
Submitted:
01 May 2023
Posted:
02 May 2023
You are already at the latest version
Abstract
Fermented food matrices, including beverages, can be defined as the result of the activity of complex microbial ecosystems where different microorganisms interact according to different biotic and abiotic factors. Certainly, in industrial production the technological processes aim to control the fermentation to place safe foods on the market. Therefore, if food safety is the essential prerogative, consumers are increasingly oriented towards a healthy and conscious diet driving the production and consequently the applied research towards natural processes. In this regard, the aim to guarantee safety, quality and diversify of products should be reached limiting or avoiding the addition of antimicrobials or synthetic additives using the biological approach. In this paper, following the recent re-evaluation of non-Saccharomyces yeasts (NSYs) has been re-viewed the bio-protectant and biocontrol activity with a particular focus on their antimicrobial power, probiotic features and promoting functional aspects. In this review, the authors would underline the contribution of NSYs in all food production chain and its role in the technological and fermentative features for their practical and useful use as biocontrol agent in food preparations.
Keywords:
biocontrol
; non-Saccharomyces yeasts
; functional yeasts
; probiotics
1. Introduction
Traditional industrial attributes to Saccharomyces cerevisiae the most important role as biotechnological organism more involved in worldwide fermentation products such as beers, cider, wines, sake, distilled spirits, bakery products, cheese, sausages, and other fermented foods. However, in the last 20 years, the world of research and in parallel the industrial one, started to re-evaluate the potential positive contribution of non-Saccharomyces yeasts (NSYs). They have found a pro-technological use in traditional fermentations, where can impart peculiar and distinctive characteristics to the product, but also in other applications as: biomedical or fundamental biological research, environmental biotechnology, heterologous protein production, biocontrol and food and feed sectors [1]. The utility of NSYs in food field is accelerating due to a wide number of metabolic abilities that confer aroma and structure to the final products but also due to their plasticity in assimilating different substrates. They generally show low fermentation yields, and higher sensitivity to ethanol stress if compared with S. cerevisiae. On the other hand. NSYs may display a great range of fermentation metabolites and end products in providing distinctive features [2].
In any case, the positive or negative contribution that each non-Saccharomyces species gives to the product strictly depends on the matrix and fermentation context (Figure1).
Figure 1.
Identification of key strengths (S), weaknesses (W), opportunities (O) and threats (T) related to the use of non-Saccharomyces yeasts (NSYs) in fermented foods and beverages.
Figure 1.
Identification of key strengths (S), weaknesses (W), opportunities (O) and threats (T) related to the use of non-Saccharomyces yeasts (NSYs) in fermented foods and beverages.
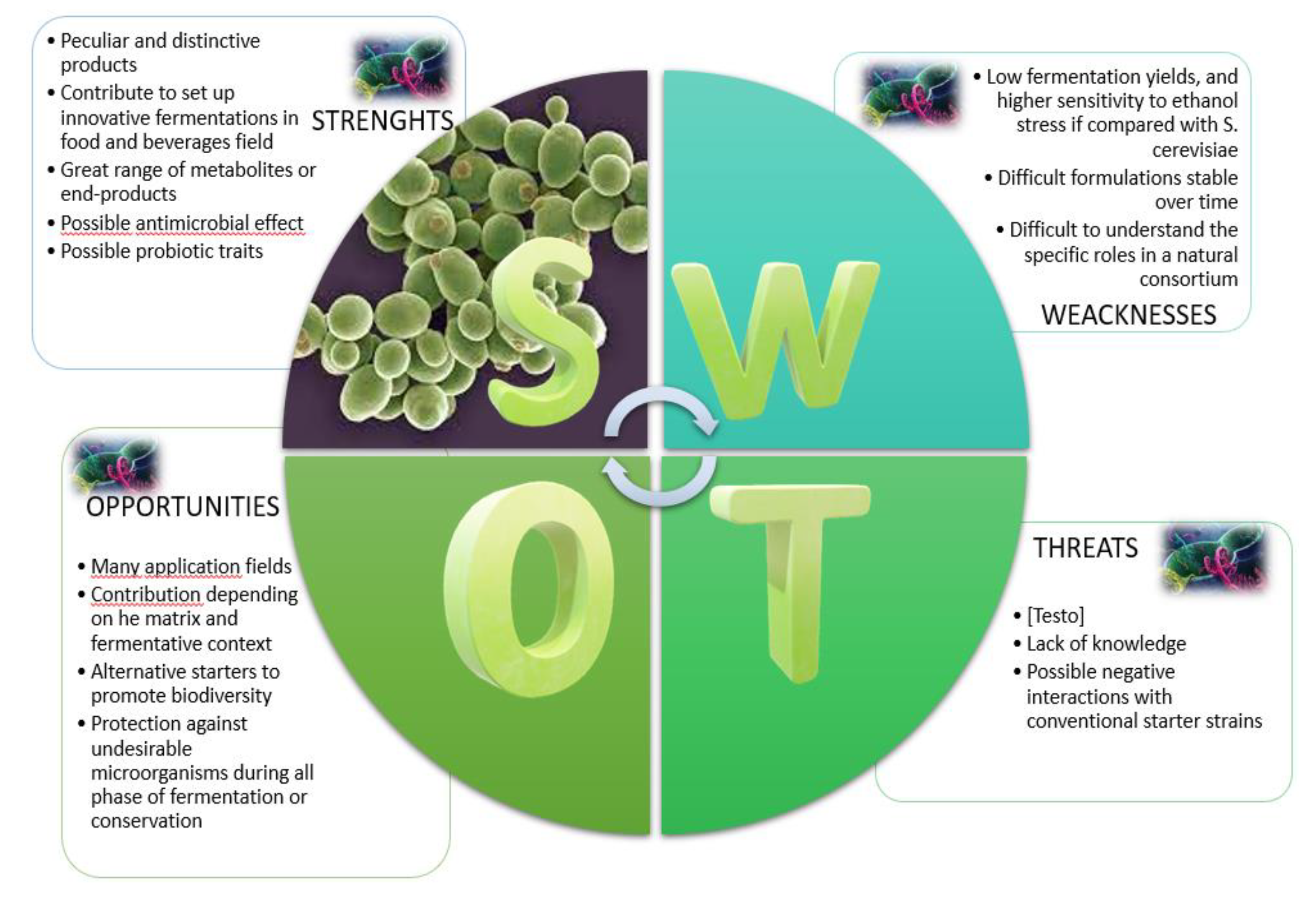
Until a few decades ago, in the traditional fermented beverages, NSYs were considered synonymous of spoilage yeasts. Differently, today many researchers clearly highlighted their potential for specie-specific or strain-specific traits, and it is unequivocally accepted that NSYs represent an important resource with economic repercussions in the market of fermented products. In the worldwide market, there is a growing interest in “non-conventional” yeast strains, another way to define NSYs, that can help generate the diversity and the complexity desired in diversified and aware consumers. Effectively, selected strains of NSYs contribute to set up innovative fermentations in food and beverages field, starting to winemaking, continuing in brewing where impart specific “bioflavouring” or functionalizing the beverages and reducing the final ethanol content. In addition, NSYs may be used as alternative starters to promote biodiversity and quality of other different fermented foods such as bakery products. In a recent paper, some selected non-Saccharomyces wine strains were assayed in the production of leavened doughs [3]. Several NSYs species as Hanseniaspora uvarum, Metschnikowia pulcherrima, Pichia kudriavzevii, Torulaspora delbruekii, and Zygotorulaspora florentina could be advantageous in the bakery industry, providing greater diversity than S. cerevisiae-based products, and may be useful in reducing or avoiding yeast intolerance.
Some NSYs including Kluyveromyces lactis, have been able to survive in the conditions that mimic the gastrointestinal environment and can form biofilms on an abiotic medium such as polystyrene. These yeast strains also exhibited highly hydrophilic cell wall surface properties and adhesion ability to intestinal Caco-2 cells, thus increasing their potential as probiotic strains. Effectively, in the pharmacological industry, the rational design of probiotics includes, in addition to their health claims, several other criteria: stability during manufacturing processes, viability during gastrointestinal transit, and functionality at the desired target site. Host safety and the management of their formulation and viability are also determining characteristics in the selection of probiotics. In relation to these multiple traits, the last decade has seen much evidence regarding NSYs as probiotic candidates. For example, a report by Li and colleagues [4] clarified the GRAS status of several Kluyveromyces species. In a general picture, yeast may offer more benefits than bacteria, as they are insensitive to antibiotics and can be easily used for the treatment of antibiotic-associated diarrhea.
The probiotic and biocontrol features as well as the general interest that revolves around the characteristics and the use of NSYs in the agri-food industry are the concerns of the present review. In this regard, an updated survey of the main recent publications focusing on this topic with scientific and market implications is prepared. The antimicrobial and probiotic attitudes is analyzed with particular attention to pointing out the recent findings, indicating the perspectives and emphasizing the fields of interest that are still poor investigated.
2. Antimicrobial activity of non-Saccharomyces yeasts (NSYs)
Already more than twenty years ago some authors [5,6] anticipated the potential use of NSYs during the first step of fermentation applied to improve the final flavor of wines. Today, knowledge has exponentially increased and the aromatic enhancement of wines due to the use of NSYs is only one of the many other features identified for their potential application. In view of the possible use in agri-food industry, it is a common prerogative that NSYs must possess specific traits to be selected. Indeed, after the general revaluation of the role of NSYs, several studies have focused the attention on multiple advantages particularly in winemaking. Among these, the antagonistic activity against undesired microorganisms is of paramount importance.
It has been widely demonstrated that some NSYs strains in winemaking can control spoilage yeasts or filamentous fungi both in vineyard and during the early stages of fermentation [7-9]. In this regard, spoilage species producing off-flavours or films in bulk wine such as Zygosaccharomyces rouxii, Kloeckera apiculata, Pichia spp., Candida spp. may be prevented by the use NSYs through active antimicrobial extracellular molecules released [10]. Effectively, the growing interest in reducing and rationalizing the use of SO2 in winemaking has urged scientific investigations on the antimicrobial activity of NSYs as alternative to chemical conventional additives [11, 12]. About this, natural control strategies may involve the use of killer toxins (mycocins), antimicrobial peptides such as Lactoferricin B or volatile compounds produced by NSYs as biological strategy to counteract contamination [13].
2.1. Biological control
The term “biocontrol” related to the use of microorganisms as natural biological agents was defined as the reduction of pathogen or disease activities through organism or their molecules. In agri-food this concept is related to an alternative strategy to the use of chemical products, and the use of microorganisms with antagonist action against other microorganisms lowering the use of pesticides and boosting food quality and safety [14-16].
In recent years, to avoid the huge losses of fruit and vegetables due to pathogens, biological strategies have been studied as alternatives to products of a chemical nature. The addition of microorganisms as bio-protective agents or their antimicrobial products have already been identified as “bio-protection”. This practice is firstly used in agriculture and then in food industries for the control of fruits decay from post-harvest spoilage microorganisms, and the relative extension of their shelf life. This strategy consists in the inoculation of viable antagonistic microorganisms [bacteria, yeasts, or a mixture of them) or the addition of their antimicrobial products in complete or partial purified form, during, at the end or after the chain production [17-19]. Biological control based on the use of beneficial microorganisms is receiving increasing recognition, although the number of registered and approved marketed bioproducts containing bioactive yeasts or bacteria remains scarce. Czajkowski and collaborators [20] argues that this situation arises from objective difficulties encountered during registration and marketing, but also from problems of understanding the specific roles of each member of a consortium and their biological activity. Regarding to the application of NSYs in bio-protection strategies against grapevine trunk diseases in the vineyard [the fungal pathogen Botrytis cinerea causing bunch sour rot] has increased significantly in recent years [21,22]. In addition, there is a growing interest in the application of this approach on the protection against undesirable microorganisms during all wine production chain as well as during the conservation and maturation phases [23,24]. The renewed interest in biological control is due to the growing attention to the use of sulphites in food and particularly in wine. In this regard NSYs has been proposed as a possible effective natural alternative. Several recent studies have been carried out by using selected strains of M. pulcherrima and T. delbrueckii at the prefermentative stage in the red winemaking process [23,25-26]. Other studies focused the attention on the use of other selected strains of M. pulcherrima during cold clarification stage of Italian Verdicchio white variety underlining the double role of this yeast as biocontrol agent and wine aroma enhancer [27].
