
Submitted:
02 May 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
Fibre-based dietary interventions are at the forefront of gut microbiome modulation research, with a wealth of 16S rRNA information to demonstrate the prebiotic effects of isolated fibres. However, there is a distinct lack of data relating to the effect of a combination of soluble and insoluble fibres in a convenient to consume fruit juice food matrix on gut microbiota structure, diversity, and function. Here, we aimed to determine the impact of the MOJU Prebiotic Shot, a fruit juice drink blend containing chicory inulin, baobab, golden kiwi, and green banana powders, on gut microbiota structure and function. Healthy adults (n = 20) were included in a randomized, double-blind, placebo-controlled, cross-over study receiving 60ml MOJU Prebiotic Shot or pla-cebo (without the fibre mix) for 3 weeks with a 3-week wash-out between interventions. Shotgun metagenomics revealed significant between-group differences in alpha and beta diversity. In addition, the relative abundance of the phyla Actinobacteria and Desulfobacteria was significantly increased as a result of the prebiotic intervention. 9 species were observed to be differentially abundant (uncorrected p-value of <0.05) as a result of the prebiotic treatment. Of these, Bifidobacterium adolescentis and CAG-81 sp900066785 (Lachnospiraceae) were present at increased abundance relative to baseline. Additionally, KEGG analysis showed an increased abundance in pathways associated with arginine biosynthesis and phenylacetate degradation during the prebiotic treatment. Our results show the effects of the daily consumption of 60ml MOJU Prebiotic Shot for 3 weeks and provide insight to the functional potential of B. adolescentis
Keywords:
shotgun metagenomics
; gut microbiota
; prebiotics
; Bifidobacterium adolescentis
; Lachnospiraceae
; arginine
; fibre
1. Introduction
Research involving interventions designed to modulate the gut microbiota with the aim to improve host health has contributed to accruing evidence of a bidirectional gut-microbiota-host-axis. Driven by technological advances and reductions in the costs associated with analyses, important advances have been made in identifying microbial communities and their influence on host health. We are now able to ask, “who is there” and “what can they do” with the aid of shotgun metagenomics that provides more detailed microbiome insights (Wang WL et al., 2015; Klimenco et al., 2022).
The collection of microorganisms inhabiting the mammalian gastrointestinal tract is now considered to have a central position in health and disease, with the ability to affect organs far beyond the abdomen (Qi et al., 2021; Novakovic et al., 2020; Maiuolo et al., 2021). This complex community that includes bacteria, archaea, viruses, fungi, and protozoa is fundamental to maintaining host physiological homeostasis (Nikolaieva et al., 2023), including programming host circadian rhythms (Mukherji et al., 2013; Butler and Gibbs, 2020; Murakami et al., 2016). Many of these gut microbes participate in several beneficial-to-host functions, including essential metabolic outputs such as short-chain fatty acids (SCFAs) (Silva et al., 2020) and neurotransmitters (Chen, Y et al., 2021).
Diet is widely agreed to be a key modulator of the gut microbiota. This is particularly true for dietary fibre, broadly defined as those carbohydrate polymers and oligomers (plus lignin) that escape digestion in the small intestine, passing into the large bowel where they are partially (insoluble dietary fibre) or more completely (soluble dietary fibre) fermented and metabolised by the gut microbiota (Wang et al., 2019). Reaching the large intestine, where the highest concentration of gut microbiota is located at an abundance of around 1011 bacterial cells per mL of content (Sender et al., 2016), dietary fibres are fermented into products that can provide a variety of localized and systemic health benefits to the host (Barber et al., 2020). Such is the interest in the impact of dietary fibre on human health, ongoing work supported by the Technical Committee on Dietary Carbohydrates of the North American Branch of the International Life Sciences Institute (ILSI North America) and Tufts University, has resulted in a publicly available research database linking fibres to a variety of health outcomes (Livingston et al., 2016). Recognized health benefits associated with dietary fibre consumption, and which have been extensively reviewed (Carlson et al., 2018; Myhrstad et al., 2020), include improved gastrointestinal function (Rao et al., 2015), moderation of circulating blood lipids and improvement of post-prandial serum glucose and insulin responses (Threapleton et al., 2013; Russel et al., 2016; Liu, F et al., 2017; Weickert et al., 2011). Furthermore, chronic low intake of dietary fibre has been associated with many deleterious health consequences including increased risk of colorectal cancer (Kim, Y et al., 2020; Khorasaniha et al., 2023), diverticular disease (Eberhardt et al., 2019), cardiovascular disease (Srour et al., 2019), obesity (Weickert et al., 2006), metabolic syndrome (Kabisch et al., 2019), pre-diabetes and type 2 diabetes (Honsek et al., 2018; Khorasaniha et al., 2023).
The definition of dietary fibres differs between countries as detailed elsewhere (Stephen et al., 2017; Swanson et al., 2019; Barber et al., 2020). Fibre solubility (dissolution capability in water), viscosity (gelling capability in water), and fermentability (degree of metabolizing capability by gut microbiota) are among the factors that determine the effect of fibre on the gut microbiota and host (Cronin et al., 2021). Despite decades of lifestyle and dietary advice and education on the importance of adequate fibre in the diet, our diets continue to lack sufficient fibre (Barber et al., 2020; Neyrinck et al., 2020; Stephen et al., 2017), even in Mediterranean countries (CEEREAL, 2022). Latest data for the UK and US for adults 19 years and over (gender combined) suggests consumption of 19.7 and 16.6g/day, respectively (NDNS, 2019; USDA ARS, 2021). Dietary fibre intakes around the world are described in detail elsewhere (Ioniță-Mîndrican et al., 2022; Barber et al., 2020; Neyrinck et al., 2020; Stephen et al., 2017). Ultimately, intake levels are low considering the fact that the European Food Safety Authority (EFSA), the Food and Agriculture Organization and World Health Organization (FAO/WHO) recommend the consumption of 25g of dietary fibre per day for adults (European Commission, 2021), while the recommended intake in the UK is 30g per day (BDA, 2023). Recently, it has been proposed that consumption of over 50g fibre per day is needed to achieve significant health benefits (Makki et al., 2018; Deehan et al., 2017; O’Keefe, 2018). An analysis of 17 prospective cohort studies reported that every 2g increase in cereal fibre intake per day was associated with a 6% reduction in the risk of developing type 2 diabetes (Yao et al., 2014), and a meta-analysis study reported that a 7 g/day increase in fibre intake can decrease the risk of cardiovascular disease, haemorrhagic and ischaemic stroke, diabetes, colorectal cancer, and rectal cancer (Threapleton et al., 2013).
Progress towards an accepted definition for prebiotics has spanned decades. First defined in 1995 as a “non-digestible food ingredient that beneficially affects the host by selectively stimulating growth and/or activity of one or a limited number of bacteria already resident in the colon” (Gibson and Roberfroid, 1995), this definition was updated in 2004 to “selectively fermented ingredients that allow specific changes, both in the composition and/or activity in the gastrointestinal microflora that confers benefits upon host well-being and health” (Gibson et al., 2004). Currently, the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus panel’s definition states that a prebiotic is “a substrate that is selectively utilized by host microorganisms conferring a health benefit” (Gibson et al., 2017).
The prebiotic properties of fermentable fibers such as inulin-type fructans, galactans and resistant starch (RS) have been the focus of much research (Hughes et al., 2022; Thompson et al., 2022; Neri-Numa et al., 2020; Colantonio et al., 2019; Fan et al., 2023; Guan et al., 2021). When these fibres enter the colon, they are degraded by the gut bacteria to oligosaccharides or monosaccharides using a variety of carbohydrate active enzymes (CAZymes) and absorbed as an energy source through specific transport systems (Singh, 2019). Symbiotic relationships between gut microbiota members, including for example representatives of Bifidobacterium and Lachnospiraceae, facilitate the optimal metabolism of fermentable intermediate molecules in a manner that is driven by the quantity, quality, and type of carbohydrate available (Lozupone et al., 2008). Bifidobacteria can utilize a diverse range of polysaccharides (Guan et al., 2021), and representatives are often seen to increase in abundance due to interventions with prebiotic fibres. Corresponding keystone species in the gut can contribute to community symbiosis by providing energy to other residential microbial communities in the form of acetate and lactate as cross-feeding metabolites (Guan et al., 2021; Moens et al., 2017).
Interventions involving prebiotic fibres are mostly limited to isolated single fibres, combinations of two, or triads. In relative terms, there is a lack of literature on the prebiotic effect of a comprehensive mix of soluble and insoluble fibres in a food readily available to the consumer. It is also important to consider that fibres may function differently than when incorporated together in a food matrix, such as a beverage (Livingston et al., 2016). Therefore, our study aimed to contribute to addressing these knowledge gaps by investigating the effect of a fruit juice drink containing a fibre blend consisting of chicory root inulin, green banana, golden kiwi fruit, and Baobab, and thus including soluble dietary fibres, pectins and fructans, insoluble resistant starch 2 (RS2), and cellulose lignin, on the gut microbiota structure and function, stool consistency, and emotional health in healthy humans.
2. Materials and Methods
2.1. Participants
2.1.1. Recruitment Procedure
Study participants were recruited through social media and poster advertisements around London South Bank University campus. A total of thirty-five individuals expressed their interest in participating. After reading the participant information sheet, fourteen individuals reported they were not eligible to participate due to not meeting the eligibility criteria, or not being available for the whole study duration. Twenty-one participants were enrolled.
To be eligible for the study participants must self-report to be healthy; over the age of 18; a BMI between 18.5 and 29.9; live in the UK; not consumed prebiotic- or probiotic-containing foods, drinks and supplements within the past 1 month; not vegan or vegetarian; experienced a significant change in weight (±5% of total body weight) or dietary intake over the past year; not taken antibiotics within the past 6 months; not pregnant or breast feeding; not a high alcohol consumer (>15 standard drinks per week for males or >10 standard drinks per week for females, and <2 days per week alcohol free); they or their immediate DNA family (mother, father, brother, sister) have not had a medical diagnosis for ulcerative colitis, Crohn’s disease, irritable bowel syndrome, bowel cancer, or any inflammatory bowel disease; not allergic to any of the ingredients in the product; not had brain cancer; no history of mental health disorders, or diagnosed with a disease or condition for which they receive NHS care/support/medication; not had a positive COVID-19 diagnosis, or displayed suspected symptoms in the last 6 months; not on hormone replacement therapy or following a low FODMAP diet. Employees of MOJU Ltd, and their family members, were not eligible to participate in this project.
A total of twenty eligible participants provided written informed consent to participate in this human intervention study (Fig. 1). Three participants withdrew after providing baseline data due to product delivery delays impacting on their availability to complete the whole study. Three further participants were withdrawn during the study, two due to testing positive for COVID-19, and one starting a course of antibiotics. Fourteen healthy participants, all non-smokers (4 male, 10 female) completed the entire 9-week study period. One participant’s (TS2) stool sample for the placebo phase (treatment order - prebiotic start) was lost in the post. Baseline characteristics (Mean ± SD) of the 14 individuals who completed the study were: age 42.7 ± 12.4 (range 30 – 68); body weight 72.6 ±13.3kg (range 102 – 58); BMI 24 ± 2.4) (range 22 – 28) (Supplemental Table S1).
The study was fully explained to the participants, in writing, and each gave their written, informed consent prior to participating.
2.1.2. Randomization and Group-Allocation Procedure
The Sealed Envelope Ltd (2022) software was used to apply the block randomization method to randomize participants into one of two intervention orders (i.e., AB - prebiotic then placebo, or BA – placebo then prebiotic).
2.1.3. Investigational product
A raspberry, lemon and baobab prebiotic shot was commercially produced by MOJU Ltd, UK, containing the following ingredients: Apple, MOJU Prebiotic* blend (Chicory Root Inulin* 4%, Green Banana Powder, Golden Kiwi Powder, Baobab Powder 1%), lemon 8%, ginger root, raspberry, apple cider vinegar, ascorbic acid. Participants consumed a 60ml portion daily, providing 2.4g of chicory inulin. The specific quantitative breakdown of the formulation is proprietary information. The placebo product contained the same ingredients, without the MOJU Prebiotic blend. Participants were recommended to consume their 60ml juice shot first thing in the morning on an empty stomach. See Supplemental Table S2 for nutritional data of the MOJU Prebiotic Shot.
2.1.4. Study design and Procedure
The study adopted a randomized controlled, crossover design (Figure 1). The sample size was determined based on the findings from previous studies with prebiotic fibre interventions in healthy humans (Kleesen et al., 2007; Bouhnik et al., 2007; Kolida et al., 2007; Grasten et al., 2003). See Supplemental Figure 1 for the CONSORT (Consolidated Standards of Reporting Trials) flow diagram.
The researcher involved in participant recruitment and data collection was unblinded to the intervention order. The participants were blinded to the intervention order.
Participation in this study was entirely online. Prior to commencing the intervention (week 0) participants provided a stool sample, completed a depression, anxiety, and stress (DASS-42) questionnaires (Lovibond and Lovibond, 1995), a Bristol Stool Scale (BSS) chart rating their average weekly consistency for the previous week (Lewis and Heaton, 1997), and completed a fructan food frequency questionnaire (F-FFQ) based on the previous week’s dietary intake (Dunn et al., 2011). Participants received their allocated products for Phase 1 via a courier. At the end of each intervention week (weeks 1 – 3) participants completed a DASS-42, F-FFQ and BSS questionnaire. In addition, at the end of Phase 1 (week 3), participants provided a second stool sample. At the end of the 3-week washout period (week 6) participants provided a third stool sample and completed the 3 questionnaires again. Participants received their allocated products for Phase 2 and completed the same questionnaires at the end of each week (weeks 7 – 9) and provided their fourth stool sample at the end of week 9. Participants received weekly emails containing the link to that week’s online questionnaires. If participants did not complete their questionnaires within 24 hours of receiving the notification email, they received daily reminder emails as well as reminder text messages to their mobile phones. Participants were also sent an email enquiring about adverse symptoms at the end of each intervention week. Participants were sent four fecal sample kits (Zymo Research, Europe) in the post. Fecal samples were taken by the participants at home and returned via special postal delivery. Samples were then sent to SeqBiome Ltd, Ireland (Seqbiome.com) for shotgun metagenomic analysis.
2.2. Gut Microbiome Analysis
2.2.1. Sample collection and analysis
Microbiome sample collection was performed using the DNA/RNA Shield-Fecal collection tube and Feces Catcher (Zymo Research, Europe).
DNA from samples was extracted with the QIAamp® Fast DNA Stool Mini Kit (Qiagen, West Sussex, United Kingdom) using an adapted protocol. A total of 200µL homogenized faecal sample was transferred to a screw-cap tube containing 1 ml of InhibitEX and a mixture of different sized sterile zirconia beads (All beads provided by Stratech). Then a 3-minutes bead beating step was performed to lyse the cells using a TissueLyser II (Qiagen) before the tube was heated for 5 min at 95°C. From this point, extraction was carried out according to the manufacturer’s instructions for “pathogen detection” (Qiagen, West Sussex, United Kingdom). DNA was eluted in 100μl and stored at −20°C until use in library preparation.
DNA was quantified using the Qubit double-stranded DNA (dsDNA) high-sensitivity assay kit (Bio-Sciences, Dublin, Ireland). All samples were prepared for shotgun metagenomic sequencing according to Illumina Nextera XT library preparation kit guidelines, with the use of combinatorial dual indexes (CDI) for multiplexing with the Nextera XT index kit (Illumina). Final clean libraries were assessed on the Agilent Bioanalyzer 2100 using a DNA High Sensitivity Assay Kit (Agilent), quantified by Qubit as before, and pooled using equimolar concentrations (15nM). Final sequencing pool quality check and quantification was performed by qPCR using the KAPA Library Quantification Kit for Illumina (Roche KAPA). Sequencing was performed using a NextSeq 500/550 Mid Output Kit v2.5 (300 Cycles).
Quality assessment of raw sequencing data was performed using FastQC which allows quality visualization and manual quality inspection of raw sequence data (Andrews, 2010). All samples were of generally good quality with minimal quality drop off at the end of sequences. All read statistics were
relatively consistent with on average ~76% of reads passing quality filtering steps and an average of ~86% of these being classified using Kraken2 (See Supplemental Figures S2 and S3). From this it was passed to Trimmomatic (Bolger et al., 2014) for trimming and quality filtering using the following parameters: SLIDINGWINDOW:5:22, MINLENGTH:75. The data was then passed to Kneaddata (https://huttenhower.sph.harvard.edu/kneaddata/ ) for contaminant removal. Pathway and gene family assignment was then performed using HUMAnN 3 (Beghini et al., 2020). The standard Uniref90 (Suzek et al., 2014) annotation of gene families was collapsed into Kyoto Encyclopedia of Gene and Genomes (KEGG) orthology annotation and further collapsed into KEGG module annotation (https://www.genome.jp/kegg/). Taxonomy assignment was performed using Kraken2, a taxonomic classification system using exact k-mer matches to achieve high accuracy and fast classification speeds, using a customized version of the Genome Taxonomy Database (GTDB), the results of which were analyzed using the R based phyloseq package (Parks et al., 2022; McMurdie and Holmes, 2013). GTDB clusters available genomes based on Average Nucleotide Identity (ANI), and assigns taxonomy based on the National Centre for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/) classifications. If a sequence does not meet clustering requirements (95% ANI), but does not have a unique taxonomic classification, a suffix will be added to indicate this (e.g., Escherichia coli_A). This allows for the most accurate taxonomic classifications, even in the case of organisms which have yet to be formally identified.
2.2.2. KEGG Pathways and Modules
Gene families and metabolic pathways were generated using HUMAnN3, which is a system for accelerated functional profiling of shotgun metagenomic and meta transcriptomic sequencing from host- and environmentally associated microbial communities (Beghini et al., 2021). The generated gene families were then converted from Uniref90 annotation to KEGG modules. KEGG is a database of orthologs organized in a hierarchical fashion into pathways and high-level modules (Kanehisa et al., 2017).
KEGG pathways and modules were assessed for differential abundance using ALDEx2. Using an uncorrected p-value of 0.05. KEGG pathways and KEGG modules were analyzed by computing PCAs using Aitchison distances. To test the significance of groupings, PERMANOVA tests were performed on these distances, using 999 permutations, with a p-value cut-off of 0.05. Dispersion homogeneity was tested using the betadisper function from vegan (Oksanen et al., 2022).
2.3. Statistical Analysis
SPSS IBM V27.0 and R v4.1.2 software was used to analyse all the data, using the packages vegan, CoDaSeq, Compositions, ggplot2, phyloseq and ALDEx2. Paired sample t-tests (or related samples Sign test) were used to determine within-group differences, and independent sample t-tests (or Mann-Whitney U test) to determine between-group differences. To account for the compositional structure of the Next Generation Sequencing (NGS) generated data and to avoid the likelihood of generating spurious correlations, we first imputed the zeros in the abundance matrices using the count zero multiplicative replacement method (CMultRepl, method = ”CZM”) implemented in the Compositions package (Palarea-Albaladejo and Martin-Fernandez, 2015) and applied a centred log-ratio transformation (CLR) using the codaSeq.clr function in the CoDaSeq package. Alpha and Beta diversity analysis was carried out using the R packages vegan and phyloseq and statistical comparison was tested using Kruskal-Wallis tests. Principle Coordinate Analysis (PCA) was carried out using the PCA function in R using Aitchison distance matrix (CLR + Euclidean distances). The differences between species-level taxonomy for every treatment group at each time point was analysed using paired Wilcoxon tests provided by ALDEx2, using an uncorrected p-value of 0.05 (Fernandes et al 2013). Uncorrected p-value was used as no significance was observed using corrected p-values, thus these results should be interpreted with some caution.
ALDEx2 was used to determine if any species or functional pathway modules were differentially abundant in any groups between timepoint or at any time point between the groups based on a Kruskal-Wallis test using 512 Monte-Carlo instances drawn from the Dirichlet distribution.
Estimation of alpha-diversity was performed using Shannon (Wagner et al., 2018), Chao 1 (Willis, 2019) and Simpson (Simpson, 1949) diversity metrics. Some carryover effect was observed, and so to account for this, the difference/delta of alpha diversities between time points were analyzed for significance using Wilcoxon tests.
Beta diversity was visualized by performing Principal Component Analysis (PCA) using Aitchison distances. To test the significance of these groupings, Permutational Multivariate Analysis of Variance (PERMANOVA) test was then performed using 999 permutations with significance cut-off value of 0.05. Dispersion homogeneity was tested using the betadisper function from vegan (Oksaken et al., 2022). DASS-42 data was analysed with a two-way repeated measures ANOVA with Bonferroni correction for multiple comparisons. Differences were considered statistically significant at P ≤ 0.05.
