Submitted:
27 April 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
Sperm gain fertilization ability and forward movement characteristics during epididymal transport. Sperm maturation is inseparable from the processing of the epididymis. Compared with other reproductive organs, relatively little is known about the epididymis. However, with the development of molecular biology and the improvement of single-cell sequence technology, we gradually realized that we should re-examine the importance of the epididymis for sperm mat-uration and for the entire male reproductive system. Recently, epididymis related reports have tended to be more regionalized and more refined. We can understand cell distribution and gene expression from the perspective of a single cell. It has been reported that different regions of the epididymis have different functions and different cell compositions, which may be determined by gene expression. Here, we comprehensively reviewed the epididymal studies. From the ana-tomical structure of the epididymis to the division of its regions, we focused on the cell compo-sition and region-specific genes differentially expressed in the epididymis. The gene expression of the epididymis extends to a series of biological processes of sperm in the epididymal cavity, and the serious effects of epididymal diseases on reproduction are discussed. Therefore, the importance of the epididymis in male reproduction is emphasized. With the maturity of single-cell spatial transcriptomics, it is hoped that the technology can be applied to epididymal research and bring about the spatiotemporal landscape of the epididymis at the single-cell level.
Keywords:
Epididymis
; Regional
; Single cell/Spatial Transcriptomics
1. Introduction
The epididymis, which is mainly composed of the epithelial propria membrane and thin annular muscle, has a secretory function. It plays a nutritional role in sperm, and promotes sperm to continue to differentiate and mature [1]. The length and morphology of the epididymis vary between the species. It is worth noting that the epididymal tube of mice is just over 1 meter (approximately 100 times its length); 3 meters in rats; and six meters in humans [2,3,4,5]. The epididymis is a member of the male duct system, developed in an androgen-dependent manner by the Wolffian duct [6]. It has been shown that the Hoxa10 and Hoxa11 genes, both members of the larger homeobox (Hox) gene family, appear to be essential in the development of Wolffian duct into the epididymis [7,8].
The function of the epididymis is closely related to the sperm maturation. It has been established that the spermatozoa in mammals undergo a ‘ripening process’ as they descend the epididymis [9]. It takes an average of 12 days for the epididymis to transport human sperm, but it can occur in as little as three days [10]. In addition to these maturation steps, the epididymis also provides a complex microenvironment that enables sperm cells to remain stationary during epididymal storage, avoiding premature sperm activation [11]. Simply put, the epididymis has four functions: to store sperm, transport sperm, protect sperm, and remove sperm.
The earliest report on the epididymis was released in the 18th century [12], which elaborated on discutient application to the indurated epididymis. Early understanding of the epididymis comes from various pathological studies[13,14,15] or anatomical structures and functions [16,17]. Related research has emerged in large numbers since the 1950s. In the past 70 years, people have explored the epididymis from different aspects. Initially, pioneers revealed the anatomy of the epididymis in horses[18], guinea pigs[19] and humans[20]. Later, relevant cellular and chemical analyses began to appear. Such as cytochemical study of mouse epididymis, histochemical study of glucose-6-phosphatase in mouse epididymis and study on the content of trace elements in sperm cells of epididymis[21,22,23]. There are also a large number of articles related to epididymitis and tumors[24,25,26,27]. Since the 1960s, research on the epididymis has emerged continuously, and the annual publication volume has increased linearly (Figure 1) (https://pubmed.ncbi.nlm.nih.gov/?term=epididymis&filter=dates.1960-2022). During this period, research on the human epididymis involved in obtaining well-preserved biological tissues, especially transcriptomics and proteomics [28,29,30]. With the development of technology and knowledge, people have begun to pay attention to its function. Then, an important role of the epididymis in sperm maturation was discovered [31,32]. More articles on the epididymal proteome, transcriptome and genetics have since emerged [33,34,35]. With the help of this new technology, some progress has been made in the study of epididymal-specific gene expression and function [36,37]. Thus far, we know that it is a highly segmented structure, and each segment expresses its own and overlapping genes, proteins and signal transduction pathways in the epididymis [38]. As shown in Figure 2, articles on region-specific gene expression in the epididymis began to appear in 1983, and the number of articles published each year increased every year until 2006, but the popularity has decreased since then(https://pubmed.ncbi.nlm.nih.gov/?term=epididymis+region+specific+gene+expression&sort=pubdate&sort_order=asc).
2. Research status of epididymis in recent years
The most commonly used epididymis research models are the mouse and rat. In recent decades, new technologies, such as the gene-knock approach[39],transcriptome sequencing[40], microarray[41], proteomics[29], and RNA-seq, which is a sequencing method of transcriptome sequencing, also known as second generation sequencing[42,43,44], have increased understanding of epididymal composition significantly. Many knockout mice exhibit phenotypes that affect male reproductive function, and in these models, males are often infertility. For example, sperm from Clgn, Adam1a, Adam2, and Adam3 knockout mice did not migrate to the fallopian tubes[45,46,47]. Whether and how these genes are expressed in the epididymis is worth exploring. In vivo and in vitro models have been used to study the function of epididymis. The technique of in vitro culture of epididymal cells has been very mature[48,49,50]. Rat caput epididymal cell line shows many characteristics of epithelial principal cells, thus providing a model for the study of the function of epididymis cells[51]. The cell line is composed of epithelial principal cells with the characteristics of cells in vivo[52]. Advances in RNA sequencing have allowed researchers to analyze the transcriptome of classified groups of cells or even individual cells[53]. In recent years, with the development of single-cell sequencing technology, the understanding of the epididymis has reached a new level. For example, single-cell sequencing techniques can be used to map the epididymal single-cell transcriptome. Correlational research revealed a new distribution pattern of mitochondria and key genes, which may be related to the first and subsequent sperm wave [54]. In the past two years, spatial transcriptomics has provided a large amount of quantitative gene expression data of mRNA distribution in tissue slices and visualized the data. These emerging technologies make new bioinformatics analysis possible and valuable in research and diagnosis[55]. At present, there is no relevant research in the field of epididymis. It is expected that future epididymal spatial transcriptome research will reveal more secretion in the epididymis. This paper summarizes the related epididymis research techniques. Representative articles on the application of new technologies in the field of epididymis research are listed in Table 1.
