
Submitted:
02 May 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
Soil-borne oomycetes include devastating plant pathogens that cause substantial losses in the agricultural sector. To better manage this important group of pathogens, it is critical to understand how they respond to common agricultural practices, such as tillage and crop rotation. Here, a long-term field experiment was established using a split-plot design with tillage as the main plot factor (conventional tillage [CT] vs. no till [NT], 2 levels) and rotation as the subplot factor (monocultures of soybean, corn, or wheat, and corn-soybean-wheat rotation, 4 levels). Post-harvest soil oomycete communities were characterized over three consecutive years (2016-2018) by metabarcoding the Internal Transcribed Spacer 1 (ITS1) region. The community contained 292 Amplicon Sequence Variants (ASVs) and was dominated by Globisporangium spp. (85.1% in abundance, 203 ASV) and Pythium spp. (10.4%, 51 ASV). NT decreased diversity and community compositional structure heterogeneity, while crop rotation only affected the community structure under CT. The interaction effects of tillage and rotation on most oomycetes species accentuated the complexity of managing these pathogens. Soil and crop health represented by soybean seedling vitality was lowest in soils under CT cultivating soybean or corn, while grain yield of the three crops responded differently to tillage and crop rotation regimes.
Keywords:
crop rotation
; metabarcoding
; soil-borne oomycetes
; soybean
; tillage
1. Introduction
The oomycetes (or Oomycota) contains a group of fungal-like microorganisms within the kingdom Stramenopila, of which 60% of the species are considered pathogenic biotrophs, hemibiotrophs, or necrotrophs [1], and of great concern in agriculture [2], aquaculture [3], forestry, and natural ecosystems [4]. Depending on their host range, plant pathogens in the oomycetes are considered generalist or specialist species, and such information often determines the control measures for pathogens in their respective categories. For example, Globisporangium ultimum is a pathogen with a wide host range that is associated with seedling damping off disease complexes. As a generalist pathogen, G. ultimum is managed in plant production systems through fungicidal seed treatments specific to oomycetes [5]. Other oomycete pathogens are more limited in their host range, such as Phytopthora (Ph.) sojae, known only to be pathogenic on soybean (Glycine max) and lupins (Lupinus). Alongside chemical control measures, Ph. sojae in soybean is managed through single gene resistance pathways, or combinations of these genes to increase pathotype resistance [6]. The diversity of pathogen-host interactions and host specificity among soil-borne plant pathogenic oomycetes makes them a difficult group to manage. Moreover, soil-borne oomycetes are susceptible to changes in soil texture and organic matter, with a preference for cool, moist soils [7,8]. Studies have shown that agronomic practices, like tillage and crop rotation, affect soil physicochemical properties and reshape the soil-borne microbiome (including oomycetes community) structure, and consequently can be disruptive to soil health and fertility. Understanding how soil-borne oomycete communities respond to these common agronomic practices, would help clarify best-practices for regions with a high incidence of oomycete plant disease and provide the fundamental basis for establishing effective pest management and mitigation strategies for these important phytopathogens.
The use of tillage and crop rotation in managing soil-borne oomycetes may reduce the pathogen inocula or improve the soil’s natural capacity to suppress pathogenicity [9-11]. Conventional tillage (CT) usually involves fall moldboard ploughing and spring cultivation, and is practiced to reduce weed establishment [12] and soil compaction [13]. Practicing CT can lead to the loss of soil tilth, increased nutrient runoff, reduced soil quality, and disruption of the soil microbiome [14]. By contrast, no-till (NT) promoted beneficial fungal and bacterial taxa, compared to CT, under which the soils were enriched with plant pathogens, as reported by Srour et al [15]. The diversity of bacterial taxa was also found to be lower in topsoil, and higher in the deeper soil layers under CT compared to NT [16]. A greater microbial species diversity in soils under NT may lead to a more complex inter-species network which may reinforce the suppression effects of beneficial microorganisms against the proliferation and growth of pathogenic species [17]. CT-mediated soil surface drying has been suggested as a management strategy since successful infection of the host by many oomycete species is dependent on zoospore mobility, supported by high soil water content [18].
Crop rotation, as a common agricultural practice, involves planting alternative crops sequentially on the same farmland for improved soil fertility and control of weeds, pests, and diseases. It has traditionally been encouraged to manage plant pathogens by mediating the availability of host plants from year to year [10,19,20]. Plant disease incidence and associated yields are impacted by the selection of crops in a cropping system and how they are rotated, with monoculture having reduced yields when compared to rotations involving other crops [21,22]. Crop rotation enriched plant growth promoting bacterial [23], and disease suppressive functional groups, such as those carrying the prnD gene that encodes the antifungal compound pyrrolnitrin [19]. Monoculture tomato soils were enriched in fungal genera containing potential pathogens, such as Pseudogymnoascus, Fusarium, and Pyrenochaeta, compared to soils under crop rotation [22].
Hwang, et al. [24] found that the levels of Pythium (Py.) inocula in monoculture soils, particularly pea, were greater than in rotation soils, which was reflected in disease incidence. Pythium and Phytophthora were the dominant genera recovered in a soybean-corn cropping system [25-27]. Oomycete pathogenicity on soybean, in particular, has been extensively studied and over 15 species of oomycetes, such as Py. Aphanidenmatum, G. ultimum, G. irregulare, G. cryptoirregulare, have been shown to be pathogenic on soybean; although, direct inoculation of soils with these recovered oomycete pathogens does not always reflect the severity of disease symptoms observed in soybean [25]. The relationship between oomycete plant pathogens and soybean seed make soybean an ideal initial crop to study disease symptoms in relation to oomycete species diversity and distribution in soils of varying rotation and tillage backgrounds. The drive for maximizing yields of high-value crops can often compete with the benefits of diversifying crop rotation systems. A thorough understanding of the potential disease ramifications due to rotation selection, especially within the oomycetes where little is known about the influence of crop rotation on community structure, may help to strengthen guidelines for more productive rotations.
This study aimed to explore the shifts in the soil-borne oomycete community in response to different combinations of tillage and crop rotation practices. More specifically, we characterized the post-harvest soil oomycete communities over three consecutive years (2016-2018) at a long-term experimental site by metabarcoding the Internal Transcribed Spacer 1 (ITS1) region. The experimental field was established using a split-plot design (two tillage levels, NT vs. CT), with four rotations arranged randomly within each tillage treatment (Supplementary Fig. S1). The rotations included monocultures of dicot soybean (SSS), monocot corn (Zea mays, CCC), and monocot wheat (Triticum aestivum, WWW), as well as corn-soybean-wheat (CSW) rotation which change the available host range for oomycetes [28]. We hypothesized that prolonged tillage and rotation practices can 1) exert a significant influence on the diversity and abundance of microbial communities, which may lead to a consistent increase or decrease trend over the course of the three-year study period, and 2) affect crop and soil health as represented by crop yield and seedling vitality of soybean. This study aimed to provide guidelines for better agricultural management practices in managing soil-born oomycetes.
2. Results
A total of 3,148,276 high-quality reads were retained in the final amplicon sequencing variants (ASV) abundance table, with 30,272 ± 5,785 (MEAN ± SD) reads per sample. In total, 292 ASVs (mean ± SD = 9 ± 3 per sample) were recovered from all of the samples.
2.1. Soil-borne oomycete community diversity and compositional structure
The alpha-diversity indices of the soil oomycete community were significantly affected by tillage practices, with a higher Shannon-based True Diversity Index (Shannon-TD; P = 0.007) and a Simpson-based True Diversity Index (Simpson-TD; P = 0.014) under CT than NT (Table 1, Fig. 1). Additionally, Chao1 index was significantly (P = 0.001) affected by the interaction of tillage and rotation, with higher values under CT than NT under CSW, but not under monoculture. We also observed that CSW increased Chao1 richness in comparison with CCC monoculture, but only under CT not NT. We did not observe significant differences in soil moisture content between the tillage and rotation treatments (Supplementary Fig. S2A). Correlation analysis also showed no significant relationship between the soil moisture content and alpha-diversity indices (P > 0.05, Supplementary Fig. S2B-D), or between the soil moisture content and the recovered oomycetes species (P > 0.05).
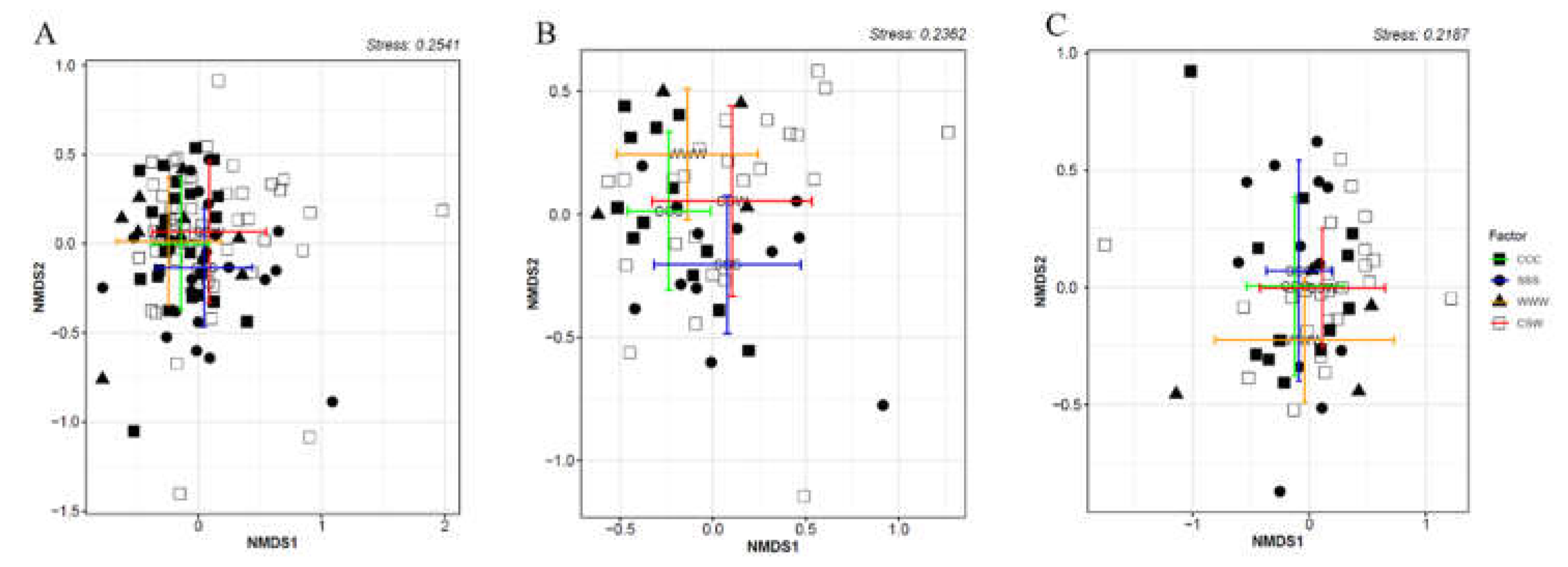
The permutational multivariate analysis of variance (PERMANOVA) results showed that soil oomycete community structure was significantly affected by tillage (F = 4.83, P = 0.001; Fig. 2B) and the interaction of tillage and rotation (F = 1.82, P = 0.01). We observed that rotation significantly affected the oomycete community structure under CT (F = 2.01, P = 0.003, Fig 2B), but not under NT (F = 1.20, P = 0.175; Fig. 2B,C). Pairwise comparison and non-metric multidimensional scaling (NMDS) results showed that the soil oomycete community structure under CT differed significantly among the three monoculture treatments (F = 2.09, P < 0.001; Fig. 2B). However, such differences were not observed between the crops within the CSW rotation (F = 0.98, P = 0.468). PERMANOVA analysis indicated insignificant impact of soil moisture content on the beta-diversity of the soil-borne oomycetes community (P > 0.05).
Figure 1.
The alpha-diversity indices of the oomycetes communities as affected by tillage (A, B) or the interaction of tillage and rotation (C). Chao1, Chao1 richness index; CT, conventional tillage, NT, no-till; CCC, monoculture of corn, SSS, monoculture of soybean, WWW, monoculture of wheat, CSW, rotation of corn-soybean-wheat. Different letters across the treatments represent significant difference at α < 0.05 based on Tukey’s HSD test. Error bars represent one standard error.