3. Results
3.1. Habitual dietary intake of inulin and oligo fructose
Long term dietary patterns and habitual food intake has significant influence in shaping individual stable gut microbiota structures, thought to explain over 50% and 20% of variability in mice and man, respectively (Leeming et al., 2019). Dietary fibre has consistently been demonstrated to impact the gut microbiome (Tanes et al., 2021) and fermentable fibres in the habitual diet would result in a gut microbiome structure designed for optimal and efficient metabolism of these fibres. Therefore, to establish potential influences for variable responses to the MOJU Prebiotic Shot, we investigated the habitual dietary intake of inulin and oligo fructose in our cohort. Participants completed a validated Fructan-FFQ (F-FFQ) (Dunn et al., 2011) questionnaire. For each food item, participants indicated their portion size (small, medium, or large) and the number of portions consumed over the previous 7 days. Portion sizes were estimated using NUTRITICS nutritional analysis software, Research Edition v5.82 (Nutritics.com). Inulin and oligo fructose amounts were calculated following the methodology previously performed (Healey et al., 2018). Composite cereal products, such as breakfast cereals and cereal bars were excluded from the calculation due to lack of detailed composite information and/or brand names.
There was a significant between-group difference in background dietary inulin intake during the placebo phase only (Table 1), with the treatment order placebo start (BA) group consuming 2.43g/day more (mean difference of 2.43g/day, 95% CI, 4.73 to 0.137, p <.05, paired t-test) (See Supplemental Table S2 for prebiotic phase). However, when we pooled the data for the whole cohort, we observed no significant difference in habitual dietary intake of inulin between the prebiotic and the placebo phase, p > .05 (See Supplemental Table S3). There were no significant within-group differences in inulin and oligofructose intakes (p > .05) between the placebo or the prebiotic phases (See Supplemental Table S4), nor between-group differences at baseline (p = .639, .645, respectively, independent t-test).
Data on population average dietary intakes of naturally occurring inulin and oligofructose was reported in 1999 for the US (2.6 and 2.5g, respectively), and 1995 for Europe (2 – 10g combined inulin and oligofructose) (Moshfegh et al., 1999). Recent data, however, remains elusive. In the present study we observed an average (Mean ± SD, n = 14) daily intake for naturally occurring inulin and oligofructose of 5.49 ± 2.5 - 6.07 ± 3.01 and 4.73 ± 2.34 - 5.12 ± 2.58 grams/day, respectively) (Supplemental Table S3). However, since we did not include composite cereal products, the average daily intakes may be higher than reported here, especially since both inulin and oligofructose are frequently used in food products such as ice cream, yoghurt, cheese, and baked goods to replace sugar and fat (Roble et al.,2011; Jackson et al., 2022).
3.2. Stool Consistency
There were no within-group significant changes in self-reported weekly stool consistency during the prebiotic fibre intervention in either treatment order group (p >0.05) (See Supplemental Table S5). Both groups reported softer stools during the prebiotic and placebo phase, and one possible explanation for this is a strong placebo effect from participating in this study (Staudacher et al., 2017). We did not collect data on stool frequency during this study. One participant in the AB group reported their stool frequency reduced during the prebiotic phase, but did not provide a number in reductions, and one participant, also AB group during the prebiotic phase, reported an increase in daily stool frequency from 1 to 2 per day for the first two weeks and from 2 to 3 per day during the third week (data not shown). Prebiotic fibres seem to impact stool consistency and frequency in those with low frequency only (Watson et al., 2019).
3.3. Safety and side effects
One participant (AB group) noted increased flatulence and a ‘grumbling tommy’ during the first week only of consumption of the prebiotic drink. One participant (AB group) noted increased intestinal bloating for the first 3 days only, and one participant (AB group) experienced increased flatulence with a sulphuric smell across the whole prebiotic intervention. There were no reported side effects from any of the BA group members during the prebiotic intervention, one reported stomach pain after the first placebo shot on an empty stomach (data not shown).
3.4. Depression, Anxiety and Stress
We observed no significant changes in depression, anxiety, or stress within any of the groups during the prebiotic or placebo phase (See Supplemental Figures S4–S7). Several participants failed to submit their DASS-42 data on either side of an intervention phase (prebiotic or placebo) which reduced the number of data sets available for analyses to ≤5 per group. Previous studies have reported prebiotic fibre amelioration of stress (Cryan and Dinan, 2017), anxiety (Schmidt et al., 2015) and depression (He et al., 2022), whereas Johnstone and colleagues observed an improvement in high anxious participants only (Johnstone et al., 2021). Subject to a high number of reviews over the past 5 years, the conclusions vary between no evidence and very little evidence for prebiotic effects on depression and anxiety (Liu, R et al., 2019; Barbosa and Veiera-Coelho, 2019; Desai et al., 2021).
3.5. Microbiome Composition Changes Due to the Intervention
3.5.1. Alpha Diversity Analysis
Both Shannon and Simpson diversity was observed to be significantly enriched as a result of prebiotic treatment when compared to placebo (p = 0.0027 and 0.0056, respectively) (Figure 2 A and B). This is in agreement with literature, as prebiotics have been shown to increase gut microbiome alpha diversity in a number of instances (Thompson et al 2017; Tandon et al 2019).
3.5.2. Beta Diversity Analysis
Treatment grouping/phase was found to be significant in regard to grouping across PCA dimension one (p = 0.001) and significant/borderline significant in regards to dispersion across PCA dimensions one to three (p = 0.0368, 0.0416 and 0.0624, respectively). This indicates that there is a considerable carryover effect, as samples tend to be grouped by treatment group and not treatment alone (Figure 3).
3.5.3. Between-Intervention Differential Abundance Analysis
Differential abundance analyses of taxa aimed to detect if there were differences in composition between the prebiotic and placebo (Lin and Peddada, 2020). We observed no significant differences in the 6 main bacterial phyla between the two groups at baseline (Table 2), however within-group responses were highly variable (Figure 4 and Table 2) and, similar to (Ramirez-Farias et al., 2009), we observed a treatment-order response. Only the participants in the prebiotic-start (AB) group responded significantly during the prebiotic phase with respect to Desulfobacteria (p = 0.016, unadjusted) and Actinobacteria (p = 0.016, unadjusted). The increase in Actinobacteria, the phylum that contains the genus Bifidobacterium, is consistent with results obtained by others (Wang et al., 2020), and substantial inter-variation responses to prebiotic fibres is frequently reported in reviews (Swanson et al 2019). After a multiple testing adjustment using the Benjamini-Hochberg method, only the changes in the Actinobacteria remained significant (p = 0.048). Actinobacteria in the treatment order placebo-start (BA) group did not increase during the prebiotic phase. A bloom in Desulfobacteria from prebiotic fibres has been reported previously (Jangid et al., 2022). (See Supplemental Figure S8 for stack bar charts of phylum level changes).
Nine species were observed to be differentially abundant between time points under prebiotic treatment (Figure 5); of these species, Bifidobacterium adolescentis (p = 0.03, unadjusted) and CAG-81 sp900066785 (p = 0.01, unadjusted) were increased relative to the baseline timepoint (Figure 5A). Three species were observed to be differentially abundant under placebo treatment, namely Coprococcus_A catus (p = 0.04, unadjusted), ER4 sp900317525 (p = 0.04, unadjusted), CAG-83 sp000431575 (p = 0.03, unadjusted) (Figure 5B). C. catus decreased relative to baseline, ER4 sp900317525 showed a slight increase and CAG-83 sp000431575 increased. However, none of these changes remained statistically significant after correction for multiple testing.
Of these species, perhaps the most relevant is Bifidobacterium adolescentis, which shows a clear pattern of enrichment under prebiotic treatments (Figure 2C). This is in agreement with literature, with Bifidobacterium species being one of the commonly enriched taxa in response to many prebiotic treatments (Wang, S et al., 2020), including pectin (Bang et al., 2018; Pascal et al., 2022; Mailing et al., 2019). CAG-81 sp900066785, from the Firmicutes phyla, and the Lachnospiraceae family, is regularly observed in healthy humans with the average abundance detected ranging from 0.06% to 2.25% (unseenbio.com, 2023). Members of this family are known carbohydrate fermenters and starch degraders (Flint et al., 2012) and demonstrated to be increased by inulin and FOS supplementation in murine models (Kleesen et al., 2001; Berger et al., 2022), including pectic oligosaccharides (POS) (Manderson et al., 2005; Bang et al., 2018; Pascal et al., 2022). Coprococcus catus, decreased during the placebo phase, another member of the Lachnospiraceae family, is one of the core members of the human gut microbiota and is demonstrated to contain fructan and starch degradation enzymes. It is a succinate, acetate, lactate, and propionate producer, and was demonstrated to grow on chicory inulin and apple pectin in vitro (Shetty et al., 2022), and enriched in the gut microbiome, alongside Bifidobacterium, of pectin fed rats (Mailing et al., 2019). It is therefore probable that the withdrawal of the prebiotic fibres during the placebo phase impacted on C. catus proliferation.
CAG-83 sp000431575 is from the Firmicutes phylum, family Oscillospiraceae, was demonstrated by the same group to be enriched in the gut microbiome in pectin fed rats (Mailing et al., 2019). Oscillospira is considered one of the next-generation probiotic candidates. ER4 sp900317525 is also from the family Oscillospiraceae. They are butyrate producers and ferment complex plant carbohydrates. Moreover, bifidobacteria (Tian et al., 2020), and FOS (Zhao et al., 2021), and an increase in plant-based meat alternatives in the diet known to contain higher levels of mixed dietary fibres (Toribio-Mateas et al., 2021), have demonstrated to have a positive effect on Oscillospira. An increase observed in these two during the placebo phase in our study could indicate a carryover effect. The precise definition of what constitutes a carryover effect is not clear, and we propose the increase in CAG-83 sp000431575 and ER4 sp900317525 during the placebo phase is an example of ‘prolonged benefit’ as opposed to a carryover effect.
Notable individual variable responses include subject TS2 who displayed a large shift in taxonomic composition between time points during prebiotic treatment, and subject VI4 who showed high levels of Akkermansia during the placebo treatment but not during prebiotic treatment (Figure 4).
3.5.4. Microbiome Functional Changes Due to the Intervention
3.5.4.1. Metagenome Functional Analysis
Several significantly differentially abundant pathways and modules were observed between time points during both placebo and prebiotic treatment, again indicating a potential carryover effect (Figure 6). The KEGG pathway associated with the citrate cycle was decreased during both placebo (p = 0.03, unadjusted) and prebiotic (p = 0.02, unadjusted) treatment. The pathway associated with vitamin B6 metabolism (p = 0.04, unadjusted) was downregulated during prebiotic treatment, and the pathways associated with microRNAs in cancer (p = 0.01, unadjusted) and cysteine/methionine (p = 0.04, unadjusted) were decreased during placebo treatment. In a similar fashion, multiple KEGG modules associated with the citrate cycle (Figure 6) were decreased during both treatments (prebiotic p = 0.05; placebo p = 0.04, unadjusted). Perhaps of most interest is the various upregulated modules associated with arginine metabolism observed during the prebiotic treatment (p <0.05, unadjusted) (Figure 6A and Figure 8). None of these changes remained statistically significant after correction for multiple testing.
The presence of a carryover effect is further supported by the fact that during PCA analysis, treatment grouping was significant in regard to grouping across all three PCA dimensions, and significant in regard to dispersion across PCA dimensions one and two. While treatment was also observed to be significant in regard to grouping and dispersion across multiple dimensions, it seems that much of this variability may be due to treatment grouping, making it difficult to determine if this significance is genuine. This can be seen in Figure 7, where the most distinct clusters consist nearly solely of samples from the prebiotic start group during placebo treatment, i.e., the samples the longest time after prebiotic treatment. This may indicate that there is a longer-term effect of prebiotic treatment which is not completely accounted for by the 21-day washout period. This pattern is consistent between both KEGG pathways and KEGG modules.
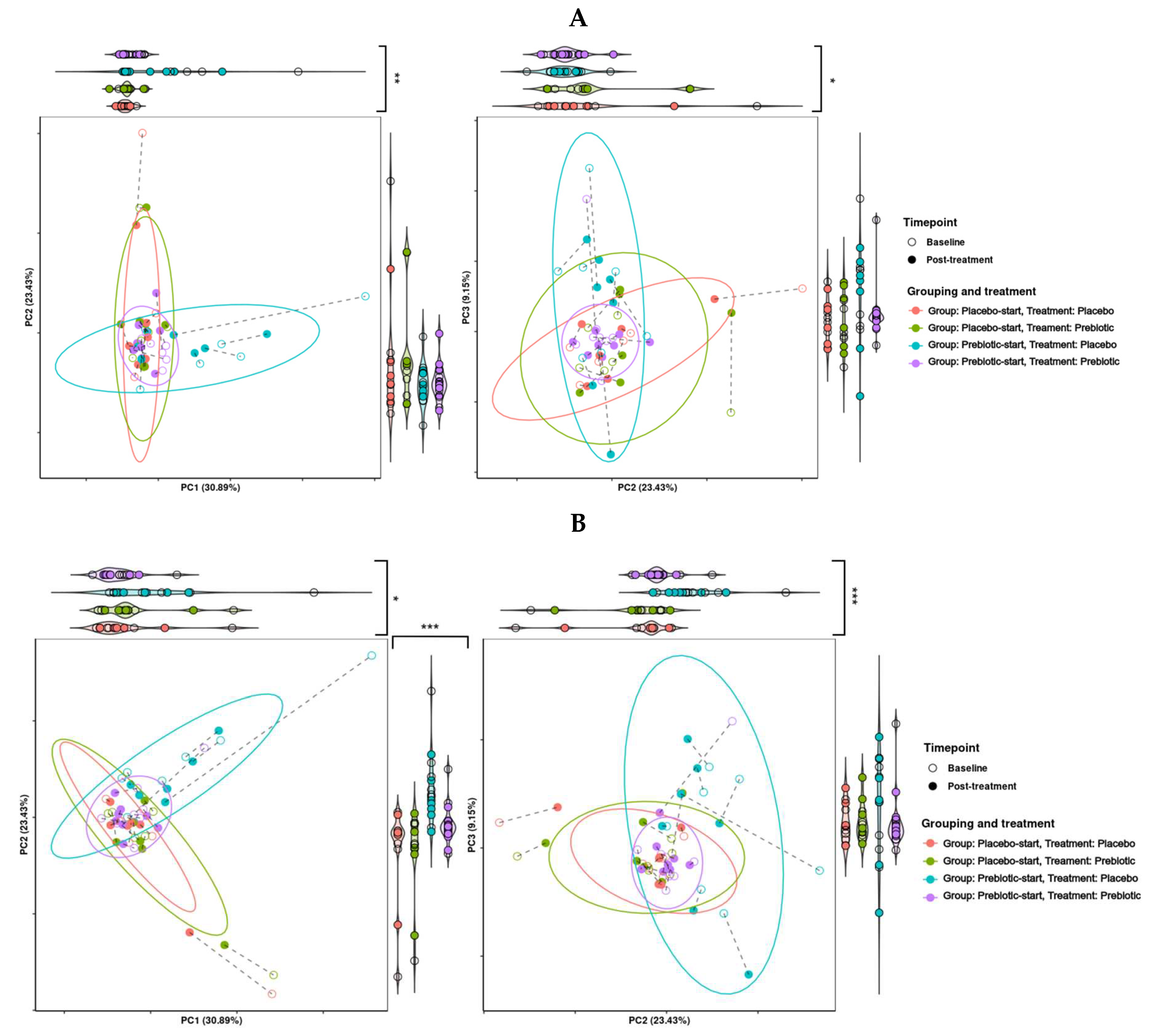
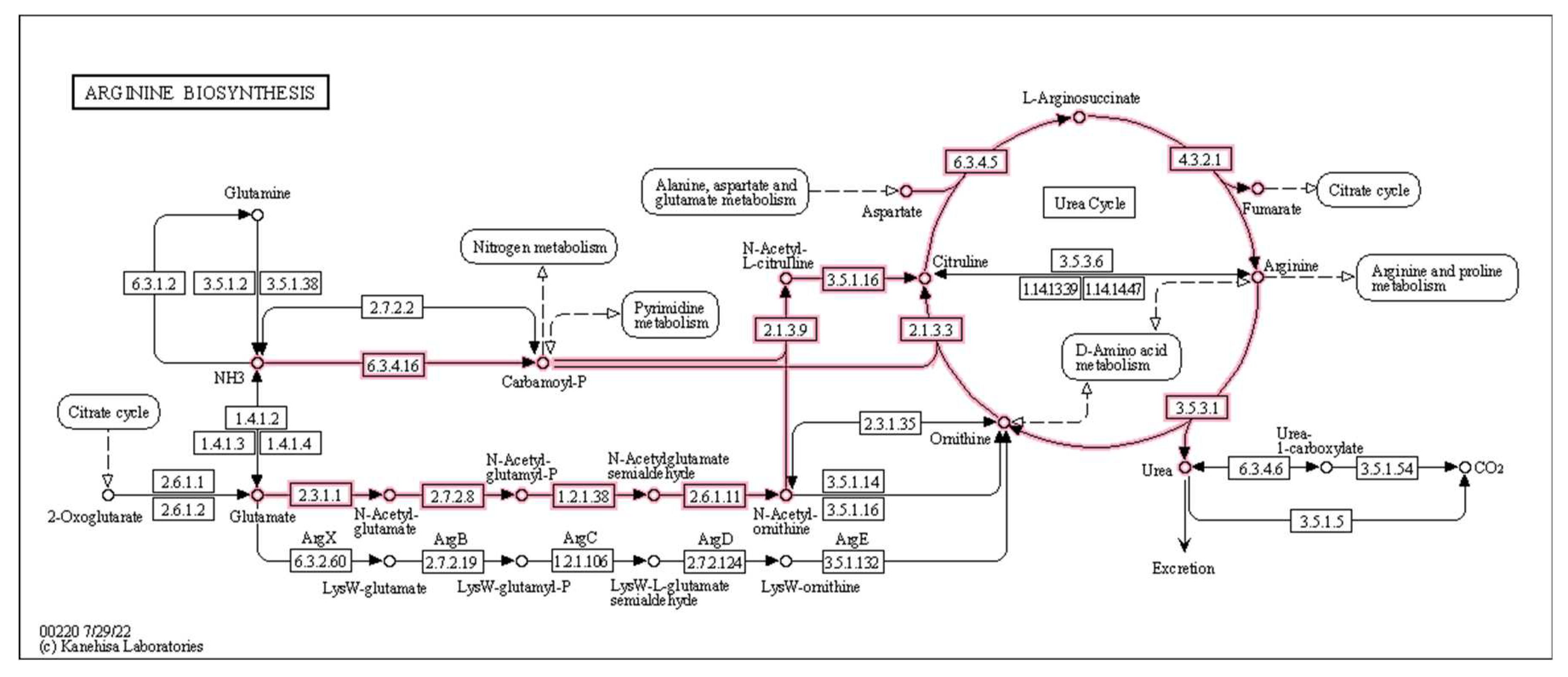
Figure 7.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults. Principle component analysis (PCoA) is based on Aitchison distance matrixes. PCA plots of PCA dimensions 1,2 and 3 of (A) KEGG modules, and (B) KEGG pathways. Ellipses represent 80% confidence intervals. Lines connect the same samples during each time point, with shapes indicating the time point, coloured by treatment and grouping. Both graphs are bordered by violin plots. Violin plots with significantly different grouping are marked, with * = p-value <0.1, ** = p-value <0.05 and *** = p-value < 0.01.
Figure 7.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults. Principle component analysis (PCoA) is based on Aitchison distance matrixes. PCA plots of PCA dimensions 1,2 and 3 of (A) KEGG modules, and (B) KEGG pathways. Ellipses represent 80% confidence intervals. Lines connect the same samples during each time point, with shapes indicating the time point, coloured by treatment and grouping. Both graphs are bordered by violin plots. Violin plots with significantly different grouping are marked, with * = p-value <0.1, ** = p-value <0.05 and *** = p-value < 0.01.
Figure 8.
Effect of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults, showing upregulated arginine biosynthesis by Bifidobacterium adolescentis KEGG Modules M00844, M00845, and the Urea Cycle M00029 during the prebiotic phase.
Figure 8.
Effect of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults, showing upregulated arginine biosynthesis by Bifidobacterium adolescentis KEGG Modules M00844, M00845, and the Urea Cycle M00029 during the prebiotic phase.
4. Discussion
In this study we investigated the effects of a fruit juice drink (MOJU Prebiotic Shot) containing a mixture of fermentable and prebiotic fibres on the structure and function of bacterial communities in the human colon, depression, anxiety and stress, and stool consistency, utilizing a cross-over approach.
The fermentable fibres in the MOJU Prebiotic Shot include non-starch polysaccharides, specifically chicory inulin and pectic-oligosaccharides from golden kiwi fruit and baobab, as well as resistant starch (RS2) from green banana. These fibres are resistant to host enzyme degradation and pass through the small intestine into the colon where they can be fermented by a variety of gut commensals. For a negative control, we used the same fruit juice drink that did not contain the mixture of fibres. A 60 ml portion contains 3.7g of dietary fibre, 2.4g, of which, is contributed by chicory root inulin.
Inulin, considered the ‘gold standard’ prebiotic, is the most extensively studied prebiotic fibre to date (Le Bastard et al., 2019; Hughes et al., 2022). Commercial baobab fruit pulp powder is rich in polyphenols and flavonoids and contains around 45 - 54% (dry weight) fibre, 50 – 75% of which are soluble fermentable fibres, including pectin-based poly- (PPOS) and oligosaccharides (POS) (42.5%) (Foltz et al., 2021; Asogwa et al., 2020). POS has previously been demonstrated to have prebiotic effects (Wongkaew et al., 2022), including bifidogenic effects (Mussatto and Mancilha 2007; Garthoff et al., 2010; Hotchkiss et al., 2003; Foltz et al., 2021).