Research on the epididymis was more focused on tissue before the emergence of transcriptome technology. With the advent of transcriptome technology, people realized the differences in genes and proteins in different parts of the epididymis[56]. Genes determine function, and the difference in function between regions is due to different gene expression. This means that different parts of the epididymis have different roles or functions in sperm maturation. Starting from the perspective of genes and cells, we have a better understanding of the spatial distribution and cell types of epididymal cells or genes in different parts of the epididymis. It deepened our understanding of the epididymis and paid more attention to regional differences.
3. Regional division and differential function
It is reported that the epididymis is a highly region-specific reproductive organ. Regions of the epididymis are artificially divided, so different regional division methods are recorded in the literature. In most studies, the epididymis is divided into three regions: caput, corpus and cauda. However, in rodents, the epididymis is divided into four anatomical regions: the initial segment, caput, corpus and cauda[57]. These regions are well recognized in rodents, but not well defined in human epididymis. To date, most studies have used the caput, corpus, and cauda as the boundaries in the epididymis for analyzing gene and protein expression patterns [58,59]. The caput, corpus, and cauda of the epididymis are proximal to distal, relative to the testis (Figure 3).
Jun Zhao et al. conducted anatomical analysis of the human epididymis based on histology, proteomics and 3D reconstruction. They subdivided the caput, corpus and cauda of the epididymis into 6, 10 and 4 parts and found that protein expression in different regions of the epididymis was different depending on the region[60]. It has been found that the protein composition of the lumen of the epididymis is different, and the proteome and secretory group of the caput, corpus and cauda of the epididymis are also different [61]. However, the regional division of the epididymis in other species is also different. There are subtle differences in the regional division of the epididymis, even in rodents. The mouse epididymis is composed of 10 different segments, while the rat epididymis is composed of 14 segments[56,62]. Besides, the size and shape of special segments of the epididymis vary slightly between different species[63].
Each region of the epididymis has a different function. By identifying miRNAs in different parts of the epididymis, some miRNAs displayed regionalized expression patterns[64]. Compared with other parts, the function and expression of the epididymal caput are more abundant, and the function of the caput is quite unique. One of its key functions is to maintain an appropriate cavity environment for normal sperm maturation[65]. Moreover, the caput is closely related to hormone stimulation. Microarray analysis showed that the epididymal caput has the highest number of androgens regulating genes [59,66,67].
The functions of the corpus and cauda are similar to a certain extent. In morphology, the physical distance between caput and cauda is the longest and the functional difference is the greatest. Specifically, immune-related biological processes were enriched in the epididymal corpus and cauda. The cauda of the epididymis is the final storage of mature sperm before ejaculation[68]. Generally, sperm are stored in the cauda. Sperm gain fertilizing capacity during transport between the corpus and cauda of the epididymis. At present, sperm in the epididymal corpus area have been proven to have a similar ability to cauda sperm, capable of capacitation, in vitro fertilization of oocytes and survival in cryopreservation[69,70,71]. Animal sperm cannot survive for more than a few days in the caput and corpus of the epididymis involved in maturation, while some animal sperm can maintain fertility for several weeks in the caudal environment[72].
Like the caput of the epididymis, the biosynthesis of secreted proteins in the cauda of the epididymis is also regulated by androgen and is also related to temperature[72,73,74,75].Compared with other large mammals, male sperm production is low, and sperm morphology and motility are generally poor. The regulatory mechanism of the cauda of the epididymis deserves further investigation. However, we do not know the mechanism of temperature in the epididymis[76]. This may be a breakthrough in future epididymis research. In general, each part of the epididymis has a unique function, which continuously processes and elaborates sperm and finally allows sperm to obtain fertilization and motility.
4. Different cell types in different epididymis regions
4.1. Epididymis epithelial and cell types
In the past twenty years, active research has been devoted to understanding the contribution of the epididymal structure and its different cell types to sperm function. The epididymal epithelium is region specific and has rich functional diversity in the epididymal caput, corpus and cauda. Each of these fragments has a specific role in ensuring that sperm can obtain full mobility and fertility. The epithelium in the epididymis presents a columnar pseudolayer over the entire length, the epithelial lining consists mainly of principal cells and basal cells[77]. In the undifferentiated stage of epididymis development, epithelial cells are composed of small undifferentiated columnar cells[78]. Then differentiated short columnar cells differentiate into basal cells and high columnar/narrow cells, which then develop into principal, clear, narrow and apical cells[79].
Epididymal epithelial cells have been successfully isolated and cultured in many species, such as bulls, mice, rabbits and humans[80,81,82,83,84]. Some studies indicate that epithelial-mesenchymal interactions are a component of epididymal morphogenesis[85]. The epididymal mesenchymal - epithelial interactions, such as the phenotypic effects of androgens, are the result of complex interactions between different and specific androgenic target cells[86]. Besides, epididymal epithelium provides an optimal acidic liquid microenvironment in the epididymal canal, enabling sperm to complete the maturation process[87,88]. All epididymal regions have a variety of epithelial cell types, each of which has different functions. The epididymal epithelium regulates the transport and absorption of ions, solutes, and macromolecules secreted by different cells and regions, providing a stable luminal environment for sperm maturation.