Figure 1.
The alpha-diversity indices of the oomycetes communities as affected by tillage (A, B) or the interaction of tillage and rotation (C). Chao1, Chao1 richness index; CT, conventional tillage, NT, no-till; CCC, monoculture of corn, SSS, monoculture of soybean, WWW, monoculture of wheat, CSW, rotation of corn-soybean-wheat. Different letters across the treatments represent significant difference at α < 0.05 based on Tukey’s HSD test. Error bars represent one standard error.
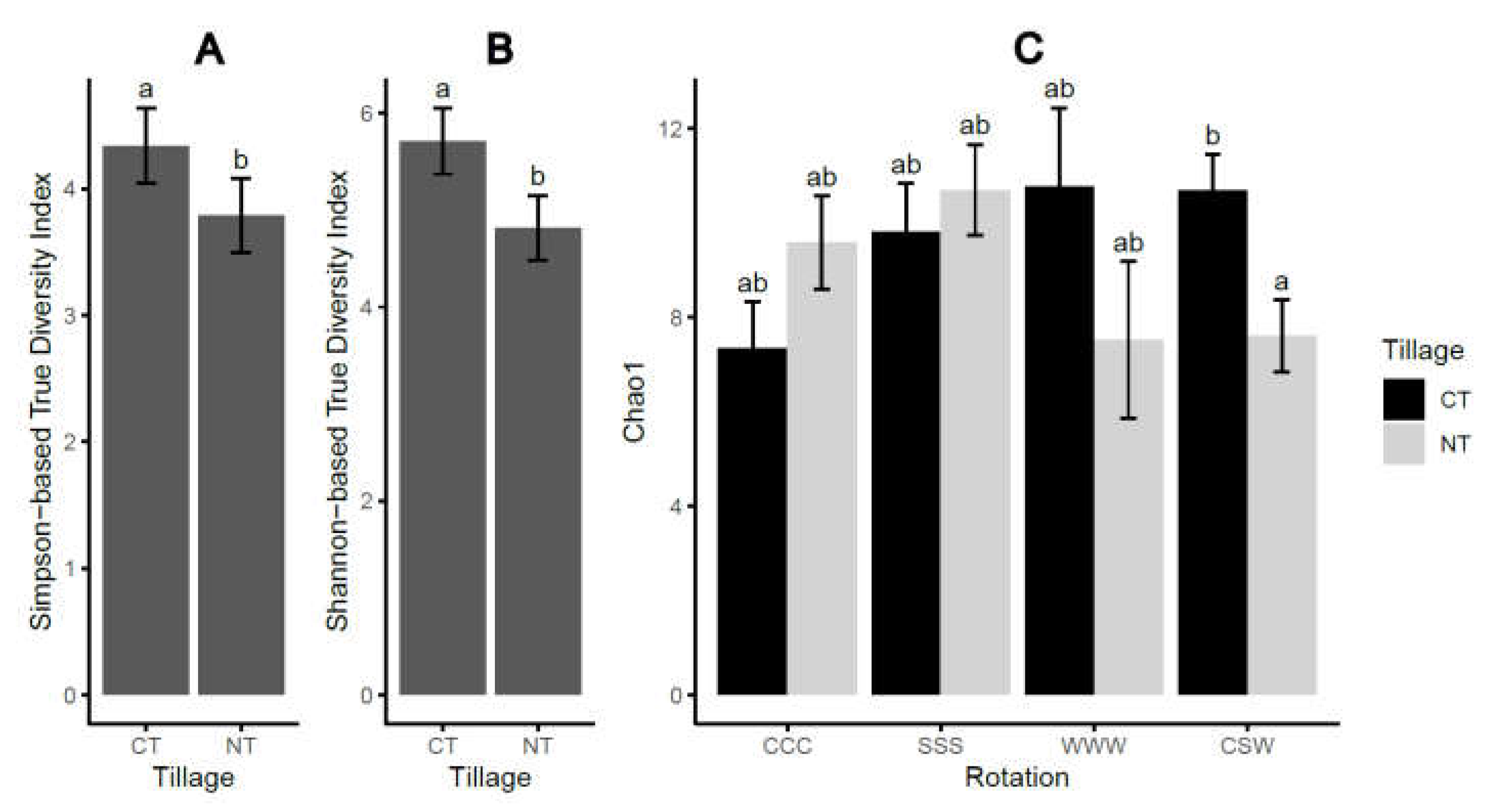
Figure 2.
The soil oomycetes community structure as affected by tillage (A) and rotation under CT (B) or under NT (C) as determined by NMDS and PERMANOVA. CT, conventional tillage; NT, no-till; CCC, monoculture of corn; SSS, monoculture of soybean; WWW, monoculture of wheat; CSW, rotation of corn-soybean-wheat. The central dots represent the means of the points on the two NMDS axes for respective groups, the bars represent one standard deviation from the mean along both axes.
Figure 2.
The soil oomycetes community structure as affected by tillage (A) and rotation under CT (B) or under NT (C) as determined by NMDS and PERMANOVA. CT, conventional tillage; NT, no-till; CCC, monoculture of corn; SSS, monoculture of soybean; WWW, monoculture of wheat; CSW, rotation of corn-soybean-wheat. The central dots represent the means of the points on the two NMDS axes for respective groups, the bars represent one standard deviation from the mean along both axes.
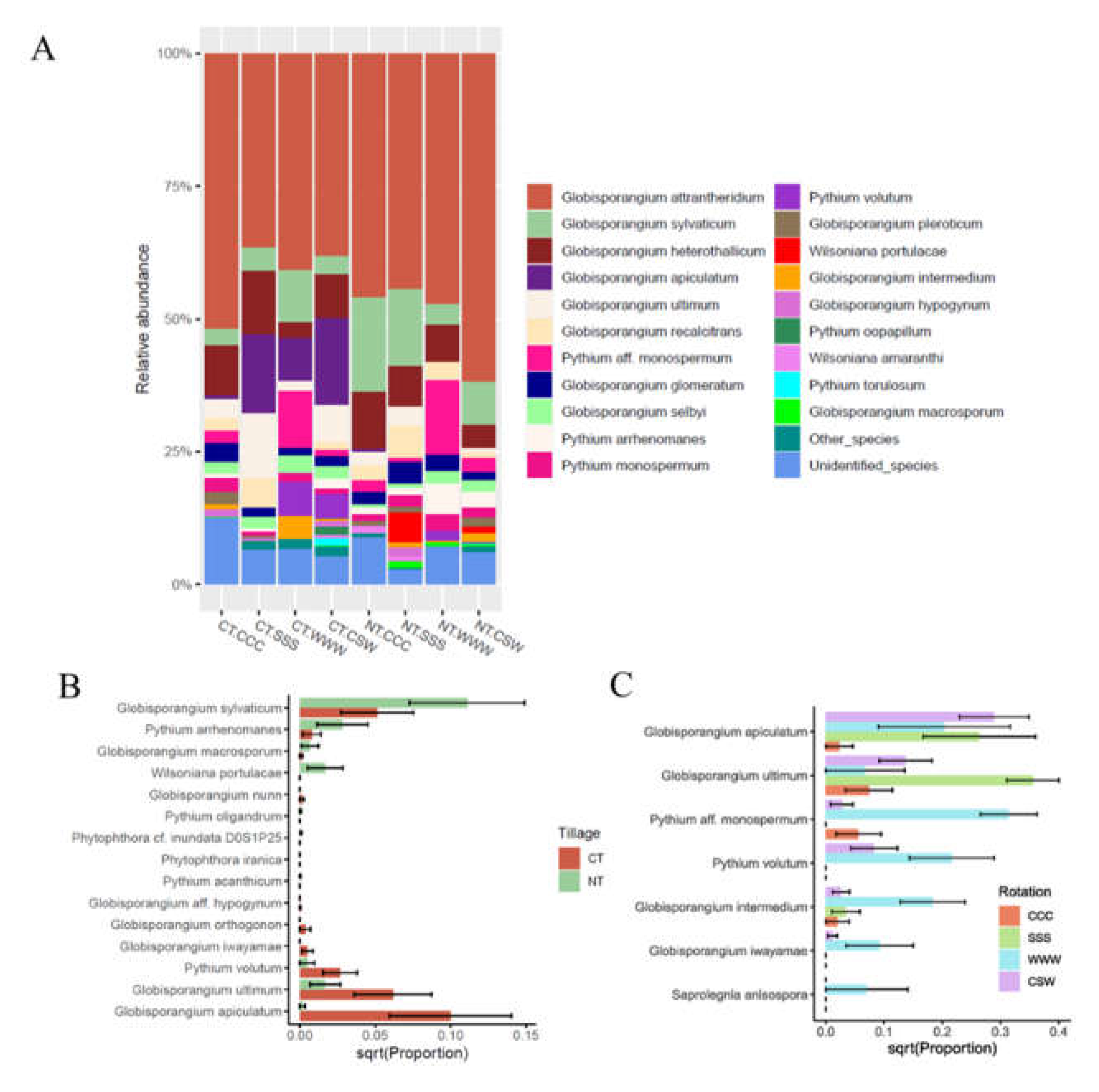
The oomycete ITS1 ASVs were assigned to six families, seven genera, and 34 species. The most predominant genera were Globisporangium (85.1%, 203 ASV), Pythium (10.4%, 51 ASV), and Wilsoniana (1.3%, 7 ASV). All other oomycete genera were represented by < 1% ASVs. Of the species identified, the most abundant were generalist species of Globisporangium, including G. attrantheridium (47.0%), G. heterothallicum (7.88%), G. sylvaticum (7.78%), G. apiculatum (5.92%), and G. ultimum (4.36%; Fig. 3A). Phylogenetic reconstruction using the representative sequences of the ASVs confirmed the accuracy of the species-level classifications (Supplementary Fig. S3). Of all identified oomycetes species, 22 were observed both under CT and under NT, six were only observed under CT, and six were only observed under NT (Table 2, Fig. 3B). Tillage had a significant (P < 0.05) effect on eight species (Table 2), with higher abundances of G. macrosporum, G. sylvaticum, Py. arrhenomanes, and Wilsoniana portulacae under NT than under CT, and higher abundances of G. iwayamae, G. ultimum, G. apiculatum, and Py. volutum under CT than under NT (Fig. 3B). Additionally, rotation had a significant effect on seven species under CT, with enriched abundances in G. ultimum under soybean monoculture, as well as G. iwayamae, Pythium sp. aff. monospermum, Py. volutum, and Saprolegnia anisospore under wheat monoculture (Table 2, Fig. 3C). We observed a low abundance of Pythium sp. aff. monospermum under CSW but a high abundance of this species (an average of 10.6%) under wheat monoculture (Table 2). G. G. iwayamae showed the same trend and was recovered in low abundance under CSW (with relative abundance of 0.2%), and in high abundance under wheat monoculture (with relative abundance of and 2.0%). By contrast, S. anisospore was abundant with 2.0% under wheat monoculture, but was not recovered under CSW (Table 2). Only Py. volutum was significantly affected by rotation under NT (Table 2).
Figure 3.
The abundance of oomycetes genera and species. (A) The 20 most abundant species as affected by tillage and rotation; Other_species, identified species that were not among the 20 most abundant species; unidentified species, sequences that were not assigned to a known species. (B) Oomycetes species that were significantly affected by tillage (P < 0.05, linear mixed effect model) and those were only found under CT or NT. (C) Oomycetes species that were significantly affected by rotation under CT (P < 0.05). Error bars represent standard error. CT, conventional tillage, NT, no-till; CCC, monoculture of corn, SSS, monoculture of soybean, WWW, monoculture of wheat, CSW, rotation of corn-soybean-wheat.
Figure 3.
The abundance of oomycetes genera and species. (A) The 20 most abundant species as affected by tillage and rotation; Other_species, identified species that were not among the 20 most abundant species; unidentified species, sequences that were not assigned to a known species. (B) Oomycetes species that were significantly affected by tillage (P < 0.05, linear mixed effect model) and those were only found under CT or NT. (C) Oomycetes species that were significantly affected by rotation under CT (P < 0.05). Error bars represent standard error. CT, conventional tillage, NT, no-till; CCC, monoculture of corn, SSS, monoculture of soybean, WWW, monoculture of wheat, CSW, rotation of corn-soybean-wheat.