Green banana flour (GBF) contains around 8.5 – 15.5g/100g of fibre, with a soluble/insoluble fibre ratio of 1:5, and 40 – 54.2g/100g resistant starch (RS2) (Bezerra et al., 2013; Li et al., 2022; Alvarado-Jasso et al., 2019, Livingston et al., 2016). Li et al (2022) demonstrated GBF restored gut permeability and intestinal barrier function over a 2-week intervention in an antibiotic gut microbiota dysbiosis model in rats faster than natural recovery, by increasing mucin secretion. Various beneficial effects of GBF have been reported in animal models, including modulating intestinal inflammation in a colitis model (Shinde et al., 2020), improvement of obesity parameters (Alvarado-Jasso et al., 2019), increase SCFAs and modulate oxidative stress (Almeida-Junior et al., 2017).
Kiwi fruit is rich in POS, polyphenols, and flavonoids. Both green and gold varieties have been investigated for their gastrointestinal (GI) symptom improvements in humans (Chan et al., 2007; Blatchford et al., 2017; Rush et al., 2002) and pigs (Han et al., 2011). Using an in vitro digestion model, Parkar and colleagues demonstrated gold kiwifruit increased Bifidobacterium, including enhancing the adhesion to intestinal epithelial cells (Parkar et al., 2021).
The MOJU Prebiotic Shot used in our study elicited a significant change in the overall composition of the gut microbiome, as demonstrated by both Shannon and Simpson diversity indices. Alpha diversity changes from prebiotic fibre interventions are not consistently reported and great variability exists amongst those who do report results (So et al., 2018). Fibre dosage does not appear to influence alpha diversity, for example chicory inulin at 20g (Tims et a.,l 2016) and 10g (Watson et al., 2019) per day observed no changes, whereas 12g (Vandeputte et al., 2017) and 6g/8g (Reimer et al., 2017) per day reported reductions in alpha diversity. One of the reasons for such variability may be due to the sequencing method used. General 16S rRNA amplicon (V3-V4) sequencing can produce false positive results, overestimating certain taxa (Leeming et al., 2021) and underreporting alpha diversity scores (Jeong, J et al., 2021), relative to shotgun sequencing (Durazzi et al., 2021; Klimenco et al., 2022), which would increase alpha diversity scores when compared with short read 16S rRNA sequencing.
We observed grouping of paired samples (beta diversity) according to treatment order rather than treatment phase, indicating potential of a carryover effect where the 3-week wash out period in the treatment order prebiotic start (AB group) was not sufficient to return the gut microbiota composition to pre-intervention baseline. Fibre intervention trial design was reported to influence the magnitude of outcomes with parallel design studies demonstrating stronger intervention effects and greater statistical heterogeneity in comparison to a crossover design, seemingly for several outcomes, and the carryover effect is proposed to be one of the reasons (So et al., 2018; Hughes et al., 2019 Part 1). Reviews on fibre interventions have reported that not all cross over trials include wash out periods (So et al., 2018; Liu et al., 2017b) and reporting of a carryover effect after a wash out period is rare. It is mainly accepted that fibre-induced changes to the gut microbiota are only maintained whilst the fibres are consumed (Makki et al., 2018; Leeming et al., 2019). We chose a 3-week washout period based on expert recommendations and previous findings (Ho et al., 2019; Boughnik et al., 2017; Liu Feitong et al., 2017; David et al., 2014; Hiel et al., 2019), including practical considerations such as duration of participant commitment and to control the influence of seasonal and holiday dietary changes. Furthermore, although we observed no significant differences between groups in habitual dietary inulin and FOS levels at baseline, and pooled data demonstrated no significant difference between the intervention phases, we did observe a between-group difference (p <.05) for inulin during the placebo phase, and the potential contribution of this difference to our treatment order groupings should not be ignored. It has been proposed to stratify participants according to their baseline microbiota enterotype and include an initial exploratory study to investigate the required washout period (Swann et al., 2020), however, beyond the cost implications and practical feasibility, it would divert from the purpose of our study which was to investigate the effect of a commercially available food product in the average healthy consumer.
Similar to other studies investigating gut microbiota changes in humans from prebiotic fibres we observed a bifidogenic effect (Kang et al., 2022; Hiel et al., 2019; Kleesen et al., 2007; Boughnik et al., 2007; Kolida et al., 2007; Baxter et al., 2019; Ramirez-Farias et al., 2008; Sawicki et al., 2017), in combination with an increase in Lachnospiraceae (Berger et al., 2021; Salonen et al., 2014). B. adolescentis appears to respond selectively to inulin (Guarino et al., 2020; Hiel et al., 2019; Joossens et al., 2011; Ramirez-Farias et al., 2009; Salazar et al., 2015; Swanson et al., 2020; Wang H et al., 2022) and resistant starch (Makki et al., 2018; Baxter et al., 2019; Venkataraman et al., 2016; Flowers et al., 2019; Kelly et al., 2016). Bifidobacteria in the gut are linked with numerous health benefits including SCFA production (Deleu et al., 2021) and improved function of the gut barrier (Al-Sadi et al., 2021). Reduced intestinal inflammation (Assimakopoulos et al., 2018), a reduction in circulating lipopolysaccharides (Ghosh et al., 2020), and healthy gut immune responses (Gasaly et al., 2021) are all linked to improved gut barrier functionality. As keystone species, Bifidobacterium spp. can apply several survival strategies in a highly competitive environment, including glycan-harvesting and glycan breakdown, leading to cross-feeding metabolites that supports community stability. Several studies have reported the symbiotic relationship between bifidobacteria and other members of the healthy gut microbiota by means of in vitro co-culturing experiments (Scott et al., 2011; Falony et al., 2006; Flint et al., 2014), including B. adolescentis (Belenguer et al., 2006). The trophic interaction between bifidobacteria and Lachnospiraceae involving lactate as a cross-feeding intermediary metabolite has previously been reported (Berger et al., 2021; Riviere et al., 2016; Schwab et al., 2017) and is of great importance to the host and the gut microbiota community (Louis and Flint, 2017). As one of the largest consortia of butyrate producers, an increase observed in Lachnospiraceae we considered a butyrogenic effect (Riviere et al., 2015). It is anticipated, in line with other studies (Cummings et al., 2001; Tsitko et al., 2019; Bang et al., 2018), the prebiotic fibres elicited an increase in butyrate production. An elegant study by Swanson and colleagues (2020) gave insight into the cross-feeding and butyrate effect of chicory inulin in the human colon. They demonstrated inulin fermentation was associated with a concurrent increase in butyrate even though acetate and lactate are the only bifidobacterial fermentation products.
Furthermore, B. adolescentis is recognised for its increased genome stability, making it particularly competitive in the gut environment. In comparison to other bifidobacteria, B. adolescentis has more genes involved in quorum sensing, biofilm formation, the two-component system and various carbon source and amino acid metabolism pathways, demonstrating its competitive advantage (Qian et al., 2022). Indeed, several of the increased functional genes observed in our study, discussed below, are expressed by B. adolescentis, demonstrating its comprehensive capability of host and gut bacterial community health promotion.
We were particularly interested in the several upregulated amino acid (AA) related functional modules during the prebiotic treatment (Figure 6A). Two pathways for arginine biosynthesis (KEGG Modules M00845 and M00844), the urea cycle, and phenylacetate degradation to acetyl-CoA/succinyl CoA. B. adolescentis express the genes for all these pathways (https://www.genome.jp) and may be the reason for the upregulation of these functions observed in our study (Figure 8).
A variety of AAs are required for gut microbial production of SCFAs (Yang, Y et al., 2021), and microbial-derived neurotransmitters (e.g., GABA, dopamine, and serotonin), and most interestingly, for amino acid-dependent environmental stress-adaptation mechanisms (Lund et al., 2014). Evidence for the de novo synthesis of some of the nutritionally essential amino acids (Metges, 2000) by gut bacteria, such as arginine, means AAs can function as important microbe-host and microbe-microbe cross-feeding metabolites. For example, arginine, as a precursor for the biosynthesis of proteins and arginine-derived cross-feeding metabolites, including polyamines, nitric oxide, glutamate, urea, and creatine (Kim et al., 2022), is essential for the maintenance of intestinal microbiota homeostasis (Rao et al 2020). In the gastrointestinal tract (GIT) it drives three predominant metabolic pathways: (i) synthesis of polyamines through the provision of intermediate ornithine and agmatine; (ii) synthesis of urea, creatine, and ornithine; and (iii) direct catabolism into nitric oxide (Yang, Y et al., 2021). Competition for arginine in the GIT is high, since it is also extensively metabolised by intestinal epithelial cells (IECs) to produce adenosine triphosphate (ATP), mucins, immunoglobins and defensins. Consequently, around 38 - 40% of dietary arginine in the intestinal lumen is catabolized in the first-pass order (Talavera et al., 2017; Wu and Morris 1998; Uyanga et al., 2021), highlighting the fundamental importance of the de novo synthesis of arginine in the colon by B. adolescentis to both host and gut microbiome health. Arginine supplementation has been demonstrated to influence gut microbiota composition in mice (Ren et al., 2014; Singh et al., 2019), and chickens (Zhang, B et al., 2019). Kim and colleagues reported an increase in Bifidobacterium spp. in mice, (Kim et al., 2022) and Van Den Abbeele and colleagues (2022) observed a significant increase in Lachnospiraceae in pigs. Taken the above, arginine functions as an important cross-feeding metabolite for beneficial gut bacteria.
Furthermore, arginine fulfils an important role in survival strategies activated in the gut microbiota under acid stress (Lund et al., 2014). Bifidobacterium can significantly upregulate genes to support growth at low pH (Lund et al., 2014) and appears to use two different acid-tolerance response (ATR) protective mechanisms against acid stress. The F1-F0 ATPase (Matsumoto et al., 2004) and the amino-acid dependent decarboxylase/antiporter system (Ventura et al., 2009), and both require arginine (Lund et al., 2014; Vracken et al., 2009). B. adolescentis is not acid tolerant and relies on these ATRs for survival and maintenance (Matsumoto et al., 2004).
B. adolescentis is known to produce acetate and lactate as intermediate fermentation products (Sheridan et al., 2022), and increased colonic SCFAs from prebiotic fibre interventions is well reported (Holscher, 2017; Liu et al., 2017), meaning the MOJU Prebiotic Shot used in our study may have resulted in an increase in the colonic SCFA pool, and the resultant increase in intestinal acidity would have triggered a requirement for an ATR (Oufir et al., 1996; Den Besten et al., 2013; Duncan et al., 2009). This may be one of the reasons why we observed the upregulation of two arginine biosynthesis pathways, as well as the urea cycle by B. adolescentis since urea is also utilised in an acid tolerance response in gram positive bacteria (Lund et al., 2014). Ureolytic gut commensals, such as Bifidobacteria spp. (LoCascio et al 2010; Ryvchin et al 2021) utilise urease (Regan et al 2021) to turn urea into ammonia and carbonic acid, which increases intestinal pH, protecting intestinal bacteria from acidity that is either self-created (for example from acetate or lactate producers such as B. adolesentis) or host-derived (for example stomach acid) (Rutherford 2014). The ammonia produced here can enter the urea cycle as a substrate to produce arginine, urea and ornithine, and ornithine can be used as an intermediate in the arginine biosynthesis pathway, leading to a perpetual recycling of important intermediates into arginine. (Figure 8). Zarei and colleagues (2022) demonstrated increased urea in the colon of germ free (GF) mice from arginine metabolism through the urea cycle in comparison to conventionally raised mice, meaning the gut microbiota is essential for urea catabolism. It is suggested the microbial community can hydrolyze between 15 and 30% of urea synthesized in healthy subjects (Mora and Arioli, 2014). Urea, synthesized from arginine in the Urea cycle, can be utilized for the de novo synthesis of microbial proteins and considering the importance of urease in nitrogen recycling, especially when diets are deficient in protein, makes the ability of the gut microbiome to produce and utilize urea particularly advantageous to both microbe and host (Mora and Arioli, 2014).
Arginine, therefore, can be seen as an important commensal gut microbiota currency, and the fact that the MOJU Prebiotic Shot elicited an increase in its production by B. adolescentis, can be considered beneficial to both gut microbiota and host.
Furthermore, the ability of B. adolescentis to degrade colonic phenylacetate to the intermediate metabolites Acetyl-CoA and Succinyl-CoA, is advantageous to both host and gut microbiota community. After carbohydrates, natural occurring aromatic compounds, found as lignin (aromatic polysaccharide), flavonoids, quinones and aromatic amino acids, are the second most abundant compounds in nature (Carmona et al., 2019). Not surprisingly, these compounds can serve as growth substrates for microorganisms (Junghare et al., 2019). After cellulose, lignin is the most abundant polymer in nature (Adler, 1977). Lignin, as part of the structural make up of plant cell walls, exists as lignocellulose, a complex mixture of carbohydrate polymers, hemicellulose and cellulose covalently bound to lignin and pectic substance, and accounts for 30 – 50% of dry matter (Popoola-Akinola et al., 2022). The cross-linked structure of hemicellulose and lignin polyphenols makes it more resistant to intestinal microbiota (Ma et al., 2023).
The fibres in the MOJU Prebiotic Shot used in our study are recognized sources of lignin. Baobab fruit cells (dried) contains 54% (Kabbashi et al., 2017), kiwifruit (wet) contains between 8 and 12% (Prathibhani et al., 2021). Elsewhere, green banana was demonstrated to be a source of lignin (Falcomer et al., 2019). Plants utilize the aromatic amino acids phenylalanine and tyrosine for the biosynthesis of phenolic compounds and lignin (Feduraev et al., 2020), and phenylacetate is a reductive intermediate of the anaerobic catabolism of these aromatic amino acids. Bifidobacteria was demonstrated to ferment lignin and metabolize the released phenolic compounds in vitro (Niemi et al., 2013), and lignin-carbohydrate complexes have demonstrated to be metabolized by intestinal microorganisms to SCFAs and increase bifidobacteria (Zhang et al., 2021; Ma et al., 2023).
Moreover, phenylacetate may also trigger virulence in pathogens (Teufel et al., 2010) and high levels of intestinal phenylacetate has been demonstrated to be implicated in colorectal cancer (Le Gall et al., 2019), and autism spectrum disorder (Zheng et al., 2021). Consequently, the removal of circulating phenylacetate in the colon by B. adolesentis, functions as an important step to host health maintenance, and the prevention of pathogen proliferation in the gut.
Furthermore, the end products of phenylacetate degradation by B. adolesentis, namely acetyl-CoA and Succinyl-CoA, function as cross-feeding metabolites to butyrogenic bacteria such as Lachnospiraceae. Both succinyl-CoA and acetyl-CoA are used to produce butyrate by butyrogenic bacteria (Louis and Flint, 2017), with the acetyl-CoA pathway demonstrated to be the most prevalent in mice (Singhal et al., 2021). Lachnospiraceae, utilizes the acetyl-CoA pathway to convert acetate to butyrate (Ge et al., 2022) and this cross-feeding example may be one of the reasons why studies with prebiotic fibres, such as ours, often report an increase in Lachnospiraceae alongside Bifidobacteria spp.
In addition, we observed an increase in Desulfobacteria (p = < 0.05, unadjusted) in one of our treatment order groups during the prebiotic intervention. Lignin and RS2 are both insoluble fibres, and Jangid and colleagues (2022) recently demonstrated an increase in Desulfovibrio from an insoluble fibre intervention in mice, the bloom, hypothesized to be the result of increased hydrogen (H2) formation from an influx of insoluble fibres available for fermentation. Key bacterial fibre fermentation products include both SCFAs and gases such as hydrogen (H2) and methane (Rowland et al., 2018), and members of the genus Desulfovibrio help in the removal of excessive H2 by converting it into hydrogen sulphide (H2S) frequently reported to have an odor like ‘eggy sulphur’, as reported by one of our subjects during the prebiotic phase in our study.
In addition, the prebiotic effect of the MOJU Prebiotic Shot used in our study is not limited to just the fermentable fibres. It contains a variety of polyphenols such as flavonoids and anthocyanins considered substances with prebiotic activity (Gibson et al., 2017; Wang et al., 2022; Ozdal et al., 2015) that has demonstrated a bifidogenic effect, modulate mucin synthesis, improve intestinal barrier structure and function, and increased SCFAs in vitro, in animals and humans (Alves-Santos et al., 2020; Nunez-Gomez et al., 2021; Silva et al., 2022; Guo et al., 2021; Islam et al., 2022; Peron et al., 2021). It is proposed the action of polyphenols on the gut microbiota relies on dual antimicrobial (Daglia, 2012; Makarewicz et al., 2021; Alvarez-Martinez et al., 2020) and growth-stimulating effects (Chang CJ et al., 2019; Boto-Ordonez et al., 2014) and has coined the term ‘duplibiotic’ (Rodriguez-Gaza et al., 2021). Moreover, the effects of polyphenols on B. adolesentis have been investigated in vitro and demonstrated to promote growth (Gwiazdowska et al., 2015), enhance anti-inflammatory activity and stimulate production of acetate and lactate (Kawabata et al., 2018). Furthermore, raspberries, rich in polyphenols such as ellagitannins and anthocyanins (Mazur et al., 2014; Tomas, 2022) increased Lachnospiraceae in an obese murine model (Garcia-Mazcorro et al., 2018), and ginger demonstrated a bifidogenic effect and increased SCFAs in mice (Wang J et al., 2019), and alpha diversity in humans (Wang X et al., 2021). It is important to note that the placebo product used in our study also contained raspberry and ginger, the only difference between the MOJU Prebiotic Shot and placebo was the fibres.
Interestingly, the synergistic relationship between fibres and polyphenols have been reported (Rodriguez-Daza et al., 2020,2021; Edwards et al., 2017; Guo et al., 2018). Unless chemically extracted, polyphenols are intimately bound through hydrophobic and hydrogen bonds to plant cell wall matrix, such as lignin cellulose, and reach the colon in the form of polyphenolic fibres (Bravo et al., 1994; Gonzales-Aquilar et al., 2017). The fibre acts as an entrapping matrix and restricts host digestive enzyme access, meaning around 80% - 95% (Aura et al., 2005; Mohsen et al., 2006; Clifford, 2004) of the polyphenols bound to these fibres reach the colon. Once the colonic microbiota ferments the fibres, resulting in SCFAs, the released polyphenols are deconjugated into microbiota-derived polyphenol metabolites (Tang et al., 2020; Silva et al., 2022) by members of the gut microbiota, such as Bifidobacteria and Lachnospiraceae (Riva et al., 2020; Amaretti et al., 2015; Dabek et al., 2008; Guo et al., 2021) that express polyphenol-associated enzymes (PAZymes) (Fan S et al., 2023). In return, SCFAs facilitate the increased absorption of these metabolites (Van Rymenant et al., 2017) demonstrating a gut microbiota-fibre-polyphenol axis that is beneficial to the host (Annunziata et al., 2021; Zhao and Juang, 2021; Zhang et al., 2023). This could be one of the reasons why conflicting results of the effects of polyphenols on gut microbiota are often reported since extracted polyphenols are not fibre-bound.
Taken the above, the phytochemicals present in the functional fruit juice drink with prebiotic fibres may have contributed to the structural changes and increased functional potential of the gut microbiota observed in our study in several ways: (i) directly by means of their proliferation effects on Lachnospiraceae and B. adolesentis, and (ii) cross-feeding effects from the microbial phenolic metabolites, and (iii) indirectly through their anti-microbial actions, and (iv) their beneficial impact on the mucosal epithelial barrier.
We consider the cross-over design of our study a strength. The cross-over design of the study increases the power to detect changes in response to the treatment with a small number of subjects and a shorter period (Kang et al., 2022), however, the design is also responsible for the main weakness of our study considered to be the small sample size available for emotional health data processing. Limited funding prevented recruiting to a number that considers for standard potential losses to follow-up and losing 3 participants at the start due to product delivery issues was not anticipated. The online-only approach was both beneficial and detrimental. Finding participants who meet the inclusion criteria and able to commit to the duration of the study is difficult, reducing the need to travel to London South Bank University improved interest to participate. Conversely, ensuring participant compliance by completing the online questionnaires at the end of each intervention week, proved a challenge and resulted in the biggest loss of data available for depression, stress and anxiety analyses.
Our results indicate the MOJU Prebiotic Shot is safe for both men and women and the daily consumption in healthy individuals provides a plausible amount of ingredients to demonstrate a prebiotic effect. Moreover, the provision of an additional 3.66g of dietary fibre in a convenient manner, means it may be beneficial to the general public, especially in terms of the increase in the relative abundance of B. adolescentis and CAG-81 sp900066785, a member of the Lachnospiraceae phyla, well known for their beneficial health effects.
To our knowledge, this is the first study providing insight into the ‘niche factor’ (Hill, 2012) strategies and functional potential of B. adolescentis brought about by a 3-week intervention of a fruit juice drink with prebiotic fibres in healthy humans. Our results provide evidence of complex cross-feeding pathways and indicate areas for future research such as the importance of the gut microbiota-arginine-host axis. In addition, we add to the lacking pool of data of fibre intervention trial wash-out duration and cross-over implications and the importance of measuring habitual dietary prebiotic fibres.