The earliest cell types were identified, respectively into four types: principal, narrow, clear and basal cells[58]. Later, it was found that the epididymal epithelium was composed of six different cell types: principal cells, narrow cells, apical cells, clear cells, halo cells and basal cells[89]. Some papers divide epididymal epithelial cells into five types: principal cells, narrow cells, clear cells, apical cells and basal cells[90]. Not all species have the same type of epididymal cells. The cell types in the epididymal epithelium are also species-specific. Some cell types vary according to species, including apical cells, narrow cells, clear cells, and halo cells[77]. Recently, different cell subtypes of epididymis were identified by single cell RNA-sequencing samples[42,91]. In 2020, Shih-Hsing Leir et al recovered 1,876, 1,309, and 2,114 cells from human epididymis aged 3,157, and 32 years, respectively, which passed quality control on the 10X Genomics Chromium System pipeline, providing scRNA-seq data on a pool of 5,299 cells. Eight humans proximal epididymal cell types were identified include principal cells, basal cells, clear cells, stromal cells, apical/narrow, immune cells, sperm cells, and efferent duct cells[91]. Each cell type can be divided into several cell subtypes based on different marker genes. Using droplet-based (10X Chromium) single-cell RNA-Seq, Gene expression in 8880 mouse epididymis and vas deferens cells was analyzed. Nine principal cell subtypes and three basal cell subtypes were analyzed from this data[42].
4.2. Principal cells
The most abundant cells are the principal cells, which are distributed throughout the whole epididymis. According to different regions, the principal cells account for 65% to 80% of the epididymal epithelium. It is reported that, the number of principal cells is at least three times more than the sum of all other cell types[92,93]. These cells are mainly responsible for the absorption and secretion of substances into the epididymal cavity, so they have high secretory and endocytotic activity[89]. In addition, the principal cells are the places where liquid, ions, antioxidants and exons are produced and released[94].
The epididymal junction complex between adjacent principal cells consists of apical spaces, adhesions and tight junctions. Which leads to the formation of the blood epididymal barrier[95]. The blood-epididymis barrier limits the molecular exchange between blood and the lumen, providing guarantee for the stability of the epididymal lumen environment. In this way, specific microenvironments are formed in the epididymal cavity, which is crucial for sperm maturation [96]. Once the principal cell of epididymis has problems, it is likely to affect sperm motility quality and fertilisation capacity. It is worth noting that Occludin (OCLN), a tight junction protein, appears at the apical junction of the principal cells of the proximal epididymis epithelium, OCLN - deficient mice had impaired sperm motility and fertilization ability and mutation of OCLN deletion can cause male sterility in mice.[97].
4.3. Basal cells
Basal cells exist in the columnar pseudostratified epithelium covering all mammalian epididymis[77]. They are present in the epididymal epithelium of all regions (caput, corpus, cauda) in all species[77,93]. Located in the basal layer, it is said to never reach the lumen. The morphology of mammalian epididymal basal cells showed similarity. They are oval or round, large, round, and are located on the basement membrane. It is generally believed that there are more basal cells in the epididymal corpus and fewer in the cauda[98,99]. Initially, basal cells are thought to have macrophage-like qualities and are associated with epididymal immunity[100,101]. The latest explanation for the role of epididymal basal cells is that they may scan and perceive the lumen environment of pseudostratified epithelium and regulate epithelial function through the mechanism of crosstalk with other epithelial cells[102]. In addition to crosstalk with other cells to regulate epithelial function, basal cells can also monitor the epididymal fluid environment. The basal cells of the epididymis act as the sensor of angiotensin 2, and regulate the lumen acidification by producing nitrous oxide[90], which may play a protective role in the blood-epididymis barrier.
Early studies showed that basal cells of epididymis can be used as stem cells[103]. Gene transcription reveals that the basal cells express a large number of genes related to the function of pluripotent stem cells[104]. Studies in rats and mice have shown that purified single epididymal basal cells cultured under 3D conditions can self-renew and differentiate into other types of epididymal cells[105]. Several signal pathways related to stem cell function, such as Notch1, Hedgehog, p63 and Wnt are also enriched in basal cells[106]. Some signaling pathways are related to sperm function. Among these signaling pathways, Wnt ligands can promote the stability of sperm proteome and sperm motility. Studies have shown that post-transcriptional Wnt signals control the maturation of epididymal sperm[107]. And the Wnt4 signaling pathway can affect the activity of spermatogonial stem cells[108]. The relationship between stem cell-related signaling pathways in basal cells and sperm function is fascinating.
In a study of the ultrastructure of the epididymal epithelium of equine animals, a single cilium was incidentally found. Primary cilia were detected in the epididymal basal cells of horses, donkeys and mules studied. These cells appeared on the surface of basal cells and infiltrated into the intercellular space. When the basal cell cilium is damaged, the epididymal epithelial cells show unbalanced apoptosis and growth rate[109].
4.4. Other cells
Apical cells are mainly located in the initial segment of the epididymal epithelium and have endocytosis activity. Narrow cells also exist only within the initial segment, as shown by the name, narrower than the adjacent principal cells. These cells have been shown to secrete H+ ions into the epididymal lumen and are responsible for endocytosis[89]. Apical cells and narrow cells are rare and only confined to the cranial region[110,111]. Halos are the primary immune cells in the epididymis [112].