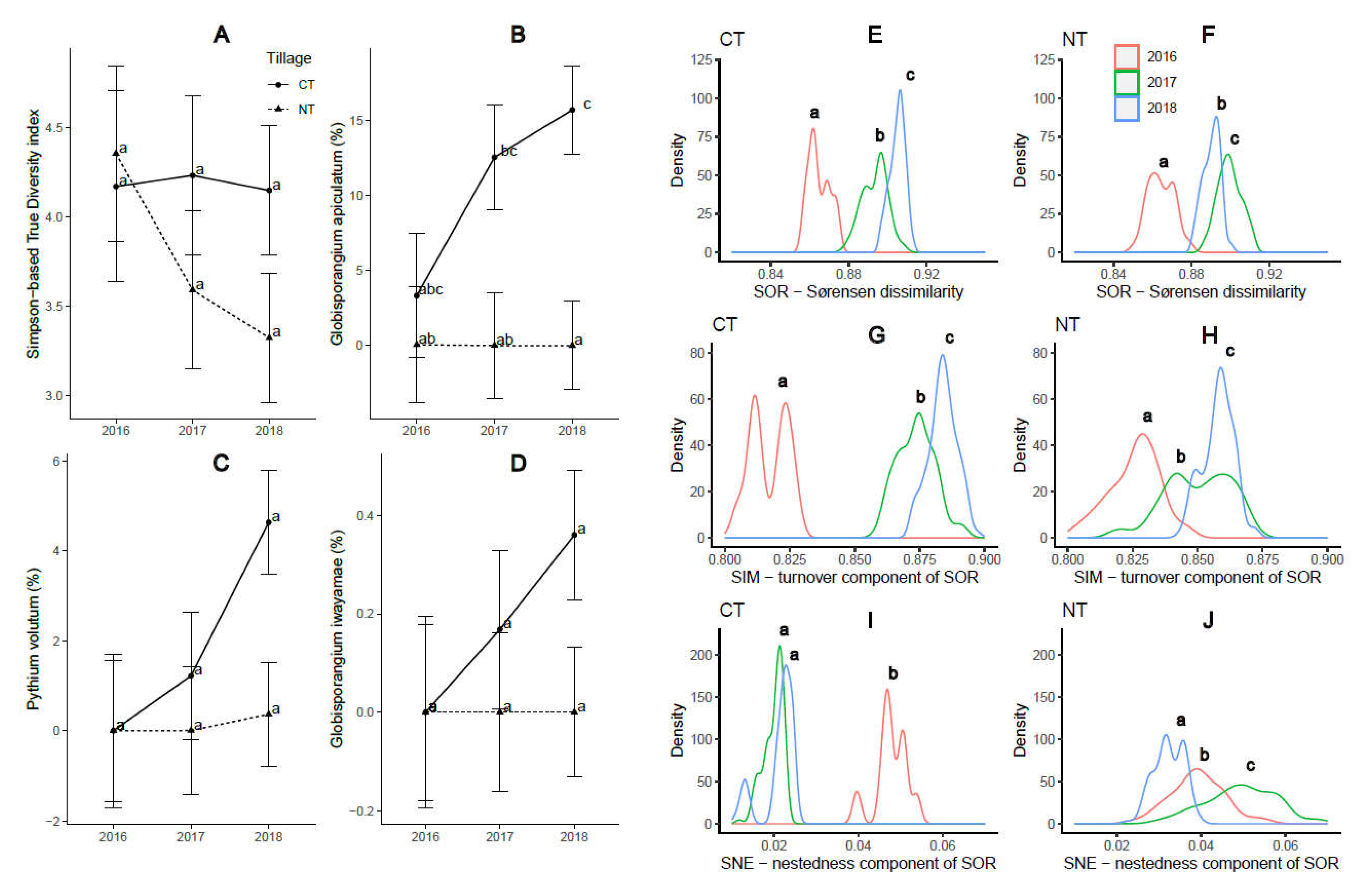
Since tillage practices showed higher impact on the oomycetes community assemblage than the rotation patterns did, one of our hypothesis was that the implementation of long-term tillage regimes would lead to a consistent trend of microbial abundance and diversity throughout the duration of the three-year study. Indeed, the Simpson-TD showed an insignificant but persistent decrease trend over the three years under NT but not under CT (Fig. 4A), suggesting a potential of NT in reducing the number of abundant oomycetes species. We also observed a persistent increase in the abundance of G. apiculatum, Py. volutum, and G. iwayamae under CT but not under NT, although such changes may not be statistically significant (Fig. 4B-D). More interestingly, under CT, the community overall dissimilarity (Sørensen dissimilarity, SOR) and the turnover component of the Sørensen dissimilarity (Simpson dissimilarity, SIM) increased over the three growing seasons, while the nestedness compoenent of the Sørensen dissimilarity (SNE) showed an opposite trend (P ≤ 0.05) (Fig. 4E, G, I). Such a trend, however, was not observed under NT. The yearly difference in SIM and SNE components, especially between 2016 and the other two years, was more than 2-fold under CT compared to under NT (Fig. 4F, H, J). These findings suggest that continuous CT practices may lead to a decreased homogeneity of the soil oomycetes community.
Figure 4.
The impact of tillage practices on oomycetes community diversity and species abundance over the three-year study period (2016-2018). A) Simpson-based True Diversity Index decreased under NT but not under CT; the relative abundance of B) Globisporangium apiculatum, C) Pythium volutum, and D) G. iwayamae increased under CT but not under NT. E-J) The impact of tillage practices on the beta-diversity of oomycetes community: E-F) the overall community dissimilarity (Sørensen dissimilarity, SOR) and G-H) the turnover component of the Sørensen dissimilarity (Simpson dissimilarity, SIM) increased over the three growing season; I-J) the nestedness compoenent of the Sørensen dissimilarity (SNE) decreased over the three years under CT but not under NT.
Figure 4.
The impact of tillage practices on oomycetes community diversity and species abundance over the three-year study period (2016-2018). A) Simpson-based True Diversity Index decreased under NT but not under CT; the relative abundance of B) Globisporangium apiculatum, C) Pythium volutum, and D) G. iwayamae increased under CT but not under NT. E-J) The impact of tillage practices on the beta-diversity of oomycetes community: E-F) the overall community dissimilarity (Sørensen dissimilarity, SOR) and G-H) the turnover component of the Sørensen dissimilarity (Simpson dissimilarity, SIM) increased over the three growing season; I-J) the nestedness compoenent of the Sørensen dissimilarity (SNE) decreased over the three years under CT but not under NT.
2.2. Crop yield and soybean seedling emergence
Crop yield and seedling emergence tests reflect soil and crop health under different tillage and crop rotation regimes. The growing seasons in 2016-2018 had highly variable precipitation (Fig. S4). Our results showed that corn yield was significantly affected by tillage, with ~18% higher yields in NT than in CT across 3 years (Table 3). Soybean and wheat yields were not significantly affected by tillage (Table 3). Crop rotation had significant effect on wheat yield, but not on corn and soybean yields. Wheat yields were ~78% higher under CSW than under wheat monoculture (Table 3).
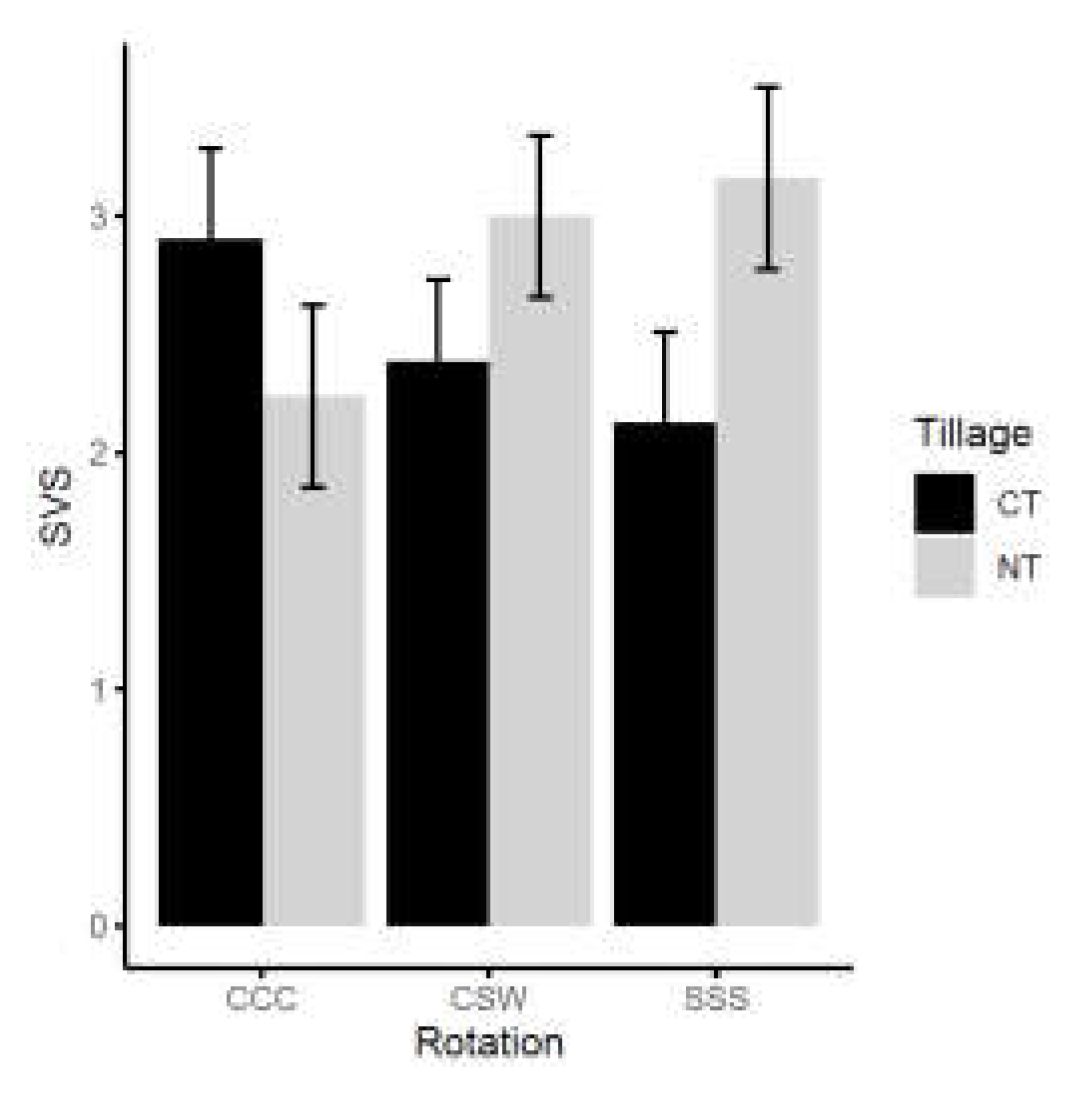
When collected field soils were planted with soybean in growth cabinet under controlled condition, seedling vitality (SVS; see section 2.6 for methodology) was not significantly affected by tillage or rotation. However, the interaction between tillage and rotation did significantly affect the SVS (Table 1, Fig. 5). We found that, compared with CT, NT decreased SVS by ~30% under corn monoculture, but increased SVS by 21% under soybean monoculture and by 33% under CSW. Under corn monoculture, SVS was highest under CT but lowest under NT.
Figure 5.
The soybean seedling vitality score (SVS) as affected by tillage and rotation (all pairwise comparisons P ≥ 0.05). CT, conventional tillage, NT, no-till; CCC, corn monoculture, SSS, soybean monoculture CSW, corn-soybean-wheat rotation.
Figure 5.
The soybean seedling vitality score (SVS) as affected by tillage and rotation (all pairwise comparisons P ≥ 0.05). CT, conventional tillage, NT, no-till; CCC, corn monoculture, SSS, soybean monoculture CSW, corn-soybean-wheat rotation.
