
Submitted:
02 May 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
Insect outbreaks are major drivers of natural disturbances in forest ecosystems. Outbreaks can have both direct and indirect effects on the composition of soil arthropod communities, through canopy opening, nutrient addition and predator-prey interactions. In this study, we aim to understand the effects of forest tent caterpillar (Malacosoma disstria; FTC) outbreaks on ant communities in both temperate and boreal forests in Canada. Using pitfall traps and Berlese funnels, we compared the ant communities as well as the surrounding arthropod communities between control and outbreak sites in boreal andboreal and temperate forests (in Québec, Canada). Using the Sørensen dissimilarity index, we determined the alpha and beta diversity of the ant community. Other arthropods collected in the traps were counted to evaluate the richness and abundance of potential prey for the ants and other potential predators of the FTC. We used an indicator species analysis to examine the species associated with sites defoliated by the outbreak. In the boreal forest, we found that FTC outbreaks caused decreases in species richness and increases in the evenness of ant communities in defoliated sites. In the boreal forest sites, species composition varied significantly between control and outbreak sites. This pattern was driven by the presence of other predators. We also saw no changes in beta diversity in the boreal forest but did see a significant decrease in the temperate forest between the outbreak sites and the control sites. A similar, but weaker pattern was observed in the temperate forest. Ant species in the boreal forest tended to exhibit a more marked preference for either control or previously defoliated sites than species in the temperate forest. Our study showed that disturbances like insect outbreaks can drive changes in the ant community. While we saw small effects of outbreaks, manipulation experiments using resource addition could help us validate the mechanisms behind these relationships.
Keywords:
ant community
; disturbance ecology
; forest ecology
; insect outbreaks
; natural disturbances
1. Introduction
Insect outbreaks are major natural disturbances in forests and have been for a long time [1,2,3]. Defoliation by insect herbivores not only slows tree growth, but it can also promote nutrient cycling and accelerate succession [4]. Caterpillars are prey and their population explosions affect trophic webs [5]. Ecosystem effects of insect outbreaks can happen through various direct and indirect mechanisms: leaf removal increases light and temperature on the forest floor, exploding caterpillar populations increase prey availability for many predators, and frass, insect corpses and dropped foliage constitute nutrient transfers from trees to soils [5,6,7,8,9,10]. Together, these effects can stimulate decomposition, enhance nutrient mineralization, increase soil respiration, promote plant growth and alter brown food webs [11,12,13,14,15]. However, the full complexity of underground food webs remains poorly understood and the role of soil arthropods in mediating the effects of leaf-eating insect outbreaks on ecosystem functioning is not clear [16].
Caldéron-Sanou et al. [16] found that direct and indirect effects of caterpillar outbreaks increase the diversity of other arthropods in the underground food web at different trophic levels. They showed that the magnitude of the effects of defoliation did not decrease at higher trophic levels, contrary to what may be assumed. The researchers expected the effect of defoliation to be diluted the higher the trophic level. Instead, they observed a more diverse food web and a greater proportion of high-trophic level taxa in defoliated than in control forests. In this kind of study in temperate and boreal regions, it is expected that the effects of disturbance will be higher for primary decomposers and producers and lessen at higher trophic levels such as predators. This finding does not support a mitigation hypothesis related to disturbances where the effects of disturbances would be lower for species at the top of the food web. Additionally, because so many of the species making up the soil community are responsible for structural and functional characteristics of the ecosystem, a change in their assemblage could have large impacts on ecosystems [17,18,19,20,21,22].
Ants are particularly important in northern forests since they are one of the few large-scale ecological engineers [23,24,25]. Ants make up a large part of the insect biomass and can play multiple ecological roles such as predators, soil engineers, nutrient cyclers and regulators of plant growth and reproduction [26,27,28,29], thus shaping both in-ground and above-ground trophic webs. In these northern forests, ants play a crucial role in arthropod communities [30,31] and nutrient fluxes [29,32]. The social organization of ant colonies means they can respond rapidly and dramatically to changes in the environment and hence can mediate ecosystem effects of disturbance [33].
Prey availability for ants can increase with defoliator outbreaks both directly due to the presence of caterpillars and indirectly due to the stimulation of brown food webs. Indeed, soil detrivores and fungivores respond to an increase in microbial biomass and activity induced by high-quality inputs leading to increased soil respiration and nutrient cycling, especially if the nutrient inputs are sustained for a few years [5,34,35,36,37]. Many of these soil microarthropods can be prey for ants [38,39], but this is not the only way in which changes in brown food webs can influence ants. Indeed, ants are deeply interconnected with boreal and temperate forest trophic webs [30,31], but their responses to changes in these communities are not well understood. For instance, increases in other arthropod predators could constrain ant responses. Ants are well-understood to exert significant predation pressure on forest defoliators [40,41,42,43]. However, the reverse, namely the effects of outbreaks on ant communities, has received less attention. Considering the keystone role of ants, this knowledge gap constrains our understanding of cascading effects of defoliator outbreaks.
The impacts of forest canopy opening on ants have mostly been studied through the effects of forest management [44,45,46,47]. Multiple studies have shown an increase in ant abundance and diversity with moderate management intensity [46,48,49]. The main driver appears to be a change in microclimate on the forest floor resulting from to canopy opening. Grevé et al. [50] found that forest management (proportion of harvested tree volume and even-aged stands) increases abundance, species richness and functional diversity in ant communities in temperate forests and that this was due to reduced canopy cover and stand structural complexity. They also found that shade-intolerant ant species were more likely to be favoured and that this was likely due to warm conditions in the stands. In Japanese temperate forests, both open habitat specialists and generalists were abundant in managed forests, but woodland specialists declined [45,51]. In boreal forests, similar trends have been observed in Europe [48,52]. In general, in temperate and boreal forests open habitats seem to have positive effects on ant diversity and abundance. Less shade-tolerant ant species were especially correlated with lower canopy coverage in European studies [48,52,53].
The forest tent caterpillar (Malacosoma disstria) is an important forest defoliator of hardwoods across much of North America. Outbreaks usually last 3-5 years and, while they slow the growth of host trees, seldom lead to widespread tree death [54]. Defoliation during forest tent caterpillar outbreaks increases canopy openness, leading to an increase in sun exposure to the forest floor, resulting in higher soil temperatures, drier soil, and increased growth of saplings and understory plants due to increased light availability [55]. Similar canopy opening has been shown to increase ant abundance and species richness in managed or recovering forests [46,48].
However, insect outbreaks, unlike most forest management practices, also involve an increase in prey abundance for ants, both directly from the outbreaking caterpillars and indirectly via the stimulation of brown food webs. Indeed, ants are common predators of caterpillars [56] and have been shown to have substantial impacts on the abundance of caterpillars [40,57,58] and other leaf-chewing herbivores [59]. Ant predation has been suggested to show a density-dependent response to caterpillar availability, to increase during an outbreak and to play a role in controlling the outbreak [60,61]. Ants could alter trophic cascades by lowering herbivory damage [62,63,64]. While thinking of trophic cascades, it is also important to consider that ants predate on soils microorganisms, such as springtails, as well, with certain groups like the ground-dwelling Dacetini being specialized predators [65], thus acting on different parts of the food web associated with caterpillar outbreaks.
In this study, we investigate the dynamic between ant communities and defoliation at the ecosystem level in both a boreal and a temperate forest. Specifically, we evaluate the effects of a forest tent caterpillar outbreak (Malacosoma disstria) on ant communities, examining drivers related to both environmental conditions on the forest floor and changes in soil arthropod communities. We hypothesize that canopy opening, the presence of high numbers of caterpillars, and an increase in soil arthropods driven by nutrient inputs will lead to higher ant species diversity. Increased energy and nutrient flow, combined with increased heterogeneity of the forest floor is predicted to open niches thus promoting higher ant diversity. In terms of evenness, we have two possibilities: either many species are favoured, thus leading to a more homogenous community (higher evenness) or only a few species can take advantage of novel conditions, thus leading to a more heterogenous community (lower evenness). We also examine associations of the ant species with control or outbreak sites. Finally, we evaluate the role of three potential drivers related to outbreaks which could affect ant populations, namely canopy opening, increased soil microarthropod populations (using collembola as a representative group), and changes in soil arthropod predator populations.
2. Materials and Methods
2.1. Study area and experimental design
This study was conducted in two forest stands that were both affected by the most recent FTC outbreak. In both regions, an FTC outbreak was detected in 2016 [66] and continued in 2017 [67], but no defoliation was observed in 2018 or 2019.
The Forêt d’enseignement et de recherche du lac Duparquet (FERLD; N48.513, W79.369) is within the boreal mixed-wood forests of eastern Canada, in post-fire (1923) regenerated stands dominated by trembling aspen (Populus tremuloides Michx). The climate is cold-temperate with a (1961-1990) mean annual temperature of 0.9 °C and mean annual precipitation of 642 mm of rain and 215 mm of snow. Fire drives the disturbance regime and large even-age trembling aspen stands arise from post-fire regeneration [68]. Soils are Grey Luvisols [69] originating from glaciolacustrine clay deposits [70]. The region was affected by an FTC outbreak for 3 years starting in 2015 [71] and continuing in 2016 and 2017 [66,67]. No FTC defoliation was observed in either 2018 or 2019 [71].
Kenauk Nature Reserve (N45.712, W74.887) is within a temperate deciduous forest comprised of stands dominated by sugar maple (Acer saccharum Marsh) and American beech (Fagus grandifolia Ehrh). The climate is cold-temperate with a mean annual temperature of 5°C and mean total annual rainfall of 807.4 mm and total annual snowfall is 178.1 cm [73]. The disturbance regime is driven by a mix of insect pests and weather events. Soils in the study area are classified as Dystric Brunisols with moder-type humus [69,74].