Clear cells are large endocytotic cells scattered among the principal cells in the caput, corpus and cauda regions. Their morphology is characterized by many covered pits, vesicles, endosomes, vesicles, lysosomes, and lipid droplets in the apical region [113,114,115]. In addition to secretion, endocytosis of lumen proteins is the main function of epithelial clear cells [116]. Narrow, apical and clear cells strongly express vacuolar proton pump ATPase (V-ATPase) in their apical membranes, which is considered to be the reason for proton secretion into the lumen, thus participating in its acidification[117,118,119].
Intraepithelial single cell types may perform separate and integrated functions in the epididymis. Recent studies have shown that basal cells regulate the transport of electrolytes in principal cells by releasing paracrine factors, especially prostaglandin PGE2[120]. It has been concluded that the interaction between cells is the basis for the formation of the epididymal environment. Therefore, the complex interactions between different cell types in epithelial cells lead to the activation of the ability of epididymal luminal acidification, which is crucial for sperm maturation and storage.
5. Different genes in different epididymis regions
5.1. Specially expressed genes and gene families
Some scholars have long proposed the regional specificity of gene expression in the epididymis[112,121]. The mRNA of the caput, corpus and cauda of the epididymis was sequenced by transcriptome sequencing. Specific mRNA synthesis supports the concept that the region-specific expression pattern of epididymal tubule transcription is the main molecular basis of the region specific expression pattern of tubule lumen proteins[112]. Furthermore, each gene has different spatial expression and regulation in the epididymis. In different mammalian species, gene expression patterns vary between epididymal segments[56,122,123]. In the following, we will specify the expression of representative epididymal genes. The genes expressed in the epididymis play an important role in sperm maturation and motility, changes in epididymal-specific gene expression may lead to infertility. Some genes are predominantly expressed in the epididymis, which thus suggests that these genes may possess epididymis-specific functions.
At the beginning of the 20th century, the mouse epididymis transcripts were analyzed on a 1,7000 oligonucleotides microarray. By comparing the transcripts of the epididymis and 22 other organs, 75 epididymal-specific transcripts were identified, and 2,168 genes were identified. These genes were up or downregulated more than four times between at least two different fragments of mouse epididymis[56]. In the end, it was determined that 77 transcripts were uniquely expressed in the epididymis[122]. Analysis based on rat microarray technology found that the number of genes detected in each segment of the epididymis varied greatly. Although 43% or 46% of the studied genes were expressed in the initial fragment (517 of 1,176 fragments) and caput (541 of 1,176 fragments), respectively, only 39% of the genes were expressed in the corpus (463 of 1,176 fragments). The most expressed genes, 53%, were located at the cauda (626 out of 1,176 genes)[58]. Recently, the region-specific expression genes were analyzed by GO and KEGG analysis to screen the key genes related to sheep sperm maturation. A total of 129, 54 and 99 specific genes were obtained in the caput, corpus and cauda, respectively. The heatmap showed that 6 genes of the LCN protein family were highly expressed in the sheep epididymal caput. RNase10 is also highly specifically expressed in caput. The same is true of other species that have identified RNase 10, such as humans, mice, and pigs[124,125,126].
5.2. Epididymal distribution and function of the Defb family
RT‒PCR showed that all but one of the rats defensin transcripts were preferentially expressed in the male reproductive tract, especially in the epididymis and testis[127]. Defensins are small cationic peptides related to innate immunity, that are part of the first line of defense against invading pathogens. β-defensins are a subgroup of the defensin family[128]. β-defensins have an antibacterial function and provide the first protection against pathogens while playing the role of immune regulation[129]. Since the β-defensins were first identified as an antibacterial drug in bovine airway epithelial cells, 30-50 new defensin genes have been found in bovines, humans and other species[127,130,131]. Some defensins are mainly expressed in the epididymis and show a region-specific expression pattern in the epididymis[132].
Studies over the past 15 years have shown that β-defensins are abundantly expressed in the postnatal epididymis of different species. When they travel along the epididymis, they secrete into the coelom and combine with sperm, indicating that they are involved in specific reproductive tasks[133]. Epididymal β–defensins not only have immune function but can also promote sperm maturation, most of which are regulated by androgens. Most of the β–defensins, such as Defb18, 19, 20, 39, 41, and 42, expressed in the epididymal cauda are affected by androgens[134,135,136]. In addition, β–defensins cooperate with other genes to regulate epididymal function. Recently, research has shown that DICER1 plays an important role in the transcription of β-defensin genes, which are associated with natural antimicrobial properties in a way that is independent of microRNAs and further affects male fertility[137].
Combined with literature analysis, Defb12, Defb35, Defb15, Defb34, Defb17, Defb18, Defb21, and Defb25 are more abundantly expressed in the caput of the epididymis[127,138,139]. Among these genes, the rat epididymis-specific Defb15 exhibits an androgen-dependent expression pattern[134], and it can bind to the acrosome of caput sperm. It also plays a dual role in rat epididymal sperm maturation and pathogen defense[140]. Defb41 is required for proper sperm maturation. Targeted inactivation of mouse epididymal Defb41 alters the sperm flagellar beat pattern and zona pellucida binding[141]. Similar to the epididymis caput, Defb9, Defb10, Defb11, Defb13, and Defb40 are highly expressed in the cauda[127,138,139].
Human DEFB126 is derived from epididymis and adsorbed on sperm surface. DEFB126 is mainly expressed in the epididymis caput, and its level is down-regulated in the epididymis caput of infertile patients with non-obstructive azoospermia[142]. Besides, DEFB126 gene mutation will lead to significant changes in sperm surface components and damage the ability of sperm to penetrate mucoid gel[143]. Immunofluorescence staining showed that the proportion of DEFB126 positive sperm in motile sperm increased significantly. In addition, the proportion of DEFB126 labeled sperm was positively correlated with sperm motility and normal morphology[144]. Sperm must pass through the female reproductive tract and undergo complex chemical reactions to obtain the ability to fertilize. DEFB126 has been suggested for coating sperm, whose overall negative charge helps sperm adhere to the epithelium of the fallopian tube and promotes the penetration of cervical mucus[145]. In a word, DEFB126 is very important in human sperm maturation, and it has potential use in vitro sperm maturation.