3. Discussion
In this study, we focused on evaluating the effects of the combined long-term tillage and rotation regimes on the soil oomycete community diversity and compositional structure, but it is important to contextualize the environment in which this study took place. To this end, we assessed the crop yield response as an indicator of soil and crop health, which helped us better understand the long-term effects of these agricultural practices on soil and crop health. The long-term tillage and rotation had a significant effect on crop yield, which varied between crops. We found that NT had no significant effect on soybean and wheat yields in 2016 to 2018, which is in agreement with a previous yield study on this site from 2001-2015 by Morrison et al, who reported that yields did not differ between CT and NT for either soybean or wheat[29]. However, corn yield was higher under NT than under CT, contrary to prior results at this site [30]. The differences between these two studies suggests that the beneficial effects of NT on crop yield may not be evident in the short term and need to be studied on a long-term basis [31]. In comparison with monoculture systems, crop rotation significantly increased wheat yield, especially in 2016 and 2018 where the growing seasons were drier, but it did not affect corn and soybean yields. This is in agreement with 2001to 2015 data showing that wheat yielded 22% more when grown in rotation than in monoculture [30], largely attributed to wheat following the nitrogen-fixing soybean in the rotation. This long-term observation supports the value of crop rotation in increasing crop productivity.
Oomycetes are vastly understudied compared to bacteria and fungi despite their importance in crop production systems, where they are responsible for severe declines in crop yields. We investigated the impact of different tillage and crop rotations on the soil oomycete communities associated with corn, soybean, and wheat across three years. From the oomycete DNA we identified 34 species from 292 Amplicon Sequence Variants (ASVs). Globisporangium spp. and Pythium spp. were most abundant recovered from the study site and accounted for over 95% of the total oomycete abundance. This was in line with a previous study on the rhizosphere-associated oomycetes of oak [32], which reported that Globisporangium and Pythium were the most abundant genera and accounted for over 60% of the total community. It was also reported that Pythium (46%) was the most abundant and Globisporangium (6%) was the second abundant genera in the soils imported to Norway attached to roots of ornamental trees and shrubs [33]. Globisporangium and Pythium species that can cause damping-off and root rot are usually generalists that can infest a large variety of agricultural crops, as shown in Table 2 [34]. Globisporangium, which recently underwent taxonomic revision and has been reclassified from within Pythium to its own genus, has inherited some of the most problematic plant pathogens formerly belonging to Pythium [33], such as the former Py. sylvaticum, and Py. ultimum [35], are now known as G. sylvaticum and G. ultimum, which were highly abundant in the soils of the present study. Zitnick-Anderson and Nelson [26] found that the G. attrantheridium, G. heterothallicum, G. hypogynum, G. intermedium, and G. irregulare, caused pre-emergence damping-off on soybean with less than 50% seedling emergence compared with 100% seedling emergence in control. G. heterothallicum was one of the most abundant oomycete species in the present study, especially under corn and soybean monoculture. This species was reported as a pathogen on many crops, including corn [36] and soybean [37], and was a dominant species in North Dakota as reported by studies focusing on oomycete pathogenicity on soybean, representing 49% of the isolates [26]. However, Radmer et al. [25] reported that G. heterothallicum can be considered less aggressive on soybean or corn, so its abundance in the population could potentially mediate the pathogenicity of more detrimental species through competition. Several Pythiaceae spp., such as Py. arrhenomanes, Py. volutum, Py. oopapillum and Py. torulosum are reported to cause diseases on the seeds and seedlings of soybean [26,28,38], corn [39,40], or wheat [41,42,43,44].
Our findings showed that tillage practices, alone or in combination with rotation regimes, had a significant impact on the oomycete community's alpha- and beta-diversity. For example, we observed that NT decreased the alpha-diversity, represented by Simpson-TD and Shannon-TD. The Chao1 richness was also reduced in NT under CSW rotation, but not in the monoculture systems. The persistent decrease trend in Simpson-TD over the three years under NT but not under CT suggests a potential of NT in reducing the number of abundant oomycetes species (Fig. 4A). These observations are not in agreement with Srour et al. [15] who found that tillage had no significant effect on Shannon’s diversity index of the oomycete community at the soybean growth phase of a corn-soybean rotation regime. A meta-analysis showed that NT significantly increased bacterial community diversity but had little effect on fungal community diversity, which could be attributed to the increase of labile carbon and water holding capacities that are essential for microbial growth under NT or the moisture loss resulting in a more conducive environment for microbial growth promoted by plough tillage [16]. Moreover, our results showed an overall decrease in homogeneity of the soil oomycetes community under continuous CT, making CT a less favorable practice for managing oomycetes pathogens.
Considering that oomycetes zoospores require wet soil for mobility and invasion of the plants, greater but not excessive soil moisture content can promote oomycetes growth and infection ability. However, contradictory to previous studies which showed that reduced tillage and crop rotation may lead to greater soil moisture and soil organic matter contents by enhancing soil aggregation, promoting biological activities, and increasing water holding capacities [45,46], this study indicated that tillage and rotation regimes did not affect the soil moisture content (Supplementary Fig. S2). The soil moisture content also did not show significant associations with the community’s alpha- and beta-diversity, or the abundance of recovered oomycetes species. In summary, we did not observe a connection between tillage and rotation regimes and soil moisture content, likely due to the fall sampling time as the soil had compacted throughout the growing season and all plots are under the same yearly precipitation, temperature, and other climate conditions.
The suppression of Pythium damping-off can be enhanced by the addition of compost to improve overall soil microbial activity, implying that soil organic matter plays an important role in the soil suppressiveness against oomycetes [47]. Hoitink and Boehm indicated that the extent of soil-borne pathogen suppression was related to the quantity and quality of soil organic matter [48]. Bongiorno et al. found that reduced tillage can potentially increase soil suppressiveness through labile carbon and the positive effect of microbial biomass [9]. Therefore, the increase in soil natural suppressiveness under NT as a result of increased soil organic matter could be an important reason for the decrease in the alpha-diversity of the oomycete community observed in our present study.
We found that CT significantly reshaped the oomycete community structure and composition. This observation is in agreement with a previous study [15], which reported that tillage was a main factor driving the soil oomycete community heterogeneity. Among the 36 identified species, six were observed under CT but absent under NT, while another eight showed an opposite trend. The impact of tillage can vary between oomycete species, likely depending on their adaptation to changes in environmental conditions. Similar results were observed by Srour et al [15] who recovered G. attantheridium only from NT plots (Fig. 3B). Furthermore, G. sylvaticum and Py. arrhenomanes, which have been confirmed as pathogens for corn and soybean [37], were more abundant under NT than CT, which is consistent with previous research showing that Pythium spp. abundance increases under reduced tillage [49,50]. In contrast, G. apiculatum was in higher abundance under CT and rarely present under NT, although this species is not a confirmed pathogen [51]. G. ultimum is another species that was more abundant under CT than NT in our study, particularly under soybean monoculture (Fig. 3A), and has been reported as one of the most damaging pathogens of corn and soybean seeds and seedlings [37,38,52,53]. In addition, Py. volutum was highly abundant under CT but was present in low abundance under NT and was reported as being one of the most damaging pathogens in wheat [41]. Similarly, G. apiculatum, Py. volutum, and G. iwayamae were found in low abundance under NT but showed a consistent increase in abundances under CT over the three-year period, although such changes were not statistically significant (Fig. 4B-D). We hypothesize that that the soils under CT have a reduced capacity to suppress the oomycetes pathogens compared to NT. To investigate this further, our next step is to examine the bacterial and fungal communities present in the same soil samples.
Previous studies have shown correlations between the abundance of some Pythium species and soil chemical properties such as pH, calcium and magnesium content, cation exchange capacity, and clay content [54,55]. The shift in soil oomycete community composition induced by tillage practices in this study is likely associated with the changes in soil physical and chemical properties induced by such disruptive practices. The occurrence of some oomycete species in no-till systems may be a result of layered crop residue on the ground, which serves as an ideal habitat for primary inoculum buildup [56,57]. It is also likely that the timing of sampling influences the oomycete community composition. In our study we sampled in the fall, at a point where the majority of the plan tissue available to the soil-borne oomycete community was dead plant tissue, favoring saprophytic oomycetes such as Pythium and Globisporangium. Asad et al found that microbiome sampling early in the growing season was most closely tied to final seed quality [58]. It is likely that spring or summer sampling of oomycete communities may result in different trends.
Our results showed that the soil oomycete community structure differed significantly between monoculture systems. Sapkota and Nicolaisen (2018) found that crops grown prior to sampling, influenced the oomycete community composition in a field survey of arable soils [59]. In this study, G. apiculatum and G. ultimum were highly abundant in soybean soils. In contrast, Pythium sp. aff. monospermum and P. volutum were highly abundant in wheat soils. This confirms the crop effect on the oomycete community, and different oomycete species prefer certain crops as hosts. The crop effect on oomycete community structure could be related to the effect of root exudates in the rhizosphere, the accumulation of crop-specific root pathogens or parasites, and also the plant-derived crop residues left over after harvest [59].
Crop rotation has been demonstrated as a good practice for reducing in plant disease caused by soil-borne pathogens [20,60-62], potentially through altering soil physical changes plus the presence of layers of crop residue on the soil surface leading to changes in pathobiome community structure and functions. The present study further showed that rotation had no significant effect on the alpha-diversity but statistically affected the beta-diversity of the oomycete community under CT. Several species, including Py. sp. aff. Monospermum, G. iwayamae, and S. anisospore, were abundant under wheat monoculture but were in low abundance or absent in wheat soils under CSW, implying that rotation may reduce some oomycete species levels by breaking disease cycles. Such breaks in pathogen cycles were observed by Bargués-Ribera et al , where the instruction of non-host crops reduced the incidence of unspecified disease [63]. The USDA fungal database (https://nt.ars-grin.gov/fungaldatabases/) indicates that G. iwayamae is present in various hosts, including wheat [64]. These two species were reported to induce plant damping-off or rot diseases. Our study shows that crop rotation has the benefit of decreasing the abundance of oomycetes plant pathogens, possibly due to improved disease suppressive capacity of soil microbiomes in more diverse rotations [19] and increased soil N-levels from the inclusion of soybean as a preceding crop; both eventually increased wheat yield as we observed. No previous studies reported Pythium sp. aff. monospermum and S. anisospore in wheat soils. Pythium sp. aff. monospermum has been isolated from grapevine [65], and S. anisospore is generally reported as an aquatic pathogen [66], their pathogenicity are unknown. We did not observe a significant effect of rotation on the beta-diversity of the soil oomycete community under NT. One possible reason is that tillage and the disruption of soil structure is the major factor driving the soil oomycete community in our study. The top ten most abundant oomycete species under NT were not significantly affected by rotation. Only Py. volutum was significantly affected by rotation, which is not a highly abundant species under NT and may have a negligible contribution to shifting the soil oomycete community structure.
Many of the oomycetes identified are pathogens associated with soybean (Table 2) [25,27], and as such, we conducted a greenhouse experiment to determine the soybean seedling vitality in the studied soils collected in 2016 and 2017, aiming to evaluate the overall health of the soils and test if there is any association with identified oomycete species. Neither tillage nor rotation showed significant impact on the seedling vitality score (SVS) independently(Table 1), however, differing trends between the rotations within the two tillage treatments highlight the effect of the interaction between tillage and rotation (Fig. 5). We did not observe significant associations between any oomycete species and SVS. The poorer emergence in NT-CCC, CT-SSS, and CT-CSW treatments could be associated with the presence of higher abundances of G. heterothallicum and G. ultimum (Table 2). These two species have been shown to be aggressively pathogenic on soybean [25,27,67]. However, without isolation and/or molecular characterization [28,53], no direct conclusions about oomycete contribution to low SVS can be made.