The antimicrobial activity of NSYs has been investigated also in other sectors since the re-evaluation of the use of these yeasts with biocontrol purposes has been extended across the food industry. Some applicative studies showed their effectiveness also in bread-making or cheesemaking. Valsaraj and colleagues [28] highlighted the biological load of some NSYs and how their toxic killer effects could be used in the food and beverage industry, maintaining food quality, and improving the safety in beer, cheese, or bread. They suggested that the biocontrol strategy of NSYs isolated from food and beverage naturally fermented, may be effective in suppressing wild yeast strains during another fermentation.
No less important is the involvement of the antimicrobial role of yeast strains in the medical field. For over thirty years, Polonelli and Morace [29] reported the killer phenomenon against bacteria, seeing the possibility of using these toxins to counteract the growing phenomenon of antibiotic resistance. In a more recent study, the yeasts D. hansenii, Pichia fermentans, Candida tropicalis and Wickerhamomyces anomalus have been shown to induce bacterial lysis [30]. Chen et al. [31] isolated mycocin-producing strains of Kluyveromyces marxianus and demonstrated that crude extracts were effective in preventing Escherichia coli disease in mice. In addition, the yeasts Kluyveromyces lactis and K. marxianus, isolated from cheese, were able to inhibit the growth of pathogenic microorganisms, such as Listeria monocytogenes and Candida albicans [32]. Finally, C. albicans strains isolated from children had inhibitory effects on strains of E. coli ATCC 25 922 and Staphylococcus aureus ATCC 25 923 [33].
2.2. Modalities of antimicrobial action of NSYs
The regulation and the control of the growth of undesired microorganisms could be exploited through different mechanisms as space and nutrient competition, cell to cell contact or antagonistic action mediated by antimicrobial compounds production as mycocins, small peptides, VOCs or extracellular vesicles.
2.2.1. Antagonistic action: competition of nutrients and space
The main mode of action of yeasts in biocontrol is the competition for space and nutrients [34]. Most organisms can starve pathogens or parasites through iron withdrawal [35]. Aureobasidium pullulans, possesses a siderophore identified as fusarinin C which has been shown to exhibit antibacterial activity [36,37]. Pulcherriminic acid is a cyclic dipeptide that complexes iron in M. pulcherrima. Studies have shown that pigment-deficient mutants of M. pulcherrima exhibited reduced or null antifungal activity, and iron deprivation of the fungal pathogen. This suggested that the production of this pigment is one of several mechanisms by which this yeast antagonizes plant pathogenic fungi [38]. However, mutants lacking the ability to synthesize pulcherriminic acid still strongly inhibited filamentous fungi, confirming that the antifungal activity was only due to iron deprivation. Therefore, the exact contribution of iron chelators to the yeast biocontrol activity remains to be clarified in detail. Another competition for nutrients was found in Saccharomycopsis schoenii that it is unable to assimilate sulfur, a specific feature of the genus Saccharomycopsis. In addition, phytopathogenic fungi and Trichoderma species exhibited a similar phenomenon, which may indicate that methionine is an important target for such organisms and has been hotly contested [39]. Pioneering experiments aimed at evaluating the suitability of an easily transformable Pichia (formerly Ogataea) angusta haploid strain to identify biocontrol-minus mutant clones: while the wild-type strain proved effective in reducing brown rot lesion caused by Monilinia fructicola on apple fruit, its derivate leucine-auxotrophic mutant L1 had no significant effect in controlling the pathogen. The addition of exogenous leucine fully restored the biocontrol capability of mutant L1, whereas a leucine stand-alone treatment showed no significant biocontrol effect [40].
Another strategy to compete for the space, is the formation of biofilm. Biofilms are microbial communities consisting of a single or more species and are considered virulence factors for pathogenic microbes [41,42]. The development of a yeast biofilm begins with the adhesion of single cells to a surface and usually involves cell wall modifications, secretion of an extracellular matrix, and often the formation of hyphae or pseudo-hyphae [43]. In yeasts to be used in biocontrol activity, biofilm formation in wounds is now considered an important mode of action even though the molecular basis of the process and the composition of different biofilms have only been studied in detail for P. fermentans. Biofilm formation of P. fermentans in apple wounds protects against post-harvest disease, while on peaches it changes from yeast-like to hyphal growth form and causes rapid decay of inoculated fruit in the absence of a plant pathogen [44]. Biofilm formation has been demonstrated besides that S. cerevisiae also in P. kudriavzevii, W. anomalus and M. pulcherrima [45-48].
The saprophytic yeast-like fungus A. pullulans has been well documented for over 60 years in microbiological literature for its ubiquitarian presence in soil, water, rock surfaces, in both cold and warm areas. A lot of A. pullulans strains are known to produce a wide range of natural antimicrobials useful for biocontrol applications against plant pathogens mediated by nutrient and space competition and/or VOC production (see paragraph 2.1.6). A. pullulans may can be used at both vineyards, for the management of grey mold disease caused by B. cinerea, and winery levels, through the synthesis of antifungal compounds, providing a versatile tool for the viticulturist/farmer as well as for the oenologist to combat problems in the field and create a high-quality wine.
2.2.2. Mycocins
Mycocins (killer toxins) are the most investigated yeast antimicrobial compounds. Since Bevan and Makower [49] discovered the killer phenomenon in S. cerevisiae, several other yeast species have been found to produce a toxic proteinaceous factor that kills sensitive strains [50,51]. Several potential applications for the killer phenomenon have been suggested in food industry to control spoilage yeasts in the preservation of food and beverages [52,53].
The mycocin Kpkt produced by Tetrapisispora phaffii was first described as anti-spoilage yeasts [52]. Kpkt acts through a specific β-glucanase activity causing irreversible modifications on the cell wall structure and it is codified by TpBGL2 chromosomal gene [17, 54,55]. The recombinant toxin (rKpkt) was recently obtained by transferring the Kpkt coding gene in Komagataella phaffii (formerly Pichia pastoris) [56]. The recombinant rKpkt, when expressed in K. phaffii, displayed a wider spectrum of action than its native yeast [57], reinforcing the idea of the possible application of mycocins in the food and beverages industries. T. phaffii was used in mixed fermentation at pre-fermentative stage to control wild yeasts such as Hanseniaspora, Zygosaccharomyces and Saccharomycodes in the place of sulfur dioxide [11].
Several works have been focused on the study of NSYs able to counteract the development of Brettanomyces spp., a relevant dangerous yeast in winemaking [58]. In this context, Pikt and Kwkt mycocins, produced by W. anomalus (formerly Pichia anomala) and Kluyveromyces wickerhamii, respectively [54], able to counteract Dekkera/Brettanomyces. Pikt is an ubiquitin-like protein of about 8 kDa able to interact with β-1,6-glucan of cell wall of sensitive yeasts [59] while Kwkt, a protein of about 72 kDa of molecular mass, without any glycosyl residue [11] and β-1,6-glucosidase activity that seems to be in involved in to act blocking the cell cycle function of sensitive yeasts [12]. Cytofluorimetric evaluation showed that both Pikt and Kwkt caused irreversible death of this yeast and in a different way from sulphur dioxide that induced a viable but non-cultivable (VBNC) state of Brettanomyces with consequence recovery of yeasts when fresh medium was replaced [12]. Another P. membranifaciens strain showed a killer action producing two mycocins denominated PMKT and PMKT2. PMKT binds linear (1→6)-β-d-glucans in the cell wall and Cwp2p plasma membrane receptor of sensitive yeasts leading alterations in ionic exchange via plasma membrane [60]. PMKT2, a protein with an apparent molecular mass of 30 kDa, binds mannoproteins and induces cell cycle blockage an early S-phase of sensitive yeasts and stimulates markers of cellular apoptosis such as the cytochrome c release, DNA strand breaks, metacaspase activation and production of reactive oxygen species at low dose [61]. The killer activity of P. membranifaciens was exploited in wine making in mixed fermentation with S. cerevisiae and B. bruxellensis [inoculum ratio of 1:1]. P. membranifaciens inhibited B. bruxellensis growth without any effects on the fermentation activity of S. cerevisiae.
Other mycocins (CpKT1 and CpKT2) active against B. bruxellensis are produced by Candida pyralidae. Both mycocins were active and stable at pH 3.5–4.5 and with the general conditions of winemaking environment [62]. Both mycocin CpKT1 and C. pyralidae viable yeasts were used in mixed fermentation in red grape juice containing B. bruxellensis determining a decrease in spoilage yeast concentration [63].
Also, a strain of W. anomalus was proposed as a biocontrol agent against Brettanomyces/Dekkera spp. [18]. The killer activity of W. anomalus is expressed through the release of KTCf20, a mycocin. This mycocin was able to counteract the growth of different spoilage yeasts such as Brettanomyces/Dekkera, Pichia guilliermondii and P. membranifaciens. Moreover, they showed that W. anomalus in mixed fermentation did not negatively affect S. cerevisiae strains. Another mycocin named WA18 and active against B. bruxellensis is produced by an autochthonous W. anomalus strain isolated from soil pit, exhibited 99 % identity with UDP-glycosyltransferase protein [64]. In accordance with de Ullivarri and co-workers [18] the compatibility of this W. anomalus strain in mixed fermentation with S. cerevisiae yeast was confirmed. Another killer strain belonging to W. anomalus has been investigated for its wide potential of antibacterial activity against numerous human pathogenic agents [65]. Interest in these antibacterial mycocins was revealed by Muccilli and Restuccia, [66] that highlighted the potential use against pathogens, resistant to conventional antibiotics, such as Staphylococcus aureus.
In a recent work, a comparative evaluation of formulates (semi-purified and lyophilized broth) of three well characterized mycocins were assessed against B. bruxellensis. The absence of dangerous effects toward human epithelial cells, opening the way for their possible commercial application [67]. Another NSYs that shows antimicrobial activity is T. delbrueckii where a mycocin exhibited glucanase and chitinase activities, it is stable in wine environmental condition and it is active against B. bruxellensis, and other potential wine spoilage yeasts. Moreover, Ramírez et al. [68] isolated and selected T. delbrueckii strain that produces a mycocin [Kbarr-1]. This toxin is encoded by a dsRNA, TdV-Mbarr-1, structurally like M dsRNAs of S. cerevisiae, both seem to be evolutionarily related [69].
2.2.3. Antimicrobial peptides (AMPs)
Some peptides produced by yeasts have shown antimicrobial effects against several grape-must/wine contaminating yeasts. In general, these peptides show lengths of up to 100 amino acids, sorted into variable sequences and the mode of action involves the disruption of the cell wall in sensitive strains. For example, small peptides with molecular mass below 5 kDa produced by Candida intermedia have shown greater antimicrobial specific effects against B. bruxellensis [70]. In addition, it was demonstrated the antibacterial activity of the same strain against Escherichia coli, L. monocytogenes and S. typhimurium [71]. Similar observations have been reported by Younis et al. [72], where three isolates of C. intermedia from raw milk and fruit yoghurt showed antimicrobial activity against E. coli, S. aureus and Pseudomonas aeruginosa.
Another mechanism of interaction in mixed fermentation is the possible involvement of extracellular vesicles (EVs). In this regard, a recent work on exo- proteome of EV-enriched fractions in pure and mixed fermentation with six different species of NSYs and S. cerevisiae showed a wide diversity of proteins secreted indicating the presence of interactions and the possible involvement of EVs [73]. The EV-enriched fractions from different species such as S. cerevisiae, T. delbrueckii and Lachancea thermotolerans showed enrichment in glycolytic enzymes and cell-wall-related proteins, particularly the enzyme exo-1,3-β-glucanase. However, this protein was not involved in the here-observed negative impact of T. delbrueckii extracellular fractions on the growth of other yeast species. These findings suggest that EVs may play a role in fungal interactions.
2.2.4. Secreted enzymes
Some enzymes secreted may be involved in biocontrol action. Indeed, the secretion of enzymes degrading cellular components such as chitinases, glucanases, or proteases is a common feature in all kinds of host–pathogen interactions and has been intensively studied. Chitinolytic enzymes allow degrading fungal cell walls [74]. Yeasts belonging to genera Aureobasidium, Candida, Debaryomyces, Metschnikowia, Meyerozyma, Pichia, Saccharomyces, Tilletiopsis, Wickerhamomyces and Saccharomycopsis, exhibited this enzymatic activity [16,39]. Chitinases from sources i.e. fungi and filamentous bacteria have demonstrated biocontrol activity against plant pathogenic fungi and chitinases are extensively studied as potential biopesticides, targets for resistance breeding or as transgenes in genetically modified plants. Chitinases, probably in indirect manner, influence biocontrol activity because Chito-oligosaccharides (CHOS) resulting by of chitin degradation, are potent inducers of plant immune responses [51].