5.3. Epididymal distribution and function of the Lcn family
The lipoprotein (LCN) family is a hydrophobic ligand binding protein with a conserved structure that is expressed in all major taxa from prokaryotes to primates [146,147]. Lipocalins show diverse functions, including the transport of hydrophobic molecules, regulation of inflammation[148], glucose and lipid metabolism[149] and immunity[150]. Notably, features of sperm maturation through the epididymis include changes in the lipid composition of the plasma membrane[18]. There are several lipids in the mouse epididymis that are thought to play a role in sperm maturation by transporting lipophilic molecules. Two lipocalin genes derived from ancestral genes, mERABP (mouse epididymal retinoic acid binding protein) and mEP 17 (mouse epididymal protein of 17 kDa), have been previously reported to be specifically expressed in the epididymis[151]. Studies have shown that the Lcn9, Lcn10, Lcn12, Lcn5, Lcn8, and Lcn13 genes are specifically expressed in the mouse epididymis[152]. However, it is still unknown what effect Lcn9 and Lcn12 have on epididymal function.
Lcn2, also known as 24p3, is expressed in the caput and corpus of the mouse epididymis [153]. Lcn6 protein is abundant in the epididymal caput epithelium, and is located in the sperm tail and the region behind the acrosome of the head[147]. In mice with Lcn6 gene deletion, sperm maturation defects were observed, including calcium overload and premature acrosome reaction. The expression of Lcn6 in the distal efferent duct and epididymal caput and the location of sperm surface proteins are consistent with the role of sperm maturation[147]. A LCN6 study on the human epididymis showed that its expression in the distal efferent ducts, initial segment and proximal caput was similar to that of rat Lcn5[154] and mouse Lcn8[151] but different from that of mouse Lcn5, which was expressed in the distal caput and corpus and accumulates in caudal fluid[155,156].
5.4. Other gene expression conditions in epididymis
Similar to the Defb family, CD52 is also an immune-related gene. CD52 is expressed in mammals such as dogs, rats, mice, monkeys and humans. The mouse CD52 antigen gene is homologous to human CD52[157]. It has been proven that CD52 is highly expressed in the cauda of the epididymis and moderately expressed in the corpus. Mouse CD52 is regulated by androgen and lumicrine factors[158].
CRISP is a highly conserved and widely distributed family in vertebrates and is called cysteine rich secretory protein. Although mutant sperm lacking CRISP1 showed obvious sperm function defects, males with CRISP1 deletion remained fertile[159,160,161]. CRISP1 and CRISP4 are also specifically expressed in the mouse epididymis[162,163]. Rat epididymal CRISP1 is expressed in the proximal part of the epididymis and binds to sperm during epididymal transport. CRISP1 is a multifunctional protein that plays different roles in the fertilization process through different connections with sperm and positioning on sperm[164]. It is worth mentioning that there is a compensatory mechanism between two homologous epididymal CRISP proteins. As CRISP1 observed, CRISP4-deficient males have in vitro fertilization defects, but animal fertility is normal[165].
Mammalian aquaporins (AQPs) are a family of 13 transmembrane aquaporins that are involved in water transport in many organs[166,167]. Aquaporin-1 (AQP-1) is a six transmembrane domain protein that is responsible for water transport. RT‒PCR showed AQP-1 mRNA expression in all mouse epididymal regions[168].A study on the epididymis of dogs showed that they were upregulated in the caput of the epididymis of dog cryptorchidism and downregulated in the cauda[169].The data shows that AQP9 is the main AQP of the epididymis, and specific mRNAs of AQP2, 5, 7 and 11 are also expressed in epididymal epithelial cells[167].
Juri Jeong et al. found that epididymal Spinks are expressed in an epididymal region-specific manner, and their expression is developmentally regulated[170]. In mice, four epididymal-specific Spink genes, Spink8, Spink10, Spink11 and Spink12, have been identified. They have unique regional expression patterns in the epididymis [171]. It has been determined that the another epididymal-specific gene Spink13 is crucial for acrosomal integrity and sperm maturation. In addition, Spink13 has been identified as significant to acrosome integrity [172]. Spink12 mutant mice showed normal fertility, indicating that Spink12 was functionally redundant in the epididymis[170]. The expression of Spink and other genes in the epididymis still needs further detailed analysis.
Therefore, we further analyzed the data provided in an article[42] with the Seurat software package, collecting the data of the mouse epididymal caput, corpus, cauda and vas deferens. Seurat is one of the most popular software suites for analyzing single-cell RNA sequencing data[173] and a computational strategy to infer cellular localization by integrating single-cell RNA-seq data with in situ RNA patterns[174]. After standardization, quality control and normalization of the data workflow were performed. We received an object of class Seurat 18097 features across 9167samples within 1 assay (Figure 4). In this data set, 1619 samples were collected from the caput of the epididymis, 2345 samples from the corpus of the epididymis, 1658 samples from the cauda of the epididymis, and 3545 samples from the vas deferens were the most. T-SNE (t-distributed stochastic neighbor embedding) is used to visualize these four parts of data, a dimensionality reduction method that allows data points to be visualized on a two-dimensional map. In the t-SNE map, features that have similar functions will come together, select each other and cluster in a coordinate system. Different colors represent different regions of the sample (Figure 4). Then, on this basis, the two main cell types in the epididymis, principal cell and basal cell, were extracted respectively and then clustered. According to the marker genes of different cell types, we specifically clustered the principal cells and basal cells by t-SNE and annotated them according to the four anatomical regions (Figure 5, Figure 6). There were 18097 features across 1410 samples within basal cells (Figure 5) and 18097 features across 2099 samples in principal cluster cells (Figure 6). Several genes of interest were selected, and their epididymal distribution in different parts was examined (Figure 7, Figure 8).