4. Materials and Methods
4.1. Study site and experimental design
The field site sampled was a tillage and crop rotation trial established in 1990 at the Central Experimental Farm in Ottawa, ON, Canada (45°23’13.6”N; 75°43’15.6”W). The soil was a Matilda sandy loam (Melanic Brunisol, Canadian classification) with a pH (in CaCl2) of 6.8 [29]. A two-replicate, split-plot tillage–rotation experiment (Supplementary Fig. S1) was established on land that had previously grown alfalfa for 1 year and corn for several years prior to that. The main plots (89.1 m × 45.7 m each) were either managed by no-tillage (NT) or conventionally tillage (CT), within which the subplots for crop rotations were 9.1 m × 45.7 m. The plots under CT were subjected to moldboard ploughing (Overum DTL 5 Furrow plow, Västervik, Sweden) in early November and cultivated in the spring with a mulch-finisher [John Deere 2310 Mulch Finisher] followed by a combination harrow (Kongskilde 2600 Triple K, Albertslund, Denmark) with rotatory baskets. Subplots were allocated to three crops (corn, soybean, or wheat) grown in monoculture (CCC, SSS, CCC) or in two 3-year rotations (corn–soybean–wheat [CSW] or corn–wheat–soybean [CWS]). The two rotations were initiated with plots grown each of the three crops, i.e., each crop in a rotation regime was grown every year, leading to a total of nine subplots (n = 3 for CSW, n = 3 for CWS, and n = 3 for monoculture) within each main plot. Within each main plot (tillage effect), the subplots (crop × rotation) were duplicated in complete blocks. Therefore, a total of 72 subplots were established, including randomized and duplicated subplots representing nine rotation sequences (n = 2 x 9) within each of the two duplicated main plots (n = 2 x 2) (Supplementary Figure S1).
Crop planting and management information were described by Morrison et al. [68]. Wheat was planted in the first two weeks of May (450 seeds m2) using a Sunflower 9312 Multifunction Drill (Beloit, KS) equipped with disc openers with a row spacing of 19 cm. Corn was seeded (7 seeds m-2) in the first two weeks of May with a John Deere 6-row corn planter set to the NT option; rows were 76 cm wide. Soybean was planted (55 seeds m-2) in the last two weeks of May with the Sunflower drill with 19 cm wide rows. Corn was fertilized with 224 kg N (as urea) ha−1 pre-plant broadcast and 40 kg ha−1 N–P2O5–K2O (18-18-18) at seeding. Wheat plots were fertilized with 100 kg N (as urea) ha−1 pre-planting. Soybean received no fertilizer. Pre-plant fertilizer was applied prior to spring tillage, therefore, it was integrated into the soil layer in CT plots but remained on the surface in the NT plots. Glyphosate was used to control weeds in commercial herbicide-resistant varieties of corn and soybean. Weeds were controlled in wheat with Buctril-M at 0.2 L ha−1 at the seedling stage.
Crop yields for soybean and wheat were measured by harvesting a central strip of each plot (6 rows) using a plot combine (NurseryMaster, Wintersteiger, Germany). Corn yields were harvested using a John Deere combine (X9, John Deere, USA). Grain yields were adjusted to a moisture content of 13%. Corn crop residues were chopped down with a Loftness 180 flail-style chopper (Loftness, Hector MN), while soybean and wheat stubble were left in place. All crop residue was fall incorporated in the CT plots but remained on the soil surface in the NT plots. A weather station 700 meters from the study site [45o22’57.34” N, 75o42’50.96” W] was used to collect precipitation and minimum and maximum daily temperature data for 2016, 2017, and 2018 growing seasons.
4.2. Soil sampling
In this study, only the corn-soybean-wheat rotation and monoculture plots were selected to investigate the rotation effect. Soil sampling was conducted at the end of the 2016 to 2018 growing seasons after the crop was harvested. Sampling in 2016 was considered as a proof of concept, for which only soils from the soybean and corn monoculture plots and CSW plots in soybean that year were sampled (24 plots). For the subsequent sampling years, additional rotation regimes were included: in 2017 the sampling was expanded to include the CSW rotation plots in soybean and wheat as well as the corn (CCC) and soybean (SSS) monoculture treatments (32 plots); in 2018 the wheat monoculture (WWW) was added to the sampled plots (46 plots). A total of 102 samples were collected over the three years. Due to the unbalanced sampling strategy across the years, we considered the treatments composed of two tillage treatments (CT and NT) and four rotations (CCC, SSS, WWW, and CSW).
Soil sampling followed a random staggered strategy, with 0-30 cm depth soil cores (diameter = 2 cm) being taken from each plot using a soil core sampler (Lamotte, Chesterton, MD, USA). In 2016 and 2017, 50 cores were collected per plot. In 2018, 15 cores were collected per plot. All equipment was rinsed with distilled water, then sterilized with 90% ethanol, and dried between sampling different plots. The soil cores were pooled by plot and were transferred to a cold room (4 °C), until sampling was completed. Prior to sub-sampling the soil samples were thoroughly mixed and sieved through a 5 mm mesh to remove rocks, plant material, and insects. A 15 ml sub-sample of the homogenized soil per plot was stored in a falcon tube at -80 °C.
Gravometric soil moisture was measured by taking a 30 mL sample of soil at sampling for each plot, weighing it wet, and drying it at 60o C until the weight was stable, approximately 7 days in this case. Dried soils were weighed, and soil moisture was calculated as shown in equation 1.
4.3. DNA extraction
The DNAs were extracted using the FastDNATM Spin Kit for Soil (MP Biomedicals, Santa Ana, CA, USA) following the manufacturer’s instructions with the following modifications: a 2 mL microcentrifuge tube was used during the binding step rather than a 15 mL falcon tube, and a Percellys® Evolution homogenizer (Bertin Instruments, Montigny-le-Bretonneux, France) was used instead of the MP Fast-Prep homogenizer. The soil was homogenized with silica and glass beads, then a column binding step was used to remove debris where the DNA was bound to beads and column elution was used to remove the protein and RNA. The purified DNA was then eluted into DNAse-free water. All samples were extracted in triplicate. The extracted DNA was stored at -25 °C.
The concentration of the DNA extract was measured using the Qubit dsDNA HS (High Sensitivity) Assay Kit on a Qubit Fluorometer (Thermo Fisher Scientific, MA, USA). Three DNA replicates were pooled to 40 µL at a concentration of 10 ng µL-1 in 96-well PCR plates, which were stored at -25 °C. The plates containing the genomic DNAs were shipped overnight on dry ice to the Génome Québec Innovation Centre (Montreal, QC, Canada) for the preparation of sequencing libraries and amplicon-based metagenomics (or metabarcoding) sequencing using the Illumina MiSeq platform target fragment length was 100-300 bp with a target output of 15 Gb.
4.4. Sequencing library preparation and Illumina MiSeq sequencing
At the Génome Québec Innovation Centre, the DNA libraries of the ITS1 region were prepared using SSU_ITS (5’-ACA CTG ACG ACA TGG TTC TAC ACG GAA GGA TCA TTA CCA CAC-3’) forward primer and the OOM_LO5.8S47c (5’-TAC GGT AGC AGA GAC TTG GTC TAT TAC GTA TCG CAG TTC GCA-3’) reverse primer (A. Levesque, personal communication). The initial PCR amplification was carried out in 8 μL reaction volume comprised of 7 μL of the master mix (Supplementary Table S1) and 1 μL of sample DNA diluted to 1/50. The following thermocycling parameters were 15 minutes at 96 °C for initial melting then 33 cycles through of 30 seconds at 96 °C, 30 seconds at 52 °C and 60 seconds at 72 °C, followed by a 10-minute cool down at 72 °C. The amplicons were verified on a 2% agarose gel quantified and were purified using the sparQ PurMag Beads (Quantabio, Beverly, MA, USA). A secondary PCR was then done to add the dual indexed barcoding adapters. The PCR was carried out in 7 μL reaction volume containing a 1/50 dilution of DNA to master mix. The PCR cycling parameters were: 15 minutes at 96 °C, 30 seconds at 96 °C, 30 seconds at 52 °C, 60 seconds at 72 °C and 10 minutes at 72 °C. The amplification was verified, and amplicons were purified as above. Indexation was done with 1 μL of undiluted amplicon product secondary PCR. Indexed samples were verified on a 2% agarose gel and quantified using Quant-iT™ PicoGreen® dsDNA Assay Kit (Life Technologies, CA, USA). The sequencing library was made with an equal quantity in ng of DNA for each sample. Library DNA was cleaned with sparQ PureMag Beads. The library was quantified using Kapa Illumina GA with Revised Primers-SYBR Fast Universal Kit (Kapa Biosystems, MA, USA), and average fragment size was determined using the LabChip GX (PerkinElmer, MA, USA). Before sequencing 10% of Phix control library was added to the amplicon pool for a final concentration of 8 pM. Sequencing was performed on the Illumina MiSeq platform with the MiSeq Reagent Kit v3 with 600 cycles. Sequencing was done with LNA™ modified custom primers.
4.5. Metabarcoding data processing and analysis
The Illumina MiSeq sequencing adapters were removed from the fastq files using Cutadapt ver.4.1 [69]. The paired-end raw reads were processed using DADA2 ver.1.14 [70] implemented in QIIME2 for denoising, chimera detection, and the amplicon sequence variants (ASVs) inference using default parameters. The raw forward and reverse reads were truncated at 200 nt.
The taxonomic assignment was performed with an In-house complied reference database of oomycetes (denoted as oomycetes-ITS1-refDB) from GenBank. The oomycetes-ITS1-refDB were first downloaded in TinySeq XML format using query “Oomycetes[Organism]” AND “150:2500[slen]” AND (“internal transcribed spacer 1” OR “ITS1”) NOT “sp.” NOT “uncultured” NOT “clone” NOT “whole genome” NOT” metagenome” (retrieved on March 2, 2022). In-house Perl and Bash scripts were developed to parse the XML file and to retrieve National Center for Biotechnology Information taxonomy for each sequence. The locations of rRNA gene regions (ITS1, 5.8S, ITS2, 28S) within each sequence were annotated by ITSx [71]. Only sequences including ITS1 region with sequence length between 150–500 bp were retained. The final oomycetes-ITS1-refDB database contained 26,220 oomycete ITS1 reference sequences. This database can be downloaded from the bitbucket repository: https://bitbucket.org/wenchen_aafc/metabarcoding_oomycetes/downloads/.
The taxonomy assignments were initially classified via the q2-feature-classifer [72] implemented in QIIME2 against the oomycetes-ITS1-refDB database. Species-level identification of the oomycetes was improved by the Automated Oligonucleotide Design Pipeline (AODP), which identified all mutations distinguishing highly conserved DNA markers between close relatives [73,74]. The final taxonomy was improved by comparing and validating the results of three classifiers: AODP, q2-feature-classifier, and BLASTn at each taxonomic rank, in particular, at species level. To assess the accuracy of the species-level assignments, we extracted the representative sequences of each ASV assigned to a specific species, which were combined with corresponding reference sequences of the species and its close relatives in oomycetes-ITS1-refDB. The combined sequence dataset was then aligned using MAFFT vers.7.407 [75], followed by the reconstruction of an approximate maximum likelihood (ML)_tree using FastTree (ver.2.1.0) with the –nt and –gtr options [76]. The ML tree was visualized in FigTree (ver.1.4.4, https://github.com/rambaut/figtree/releases).
4.6. Soybean seedling vitality experiment
To evaluate the impact of tillage and rotation regimes on the potential pathogenicity of soil-borne microorganisms, including oomycetes, on soybean seedling emergence, we carried out seedling emergence tests under controlled growth cabinet conditions in 2016 and 2017. In this trial, 5 kg of soil were collected from selected subplots under different tillage × rotation treatments following the same procedure as the soil samples that were submitted for DNA extraction and sequencing. We collected a total of 24 soil samples in 2016 and 32 in 2017. The soils from each subplot were placed in a plastic tray (9×12×35 cm). Thirty-two soybean seeds (variety: AC Mandor) were planted in a four column 8 row grid, 2 cm deep. The growth cabinet was set at 15 ℃ with a 10-hour photoperiod. Field capacity was calculated by filling a 2-inch plastic pot with field soil, saturating the soil, and weighing after 16 hours of draining. Soil moisture was maintained by weighing the trays and adjusting the water content to 80% of field capacity twice a day with distilled water. A final seedling vitality score was used to asses plant health at harvest: a score of “1” was given to seeds that did not germinate; “2“ to seeds that germinated but had broken off cotyledons and signs of rot on the stem and roots; “3“ to seedlings that had emerged but had signs of necrosis on the cotyledons and delayed unifoliate emergence; “4“ to seedlings with signs of necrosis on the stems and cotyledons as well as stunted and damaged unifoliates; “5“ to seedlings with healthy unifoliates but signs of necrosis on the stems and cotyledons; and “6“ to healthy seedlings (Supplementary Fig. S5). To minimize edge effects, the seedlings of the end rows proximity to the edge of the plug tray were not evaluated. The overall seedling vitality score (SVS) for each subplot was calculated using equation 2, where i is the vitality score ranging from 1 to 6, xi is the number of seedlings with a given vitality score, and n is the total number of seedlings.