Glucans are major cell wall components in fungi and exoglucanases are involved in cell wall modification, cell adhesion and resistance to mycocins [75]. A 1,3-β-glucanase from Candida oleophila was the first gene cloned in this organism and overexpression or deletion of this gene did not significantly affect Penicillium digitatum spore germination, but subsequent studies have documented a reduced inhibitory activity of the β-exoglucanase deletion mutant compared to the wild type and overexpressing strain [in vitro and in fruit], thus demonstrating the involvement of glucanases in the biocontrol activity of yeast [76].
In W. anomalus the deletion of two exo-β-glucanases (PaEXG1 and PaEXG2) significantly reduced the fruit biocontrol activity against B. cinerea [Friel et al. 2007], while the single deletion of PaEXG2 did not reduce biocontrol performance. Exoglucanase activity was also detected in several biocontrol yeasts and was linked to antagonist activity, but without demonstrating a causal involvement.
It has long been proven [77] that in Rhodotorula glutinis and Cryptococcus laurentii, β-1,3-glucanase activity did not correlate with their respective inhibitory activity against B. cinerea. The pathogenicity of yeast species belonging to Candida, Cryptococcus or Malassezia is related to lipase activity. Several studies have also correlated the role of lipases with the biocontrol action of fungi and bacteria against plate diseases. For this reason, the lipolytic activity of yeasts may represent an aspect to be investigated in relation to biological control [78].
The alkaline serine protease Alp5 of A. pullulans reduced spore germination and germ tube length of Penicillium expansum, B. cinerea, M. fructicola and Alternaria alternata in vitro and showed a concentration-dependent inhibitory effect on these pathogens on the apple tree [79]. Protease activity has been reported but not confirmed or investigated in the genera Metschnikowia, Pichia, and Wickerhamomyces, but has not been further studied or confirmed.
2.2.5. Mycoparasitism
Mycoparasitism is little studied in yeasts, some studies have shown that P. guilliermondii adheres to the hyphae of B. cinerea and causes the collapse of the hyphae, presumably due to the secretion of hydrolytic enzymes such as glucanases [80].
Species belonging to the genus Saccharomycopsis have been studied against biocontrol of several clinically relevant Penicillium species and yeasts [39].
2.2.6. Volatile organic compounds (VOCs)
Volatile organic compounds (VOCs) are the most relevant metabolites that show a biocontrol action. These metabolites are molecules <300 Da with a low solubility in water and a high vapor pressure and include molecules such as hydrocarbons, alcohols, thioalcohols, aldehydes, ketones, thioesters, cyclohexenes, heterocyclic compounds, phenols, and benzene derivatives. Volatiloma is specific for each yeast species as well as the spectrum of action against pathogenic microorganisms [81,82]. VOCs are species specific and are produced by fungi, bacteria and yeast during and their primary and secondary metabolism, limiting the growth of other microorganisms. The chemical composition of VOCs strongly depends on the environment and the pathogen being antagonized. The chemical composition includes alcohols, aldehydes, cyclohexenes, benzene derivatives, heterocyclic compounds, hydrocarbons, ketones, phenols, thioalcohols and thioesters. VOCs are produced by several yeasts species to reduce the growth of pathogens moulds. In this context, Di Francesco and co- workers [83] have demonstrated both in vitro and in vivo that the VOCs produced by A. pullulans reduced the growth and the infection of B. cinerea, Colletotrichum acutatum, P. expansum, P. digitatum and P. italicum.
Also VOCs formed by NSYs W. anomalus, M. pulcherrima and A. pullulans as well as S. cerevisiae showed a biocontrol action against B. cinerea on table grape berries [84]. Selected strains of Cyberlindnera jadinii, Candida friedrichii, C. intermedia, and L. thermotolerans inhibited the formation of both mycelial growth and ochratoxin A in Aspergillus carbonarius and Aspergillus ochraceus identifying β-phenyl ethanol as the active compound [85,86]. The VOCs by W. anomalus, preventing spore germination, mycelial growth and toxin production of Aspergillus flavus [87]. Similarly, VOCs released by Candida sake reduced the incidence of apple rot caused by P. expansum and B. cinerea [88]. The inhibitory activity of Sporidiobolus pararoseus on spore germination and mycelial growth of B. cinerea was mainly attributed to 2-ethyl-1-hexanol, whereas C. intermedia produced 1,3,5,7-cyclooctatetraene, 3-methyl1-butanol, 2-nonanone, and phenylethyl alcohol as the major components of its volatilome during the interaction with this pathogen. VOCs released by W. anomalus, Pichia kluyveri, and H. uvarum inhibited A. ochraceus growth and ochratoxin A production during the fermentation process of coffee [89].
Recently, Ruiz-Moyano et al., [90] screened yeasts isolated from figs producing antifungal VOCs. Eleven out of 34 yeasts, belonging to A. pullulans, Filobasidium oeirense, H. uvarum and Hanseniaspora opuntiae showed a reduction in the growth of B. cinerea correlated with the production of 10 volatile compounds: two acids [acetic acid and octanoic acid], 7 esters [ethyl propionate, n-propyl acetate, isobutyl acetate, 2-methylbutyl acetate, furfuryl acetate, phenylmethyl acetate, 2-phenylethyl acetate] and a ketone (heptane-2-one).
A strain Torulaspora spp. showed biological action against Alternaria arborescens causal agent of tomato fruits decay. The mechanisms involved were both the production of volatile compounds and the competition for nutrients [91]. In a screening of 147 yeasts and yeast-like fungi five strains belonging to Anthracocystis sp., two strains of Aureobasidium sp. Rhodotorula sp. and Solicococcus keelungensis produced VOCs and were active against Aspergillus flavus that produce aflatoxin B1. Alcohols, alkenes, aromatics, esters and furans. 2-phenyl ethanol and methyl benzene acetate were the most abundant compounds generated by Aureobasidium sp. On the other hand, 2-methyl-1-butanol and 3-methyl-1-butanol were significant compounds produced by the other three genera [92]. In another recent work, Candida pseudolambica showed a significant reduction such as 41.2 % of the disease incidence of gray mold in peach inoculated with B. cinerea [93]. Several modalities of action were identified: the growth dynamics, VOCs effects, parasitism, and biofilm formation. VOCs released from C. pseudolambica inhibited the mycelial growth and conidia germination of B. cinerea. Fourteen VOCs were identified with the main compounds being 3-methyl-1-butanol and 2-phenylethanol, which made up 85.90 % of the relative peak area.
2.3. Registered formulations of biocontrol NSYs species
Nowadays, there are few formulations based on yeasts suitable for plant protection that are registered and marketed such as C. oleophila, A. pullulans, Metschnikowia fructicola, C. albidus, and S. cerevisiae. However, selected NSYs are also used in postharvest to extend the shelf life of vegetable or in winery to limit the common use of conventional chemical antimicrobials, such as sulfur dioxide. For example, strains of M. pulcherrima and T. delbrueckii are commercialized as a natural biological control agents with potential to partially or fully replace sulphites. A brief description of the NSYs strains used in registred formulations present in the market is reported.
2.3.1. Candida oleophila
Species of the genus Candida strongly inhibit plant pathogens are: Candida diverse [94], Candida ernobii [95], Candida guillermondii [96], C. oleophila [37], Candida saitoana, C. sake [88,97], or Candida subhashii. These species are biocontrol agents against molds and post-harvest diseases of hazelnut and citrus fruits.
C. oleophila was the first yeast to be developed into a commercial plant protection agent and the basis for its antifungal activity several different mechanisms of action have been demonstrated. In addition to competition for nutrients and space, studies of several Candida species have identified hydrolytic enzymes such as proteases, chitinases and glucanases, as well as volatile compounds, which have been implicated in antifungal activity [98].
Furthermore, bioflm formation, high osmotolerance, induction of resistance in the plant/fruit, and direct parasitism of hyphae were shown to contribute to the biocontrol activity of Candida species [99]. To overcome the inconsistent performance of the initial Candida-based biocontrol products combinations with fungicides, different buffers [e.g., calcium chloride, bicarbonate], chitosan, or lysozyme were studied [100]. The C. oleophila strains I-182 and strains O have been developed into the biocontrol products Aspire® and Nexy®, respectively. The latter was the first biocontrol yeast to be registered against a postharvest disease [99] and C. oleophila strain O has been approved as a plant protection agent in Europe in 2015 (European Commission Health & Consumers Directorate-General 2013; European Food Safety Authority, EFSA) [101].
2.3.2. Aureobasidium pullulans
The biocontrol activity of A. pullulans has been documented in several different strains, but only DSM 14940 and DSM 14941 are registered, in admixture Botector-New (Manica), as active ingredients of plant protection products against the disease caused by the bacterium Erwinia amylovora and post-harvest diseases (European Food Safety Authority EFSA, 2013) [102]. These two strains of A. pullulans were selected based on their strong inhibition against E. amylovora. The two strains were formulated into the Blossom-Protect® product and tested over several years in different sites under field conditions [103] and also registered for post-harvest apple tree disease control as a Boni-Protect® product [104]. As with many other yeasts with a biocontrol action, the mode of action of A. pullulans involves competition for space and nutrients, but enzymatic activities such as proteases, chitinases or secreted molecules may also be involved (see above). Specific metabolites or enzymes and their contribution to the biocontrol activity of DSM 14940 and DSM 14941 have not been identified.
2.3.3. Metschnikowia spp.
M. fructicola and M. pulcherrima are the most studied yeast species regarding biocontrol action. Indeed, they can inhibit a range of postharvest and plant rot diseases, [84, 105,106]. The antifungal activity of Metschnikowia species is mediated by a range of mechanisms that involve competition for nutrients (e.g.; amino acids, iron), secretion of glucanases and chitinases, and the production of volatile organic compounds [38,74,79]. Originally, M. fructicola was isolated and discovered in Israel and developed and registered as a biocontrol product to prevent postharvest diseases, particularly in sweet potato and carrot [107]. M. fructicola has also been patented as an antagonist of plant pathogenic microorganisms [108].
2.3.4. Cryptococcus spp.
Basidiomycetes belonging to the genus Cryptococcus are isolated from water sources, soil and decomposing plant material. Criptococcus albidus, C. laurentii and Cryptococcus flavus strains have been shown to have a post-harvest protective action in peach, cherry, strawberry, tomato, citrus and pome fruit from post-harvest decay [109-111]. C. albidus was used in the preparation of Yieldplus® (Anchor Bio-Technologies in South Africa) against B. cinerea, P. expansum of pome and citrus friut and B. cinerea during post-harvest cold storage of strawberries [112]. The modality of action of this yeast is related to competition of nutrient and space. Moreover, C.albidus exhibits glucanase, chitinase and protease activity and produces unidentifed volatile compounds that inhibit fungal growth and can display killer activity against C. glabrata [113]. However, none of these mechanisms have been directly linked to the inhibitory activity of the target plant pathogens.
2.3.5. Torulaspora delbrueckii and Metschnikowia pulcherrima
A strain of M. pulcherrima registered as LEVEL2 INITIA™ is a biological tool selected with purpose of preserving the aromatic potential and protecting from oxidative phenomena in the pre-fermentation phases, limiting the use of SO2. LEVEL2 GUARDIA™ is another strain of M. pulcherrima that added at very early stages of the production process, colonizes and grows rapidly in the medium ensuring prompt and efficient bioprotection of red musts. The mode of action is based on the ability to secrete high concentrations of pulcherriminic acid able of chelating the iron present in the medium which makes the environment unsuitable for the growth of the contaminating microbiota, facilitating the alcoholic fermentation driven by S. cerevisiae sequentially inoculated. A multistarter formulated based on two strains of the species T. delbrueckii and M. pulcherrima are marketed under the name of ZYMAFLORE® ÉGIDETDMP to limit the prevalence of unwanted microorganisms on the surface of the harvesting equipment in contact with the grapes.