2.2. Experimental design
In the boreal forest, we selected 28 sites in trembling aspen-dominated stands (generally over 50% of aspen and where other trees were conifers) including 14 that were heavily defoliated in 2016 - 2017 (outbreak sites) and 14 control sites. This variable will be referred to as defoliation history for the rest of the article. In the temperate forest, twelve (12) sites in sugar maple stands that were heavily defoliated in 2016 and 2017 (outbreak sites), and 12 control sites in similar age stands having escaped defoliation were sampled in 2018 and 2019. In both cases, each site is characterized by a focal tree on which we put experimental devices to study populations of FTC (see [75] for a full description of the methods). The arthropod sampling was done within 5 meters of the focal tree at each site. We sampled all sites 2 times per field season over two years. The traps, sampling period and year for each site.
2.3. Environmental variables
To confirm the defoliation history of control and outbreak sites, we examined twenty saplings of the respective focus tree species in each region between 1 and 3 m tall in a 100 m radius around the focal tree for forest tent caterpillar colonies in 2017, 2018 and 2019. At each site, we also measured canopy openness using a densiometer in 2018 and 2019 (every two weeks during the months of May, June and July) and used an average of four measurements taken on the densiometer, as per the methodology recommended with the tool. Then the average of the measurements taken at the same sites since the bud burst during different periods.
2.4. Ant survey
Ants were sampled in control stands at FERLD in 2016 by Despland and Lessard [61] and these data were only included in the species list. since the sampling method was quite different from the ones used the following year. They, however, helped get a snapshot of the ant fauna. Standardized ant sampling in control and outbreak sites was conducted in 2017-2019 at FERLD and in 2018-2019 at Kenauk.
At each site, five pitfall traps of 50 mm diameter filled with propylene glycol and one drop of both ethanol and unscented liquid soap were set in the ground for 48 h [76]. Pitfall traps were positioned in a radius around the focal tree, 5 meters from one another. The process was only repeated two times, once in late May and once in late June, to coincide with the period during which FTC is most active. Note that ants are not very active in May, but late June is near the peak activity period.
All ants sampled in the pitfall traps were identified to species level and confirmed by Dr. André Francoeur [77,78]. Morphospecies were used for the specimens that were either too deteriorated or could not be identified using taxonomic keys. Ant abundance was transformed to presence-absence data to account for bias that could come from nest proximity. Ant occurrence was evaluated as the number of traps in which a given species of ant was collected.
For further analyses, we pooled data from all the traps at each site. We calculated ant species richness as the number of ant species found per plot.
2.5. Survey of other arthropods
We collected collembolas to use them as a proxy of other potential food resources for ants on top of the FTC larvae. Collembolas were sampled as well in a subset (N=8) of outbreak and control sites. Samples were taken within a diameter of 1m around the focal tree. At each tree, two replicates of litter were taken within a 20x20cm quadrat, two soil samples from 0-5cm and two samples from 5-10 cm using a 5 cm corer. The two replicates for each sampling type were combined and conserved at 4°C and extracted less than 48h after collecting in the laboratory. Extraction of the soil fauna was done using Berlese-Tullgren extractors running for 7 days with temperatures gradually increasing from 20° C to 50°C. The specimens were conserved in 70% ethanol and sorted to separate collembolas from other soil organisms. Collembolas were prepared from identification and identified using published keys [79,80,81,82] as well as collembola.org and ecotaxonomy.org. Specimens were identified to the lowest taxonomic level, if possible, but were at least identified to family. Specimens were also grouped into morphospecies for hard-to-identify species (Gagnon-Koudji et al. In prep).
Additionally, all other potential predators found in the pitfall traps, such as spiders, wasps, beetles, and stinkbugs, were counted and total predator abundance was included in statistical analyses.
2.6. Statistical analysis
An ANOVA was performed to evaluate the effect of defoliation history (independent variable) on ant species richness and evenness (dependent variables). All years were pooled together in each region (2017-2019 for the boreal forest and 2018-2019 for the temperate forest) in all analyses since preliminary analysis showed no effect or interaction of year.
To investigate the differences in community composition between sites with different defoliation histories, we calculated a dissimilarity matrix based on the Sørensen dissimilarity index [83]. This index was used for the community analysis (for both ordination and beta diversity analyses) because it is one of the widely used indices for presence-absence datasets and examines the number of species shared by two sites and the number of species unique to each. The maximum and minimum values of Sørensen are 0 (the same species composition) and 1 (no shared species). We used nonparametric multidimensional scaling (NMDS) plots via the metaMDS function in the vegan package for R [85] to visualize the dissimilarity matrices. We then tested the differences in the taxonomic position of the community centroids (multivariate location) between defoliation histories using ‘Permutational Multivariate Analysis of Variance’ (PERMANOVA) using the adonis2 function in the vegan package with 999 permutations. These vectors of our chosen variables (defoliation history, canopy openness, predator abundance, prey abundance and prey species richness) were tested for significant effects in shaping the observed ant communities. Since we cannot use AICs or similar methods for model selection with PERMANOVAs, we opted to choose the variables to retain in our analysis by selecting variables that had an R2 of more than 0.02 when doing a PERMANOVA with all possible variables.
To further explore the community changes due to the different regimes of defoliation, we performed an indicator species analysis [86]. This method allows us to assess quantitatively the association of specific species of ants to either of our two groups, in this case, control and outbreak sites. It assesses the predictive values of species as indicators of the conditions prevailing under different defoliation regimes [87]. Component ‘A’ is a conditional probability referring to the positive predictive value of the species as an indicator of the defoliation history. Component ‘B’ is another conditional probability referring to the probability of finding a species in sites experiencing the defoliation history. Both components vary from 0 to 1 [86]. All the analyses were carried out using R [88].
3. Results
We collected and identified a total of 2944 individual worker ants belonging to 54 species and morphospecies from 3 subfamilies and 10 genera across our 28 sites in the boreal forest and 24 sites in the temperate forest.
Ants were not present in all pitfall traps, even though they collected other arthropods. From the sites sampled in the boreal forest, we were able to collect ants from 100% of the control sites and 92% of the outbreak sites. In the temperate forest, we collected ants from 83% of the control sites and 50% of the outbreak sites. Species accumulation curves validated our experimental design in terms of species sampling for both defoliation histories and both regions.
Forest tent caterpillar colonies were observed in 2017 in outbreak sites (3.36 ± 1.86 (mean ± SD) colonies of 20 saplings) but not in control sites, and none were observed at all in 2018 and 2019 in the boreal forest. In the temperate forest, we observed colonies both in the outbreak sites (5.5 ± 2.85 (mean ± SD) colonies of 20 saplings) and in the control sites (4.01 ± 2.50 (mean ± SD) colonies of 20 saplings).
3.1. Species richness and evenness
Three species were shared between the two regions. We found a total of 30 species in the boreal forest sites, with the species occurring most often being Camponotus novaeboracensis (23% of traps), Myrmica alaskensis and Formica subanescens. In the temperate forest sites, we found a total of 18 species with the species occurring most often being Aphaenogaster picea (25% of traps), Lasius americanus and Stenamma diecki. Many rare species, with only one occurrence, were observed.
In the boreal forest, we observed significantly lower species richness (df =1, F=9.901, P=0.003) and significantly higher evenness in the outbreak than in the control sites (df=1, F=8.667, P=0.005). We identify similar trends in the temperate forest sites, however, the high proportion of traps that did not collect ants (90 %) reduced the sample size and hence the power of the analyses. At least one trap per site had ants, therefore, when pooled, we had ants in 83% of the outbreak sites and 50% of the control sites. Species richness did not differ between control and outbreak sites (df=1, F=1.53, P=0.234), but evenness was significantly higher in outbreak sites (df=1, F=5.008, P=0.056) (Figure 1, Table 1).
3.2. Species composition
3.2.1. Ordination
In the boreal forest sites, species composition varied significantly between control and outbreak sites (F = 5.391, p = 0.002, R2 = 0.091 df = 1) (Figure 2a; Table 2). Canopy openness, collembola abundance and diversity all align with defoliation showing that they all increase with increased defoliation, but do not contribute significantly to predicting ant communities. Predators also increase with defoliation and were significantly associated with changes in the ant community.
3.3. Beta diversity
The taxonomic multivariate dispersion (i.e., homogenization) did not differ significantly between defoliation histories in the boreal forest (F = 1.021, df = 1, p = 0.27), but did in the temperate forest (F = 7.60, df = 1, p = 0.01) (Figure 3). However, it is important to note that homogenization is quite low (i.e. beta diversity is high) for all our sites since the distance to the centroid is high above the null expectation.
3.4. Indicator species
The indicator Value (IndVal) index measures the association between a species and a site group, in our case, defoliation history. Out of the 30 species found in the selected sites in the boreal forest, 26% (5 species) were significantly associated with one group, with 4 representing the control sites and 1 the outbreak sites. (Table 3) Another 4 species were found to be as likely to occur in both groups.
In the temperate forest, however, we found no significant pattern of association with the defoliation history groups. Out of the 15 species found in the selected sites in the temperate forest, 4 species were as likely to occur in both groups, but even for others showing a preference for one of the two groups, none was strong enough to be considered as an indicator.
4. Discussion
Contrary to our hypothesis that disturbance would increase the diversity present at defoliated sites by creating new ecological niches, the sites where the outbreak occurred, did not contain a more diverse ant community. In the boreal forest, defoliation slightly lowered ant alpha diversity, but did not alter beta diversity or the proportion of singletons. The increase in evenness was due to no species being overwhelmingly dominant. The only significant trend identified in the temperate forest was a decrease in beta diversity with defoliation. In the boreal forest, observed changes in ant communities were not significantly linked to changes in canopy openness, prey abundance or diversity, but did correlate with an overall increase in predator abundance in defoliated sites. In the boreal forest, four ant species are identified as indicators of control sites and one as an indicator of outbreak sites.
4.1. Ant community responses to disturbance
In both temperate and boreal forests, sites that had experienced defoliation due to an outbreak were more even and less species-rich than the control sites; however, this effect only attained statistical significance in the boreal. Disturbances generally have a greater impact on arthropod communities in structurally simple habitats than in complex ones [89,90].