4.7. Statistical analysis
All statistical analyses were performed in R (ver. 4.2.0) [77]. To avoid the risk of losing rare taxa (ASVs with low sequence counts), the ASV table was not rarefied but was Hellinger-transformed for multivariate analysis. The alpha-diversity indices are quantitative measures representing the diversity of ASVs in a sample. The Shannon Index (H), Simpson Index (D), and Chao1 index were calculated using vegan [78] and biodiversityR [79] packages. The Shannon-based True Diversity (Shannon-TD = exp(H)) and the Simpson-based True Diversity (Simpson-TD = 1/D) were calculated as suggested by Jost [80].
The alpha-diversity indices, crop yield, and SVS were checked for normality using the shapiro.test function and were transformed as needed. Linear mixed models were used to assess the main effects of tillage and rotation, and their interaction on alpha-diversity indices, the relative abundances of species, and the SVS using the lme function in the nlme package [81] at a significance level of P ≤ 0.05. Tillage and rotation were treated as fixed effects, and year as random effect. Linear mixed models were also used to assess the effect of rotation under each tillage treatment on the relative abundances of taxa at species level, with rotation as a fixed effect, and blocks and years as random effects. Linear mixed models were used to evaluate the tillage, rotation, and their interaction on corn, soybean and wheat crop yields separately, with tillage and rotation as fixed effects, and blocks as random effect, and year as repeated measurements.
The non-metric multidimensional scaling (NMDS) was done to evaluate the overall impact of tillage and crop rotation on oomycete community heterogeneity using the metaMDS function in vegan [78]. The adonis function from the vegan package was used to perform permutational multivariate analysis of variance (PERMANOVA) [82] for determining the main effects of tillage, rotation, and the interaction effects, and also the effect of current crop under CSW treatment on the community heterogeneity of oomycete community based on the Bray-Curtis dissimilarity. Pairwise comparisons between treatments (tillage, rotation, or their combination) were conducted by using the pairwise.perm.manova function from the RVAideMemoire package [83] when one factor or interaction effect is significant. The community dissimilarity over the study years under CT and NT was evaluated by using the beta.sample function in the betapart package [84].
5. Conclusions
Our study shows that combined tillage and rotation regimes have pronounced impacts on soil and crop health and the oomycete communities. We found that no-tillage (NT) is an effective way to suppress the overall soil oomycete community, while the selection of crop rotations is crucial under conventional tillage (CT), where the oomycete community is more responsive to changes in management practices. Our findings suggest that continuous CT practices may lead to a decreased homogeneity of the soil oomycetes community, while NT may be a more sustainable approach to farming that helps maintain soil health and biodiversity. It's important to note that different oomycete species respond differently to tillage and rotation practices, which could be due to host availability and their unique adaptations to specific soil and environmental conditions. However, to gain a comprehensive understanding of the potential of tillage and rotation in mediating the pathogenicity of soil-borne oomycetes, future studies should consider investigating the effects of soil physicochemical properties on various microbial communities. Additionally, constructing cross-kingdom co-occurrence networks and conducting culture-dependent and molecular diagnostic assays to confirm causal agents would be beneficial.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Fig. S1: Field maps of the split-plot experimental design in each field sampling year. Fig. S2: A) The impact of tillage and rotation regimes on the soil moisture content, as measured by weighing soils at sampling, drying down the soils at 60oC and calculating gravometric soil moisture content. The correlation relationships between the soil moisture content and the B) Shannon-based true diversity index, C) Simpson-based true diversity index, and D) the chao1 index of the soil-borne oomycetes community. Fig. S3: Validation of the classification accuracy of oomycetes ASVs at the species level using phylogenetic tree reconstruction. The 236 oomycetes ASVs (highlighted in red) was combined with reference sequences downloaded from the Genbank for multiple sequence alignment and Approximate Maximum Likelihood tree reconstruction. Fig. S4: Cumulative precipitation at the experimental site over the study years (2016-2018). Fig. S5: Examples of soybean plants from each vitality score (1-6) rating where: 1, no germination; 2, broken-off cotyledons and signs of necrosis on roots; 3, seedlings with signs of necrosis on the cotyledons and stunted unifoliate emergence; 4, seedlings with signs of necrosis on the cotyledons and minimal stunting of the unifoliates; 5 seedlings with minimal cotyledon necrosis and healthy unifoliate development; 6, healthy seedlings with no signs of disease. Soybean plants were harvested after 21 days of growth in field soil at 15 oC constant temperature at 80% of saturated pot capacity. Scale bar = 1 cm.
Author Contributions
Conceptualization, C.G., M.M, W.C.; methodology, C.G. M.M., E.G.; Software, W.C., D.R; formal analysis, C.G. and Y.S; data curation, W.C, D.R; writing—original draft preparation, C.G, Y.S.; writing—review and editing, W.C., S.A.B, M.M., E.G.; visualization, C.G., Y.S.; supervision, W.C., A.S., M.M.; project administration, W.C.; funding acquisition, W.C.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by Agriculture and Agri-Food Canada (grants J-002272, J-002305, and J-002366).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw paired-end metabarcoding sequencing data have been deposited in the Sequence Read Archive (SRA) under the BioProject accession PRJNA932837, Biosample accession SAMN33294720 to SAMN33294793.
Acknowledgments
We thank A. Levesque for providing the new primer for oomycetes ITS1; H. Nguyen for advising on oomycetes taxonomy. Ryan Tobalt and Thomas Hotte for their assistance in sample collection, Hai Nguyen and Ekaterina Ponomareva for providing lab space and sample storage throughout the project.
Conflicts of Interest
The authors declare no conflict of interest.
Crown Copyright statement
© His Majesty the King in Right of Canada, as represented by the Minister of Agriculture and Agri-Food Canada 2023.
References
- Thines, M.; Voglmayr, H.; Göker, M. Taxonomy and Phylogeny of the Downy Mildews (Peronosporaceae). In Oomycete Genetics and Genomics; 2009; pp. 47–75. [Google Scholar]
- Fry, W. Phytophthora infestans: the plant (and R gene) destroyer. Molecular Plant Pathology 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Anderson, V.L.; Robertson, E.J.; Secombes, C.J.; van West, P. New insights into animal pathogenic oomycetes. Trends in Microbiology 2008, 16, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Cahill, D.M.; Rookes, J.E.; Wilson, B.A.; Gibson, L.; McDougall, K.L. Phytophthora cinnamomi and Australia’s biodiversity: impacts, predictions and progress towards control. Australian Journal of Botany 2008, 56. [Google Scholar] [CrossRef]
- Cohen, Y.; Coffey, M.D. SYSTEMIC FUNGICIDES AND THE CONTROL OF OOMYCETES. Annual Reviews in Phytopathology 1986, 24, 311–338. [Google Scholar] [CrossRef]
- Dorrance, A.; Grünwald, N.J. Phytophthora sojae: Diversity among and within Populations. In Oomycete Genetics and Genomics; 2009; pp. 197–212. [Google Scholar]
- Cook, R.; Sitton, J.; Haglund, W. Influence of soil treatments on growth and yield of wheat and implications for control of Pythium root rot. Phytopathology 1987, 77, 1192–1198. [Google Scholar] [CrossRef]
- Cook, R.J.; Haglund, W.A. Wheat yield depression associated with conservation tillage caused by root pathogens in the soil not phytotoxins from the straw. Soil Biology and Biochemistry 1991, 23, 1125–1132. [Google Scholar] [CrossRef]
- Bongiorno, G.; Postma, J.; Bünemann, E.K.; Brussaard, L.; de Goede, R.G.M.; Mäder, P.; Tamm, L.; Thuerig, B. Soil suppressiveness to Pythium ultimum in ten European long-term field experiments and its relation with soil parameters. Soil Biology and Biochemistry 2019, 133, 174–187. [Google Scholar] [CrossRef]
- Peters, R.D.; Sturz, A.V.; Carter, M.R.; Sanderson, J.B. Developing disease-suppressive soils through crop rotation and tillage management practices. Soil and Tillage Research 2003, 72, 181–192. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, B.; Hong, S.; Xiong, W.; Shen, Z.; Ruan, Y.; Li, R.; Shen, Q.; Dini-Andreote, F. Promoting soil microbial-mediated suppressiveness against Fusarium wilt disease by the enrichment of specific fungal taxa via crop rotation. Biology and Fertility of Soils 2021, 57, 1137–1153. [Google Scholar] [CrossRef]
- Santín-Montanyá, M. I.; Sombrero Sacristán, A. The effects of soil tillage techniques on weed flora in high input barley systems in northern Spain. Canadian Journal of Plant Science 2020, 100, 245–252. [Google Scholar] [CrossRef]
- Bogunovic, I.; Pereira, P.; Kisic, I.; Sajko, K.; Sraka, M. Tillage management impacts on soil compaction, erosion and crop yield in Stagnosols (Croatia). CATENA 2018, 160, 376–384. [Google Scholar] [CrossRef]
- Abdollahi, L.; Schjønning, P.; Elmholt, S.; Munkholm, L.J. The effects of organic matter application and intensive tillage and traffic on soil structure formation and stability. Soil and Tillage Research 2014, 136, 28–37. [Google Scholar] [CrossRef]
- Srour, A.Y.; Ammar, H.A.; Subedi, A.; Pimentel, M.; Cook, R.L.; Bond, J.; Fakhoury, A.M. Microbial Communities Associated With Long-Term Tillage and Fertility Treatments in a Corn-Soybean Cropping System. Front Microbiol 2020, 11, 1363. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, D.; Liang, S.; Dang, P.; Qin, X.; Liao, Y.; Siddique, K.H.M. Effect of no-tillage on soil bacterial and fungal community diversity: A meta-analysis. Soil Tillage Res. 2020, 204, 104721. [Google Scholar] [CrossRef]
- Sipilä Timo, P.; Yrjälä, K.; Alakukku, L.; Palojärvi, A. Cross-Site Soil Microbial Communities under Tillage Regimes: Fungistasis and Microbial Biomarkers. Applied and Environmental Microbiology 2012, 78, 8191–8201. [Google Scholar] [CrossRef]
- Liu, B.; Gumpertz, M.L.; Hu, S.; Ristaino, J.B. Effect of prior tillage and soil fertility amendments on dispersal of Phytophthora capsici and infection of pepper. European Journal of Plant Pathology 2008, 120, 273–287. [Google Scholar] [CrossRef]
- Peralta, A.L.; Sun, Y.; McDaniel, M.D.; Lennon, J.T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. 2018, 9, e02235. [Google Scholar] [CrossRef]
- Pedersen, E.A. The Effect of Crop Rotation on Development of the Septoria Disease Complex on Spring Wheat in Saskatchewan. Canadian Journal of Plant Pathology 1992, 14, 152–158. [Google Scholar] [CrossRef]
- Larkin, R.P.; Halloran, J.M. Management Effects of Disease-Suppressive Rotation Crops on Potato Yield and Soilborne Disease and Their Economic Implications in Potato Production. American Journal of Potato Research 2014, 91, 429–439. [Google Scholar] [CrossRef]