2.4. Application of NSYs in biopackaging
In recent years, the need to reduce the production and therefore the disposal of plastic and other non-ecofriendly materials has led the world of research towards the development and application of natural systems as green packaging in agri-food industry [114]. The prefix bio- associated to packaging has a double meaning: biodegradable and/or coming from natural materials. In any case, the objective of food bio packaging is to protect and preserve food products in a sustainable way. Indeed, appropriate packaging has to be capable of maintaining food sensorial and nutritional quality and guaranteeing food safety both in international and national markets, for long time and even in complex conditions [115].
The observation that contaminating microorganisms in food can be controlled during their shelf-life by antimicrobial compounds release from packaging has led to focused efforts to active bio packaging systems that incorporate natural antimicrobial compounds. Castelan and coworkers [116] named this bioactive antimicrobial molecules NACs and reviewed the principal natural sources suitable for this purpose. The NACs are mainly represented by minerals, fruit extracts, essential oils, animal shells, but also some microorganisms or their active molecules. Regarding to this last group, there is a growing interest in the development of antimicrobial packaging materials containing natural antimicrobial agents [117]. The same interest has been driven by consumer concerns about health-related issues, such as the use of synthetic antimicrobial agents. Incorporating synthetic antimicrobial agents directly into foods can effectively inhibit the growth and survival of various microorganisms, but consumers demand minimally processed, preservative-free food products with a longer shelf life. Active antimicrobial bio packaging provides a suitable natural alternative to chemical additives as advanced barrier able to prevent the develop pf food pathogens.
Since 2012, many articles have appeared regarding the use of antimicrobials of bacterial origin [118] such as bacteriocins synthesized by Archaebacteria, Lactococcus, Streptococcus, and Lactobacillus. For example, the incorporation of nisin and pediocin produced by Lactococcus lactis and Pediococcus acidilactici, respectively, into polymer matrices were applied in bio packaging, exploiting their bactericidal and bacteriostatic efficacy against L. monocytogenes [119,120] for mozzarella preservation.
The effectiveness of yeast in preventing food decay has already been established over the last decades, particularly in postharvest treatments of fruits to control mould decay [21,121]. Recently, Zhang et al., [16] reviewed the recent literature about the antagonistic role of yeasts in food industry, highlighting their suitable role among various microbial antagonists. Indeed, yeast and yeast-like fungi as biocontrol tools against pathogens are environmentally friendly, possess greater stress tolerance then bacteria, and can potentially be genetically improved. Moreover, the utilization of yeasts is generally considered safe, and easily acceptable by market, and growing antagonistic yeasts with excellent biocontrol performance have been developed and registered as commercial products.
The incorporation of yeast into the packaging material could impart antimicrobial activity as well as enhancing its nutritional value and serving as the probiotic. For instance, the yeast Meyerozyma guilliermondii is non-pathogenic and demonstrates good antimicrobial activity and it is a rich source of vitamins and proteins [122]. In 2013, Coda and collegues [123] demonstrated the efficacy of M. guilliermondii strain LCF1353 for the effective antifungal activity on long-term storage in wheat bread. Therefore, the direct incorporation of M. guilliermondii or its metabolites to the packaging material could contribute to increasing the shelf life of food. On this, Atta at al., [124] published a study aimed to develop edible and bioactive food packaging films comprising yeast incorporated into bacterial cellulose in conjunction with carboxymethyl cellulose and glycerol to extend the shelf life of packaged food materials. M. guilliermondii strain MT502203.1 and Gluconacetobacter xylinus strain ATCC53582, biofilms were developed ex situ then incorporated into the fibrous cellulose matrix. The findings of this study indicate that the developed biofilm could be used as edible packaging material with high nutritional value and distinctive properties related to the film component, which would provide protection and extended shelf life of foods.
Recently, Guimaraes and colleagues [125] have provided a comprehensive review of the use of edible films and coatings for the incorporation of living microorganisms, aiming at the biopreservation and probiotic capacity of food products. They summarized six benefits and advantages of edible films and coatings containing living microorganisms: physical barrier to protect food, antimicrobial and/or probiotic add value, protection against mechanical damages, increasement of shelf-life and green approach.
The ability of two bio-based films of sodium alginate and locust bean gum to deliver the antimicrobial yeasts W. anomalus cells for the growth control of P. digitatum was investigated [126]. The authors confirmed the efficacy of yeasts incorporation in preserving the postharvest quality of 'Valencia' oranges artificially infected, where the reduction of green mold were more than 73% after 13 days of shelf-life. The edible coating also represents a strategy for conveying probiotic microorganisms and making them readily available in foods which would therefore have an improved nutraceutical value. Tripathi and Giri, [127] catalogued about 500 probiotic food products introduced in the market during the last years mainly yoghurt and other fermented dairy products, such as cheese, ice cream and fermented children formula. Since the criteria for a microbial strain to be used as a probiotic includes the ability of remaining viable at high cell count throughout the manufacture and storage of the product, conveying probiotic strains in non-fermented foods through edible coatings could represent a cutting-edge strategy. The first study of edible coating with yeast dates back to 1994 and concerns the use of C. oleophila in cellulose films to extend the storage of table grapefruits [128]. When C. oleophila was incorporated in methylcellulose or hydroxypropylcellulose the storage time was increased 11 days moreover, C. oleophila was not adversely affected by the incorporation of 0.15% potassium sorbate, the maximum concentration allowed in food products. In the following years, C. guillermondii and Debaryomyces spp. were applied in formulation to extend the shelflife of oranges. Sharma et al. [129] used chitosan films containing Candida utilis for the control of decay in tomatoes caused by Alternaria alternata and Geotrichum candidum, while Fan et al. [130] tested the incorporation of C. laurentii in alginate-based coatings to extend the shelf life of strawberries. These last results showed that the addition of C. laurentii had an antagonistic effect and inhibited the growth of molds as well as maintaining the overall qualities of the strawberries during prolonged storage. Furthermore, sodium alginate films incorporated with C. laurentii did not show any significant effect on color parameters or anthocyanin concentrations in strawberries, but were able to maintain fruit firmness during storage. More recently, Yinzhe and Shaoying [131] investigated the effect of carboxymethylcellulose and alginate-based coatings incorporating brewer yeast on grape preservation highlighting an effective reduced decay compared with uncoated control and an increase of general quality of the grape since, coatings decreased weight loss. Again, Parafati et al., [132] tested the survival and biocontrol ability of W. anomalus, M. pulcherrima, and A. pullulans in coated mandarins, where the incorporation of yeasts reduced the incidence of P. digitatum in mandarins.
Although many positive effects of active food packaging have been proven by research, there are still few applications. Theoretically, regulation has been well stablished in EU, where active packaging are considered as active materials and articles; they are framed in the Regulation 1935/2004/EC (European Commission, 2004) and Regulation 450/2009/EC (European Commission, 2009), however bioackaging materials are still limited in commercial applications if compared with traditional packaging applications reported in the literature. The main principle for the successful packaging of fresh and fresh-cut produce specific, since each produce varies and hence the requirements for packaging and storage vary.
3. NSYs as probiotic yeasts
Probiotics are defined, by an Expert Panel in 2001, as “live microorganisms which when administered in adequate amounts confer a health benefit on the host”, according with the international organizations FAO (Food and Agriculture Organization of the United Nations) and WHO (World Health Organization). Since then, many researchers focused the attention on this topic, going from about 750 papers indexed prior to 2001, to more than 20,000 until early 2019 [133]. However, most of the papers use the general term “probiotics” and only a minor part (< 3 %) use the term probiotic yeasts [134]. Most of the studies conducted so far have been almost exclusively focused on the bacteria population as probiotics and only recently the scenario is opening to yeasts as new probiotics. Effectively, the subject of probiotic and potentially probiotic yeasts has been developing and arising potential for new probiotic products with novel properties, which are not offered by bacteria-based probiotics available on the current market, showing a lot of technological usefull traits (Figure 2).
Initial in vitro screening is necessary to propose possible new probiotic candidates. Mainly, the screening need to include the ability to survive in the host. Indeed generally probiotics are taken orally reaching the gastrointestinal tract, thus it is important to evaluate their resistance to gastrointestinal conditions (the presence of digestive enzymes, gastric and pancreatic juices, bile salts, pH and body temperature of host). To these conditions are added the ability to colonize intestinal mucosal surfaces [correlated to auto- and coaggregation capability and surface hydrophobicity], the interaction with the existent gut microbiota (antimicrobial activity toward pathogenic microorganisms) and the exibition of the antibiotic resistance [135,136]. No less important is also the assessment of the safety of the new candidate against the host: they must not produce toxins, be pathogenic or exhibit hemolytic and DNase activities and gelatinase production [137-139]. Lastly, the evaluation of technological features that include the ability to easily cultivation and biomass production, and resistance to preservation procedures such as lyophilisation, genetic stability and no deterioration, are also important [134].
Actually, the only recognized and commercialized yeast for the human applications, which fully satisfies the presence of probiotic characteristics, is S. cerevisiae var. boulardii [140]. According to the current literature and Index Fungrum, 2021, it is closely related to the S. cerevisiae wine strains [141], therefore currently it has been re-classified as S. cerevisiae var. boulardii, although it exhibits some unique properties. This yeast is commonly used as biotherapeutic in humans, especially for the treatment of gastrointestinal tract disorders [142] and, more recently, as starting-culture to produce functional and probiotic foods/beverages characterized by health-promoting properties [143].
The recent growing interest for functional and probiotic foods and beverages has prompted scientists to focalize the research on the selection of new strains with both technological and innovative traits such as functional and probiotic properties and bioactive compound production [144,145]. Yeasts are widespread in nature and their relatively easy availability and cultivation make them promising probiotic candidates, especially NSYs, still poorly investigated. However, a recent report from Li et al. [146], described the GRAS status of some NSYs, necessary prerequisite for their evaluation as probiotics. Generally, these yeasts are isolated from fermented foods and drinks, vegetables, fruit juices, fruits, dairy products, industrial food waste and grains [147,148]. The isolation of these yeasts from human-related food matrices could represent a way to guarantee their safety.
Un-anthropized natural environments and spontaneous processed foods could represent valid sources for the isolation of new strains with probiotic features. Thirteen out of 180 yeasts isolated from moss on oak, beech tree bark, wine, wineries and grapes, sugar cane juice and papaya leaves revealed the potential probiotic and antimicrobial activity against some pathogenic yeasts. The isolated yeasts were belonging to L. thermotolerans, Metschnikowia ziziphicola, S. cerevisiae and T. delbrueckii species [149]. Findings highlighted their promising probiotic characteristics, although these aptitudes were strictly strain-dependent [150,151]. Also naturally fermented table olives represented a source of potential probiotic yeasts: five NSYs belonging to Candida orthopsilosis, C. tropicalis, D. hansenii, P. guillermondii and Meyerozyma carribica showed resistance to 37 °C, pH 2.0, in presence of bile salts and antimicrobial activity against Salmonella enteritidis [136].
Several studies described similar properties in other different NSYs including H. osmophila, H. guilliermondii, H. uvarum, H. osmopholia, K. thermotolerans, K. marxianus, P. membranifaciens, P. kudriavzevii, Pichia masmurika, Pichia occidentalis, Candida quercitresa, C. intermedia, C. sake, T. delbrueckii, W. anomalus and M. guillermondii [140]. They were unaffected by conditions that mimicking a dynamic gastrointestinal system and, thus, could be proposed for new commercial functional foods [152,153]. In a study conducted to compare functional and biotechnological characteristics of potential probiotic yeast strains belonging to Saccharomyces and NSYs (genera Pichia, Lachancea, Hanseniaspora, Candida, and Zygosaccharomyces) it was reported that in aerobic conditions all NSYs showed higher capacity for adhesion to Caco-2/TC7 intestine-derived cell line and the higher assimilation for nine prebiotics (cellulose, inulin, melibiose, raffinose, xylan, trehalose, pectin, cellobiose and beta-glucans) than Saccharomyces strains. Furthermore, considering other properties, the presence of digestive enzymes, antioxidant activity, antifungal resistance and vitality after sonication treatment two strains of H. osmophila and one of L. thermotolerans were identified as health-promoting probiotics [154]. The evaluation of non-pathogenicity and safe for consumption aspectes excluded the strain H. osmophila 1094 for the high production of biogenic amines, especially tyramine [152].
