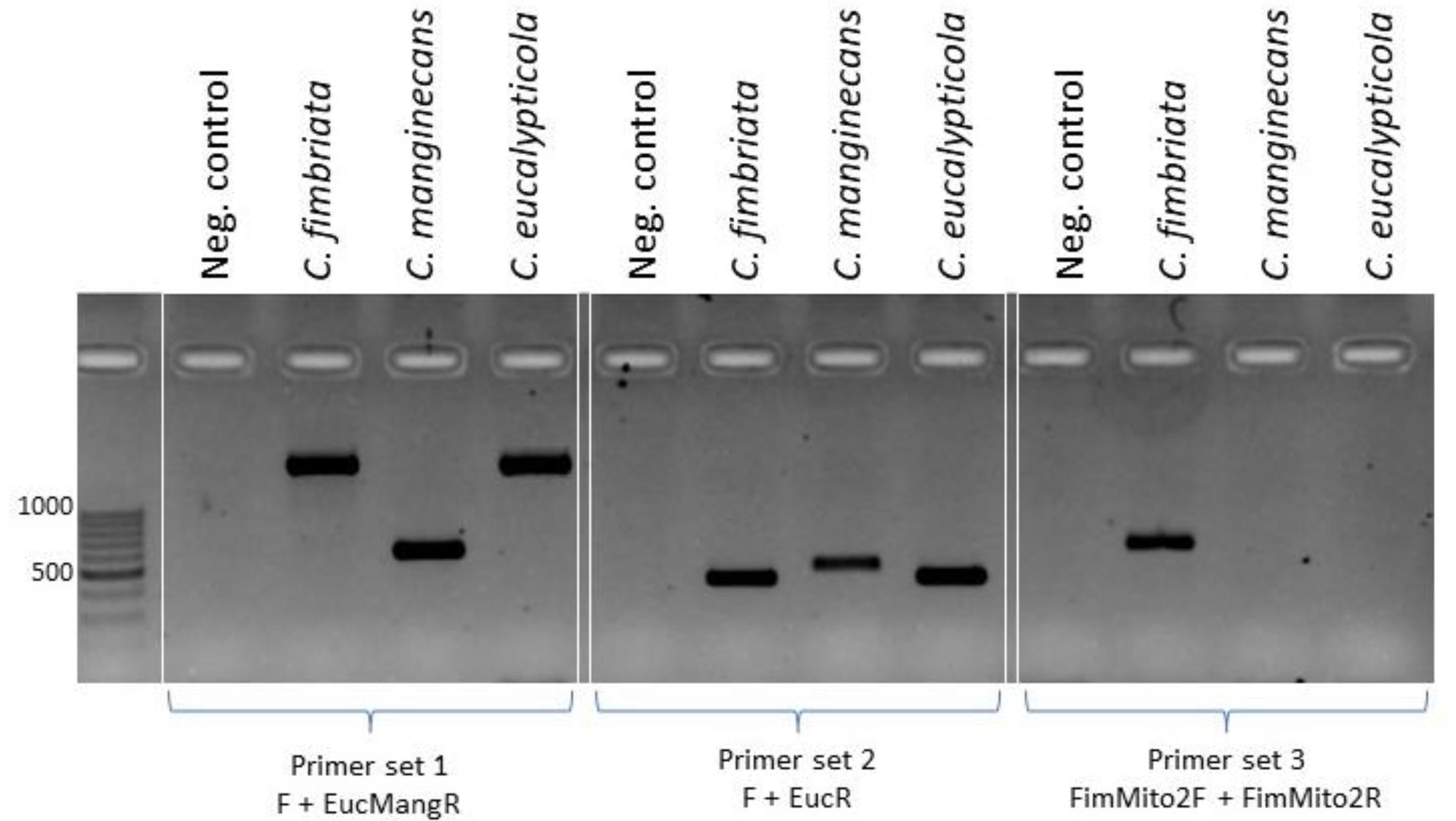

Table 1.