5. Conclusion
Three weeks of supplementation with 60ml of the MOJU Prebiotic Shot, a fruit juice drink with a mixture of prebiotic fibres rich in secondary metabolites including polyphenols, significantly changed gut microbiota alpha and beta diversity, and resulted in measurable increases in beneficial organisms such as B. adolesentis and CAG-81 sp900066785, a member from the Lachnospiraceae phyla, in generally healthy individuals. Our metagenomic data revealed upregulated pathways and modules during the prebiotic intervention phase shedding light on the specific functions assigned to B. adolescentis. In a world where we are not consuming enough dietary fibre together with the knowledge that variety of fibres in the diet matter, and environmental influences to fibre content in food crops means there is disparity between what we eat and what we think we eat (Ioniță-Mîndrican et al., 2022), our results demonstrate the tangible benefit of a convenient daily fibre top-up in the form of a functional fruit juice drink with a mixture of prebiotic fibres.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary information includes 1. Raw metagenome data and diversity statistics. 2. Subject characteristics. 3. MOJU Prebiotic Shot nutritional information. 4. Inulin and oligofructose habitual dietary information. 5. BSS consistency changes. 6. CONSORT Flow diagram. 7. Stool sample quality filtering data. 8. DASS-42 results graphs. 9. Gut microbiome phyla stack bar charts.
Author Contributions
Conceptualization, A.B; methodology, A.B; project administration A.B; resources A.B; software M.O., A.B, and C.C; formal analyses M.O., S.D., A.B., and C.C; visualization, M.O., A.B., and C.C., writing – original draft preparation, A.B. and M.O.; writing – review and editing, A.B., M.O., P.C., S.D., C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by South Bank Innovation’s London Agri-Food Innovation Clinic (LAFIC) which was set up to support and promote research and innovation in London’s food businesses. It is co-funded by London South Bank University and the European Regional Development Fund (ERDF). MOJU Ltd (UK) provided the intervention products.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the School of Applied Sciences Ethics Committee (SASEC) of London South Bank University, ETH2021-0175.
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the study. None of the data shared in this article can lead to the identification of any of the participants.
Data Availability Statement
All data described in the manuscript, not available in the Supplementary Materials, will be made available upon reasonable request.
Acknowledgments
We thank Ross Austen, Research and Nutrition Lead at MOJU Ltd, for his support with the logistics and delivery of the intervention products to the participants.
Author disclosures: A.B. and C.C. are employees of LAFIC, London South Bank University, that funded and managed the trial. P.D.C., M.O. and S.D. are employees of SeqBiome, Ireland, which was contracted by the LAFIC to carry out the gut microbiota shotgun metagenome analyses. We confirm that MOJU Ltd had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Adler, E. (1977) Adler 1977, Wood Sci. Technol, 11 , pp. 169.
- Agostoni, C., Bresson, J., Fairweather-Tait, S., Flynn, A., Golly, I., Korhonen, H., Lagiou, P., Løvik, M., Marchelli, R., Martin, A., Moseley, B., Neuhäuser-Berthold, M., Przyrembel, H., Salminen, S., Sanz, Y., , S., Strain, J. J. )., Strobel, S., Tetens, I., Tomé, D., Van Loveren, H., Verhagen, H., Asp, G., Becker, W., Van Den Berg, H., Hulshof, K., Flynn, A., Martin, A., Przyrembel, H., Tetens, I., , E., Valtueña, S. and , M. (2010) Scientific opinion on dietary reference values for carbohydrates and dietary fibre, EFSA Journal, 8 (3), pp. n/a. [CrossRef]
- Almeida-Junior, L. D., Curimbaba, T. F. S., Chagas, A. S., Quaglio, A. E. V. and Di Stasi, L. C. (2017) Dietary intervention with green dwarf banana flour (musa sp. AAA) modulates oxidative stress and colonic SCFAs production in the TNBS model of intestinal inflammation, Journal of Functional Foods, 38 , pp. 497-504. [CrossRef]
- Al-Sadi, R., Dharmaprakash, V., Nighot, P., Guo, S., Nighot, M., Do, T. and Ma, T. Y. (2021) Bifidobacterium bifidum enhances the intestinal epithelial tight junction barrier and protects against intestinal inflammation by targeting the toll-like receptor-2 pathway in an NF-κB-independent manner, International Journal of Molecular Sciences, 22 (15), pp. 8070. [CrossRef]
- Alvarado-Jasso, G. M., Camacho-Díaz, B. H., Arenas Ocampo, M. L., Jiménez-Ferrer, J. E., Mora-Escobedo, R. and Osorio-Díaz, P. (2020) Prebiotic effects of a mixture of agavins and green banana flour in a mouse model of obesity, Journal of Functional Foods, 64 , pp. 103685. [CrossRef]
- Alves-Santos, A. M., Sugizaki, C. S. A., Lima, G. C. and Naves, M. M. V. (2020) Prebiotic effect of dietary polyphenols: A systematic review, Journal of Functional Foods, 74 , pp. 104169. [CrossRef]
- Amaretti, A., Raimondi, S., Leonardi, A., Quartieri, A. and Rossi, M. (2015) Hydrolysis of the rutinose-conjugates flavonoids rutin and hesperidin by the gut microbiota and bifidobacteria, Nutrients, 7 (4), pp. 2788-2800. [CrossRef]
- Andrews S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Annunziata, G., Sureda, A., Orhan, I. E., Battino, M., Arnone, A., Jiménez-García, M., Capó, X., Cabot, J., Sanadgol, N., Giampieri, F., Tenore, G. C., Kashani, H. R. K., Silva, A. S., Habtemariam, S., Nabavi, S. F. and Nabavi, S. M. (2021) The neuroprotective effects of polyphenols, their role in innate immunity and the interplay with the microbiota, Neuroscience and Biobehavioral Reviews, 128 , pp. 437-453. [CrossRef]
- Asogwa, I. S., Ibrahim, A. N. and Agbaka, J. I. (2021) African baobab: Its role in enhancing nutrition, health, and the environment, Trees, Forests and People (Online), 3, pp. 100043. [CrossRef]
- Assimakopoulos, S. F., Triantos, C., Maroulis, I. and Gogos, C. (2018) The role of the gut barrier function in health and disease, Gastroenterology Research, 11 (4), pp. 261-263. [CrossRef]
- AURA, A., MARTIN-LOPEZ, P., OLEARY, K. A., WILLIAMSON, G., OKSMAN-CALDENTEY, K., POUTANEN, K. and SANTOS-BUELGA, C. (2005) In vitro metabolism of anthocyanins by human gut microflora, European Journal of Nutrition, 44 (3), pp. 133-142. [CrossRef]
- Bang, S., Kim, G., Lim, M. Y., Song, E., Jung, D., Kum, J., Nam, Y., Park, C. and Seo, D. (2018) The influence of in vitro pectin fermentation on the human fecal microbiome, AMB Express, 8 (1), pp. 98. [CrossRef]
- Barber, T. M., Kabisch, S., Pfeiffer, A. F. H. and Weickert, M. O. (2020) The Health Benefits of Dietary Fibre, Nutrients, -10-21, 12 (10). [CrossRef]
- Barbosa, R. S. D. and Vieira-Coelho, M. A. (2020) Probiotics and prebiotics: Focus on psychiatric disorders – a systematic review, Nutrition Reviews, 78 (6), pp. 437-450. [CrossRef]
- Baxter, N. T., Schmidt, A. W., Venkataraman, A., Kim, K. S., Waldron, C. and Schmidt, T. M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers, .
- Beghini, F., Mciver, L. J., Blanco-Míguez, A., Dubois, L., Asnicar, F., Maharjan, S., Mailyan, A., Manghi, P., Scholz, M., Thomas, A. M., Valles-Colomer, M., Weingart, G., Zhang, Y., Zolfo, M., Huttenhower, C., Franzosa, E. A. and Segata, N. (2021) Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery 3, eLife, -05-04, 10 . [CrossRef]
- Belenguer, A., Duncan, S. H., Calder, A. G., Holtrop, G., Louis, P., Lobley, G. E. and Flint, H. J. (2006) Two routes of metabolic cross-feeding between bifidobacterium adolescentis and butyrate-producing anaerobes from the human gut, Applied and Environmental Microbiology, 72 (5), pp. 3593-3599. [CrossRef]
- Berger, K., Burleigh, S., Lindahl, M., Bhattacharya, A., Patil, P., Stålbrand, H., Nordberg Karlsson, E., Hållenius, F., Nyman, M. and Adlercreutz, P. (2021) Xylooligosaccharides Increase Bifidobacteria and Lachnospiraceae in Mice on a High-Fat Diet, with a Concomitant Increase in Short-Chain Fatty Acids, Especially Butyric Acid, Journal of Agricultural and Food Chemistry, 69 (12), pp. 3617. [CrossRef]
- Blatchford, P., Stoklosinski, H., Eady, S., Wallace, A., Butts, C., Gearry, R., Gibson, G. and Ansell, J. (2017) Consumption of kiwifruit capsules increases faecalibacterium prausnitzii abundance in functionally constipated individuals: A randomised controlled human trial, Journal of Nutritional Science, 6 , pp. e52. [CrossRef]
- Bolger, A. M., Lohse, M. and Usadel, B. (2014) Trimmomatic: A flexible trimmer for illumina sequence data, Bioinformatics, 30 (15), pp. 2114-2120. [CrossRef]
- Boto-Ordóñez M, Urpi-Sarda M, Queipo-Ortuño MI, Tulipani S, Tinahones FJ, Andres-Lacueva C. High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: a randomized clinical trial. Food Funct. 2014 Aug;5(8):1932-8. PMID: 24958563. [CrossRef]
- Bouhnik, Y., Raskine, L., Champion, K., Andrieux, C., Penven, S., Jacobs, H. and Simoneau, G. (2007) Prolonged administration of low-dose inulin stimulates the growth of bifidobacteria in humans, Nutrition Research (New York, N.Y.), 27 (4), pp. 187-193. [CrossRef]
- Bravo, L., Abia, R. and Saura-Calixto, F. (1994) Polyphenols as dietary fiber associated compounds. comparative study on in vivo and in vitro properties, Journal of Agricultural and Food Chemistry, 42 (7), pp. 1481-1487. [CrossRef]
- Butler, T. D. and Gibbs, J. E. (2020) Circadian host-microbiome interactions in immunity, Frontiers in Immunology, 11 , pp. 1783. [CrossRef]
- Carlson, J. L., Erickson, J. M., Lloyd, B. B. and Slavin, J. L. (2018) Health effects and sources of prebiotic dietary fiber, Current Developments in Nutrition, 2 (3), pp. nzy005. [CrossRef]
- Carrillo-Cedillo, E. G., Rodríguez-Avila, J. A., Arredondo-Soto, K. C. and Cornejo-Bravo, J. M. (2019) Advantages, Disadvantages, and Future Trends on the Use of Design of Experiments in Cross-Over Trials in Nutritional Clinical Investigation, in: Advantages, Disadvantages, and Future Trends on the Use of Design of Experiments in Cross-Over Trials in Nutritional Clinical Investigation, in: Design of Experiments for Chemical, Pharmaceutical, Food, and Industrial Applications, IGI Global, pp. 158-173. [CrossRef]
- CEEREAL Statement on Dietary Fibre, (2022) -02nd, https://ceereal.eu/images/technical-docs/20220210_Statement_on_FIBRE-FINAL.pdf, Accessed 09/03/2023.
- Chan AO, Leung G, Tong T, Wong NY. Increasing dietary fiber intake in terms of kiwifruit improves constipation in Chinese patients. World J Gastroenterol. 2007 Sep 21;13(35):4771-5. PMID: 17729399; PMCID: PMC4611199. [CrossRef]
- Chang, C., Lin, T., Tsai, Y., Wu, T., Lai, W., Lu, C. and Lai, H. (2019) Next generation probiotics in disease amelioration, Journal of Food and Drug Analysis, 27 (3), pp. 615-622. [CrossRef]
- Chen, Y., Xu, J. and Chen, Y. (2021) Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders, Nutrients, 13 (6), pp. 2099. [CrossRef]
- Clifford, M. N. (2004) Diet-derived phenols in plasma and tissues and their implications for health, Planta Medica, 70 (12), pp. 1103-1114. [CrossRef]
- Colantonio, A. G., Werner, S. L. and Brown, M. (2020) The effects of prebiotics and substances with prebiotic properties on metabolic and inflammatory biomarkers in individuals with type 2 diabetes mellitus: A systematic review, Journal of the Academy of Nutrition and Dietetics, 120 (4), pp. 587-607.e2. [CrossRef]
- Conway de Macario, E. and Macario, A. J. L. (2009) Methanogenic archaea in health and disease: A novel paradigm of microbial pathogenesis, International Journal of Medical Microbiology, 299 (2), pp. 99-108. [CrossRef]
- Cronin, P., Joyce, S. A., O’Toole, P. W. and O’Connor, E. M. (2021) Dietary fibre modulates the gut microbiota, Nutrients, 13 (5), pp. 1655. [CrossRef]
- 20 February; 73, 36. John H Cummings, George T Macfarlane, Hans N Englyst, Prebiotic digestion and fermentation, The American Journal of Clinical Nutrition, Volume 73, Issue 2, February 2001, Pages 415s–420s, https://doi.org/10.1093/ajcn/73.2.415s. [CrossRef]
- Daglia, M. (2012) Polyphenols as antimicrobial agents, Current Opinion in Biotechnology, 23 (2), pp. 174-181. [CrossRef]
- David, L. A., Maurice, C. F., Carmody, R. N., Gootenberg, D. B., Button, J. E., Wolfe, B. E., Ling, A. V., Devlin, A. S., Varma, Y., Fischbach, M. A., Biddinger, S. B., Dutton, R. J. and Turnbaugh, P. J. (2014) Diet rapidly and reproducibly alters the human gut microbiome, Nature, 505 (7484), pp. 559-563. [CrossRef]
- Deehan, E. C., Duar, R. M., Armet, A. M., Perez-Muñoz, M. E., Jin, M. and Walter, J. (2019) Modulation of the Gastrointestinal Microbiome with Nondigestible Fermentable Carbohydrates To Improve Human Health, Microbiology Spectrum, -06-05, 5 (5). [CrossRef]
- Deleu, S., Machiels, K., Raes, J., Verbeke, K. and Vermeire, S. (2021) Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? EBioMedicine, 66 , pp. 103293. [CrossRef]
- den Besten, G., van Eunen, K., Groen, A. K., Venema, K., Reijngoud, D. and Bakker, B. M. (2013) The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism, Journal of Lipid Research, 54 (9), pp. 2325-2340. [CrossRef]
- Desai, V., Kozyrskyj, A. L., Lau, S., Sanni, O., Dennett, L., Walter, J. and Ospina, M. B. (2021) Effectiveness of Probiotic, Prebiotic, and Synbiotic Supplementation to Improve Perinatal Mental Health in Mothers: A Systematic Review and Meta-Analysis, Frontiers in Psychiatry, -04th, 12 . [CrossRef]
- Dinan, T. G. and Cryan, J. F. (2017) Microbes, Immunity, and Behavior: Psychoneuroimmunology Meets the Microbiome, Neuropsychopharmacology, 42 (1), pp. 178. [CrossRef]
- Duncan, S. H., Louis, P., Thomson, J. M. and Flint, H. J. (2009) Role of pH in determining the species composition of the human colonic microbiota, Environmental Microbiology, 11 (8), pp. 2112-2122. [CrossRef]
- DUNN, S., DATTA, A., KALLIS, S., LAW, E., MYERS, C. E. and WHELAN, K. (2011) Validation of a food frequency questionnaire to measure intakes of inulin and oligofructose, European Journal of Clinical Nutrition, 65 (3), pp. 402-408. [CrossRef]
- Durazzi, F., Sala, C., Castellani, G., Manfreda, G., Remondini, D. and De Cesare, A. (2021) Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota, Scientific Reports, 11 (1), pp. 3030. [CrossRef]
- Eberhardt, F., Crichton, M., Dahl, C., Nucera, R., Jenkins, J., Marx, W. and Marshall, S. (2019) Role of dietary fibre in older adults with asymptomatic (AS) or symptomatic uncomplicated diverticular disease (SUDD): Systematic review and meta-analysis, Maturitas, 130 , pp. 57-67. [CrossRef]
- Edwards, C. A., Havlik, J., Cong, W., Mullen, W., Preston, T., Morrison, D. J. and Combet, E. (2017) Polyphenols and health: Interactions between fibre, plant polyphenols and the gut microbiota, Nutrition Bulletin, 42 (4), pp. 356-360. [CrossRef]
- El Mohsen, M. A., Marks, J., Kuhnle, G., Moore, K., Debnam, E., Srai, S. K., Rice-Evans, C. and Spencer, J. P. E. (2006) Absorption, tissue distribution and excretion of pelargonidin and its metabolites following oral administration to rats, British Journal of Nutrition, 95 (1), pp. 51-58. [CrossRef]
- El Oufir, L., Flourié, B., Bruley des Varannes, S., Barry, J. L., Cloarec, D., Bornet, F. and Galmiche, J. P. (1996) Relations between transit time, fermentation products, and hydrogen consuming flora in healthy humans, Gut, 38 (6), pp. 870-877. [CrossRef]
- Falcomer, A. L., Riquette, R. F. R., de Lima, B. R., Ginani, V. C. and Zandonadi, R. P. (2019) Health benefits of green banana consumption: A systematic review, Nutrients, 11 (6), pp. 1222. [CrossRef]
- Falony, G., Vlachou, A., Verbrugghe, K. and Vuyst, L. D. (2006) Cross-feeding between bifidobacterium longum BB536 and acetate-converting, butyrate-producing colon bacteria during growth on oligofructose, Applied and Environmental Microbiology, 72 (12), pp. 7835-7841. [CrossRef]
- Fan, S., Zhang, Z., Zhao, Y., Daglia, M., Zhang, J., Zhu, Y., Bai, J., Zhu, L. and Xiao, X. (2023) Recent advances in targeted manipulation of the gut microbiome by prebiotics: From taxonomic composition to metabolic function, Current Opinion in Food Science, 49 , pp. 100959. [CrossRef]
- Feduraev, P., Skrypnik, L., Riabova, A., Pungin, A., Tokupova, E., Maslennikov, P. and Chupakhina, G. (2020) Phenylalanine and tyrosine as exogenous precursors of wheat (triticum aestivum L.) secondary metabolism through PAL-associated pathways, Plants, 9 (4), pp. 476. [CrossRef]
- Flint, H. J., Duncan, S. H., Scott, K. P. and Louis, P. (2015) Links between diet, gut microbiota composition and gut metabolism, Proceedings of the Nutrition Society, 74 (1), pp. 13-22. [CrossRef]
- Flint, H. J., Scott, K. P., Duncan, S. H., Louis, P. and Forano, E. (2012) Microbial degradation of complex carbohydrates in the gut, Gut Microbes, 3 (4), pp. 289-306. [CrossRef]
- Flowers, S. A., Baxter, N. T., Ward, K. M., Kraal, A. Z., McInnis, M. G., Schmidt, T. M. and Ellingrod, V. L. (2019) Effects of atypical antipsychotic treatment and resistant starch supplementation on gut microbiome composition in a cohort of patients with bipolar disorder or schizophrenia, Pharmacotherapy, 39 (2), pp. 161-170. [CrossRef]
- Foltz, M., Zahradnik, A. C., Van den Abbeele, P., Ghyselinck, J. and Marzorati, M. (2021) A pectin-rich, baobab fruit pulp powder exerts prebiotic potential on the human gut microbiome in vitro, Microorganisms (Basel), 9 (9), pp. 1981. [CrossRef]
- Francisco Javier Álvarez-Martínez, Enrique Barrajón-Catalán, José Antonio Encinar, Juan Carlos Rodríguez-Díaz and Vicente Micol (2020) Antimicrobial capacity of plant polyphenols against gram-positive bacteria: A comprehensive review, Current Medicinal Chemistry, 27 (15), pp. 2576-2606. [CrossRef]
- Garcia-Mazcorro, J. F., Pedreschi, R., Chew, B., Dowd, S. E., Kawas, J. R. and Noratto, G. D. (2018) Dietary supplementation with raspberry extracts modifies the fecal microbiota in obese diabetic db/db mice, Journal of Microbiology and Biotechnology, 28 (8), pp. 1247-1259. [CrossRef]
- Garthoff, J. A., Heemskerk, S., Hempenius, R. A., Lina, B. A. R., Krul, C. A. M., Koeman, J. H. and Speijers, G. J. A. (2010) Safety evaluation of pectin-derived acidic oligosaccharides (pAOS): Genotoxicity and sub-chronic studies, Regulatory Toxicology and Pharmacology, 57 (1), pp. 31-42. [CrossRef]
- Gasaly, N., de Vos, P. and Hermoso, M. A. (2021) Impact of bacterial metabolites on gut barrier function and host immunity: A focus on bacterial metabolism and its relevance for intestinal inflammation, Frontiers in Immunology, 12 , pp. 658354. [CrossRef]
- Ge, L., Qi, J., Shao, B., Ruan, Z., Ren, Y., Sui, S., Wu, X., Sun, X., Liu, S., Li, S., Xu, C. and Song, W. (2022) Microbial hydrogen economy alleviates colitis by reprogramming colonocyte metabolism and reinforcing intestinal barrier, Gut Microbes, -01-13, 14 (1). [CrossRef]
- Ghosh, S. S., Wang, J., Yannie, P. J. and Ghosh, S. (2020) Intestinal barrier dysfunction, LPS translocation, and disease development, Journal of the Endocrine Society, 4 (2), pp. bvz039. [CrossRef]
- Gibson, G. R., Probert, H. M., Loo, J. V., Rastall, R. A. and Roberfroid, M. B. (2004) Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics, Nutrition Research Reviews, 17 (2), pp. 259-275. [CrossRef]