- Lyu, J.; Jin, L.; Jin, N.; Xie, J.; Xiao, X.; Hu, L.; Tang, Z.; Wu, Y.; Niu, L.; Yu, J. Effects of Different Vegetable Rotations on Fungal Community Structure in Continuous Tomato Cropping Matrix in Greenhouse. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Sui, P.; Liu, X.; Zhang, T.; Li, X. Crop rotation to diversify the soil microbiome in the semi-arid area of Inner Mongolia, China. Archives of Agronomy and Soil Science 2022, 1–16. [Google Scholar] [CrossRef]
- Hwang, S.; Ahmed, H.; Gossen, B.; Kutcher, H.; Brandt, S.; Strelkov, S.; Chang, K.; Turnbull, G. Effect of crop rotation on the soil pathogen population dynamics and canola seedling establishment. Plant Pathology Journal 2009, 8, 106–112. [Google Scholar] [CrossRef]
- Radmer, L.; Anderson, G.; Malvick, D.M.; Kurle, J.E.; Rendahl, A.; Mallik, A. Pythium, Phytophthora, and Phytopythium spp. Associated with Soybean in Minnesota, Their Relative Aggressiveness on Soybean and Corn, and Their Sensitivity to Seed Treatment Fungicides. 2017, 101, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Zitnick-Anderson, K.K.; Nelson, B.D. Identification and Pathogenicity of Pythium on Soybean in North Dakota. 2015, 99, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.A.; Jacobs, J.L.; Napieralski, S.; Karaj, B.; Bradley, C.A.; Chase, T.; Esker, P.D.; Giesler, L.J.; Jardine, D.J.; Malvick, D.K.; et al. Oomycete Species Associated with Soybean Seedlings in North America—Part I: Identification and Pathogenicity Characterization. 2017, 107, 280–292. [Google Scholar] [CrossRef]
- Rojas, J.A.; Jacobs, J.L.; Napieralski, S.; Karaj, B.; Bradley, C.A.; Chase, T.; Esker, P.D.; Giesler, L.J.; Jardine, D.J.; Malvick, D.K.; et al. Oomycete Species Associated with Soybean Seedlings in North America—Part II: Diversity and Ecology in Relation to Environmental and Edaphic Factors. 2017, 107, 293–304. [Google Scholar] [CrossRef]
- Morrison, M.J.; Cober, E.R.; Gregorich, E.; Voldeng, H.D.; Ma, B.; Topp, G.C. Tillage and crop rotation effects on the yield of corn, soybean and wheat in eastern Canada. Canadian Journal of Plant Science 2017, 98, 183–191. [Google Scholar] [CrossRef]
- Morrison, M.J.; Cober, E.R.; Gregorich, E.G.; Voldeng, H.D.; Ma, B.; Topp, G.C. Tillage and crop rotation effects on the yield of corn, soybean, and wheat in eastern Canada. 2018, 98, 183–191. [Google Scholar] [CrossRef]
- Büchi, L.; Wendling, M.; Amossé, C.; Jeangros, B.; Sinaj, S.; Charles, R. Long and short term changes in crop yield and soil properties induced by the reduction of soil tillage in a long term experiment in Switzerland. Soil and Tillage Research 2017, 174, 201–129. [Google Scholar] [CrossRef]
- Sapp, M.; Tyborski, N.; Linstädter, A.; López Sánchez, A.; Mansfeldt, T.; Waldhoff, G.; Bareth, G.; Bonkowski, M.; Rose, L.E. Site-specific distribution of oak rhizosphere-associated oomycetes revealed by cytochrome c oxidase subunit II metabarcoding. 2019, 9, 10567–10581. [Google Scholar] [CrossRef]
- Rossmann, S.; Lysøe, E.; Skogen, M.; Talgø, V.; Brurberg, M.B. DNA Metabarcoding Reveals Broad Presence of Plant Pathogenic Oomycetes in Soil From Internationally Traded Plants. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.N.; Loper, J.E. Soilborne Plant Diseases Caused by Pythium spp.: Ecology, Epidemiology, and Prospects for Biological Control. Critical Reviews in Plant Sciences 1999, 18, 111–181. [Google Scholar] [CrossRef]
- Uzuhashi, S.; Kakishima, M.; Tojo, M. Phylogeny of the genus Pythium and description of new genera. Mycoscience 2010, 51, 337–365. [Google Scholar] [CrossRef]
- Gan, H.; Chai, Z.; Lou, B.; Li, J. Pythium heterothallicum new to China and its pathogenicity. Mycosystem 2010, 29, 494–501. [Google Scholar]
- Coffua, L.S.; Veterano, S.T.; Clipman, S.J.; Mena-Ali, J.I.; Blair, J.E. Characterization of Pythium spp. Associated with Asymptomatic Soybean in Southeastern Pennsylvania. 2016, 100, 1870–1879. [Google Scholar] [CrossRef]
- Broders, K.D.; Lipps, P.E.; Paul, P.A.; Dorrance, A.E. Characterization of Pythium spp. Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Disease 2007, 91, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.G.; Moorman, T.B.; Kaspar, T.C.; Manter, D.K. Isolation of Cultivation-Resistant Oomycetes, First Detected as Amplicon Sequences, from Roots of Herbicide-Terminated Winter Rye. 2017, 1, 24–35. [Google Scholar] [CrossRef]
- Reyes-Tena, A.; Vallejo-González, R.; Santillán-Mendoza, R.; Rodríguez-Alvarado, G.; Larsen, J.; Fernández-Pavía, S.P. Pythium arrhenomanes causal agent of root rot on yellow maize in Mexico. Australasian Plant Disease Notes 2018, 13, 6. [Google Scholar] [CrossRef]
- Chamswarng, C.; Cook, R. Identification and comparative pathogenicity of pythium species from wheat roots and wheat-field soils in the Pacific Northwest. Phytopathology 1985, 75, 821–827. [Google Scholar] [CrossRef]
- Paulitz, T.C. Low Input No-till Cereal Production in the Pacific Northwest of the U.S.: The Challenges of Root Diseases. European Journal of Plant Pathology 2006, 115, 271–281. [Google Scholar] [CrossRef]
- Waller, J.M. Observations on Pythium root rot of wheat and barley. 1979, 28, 17–24. [Google Scholar] [CrossRef]
- Yin, C.; McLaughlin, K.; Paulitz, T.C.; Kroese, D.R.; Hagerty, C.H. Population Dynamics of Wheat Root Pathogens Under Different Tillage Systems in Northeast Oregon. 2020, 104, 2649–2657. [Google Scholar] [CrossRef] [PubMed]
- Shittu, K.A.; Oyedele, D.J.; Babatunde, K.M. The effects of moisture content at tillage on soil strength in maize production. Egyptian Journal of Basic and Applied Sciences 2017, 4, 139–142. [Google Scholar] [CrossRef]
- Yang, T.; Siddique, K.H.M.; Liu, K. Cropping systems in agriculture and their impact on soil health-A review. Global Ecology and Conservation 2020, 23. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends in Plant Science 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Hoitink, H.; Boehm, M. Biocontrol within the context of soil microbial communities: a substrate-dependent phenomenon. Annual review of phytopathology 1999, 37, 427. [Google Scholar] [CrossRef]
- Cook, R.; Sitton, J.; Waldher, J. Evidence for Pythium as a pathogen of direct-drilled wheat in the Pacific Northwest. Plant Disease 1980, 64, 61–61. [Google Scholar] [CrossRef]
- Pankhurst, C.E.; McDonald, H.J.; Hawke, B.G. Influence of tillage and crop rotation on the epidemiology of Pythium infections of wheat in a red-brown earth of South Australia. Soil Biology and Biochemistry 1995, 27, 1065–1073. [Google Scholar] [CrossRef]
- Paul, B. A new species of Pythium isolated from a vineyard in France. FEMS Microbiology Letters 2006, 263, 194–199. [Google Scholar] [CrossRef]
- Jiang, Y.N.; Haudenshield, J.S.; Hartman, G.L. Characterization of Pythium spp. from soil samples in Illinois. Canadian Journal of Plant Pathology 2012, 34, 448–454. [Google Scholar] [CrossRef]
- Wei, L.; Xue, A.; Cober, E.; Babcock, C.; Zhang, J.; Zhang, S.; Li, W.; Wu, J.; Liu, L.J.P. Pathogenicity of Pythium species causing seed rot and damping-off in soybean under controlled conditions. 2010, 91, 3–10. [Google Scholar] [CrossRef]
- Broders, K.D.; Wallhead, M.W.; Austin, G.D.; Lipps, P.E.; Paul, P.A.; Mullen, R.W.; Dorrance, A.E. Association of Soil Chemical and Physical Properties with Pythium Species Diversity, Community Composition, and Disease Incidence. 2009, 99, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Hancock, J. Association of chemical and biological factors in soils suppressive to Pythium ultimum. Phytopathology 1986, 76, 1221–1231. [Google Scholar] [CrossRef]
- Rojas, J.A.; Witte, A.; Noel, Z.A.; Jacobs, J.L.; Chilvers, M.I. Diversity and Characterization of Oomycetes Associated with Corn Seedlings in Michigan. 2019, 3, 224–234. [Google Scholar] [CrossRef]
- Bockus, W.; Shroyer, J. The impact of reduced tillage on soilborne plant pathogens. Annual review of phytopathology 1998, 36, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Asad, N.I.; Wang, X.-B.; Dozois, J.; Azarbad, H.; Constant, P.; Yergeau, E. Early season soil microbiome best predicts wheat grain quality. FEMS Microbiology Ecology 2023, 99, fiac144. [Google Scholar] [CrossRef]
- Sapkota, R.; Nicolaisen, M. Cropping history shapes fungal, oomycete and nematode communities in arable soils and affects cavity spot in carrot. Agric. Ecosyst. Environ. 2018, 257, 120–131. [Google Scholar] [CrossRef]
- EMMOND, G.S.; LEDINGHAM, R.J. EFFECTS OF CROP ROTATION ON SOME SOIL-BORNE PATHOGENS OF POTATO. 1972, 52, 605–611. [Google Scholar] [CrossRef]
- Frank, J.A.; Murphy, H. The effect of crop rotations on Rhizoctonia disease of potatoes. American Potato Journal 1977, 54, 315–322. [Google Scholar] [CrossRef]
- Scholte, K.H. The effect of crop rotation and granular nematicides on the incidence ofRhizoctonia solani in potato. J Potato Research 1987, 30, 187–199. [Google Scholar] [CrossRef]
- Bargués-Ribera, M.; Gokhale, C.S. Eco-evolutionary agriculture: Host-pathogen dynamics in crop rotations. PLoS computational biology 2020, 16, e1007546. [Google Scholar] [CrossRef]
- Lipps, P.; Bruehl, G. Snow rot of winter wheat in Washington. Phytopathology 1978, 68, 120–121. [Google Scholar] [CrossRef]
- Spies, C.F.J.; Mazzola, M.; McLeod, A. Characterisation and detection of Pythium and Phytophthora species associated with grapevines in South Africa. European journal of plant pathology 2011, 131, 103–119. [Google Scholar] [CrossRef]
- Masigol, H.; Khodaparast, S.A.; Mostowfizadeh-Ghalamfarsa, R.; Rojas-Jimenez, K.; Woodhouse, J.N.; Neubauer, D.; Grossart, H.-P. Taxonomical and functional diversity of Saprolegniales in Anzali lagoon, Iran. Aquatic Ecology 2020, 54, 323–336. [Google Scholar] [CrossRef]
- Rizvi, S.; Yang, X. Fungi associated with soybean seedling disease in Iowa. Plant disease 1996, 80, 57–60. [Google Scholar] [CrossRef]
- Morrison, M.J.; Cober, E.R.; Gregorich, E.G.; Voldeng, H.D.; Ma, B.; Topp, G.C. Tillage and crop rotation effects on the yield of corn, soybean, and wheat in eastern Canada. Canadian Journal of Plant Science 2017, 98, 183–191. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EBMnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler2, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6. [Google Scholar] [CrossRef]
- Zahariev, M.A.-O.; Chen, W. Cluster oligonucleotide signatures for rapid identification by sequencing. BMC Bioinformatics 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Radford, D.; Hambleton, S. Towards improved detection and identification of rust fungal pathogens in environmental samples using a metabarcoding approach. Phytopathology 2021, 112, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS One 2010, 5, e9490. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. 2021.
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O'Hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. vegan: Community Ecology Package. R package version 2.6-2. https://CRAN.R-project.org/package=vegan 2022, 2. [Google Scholar]
- Kindt, R. BiodiversityR: Package for community ecology suitability analysis. 2020. Available online: https://github.com/cran/BiodiversityR.
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. nlme: Linear and Nonlinear Mixed Effects Models. 2023. Available online: https://CRAN.R-project.org/package=nlme.