In 2018 it was firstly described a strain of Cryptococcus (95% of ITS sequence similarity with C. albidus) from the Red Sea with probiotic traits: tolerance to low pH and gastric juice, resistance to bile salts, hydrophobicity, antimicrobial activity and ability to degrade cholesterol [155]. Other NSYs with probiotic properties was the strain of P. pastoris X-33 wild type that showed the ability to survive the stresses of the gastrointestinal tract and caused no lesions when provided to mice thorough the diet exibiting a high antibacterial activity against S. enterica serovar typhimurium[156]. Mice infected by a virulent strain of S. typhimurium showed high survival when supplemented with P. pastoris by gavage or via diet. These results concluded that the yeast P. pastoris X-33 has probiotic properties with considerable antibacterial activity against S. typhimurium. Also, P. kluyveri was described as potential probiotic yeast proposed in co-fermentation with S. cerevisiae strains and Lactobacillus paracasei to obtain a novel, functional fermented maize-based beverage. Similarly, Canonico and co-workers [157] proposed the application of wild non-Saccharomyces potential probiotic yeasts to produce a premium craft beer Kazachstania unispora, isolated from artisanal sourdough, capable of produce a craft beer with low ethanol content and distinctive aromatic notes. Still, P. kudriavzevii DCNa1 and Wickerhamomyces subpelliculosus DFNb6 were proposed to ferment cornelian cherry fruits puree with the aim to obtain a functional beverage characterized by low ethanol content, high amounts of alcohols and esters, and low levels of aldehydes and alkanes. Moreover, these yeasts remained alive in a dose recommended for a probiotic beverage after 21 days of cold storage and after in vitro simulated digestion system. They were able to modulate the intestinal microbiota composition, when they were ingested through this beverage, in an in vitro gastrointestinal simulator [158].
Retracing all the aspects here reported, a general reflection emerges: the long history of yeasts in science, their useful contributions to research and above all to human life, as well as the broad prospects for their use in new fields of application, lead us to assume that they will continue to accompany scientists for many years, contributing to the improvement of human life, as they have done since the beginning. The recently highlighted potential of NSYs, their specific contribution in biotechnological food matrices represent the central themes of this review, where the recently highlighted potential of NSYs, their specific contribution in biotechnological food matrices represent the central themes. In this regard NSYs constitute a great variety of species that have shown effective antagonistic capacity against several spoilage and pathogenic microorganisms, as attested by this review in which references related to more than 70 yeast species involved in the central topic of biocontrol or probiotic trait, were considered (Table 1).
4. Conclusion and perspectives
In the last two decades there was growing interest on NSYs with its improving application in agri-food industry with consequent commercial relevance. In addition to the features regarding quality of food and beverages, as the enhancement of flavor and aroma complexity or ethanol reduction, there are growing interest on NSYs concerning antimicrobial issue. Furthermore, the interest in NSYs, triggered by their rich and diverse reservoir of enzymes and secondary metabolites typically not produced by the conventional S. cerevisiae yeast, as well as their metabolic diversity, is now fundamental to meet consumer demands for novel sensory properties and health benefits. Effectively, NSYs can be used against pathogenic microorganisms in all producing chain step, starting to primary production in field, ongoing in postharvest or during the transformation processes and the packaging and storage phase of food and beverages. But also, enriched food with the addition of NSYs with probiotics traits represents a safe and easy way to bring to market fermented foods and beverages with health attributes as well, without increasing costs. Therefore, the consumer would have the possibility of simultaneously consuming a fermented food supplemented with a source of viable probiotics, bypassing the purchase of pharmaceutical formulations with high costs and dubious vitality. For these reasons, the metabolic diversity provided by NSYs promises to meet the consumer demands for new sensorial and beneficial health properties.
Moreover, the growing interest in ecological, economical and health sustainability of agri-food industry in reducing agrochemical treatments but also conventional synthetic additives and antibiotics or antiseptics during technological food transformation, represent the starting point to explore in this topic. On the other hand, this field of research is still little explored and deserves to be investigated. Little is known about the activities of yeasts and NSYs against very dangerous pathogenic bacteria involved in food processing and preservation. In this way, more in-depth knowledge on the spectrum of action of NSYs towards pathogenic or alterative bacteria together with the study of the mode of action could represent an opportunity to counteract the antibiotic resistance phenomenon, which afflicts not only the medical field, but also food sector. It would therefore be desirable to direct the research towards studies aimed at the interactions between the safe and easily handled NSYs and bacteria, even in complex fermentations involving the interaction of several microbial species, with the aim of clarifying the contribution of the antagonist and the sensitive microorganisms.
Finally integrated studies of applied microbiology and food technology would be desirable to increase the possibilities of supplying formulations in which the microorganisms remain alive for long time and in which the management of the inoculum and fermentation process is as flexible, economical and easy as possible.
Author Contributions
conceptualization, writing, review, and editing, F.C. and M.C.; original draft preparation, L.C. and A.A.; review and editing F.C. and M.C.; all authors have read and agreed to the published version of the manuscript.
Funding
no funds were used to produce this review.
Institutional Review Board Statement
not applicable.
Informed Consent Statement
not applicable.
Data Availability Statement
data is contained within the article.
Conflicts of Interest
the authors declare no conflict of interest.
References
- Johnson, L.; Horsman, S. R.; Charron-Mazenod, L.; Turnbull, A. L.; Mulcahy, H.; Surette, M. G.; Lewenza, S. Extracellular DNA-induced antimicrobial peptide resistance in Salmonella enterica serovar Typhimurium. BMC microbiol 2013, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl Microbiol Biotechnol 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Di Renzo, T.; Sorrentino, A.; Reale, A.; Boscaino, F. Selection of Non-Saccharomyces Wine Yeasts for the Production of Leavened Doughs. Microorganisms 2022, 10, 1849. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ma, X.; Deng, L.; Zhao, X.; Wei, Y.; Gao, Z.; Sun, C. Fresh garlic extract enhances the antimicrobial activities of antibiotics on resistant strains in vitro. Jundishapur J Microbiol 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with winemaking. World J Microbiol Biotechnol 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Manzanares, P.; Ramón, D.; Querol, A. The role of non-Saccharomyces yeasts in industrial winemaking. Int Microbiol 1998, 143–148. [Google Scholar]
- de Ullivarri, M.F.; Mendoza, L.M.; Raya, R.R. Characterization of the killer toxin KTCf20 from Wickerhamomyces anomalus, a potential biocontrol agent against wine spoilage yeasts. Biological control 2018, 223–228. [Google Scholar] [CrossRef]
- Kuchen, B.; Maturano, Y.P.; Mestre, M.V.; Combina, M.; Toro, M.E.; Vazquez, F. Selection of native non-Saccharomyces yeasts with biocontrol activity against spoilage yeasts in order to produce healthy regional wines. Fermentation 2019, 5, 60. [Google Scholar] [CrossRef]
- Velázquez, A.E.; Elghandour, M.M.; Adegbeye, M.J.; Pilego, A.B.; Vallejo, L.H.; Salem, A.Z.; Salazar, M.C. Influence of dietary inclusion with corn and soybean oils, in combination with live yeast culture, on horse fecal methane, carbon dioxide and hydrogen production. JEVS 2019, 74, 42–50. [Google Scholar] [CrossRef]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Suárez-Lepe, J.A. Zygosaccharomyces rouxii: Control strategies and applications in food and winemaking. Fermentation 2018, 4(3), 69. [Google Scholar] [CrossRef]
- Comitini, F.; Ciani, M. Kluyveromyces wickerhamii killer toxin: purification and activity towards Brettanomyces/Dekkera yeasts in grape must. FEMS microbiol lett 2011, 316, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M. , Bizzaro, D.; Comitini, F. Evaluation of damage induced by Kwkt and Pikt zymocins against Brettanomyces/Dekkera spoilage yeast, as compared to sulphur dioxide. J appl microbiol 2016, 121, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; Loira, I.; Morata, A.; Bañuelos, M.A.; Suárez-Lepe, J.A. Wine spoilage yeasts: Control strategy. Yeast-ind appl 2017, 89–116. [Google Scholar]
- Rodriguez-Navarro, C.; González-Muñoz, M.T.; Jimenez-Lopez, C.; Rodriguez-Gallego, M. Bioprotection. In Encyclopedia of Earth Sciences Series; Finkl, C.W., Ed.; Springer: Berlin, Germany, 2011; pp. 185–189. [Google Scholar]
- Rahman, M.A.; Mostofa, M.G. ; Ushimaru, TThe Nem1/Spo7–Pah1/lipin axis is required for autophagy induction after TORC 1 inactivation. The FEBS Journal 2018, 285, 1840–1860. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic yeasts: A promising alternative to chemical fungicides for controlling postharvest decay of fruit. J fungi 2020, 6(3), 158. [Google Scholar] [CrossRef]
- Oro, L.; Zara, S.; Fancellu, F.; Mannazzu, I.; Budroni, M.; Ciani, M.; Comitini, F. Tp BGL2 codes for a Tetrapisispora phaffii killer toxin active against wine spoilage yeasts. FEMS yeast res 2014, 14, 464–471. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int j food microbiol 2018, 265, 18–22. [Google Scholar] [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and characterization of WA18, a new mycocin produced by Wickerhamomyces anomalus active in wine against Brettanomyces bruxellensis spoilage yeasts. Microorganisms 2020, 9, 56. [Google Scholar] [CrossRef]
- Czajkowski, C.; Nowak, A.I.; Błasiak, P.; Ochman, A.; Pietrowicz, S. Experimental study on a large scale pulsating heat pipe operating at high heat loads, different adiabatic lengths and various filling ratios of acetone, ethanol, and water. Applied Therm Eng 2020, 165, 114534. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Pecci, T.; Romanazzi, G.; Ciani, M.; Comitini, F. Biocontrol of non-Saccharomyces yeasts in vineyard against the gray mold disease agent Botrytis cinerea. Microorganisms 2022, 10, 200. [Google Scholar] [CrossRef]
- Di Gianvito, P.; Englezos, V.; Rantsiou, K.; Cocolin, L. Bioprotection strategies in winemaking. Int J Food Microbiol 2022. [Google Scholar] [CrossRef] [PubMed]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Tourdot-Maréchal, R. Bio-protection as an alternative to sulphites: impact on chemical and microbial characteristics of red wines. Frontiers in microbiology 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; Fernández, L.; López, R.; Santamaría, P.; Gutiérrez, A.R. Bioprotective effect of a Torulaspora delbrueckii/Lachancea thermotolerans-mixed inoculum in red winemaking. Fermentation 2022, 8, 337. [Google Scholar] [CrossRef]
- Chacon-Rodriguez, L.; Joseph, C.L.; Nazaris, B.; Coulon, J.; Richardson, S.; Dycus, D.A. Innovative use of non-Saccharomyces in bio-protection: T. delbrueckii and M. pulcherrima applied to a machine harvester. Catalyst: Discovery into Practice 2020, 4, 82–90. [Google Scholar] [CrossRef]
- Windholtz, S.; Dutilh, L.; Lucas, M.; Maupeu, J.; Vallet-Courbin, A.; Farris, L.; Masneuf-Pomarède, I. Population dynamics and yeast diversity in early winemaking stages without sulfites revealed by three complementary approaches. Appl Sci 2021, 11(6), 2494. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Galli, E.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima as biocontrol agent and wine aroma enhancer in combination with a native Saccharomyces cerevisiae. LWT 2023, 181, 114758. [Google Scholar] [CrossRef]
- Valsaraj, P.; Dubash, T.; Prakash, P.Y. Biocontrol of yeast spoilage in selected food and beverages by yeast mycocin. Acta Biologica Indica 2012, 1, 109–112. [Google Scholar]
- Polonelli, L.; Morace, G. Reevaluation of the yeast killer phenomenon. J Clin Microbiol 1986, 24, 866–869. [Google Scholar] [CrossRef]
- Hatoum, R.M.; Labrie, S.; Fliss, I. Identification and partial characterization of antilisterial compounds produced by dairy yeasts. Probiotics and Antimicrobial Proteins 2013, 5, 8–17. [Google Scholar] [CrossRef]