- Gibson, G. R., Hutkins, R., Sanders, M. E., Prescott, S. L., Reimer, R. A., Salminen, S. J., Scott, K., Stanton, C., Swanson, K. S., Cani, P. D., Verbeke, K. and Reid, G. (2017) Expert consensus document: The international scientific association for probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics, Nature Reviews. Gastroenterology & Hepatology, 14 (8), pp. 491-502. [CrossRef]
- González-Aguilar, G. A., Blancas-Benítez, F. J. and Sáyago-Ayerdi, S. G. (2017) Polyphenols associated with dietary fibers in plant foods: Molecular interactions and bioaccessibility, Current Opinion in Food Science, 13 , pp. 84-88. [CrossRef]
- Gråsten, S., Liukkonen, K., Chrevatidis, A., El-Nezami, H., Poutanen, K. and Mykkänen, H. (2003) Effects of wheat pentosan and inulin on the metabolic activity of fecal microbiota and on bowel function in healthy humans, Nutrition Research, -11th, 23 (11), pp. 1503. [CrossRef]
- Guan, J., Liu, F., Zhao, S., Evivie, S. E., Shi, J., Li, N., Zhao, L., Yue, Y., Xie, Q., Huo, G. and Li, B. (2021) Effect of bifidobacterium longum subsp. longum on the proliferative and tight-junction activities of human fetal colon epithelial cells, Journal of Functional Foods, 86 , pp. 104715. [CrossRef]
- Guarino, M. P. L., Altomare, A., Emerenziani, S., Di Rosa, C., Ribolsi, M., Balestrieri, P., Iovino, P., Rocchi, G. and Cicala, M. (2020) Mechanisms of action of prebiotics and their effects on gastro-intestinal disorders in adults, Nutrients, 12 (4), pp. 1037. [CrossRef]
- Guo, C., Cui, Q., Cheng, J., Chen, J., Zhao, Z., Guo, R., Dai, X., Wei, Z. and Li, W. (2021) Probiotic-fermented chinese dwarf cherry [cerasus humilis (bge.) sok.] juice modulates the intestinal mucosal barrier and increases the abundance of akkermansia in the gut in association with polyphenols, Journal of Functional Foods, 80 , pp. 104424. [CrossRef]
- Guo, Y., Xie, J. Y., Deng, K., Li, X. K., Yuan, Y., Xuan, Q., Xie, J. Y., He, X. J., Wang, Q., Li, J. K. and Luo, H. (2019) Prophylactic Effects of Bifidobacterium adolescentis on Anxiety and Depression-Like Phenotypes After Chronic Stress: A Role of the Gut Microbiota-Inflammation Axis, Frontiers in Behavioral Neuroscience, -06-18, 13 . [CrossRef]
- Gwiazdowska, D., Juś, K., Jasnowska-Małecka, J. and Kluczyńska, K. (2015) The impact of polyphenols on bifidobacterium growth, Acta Biochimica Polonica, 62 (4), pp. 895-901. [CrossRef]
- Han, K. S., Balan, P., Molist Gasa, F. and Boland, M. (2011) Green kiwifruit modulates the colonic microbiota in growing pigs, Letters in Applied Microbiology, 52 (4), pp. 379-385. [CrossRef]
- He, Q., Si, C., Sun, Z., Chen, Y. and Zhang, X. (2022) The intervention of prebiotics on depression via the Gut–Brain axis, Molecules (Basel, Switzerland), 27 (12), pp. 3671. [CrossRef]
- Healey, G., Murphy, R., Butts, C., Brough, L., Whelan, K. and Coad, J. (2018) Habitual dietary fibre intake influences gut microbiota response to an inulin-type fructan prebiotic: A randomised, double-blind, placebo-controlled, cross-over, human intervention study, British Journal of Nutrition, 119 (2), pp. 176-189. [CrossRef]
- Hiel, S., Bindels, L. B., Pachikian, B. D., Kalala, G., Broers, V., Zamariola, G., Chang, B. P., Kambashi, B., Rodriguez, J., Cani, P. D., Neyrinck, A. M., Thissen, J., Luminet, O., Bindelle, J. and Delzenne, N. M. (2019) Effects of a diet based on inulin-rich vegetables on gut health and nutritional behavior in healthy humans, The American Journal of Clinical Nutrition, 109 (6), pp. 1683. [CrossRef]
- Hill, C. (2012) Virulence or niche factors: What's in a name? Journal of Bacteriology, 194 (21), pp. 5725-5727. [CrossRef]
- Ho, J., Nicolucci, A. C., Virtanen, H., Schick, A., Meddings, J., Reimer, R. A. and Huang, C. (2019) Effect of prebiotic on microbiota, intestinal permeability, and glycemic control in children with type 1 diabetes, The Journal of Clinical Endocrinology and Metabolism, 104 (10), pp. 4427-4440. [CrossRef]
- Holscher, H. D., Bauer, L. L., Gourineni, V., Pelkman, C. L., Fahey, J., George C and Swanson, K. S. (2015) Agave inulin supplementation affects the fecal microbiota of healthy adults participating in a randomized, double-blind, placebo-controlled, crossover trial, The Journal of Nutrition, 145 (9), pp. 2025-2032. [CrossRef]
- Hotchkiss, A. T., Olano-Martin, E., Grace, W. E., Gibson, G. R. and Rastall, R. A. (20032-04-204) Pectic Oligosaccharides as Prebiotics, ACS Symposium Series, 20032-04-204, , pp. 54. [CrossRef]
- Hughes, R. L., Alvarado, D. A., Swanson, K. S. and Holscher, H. D. (2022) The prebiotic potential of inulin-type fructans: A systematic review, Advances in Nutrition (Bethesda, Md.), 13 (2), pp. 492-529. [CrossRef]
- Hughes, R. L., Marco, M. L., Hughes, J. P., Keim, N. L. and Kable, M. E. (2019) The role of the gut microbiome in predicting response to diet and the development of precision nutrition Models—Part I: Overview of current methods, Advances in Nutrition (Bethesda, Md.), 10 (6), pp. 953-978. [CrossRef]
- Ioniță-Mîndrican, C., Ziani, K., Mititelu, M., Oprea, E., Neacșu, S. M., Moroșan, E., Dumitrescu, D., Roșca, A. C., Drăgănescu, D. and Negrei, C. (2022) Therapeutic benefits and dietary restrictions of fiber intake: A state of the art review, Nutrients, 14 (13), pp. 2641. [CrossRef]
- Islam, T., Albracht-Schulte, K., Ramalingam, L., Schlabritz-Lutsevich, N., Park, O., Zabet-Moghaddam, M., Kalupahana, N. S. and Moustaid-Moussa, N. (2023) Anti-inflammatory mechanisms of polyphenols in adipose tissue: Role of gut microbiota, intestinal barrier integrity and zinc homeostasis, The Journal of Nutritional Biochemistry, 115 , pp. 109242. [CrossRef]
- Jackson, P. P. J., Wijeyesekera, A. and Rastall, R. A. (2023) Inulin-type fructans and short-chain fructooligosaccharides—their role within the food industry as fat and sugar replacers and texture modifiers—what needs to be considered, Food Science & Nutrition, 11 (1), pp. 17-38. [CrossRef]
- Jangid, A., Fukuda, S., Suzuki, Y., Taylor, T. D., Ohno, H. and Prakash, T. (123456789) Shotgun metagenomic sequencing revealed the prebiotic potential of a grain-based diet in mice, Scientific Reports, 123456789th, 12 (1). [CrossRef]
- Jeong, J., Yun, K., Mun, S., Chung, W., Choi, S., Nam, Y., Lim, M. Y., Hong, C. P., Park, C., Ahn, Y. J. and Han, K. (123456789) The effect of taxonomic classification by full-length 16S rRNA sequencing with a synthetic long-read technology, Scientific Reports, 123456789th, 11 (1). [CrossRef]
- Jiang, R., Zhao, S., Wang, R., Feng, H., Zhang, J., Li, X., Mao, Y., Yuan, X., Fei, Z., Zhao, Y., Yu, X., Poon, W. S., Zhu, X., Liu, N., Kang, D., Sun, T., Jiao, B., Liu, X., Yu, R., Zhang, J., Gao, G., Hao, J., Su, N., Yin, G., Zhu, X., Lu, Y., Wei, J., Hu, J., Hu, R., Li, J., Wang, D., Wei, H., Tian, Y., Lei, P., Dong, J. and Zhang, J. (2018) Safety and efficacy of atorvastatin for chronic subdural hematoma in chinese patients: A randomized ClinicalTrial, JAMA Neurology, 75 (11), pp. 1338-1346. [CrossRef]
- Johnstone, N., Milesi, C., Burn, O., Van Den Bogert, B., Nauta, A., Hart, K., Sowden, P., Burnet, P. W. J. and Cohen Kadosh, K. (123456789) Anxiolytic effects of a galacto-oligosaccharides prebiotic in healthy females (18–25 years) with corresponding changes in gut bacterial composition, Scientific Reports, 123456789th, 11 (1). [CrossRef]
- Joossens, M., Huys, G., Van Steen, K., Cnockaert, M., Vermeire, S., Rutgeerts, P., Verbeke, K., Vandamme, P. and De Preter, V. (2011) High-throughput method for comparative analysis of denaturing gradient gel electrophoresis profiles from human fecal samples reveals significant increases in two bifidobacterial species after inulin-type prebiotic intake, FEMS Microbiology Ecology, 75 (2), pp. 343-349. [CrossRef]
- Junghare, M., Spiteller, D. and Schink, B. (2019) Anaerobic degradation of xenobiotic isophthalate by the fermenting bacterium syntrophorhabdus aromaticivorans, The ISME Journal, 13 (5), pp. 1252-1268. [CrossRef]
- Kabbashi, N. A., Mirghani, M. E. S. and Bello, I. (2017) Characterization of the Baobab fruit shells as adsorption material, -12th, .
- Kanehisa, M., Furumichi, M., Tanabe, M., Sato, Y. and Morishima, K. (2017) KEGG: New perspectives on genomes, pathways, diseases and drugs, Nucleic Acids Research, 45 (D1), pp. D353-D361. [CrossRef]
- Kang, J. W., Tang, X., Walton, C. J., Brown, M. J., Brewer, R. A., Maddela, R. L., Zheng, J. J., Agus, J. K. and Zivkovic, A. M. (2022) Multi-Omic Analyses Reveal Bifidogenic Effect and Metabolomic Shifts in Healthy Human Cohort Supplemented With a Prebiotic Dietary Fiber Blend. Frontiers Media SA.
- Kawabata, K., Baba, N., Sakano, T., Hamano, Y., Taira, S., Tamura, A., Baba, S., Natsume, M., Ishii, T., Murakami, S. and Ohigashi, H. (2018) Functional properties of anti-inflammatory substances from quercetin-treated bifidobacterium adolescentis, Bioscience, Biotechnology, and Biochemistry, 82 (4), pp. 689-697. [CrossRef]
- Kelly, E. D., Bottacini, F.,Callaghan, J., Motherway, M. O.,Connell, K. J., Stanton, C. and van Sinderen, D. (2016) Glycoside hydrolase family 13 α-glucosidases encoded by bifidobacterium breve UCC2003; A comparative analysis of function, structure and phylogeny, International Journal of Food Microbiology, 224 , pp. 55-65. [CrossRef]
- Khorasaniha, R., Olof, H., Voisin, A., Armstrong, K., Wine, E., Vasanthan, T. and Armstrong, H. (2023) Diversity of fibers in common foods: Key to advancing dietary research, Food Hydrocolloids, 139 , pp. 108495. [CrossRef]
- Kim, Y., Je, Y. and Giovannucci, E. L. (2021) Association between dietary fat intake and mortality from all-causes, cardiovascular disease, and cancer: A systematic review and meta-analysis of prospective cohort studies, Clinical Nutrition (Edinburgh, Scotland), 40 (3), pp. 1060-1070. [CrossRef]
- Kleessen, B., Hartmann, L. and Blaut, M. (2001) Oligofructose and long-chain inulin: Influence on the gut microbial ecology of rats associated with a human faecal flora, British Journal of Nutrition, 86 (2), pp. 291-300. [CrossRef]
- Kleessen, B., Schwarz, S., Boehm, A., Fuhrmann, H., Richter, A., Henle, T. and Krueger, M. (2007) Jerusalem artichoke and chicory inulin in bakery products affect faecal microbiota of healthy volunteers, British Journal of Nutrition, 98 (3), pp. 540-549. [CrossRef]
- Klimenko, N. S., Odintsova, V. E., Revel-Muroz, A. and Tyakht, A. V. (2022) The hallmarks of dietary intervention-resilient gut microbiome, NPJ Biofilms and Microbiomes, 8 (1), pp. 77. [CrossRef]
- Kolida, S., Meyer, D. and Gibson, G. R. (2007) Double-blind placebo-controlled study to establish the bifidogenic dose of inulin in healthy humans, European Journal of Clinical Nutrition, 61 (10), pp. 1189-1195. [CrossRef]
- Kumarihami, H. M. P. C., Kim, J. G., Kim, Y. G., Lee, M., Lee, Y., Kwack, Y. and Kim, J. G. (2021) Preharvest Application of Chitosan Improves the Postharvest Life of ‘Garmrok’ Kiwifruit through the Modulation of Genes Related to Ethylene Biosynthesis, Cell Wall Modification and Lignin Metabolism, Foods, -02-09, 10 (2). [CrossRef]
- Le Bastard, Q., Chapelet, G., Javaudin, F., Lepelletier, D., Batard, E. and Montassier, E. (2020) The effects of inulin on gut microbial composition: A systematic review of evidence from human studies, European Journal of Clinical Microbiology & Infectious Diseases, 39 (3), pp. 403-413. [CrossRef]
- Le Gall, G., Guttula, K., Kellingray, L., Tett, A. J., Ten Hoopen, R., Kemsley, E. K., Savva, G. M., Ibrahim, A. and Narbad, A. (2018) Metabolite quantification of faecal extracts from colorectal cancer patients and healthy controls, Oncotarget, 9 (70), pp. 33278-33289. [CrossRef]
- Leeming, E. R., Johnson, A. J., Spector, T. D. and Le Roy, C. I. (2019) Effect of diet on the gut microbiota: Rethinking intervention duration, Nutrients, 11 (12), pp. 2862. [CrossRef]
- Lewis, S.J. and K.W., Heaton (1997) Stool From Scale as a Useful guide to Intestinal Transit Time, Scandinavian Journal of Gastroenterology, 32:9, 920-924. [CrossRef]
- Li, P., Li, M., Song, Y., Huang, X., Wu, T., Xu, Z. Z. and Lu, H. (2022) Green banana flour contributes to gut microbiota recovery and improves colonic barrier integrity in mice following antibiotic perturbation, Frontiers in Nutrition (Lausanne), 9 , pp. 832848. [CrossRef]
- Lin, H. and Peddada, S. D. (2020) Analysis of microbial compositions: A review of normalization and differential abundance analysis, NPJ Biofilms and Microbiomes, 6 (1), pp. 60. [CrossRef]
- Liu, F., Prabhakar, M., Ju, J., Long, H. and Zhou, H. (2017) Effect of inulin-type fructans on blood lipid profile and glucose level: a systematic review and meta-analysis of randomized controlled trials, European Journal of Clinical Nutrition, 71 (1), pp. 9. [CrossRef]
- Liu, F., Li, P., Chen, M., Luo, Y., Prabhakar, M., Zheng, H., He, Y., Qi, Q., Long, H., Zhang, Y., Sheng, H. and Zhou, H. (2017) Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population, Scientific Reports, -09-18, 7 (1). [CrossRef]
- Liu, R. T., Walsh, R. F. L. and Sheehan, A. E. (2019) Prebiotics and probiotics for depression and anxiety: A systematic review and meta-analysis of controlled clinical trials, Neuroscience and Biobehavioral Reviews, 102 , pp. 13-23. [CrossRef]
- Livingston, K. A., Chung, M., Sawicki, C. M., Lyle, B. J., Wang, D. D., Roberts, S. B. and Mckeown, N. M. (2016) Development of a Publicly Available, Comprehensive Database of Fiber and Health Outcomes: Rationale and Methods, Plos One, -06-27, 11 (6). [CrossRef]
- LoCascio, R. G., Desai, P., Sela, D. A., Weimer, B. and Mills, D. A. (2010) Broad conservation of milk utilization genes in bifidobacterium longum subsp. infantis as revealed by comparative genomic hybridization, Applied and Environmental Microbiology, 76 (22), pp. 7373-7381. [CrossRef]
- Louis, P. and Flint, H. J. (2017) Formation of propionate and butyrate by the human colonic microbiota, Environmental Microbiology, 19 (1), pp. 29-41. [CrossRef]
- Lovibond, P. F. and Lovibond, S. H. (1995) The structure of negative emotional states: Comparison of the Depression Anxiety Stress Scales (DASS) with the Beck Depression and Anxiety Inventories, Behaviour Research and Therapy, 33 (3), pp. 335. [CrossRef]
- Lozupone, C. A., Hamady, M., Cantarel, B. L., Coutinho, P. M., Henrissat, B., Gordon, J. I. and Knight, R. (2008) Convergence of carbohydrate active gene repertoires in human gut microbes, Proceedings of the National Academy of Sciences - PNAS, 105 (39), pp. 15076-15081. [CrossRef]
- Lund, P., Tramonti, A. and De Biase, D. (2014) Coping with low pH: Molecular strategies in neutralophilic bacteria, FEMS Microbiology Reviews, 38 (6), pp. 1091-1125. [CrossRef]
- Ma, X., Liu, S., Wang, H., Wang, Y., Li, Z., Gu, T., Li, Y., Xin, F. and Wen, B. (2023) In vitro fermentation of beechwood lignin-carbohydrate complexes provides evidence for utilization by gut bacteria, Nutrients, 15 (1), pp. 220. [CrossRef]
- Mailing, L. J., Allen, J. M., Pence, B. D., Rytych, J., Sun, Y., Bhattacharya, T. K., Park, P., Cross, T. L., McCusker, R. H., Swanson, K. S., Fahey, G. C., Rhodes, J. S., Kelley, K. W., Johnson, R. W. and Woods, J. A. (2019) Behavioral response to fiber feeding is cohort-dependent and associated with gut microbiota composition in mice, Behavioural Brain Research, 359 , pp. 731-736. [CrossRef]
- Maiuolo, J., Gliozzi, M., Musolino, V., Carresi, C., Scarano, F., Nucera, S., Scicchitano, M., Oppedisano, F., Bosco, F., Ruga, S., Zito, M. C., Macri, R., Palma, E., Muscoli, C. and Mollace, V. (2021) The Contribution of Gut Microbiota–Brain Axis in the Development of Brain Disorders, Frontiers in Neuroscience, -03-23, 15 . [CrossRef]
- Makarewicz, M., Drożdż, I., Tarko, T. and Duda-Chodak, A. (2021) The interactions between polyphenols and microorganisms, especially gut microbiota, Antioxidants, 10 (2), pp. 188. [CrossRef]
- Makki, K., Deehan, E. C., Walter, J. and Bäckhed, F. (2018) The impact of dietary fiber on gut microbiota in host health and disease, Cell Host & Microbe, 23 (6), pp. 705-715. [CrossRef]
- Manderson, K., Pinart, M., Tuohy, K. M., Grace, W. E., Hotchkiss, A. T., Widmer, W., Yadhav, M. P., Gibson, G. R. and Rastall, R. A. (2005) In vitro determination of prebiotic properties of oligosaccharides derived from an orange juice manufacturing by-product stream, Applied and Environmental Microbiology, 71 (12), pp. 8383-8389. [CrossRef]
- Matsumoto, M., Ohishi, H. and Benno, Y. (2004) H +-ATPase activity in bifidobacterium with special reference to acid tolerance, International Journal of Food Microbiology, 93 (1), pp. 109-113. [CrossRef]
- Mazur, S. P., Nes, A., Wold, A., Remberg, S. F. and Aaby, K. (2014) Quality and chemical composition of ten red raspberry (rubus idaeus L.) genotypes during three harvest seasons, Food Chemistry, 160 , pp. 233-240. [CrossRef]
- McMurdie, P. J. and Holmes, S. (2013) Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data, PLoS ONE, 8 (4), pp. e61217. [CrossRef]
- Metges, C. C. (2000) Contribution of microbial amino acids to amino acid homeostasis of the host, The Journal of Nutrition, 130 (7), pp. 1857S-1864S. [CrossRef]
- Míguez, B. (2016) Pectic Oligosaccharides and Other Emerging Prebiotics, in: Pectic Oligosaccharides and Other Emerging Prebiotics, in: IntechOpen. [CrossRef]
- Moens, F., Verce, M. and De Vuyst, L. (2017) Lactate- and acetate-based cross-feeding interactions between selected strains of lactobacilli, bifidobacteria and colon bacteria in the presence of inulin-type fructans, International Journal of Food Microbiology, 241 , pp. 225-236. [CrossRef]
- Mora, D. and Arioli, S. (2014) Microbial urease in health and disease, PLoS Pathogens, 10 (12), pp. e1004472. [CrossRef]
- Moshfegh, A. J., Friday, J. E., Goldman, J. P. and Chug Ahuja, J. K. (1999) Presence of inulin and oligofructose in the diets of americans, The Journal of Nutrition, 129 (7S), pp. 1407-1411. [CrossRef]
- Mukherji, A., Kobiita, A., Ye, T. and Chambon, P. (2013) Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by TLRs, Cell, 153 (4), pp. 812-827. [CrossRef]
- Murakami, M., Tognini, P., Liu, Y., Eckel-Mahan, K. L., Baldi, P. and Sassone-Corsi, P. (2016) Gut microbiota directs PPARγ-driven reprogramming of the liver circadian clock by nutritional challenge, EMBO Reports, 17 (9), pp. 1292-1303. [CrossRef]
- Mussatto, S. I. and Mancilha, I. M. (2007) Non-digestible oligosaccharides: A review, Carbohydrate Polymers, 68 (3), pp. 587-597. [CrossRef]
- Myhrstad, M. C. W., Tunsjø, H., Charnock, C. and Telle-Hansen, V. H. (2020) Dietary fiber, gut microbiota, and metabolic Regulation—Current status in human randomized trials, Nutrients, 12 (3), pp. 859. [CrossRef]
- Nakamura, A., Kurihara, S., Takahashi, D., Ohashi, W., Nakamura, Y., Kimura, S., Onuki, M., Kume, A., Sasazawa, Y., Furusawa, Y., Obata, Y., Fukuda, S., Saiki, S., Matsumoto, M. and Hase, K. (2021) Symbiotic polyamine metabolism regulates epithelial proliferation and macrophage differentiation in the colon, Nature Communications, -04-08, 12 (1). [CrossRef]
- NDNS (2016. https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/943114/NDNS_UK_Y9-11_report.pdf, Accessed 9/03/2023.