Ants are typically impacted indirectly by disturbances through effects on habitat structure, microclimate, resource availability and competitive interactions [94,95,96,97,98]. In our study, we know (Lafleur, pers. obs. (boreal) and Nowell, pers. obs. (temperate)) there was a large amount of light reaching the understory during the outbreak, therefore creating a disturbance in the light regime. Outbreaks of species like the FTC are what could be considered a moderate chronic disturbance [99] meaning that an outbreak event in itself causes moderate damage in an area since leaves can reflush during the summer and that the epidemic cycle is relatively short (10 years) in contrast to the timescale on which a forest operates. However, since these outbreaks are not static geographically and transient in time [100], the impacts might be less dramatic especially since impacts on ant communities are mostly driven by long-term disturbance regimes [98,101,102,103,104]. While FTC outbreaks are part of this disturbance regime and can alter forest succession in the boreal forest [55], they are relatively short-lived compared to those of the spruce budworm [105].
Research suggests that ant foraging in temperate regions is sensitive to sun exposure on the forest floor [61,106]. This influence on foraging could then translate to changes in communities as foraging and competition for it are some of the main drivers of community assemblage [107]. Canopy openness was higher in defoliated sites, but it was not a significant contributor to predicting ant communities in our study, suggesting that the observed canopy opening was not significant enough to directly affect ant communities. However, it is important to mention that we did not have the canopy openness data for the years of the outbreak and, thus, this measure relates to the canopy 1- and 2 years post-outbreak. In the boreal region, rarer shade species, however, were excluded from outbreak sites and replaced with more common species. Therefore, while canopy openness was not retained as an indicator in our model, we nonetheless see a change in the community that is consistent with a role for opening. Our study system, therefore, resembles uneven-aged forest management with a strong vertical structure and small gaps that close quickly. These small gaps are less favourable to shade-intolerant ants than large open areas created by forest harvesting.
4.2. Indicator species
Taxa that favour open habitats such as generalist Myrmicinae usually do well in disturbed habitats whereas groups favouring closed habitats such as specialist predators are often at a disadvantage [108,109,110,111]. In our study, species that were only present in outbreak sites are known from the literature to be associated with more open habitats such as managed forests, bogs and open areas [112,113,114,115,116,117].
In the boreal forest, Camponotus novaeboracensis was the species occurring most often in both control and defoliated stands. C. novaeboracensis is a behaviourally dominant ant [118] and has been observed successfully attacking forest tent caterpillars during the outbreak preceding our study [61]. However, our results do not show this species becoming more dominant in outbreak sites.
The main species that were retained as indicators of control sites in the boreal forest were Camponotus herculeanus and Myrmica detritinodis. C. herculeanus is a very common species in boreal forests [119,120]. M. detritinodis is classified as a shade species and prefers shelter under moss and lichens, preferably in high moisture conditions [121]. It is expected to prefer a high canopy cover. C. herculeanus is more surprising since it has not been seen to be previously associated with either closed or open habitats [119,120]. In our system, we conclude that they might be more associated with closed habitats, but may depend on other environmental characteristics.
The one species associated with outbreak sites is Formica integra, but limited resources are available on their ecology due to their low occurrences. However, they are part of the Formica rufa group (Red wood ants) whose ecology at large has been well-studied in European boreal and temperate forests. In particular, red wood ants feed on invertebrate prey from both the canopy and forest floor [25,28,122] and have been suggested to have the potential to control outbreaks of insect pests [123]. They also build large mounds on the forest floor for their nests. The distribution of these mounds is governed by multiple factors (climate, ecosystem productivity, food resources) [124,125], but light availability stood out as the most important factor [126] signifying the importance of canopy openness. In our study, their increase in outbreak sites suggests that this ant species is uniquely positioned to benefit from a short-term canopy opening and resource pulse associated with a defoliator outbreak and to carry it over temporally.
Since these mounds concentrate nutrients and can affect the biotic and abiotic components of the surrounding forest [29,43,127,128,129], they can increase the presence of associated species e.g. spiders, beetles, and millipedes, living near them, such as parasites, predators, and scavengers as they provide a stable habitat with consistently higher temperatures than the rest of the environment [130].
Formica and other Camponotus species have been observed to attack other Lepidopterans in eastern Canada [61]. While studying ants' predation of the spongy moth, Weseloh [131] found that both Formica and Camponotus spp. workers attacked caterpillars, but especially at the first instar, decreasing as the larvae grew. The use of forest tent caterpillars as prey could thus contribute to populations of F. integra and C. novaeboracensis in outbreak sites.
In the temperate forest, the species most occurring were smaller-bodied ants from the Aphaenogaster and Lasius genera. However, no species were found to be indicators of either control or outbreak sites. Aphaenogaster species from the species complex A. picea are omnivorous feeders and generally feed on small invertebrates as well as some mushrooms [132]. Lasius americanus is omnivorous and feeds on seeds, and live and dead insects [78]. Both the Aphaenogaster and Lasius genera were widely distributed in the sites studied, but their distribution did not appear to be influenced by a history of defoliation.
4.3. Prey, predators, and outbreaks
Results from Grevé et al. [50] support our finding that overall prey abundance did not influence ant species composition through increased food resources. However, our results show a positive relationship between ants and other predators in the boreal forests. But the basis for this relationship is not clear. These arthropods could be responding to disturbance similarly to ants but without any direct trophic interactions between them [50]. Generalist predators tend to increase slowly in response to increased prey availability during a caterpillar outbreak, thus showing up as delayed density dependence scenarios [133]. High numbers can thus be maintained in the year following the outbreak crash [134,135].
Previous work also confirms that increased prey abundance does not drive ant community responses to disturbance [50] and that ants do not respond to resource pulses of arthropod prey [136]. Resources pulses tend to satiate predators and aboveground consumers [137,138,139]. This pulse can also lead to changes in both structure and dynamics of communities [140] This phenomenon is particularly present in boreal forest communities which were affected by outbreaks of spruce budworm. This outbreak and the resulting pulse increase the relative abundance of mobile predators and parasitoids [141]. It is, however, important to note that spruce budworm outbreaks such as the ones in the cited studies last much longer than FTC. Therefore, it is not surprising that the effects are larger. The decrease in ant diversity at outbreak sites did correlate with an increase in other arthropod predators, suggesting that these predators might have been more effective than ants in responding to the prey pulse generated by the insect outbreak [50]. This trend could then emphasize an indirect effect of other predators on the ant population but was not directly studied.
5. Conclusions
Changes in the ant community were only observed in the boreal forest sites. Indeed, less ecologically complex ecosystems are often less resilient to disturbances such as FTC defoliation [50]. In the boreal forest, ant community composition did differ depending on defoliation history, albeit with the opposite effect we expected, with lower species richness and no clear association with either increased sunlight or prey availability. The increase in other potential predators of FTC shows a path toward the hypothesis of the resource pulse since this functional group also increased with defoliation. The effect of resource addition to ants and other predators would provide us with a better understanding of the large disturbance event that are insect outbreaks. In 2018, neither soil respiration [142], nor collembola (Gagnon-Koudji et al., in prep) were higher in defoliated sites. If a peak occurred during defoliation, it was finished by the following year. Therefore, predators including ants are showing a delayed response to previous high resource availability. If boreal defoliated sites have been pushed on a successional trajectory toward increased coniferous dominance [55] then shade-tolerant specialist ants will likely return but communities will continue to diverge. Further research exploring microclimatic changes and resources related to FTC outbreaks and ants is needed to decipher the mechanisms at play. Natural history knowledge for specific ant species is also needed to be able to make light of these mechanisms. Additionally, as mentioned by Kristensen, Rousk, and Metcalfe [143], a distinction between low-intensity chronic herbivory and intense pulses like those present during outbreaks in similar systems would be beneficial in deciphering the underlying mechanisms leading to changes in food webs.
Author Contributions
ASC, JJJ, ECG, ITH and ED made contributions to the conception and design, ASC, ECG and JJJ acquired the data, and ASC analyzed and interpreted the data with the help of ED, MMG, ECG and IRH. ASC wrote the manuscript and MMG and ED revised it critically. MMG and ED provided resources and supervision. All authors gave final approval and agreed to be accountable for all aspects of the work, such as the accuracy and integrity of the work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministère de la Forêt de la Faune et des Parcs (Quebec), the Natural Sciences and Engineering Research Council (Canada), Norbord Inc., Kenauk Institute, and SERG-international.
Data Availability Statement
Data can be found in Spectrum, Concordia University's open-access research repository.