- Wu, W.H.; Hung, W.C.; Lo, K.Y.; Chen, Y.H.; Wan, H.P.; Cheng, K.C. Bioethanol production from taro waste using thermo-tolerant yeast Kluyveromyces marxianus K21. Biores technol 2016, 201, 27–32. [Google Scholar] [CrossRef]
- Ceugniez, A.; Drider, D.; Jacques, P.; Coucheney, F. Yeast diversity in a traditional French cheese “Tomme d'orchies” reveals infrequent and frequent species with associated benefits. Food microbiol 2015, 52, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Alzaeem, I.; Salama, A.R.; Sedik, M.Z. Incidence of Listeria monocytogenes, Staphylococcus aureus and Escherichia coli in fresh white cheese in Gaza city markets. Asian J Agrice Food Sci 2016, 4. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: the importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci Technol 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Barber, M.F.; Elde, N.C. Buried treasure: evolutionary perspectives on microbial iron piracy. Trends Genet 2015, 31, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chi, Z.; Liu, G.; Buzdar, M.A.; Chi, Z.; Gu, Q. Chemical and biological characterization of siderophore produced by themarine-derived Aureobasidium pullulans HN6.2 and its antibacterial activity. Biometals 2009, 22, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, A.; Li, H. Using Candida oleophila as a biocontrol agent to prevent foodborne Escherichia coli O157 EHEC infections. Springerplus 2012, 1, 82. [Google Scholar] [CrossRef]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Freimoser, F.M. Snf2 controls pulcherriminic acid biosynthesis and antifungal activity of the biocontrol yeast Metschnikowia pulcherrima. Mol Microbiol 2019, 112, 317–332. [Google Scholar] [CrossRef]
- 39. Junker, K:; Chailyan, A.; Hesselbart, A.; Forster, J;, Wendland, J. Multi-omics characterization of the necrotrophic mycoparasite Saccharomycopsis schoenii. PLoS Pathog 2019, 15, e1007692. [CrossRef]
- Fiori, S.; Fadda, A.; Giobbe, S.; Berardi, E.; Migheli, Q. Pichia angusta is an effective biocontrol yeast against postharvest decay of apple fruit caused by Botrytis cinerea and Monilia fructicola. FEMS Yeast Res 2008, 8, 961–963. [Google Scholar] [CrossRef]
- Desai, J.V.; Mitchell, A.P.; Andes, D.R. Fungal biofilms, drug resistance, and recurrent infection. Cold Spring Harb Perspect Med 2014. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B. Fungal biofilms and polymicrobial diseases. J Fungi (Basel) 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: threats, challenges, and promising strategies. Front Med (Lausanne) 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: mechanisms and applications. World J Microbiol Biotechnol 2019, 35, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pu, L.; Jingfan, F.; Kai, C.; Chao-an, L.; Yunjiang, C. Phenylethanol promotes adhesion and biofilm formation of the antagonistic yeast Kloeckera apiculata for the control of blue mold on citrus. FEMS Yeast Res 2014, 14, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.; Li, G.; Liu, Y. , Liu; G., Li; M., Zhang, X.; Liu, J. Increase in antioxidant enzyme activity, stress tolerance and biocontrol efficacy of Pichia kudriavzevii with the transition from a yeast-like to biofilm morphology. Biol Cont 2015, 90, 113–119. [Google Scholar] [CrossRef]
- Wachowska, U.; Głowacka, K.; Mikołajczyk, W.; Kucharska, K. Biofilm of Aureobasidium pullulans var. pullulans on winter wheat kernels and its effect on other microorganisms. Microbiology 2016, 85, 523–530. [Google Scholar]
- Klein, M.N. , Kupper, K.C. Biofilm production by Aureobasidium pullulans improves biocontrol against sour rot in citrus. Food Microbiol 2018, 69, 1–10. [Google Scholar] [CrossRef]
- Bevan, E.; Makower, M. The physiological basis of the killer character in yeast. In Genetics today, XIth International Congress of Genetics; 1963; Volume 1, pp. 202–203. [Google Scholar]
- Marquina, D.; Santos, A.; Peinado, J. Biology of killer yeasts. Internat Microbiol 2002, 5, 65–71. [Google Scholar] [CrossRef]
- Liu, G.L.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Li, Y.; Chi, Z.M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit rev biotechnol 2015, 35, 222–234. [Google Scholar] [CrossRef]
- Ciani, M.; Fatichenti, F. Killer toxin of Kluyveromyces phaffii DBVPG 6076 as a biopreservative agent to control apiculate wine yeasts. Appli Environ Microbiol 2001, 67, 3058–3063. [Google Scholar] [CrossRef]
- Mazzucco, M.B.; Ganga, M.A.; Sangorrín, M.P. Production of a novel killer toxin from Saccharomyces eubayanus using agro-industrial waste and its application against wine spoilage yeasts. Antonie van Leeuwenhoek 2019, 112(7), 965–973. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; de Ingeniis, J.M.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microb lett 2004, 238(1), 235–240. [Google Scholar] [CrossRef]
- Comitini, F.; Mannazzu, I.; Ciani, M. Tetrapisispora phaffii killer toxin is a highly specific β-glucanase that disrupts the integrity of the yeast cell wall. Microb Cell Factories 2009, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Carboni, G.; Fancello, F.; Zara, G.; Zara, S.; Ruiu, L.; Marova, I.; Mannazzu, I. Production of a lyophilized ready-to-use yeast killer toxin with possible applications in the wine and food industries. Int j food microbiol 2020, 335, 108883. [Google Scholar] [CrossRef] [PubMed]
- Carboni, G.; Marova, I.; Zara, G.; Zara, S.; Budroni, M.; Mannazzu, I. Evaluation of Recombinant Kpkt Cytotoxicity on HaCaT Cells: Further Steps towards the Biotechnological Exploitation Yeast Killer Toxins. Foods 2021, 10(3), 556. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging technologies to control Brettanomyces spp. in wine: Recent advances and future trends. Trends Food Sci Technol 2020, 99, 88–100. [Google Scholar] [CrossRef]
- De Ingeniis, J.; Raffaelli, N.; Ciani, M.; Mannazzu, I. Pichia anomala DBVPG 3003 secretes a ubiquitin-like protein that has antimicrobial activity. Appl environ microbiol 2009, 75(4), 1129–1134. [Google Scholar] [CrossRef]
- Santos, A.; San Mauro, M.; Bravo, E.; Marquina, D. PMKT2, a new killer toxin from Pichia membranifaciens, and its promising biotechnological properties for control of the spoilage yeast Brettanomyces bruxellensis. Microbiology 2009, 155, 624–634. [Google Scholar] [CrossRef]
- Santos, E.O.; Michelon, M.; Gallas, J.A.; Kalil, S.J.; André Veiga Burkert, C. Raw glycerol as substrate for the production of yeast biomass. Int J Food Eng 2013, 9(4), 413–420. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int j food microbiol 2014, 188, 83–91. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Prior, K.J.; Setati, M.E.; Divol, B. Candida pyralidae killer toxin disrupts the cell wall of Brettanomyces bruxellensis in red grape juice. J appl microbiol 2017, 122(3), 747–758. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast interactions and molecular mechanisms in wine fermentation: a comprehensive review. Int J Mol Sci 2021, 22, 7754. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Magliani, W.; Ciociola, T.; Giovati, L.; Conti, S. From Pichia anomala killer toxin through killer antibodies to killer peptides for a comprehensive anti-infective strategy. Antonie Van Leeuwenhoek 2011, 99, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Ciani, M.; Esin, S.; Agnolucci, M.; Marcheggiani, F.; Tiano, L.; Comitini, F. Comparative Zymocidial Effect of Three Different Killer Toxins against Brettanomyces bruxellensis spoilage yeasts. Int J Mol Sci 2023, 24, 1309. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; López-Piñeiro, A.; Ribas, J. C. A new wine Torulaspora delbrueckii killer strain with broad antifungal activity and its toxin-encoding double-stranded RNA virus. Front microbiol 2015, 6, 983. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, A.M.; Amézquita, A.; Cardona Jaramillo, J.E.C.; Matiz-Cerón, L.F.; Andrade-Martínez, J.S.; Triana, S.; Cock, H.D. Analysis of Malassezia lipidome disclosed differences among the species and reveals presence of unusual yeast lipids. Front cell inf microbiol 2020, 10, 338. [Google Scholar]
- Peña, R.; Vílches, J.; Poblete, C.; Ganga, M.A. Effect of Candida intermedia LAMAP1790 antimicrobial peptides against wine-spoilage yeasts Brettanomyces bruxellensis and Pichia guilliermondii. Fermentation 2020, 6, 65. [Google Scholar] [CrossRef]
- Acuña-Fontecilla, A.; Silva-Moreno, E.; Ganga, M.A.; Godoy, L. Evaluation of antimicrobial activity from native wine yeast against food industry pathogenic microorganisms. CyTA-J Food 2017, 15, 457–465. [Google Scholar] [CrossRef]
- Younis, G.; Awad, A.; Dawod, R.E.; Yousef, N.E. Antimicrobial activity of yeasts against some pathogenic bacteria. Vet world 2017, 10, 979. [Google Scholar] [CrossRef]
- Mencher, A.; Morales, P.; Valero, E.; Tronchoni, J.; Patil, K.R.; Gonzalez, R. Proteomic characterization of extracellular vesicles produced by several wine yeast species. Microb biotechnol 2020, 13(5), 1581–1596. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Gostincar, C.; Cernosa, A.; Gunde-Cimerman, N. ; Stress tolerant yeasts: opportunistic pathogenicity versus biocontrol potential. Genes (Basel) 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Roach, W.; Sun, T.; Jain, T.; Prinz, B.; Yu, T.Y.; Krauland, E. Addressing polyspecificity of antibodies selected from an in vitro yeast presentation system: a FACS-based, high-throughput selection and analytical tool. PEDS 2013, 26(10), 663–670. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shimon, M.; Yehuda, H.; Cohen, L.; Weiss, B.; Kobeshnikov, A.; Daus, A.; Droby, S. Characterization of extracellular lytic enzymes produced by the yeast biocontrol agent Candida oleophila. Curr Genet 2004, 45, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Castoria, R.; De Curtis, F.; Lima, G.; De Cicco, V. β-1, 3-glucanase activity of two saprophytic yeasts and possible mode of action as biocontrol agents against postharvest diseases. Postharvest Biol Technol 1997, 12(3), 293–300. [Google Scholar] [CrossRef]
- Park, H.W.; Choi, K.D.; Shin, I.S. Antimicrobial activity of isothiocyanates (ITCs) extracted from horseradish (Armoracia rusticana) root against oral microorganisms. Biocontrol sci 2013, 18(3), 163–168. [Google Scholar] [CrossRef]
- Banani, H.; Spadaro, D.; Zhang, D.; Matic, S.; Garibaldi, A.; Gullino, M.L. Postharvest application of a novel chitinase cloned from Metschnikowia fructicola and overexpressed in Pichia pastoris to control brown rot of peaches. Int J Food Microbiol 2015, 199, 54–61. [Google Scholar] [CrossRef]
- Wisniewski, M.; Biles, C.; Droby, S.R.; Wilson, C.; Chalutz, E. Mode of action of the postharvest biocontrol yeast, Pichia guilliermondii. Characterization of attachment to Botrytis cinerea. Physiol Mol Plant 1991, 39, 245–258. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Performance evaluation of volatile organic compounds by antagonistic yeasts immobilized on hydrogel spheres against gray, green and blue postharvest decays. Food Microbiol 2017, 63, 191–198. [Google Scholar] [CrossRef]
- Lemos, W.J.; Treu, L.; da Silva Duarte, V.; Carlot, M.; Nadai, C.; Campanaro, S.; Giacomini, A.; Corich, V. Biocontrol ability and action mechanism of Starmerella bacillaris (synonym Candida zemplinina) isolated from wine musts against gray mold disease agent Botrytis cinerea on grape and their effects on alcoholic fermentation. Front Microbiol 2016, 7, 1249. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biological Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol 2015, 47, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.I.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int J Food Microbiol 2018, 284, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int J Food Microbiol 2014, 189, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.S.; Beck, J.J.; Sarreal, S.B.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res 2014, 30, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile organic compounds produced by Antarctic strains of Candida sake play a role in the control of postharvest pathogens of apples. Biol Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Masoud, W.; Poll, L.; Jakobsen, M. Influence of volatile compounds produced by yeasts predominant during processing of Coffea arabica in East Africa on growth and ochratoxin A (OTA) production by Aspergillus ochraceus. Yeast 2005, 22, 1133–1142. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Hernández, A.; Galvan, A.I.; Córdoba, M.G.; Casquete, R.; Serradilla, M.J.; Martín, A. Selection and application of antifungal VOCs-producing yeasts as biocontrol agents of grey mould in fruits. Food microbiol 2020, 92, 103556. [Google Scholar] [CrossRef]
- Bosqueiro, A.S. ; Bizarria, Jr R.; Rosa-Magri, M.M. Biocontrol of post-harvest tomato rot caused by Alternaria arborescens using Torulaspora indica. Biocontrol Sci Technol 2023, 1–18. [Google Scholar]
- Nasanit, R.; Jaibangyang, S.; Onwibunsiri, T.; Khunnamwong, P. Screening of Volatile Organic Compound-Producing Yeasts and Yeast-Like Fungi against Aflatoxigenic Aspergillus flavus 2022.