- Neri-Numa, I. A. and Pastore, G. M. (2020) Novel insights into prebiotic properties on human health: A review, Food Research International, 131 , pp. 108973. [CrossRef]
- Neyrinck, A. M., Nazare, J., Rodriguez, J., Jottard, R., Dib, S., Sothier, M., Berghe, L. V. D., Alligier, M., Alexiou, H., Maquet, V., Vinoy, S., Bischoff, S. C., Walter, J., Laville, M. and Delzenne, N. M. (2020) Development of a Repertoire and a Food Frequency Questionnaire for Estimating Dietary Fiber Intake Considering Prebiotics: Input from the FiberTAG Project, Nutrients, 12 (9). [CrossRef]
- Niemi, Pritta, Anna-Marja Aura, Johanna Maukonen, Annika I. Smeds, Ismo Mattila, Klaus Niemelä, Tarja Tamminen, Craig B. Faulds, Johanna Buchert, and Kaisa Poutanen (2013) Interactions of a Lignin-Rich Fraction from Brewer’s Spent Grain with Gut Microbiota in Vitro, Journal of Agricultural and Food Chemistry, 61 (27), 6754-6762. [CrossRef]
- Nikolaieva, N., Sevcikova, A., Omelka, R., Martiniakova, M., Mego, M. and Ciernikova, S. (2022) Gut microbiota-MicroRNA interactions in intestinal homeostasis and cancer development, Microorganisms (Basel), 11 (1), pp. 107. [CrossRef]
- Novakovic, M., Rout, A., Kingsley, T., Kirchoff, R., Singh, A., Verma, V., Kant, R. and Chaudhary, R. (2020) Role of gut microbiota in cardiovascular diseases, World Journal of Cardiology, 12 (4), pp. 110-122. [CrossRef]
- Núñez-Gómez, V., Periago, M. J., Navarro-González, I., Campos-Cava, M. P., Baenas, N. and González-Barrio, R. (2021) Influence of raspberry and its dietary fractions on the in vitro activity of the colonic microbiota from normal and overweight subjects, Plant Foods for Human Nutrition (Dordrecht), 76 (4), pp. 494-500. [CrossRef]
- O’Keefe, S. J. D. (2018) The need to reassess dietary fiber requirements in healthy and critically ill patients, Gastroenterology Clinics of North America, 47 (1), pp. 219-229. [CrossRef]
- Oksanen J, Blanchet GF, Kindt R, Legendre P, Minchin PR, B. O’Hara R, et al. Community Ecology Package: Ordination, Diversity and Dissimilarities. https://CRANR-project.org/package=vegan. 2022. Available: https://CRAN.R-project.org/package=vegan.
- Ozdal, T., Sela, D. A., Xiao, J., Boyacioglu, D., Chen, F. and Capanoglu, E. (2016) The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility, Nutrients, 8 (2), pp. 78. [CrossRef]
- Parkar, S. G., Rosendale, D., Paturi, G., Herath, T. D., Stoklosinski, H., Phipps, J. E., Hedderley, D. and Ansell, J. (2012) In vitro utilization of gold and green kiwifruit oligosaccharides by human gut microbial populations, Plant Foods for Human Nutrition (Dordrecht), 67 (3), pp. 200-207. [CrossRef]
- Parks, D. H., Chuvochina, M., Rinke, C., Mussig, A. J., Chaumeil, P. and Hugenholtz, P. (2022) GTDB: An ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy, Nucleic Acids Research, 50 (D1), pp. D785-D794. [CrossRef]
- Pascale, N., Gu, F., Larsen, N., Jespersen, L. and Respondek, F. (2022) The potential of pectins to modulate the human gut microbiota evaluated by in vitro fermentation: A systematic review, Nutrients, 14 (17), pp. 3629. [CrossRef]
- Peron, G., Gargari, G., Meroño, T., Miñarro, A., Lozano, E. V., Escuder, P. C., González-Domínguez, R., Hidalgo-Liberona, N., Del Bo’, C., Bernardi, S., Kroon, P. A., Carrieri, B., Cherubini, A., Riso, P., Guglielmetti, S. and Andrés-Lacueva, C. (2021) Crosstalk among intestinal barrier, gut microbiota and serum metabolome after a polyphenol-rich diet in older subjects with “leaky gut”: The MaPLE trial, Clinical Nutrition (Edinburgh, Scotland), 40 (10), pp. 5288-5297. [CrossRef]
- Popoola-Akinola, O. O., Raji, T. J. and Olawoye, B. (2022) Lignocellulose, dietary fibre, inulin and their potential application in food, Heliyon, 8 (8), pp. e10459. [CrossRef]
- Qi, X., Yun, C., Pang, Y. and Qiao, J. (2021) The impact of the gut microbiota on the reproductive and metabolic endocrine system, Gut Microbes, -01-01, 13 (1). [CrossRef]
- Qian, X., Si, Q., Lin, G., Zhu, M., Lu, J., Zhang, H., Wang, G. and Chen, W. (2022) Bifidobacterium adolescentis is effective in relieving type 2 diabetes and may be related to its dominant core genome and gut microbiota modulation capacity, Nutrients, 14 (12), pp. 2479. [CrossRef]
- Ramirez-Farias, C., Slezak, K., Fuller, Z., Duncan, A., Holtrop, G. and Louis, P. (2009) Effect of inulin on the human gut microbiota: Stimulation of bifidobacterium adolescentis and faecalibacterium prausnitzii, British Journal of Nutrition, 101 (4), pp. 541-550. [CrossRef]
- Rao, S. S. C., Yu, S. and Fedewa, A. (2015) Systematic review: Dietary fibre and FODMAP-restricted diet in the management of constipation and irritable bowel syndrome, Alimentary Pharmacology & Therapeutics, 41 (12), pp. 1256-1270. [CrossRef]
- Rao, J. N., Xiao, L. and Wang, J. (2020) Polyamines in Gut Epithelial Renewal and Barrier Function, Physiology, -09-01, 35 (5), pp. 328. [CrossRef]
- Regan, M., Chiang, E., Liu, Y., Tonelli, M., Verdoorn, K., Gugel, S., Suen, G., Carey, H. and Assadi-Porter, F. (2021) Urea nitrogen recycling via gut symbionts increases in hibernators over the winter fast. Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
- Ren, W., Chen, S., Yin, J., Duan, J., Li, T., Liu, G., Feng, Z., Tan, B., Yin, Y. and Wu, G. (2014) Dietary arginine supplementation of mice alters the microbial population and activates intestinal innate immunity, The Journal of Nutrition, 144 (6), pp. 988-995. [CrossRef]
- Riva, A., Kolimár, D., Spittler, A., Wisgrill, L., Herbold, C. W., Abrankó, L. and Berry, D. (2020) Conversion of Rutin, a Prevalent Dietary Flavonol, by the Human Gut Microbiota, Frontiers in Microbiology, -12-21, 11 . [CrossRef]
- Rivière, A., Gagnon, M., Weckx, S., Roy, D. and De Vuyst, L. (2015) Mutual cross-feeding interactions between bifidobacterium longum subsp. longum NCC2705 and eubacterium rectale ATCC 33656 explain the bifidogenic and butyrogenic effects of arabinoxylan oligosaccharides, Applied and Environmental Microbiology, 81 (22), pp. 7767-7781. [CrossRef]
- Rivière, A., Selak, M., Lantin, D., Leroy, F. and De Vuyst, L. (2016) Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut, Frontiers in Microbiology, 7 , pp. 979. [CrossRef]
- Rodríguez-Daza, M. C., Pulido-Mateos, E. C., Lupien-Meilleur, J., Guyonnet, D., Desjardins, Y. and Roy, D. (2021) Polyphenol-mediated gut microbiota modulation: Toward prebiotics and further, Frontiers in Nutrition (Lausanne), 8 , pp. 689456. [CrossRef]
- Rodríguez-Daza, M., Roquim, M., Dudonné, S., Pilon, G., Levy, E., Marette, A., Roy, D. and Desjardins, Y. (2020) Berry polyphenols and fibers modulate distinct microbial metabolic functions and gut microbiota enterotype-like clustering in obese mice, Frontiers in Microbiology, 11 , pp. 2032. [CrossRef]
- Rößle, C., Ktenioudaki, A. and Gallagher, E. (2011) Inulin and oligofructose as fat and sugar substitutes in quick breads (scones): A mixture design approach, European Food Research & Technology, 233 (1), pp. 167-181. [CrossRef]
- Rowland, I., Gibson, G., Heinken, A., Scott, K., Swann, J., Thiele, I. and Tuohy, K. (2018) Gut microbiota functions: Metabolism of nutrients and other food components, European Journal of Nutrition, 57 (1), pp. 1-24. [CrossRef]
- Rush, E. C., Patel, M., Plank, L. D. and Ferguson, L. R. (2002) Kiwifruit promotes laxation in the elderly, Asia Pacific Journal of Clinical Nutrition, 11 (2), pp. 164-168. [CrossRef]
- Rutherford, J. C. (2014) The emerging role of urease as a general microbial virulence factor, PLoS Pathogens, 10 (5), pp. e1004062. [CrossRef]
- Ryvchin, R., Dubinsky, V., Rabinowitz, K., Wasserberg, N., Dotan, I. and Gophna, U. (2021) Alteration in urease-producing bacteria in the gut microbiomes of patients with inflammatory bowel diseases, Journal of Crohn's and Colitis, 15 (12), pp. 2066-2077. [CrossRef]
- Salazar, N., Dewulf, E. M., Neyrinck, A. M., Bindels, L. B., Cani, P. D., Mahillon, J., de Vos, W. M., Thissen, J., Gueimonde, M., de los Reyes-Gavilán, C. G. and Delzenne, N. M. (2015) Inulin-type fructans modulate intestinal bifidobacterium species populations and decrease fecal short-chain fatty acids in obese women, Clinical Nutrition, 34 (3), pp. 501-507. [CrossRef]
- Salonen, A., Lahti, L. M., Salojärvi, J., Holtrop, G., Korpela, K., Duncan, S. H., Date, P., Farquharson, F., Johnstone, A. M., Lobley, G. E., Louis, P., Flint, H. J. and de Vos, W. M. (2014) Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men, The ISME Journal, 8 (11), pp. 2218-2230. [CrossRef]
- Sawicki, C. M., Livingston, K. A., Obin, M., Roberts, S. B., Chung, M. and Mckeown, N. M. (2017) Dietary Fiber and the Human Gut Microbiota: Application of Evidence Mapping Methodology, Nutrients, -02-10, 9 (2). [CrossRef]
- Schmidt, K., Cowen, P. J., Harmer, C. J., Tzortzis, G., Errington, S. and Burnet, P. W. J. (2015) Prebiotic intake reduces the waking cortisol response and alters emotional bias in healthy volunteers, Psychopharmacology, 232 (10), pp. 1793-1801. [CrossRef]
- Schwab, C., Ruscheweyh, H., Bunesova, V., Pham, V. T., Beerenwinkel, N. and Lacroix, C. (2017) Trophic interactions of infant bifidobacteria and eubacterium hallii during L-fucose and fucosyllactose degradation, Frontiers in Microbiology, 8 , pp. 95. [CrossRef]
- Scott, K. P., Martin, J. C., Chassard, C., Clerget, M., Potrykus, J., Campbell, G., Mayer, C., Young, P., Rucklidge, G., Ramsay, A. G. and Flint, H. J. (2011) Substrate-driven gene expression in roseburia inulinivorans: Importance of inducible enzymes in the utilization of inulin and starch, Proceedings of the National Academy of Sciences - PNAS, 108 (Supplement 1), pp. 4672-4679. [CrossRef]
- Sealed Envelope Ltd (2022) Simple randomisation service. [Online] Available from: https://www.sealedenvelope.com/simple-randomiser/v1. Accessed 27.02.2023.
- Sender, R., Fuchs, S. and Milo, R. (2016) Revised estimates for the number of human and bacteria cells in the body, PLoS Biology, 14 (8), pp. e1002533. [CrossRef]
- Sheridan, P., Louis, P., Tsompanidou, E., Shaw, S., Harmsen, H., Duncan, S., Flint, H. and Walker, A. (2021) Distribution, organization and expression of genes concerned with anaerobic lactate-utilization in human intestinal bacteria. Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
- Shetty, S. A., Kuipers, B., Atashgahi, S., Aalvink, S., Smidt, H. and Vos, W. M. d. (2022) Inter-species metabolic interactions in an in-vitro minimal human gut microbiome of core bacteria, NPJ Biofilms and Microbiomes, 8 (1), pp. 1-13. [CrossRef]
- Shinde, T., Perera, A. P., Vemuri, R., Gondalia, S. V., Beale, D. J., Karpe, A. V., Shastri, S., Basheer, W., Southam, B., Eri, R. and Stanley, R. (2020) Synbiotic supplementation with prebiotic green banana resistant starch and probiotic bacillus coagulans spores ameliorates gut inflammation in mouse model of inflammatory bowel diseases, European Journal of Nutrition, 59 (8), pp. 3669-3689. [CrossRef]
- Silva, M., Cueva, C., Alba, C., Rodriguez, J. M., de Pascual-Teresa, S., Jones, J., Caturla, N., Victoria Moreno-Arribas, M. and Bartolomé, B. (2022) Gut microbiome-modulating properties of a polyphenol-enriched dietary supplement comprised of hibiscus and lemon verbena extracts. monitoring of phenolic metabolites, Journal of Functional Foods, 91 , pp. 105016. [CrossRef]
- Silva, Y. P., Bernardi, A. and Frozza, R. L. (2020) The role of short-chain fatty acids from gut microbiota in gut-brain communication, Frontiers in Endocrinology, 11 , pp. 25. [CrossRef]
- Simpson (1949) Measurements of diversity, Nature, Vol. 163.
- Singh, R. P. (2019) Glycan utilisation system in bacteroides and bifidobacteria and their roles in gut stability and health, Applied Microbiology and Biotechnology, 103 (18), pp. 7287-7315. [CrossRef]
- Singhal, R., Donde, H., Ghare, S., Stocke, K., Zhang, J., Vadhanam, M., Reddy, S., Gobejishvili, L., Chilton, P., Joshi-Barve, S., Feng, W., Mcclain, C., Hoffman, K., Petrosino, J., Vital, M. and Barve, S. (2021) Decrease in acetyl-CoA pathway utilizing butyrate-producing bacteria is a key pathogenic feature of alcohol-induced functional gut microbial dysbiosis and development of liver disease in mice, Gut Microbes, -01-01, 13 (1). [CrossRef]
- So, D., Whelan, K., Rossi, M., Morrison, M., Holtmann, G., Kelly, J. T., Shanahan, E. R., Staudacher, H. M. and Campbell, K. L. Dietary fiber intervention on gut microbiota composition in healthy adults: a systematic review and meta-analysis, . [CrossRef]
- Srour, B., Fezeu, L. K., Kesse-Guyot, E., Allès, B., Méjean, C., Andrianasolo, R. M., Chazelas, E., Deschasaux, M., Hercberg, S., Galan, P., Monteiro, C. A., Julia, C. and Touvier, M. (2019) Ultra-processed food intake and risk of cardiovascular disease: prospective cohort study (NutriNet-Santé), Bmj, -05-29, . [CrossRef]
- Staudacher, H. M., Irving, P. M., Lomer, M. C. E. and Whelan, K. (2017) The challenges of control groups, placebos and blinding in clinical trials of dietary interventions, Proceedings of the Nutrition Society, 76 (3), pp. 203-212. [CrossRef]
- Stephen, A. M., Champ, M. M. -., Cloran, S. J., Fleith, M., van Lieshout, L., Mejborn, H. and Burley, V. J. (2017) Dietary fibre in europe: Current state of knowledge on definitions, sources, recommendations, intakes and relationships to health, Nutrition Research Reviews, 30 (2), pp. 149-190. [CrossRef]
- Suzek, B. E., Wang, Y., Huang, H., McGarvey, P. B. and Wu, C. H. (2015) UniRef clusters: A comprehensive and scalable alternative for improving sequence similarity searches, Bioinformatics, 31 (6), pp. 926-932. [CrossRef]
- Swann, J. R., Rajilic-Stojanovic, M., Salonen, A., Sakwinska, O., Gill, C., Meynier, A., Fança-Berthon, P., Schelkle, B., Segata, N., Shortt, C., Tuohy, K. and Hasselwander, O. (2020) Considerations for the design and conduct of human gut microbiota intervention studies relating to foods, European Journal of Nutrition, 59 (8), pp. 3347-3368. [CrossRef]
- Swanson, K. S., de Vos, W. M., Martens, E. C., Gilbert, J. A., Menon, R. S., Soto-Vaca, A., Hautvast, J., Meyer, P. D., Borewicz, K., Vaughan, E. E. and Slavin, J. L. (2020) Effect of fructans, prebiotics and fibres on the human gut microbiome assessed by 16S rRNA-based approaches : A review, Beneficial Microbes, 11 (2), pp. 101-129. [CrossRef]
- Talavera, M. M., Nuthakki, S., Cui, H., Jin, Y., Liu, Y. and Nelin, L. D. (2017) Immunostimulated arginase II expression in intestinal epithelial cells reduces nitric oxide production and apoptosis, Frontiers in Cell and Developmental Biology, 5 , pp. 15. [CrossRef]
- Tandon, D., Haque, M. M., Gote, M., Jain, M., Bhaduri, A., Dubey, A. K. and Mande, S. S. (2019) A prospective randomized, double-blind, placebo-controlled, dose-response relationship study to investigate efficacy of fructo-oligosaccharides (FOS) on human gut microflora, Scientific Reports, 9 (1), pp. 5473. [CrossRef]
- Tanes, C., Bittinger, K., Gao, Y., Friedman, E. S., Nessel, L., Paladhi, U. R., Chau, L., Panfen, E., Fischbach, M. A., Braun, J., Xavier, R. J., Clish, C. B., Li, H., Bushman, F. D., Lewis, J. D. and Wu, G. D. (2021) Role of dietary fiber in the recovery of the human gut microbiome and its metabolome, Cell Host & Microbe, 29 (3), pp. 394-407.e5. [CrossRef]
- Tang, H., Fang, Z. and Ng, K. (2020) Dietary fiber-based colon-targeted delivery systems for polyphenols, Trends in Food Science & Technology, 100 , pp. 333-348. [CrossRef]
- Teufel, R., Mascaraque, V., Ismail, W., Voss, M., Perera, J., Eisenreich, W., Haehnel, W., Fuchs, G. and Harwood, C. S. (2010) Bacterial phenylalanine and phenylacetate catabolic pathway revealed, Proceedings of the National Academy of Sciences - PNAS, 107 (32), pp. 14390-14395. [CrossRef]
- Thompson, M. S., Hui Yan, T., Saari, N. and Sarbini, S. R. (2022) A review: Resistant starch, a promising prebiotic for obesity and weight management, Food Bioscience, 50 , pp. 101965. [CrossRef]
- Thompson, R. S., Roller, R., Mika, A., Greenwood, B. N., Knight, R., Chichlowski, M., Berg, B. M. and Fleshner, M. (2017) Dietary prebiotics and bioactive milk fractions improve NREM sleep, enhance REM sleep rebound and attenuate the stress-induced decrease in diurnal temperature and gut microbial alpha diversity, Frontiers in Behavioral Neuroscience, 10 , pp. 240. [CrossRef]
- Threapleton, D. E., Greenwood, D. C., Evans, C. E. L., Cleghorn, C. L., Nykjaer, C., Woodhead, C., Cade, J. E., Gale, C. P. and Burley, V. J. (2013) Dietary fibre intake and risk of cardiovascular disease: systematic review and meta-analysis, Bmj, -12-19, 347 (dec19 2), pp. f6879. [CrossRef]
- Tian, X., Li, R., Jiang, Y., Zhao, F., Yu, Z., Wang, Y., Dong, Z., Liu, P. and Li, X. (2020) Bifidobacterium breve ATCC15700 pretreatment prevents alcoholic liver disease through modulating gut microbiota in mice exposed to chronic alcohol intake, Journal of Functional Foods, 72 , pp. 104045. [CrossRef]
- Tomas, M. (2022) Effect of dietary fiber addition on the content and in vitro bioaccessibility of antioxidants in red raspberry puree, Food Chemistry, 375 , pp. 131897. [CrossRef]
- Toribio-Mateas, M. A., Bester, A. and Klimenko, N. (2021) Impact of plant-based meat alternatives on the gut microbiota of consumers: A real-world study, Foods, 10 (9), pp. 2040. [CrossRef]
- Tsitko, Wiik-Miettinen, Mattila, Rosa-Sibakov, Maukonen, Nordlund and Saarela (2019) A Small In Vitro Fermentation Model for Screening the Gut Microbiota Effects of Different Fiber Preparations, International Journal of Molecular Sciences, -04-18, 20 (8). [CrossRef]
- Unseenbio.com (2023) https://my.unseenbio.com/en/taxa/species/cag-81-sp900066785 Accessed 28.02.2023.