6. Gene expression varies at different epididymal positions and in different cell types
6.1. Comparison of gene expression in different parts of the epididymis
At present, the gene expression of human, mouse and ram epididymis from caput, corpus and cauda has been compared. By comparing the differentially expressed genes (DEGs) of human caput, corpus and cauda cells in different parts of the epididymis, it was found that the gene expression profiles of the corpus and cauda are very similar, and both differ from the caput to a similar degree. Gene ontology process enrichment analyses showed that ion transport, hormone stimulation and urogenital tract development were more obvious in the caput, and more defensive response in the corpus and cauda[65].
Microarray analysis of the epididymis in human shows that the cauda transcriptome is specialized in muscle contraction and establishment of localization[121]. Mouse microarray analysis find the relative expression of more than 60 genes between any two fragments was at least 100 times different. Defensin-β 9 (1832 times), adam28 (1790 times) and defensin-β 11 (981 times) are the highest in this category[56]. One study revealed expression in the human epididymis. More than 1,500,0 transcripts, almost one-third of which are found in the whole epididymis. Among them, 65% were detected in all three regions of the epididymis, 2.6% were present only in one region, and the remaining 32.4% were distributed in two regions. The region-specific transcripts observed in the caput (264), corpus (61) and cauda (81) epididymis are further classified as empirically determined reporter genes or expression sequence tags of some region-specific genes[175]. Another survey of rams obtained 1321 DEGs between the caput and the corpus (535 genes were more highly expressed in the caput and 786 genes in the corpus). Similarly, there were 855 DEGs between the corpus and the cauda (386 genes more highly expressed in the corpus and 469 genes in the cauda). A total of 1564 DEGs were identified between the caput and cauda (519 genes were more highly expressed in the caput, and 1045 genes were expressed in the cauda)[40].
6.2. Comparison of gene expression in different cell types of epididymis
As can be seen from the description above, the regional expression of epididymal genes is very complex and varies greatly among different species. Gene expression is different in different regions of epididymis, and the cell composition of each region is also different. This shows that we can explore the expression of genes in different types of epididymis cells. However, there is very little literature for reference. Based on document published by Rinaldi et al. in 2020 related to single-cell data analysis of the epididymis. Their team obtained sperm-depleted single-cell suspensions from eight epididymis of four 10-12 week-old FVB mice. The organ is cut into four parts, roughly corresponding to caput, corpus, cauda, and vas deferens. A single cell RNA sequencing library was prepared using the Chrome single cell 3' kit V2 (10X Genomics). Using single-cell data that their team uploaded to the GEO (Gene Expression Omnibus) website, we visualized the expression of several genes in basal cells and principal cells.
Principal cells and basal cells were identified in the integrated data of the caput corpus cauda and vas deferens. Interestingly, we found that the principal cells were very poorly distributed in the epididymis corpus, but the caput of the epididymis has very few basal cells and there were many basal cells in the epididymis corpus. Then, we selected the representative genes of interest and observed their epididymal expression in principal cells and basal cells. As shown in Figure 7, Lcn8 is reported to be an epididymis-specific lipocalin[176] which is highly expressed in caput basal cells (Figure 7A). Spag11b is associated with androgen receptor binding sites in its promoter or intron regions[134]. Its expression in basal cells is the same as that of Lcn8 (Figure 7D). Lcn5 is an epididymal retinoic acid-binding protein that belongs to the lipoprotein family. Its expression regulation in the epididymis has a unique gene expression mode[176]. The analysis results show that Lcn5 is expressed in caput and corpus basal cells (Figure 7B). However, Defb28 is expressed in all sections except the vas deferens of epididymis basal cells (Figure 7C). Unlike basal cells, Lcn5, Lcn8 and Spag11b are highly expressed in caput principal cells (Figure 8A, 8B, 8D), while Defb28 is only expressed in cauda principal cells (Figure 8C). Obviously, the expression of the same gene in different cell types is also different.
7. Epididymal spermatozoa and epididymis fluid
It has been recognized that the environment of the epididymal canal might influence the sperm maturation steps[177]. In the process of mammalian sperm transport along the epididymis, it encounters intraluminal fluid with different protein compositions, which interacts with the sperm surface and endows the male gamete with fertilization activity[178]. Many previous studies have shown that most sperm in the cauda of the epididymis are mature, capable of capacitation and acrosome reaction, and are fertile when used for in vitro fertilization (IVF) or artificial insemination (AI)[179,180,181].
The cell component of semen is mainly sperm, accounting for approximately 5% of the semen volume, while the cell-free part or semen contains secretions from different accessory gonads, accounting for 95% of the semen. In addition, 70-80% of the secretory body is composed of 10-20 major secretory proteins in epididymal fluid[34]. At the distal end of the epididymis, the sperm is fully mature and can be fertilized. There, they are preserved until ejaculation, and epididymal caudate fluid may be considered a natural semen preservation medium. Seminal plasma is a compound liquid composed of secretions from the testes, epididymis and accessory gonads. It has obvious species differences and forms ejaculation together with a small amount of epididymal sperm reserves[182]. In some species (such as pigs), seminal plasma accounts for the majority of total ejaculation, while epididymal lining cells and other cellular components (including a small amount of epididymal caudate fluid) account for only 5%[183]. Some animal species, such as poultry, ejaculate very little, but they are highly concentrated in the liquid formed by the testes, residual epididymal tubes and vas deferens secretion[184].