Acknowledgments
We want to acknowledge that this work was done on the treaty and traditional territory of the Abitibiwinni and Algonquin Anishinabeg people in the boreal forest as well as the Kanienʼkehá꞉ka, Omàmìwininìwag (Algonquin) and Anishinabewaki in the temperate forest. We thank Liane Nowell from The Kenauk Institute for access to the Kenauk territory and the invaluable support during data collection from Liane Nowell and the Kenauk Institute. We would like to thank Gaspar Legendre who set up the experiment and collected data in 2017. Dr. André Francoeur for his help with the identification of the ant specimens. Dr. Jean-Philippe Lessard for his guidance through the set-up and analysis of the study as well as valuable comments along with Dr. Carly Ziter. Javier Ibarra Isassi and Dr. Eric Pedersen for their help with the statistical analysis. Pamela Yataco Marquez for the sampling, as well as April Mansfield, Sarah Farhat and Andrew Cormier for their help with sorting the samples and identifying the samples.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Aakala, T.; Remy, C.C.; Arseneault, D.; Morin, H.; Girardin, M.P.; Gennaretti, F.; Navarro, L.; Kuosmanen, N.; Ali, A.A.; Boucher, É.; et al. Millennial-Scale Disturbance History of the Boreal Zone. In Boreal Forests in the Face of Climate Change: Sustainable Management; Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y., Eds.; Advances in Global Change Research; Springer International Publishing: Cham, 2023; pp. 53–87. ISBN 978-3-031-15988-6. [Google Scholar]
- Montoro Girona, M.; Navarro, L.; Morin, H. A Secret Hidden in the Sediments: Lepidoptera Scales. Front. Ecol. Evol. 2018, 6, 2. [Google Scholar] [CrossRef]
- Navarro, L.; Morin, H.; Bergeron, Y.; Girona, M.M. Changes in Spatiotemporal Patterns of 20th Century Spruce Budworm Outbreaks in Eastern Canadian Boreal Forests. Frontiers in Plant Science 2018, 9, 1905. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.; Montoro Girona, M.; Grosbois, G.; Morin, H. Does the Type of Silvicultural Practice Influence Spruce Budworm Defoliation of Seedlings? Ecosphere 2021, 12, e03506. [Google Scholar] [CrossRef]
- De Grandpré, L.; Marchand, M.; Kneeshaw, D.D.; Paré, D.; Boucher, D.; Bourassa, S.; Gervais, D.; Simard, M.; Griffin, J.M.; Pureswaran, D.S. Defoliation-Induced Changes in Foliage Quality May Trigger Broad-Scale Insect Outbreaks. Commun Biol 2022, 5, 463. [Google Scholar] [CrossRef] [PubMed]
- Swank, W.T.; Waide, J.B.; Crossley, D.A.; Todd, R.L. Insect Defoliation Enhances Nitrate Export from Forest Ecosystems. Oecologia 1981, 51, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.D. Insect Population Dynamics Meets Ecosystem Ecology: Effects of Herbivory on Soil Nutrient Dynamics. Agric Forest Ent 2001, 3, 77–84. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Asner, G.P.; Martin, R.E.; Silva Espejo, J.E.; Huasco, W.H.; Farfán Amézquita, F.F.; Carranza-Jimenez, L.; Galiano Cabrera, D.F.; Baca, L.D.; Sinca, F.; et al. Herbivory Makes Major Contributions to Ecosystem Carbon and Nutrient Cycling in Tropical Forests. Ecol Lett 2014, 17, 324–332. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Crutsinger, G.M.; Kumordzi, B.B.; Wardle, D.A. Nutrient Fluxes from Insect Herbivory Increase during Ecosystem Retrogression in Boreal Forest. Ecology 2016, 97, 124–132. [Google Scholar] [CrossRef]
- Lovett, G.M.; Christenson, L.M.; Groffman, P.M.; Jones, C.G.; Hart, J.E.; Mitchell, M.J. Insect Defoliation and Nitrogen Cycling in Forests. BioScience 2002, 52, 335. [Google Scholar] [CrossRef]
- Gravel, D.; Albouy, C.; Thuiller, W. The Meaning of Functional Trait Composition of Food Webs for Ecosystem Functioning. Philosophical Transactions of the Royal Society B: Biological Sciences 2016, 371, 20150268. [Google Scholar] [CrossRef]
- Barnes, A.D.; Jochum, M.; Lefcheck, J.S.; Eisenhauer, N.; Scherber, C.; O’Connor, M.I.; de Ruiter, P.; Brose, U. Energy Flux: The Link between Multitrophic Biodiversity and Ecosystem Functioning. Trends in ecology & evolution 2018, 33, 186–197. [Google Scholar] [CrossRef]
- Kristensen, J.A.; Metcalfe, D.B.; Rousk, J. The Biogeochemical Consequences of Litter Transformation by Insect Herbivory in the Subarctic: A Microcosm Simulation Experiment. Biogeochemistry 2018, 138, 323–336. [Google Scholar] [CrossRef]
- Lovett, G.M.; Ruesink, A.E. Carbon and Nitrogen Mineralization from Decomposing Gypsy Moth Frass. Oecologia 1995, 104, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Debaly, Z.M.; Marchand, P.; Girona, M.M. Autoregressive Models for Time Series of Random Sums of Positive Variables: Application to Tree Growth as a Function of Climate and Insect Outbreak. Ecological Modelling 2022, 471, 110053. [Google Scholar] [CrossRef]
- Calderón-Sanou, I.; Münkemüller, T.; Zinger, L.; Schimann, H.; Yoccoz, N.G.; Gielly, L.; Foulquier, A.; Hedde, M.; Ohlmann, M.; Roy, M.; et al. Cascading Effects of Moth Outbreaks on Subarctic Soil Food Webs. Sci Rep 2021, 11, 15054. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Caruso, T. Soil Microbial Community Responses to Climate Extremes: Resistance, Resilience and Transitions to Alternative States. Philosophical Transactions of the Royal Society B 2020, 375, 20190112. [Google Scholar] [CrossRef] [PubMed]
- De Ruiter, P.C.; Neutel, A.-M.; Moore, J.C. Energetics, Patterns of Interaction Strengths, and Stability in Real Ecosystems. Science 1995, 269, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B. Trophic Downgrading of Planet Earth. science 2011, 333, 301–306. [Google Scholar] [CrossRef]
- Saravesi, K.; Aikio, S.; Wäli, P.R.; Ruotsalainen, A.L.; Kaukonen, M.; Huusko, K.; Suokas, M.; Brown, S.P.; Jumpponen, A.; Tuomi, J. Moth Outbreaks Alter Root-Associated Fungal Communities in Subarctic Mountain Birch Forests. Microbial Ecology 2015, 69, 788–797. [Google Scholar] [CrossRef]
- Vindstad, O.P.L.; Schultze, S.; Jepsen, J.U.; Biuw, M.; Kapari, L.; Sverdrup-Thygeson, A.; Ims, R.A. Numerical Responses of Saproxylic Beetles to Rapid Increases in Dead Wood Availability following Geometrid Moth Outbreaks in Sub-Arctic Mountain Birch Forest. PLoS ONE 2014, 9, e99624. [Google Scholar] [CrossRef]
- Sandén, H.; Mayer, M.; Stark, S.; Sandén, T.; Nilsson, L.O.; Jepsen, J.U.; Wäli, P.R.; Rewald, B. Moth Outbreaks Reduce Decomposition in Subarctic Forest Soils. Ecosystems 2020, 23, 151–163. [Google Scholar] [CrossRef]
- Jurgensen, M.F.; Finér, L.; Domisch, T.; Kilpeläinen, J.; Punttila, P.; Ohashi, M.; Niemelä, P.; Sundström, L.; Neuvonen, S.; Risch, A.C. Organic Mound-Building Ants: Their Impact on Soil Properties in Temperate and Boreal Forests. Journal of Applied Entomology 2008, 132, 266–275. [Google Scholar] [CrossRef]
- Risch, A.C.; Jurgensen, M.F. Ants in the Soil System-a Hydrological, Chemical and Biological Approach. Journal of Applied Entomology 2008, 132, 265. [Google Scholar] [CrossRef]
- Domisch, T.; Finér, L.; Neuvonen, S.; Niemelä, P.; Risch, A.C.; Kilpeläinen, J.; Ohashi, M.; Jurgensen, M.F. Foraging Activity and Dietary Spectrum of Wood Ants ( Formica Rufa Group) and Their Role in Nutrient Fluxes in Boreal Forests. Ecological Entomology 2009, 34, 369–377. [Google Scholar] [CrossRef]
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The Little Things That Run the World Revisited: A Review of Ant-Mediated Ecosystem Services and Disservices (Hymenoptera: Formicidae). Myrmecological News 2012, 17, 133–146. [Google Scholar]
- Folgarait, P.J. Ant Biodiversity and Its Relationship to Ecosystem Functioning: A Review. Biodiversity & Conservation 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wardle, D.A.; Hyodo, F.; Bardgett, R.D.; Yeates, G.W.; Nilsson, M.-C. Long-Term Aboveground and Belowground Consequences of Red Wood Ant Exclusion in Boreal Forest. Ecology 2011, 92, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Laine, K.J.; Niemelä, P. The Influence of Ants on the Survival of Mountain Birches during an Oporinia Autumnata (Lep., Geometridae) Outbreak. Oecologia 1980, 47, 39–42. [Google Scholar] [CrossRef]
- Punttila, P.; Niemelä, P.; Karhu, K. The Impact of Wood Ants (Hymenoptera: Formicidae) on the Structure of Invertebrate Community on Mountain Birch (Betula Pubescens Ssp. Czerepanovii). In Proceedings of the Annales Zoologici Fennici; JSTOR, 2004; pp. 429–446. [Google Scholar]
- Finér, L.; Jurgensen, M.F.; Domisch, T.; Kilpeläinen, J.; Neuvonen, S.; Punttila, P.; Risch, A.C.; Ohashi, M.; Niemelä, P. The Role of Wood Ants (Formica Rufa Group) in Carbon and Nutrient Dynamics of a Boreal Norway Spruce Forest Ecosystem. Ecosystems 2013, 16, 196–208. [Google Scholar] [CrossRef]
- Andersen, A.N. Responses of Ant Communities to Disturbance: Five Principles for Understanding the Disturbance Dynamics of a Globally Dominant Faunal Group. J Anim Ecol 2019, 88, 350–362. [Google Scholar] [CrossRef]
- Kaukonen, M.; Ruotsalainen, A.L.; Wäli, P.R.; Männistö, M.K.; Setälä, H.; Saravesi, K.; Huusko, K.; Markkola, A. Moth Herbivory Enhances Resource Turnover in Subarctic Mountain Birch Forests? Ecology 2013, 94, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Mikola, J.; Yeates, G.W.; Barker, G.M.; Wardle, D.A.; Bonner, K.I. Effects of Defoliation Intensity on Soil Food-Web Properties in an Experimental Grassland Community. Oikos 2001, 92, 333–343. [Google Scholar] [CrossRef]
- Pitman, R.M.; Vanguelova, E.I.; Benham, S.E. The Effects of Phytophagous Insects on Water and Soil Nutrient Concentrations and Fluxes through Forest Stands of the Level II Monitoring Network in the UK. Science of the total environment 2010, 409, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.K.; Hart, S.C.; Cobb, N.S.; Whitham, T.G.; Koch, G.W. Insect Herbivory Increases Litter Quality and Decomposition: An Extension of the Acceleration Hypothesis. Ecology 2003, 84, 2867–2876. [Google Scholar] [CrossRef]
- Moya-Laraño, J.; Wise, D.H. Direct and Indirect Effects of Ants on a Forest-Floor Food Web. Ecology 2007, 88, 1454–1465. [Google Scholar] [CrossRef] [PubMed]
- Çakır, M. The Negative Effect of Wood Ants (Formica Rufa) on Microarthropod Density and Soil Biological Quality in a Semi-Arid Pine Forest. Pedobiologia 2019, 77, 150593. [Google Scholar] [CrossRef]
- Karhu, K.J.; Neuvonen, S. Wood Ants and a Geometrid Defoliator of Birch: Predation Outweighs Beneficial Effects through the Host Plant. Oecologia 1998, 113, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Way, M.J.; Khoo, K.C. Role of Ants in Pest Management. Annu. Rev. Èntomol. 1992, 37, 479–503. [Google Scholar] [CrossRef]
- Tilman, D. Cherries, Ants and Tent Caterpillars: Timing of Nectar Production in Relation to Susceptibility of Caterpillars to Ant Predation. Ecology 1978, 59, 686–692. [Google Scholar] [CrossRef]
- Gösswald, K Die Waldameise. Band 2. Die Waldameise in Okosystem Wald, Ihr Nutzen Und Ihre Hege. Wiesbaden: Aula-Verlag. 1990, 510 S.