- USDA ARS (2021). https://www.ars.usda.gov/ARSUserFiles/80400530/pdf/1720/Table_1_NIN_GEN_1720.pdf Accessed 9.03.2023.
- Uyanga, V. A., Amevor, F. K., Liu, M., Cui, Z., Zhao, X. and Lin, H. (2021) Potential Implications of Citrulline and Quercetin on Gut Functioning of Monogastric Animals and Humans: A Comprehensive Review, Nutrients, -10-25, 13 (11). [CrossRef]
- Van Den Abbeele, P., Ghyselinck, J., Marzorati, M., Koch, A., Lambert, W., Michiels, J. and Chalvon-Demersay, T. (2022) The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota, Microorganisms, -03-31, 10 (4). [CrossRef]
- Van Rymenant, E., Abrankó, L., Tumova, S., Grootaert, C., Van Camp, J., Williamson, G. and Kerimi, A. (2017) Chronic exposure to short-chain fatty acids modulates transport and metabolism of microbiome-derived phenolics in human intestinal cells, The Journal of Nutritional Biochemistry, 39 , pp. 156-168. [CrossRef]
- Vandeputte, D., Falony, G., Vieira-Silva, S., Wang, J., Sailer, M., Theis, S., Verbeke, K. and Raes, J. (2017) Prebiotic inulin-type fructans induce specific changes in the human gut microbiota, Gut, -02-17, 66 (11), pp. 1968. [CrossRef]
- Venkataraman, A., Sieber, J. R., Schmidt, A. W., Waldron, C., Theis, K. R. and Schmidt, T. M. (2016) Variable responses of human microbiomes to dietary supplementation with resistant starch, Microbiome, -06-29, 4 (1). [CrossRef]
- Ventura, M., Turroni, F., Zomer, A., Foroni, E., Giubellini, V., Bottacini, F., Canchaya, C., Claesson, M. J., He, F., Mantzourani, M., Mulas, L., Ferrarini, A., Gao, B., Delledonne, M., Henrissat, B., Coutinho, P., Oggioni, M., Gupta, R. S., Zhang, Z., Beighton, D., Fitzgerald, G. F., O'Toole, P. W. and van Sinderen, D. (2009) The bifidobacterium dentium Bd1 genome sequence reflects its genetic adaptation to the human oral cavity, PLoS Genetics, 5 (12), pp. e1000785. [CrossRef]
- Vieira Bezerra, C., Da, A. M., Rodrigues3, C., Amante, E. R., Luiza, H., Meller, D. A. and Silva (2013) NUTRITIONAL POTENTIAL OF GREEN BANANA FLOUR OBTAINED BY DRYING IN SPOUTED BED 1 POTENCIAL NUTRICIONAL DA FARINHA DE BANANA VERDE OBTIDA POR SECAGEM EM LEITO DE JORRO, Rev. Bras. Frutic., Jaboticabal -SP, , pp. 1140.
- Vrancken, G., Rimaux, T., Wouters, D., Leroy, F. and De Vuyst, L. (2009) The arginine deiminase pathway of lactobacillus fermentum IMDO 130101 responds to growth under stress conditions of both temperature and salt, Food Microbiology, 26 (7), pp. 720-727. [CrossRef]
- Wagner, B. D., Grunwald, G. K., Zerbe, G. O., Mikulich-Gilbertson, S. K., Robertson, C. E., Zemanick, E. T. and Harris, J. K. (2018) On the Use of Diversity Measures in Longitudinal Sequencing Studies of Microbial Communities, Frontiers in Microbiology, -05th, 9 . [CrossRef]
- Wang, H., Huang, X., Tan, H., Chen, X., Chen, C. and Nie, S. (2022) Interaction between dietary fiber and bifidobacteria in promoting intestinal health, Food Chemistry, 393 , pp. 133407. [CrossRef]
- Wang, J., Wang, P., Li, D., Hu, X. and Chen, F. (2020) Beneficial effects of ginger on prevention of obesity through modulation of gut microbiota in mice, European Journal of Nutrition, 59 (2), pp. 699-718. [CrossRef]
- Wang, M., Wichienchot, S., He, X., Fu, X., Huang, Q. and Zhang, B. (2019) In vitro colonic fermentation of dietary fibers: Fermentation rate, short-chain fatty acid production and changes in microbiota, Trends in Food Science & Technology, 88 , pp. 1-9. [CrossRef]
- Wang, S., Xiao, Y., Tian, F., Zhao, J., Zhang, H., Zhai, Q. and Chen, W. (2020) Rational use of prebiotics for gut microbiota alterations: Specific bacterial phylotypes and related mechanisms, Journal of Functional Foods, 66 , pp. 103838. [CrossRef]
- Wang, W., Xu, S., Ren, Z., Tao, L., Jiang, J. and Zheng, S. (2015) Application of metagenomics in the human gut microbiome, World Journal of Gastroenterology : WJG, 21 (3), pp. 803-814. [CrossRef]
- Wang, X., Zhang, D., Jiang, H., Zhang, S., Pang, X., Gao, S., Zhang, H., Zhang, S., Xiao, Q., Chen, L., Wang, S., Qi, D. and Li, Y. (2021) Gut Microbiota Variation With Short-Term Intake of Ginger Juice on Human Health, Frontiers in Microbiology, -02nd, 11 . [CrossRef]
- Wang, Y., Ramos, M., Jefferson, M., Zhang, W., Beraza, N., Carding, S., Powell, P. P., Stewart, J. P., Mayer, U. and Wileman, T. (2022) Control of infection by LC3-associated phagocytosis, CASM, and detection of raised vacuolar pH by the V-ATPase-ATG16L1 axis, Science Advances, 8 (43), pp. eabn3298. [CrossRef]
- Watson, A. W., Houghton, D., Avery, P. J., Stewart, C., Vaughan, E. E., Meyer, P. D., de Bos Kuil, M. J. J., Weijs, P. J. M. and Brandt, K. (2019) Changes in stool frequency following chicory inulin consumption, and effects on stool consistency, quality of life and composition of gut microbiota, Food Hydrocolloids, 96 , pp. 688-698. [CrossRef]
- Weickert, M. O., Möhlig, M., Schöfl, C., Arafat, A. M., Otto, B., Viehoff, H., Koebnick, C., Kohl, A., Spranger, J. and Pfeiffer, A. F. H. (2006) Cereal fiber improves whole-body insulin sensitivity in overweight and obese women, Diabetes Care, 29 (4), pp. 775-780. [CrossRef]
- Weickert, M. O., Roden, M., Isken, F., Hoffmann, D., Nowotny, P., Osterhoff, M., Blaut, M., Alpert, C., Gögebakan, Ö, Bumke-Vogt, C., Mueller, F., Machann, J., Barber, T. M., Petzke, K. J., Hierholzer, J., Hornemann, S., Kruse, M., Illner, A., Kohl, A., Loeffelholz, C. V., Arafat, A. M., Möhlig, M. and Pfeiffer, A. F. (2011) Effects of supplemented isoenergetic diets differing in cereal fiber and protein content on insulin sensitivity in overweight humans, The American Journal of Clinical Nutrition, 94 (2), pp. 459-471. [CrossRef]
- Willis, A. D. (2019) Rarefaction, alpha diversity, and statistics, Frontiers in Microbiology, 10 , pp. 2407. [CrossRef]
- Wongkaew, M., Tangjaidee, P., Leksawasdi, N., Jantanasakulwong, K., Rachtanapun, P., Seesuriyachan, P., Phimolsiripol, Y., Chaiyaso, T., Ruksiriwanich, W., Jantrawut, P. and Sommano, S. R. (2022) Mango Pectic Oligosaccharides: A Novel Prebiotic for Functional Food, Frontiers in Nutrition, -03rd, 9 . [CrossRef]
- Wu, G. and Morris, J., S M (1998) Arginine metabolism: Nitric oxide and beyond, Biochemical Journal, 336 ( Pt 1) (1), pp. 1-17. [CrossRef]
- Wu, M., Xiao, H., Shao, F., Tan, B. and Hu, S. (2020) Arginine accelerates intestinal health through cytokines and intestinal microbiota, International Immunopharmacology, 81 , pp. 106029. [CrossRef]
- Gibson, M. B. and Roberfroid (1995) Dietary Modulation of the Human Colonie Microbiota: Introducing the Concept of Prebiotics, J. Nutr, 125 , pp. 1401.
- Yang, Y., Bin, P., Tao, S., Zhu, G., Wu, Z., Cheng, W., Ren, W. and Wei, H. (2021) Evaluation of the mechanisms underlying amino acid and microbiota interactions in intestinal infections using germ-free animals, Infectious Microbes & Diseases, 3 (2), pp. 79-86. [CrossRef]
- Yao, B., Fang, H., Xu, W., Yan, Y., Xu, H., Liu, Y., Mo, M., Zhang, H. and Zhao, Y. (2014) Dietary fiber intake and risk of type 2 diabetes: A dose-response analysis of prospective studies, European Journal of Epidemiology, 29 (2), pp. 79-88. [CrossRef]
- Zarei, I., Koistinen, V. M., Kokla, M., Klåvus, A., Babu, A. F., Lehtonen, M., Auriola, S. and Hanhineva, K. (2022) Tissue-wide metabolomics reveals wide impact of gut microbiota on mice metabolite composition, Scientific Reports, 12 (1), pp. 15018. [CrossRef]
- Zhang, B., Li, G., Shahid, M. S., Gan, L., Fan, H., Lv, Z., Yan, S. and Guo, Y. (2020) Dietary l-arginine supplementation ameliorates inflammatory response and alters gut microbiota composition in broiler chickens infected with salmonella enterica serovar typhimurium, Poultry Science, 99 (4), pp. 1862-1874. [CrossRef]
- Zhang, Ran, Lei Wang, Chengcheng Shi, Qipeng Shi, Fuying Ma, Xiaoyu Zhang, Wen Yu, and Hongbo Yu (2021) Structural Characterization of Lignin-Carbohydrate Complexes (LCCs) and Their Biotransformation by Intestinal Microbiota In Vitro, Journal of Agricultural and Food Chemistry 69 (43), 12880-12890. [CrossRef]
- Zhang, Y., Chen, R., Zhang, D., Qi, S. and Liu, Y. (2023) Metabolite interactions between host and microbiota during health and disease: Which feeds the other? Biomedicine & Pharmacotherapy, 160 , pp. 114295. [CrossRef]
- Zhao, S., Peng, X., Zhou, Q., Huang, Y., Rao, X., Tu, J., Xiao, H. and Liu, D. (2021) Bacillus coagulans 13002 and fructo-oligosaccharides improve the immunity of mice with immunosuppression induced by cyclophosphamide through modulating intestinal-derived and fecal microbiota, Food Research International, 140 , pp. 109793. [CrossRef]
- Zhao, Y. and Jiang, Q. (2021) Roles of the Polyphenol–Gut microbiota interaction in alleviating colitis and preventing colitis-associated colorectal cancer, Advances in Nutrition (Bethesda, Md.), 12 (2), pp. 546-565. [CrossRef]
- Zheng, Y., Bek, M. K., Prince, N. Z., Peralta Marzal, L. N., Garssen, J., Perez Pardo, P., Kraneveld, A. D., Afd Pharmacology and Pharmacology (2021) The role of bacterial-derived aromatic amino acids metabolites relevant in autism spectrum disorders: A comprehensive review, Frontiers in Neuroscience, 15 , pp. 1-738220. [CrossRef]
Figure 1.
Study design.
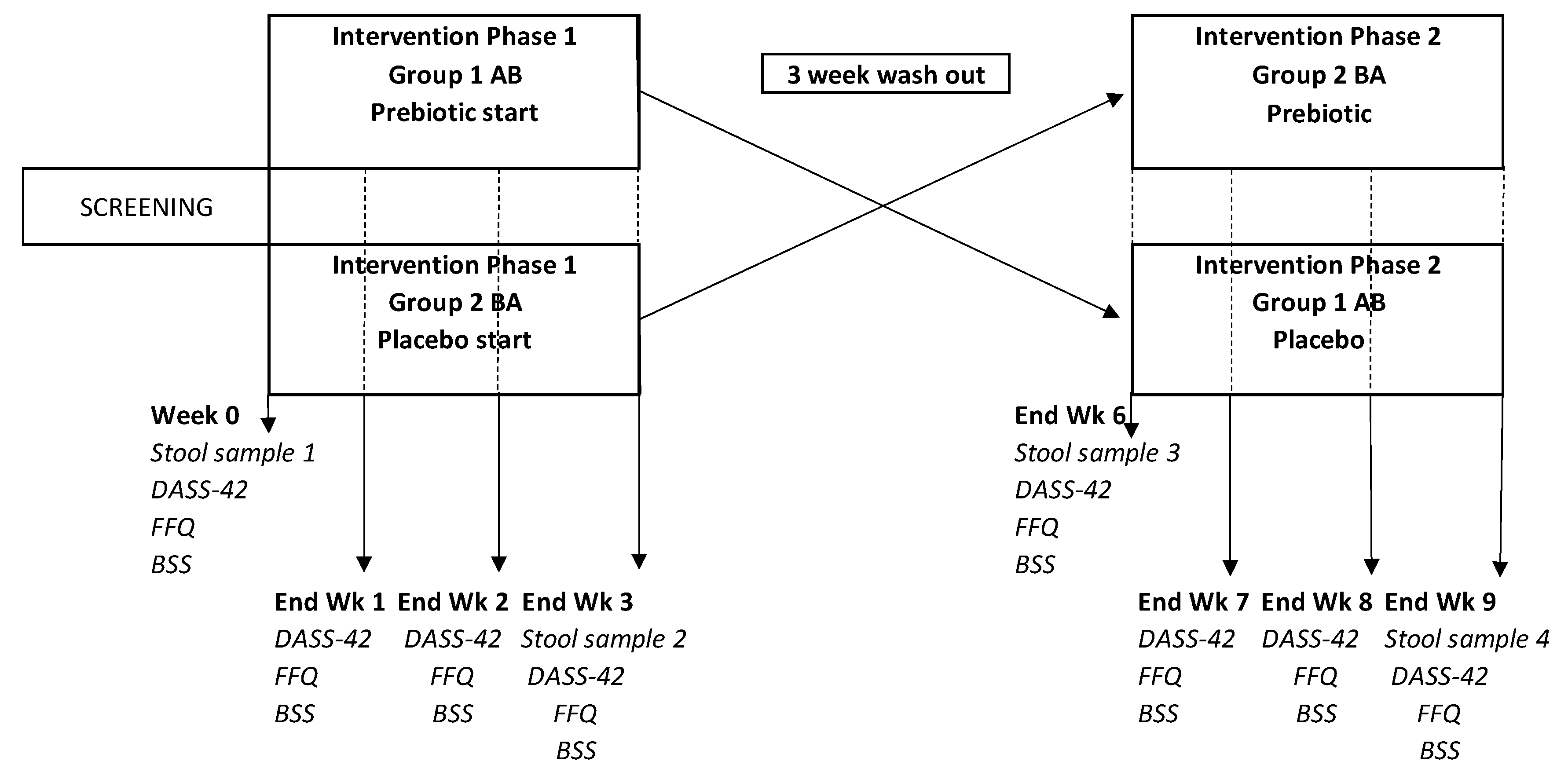
Figure 2.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota in healthy adults. Boxplot showing the (A) Shannon and (B) Simpson alpha diversity, grouped by treatment, colored by timepoint, and faceted by phase. (C) Box plot showing the percent abundance of B. adolescentis, colored by treatment, grouped by time point, and faceted by treatment phase.
Figure 2.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota in healthy adults. Boxplot showing the (A) Shannon and (B) Simpson alpha diversity, grouped by treatment, colored by timepoint, and faceted by phase. (C) Box plot showing the percent abundance of B. adolescentis, colored by treatment, grouped by time point, and faceted by treatment phase.
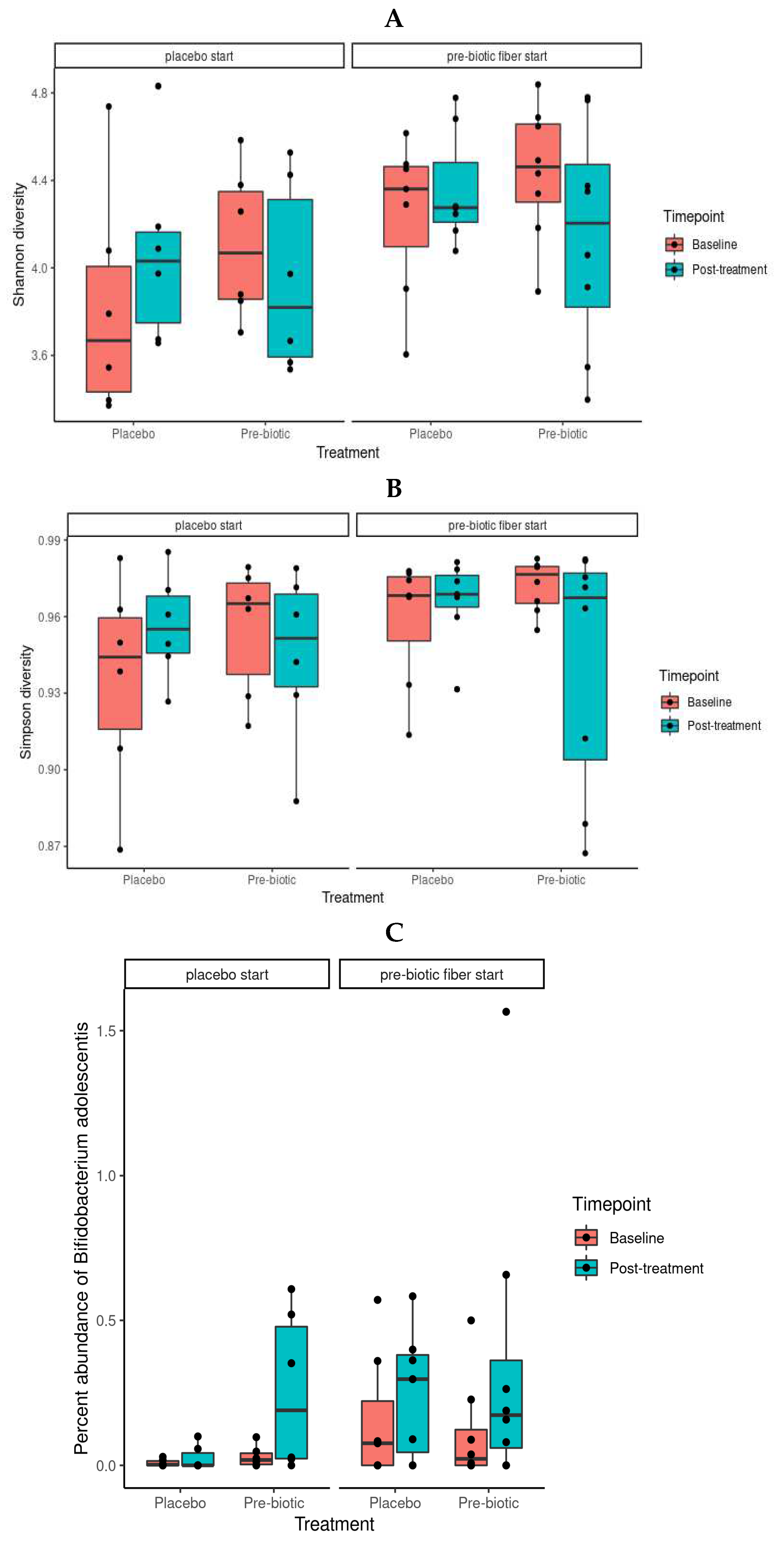
Figure 3.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota beta diversity in healthy adults. Principle component analysis (PCoA) is based on Aitchison distance matrixes. PCA plots of PCA dimensions 1, 2 and 3 of Beta diversity analysis. The ellipses represent 80% confidence intervals. Lines connect the same samples during each time point, with shapes indicating the time point, colored by treatment and grouping. Both graphs are bordered by violin plots showing projections onto the principal component axes. Violin plots with significantly different grouping are marked, with * = p-value <0.1, ** = p-value <0.05 and *** = p-value < 0.01.