8. Dysfunction of epididymis and reproductive health
Obstructive azoospermia and non-obstructive azoospermia account for 20-30% of male infertility[185,186]. Moreover, the formation and function of the epididymis is partly androgen dependent. Studies have shown that the epithelial androgen receptor signaling pathway is critical to the postnatal development and function of the epididymal epithelium, and the destruction of this signaling pathway may lead to obstructive azoospermia[187]. Obstructive azoospermia can be an acquired obstruction of epididymis, or congenital due to mutations in the CFTR or ADGRG2 gene. Non-obstructive azoospermia is associated with abnormal gene expression, and some abnormal gene expression proteins are also expressed in the epididymis. However, at present, the focus of attention on non-obstructive azoospermia is focused on the study of testis, while there are few articles related to epididymis. In the future, we can integrate the research of epididymis transcriptome gene expression, focus on the influence of epididymis on non-obstructive azoospermia, providing a more detailed understanding of the molecular mechanism of non-obstructive azoospermia and potential therapeutic targets for its treatment. Therefore, we can start with the study of epididymis to break through male infertility diseases such as obstructive azoospermia and non-obstructive azoospermia.
The epididymis stores and transports sperm, so there is a unique immune environment in which the underlying autoimmune response must be suppressed in order to establish immune tolerance to sperm present after puberty[188]. Infection and inflammation, including epididymitis and epididymo-orchitis orchitis, account for 13-15% of all cases of male-related infertility[189]. Maintaining proinflammatory and immune tolerance is essential for the balance of male fertility. Fortunately, the epididymis has a certain barrier function called the blood epididymal barrier, which creates a suitable environment for sperm to mature.
Sperm are very sensitive to reactive oxygen species (ROS), which are the end product of normal metabolism of aerobic cells. At elevated levels it promotes oxidative damage and impairs sperm function[190]. Epididymal epithelial cells contain different antioxidant enzymes involved in protecting epididymal sperm from oxidative damage. The glutathione peroxidase (GPX) family is thought to be antioxidants and participate in the protection of the maturing spermatozoa in the epididymis. GPX5, as a member of GPX, is secreted by the epididymal caput principal cell and regulated by androgens[191]. Another antioxidant enzymes include superoxide dismutase (SOD), catalase (CAT)[192]. Activins play a complex role in controlling inflammation and immunity of the epididymis and vas deferens. The epididymis caput is relatively resistant to inflammatory damage caused by autoimmune or bacterial infection, but the caput shows low activin expression and high level of activin binding protein folliclestatin, which is highly susceptible to inflammatory damage [193].
9. Discussion
The epididymis is a complex convoluted tube that connects the vas deferens and the efferent ducts in the male reproductive tract and plays a vital role in regulating sperm maturation and fertilization. Sperm capacitation requires epididymal processing, which includes changes in the structure and molecules of spermatozoa to fertilize an ovum[194]. This includes the immune sterilization process and various metabolic physiological processes. Some sperm can be stored in the cauda of the epididymis. For the whole male reproductive system, the epididymis plays an important role in connecting the former and the latter. However, it seems that the effect of the epididymis on male infertility has been ignored[195].
Before the 20th century, the transcriptome did not emerge; therefore, epididymis research was limited to pathology and histology[196,197,198,199]. For example, specialists studied the sperm quality of the caput, corpus and cauda area of the epididymis in healthy and sexually mature Landrace pigs[200]. In such studies, the characteristics of epididymal sperm were examined by light microscopy (LM), scanning electron microscopy (SEM) and transmission electron microscopy (TEM). In addition, the weight, histology and biochemical changes in the rat epididymis before and after puberty were studied[201]. With the development of new technologies, especially transcriptomics[35,40,202,203], more molecular and genetic knowledge of the epididymis revealed. The role of the epididymis in the reproductive system has received increasing attention.
Moreover, regional differences in the epididymis have been found[204,205,206,207]. For instance, the caput epididymis is the most metabolically active region, and the secreted protein accounts for 70%-80% of the total protein secreted by the epididymal lumen[89]. The epididymis corpus plays a more important role in immune defense. In addition to its defensive function, the cauda epididymis is also very muscular. In general, in the history of epididymis research, our understanding of the epididymis has gradually deepened from the initial morphology and structure[60,208] to the regional division of the epididymis[209,210]. With the application of transcriptome technology in the epididymis, we realized that different regions have distinct gene compositions[205], which can be seen from microarray and single cell RNA analysis. This is the regional gene expression of the epididymis.
The male reproductive system shows great variability among different mammal species. In adult rats and mice epididymis, these segments are highly differentiated[211]. Due to the advantages of a short generation cycle and close biological evolution to human beings, rodents have become the most common animal model for epididymis research. Recently, it has been found that it is feasible to use dogs as biological models to study the molecular function of the human epididymis[212]. However, in terms of reproductive biology, it may be dangerous to infer other mammalian species that practice different reproductive strategies based on specific mammalian species. This is especially true for humans.