- Carvalho, K.S.; Vasconcelos, H.L. Forest Fragmentation in Central Amazonia and Its Effects on Litter-Dwelling Ants. Biological Conservation 1999, 91, 151–157. [Google Scholar] [CrossRef]
- Maeto, K.; Sato, S. Impacts of Forestry on Ant Species Richness and Composition in Warm-Temperate Forests of Japan. Forest Ecology and Management 2004, 187, 213–223. [Google Scholar] [CrossRef]
- Palladini, J.D.; Jones, M.G.; Sanders, N.J.; Jules, E.S. The Recovery of Ant Communities in Regenerating Temperate Conifer Forests. Forest Ecology and Management 2007, 242, 619–624. [Google Scholar] [CrossRef]
- Ewers, R.M.; Boyle, M.J.; Gleave, R.A.; Plowman, N.S.; Benedick, S.; Bernard, H.; Bishop, T.R.; Bakhtiar, E.Y.; Chey, V.K.; Chung, A.Y. Logging Cuts the Functional Importance of Invertebrates in Tropical Rainforest. Nature communications 2015, 6, 1–7. [Google Scholar] [CrossRef]
- Punttila, P.; Haila, Y.; Niemelä, J.; Pajunen, T. Ant Communities in Fragments of Old-Growth Taiga and Managed Surroundings. In Proceedings of the Annales Zoologici Fennici; JSTOR; 1994; pp. 131–144. [Google Scholar]
- Véle, A.; Holuša, J.; Horák, J. Ant Abundance Increases with Clearing Size. Journal of forest research 2016, 21, 110–114. [Google Scholar] [CrossRef]
- Grevé, M.E.; Hager, J.; Weisser, W.W.; Schall, P.; Gossner, M.M.; Feldhaar, H. Effect of Forest Management on Temperate Ant Communities. Ecosphere 2018, 9, e02303. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. Forest Gap Dynamics and Tree Regeneration. Journal of forest research 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Punttila, P.; Haila, Y.; Pajunen, T.; Tukia, H. Colonisation of Clearcut Forests by Ants in the Southern Finnish Taiga: A Quantitative Survey. Oikos 1991, 61, 250–262. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S. The Impact of Even-Aged and Uneven-Aged Forest Management on Regional Biodiversity of Multiple Taxa in European Beech Forests. Journal of applied Ecology 2018, 55, 267–278. [Google Scholar] [CrossRef]
- Fitzgerald, T. The Tent Caterpillars; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Moulinier, J. Impacts de La Défoliation Par La Livrée Des Forêts Sur La Mortalité Du Peuplier Faux-Tremble et La Dynamique Forestière Post-Épidémie En Forêt Boréale. 2013.
- Lach, L.; Parr, C.; Abbott, K. Ant Ecology; Oxford university press, 2010.
- Montllor, C.B.; Bernays, E.A. Invertebrate Predators and Caterpillar Foraging. 1993.
- Piñol, J.; Espadaler, X.; Cañellas, N.; MARTÍNEZ-VILALTA, J.; Barrientos, J.A.; Sol, D. Ant versus Bird Exclusion Effects on the Arthropod Assemblage of an Organic Citrus Grove. Ecological Entomology 2010, 35, 367–376. [Google Scholar] [CrossRef]
- Clark, R.E.; Farkas, T.E.; Lichter-Marck, I.; Johnson, E.R.; Singer, M.S. Multiple Interaction Types Determine the Impact of Ant Predation of Caterpillars in a Forest Community. Ecology 2016, 97, 3379–3388. [Google Scholar] [CrossRef]
- Green, G.; Sullivan, C. Ants Attacking Larvae of the Forest Tent Caterpillar, Malacosoma Disstria Hbn. (Lepidoptera: Lasiocampidae). The Canadian Entomologist 1950, 82, 94–195. [Google Scholar] [CrossRef]
- Despland, E.; Lessard, J.-P. Social Predation by Ants as a Mortality Source for an Arboreal Gregarious Forest Pest. Basic and Applied Ecology 2022, 59, 82–91. [Google Scholar] [CrossRef]
- Rosumek, F.B.; Silveira, F.A.O.; de, S. Neves, F.; de U. Barbosa, N.P.; Diniz, L.; Oki, Y.; Pezzini, F.; Fernandes, G.W.; Cornelissen, T. Ants on Plants: A Meta-Analysis of the Role of Ants as Plant Biotic Defenses. Oecologia 2009, 160, 537–549. [Google Scholar] [CrossRef]
- Romero, G.Q.; Antiqueira, P.A.; Koricheva, J. A Meta-Analysis of Predation Risk Effects on Pollinator Behaviour. PLoS ONE 2011, 6, e20689. [Google Scholar] [CrossRef]
- Moreira, X.; Mooney, K.A.; Zas, R.; Sampedro, L. Bottom-up Effects of Host-Plant Species Diversity and Top-down Effects of Ants Interactively Increase Plant Performance. Proceedings of the Royal Society B: Biological Sciences 2012, 279, 4464–4472. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, X.; Dejean, A. 3. Predation by Ants on Arthropods and Other Animals. In Predation in the hymenoptera: An evolutionary perspective; Transworld Research Network: 2011.
- MFFP Aires infestées par la livrée des forêts au Québec en 2016; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2016; p. 10.
- MFFP Aires infestées par la livrée des forêts au Québec en 2017; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2017; p. 14.
- Bergeron, Y. Species and Stand Dynamics in the Mixed Woods of Quebec’s Southern Boreal Forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Canada, A. and A.-F. The Canadian System of Soil Classification (Second Edition). Available online: https://sis.agr.gc.ca/cansis/publications/manuals/1987-cssc-ed2/index.html (accessed on 3 November 2022).
- Vincent, J.-S.; Hardy, L. L’évolution et l’extension Des Lacs Glaciaires Barlow et Ojibway En Territoire Québécois. Géographie physique et Quaternaire 1977, 31, 357–372. [Google Scholar] [CrossRef]
- MFFP Aires infestées par la livrée des forêts au Québec en 2015; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2015; p. 8.
- MFFP Aires infestées par la livrée des forêts au Québec en 2018; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2019; p. 14.
- Environment Canada National Climate Data and Information Archive; Environment Canada: Ottawa, On tario, Canada, 2017.
- Lajoie, P.G. Étude Pédologique Des Comtés de Hull, Labelle et Papineau. Québec)(Ottawa: Ministère I’agriculture) 1967.
- Caron, A.; Jarry, J.J.; Despland, E. Early Instar Mortality of a Forest Pest Caterpillar: Which Mortality Sources Increase during an Outbreak Crash? Entomologia Exp Applicata 2022, 170, 268–276. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Agosti, D.; Alonso, L.E.; Brandão, C.R.F.; Brown, W.L.; Delabie, J.H.; Silvestre, R. Field Techniques for the Study of Ground-Dwelling Ant: An Overview, Description, and Evaluation. In Ants: Standard methods for measuring and monitoring biodiversity; 2000.
- Francœur, A. Les fourmis de la forêt boréale du Québec (Formicidae, Hymenoptera). Le Naturaliste Canadien 2001, 125, 8. [Google Scholar]
- Ellison, A.M.; Gotelli, N.J.; Farnsworth, E.J.; Alpert, G.D. A Field Guide to the Ants of New England; Yale University Press, 2012; ISBN 978-0-300-16930-0. [Google Scholar]
- Hopkin, S.P. A Key to the Collembola (Springtails) of Britain and Ireland.; FSC publications, 2007.
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part 1: Poduromorpha Fauna. Entomologica Scandinavica. Brill Academic, Leiden 1998.
- Fjellberg, A. The Collembola of Fennoscandia and Denmark, Part II: Entomobryomorpha and Symphypleona; Brill, 2007.
- Christiansen, K.; Bellinger, P. The Collembola of North America North of the Rio Grande. A Taxonomic Analysis. Part 1. Introduction. General. Families Poduridae and Hypogastruridae. Part 2. Families Onychiuridae and Isotomidae. Part 3. Family Entomobrydae. Part 4. Families Neelidae and Sminthuridae. Glossary. Bibliography. Index. The Collembola of North America north of the Rio Grande. A taxonomic analysis. Part 1. Introduction. General. Families Poduridae and Hypogastruridae. Part 2. Families Onychiuridae and Isotomidae. Part 3. Family Entomobrydae. Part 4. Families Neelidae and Sminthuridae. Glossary. Bibliography. Index. 1980.
- Sorensen, T.A. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content and Its Application to Analyses of the Vegetation on Danish Commons. Biol. Skar. 1948, 5, 1–34. [Google Scholar]
- Jaccard, P. Étude Comparative de La Distribution Florale Dans Une Portion Des Alpes et Des Jura. Bull Soc Vaudoise Sci Nat 1901, 37, 547–579. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 1.17-4. http://cran. r-project. org>. Acesso em 2010, 23, 2010. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecological monographs 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Caceres, M.; Jansen, F. Package ‘Indicspecies. ’ indicators 2016, 8. [Google Scholar]
- R Development Core, T. A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing, Department of Agronomy, Faculty …, 2017.