List of primers used in this study and notes on their specific usage in the various experiments.
Table 1.
List of primers used in this study and notes on their specific usage in the various experiments.
| Primer name* | Sequence (5’ → 3’) | Size (bp) | Notes |
|---|---|---|---|
| marker1_R | TTTCTGCTGTCCACACCTTG | 692 | Nuclear primer set 1. Region amplified from all three species but restriction enzyme (RE) digestion produces unique banding pattern for C. manginecans. |
| marker1_F | TTTCTGCTGTCCACACCTTG | ||
| marker2_R | GACCGCATGGTTGAGGTTAC | 836 | Nuclear primer set 2. Region amplified from all three species but RE digestion produces unique banding pattern for C. eucalypticola. |
| marker2_F | GTTCATGATGCCATCGACAC | ||
| marker3_R | GTGTAGCCGTCGGAAAATGT | 908 | Nuclear primer set 3. Region amplified from all three species but RE digestion produces unique banding pattern for C. fimbriata. |
| marker3_F | TGTTGGATGGGCTGTATTGA | ||
| Cf_Mt_F1 | GAAGTGCCTTCGCTTTAT | 436 | Targets the MAT1-1-1 gene in all three species. |
| Cf_Mt_R1 | GACCGCGATTCTAACCAAAA | ||
| MAT1-2-1F | AAGATGCTCTTTAATACCCACCA | 495 | Targets the MAT1-2-1 gene in all three species. |
| MAT1-2-1R | TGCCGCTAATAAGCTAGGAA | ||
| F | AATTGGATCTTCAACGACTAAAC | 562 1886 1895 |
Mitochondrial primer set 1. Amplicon of 562 in C. manginecans, and amplicons of 1886 and 1895 bp in C. eucalypticola and C. fimbriata. |
| EucMangR | GGGCTCTGTTAGTCTCTGCA | ||
| F | AATTGGATCTTCAACGACTAAAC | 387 387 |
Mitochondrial primer set 2. Amplicons of 387 and 387 bp in C. eucalypticola and C. fimbriata. |
| EucR | TCCGCGATCATCCATTTCTC | ||
| FimMito2F | CCTGCATCTCGTCCTATCGT | 559 | Mitochondrial primer set 3 amplifies only in C. fimbriata. |
| FimMito2R | TCGCCTTTCAAGTTCCATGC | ||
| feMitSeq_F | CATTTGGGGGCTTTTTGTAA | n/a | Primer used to sequence amplicons produced with mitochondrial primer set 1 in order to score the region of variability between C. eucalypticola and C. fimbriata. |
* Primer F was used in both mitochondrial primer sets 1 and 2.
Table 2.
Summary of the results for the PCR-RFLP and mitochondrial PCR analyses for the two rounds of crossing experiments conducted in this study (see Tables S1 and S2 for detail).
Table 2.
Summary of the results for the PCR-RFLP and mitochondrial PCR analyses for the two rounds of crossing experiments conducted in this study (see Tables S1 and S2 for detail).
| Interspecific cross | Hybridization | Selfing | Number of cultures containing hybrids | Mitochondrial inheritance in hybrids |
|---|---|---|---|---|
| C. fimbriatax C. eucalypticola | ||||
| Only one nuclear marker | 11 | 24 | 26 of 50 | Both parents |
| Both nuclear markers | 15 | 24 | ||
| Both mitochondrial markers | 6 | n/a | ||
| C. fimbriatax C. manginecans | ||||
| Only one nuclear marker | 8 | 41 | 8 of 49 | Both parents |
| Both nuclear markers | 0 | 41 | ||
| Both mitochondrial markers | 8 | n/a | ||
| C. manginecansx C. eucalypticola | ||||
| Only one nuclear marker | 0 | 42 | 0 of 42 | n/a |
| Both nuclear markers | 0 | 7 | ||
| Both mitochondrial markers | n/a | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