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; 2014; pp. 1–15. [Google Scholar]
- Hervé, M. RVAideMemoire: Testing and plotting procedures for biostatistics. R package version 0.9-81-2 2022, https://CRAN.R-project.org/package=RVAideMemoire, doi:https://CRAN.Rproject. org/package=RVAideMemoire.
- Baselga, A.; Orme, C.D.L. betapart: an R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]

Table 1.
The P values of the analysis of variance for the effects of tillage and rotation on the alpha-diversity indices.
Table 1.
The P values of the analysis of variance for the effects of tillage and rotation on the alpha-diversity indices.
| DF | Simpson-TD1 | Shannon-TD2 | Chao1 | |
| Tillage (T) | 1 | 0.014 | 0.007 | 0.160 |
| Rotation (R) | 3 | 0.613 | 0.563 | 0.4292 |
| T×R | 3 | 0.217 | 0.041 | <0.001 |
1 Simpson-based True Diversity Index. 2 Shannon-based True Diversity Index.
Table 2.
Soil oomycete species relative abundance (%) as affected by tillage (T), either conventional tillage (CT) or no tillage (NT) and rotation (R) (including soybean monoculture (SSS), corn monoculture (CCC), wheat monoculture (WWW) and corn-soybean-wheat rotation (CSW), the associated P-values from the analysis of variance, and potential hosts and disease symptoms. Species below 0.001% relative abundance not included.
Table 2.
Soil oomycete species relative abundance (%) as affected by tillage (T), either conventional tillage (CT) or no tillage (NT) and rotation (R) (including soybean monoculture (SSS), corn monoculture (CCC), wheat monoculture (WWW) and corn-soybean-wheat rotation (CSW), the associated P-values from the analysis of variance, and potential hosts and disease symptoms. Species below 0.001% relative abundance not included.
| Species a | Relative abundance (%) | Analysis of variance (P value) b | Disease Note c | Known Hosts c,d | ||||||||||||||
| CT | NT | T | R | T*R | R CT | R NT | ||||||||||||
| No. ASV | CCC | SSS | WWW | CSW | CCC | SSS | WWW | CSW | ||||||||||
| Globisporangium aff. hypogynum | 1 | 0.000 | 0.000 | 0.000 | 0.153 | 0.000 | 0.000 | 0.000 | 0.000 | 0.320 | 0.78 | 0.772 | 0.7817 | N/A | ||||
| Globisporangium apiculatum | 20 | 0.664 | 16.294 | 7.970 | 16.522 | 0.000 | 0.000 | 0.000 | 0.002 | <0.001 | 0.036 | 0.031 | 0.041 | 0.761 | grape* | |||
| Globisporangium attrantheridium | 49 | 57.290 | 33.493 | 40.079 | 41.743 | 38.209 | 46.590 | 47.258 | 59.105 | 0.230 | 0.448 | 0.021 | 0.066 | 0.138 | Cavity spot lesions | Daucus carota, Prunus, soybean | ||
| Globisporangium glomeratum | 11 | 2.273 | 2.341 | 1.381 | 2.454 | 1.770 | 2.658 | 3.176 | 2.208 | 0.96 | 0.833 | 0.936 | 0.988 | 0.734 | soybean | |||
| Globisporangium heterothallicum | 27 | 10.438 | 11.844 | 2.942 | 6.979 | 13.878 | 7.603 | 7.139 | 3.802 | 0.539 | 0.039 | 0.558 | N/A | 0.076 | Damping off | Pepper, soybean, corn, spinach, wheat, lentils | ||
| Globisporangium hypogynum | 3 | 0.000 | 0.930 | 0.000 | 0.000 | 1.115 | 2.277 | 0.000 | 0.728 | 0.055 | 0.075 | 0.902 | 0.061 | 0.381 | Root rots | soybean | ||
| Globisporangium intermedium | 3 | 1.000 | 0.000 | 4.290 | 0.388 | 0.000 | 0.995 | 0.281 | 1.700 | 0.755 | 0.074 | 0.001 | 0.004 | 0.407 | Damping-off, rots | abutilon, antirrhinum, arabis, beet, begonia, carrot, cauliflower, chamaecyparis, cherry laurel, chrysanthemum, cotoneaster, cucumber, erica, ferns, godetia, hazel, hop, hyacinth, lettuce, leyland cypress, lupin, nasturtium, pea, pear, pelargonium, pepper, saintpaulia, strawberry, fragaria vesca, tomato, violet, yew, soybean, corn | ||
| Globisporangium irregulare | 2 | 0.000 | 0.000 | 0.000 | 0.247 | 0.074 | 0.000 | 0.000 | 0.000 | 0.559 | 0.867 | 0.563 | 0.782 | 0.347 | Blight, damping off, root and other rots, etc | soybean, wheat, corn | ||
| Globisporangium iwayamae | 2 | 0.000 | 0.000 | 1.860 | 0.163 | 0.000 | 0.000 | 0.000 | 0.000 | 0.024 | 0.001 | 0.0007 | 0.002 | N/A | Rots | Poaceae, wheat | ||
| Globisporangium macrosporum | 2 | 0.000 | 0.000 | 0.000 | 0.181 | 0.055 | 1.220 | 0.797 | 0.546 | 0.029 | 0.658 | 0.738 | 0.782 | 0.682 | Damping-off, root rot | iris, soybean, corn | ||
| Globisporangium nunn | 1 | 0.000 | 0.000 | 0.000 | 0.000 | 1.042 | 0.000 | 0.000 | 0.000 | 0.333 | 0.349 | 0.339 | N/A | 0.3415 | soybean | |||
| Globisporangium orthogonon | 1 | 0.000 | 1.578 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.305 | 0.323 | 0.264 | 0.3061 | N/A | corn | |||
| Globisporangium pleroticum | 5 | 2.38 | 0.3683 | 0.000 | 0.193 | 0.952 | 1.170 | 0.000 | 1.760 | 0.7545 | 0.074 | 0.009 | 0.711 | 0.841 | peas, soybean, lupins | |||
| Globisporangium recalcitrans | 14 | 1.103 | 6.190 | 0.000 | 1.321 | 3.613 | 4.969 | 3.326 | 1.693 | 0.131 | 0.194 | 0.566 | 0.219 | 0.435 | Root rot, damping off. | beet, hebe | ||
| Globisporangium rostratifingens | 5 | 0.000 | 0.000 | 0.000 | 0.329 | 0.725 | 0.233 | 0.000 | 0.000 | 0.699 | 0.671 | 0.107 | 0.529 | 0.174 | Root rot | pea, soybean, corn, wheat | ||
| Globisporangium selbyi | 8 | 2.200 | 2.223 | 3.130 | 2.350 | 0.591 | 0.774 | 2.410 | 2.290 | 0.702 | 0.859 | 0.996 | 0.943 | 0.750 | Lesions | corn, soybean | ||
| Globisporangium sylvaticum | 25 | 4.970 | 2.344 | 9.556 | 3.796 | 19.620 | 12.984 | 3.839 | 7.282 | 0.006 | 0.330 | 0.086 | 0.334 | 0.063 | Root disease, rots | apples, carrot, cherry laurel, cress, chrysanthemum, cucumber, garlic, lettuce, onion, pea, radish, rhododendron, spinach, strawberry, yew, wheat | ||
| Globisporangium ultimum | 11 | 2.346 | 14.656 | 1.834 | 6.589 | 2.490 | 2.820 | 0.000 | 1.348 | <0.001 | 0.008 | 0.011 | 0.002 | 0.991 | Blight, damping off, root rot, etc | soybean, garlic, grape, hyacinth, lettuce, lily, lupin, melon, mustard, onion, parsley, pea, pear, pelargonium, pepper, poinsettia, primula, radish, rhododendron, rhubarb, spinach, strawberry, sweet pea, tomato, tulip, wallflower, yew | ||
| Phytophthora cf. inundata D0S1P25 | 1 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.237 | 0.000 | 0.000 | 0.180 | 0.093 | 0.152 | N/A | 0.092 | ||||
| Pythium acanthicum | 1 | 0.000 | 0.000 | 0.000 | 0.073 | 0.000 | 0.000 | 0.000 | 0.000 | 0.315 | 0.724 | 0.763 | 0.711 | N/A | Downy mildew. Blight, damping off, rots, etc. | soybean, corn | ||
| Pythium aff. monospermum | 3 | 1.956 | 0.000 | 10.579 | 0.949 | 2.768 | 0.774 | 14.025 | 2.828 | 0.263 | <0.001 | 0.794 | <0.001 | 0.073 | grapevine | |||
| Pythium arrhenomanes | 8 | 1.116 | 0.000 | 0.000 | 0.872 | 1.257 | 3.545 | 5.713 | 2.795 | 0.014 | 0.782 | 0.223 | 0.492 | 0.409 | Blight, root rot | corn, rice, barley, wheat | ||
| Pythium monospermum | 12 | 2.109 | 0.872 | 1.683 | 1.187 | 1.274 | 1.928 | 3.108 | 2.005 | 0.367 | 0.857 | 0.725 | 0.476 | 0.962 | Downy mildew. Root necrosis, not known as a strong pathogen | cherry, juniper, spinach | ||
| Pythium oligandrum | 1 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.200 | 0.338 | 0.763 | 0.772 | N/A | 0.762 | Damping-off; root, stem, and fruit rots | soybean, wheat | ||
| Pythium oopapillum | 2 | 0.000 | 0.158 | 0.000 | 1.536 | 0.000 | 0.000 | 0.000 | 0.404 | 0.35 | 0.48 | 0.931 | 0.678 | 0.761 | Root rot | soybean | ||
| Pythium torulosum | 1 | 0.000 | 0.000 | 0.000 | 1.326 | 0.000 | 0.000 | 0.000 | 0.000 | 0.144 | 0.503 | 0.477 | 0.469 | N/A | damping-off, root rot | pea, soybean, corn | ||
| Pythium volutum | 6 | 0.000 | 0.000 | 6.287 | 4.399 | 0.000 | 0.002 | 1.895 | 0.053 | 0.020 | 0.004 | 0.143 | 0.034 | 0.014 | Root rot | melon, morning glory, wheat, barley, turfgrass | ||
| Saprolegnia anisospora | 1 | 0.000 | 0.000 | 1.997 | 0.000 | 0.000 | 0.238 | 0.000 | 0.000 | 0.704 | 0.024 | 0.01 | 0.006 | 0.379 | ||||
| Saprolegnia torulosa | 1 | 0.000 | 0.175 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1.093 | 0.142 | 0.384 | 0.181 | 0.306 | 0.265 | ||||
| Wilsoniana amaranthi | 1 | 0.000 | 0.000 | 0.000 | 0.638 | 1.252 | 0.743 | 0.000 | 0.000 | 0.646 | 0.876 | 0.369 | 0.782 | 0.515 | White blister rust | amaranth | ||
| Wilsoniana portulacae | 6 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 5.157 | 0.000 | 1.132 | 0.031 | 0.19 | 0.211 | N/A | N/A | White blister rust | portulacaceae | ||
a Species are in alphabetic order. b P values are in bold for significant impact of treatments. c Lifestyle and host information are acquired from https://nt.ars-grin.gov/fungaldatabases/. d Hosts included in the study are in bold.
Table 3.
The effects1 of tillage (T) (conventional tillage; CT vs. no-till; NT), and rotation (rotation vs. monoculture for each individual crop) on corn, soybean, and wheat yields (kg ha-1) during 2016-2018.
Table 3.
The effects1 of tillage (T) (conventional tillage; CT vs. no-till; NT), and rotation (rotation vs. monoculture for each individual crop) on corn, soybean, and wheat yields (kg ha-1) during 2016-2018.
| Corn yield | Soybean yield | Wheat yield | |
| Tillage | |||
| CT | 9177b1 | 2257 | 1854 |
| NT | 10806a | 2652 | 1795 |
| Rotation | |||
| Monoculture | 9735 | 2404 | 1298b |
| Rotation | 10248 | 2446 | 2305a |
| Analysis of variance (P-values) | |||
| Tillage (T) | 0.018 | 0.226 | 0.548 |
| Rotation (R) | 0.442 | 0.577 | <0.001 |
| T×R | 0.163 | 0.343 | 0.652 |
1 Letters represent significant difference between treatments in tillage or rotation category at P = 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