- Broza, M.; Izhaki, I. Post-Fire Arthropod Assemblages in Mediterranean Forest Soils in Israel. International Journal of Wildland Fire 1997, 7, 317–325. [Google Scholar] [CrossRef]
- Gardner, S.M.; Cabido, M.R.; Valladares, G.R.; Diaz, S. The Influence of Habitat Structure on Arthropod Diversity in Argentine Semi-Arid Chaco Forest. Journal of Vegetation Science 1995, 6, 349–356. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A Temporal Approach to Linking Aboveground and Belowground Ecology. Trends in ecology & evolution 2005, 20, 634–641. [Google Scholar] [CrossRef]
- Hedlund, K.; Griffiths, B.; Christensen, S.; Scheu, S.; Setälä, H.; Tscharntke, T.; Verhoef, H. Trophic Interactions in Changing Landscapes: Responses of Soil Food Webs. Basic and Applied Ecology 2004, 5, 495–503. [Google Scholar] [CrossRef]
- Anderson, K.E.; Russell, J.A.; Moreau, C.S.; Kautz, S.; Sullam, K.E.; Hu, Y.I.; Basinger, U.; Mott, B.M.; Buck, N.; Wheeler, D.E. Highly Similar Microbial Communities Are Shared among Related and Trophically Similar Ant Species. Molecular Ecology 2012, 21, 2282–2296. [Google Scholar] [CrossRef]
- Souza, R.F.; Anjos, D.V.; Carvalho, R.; Del-Claro, K. Availability of Food and Nesting-Sites as Regulatory Mechanisms for the Recovery of Ant Diversity after Fire Disturbance. Sociobiology 2015, 62, 1–9. [Google Scholar] [CrossRef]
- Frizzo, T.L.; Campos, R.I.; Vasconcelos, H.L. Contrasting Effects of Fire on Arboreal and Ground-Dwelling Ant Communities of a Neotropical Savanna. Biotropica 2012, 44, 254–261. [Google Scholar] [CrossRef]
- Silveira, J.M.; Louzada, J.; Barlow, J.; Andrade, R.; Mestre, L.; Solar, R.; Lacau, S.; Cochrane, M.A. A Multi-Taxa Assessment of Biodiversity Change after Single and Recurrent Wildfires in a Brazilian Amazon Forest. Biotropica 2016, 48, 170–180. [Google Scholar] [CrossRef]
- Vasconcelos, H.L.; Maravalhas, J.B.; Cornelissen, T. Effects of Fire Disturbance on Ant Abundance and Diversity: A Global Meta-Analysis. Biodiversity and Conservation 2017, 26, 177–188. [Google Scholar] [CrossRef]
- Calizza, E.; Rossi, L.; Careddu, G.; Sporta Caputi, S.; Costantini, M.L. Species Richness and Vulnerability to Disturbance Propagation in Real Food Webs. Sci Rep 2019, 9, 19331. [Google Scholar] [CrossRef]
- Cooke, B.J.; Sturtevant, B.R.; Robert, L.-E. The Forest Tent Caterpillar in Minnesota: Detectability, Impact, and Cycling Dynamics. Forests 2022, 13, 601. [Google Scholar] [CrossRef]
- Andersen, A.N. ; Penman; Debas; Houadria Ant Community Responses to Experimental Fire and Logging in a Eucalypt Forest of South-Eastern Australia. Forest Ecology and Management 2009, 258, 188–197. [Google Scholar] [CrossRef]
- York, A. Long-Term Effects of Frequent Low-Intensity Burning on Ant Communities in Coastal Blackbutt Forests of Southeastern Australia. Austral Ecology 2000, 25, 83–98. [Google Scholar] [CrossRef]
- Parr, C.L.; Robertson, H.G.; Biggs, H.C.; Chown, S.L. Response of African Savanna Ants to Long-Term Fire Regimes. Journal of Applied Ecology 2004, 630–642. [Google Scholar] [CrossRef]
- Maravalhas, J.; Vasconcelos, H.L. Revisiting the Pyrodiversity–Biodiversity Hypothesis: Long-Term Fire Regimes and the Structure of Ant Communities in a N Eotropical Savanna Hotspot. Journal of Applied Ecology 2014, 51, 1661–1668. [Google Scholar] [CrossRef]
- Johns, R.C.; Bowden, J.J.; Carleton, D.R.; Cooke, B.J.; Edwards, S.; Emilson, E.J.S.; James, P.M.A.; Kneeshaw, D.; MacLean, D.A.; Martel, V.; et al. A Conceptual Framework for the Spruce Budworm Early Intervention Strategy: Can Outbreaks Be Stopped? Forests 2019, 10, 910. [Google Scholar] [CrossRef]
- Lessard, J.-P.; Dunn, R.R.; Sanders, N.J. Temperature-Mediated Coexistence in Temperate Forest Ant Communities. Insect. Soc. 2009, 56, 149–156. [Google Scholar] [CrossRef]
- Despland, E.; Lessard, J.-P. Social Predation by Ants as a Mortality Source for an Arboreal Gregarious Forest Pest. Basic and Applied Ecology 2022, 59, 82–91. [Google Scholar] [CrossRef]
- Hoffmann, B.D.; Andersen, A.N. Responses of Ants to Disturbance in Australia, with Particular Reference to Functional Groups. Austral Ecology 2003, 28, 444–464. [Google Scholar] [CrossRef]
- Fotso Kuate, A.; Hanna, R.; Tindo, M.; Nanga, S.; Nagel, P. Ant Diversity in Dominant Vegetation Types of Southern Cameroon. Biotropica 2015, 47, 94–100. [Google Scholar] [CrossRef]
- Leal, I.R.; Filgueiras, B.K.C.; Gomes, J.P.; Iannuzzi, L.; Andersen, A.N. Effects of Habitat Fragmentation on Ant Richness and Functional Composition in Brazilian Atlantic Forest. Biodivers Conserv 2012, 21, 1687–1701. [Google Scholar] [CrossRef]
- Solar, R.R.d.C.; Barlow, J.; Andersen, A.N.; Schoereder, J.H.; Berenguer, E.; Ferreira, J.N.; Gardner, T.A. Biodiversity Consequences of Land-Use Change and Forest Disturbance in the Amazon: A Multi-Scale Assessment Using Ant Communities. Biological Conservation 2016, 197, 98–107. [Google Scholar] [CrossRef]
- Fairweather, A.D.; Lewis, J.H.; Hunt, L.; McAlpine, D.F.; Smith, M.A. Ants (Hymenoptera: Formicidae) of Rockwood Park, New Brunswick: An Assessment of Species Richness and Habitat. Northeastern Naturalist 2020, 27, 576. [Google Scholar] [CrossRef]
- Milford, E.R. Ant Communities in Flooded and Unflooded Riparian Forest of the Middle Rio Grande. The Southwestern Naturalist 1999, 278–286. [Google Scholar]
- Ellison, A.M.; Farnsworth, E.J.; Gotelli, N.J. Ant Diversity in Pitcher-Plant Bogs of Massachusetts. Northeastern Naturalist 2002, 9, 267–284. [Google Scholar] [CrossRef]
- Francoeur, A. Deux Nouvelles Fourmis Néarctiques: Leptothorax Retractus et L. Sphagnicolus (Formicidae, Hymenoptera). the Canadian entomologist 1986, 118, 1151–1164. [Google Scholar] [CrossRef]
- Francoeur, A. Revision Taxonomique Des Especes Nearctiques Du Groupe/Usca, Genre Formica (Formicidae, Hymenoptera). Mem. Soc. Ent. Queb. 3. 316 Pp.. 1979. Formicoidea. Canada and its Insect Fauna. Mem. ent. Soc. Can 1973, 108, 502–503. [Google Scholar]
- Francoeur, A. Extension de l’aire Connue de La Fourmi Myrmica Quebecensis (Formicidae, Hymenoptera). 2011.
- Oberg, E.; Del Toro, I.; Pelini, S. Characterization of the Thermal Tolerances of Forest Ants of New England. Insectes sociaux 2012, 59, 167–174. [Google Scholar] [CrossRef]
- Wheeler, G.C.; Wheeler, J. others The Ants of North Dakota; University of North Dakota Grand Forks, 1963.
- MacKay, W.P.; Mackay, E. The Ants of New Mexico (Hymenoptera: Formicidae); Edwin Mellen Press Lewiston, NY, 2002.
- Sirois, L. Impact of Fire on Picea Mariana and Pinus Banksiana Seedlings in Subarctic Lichen Woodlands. Journal of Vegetation Science 1993, 4, 795–802. [Google Scholar] [CrossRef]
- Rosengren, R. The Interaction between Red Wood Ants, Cinara Aphids, and Pines. A Ghost of Mutualism Past? In Ant-plant interactions; 1991; pp. 80–91.
- Parmentier, T.; Dekoninck, W.; Wenseleers, T. A Highly Diverse Microcosm in a Hostile World: A Review on the Associates of Red Wood Ants (Formica Rufa Group). Insect. Soc. 2014, 61, 229–237. [Google Scholar] [CrossRef]
- Kilpeläinen, J.; Finér, L.; Niemelä, P.; Domisch, T.; Neuvonen, S.; Ohashi, M.; Risch, A.C.; Sundström, L. Carbon, Nitrogen and Phosphorus Dynamics of Ant Mounds (Formica Rufa Group) in Managed Boreal Forests of Different Successional Stages. Applied Soil Ecology 2007, 36, 156–163. [Google Scholar] [CrossRef]
- Serttaş, A.; Bakar, Ö.; Alkan, U.M.; Yılmaz, A.; Yolcu, H.I.; Ipekdal, K. Nest Survival and Transplantation Success of Formica Rufa (Hymenoptera: Formicidae) Ants in Southern Turkey: A Predictive Approach. Forests 2020, 11, 533. [Google Scholar] [CrossRef]
- Stockan, J.A.; Rao, S.; Pakeman, R. Nesting Preferences of the Threatened Wood Ant Formica Exsecta (Hymenoptera: Formicidae); Implications for Conservation in Scotland. J Insect Conserv 2010, 14, 269–276. [Google Scholar] [CrossRef]
- Frouz, J. The Effect of Nest Moisture on Daily Temperature Regime in the Nests of Formica Polyctena Wood Ants. Insectes sociaux 2000, 47, 229–235. [Google Scholar] [CrossRef]
- Frouz, J.; Rybníček, M.; Cudlín, P.; Chmelíková, E. Influence of the Wood Ant, Formica Polyctena, on Soil Nutrient and the Spruce Tree Growth. Journal of Applied Entomology 2008, 132, 281–284. [Google Scholar] [CrossRef]
- Domisch, T.; Ohashi, M.; Finér, L.; Risch, A.C.; Sundström, L.; Kilpeläinen, J.; Niemelä, P. Decomposition of Organic Matter and Nutrient Mineralisation in Wood Ant (Formica Rufa Group) Mounds in Boreal Coniferous Forests of Different Age. Biol Fertil Soils 2008, 44, 539–545. [Google Scholar] [CrossRef]
- Rosengren, R.; Fortelius, W.; Lindström, K.; Luther, A. Phenology and Causation of Nest Heating and Thermoregulation in Red Wood Ants of the Formica Rufa Group Studied in Coniferous Forest Habitats in Southern Finland. In Proceedings of the Annales Zoologici Fennici; JSTOR; 1987; pp. 147–155. [Google Scholar]
- Weseloh, R.M. Simulation of Predation by Ants Based on Direct Observations of Attacks on Gypsy Moth Larvae. 1989. [CrossRef]
- Mokadam, C. Native and Non-Native Ant Impacts on Native Fungi. 2021.