- Zou, X.; Wei, Y.; Jiang, S.; Cao, Z.; Xu, F.; Wang, H.; Zhan, P.; Shao, X. Volatile organic compounds and rapid proliferation of Candida pseudolambica W16 are modes of action against gray mold in peach fruit. Postharvest Biol Technol 2022, 183, 111751. [Google Scholar] [CrossRef]
- Liu, J.; Li, G.; Sui, Y. Optimization of culture medium enhances viable biomass production and biocontrol efficacy of the antagonistic yeast, Candida diversa. Front Microbiol 2017, 8, 2021. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Guo, J.H.; Liu, P.; Cheng, Y.J.; Wang, B.Q.; Long, C.A.; Deng, B.X. Inhibitory activity of tea polyphenol and Candida ernobii against Diplodia natalensis infections. J Appl Microbiol 2010, 108, 1066–1072. [Google Scholar] [CrossRef] [PubMed]
- Papon, N.; Savini, V.; Lanoue, A.; Simkin, A.J.; Crèche, J.; Giglioli-Guivarc’h, N.; Clastre, M.; Courdavault, V.; Sibirny, A.A. Candida guilliermondii: biotechnological applications, perspectives for biological control, emerging clinical importance and recent advances in genetics. Curr Genet 2013, 59, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Carbo, A.; Torres, R.; Teixido, N.; Usall, J.; Medina, A.; Magan, N. Impact of climate change environmental conditions on the resilience of different formulations of the biocontrol agent Candida sake CPA-1 on grapes. Lett Appl Microbiol 2018, 67, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of postharvest Botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia. Phytopathol 2011, 101, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Wilson, C.; Droby, S.; Chalutz, E.; El Ghaouth, A.; Stevens, C. Postharvest biocontrol: new concepts and applications. In Biological control: A global perspective; CABI: Wallingford, UK, 2007; pp. 262–273. [Google Scholar]
- Scherm, B.; Ortu, G.; Muzzu, A.; Budroni, M.; Arras, G.; Migheli, Q. Biocontrol activity of antagonistic yeasts against Penicillium expansum on apple. J Plant Pathol 2003, 85, 205–213. [Google Scholar]
- European Food Safety Authority (EFSA) Conclusion on the peer review of the pesticide risk assessment of the active substance Candida oleophila strain O. EFSA J 2015, 10, 2944.
- European Commission Health & Consumers Directorate-General (2013) Review report for the active substance Candida oleophila strain O. vol SANCO/10395/2013 rev 1.
- Kunz, S.; Schmitt, A.; Haug, P. Field testing of strategies for fire blight control in organic fruit growing. Acta Hortic 2011, 896, 431–436. [Google Scholar] [CrossRef]
- Weiss, A.; Mögel, G. Kunz S Development of “Boni-Protect”- a yeast preparation for use in the control of post-harvest diseases of apples. In: Boos M (ed) 12th International conference on cultivation technique and phytopathologtical problems in organic fruit-growing. Weinsberg, Germany, 2006; pp 113–117.
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition assays and physiological experiments of soil and phyllosphere yeasts identify Candida subhashii as a novel antagonist of filamentous fungi. BMC Microbiol 2017, 17, 4. [Google Scholar] [CrossRef]
- Turkel, S.; Korukluoglu, M.; Yavuz, M. Biocontrol activity of the local strain of Metschnikowia pulcherrima on different postharvest pathogens. Biotechnol Res Int 2014. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S. Biopreservation of food and feed by postharvest biocontrol with microorganisms. In: Sundh I, Wilcks A, Goettel MS (eds) Beneficial microorganisms in agriculture, food and the environment. CABI International, Oxfordshire, 2012; pp. 1234-1254.
- Droby, S.; El-Gerberia, B. Yeast Metschnikowia fructicola NRRL Y-30752 for inhibiting deleterious microorganisms on plants. USA Patent 7 February 2006. [Google Scholar]
- Tian, S.P.; Qin, G.Z.; Xu, Y.; Wang, Y.S. Application of antagonistic yeasts under field conditions and their biocontrol ability against postharvest diseases of sweet cherry. Acta Bot Sin 2004, 46, 1324–1330. [Google Scholar]
- Zhang, H.Y.; Zheng, X.D.; Wang, L.; Li, S.S; Liu, R.F. Effect of yeast antagonist in combination with hot water dips on postharvest Rhizopus rot of strawberries. J Food Eng 2007, 78, 281–287. [Google Scholar] [CrossRef]
- Zhang, H.Y. , Zheng, X.D., Yu, T. Biological control of postharvest diseases of peach with Cryptococcus laurentii. Food Control 2007, 18, 287–291. [Google Scholar] [CrossRef]
- Kowalska, J.; Drożdżyński, D.; Remlein-Starosta, D.; Sas-Paszt, L.; Malusá, E. Use of Cryptococcus albidus for controlling grey mould in the production and storage of organically grown strawberries. J Plant Dis Protect 2012, 119, 174–178. [Google Scholar] [CrossRef]
- Lutz, M.C.; Lopes, C.A.; Rodriguez, M.E.; Sosa, M.C.; Sangorrin, M.P. Efficacy and putative mode of action of native and commercial antagonistic yeasts against postharvest pathogens of pear. Int J Food Microbiol 2013, 164, 166–172. [Google Scholar] [CrossRef]
- Silvestri, D.; Wacławek, S.; Venkateshaiah, A.; Krawczyk, K.; Sobel, B.; Padil, V.V.; Varma, R.S. Synthesis of Ag nanoparticles by a chitosan-poly (3-hydroxybutyrate) polymer conjugate and their superb catalytic activity. Carbohydr polym 2020, 232, 115806. [Google Scholar] [CrossRef]
- Di Matteo, G.; Di Matteo, P.; Sambucci, M.; Tirillò, J.; Giusti, A.M.; Vinci, G.; Valente, M. Commercial bio-packaging to preserve the quality and extend the shelf-life of vegetables: The case-study of pumpkin samples studied by a multimethodological approach. Foods 2012, 10, 2440. [Google Scholar] [CrossRef]
- Castelán, E.; Rodríguez-Hernández, A.I.; Chavarría-Hernández, N.; López-Ortega, M.A.; del Rocio López-Cuellar, M. Natural antimicrobial systems protected by complex polyhydroxyalkanoate matrices for food biopackaging applications—A review. Int J Bioll Macromol 2023, 123418. [Google Scholar]
- Maisanaba, S.; Llana-Ruiz-Cabello, M.; Gutiérrez-Praena, D.; Pichardo, S.; Puerto, M.; Prieto, A.I.; Jos, A.; Cameán, A.M. New advances in active packaging incorporated with essential oils or their main components for food preservation. Food Rev Int 2017, 33, 447–515. [Google Scholar] [CrossRef]
- Gálvez, A.; Abriouel, H.; López, R.L.; Omar, N.B. Bacteriocin-based strategies for food biopreservation. Int j food microbiol 2007, 120, 51–70. [Google Scholar] [CrossRef]
- Espitia, P.J.P.; Otoni, C.G.; Soares, N.F.F. Pediocin applications in antimicrobial food packaging systems. In Antimicrob food pack, pp. 445-454. Academic Press (2016).
- dos Santos Pires, A.C.; de Fátima Ferreira Soares, N.; de Andrade, N.J.; da Silva, L.H.M.; Camilloto, G.P.; Bernardes, P.C. Development and evaluation of active packaging for sliced mozzarella preservation. Packag Technol Sci 2008, 21, 375–383. [Google Scholar] [CrossRef]
- Hammami, R.; Oueslati, M.; Smiri, M.; Nefzi, S.; Ruissi, M.; Comitini, F. , Sadfi Zouaoui, N. Epiphytic yeasts and bacteria as candidate biocontrol agents of green and blue molds of citrus fruits. J Fungi 2022, 8, 818. [Google Scholar] [CrossRef] [PubMed]
- Bertini, E.V.; Leguina, A.C.D.V.; Castellanos, L.I.; Nieto Peñalver, C.G. Endophytic microorganisms Agrobacterium tumefaciens 6N2 and Meyerozyma guilliermondii 6N serve as models for the study of microbial interactions in colony biofilms. 2019, 2, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Rizzello, C.G.; Di Cagno, R.; Trani, A.; Cardinali, G.; Gobbetti, M. Antifungal activity of Meyerozyma guilliermondii: identification of active compounds synthesized during dough fermentation and their effect on long-term storage of wheat bread. Food microbiol 2013, 33(2), 243–251. [Google Scholar] [CrossRef]
- Atta, O.M.; Manan, S.; Ul-Islam, M.; Ahmed, A.A.Q.; Ullah, M.W.; Yang, G. Silver decorated bacterial cellulose nanocomposites as antimicrobial food packaging materials. ES Food Agroforestry 2021, 6, 12–26. [Google Scholar] [CrossRef]
- da Costa Guimarães, N.; Freitas-de-Sousa, L.A.; de Souza, M.C.S.; de Almeida, P.D.O.; Dos-Santos, M.C.; Nunez, C.V.; de Moura, V.M. Evaluation of the anti-snakebite, antimicrobial and antioxidant potential of Philodendron megalophyllum Schott (Araceae), traditionally used in accidents caused by snakes in the western region of Pará, Brazil. Toxicon 2020, 184, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Aloui, H.; Licciardello, F.; Khwaldia, K.; Hamdi, M.; Restuccia, C. Physical properties and antifungal activity of bioactive films containing Wickerhamomyces anomalus killer yeast and their application for preservation of oranges and control of postharvest green mold caused by Penicillium digitatum. Int J Food Microbiol 2015, 200, 22–30. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J funct foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- McGuire, R.G.; Baldwin, E.A. Compositions of cellulose coatings affect populations of yeasts in the liquid formulation and on coated grapefruits. In PROCEEDINGS-FLORIDA STATE HORTICULTURAL SOCIETY (Vol. 107, pp. 293–293). FLORIDA STATE HORTICULTURAL SOCIETY,1994.
- Sharma, N.; Brandis, K.A.; Herrera, S.K.; Johnson, B.E.; Vaidya, T.; Shrestha, R.; DebBurman, S.K. α-Synuclein budding yeast model: toxicity enhanced by impaired proteasome and oxidative stress. JMB 2006, 28, 161–178. [Google Scholar] [CrossRef]
- Fan, X.; Sokorai, K.J.; Liao, C.H.; Cooke, P.; Zhang, H.Q. Antibrowning and antimicrobial properties of sodium acid sulfate in apple slices. J food sci 2009, 74, M485–M492. [Google Scholar] [CrossRef]
- Yinzhe, R.; Shaoying, Z. Effect of carboxymethyl cellulose and alginate coating combined with brewer yeast on postharvest grape preservation. Int Sch Res Notices 2013. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. The effect of locust bean gum (LBG)-based edible coatings carrying biocontrol yeasts against Penicillium digitatum and Penicillium italicum causal agents of postharvest decay of mandarin fruit. Food microbiol 2016, 58, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: reiterating what they are and what they are not. Front microbiol 2019, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Staniszewski, A.; Kordowska-Wiater, M. Probiotic and potentially probiotic yeasts—characteristics and food application. Foods 2021, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V. From yaks to yogurt: The history, development, and current use of probiotics. Clin Infect Dis 2015, 60, S85–S90. [Google Scholar] [CrossRef] [PubMed]
- Simões, L.A.; Cristina de Souza, A.; Ferreira, I.; Melo, D.S.; Lopes, L.A.A.; Magnani, M.; Dias, D.R. Probiotic properties of yeasts isolated from Brazilian fermented table olives. J Appl Microbiol 2021, 131, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Da, R.; Tuo, X. Probiotic and safety properties screening of Enterococcus faecalis from healthy Chinese infants. Probiotics Antimicrob Proteins 2020, 12, 1115–1125. [Google Scholar] [CrossRef]
- Pereira, R.P.; Jadhav, R.; Baghela, A.; Barretto, D.A. In Vitro Assessment of probiotic potential of Saccharomyces cerevisiae DABRP5 isolated from bollo batter, a traditional Goan fermented food. Probiotics Antimicrob Prot 2021, 13, 796–808. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food: Report of a Joint FAO/WHO. Working group on drafting guidelines for the evaluation of probiotics in food, 2002.