Figure 3.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota beta diversity in healthy adults. Principle component analysis (PCoA) is based on Aitchison distance matrixes. PCA plots of PCA dimensions 1, 2 and 3 of Beta diversity analysis. The ellipses represent 80% confidence intervals. Lines connect the same samples during each time point, with shapes indicating the time point, colored by treatment and grouping. Both graphs are bordered by violin plots showing projections onto the principal component axes. Violin plots with significantly different grouping are marked, with * = p-value <0.1, ** = p-value <0.05 and *** = p-value < 0.01.
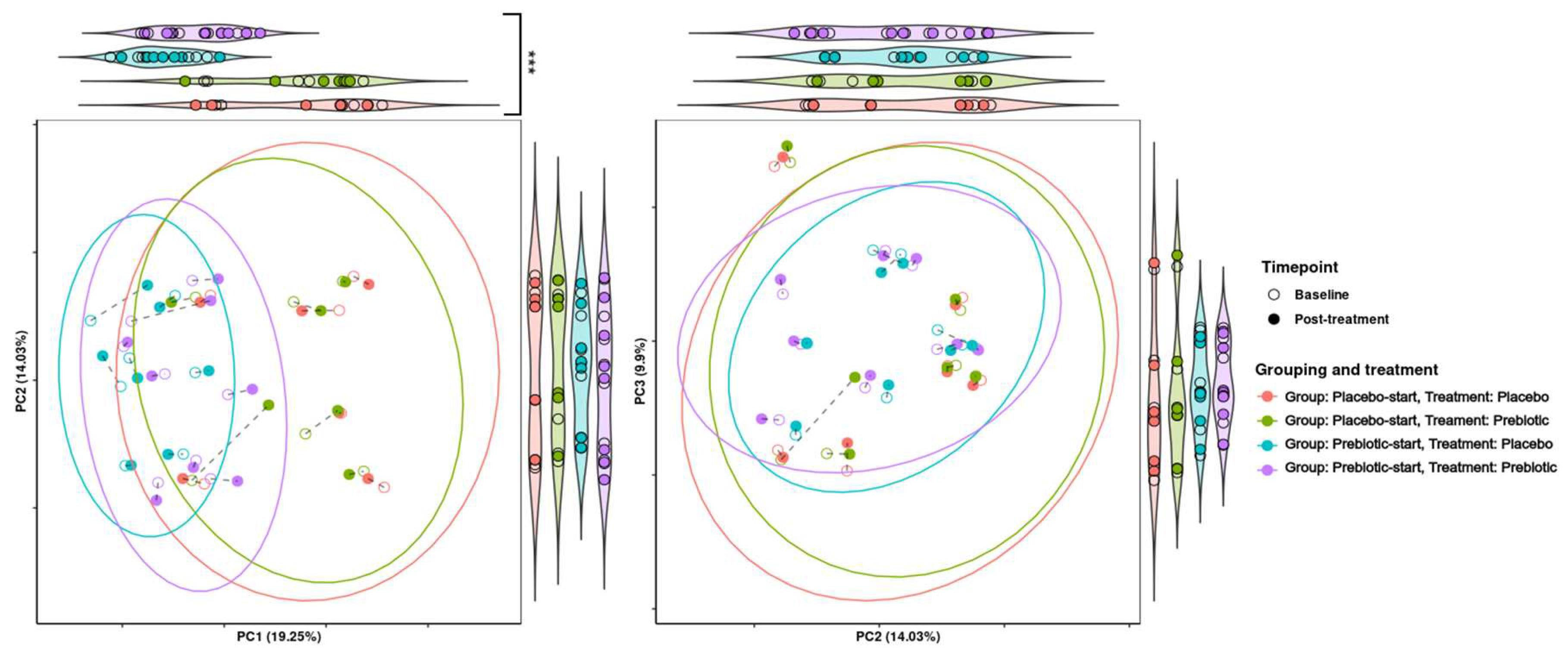
Figure 4.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota in healthy adults. Bar chart showing the percentage composition of the 20 most abundant species, colored by Genus, grouped by Phylum, faceted by Subject, Ordered by Treatment and timepoint. Note: Firmicutes_A is a placeholder phylum which has been classified as Firmicutes in the NCBI database but does not meet the clustering requirements for GTDB. Unused space represents other species not in the top 20 most abundant.
Figure 4.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota in healthy adults. Bar chart showing the percentage composition of the 20 most abundant species, colored by Genus, grouped by Phylum, faceted by Subject, Ordered by Treatment and timepoint. Note: Firmicutes_A is a placeholder phylum which has been classified as Firmicutes in the NCBI database but does not meet the clustering requirements for GTDB. Unused space represents other species not in the top 20 most abundant.
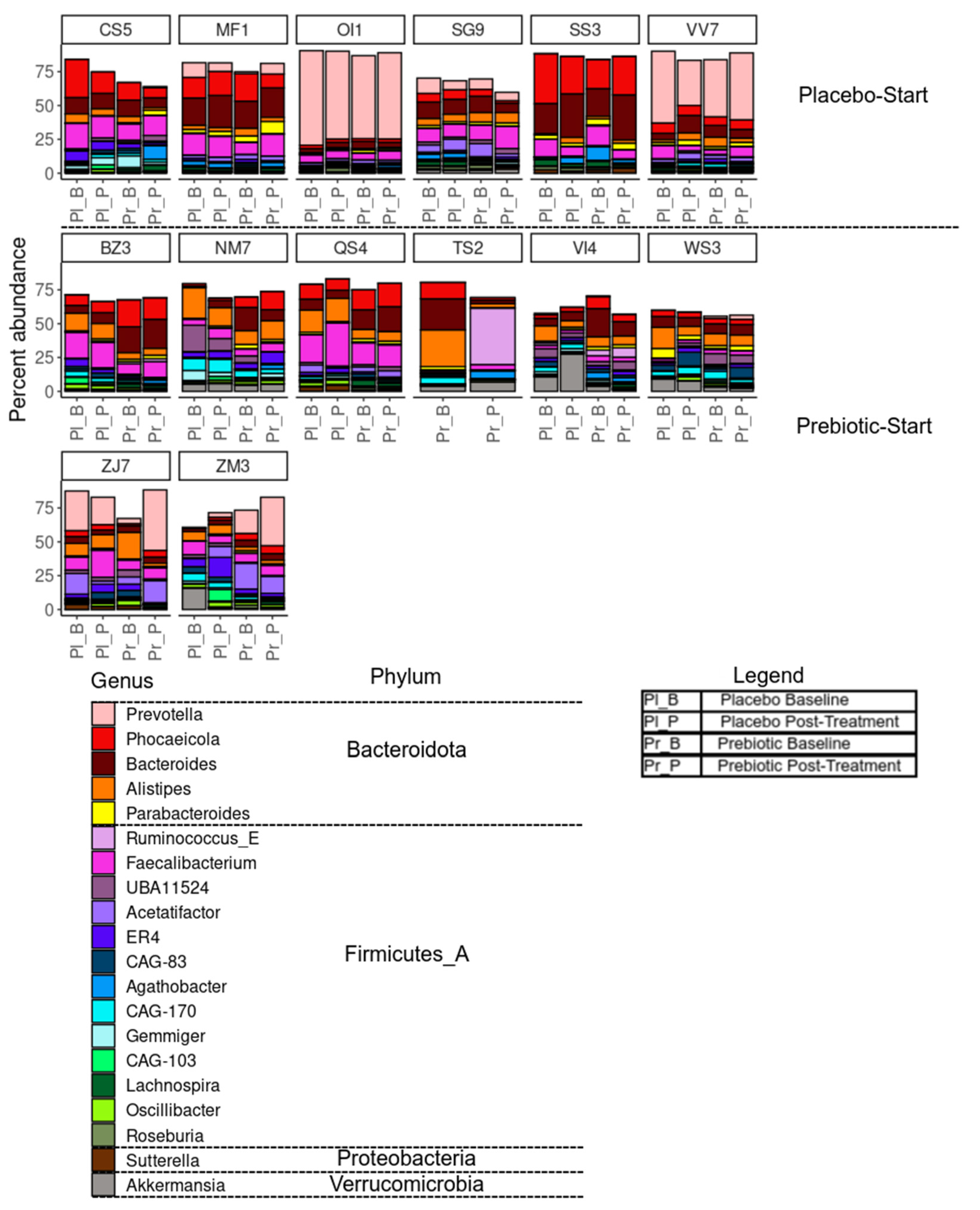
Figure 5.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota composition in healthy adults. Data represents a volcano plot of differential abundant gut microbiota species showing effect size and -log10 of the uncorrected p-value taxa between time points under (A) prebiotic fibres and (B) placebo (without the fibres). Colored by change relative to baseline and species with an uncorrected p-value <0.05 labelled. The horizontal line represents the unadjusted p-value cut-off at 0.05. N = 14.
Figure 5.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota composition in healthy adults. Data represents a volcano plot of differential abundant gut microbiota species showing effect size and -log10 of the uncorrected p-value taxa between time points under (A) prebiotic fibres and (B) placebo (without the fibres). Colored by change relative to baseline and species with an uncorrected p-value <0.05 labelled. The horizontal line represents the unadjusted p-value cut-off at 0.05. N = 14.
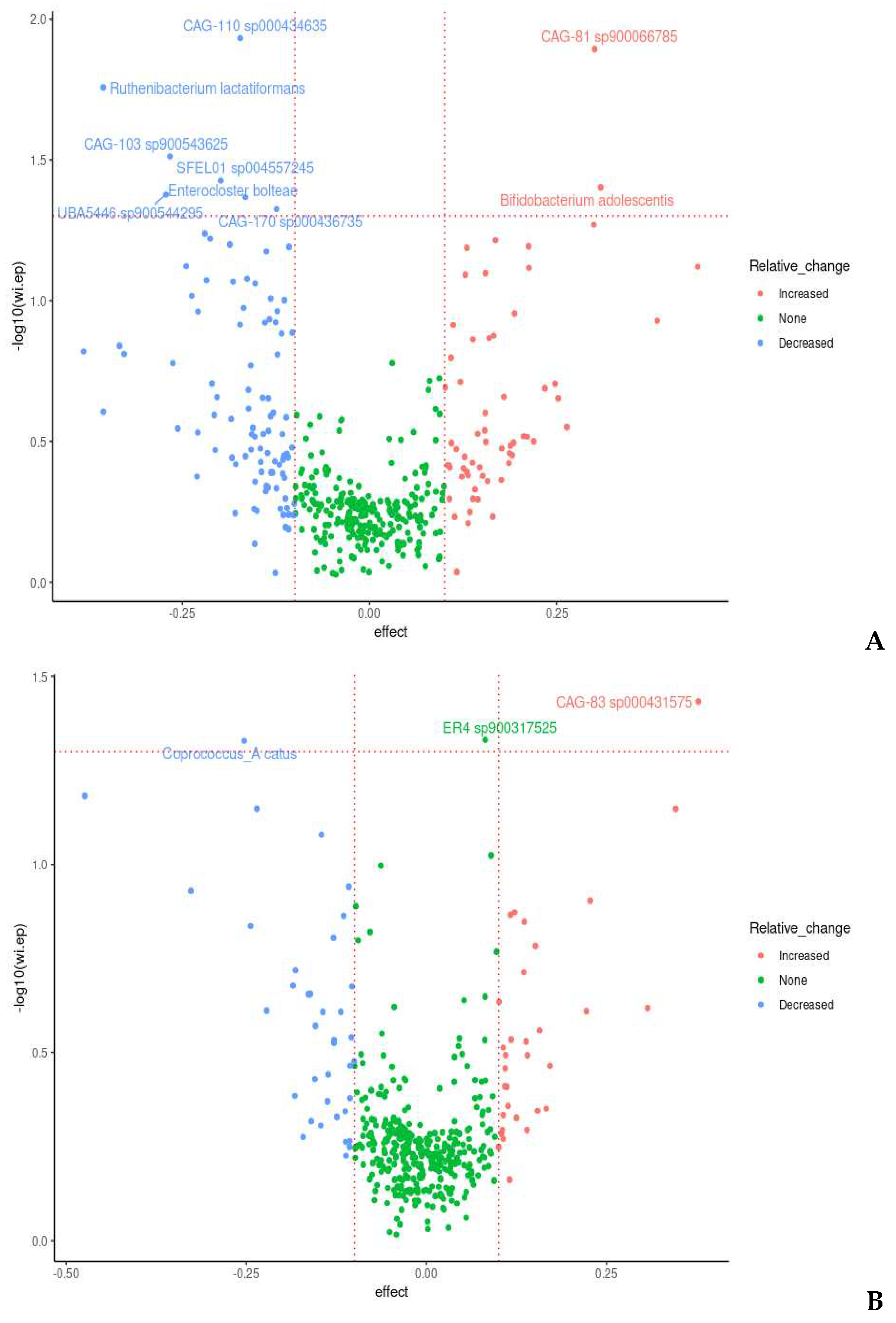
Figure 6.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults. Data represent a volcano plot of differential abundant KEGG Modules (A) with prebiotic fibres and (C) placebo, and KEGG Pathways (B) with prebiotic fibres and (D) placebo, between groups. The X-axis position of each point represents effect size differences at the end of the intervention, colored by change relative to baseline. The horizontal line represents the unadjusted p-value cut-off at 0.05; the vertical lines represent an effect size of -0.1 and 0.1. N = 14.
Figure 6.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on gut microbiota functional metagenome in healthy adults. Data represent a volcano plot of differential abundant KEGG Modules (A) with prebiotic fibres and (C) placebo, and KEGG Pathways (B) with prebiotic fibres and (D) placebo, between groups. The X-axis position of each point represents effect size differences at the end of the intervention, colored by change relative to baseline. The horizontal line represents the unadjusted p-value cut-off at 0.05; the vertical lines represent an effect size of -0.1 and 0.1. N = 14.
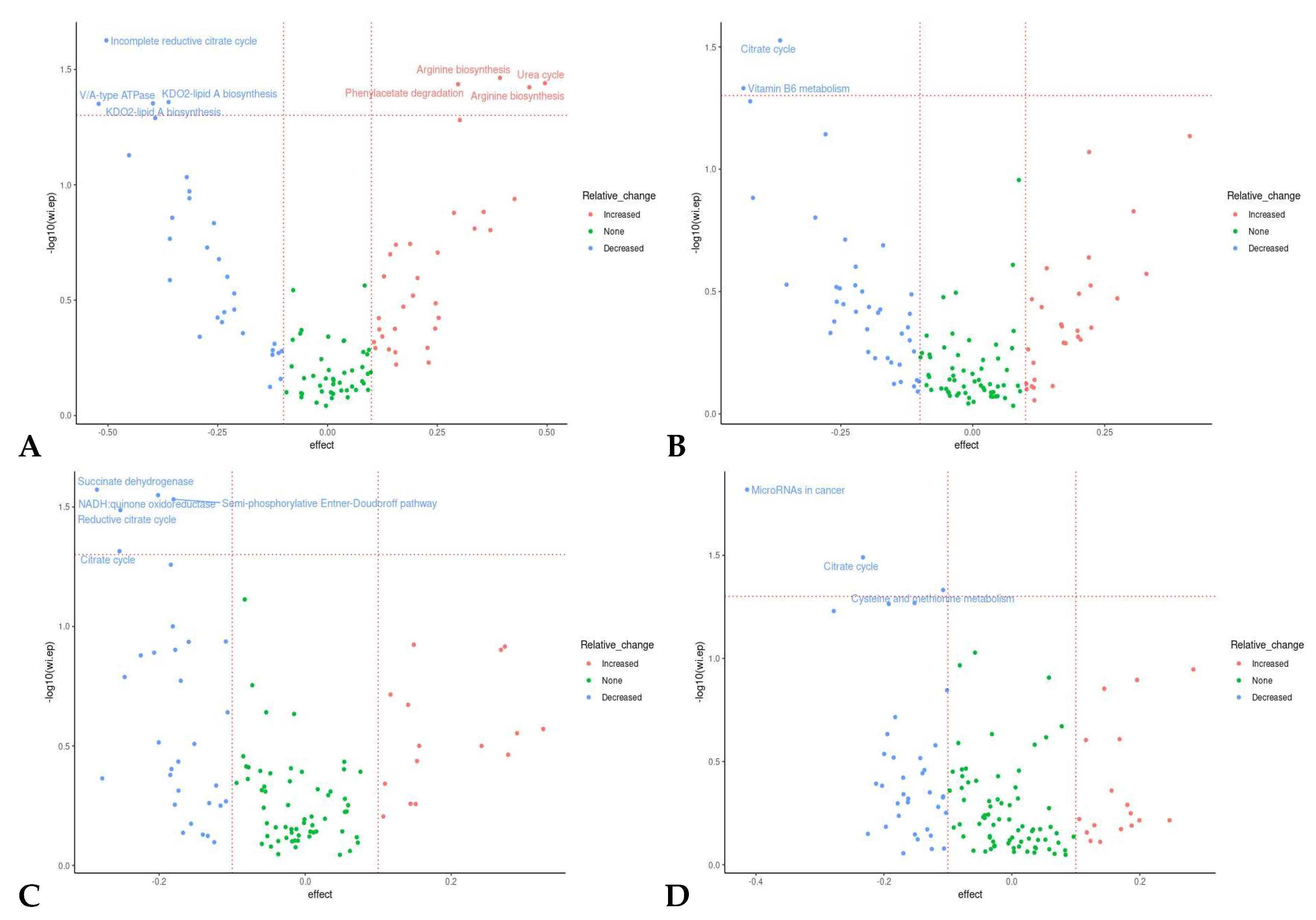

Table 1.
Habitual dietary inulin and oligofructose during the placebo supplement Phase. Data are gram per day (Mean ± SD), independent t-test.
Table 1.
Habitual dietary inulin and oligofructose during the placebo supplement Phase. Data are gram per day (Mean ± SD), independent t-test.
| Group/Treatment order | Phase | Inulin | Between group p value | Oligofructose | Between group p value |
|---|---|---|---|---|---|
| AB | 2 | 4.0 ± 1.65 | .04* | 3.47 ± 1.28 | .78 |
| BA | 1 | 6.44 ± 2.1 | 5.62 ± 2.52 |
* 2-tailed.
Table 2.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on individual responses of gut microbiota at genus level in healthy adults. AB is treatment order prebiotic start, and BA is treatment order placebo start.
Table 2.
Effects of 3-week supplementation of the MOJU Prebiotic Shot or placebo (without the fibres) on individual responses of gut microbiota at genus level in healthy adults. AB is treatment order prebiotic start, and BA is treatment order placebo start.
| Treatment order | AB Prebiotic phase | AB Placebo phase | BA Placebo phase | BA Prebiotic phase | Baseline between-group differences (p values) | ||||
| Results | Pre-post p value | Individual responsesN=8 | Pre-post p value | Individual responsesN = 7 | Pre-post p value | Individual responsesN = 6 | Pre-post p value | Individual responsesN = 6 | |
| Verrucomicrobiota | .125.25 | 7 ↑, 1↓ | .4531.0 | 5 ↑, 2↓ | .062.372 | 5 ↑, 1↔ | 1.0 | 3 ↑, 2↓, 1↔ | .181 |
| Proteobactera | .453.67 | 5 ↑, 3↓ | .4530.9 | 5 ↑, 2↓ | .6881.0 | 4 ↑, 2↓ | .6881.0 | 4 ↑, 2↓ | .534 |
| Firmicutes | 1.0 | 4 ↑, 4↓ | 1.0 | 3 ↑, 4↓ | .6881.0 | 2 ↑, 4↓ | 1.0 | 3 ↑, 3↓ | .534 |
| Desulfobacteria | .016*.09 | 8 ↑ | .453.67 | 5 ↑, 2↓ | 1.0 | 3 ↑, 3↓ | .6881.0 | 4 ↑, 2↓ | .628 |
| Bacteroidetes | .453.54 | 2 ↑, 6↓ | .125.75 | 1 ↑, 6↓ | .219.65 | 1 ↑, 5↓ | .6881.0 | 4 ↑, 2↓ | .101 |
| Actinobacteria | .016*.048* | 8 ↑ | .453.54 | 5 ↑, 2↓ | 1.0 | 3 ↑, 3↓ | 1.0 | 3 ↑, 3↓ | .366 |
* Denotes significant p value; Related-Samples Sign test; Benjamini-Hochberg adjusted p values in bold; baseline between-group differences were analysed using the Mann-Whitney U test; ↑= increase, ↓= decrease, ↔= no change.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