- Sabelis, M.W. Predatory Arthropods. Natural enemies: The population biology of predators, parasites and diseases 1992, 225–264.
- Settle, W.H.; Ariawan, H.; Astuti, E.T.; Cahyana, W.; Hakim, A.L.; Hindayana, D.; Lestari, A.S. Managing Tropical Rice Pests through Conservation of Generalist Natural Enemies and Alternative Prey. Ecology 1996, 77, 1975–1988. [Google Scholar] [CrossRef]
- Symondson, W.; Sunderland, K.; Greenstone, M. Can Generalist Predators Be Effective Biocontrol Agents. Annu. Rev. Èntomol. 2002, 47, 561–594. [Google Scholar] [CrossRef]
- Yang, L.H. Interactions between a Detrital Resource Pulse and a Detritivore Community. Oecologia 2006, 147, 522–532. [Google Scholar] [CrossRef]
- Karban, R. Increased Reproductive Success at High Densities and Predator Satiation for Periodical Cicadas. Ecology 1982, 63, 321–328. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Vannote, R.L. Population Synchrony in Mayflies: A Predator Satiation Hypothesis. Evolution 1982, 810–821. [Google Scholar] [CrossRef]
- Williams, K.S.; Smith, K.G.; Stephen, F.M. Emergence of 13-Yr Periodical Cicadas (Cicadidae: Magicicada): Phenology, Mortality, and Predators Satiation. Ecology 1993, 74, 1143–1152. [Google Scholar] [CrossRef]
- Yang, L.H.; Edwards, K.F.; Byrnes, J.E.; Bastow, J.L.; Wright, A.N.; Spence, K.O. A Meta-Analysis of Resource Pulse–Consumer Interactions. Ecological Monographs 2010, 80, 125–151. [Google Scholar] [CrossRef]
- Eveleigh, E.S.; McCann, K.S.; McCarthy, P.C.; Pollock, S.J.; Lucarotti, C.J.; Morin, B.; McDougall, G.A.; Strongman, D.B.; Huber, J.T.; Umbanhowar, J.; et al. Fluctuations in Density of an Outbreak Species Drive Diversity Cascades in Food Webs. Proceedings of the National Academy of Sciences 2007, 104, 16976–16981. [Google Scholar] [CrossRef]
- Dansereau-Macias, É.; Despland, E.; Handa, I.T. Decreased Soil Microbial Biomass and Changed Microbial Community Composition Following a Defoliation Event by the Forest Tent Caterpillar. Forests 2023, 14, 792. [Google Scholar] [CrossRef]
- Kristensen, J.Å.; Rousk, J.; Metcalfe, D.B. Below-ground Responses to Insect Herbivory in Ecosystems with Woody Plant Canopies: A Meta-analysis. J Ecol 2020, 108, 917–930. [Google Scholar] [CrossRef]
Figure 1.
Species richness and evenness. The two panels on the right refer to the boreal forest data while the two left panels refer to the temperate forest data. Box plots represent data from pitfall traps only and from a subset of years for the boreal forest samples (2017-2018) to limit the variation in sampling method from 2016 and 2019 respectively. In the box plots, the lower boundary of the box indicates the 25th percentile, the bold line within the box marks the median, and the upper boundary of the box indicates the 75th percentile. The whiskers indicate the 10th and 90th percentiles. Dots are outliers (>Q3 + 1.5 × interquartile range).
Figure 1.
Species richness and evenness. The two panels on the right refer to the boreal forest data while the two left panels refer to the temperate forest data. Box plots represent data from pitfall traps only and from a subset of years for the boreal forest samples (2017-2018) to limit the variation in sampling method from 2016 and 2019 respectively. In the box plots, the lower boundary of the box indicates the 25th percentile, the bold line within the box marks the median, and the upper boundary of the box indicates the 75th percentile. The whiskers indicate the 10th and 90th percentiles. Dots are outliers (>Q3 + 1.5 × interquartile range).
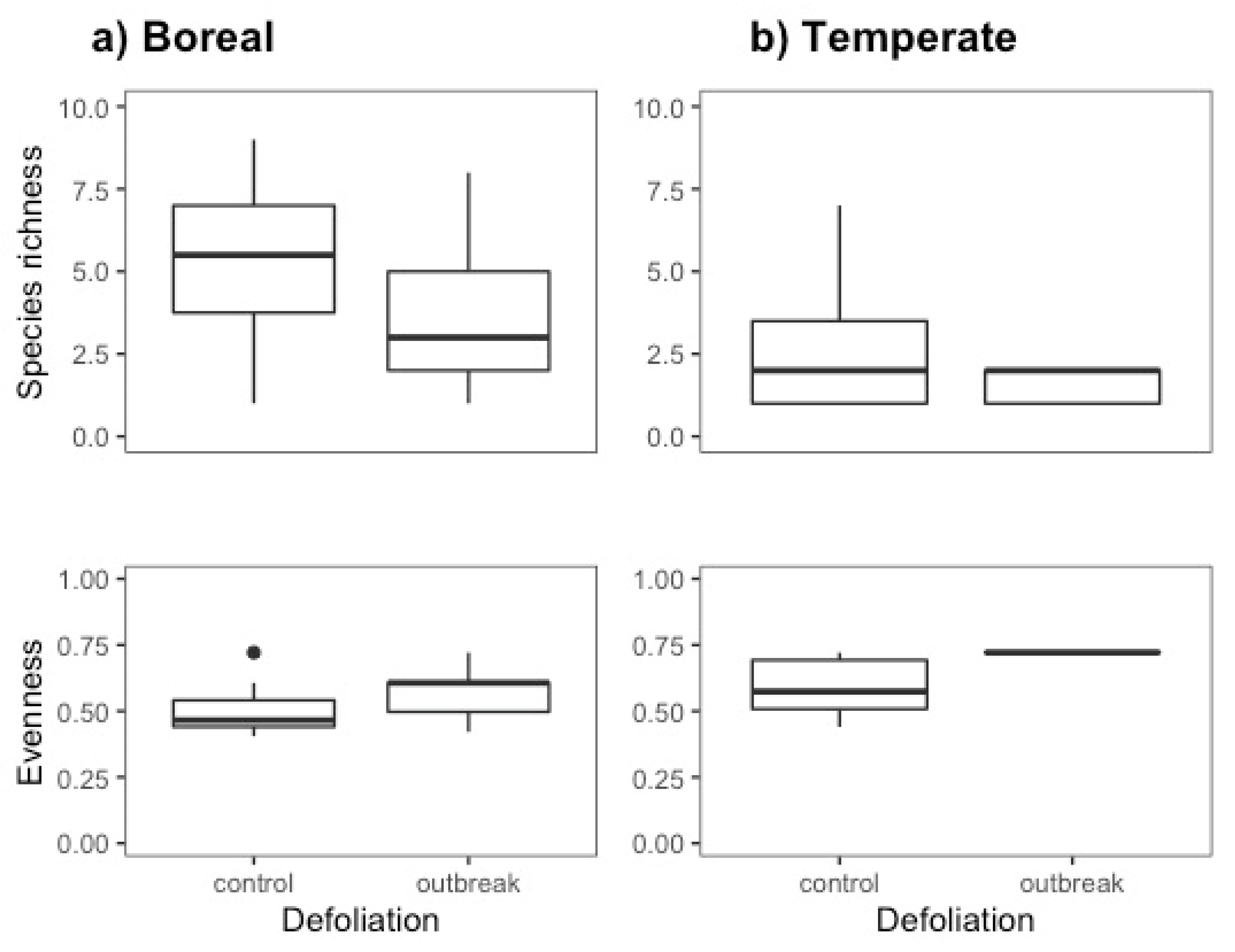
Figure 2.
NMDS ordination of boreal forest sites within control and outbreak stands based on the Sørensen dissimilarity index using taxonomic data. The abundance of predators was the only covariate that aligned significantly with ant community composition.
Figure 2.
NMDS ordination of boreal forest sites within control and outbreak stands based on the Sørensen dissimilarity index using taxonomic data. The abundance of predators was the only covariate that aligned significantly with ant community composition.