- Shruthi, B.; Deepa, N.; Somashekaraiah, R.; Adithi, G.; Divyashree, S.; Sreenivasa, M.Y. Exploring biotechnological and functional characteristics of probiotic yeasts: A review. Biotechnol Rep 2022, e00716. [Google Scholar] [CrossRef]
- Khatri, I.; Tomar, R.; Ganesan, K.; Prasad, G.S.; Subramanian, S. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: what makes it tick as successful probiotic? J Fungus 2020, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lazo-Vélez, M.A.; Serna-Saldívar, S.O.; Rosales-Medina, M.F.; Tinoco-Alvear, M.; Briones-García, M. Application of Saccharomyces cerevisiae var. boulardii in food processing: a review. J Appl Microbiol 2018, 125, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, D.M.; Leem, C. The bioactive compounds of probiotic foods/ supplements and their application in managing mental disorders. BCHD 2019, 2, 206–220. [Google Scholar] [CrossRef]
- Rai, A.K. Pandey, A.; Sahoo, D. Biotechnological potential of yeasts in functional food industry. Trends Food Sci Technol 2019, 83, 129–137. [Google Scholar] [CrossRef]
- Li, Y.; Sadiq, F.A.; Liu, T.; Chen, J.; He, G. Purification and identification of novel peptides with inhibitory effect against angiotensin I-converting enzyme and optimization of process conditions in milk fermented with the yeast Kluyveromyces marxianus. J Funct Foods 2015, 16, 278–288. [Google Scholar] [CrossRef]
- Agarbati, A.; Marini, E.; Galli, E.; Canonico, L.; Ciani, M.; Comitini, F. Characterization of wild yeasts isolated from artisan dairies in the Marche region, Italy, for selection of promising functional starters. LWT 2021, 139, 110531. [Google Scholar] [CrossRef]
- Fernández-Pacheco, P.; García-Bejar, B.; Jimenez-del Castillo, M.; Carreno- Domínguez, J.; Briones Perez, A.; Arevalo-Villena, M. Potential probiotic and food protection role of wild yeasts isolated from pistachio fruits (Pistacia vera). J Sci Food Agric 2021, 101, 2201–2209. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Marini, E.; Zannini, E.; Ciani, M.; Comitini, F. Potential probiotic yeasts sourced from natural environmental and spontaneous processed foods. Foods 2020, 9–287. [Google Scholar] [CrossRef]
- Fadda, M.E.V.; Mossa, M.; Deplano, M.B.; Pisano, S.; Cosentino, D. In vitro screening of Kluyveromyces strains isolated from Fiore Sardo cheese for potential use as probiotics. LWT 2017, 75, 100–106. [Google Scholar] [CrossRef]
- Alvarez, S.C.V.; Alaniz, M.J.L.; Furlani, M.V.M.; Vazquez, F.; Agresti, P.M.; Nally, M.C.; Maturano, Y.P. Bioprospecting of the probiotic potential of yeasts isolated from a wine environment. Fungal Genet Biol 1037, 164, 103767. [Google Scholar] [CrossRef]
- Fernández-Pacheco, C.; Cueva, M.; Arevalo-Villena, M.V.; Moreno-Arribas, A.; Briones Perez, G. Saccharomyces cerevisiae and Hanseniaspora osmophila strains as yeast active cultures for potential probiotic applications. Food Funct 2019, 4924–4931. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, F.C.; Sooklim, P.; Songdech, O.; Duangpakdee, N.; Soontorngun, O. Selection of potential yeast probiotics and a cell factory for xylitol or acid production from honeybee samples. Metabolites 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pacheco, P.; Pintado, C.; Briones Pérez, A.; Arévalo-Villena, M. Potential probiotic strains of Saccharomyces and non-Saccharomyces: functional and biotechnological characteristics. J Fungus 2021, 7, 177. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, A.F.; El-Enshasy, H.A.; Shetaia, Y.M.; Mahrous, H.; Othman, N.Z.; Yousef, A.E. Semi-industrial scale production of a new yeast with probiotic traits, Cryptococcus sp. YMHS, isolated from the Red Sea. Probiotics antimicrob 2018, 10, 77–88. [Google Scholar] [CrossRef]
- França, R.C.; Conceição, F.R.; Mendonça, M.; Haubert, L.; Sabadin, G.; de Oliveira, P.D.; Moreira, Â.N. Pichia pastoris X-33 has probiotic properties with remarkable antibacterial activity against Salmonella typhimurium. Appl microbiol biotechnol 2015, 99, 7953–7961. [Google Scholar] [CrossRef]
- Canonico, L.; Zannini, E.; Ciani, M.; Comitini, F. Assessment of non-conventional yeasts with potential probiotic for protein-fortified craft beer production. LWT 2021, 145, 111361. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Cantatore, V.; Polo, A.; Celano, G.; Martinovic, A.; Gobbetti, M. Design of potential probiotic yeast starters tailored for making a cornelian cherry (Cornus mas L.) functional beverage. Int j food microbiol 2020, 323, 108591. [Google Scholar] [CrossRef]
Figure 2.
Graphical representation of the main probiotic technological characteristics.
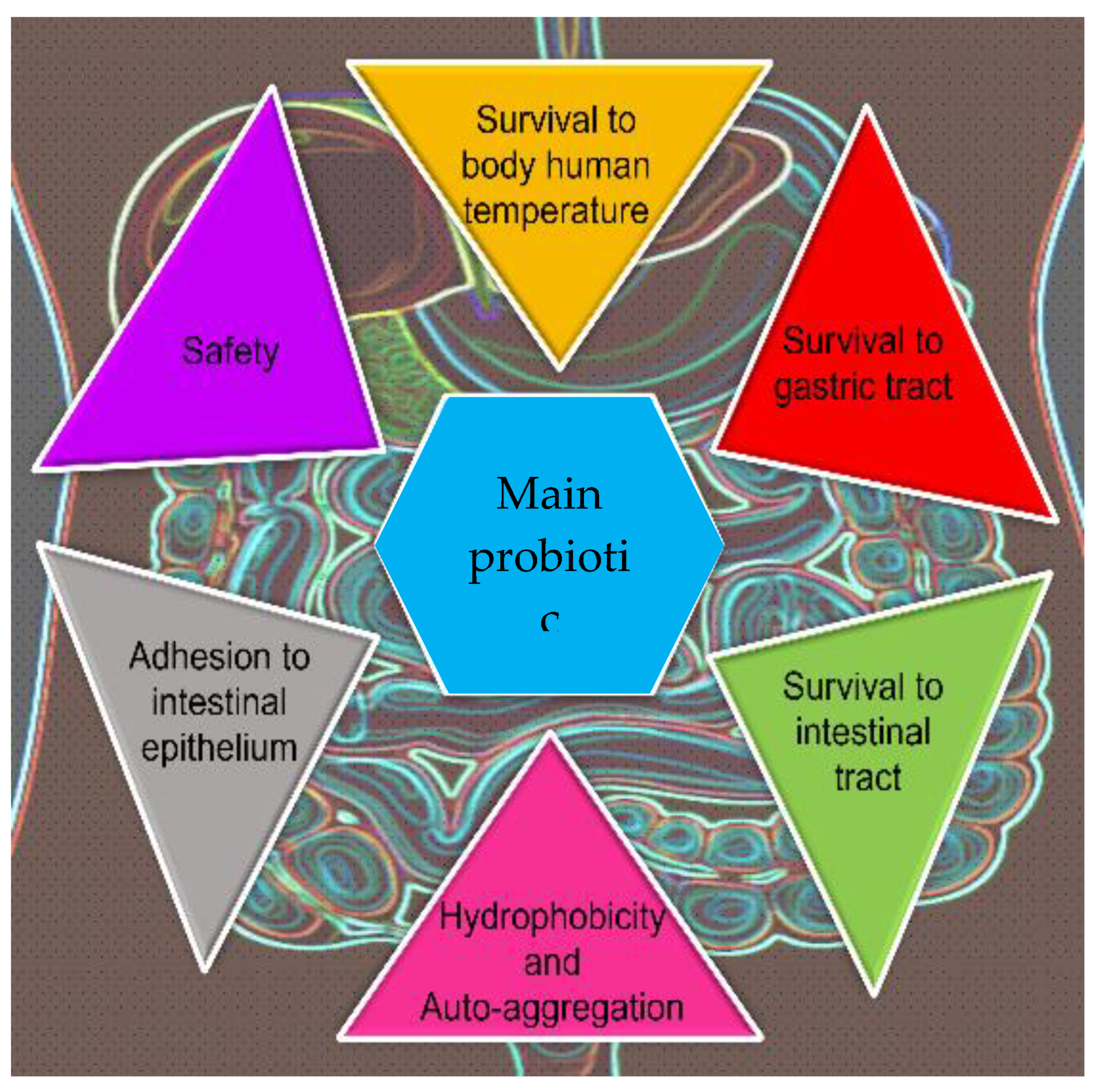

Table 1.
List of all NSYs reported in the review and relative matrices, specific activity, and reference.
Table 1.
List of all NSYs reported in the review and relative matrices, specific activity, and reference.
| NSYs | Products | Distinctive antimicrobial features |
References |
|---|---|---|---|
| Aureobasidium pullulans | Guava fruit | Aantibacterial activity mediated by fusarinin C | [37] |
| Aureobasidium pullulans | Citrus | Antimicrobial biofilm formation | [48] |
| Aureobasidium pullulans | Plants | biocontrol mediated by nutrient, space or VOC | [18,52,54-56] |
| Aureobasidium pullulans | Mandarins edible coating | Anti-mould activity | [80] |
| Aureobasidium pullulans | Table grape berries | Biocontrol VOC mediated | [80] |
| Aureobasidium pullulans | Fruits | Biocontrol VOC mediated against A. flavus | [86] |
| Candida spp. | Top, hazel and citrus fruits | Antagonist of filamentous fungi | [104] |
| Candida albicans | Medical field | Inhibitory effect on S. aureus and E. coli | [33] |
| Candida guillermondii | Oranges biopackaging | Anti-mould activity | [128] |
| Candida inetrmedia | Milk | Probiotic activity | [139] |
| Candida intermedia | Wine environment | Anti spoilage yeasts (wide spectrum) AMPs mediated | [69] |
| Candida intermedia | Medical field | Antimicrobial against E. coli, S.aureus AMPs mediated | [71, 70] |
| Candida laurentii | Strawberries edible coating | Antimould activity | [129] |
| Candida oleophyla | Fruits | Antimould activity (P. digitatum) EVs mediated | [75] |
| Candida oleophyla | Table grapefuits biopackaging | Ant-imould activity | [127] |
| Candida orthopsilosis | Table olives | Probiotic activity | [135] |
| Candida pseudolambica | Peach | Biocontrol VOC mediated against B. cinerea | [92] |
| Candida pyralidale | Wine | Anti-Brettanomyces activity mycocin mediated | [63] |
| Candida quercitresa | Indigenous fermented food | Probiotic activity | [139] |
| Candida sake | apple rot | Biocontrol VOC mediated against P.expansum and B.cinerea | [87] |
| Candida sake | Milk products | Probiotic activity | [139] |
| Candida tropicalis | Medical field | Bacterial lysis | [30] |
| Candida tropicalis | Table olives | Probiotic activity | [135] |
| Candida utilis | Tomatoes biopackaging | Anti-mould activity | [128] |
| Debaryomyces hansenii | Medical field | Bacterial lysis | [30] |
| Debaryomyces hansenii | Table olives | Probiotic activity | [135] |
| Hanseniaspora guillermondii | N.D.1 | Probiotic activity | [139] |
| Hanseniaspora uvarum | Coffee | Biocontrol VOC mediated against A. ochraceus | [88] |
| Hanseniaspora uvarum | N.D.1 | Probiotic activity | [139] |
| Kluyveromyces lactis | Medical field | L. monocytogenes and C. albicans growth inhibition | [32] |
| Kluyveromyces marxianus | Medical field | E. coli diseases prevention in mice | [31] |
| Kluyveromyces marxianus | Medical field | L. monocytogenes and C. albicans growth inhibition | [32] |
1 ND: Not Determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