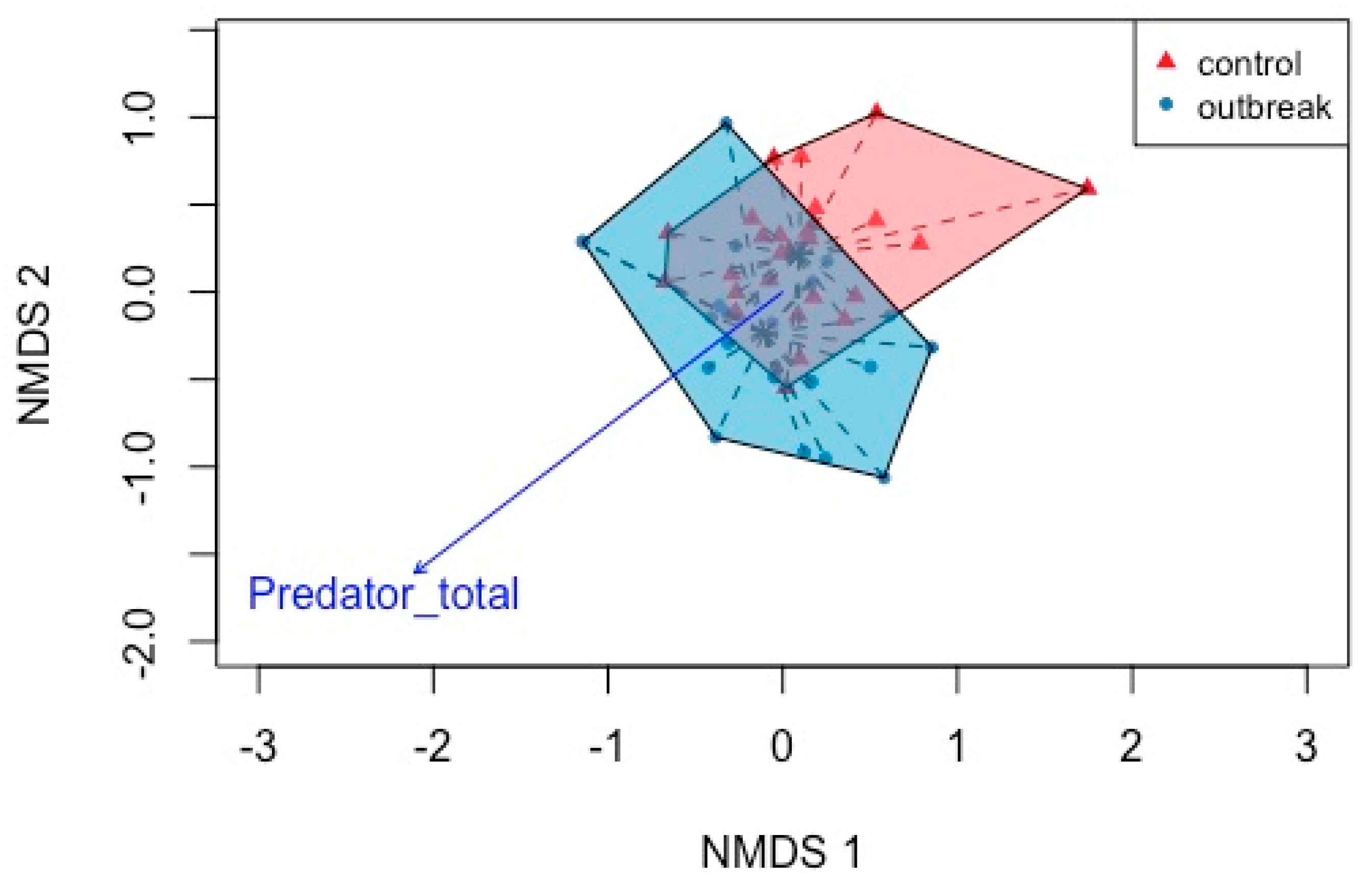
Figure 3.
Boxplot of the standardized effect size of the distance to multivariate space centroid (SES Dcentroid) of each site in a) boreal and b) temperate. The black dashed line represents null expectations. In the box plots, the lower boundary of the box indicates the 25th percentile, the bold line within the box marks the median, and the upper boundary of the box indicates the 75th percentile. The whiskers indicate the 10th and 90th percentiles. Dots are outliers (>Q3 + 1.5 × interquartile range).
Figure 3.
Boxplot of the standardized effect size of the distance to multivariate space centroid (SES Dcentroid) of each site in a) boreal and b) temperate. The black dashed line represents null expectations. In the box plots, the lower boundary of the box indicates the 25th percentile, the bold line within the box marks the median, and the upper boundary of the box indicates the 75th percentile. The whiskers indicate the 10th and 90th percentiles. Dots are outliers (>Q3 + 1.5 × interquartile range).
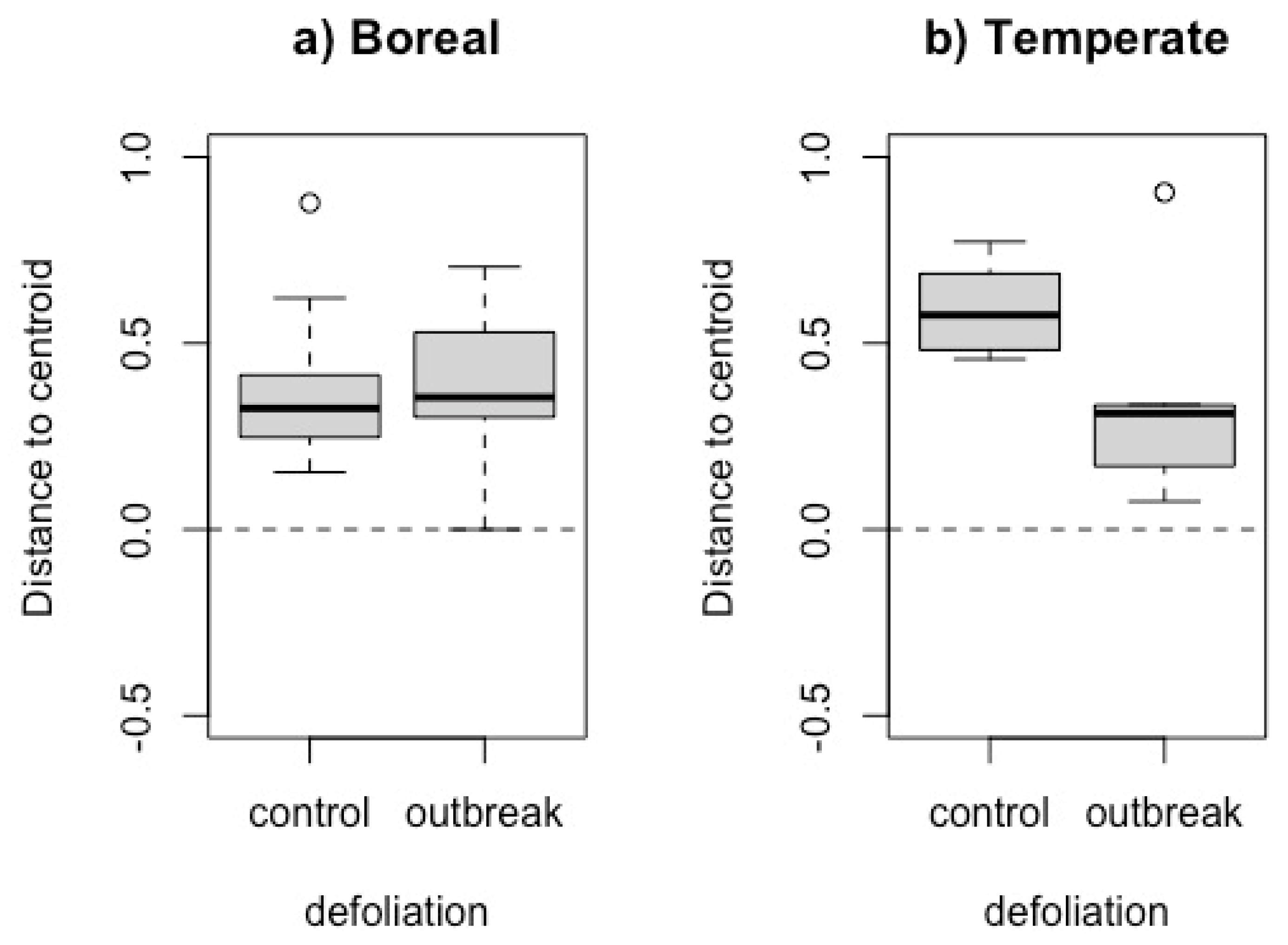

Table 2.
Summary of statistics for PERMANOVAs of the effect of defoliation history on taxonomic compositional turnover of ant communities. Significant values (p < 0.05) are highlighted in bold.
Table 2.
Summary of statistics for PERMANOVAs of the effect of defoliation history on taxonomic compositional turnover of ant communities. Significant values (p < 0.05) are highlighted in bold.
| Boreal forest | Temperate forest | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | R2 | p-value | df | F | R2 | p-value | ||
| Defoliation history | 1 | 5.3914 | 0.09096 | 0.002 | Defoliation history | 1 | 1.4536 | 0.10198 | 0.249 |
| Canopy openness | 1 | 0.9813 | 0.01656 | 0.434 | Canopy openness | 1 | 0.3802 | 0.02667 | 0.830 |
| Predators | 1 | 4.0690 | 0.06865 | 0.004 | Predators | 1 | 0.7809 | 0.05478 | 0.567 |
| Collembola (abundance) | 1 | 0.5300 | 0.00894 | 0.764 | Collembola (abundance) | 1 | 1.6389 | 0.11498 | 0.181 |
| Collembola (species richness) | 1 | 1.2997 | 0.02193 | 0.283 | |||||
| Residuals | 47 | 0.79296 | 14 | 0.70158 | |||||
Table 3.
Indicator species analysis for the boreal forest data only at alpha = 0.05. The “Indicator value index” measures the association between a species and a site group. “A” is the positive predictive value of the species as an indicator of the site group and “B” is the sensitivity of the species as an indicator of the target site group.
Table 3.
Indicator species analysis for the boreal forest data only at alpha = 0.05. The “Indicator value index” measures the association between a species and a site group. “A” is the positive predictive value of the species as an indicator of the site group and “B” is the sensitivity of the species as an indicator of the target site group.
| Groups | Species | A | B | Indicator value index | p-value |
|---|---|---|---|---|---|
| Control | Camponotus herculeanus | 0.7812 | 0.5714 | 0.668 | 0.002 |
| Myrmica sp1 | 0.9146 | 0.4286 | 0.626 | 0.003 | |
| Myrmica detritinodis | 0.8893 | 0.3214 | 0.535 | 0.011 | |
| Formica sp1 | 1.0000 | 0.2857 | 0.535 | 0.004 | |
| Outbreak | Formica integra | 1.0 | 0.2 | 0.447 s | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