Cell classifications of the epididymal epithelium in each region of the epididymis are different[213]. It can be concluded from the foregoing that the main cell types, such as principal cells and basal cells, are expressed in all regions of the epididymis. The primary cell type of the entire tubule is the principal cells that constitute 80% of epithelial cells, which are responsible for most of the proteins secreted into the lumen. The blood–epididymis barrier formed by the tight connection of principal cells plays an immunoprotective role in the epididymal lumen. Morphological and physiological components of blood–epididymis barrier represent the basic aspects of male fertility. The disorder of epididymal epithelium impairs the integrity and function of blood–epididymis barrier and is related to the decline of human fertility. In addition, different cell types in the epididymis interact with androgen to promote sperm maturation. The principal cells of the epididymal caput absorb testosterone bound to androgen binding protein through a receptor-mediated process[214]. Different cell type compositions in different parts promote different functions. As a whole, the luminal environment formed in the epididymis is necessary for sperm maturation. Different cell compositions make each region have unique functions for sperm maturation. Gene ontology shows that caput transcripts are committed to cell‒cell adhesion according to their similar tissue qualifiers, while the corpus is characterized by genes participating in reactions to other organisms (defense mechanisms), and the cauda transcriptome is dedicated to the establishment of muscle contraction and localization. The cauda and corpus conduct early and late sperm maturation events, respectively, while the cauda is mainly used as the storage site of functional mature sperm[89]. These functions are well studied in rodents[215]and large animal models[216] but are less well characterized in humans.
Epithelial cells in various regions of the epididymis have unique transcriptomes and unique functions[89,215]. After further understanding, we found that the gene expression of the same cell type in different parts of the epididymis is also different. No relevant report has been found before the emergence of single-cell sequencing technology. Referring to the previous data, we briefly looked at the expression of several genes in the mouse epididymis and vas deferens (Figure 7, Figure 8). The analysis in this paper refers to the single-cell data of the mouse epididymis and vas deferens in the open database. Different epididymal cell types had different marker genes at different locations. RNase10, Cst11, Lcn2 and other genes are marker genes of epididymal caput principal cells. Differentiate the principal cells of the mice epididymis corpus according to Lcn5, RNase9, Plac8 and other marker genes. In addition, the principal cells in the cauda of the epididymis are also distinguished according to Gpx3, Klk1b27, Hint1, Gstm2 and other marker genes[56].The basal cells were identified according to the Itga6 and Krt14 genes[42]. Our analysis results showed that the same gene is expressed differently in the principal cells and basal cells. Lcn5 was highly expressed in caput principal cells (Figure 8B); however, it was highly expressed in caput and corpus basal cells (Figure 7B). Defb28 is highly expressed in cauda principal cells (Figure 8C) but highly expressed in all epididymis basal cells (Figure 7C). It has been reported that in epididymis caput principals, highly expressed genes include RNase10, Cst11, and Lcn2[42]. The results showed that the role of Rnase10 in the proximal epididymis was necessary to obtain sperm adhesion, which is a characteristic of mouse sperm and seems to be related to the mode of sperm transport in the female reproductive tract[124]. Combined with our own data analysis on Lcn5 in the principal cells (Figure 8B), we suspect that the caput of the epididymis is related to lipid metabolism and adhesion. Similarly, genes related to membrane transport and lipid metabolism are also highly expressed in basal cells[42]. This shows that different cells also have similarities, which together contribute to the formation of an epididymal cavity environment suitable for sperm maturation. The complex gene expression and cell composition of the caput, corpus and cauda of the epididymis make each region have unique functions for sperm maturation. Some coregulatory factors showed fragment enrichment in their expression, indicating that these factors may contribute to fragment-specific responses[65]. Each epididymal fragment has its own transcriptomic signature, regulating luminal composition and thus dominating mature male sperm.
10. Conclusions
The coordination of gene expression in different cell types promotes sperm maturation. The more we know about the gene expression in the epididymal region, the more likely it is to use transgenic technology to change epididymal traits at the gene level and improve fertility and sperm quality. It is warmly expected that one day can extract the molecules and proteins in epididymal fluid that play a key role in regulating spermatogenesis and apply them to fresh sperm and frozen sperm to improve semen quality and pregnancy rate during estrus.
Author Contributions
The first author, Meng-Meng Liu, had the original idea of writing a review article on the work and wrote the first draft. Corresponding authors, Guo-Liang Zhang, read and reviewed the first and subsequent drafts of the article and assisted the first author to design the review work. Corresponding authors also revised and put together figures and tables on the various revised versions of the review and checked out the correct citations of all 190 references. Authors Hao Shang, Xin-Lei Feng, Chao Qi, Shu-Er Zhang, Yuan-Chao Sun, Yu-Jiang Sun substantially revised and corrected several versions of the manuscript and checked that reference numbers and references list was correctly written in the various version. The author(s) read and approved the final manuscript.
Funding
This work was supported by National Natural Science Foundation of China (31902158), Porcine Industry Technology System of Shandong Province (SDAIT-08-16), Key Technology R&D Program of Shandong Province (2021LZGC001), Research Foundation for Advanced Talents of Qingdao Agricultural University (665/1119013) and National Training Programs of Innovation and Entrepreneurship for Undergraduates (202210435036).
Data Availability Statement
The datasets analysed during the current study are available in the GEO, Accession#GSE145443,https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE145443. The DOI of the data source article is 10.7554/eLife.55474. Operating system: x86_64, mingw32, Programming language: R, R version 4.1.2 (2021-11-01).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Hox | homeobox |
| OCLN | Occludin |
| PGE2 | ProstaglandinE2 |
| LCN | lipocalin |
| DEFB | defensin, beta |
| DICER1 | dicer1, ribonuclease III |
| mERABP | mouse epididymal retinoic acid binding protein |
| mEP17 | mouse epididymal protein of 17 kDa |
| CD52 | CD52 molecule |
| CRISP | cysteine rich secretory protein |
| AQPs | aquaporins |
| SPINK | serine peptidase inhibitor Kazal type |
| t-SNE | t-distributed stochastic neighbor embedding |
| DEGs | differentially expressed genes |
| IVF | in vitro fertilization |
| AI | artificial insemination |
| LM | light microscopy |
| SEM | scanning electron microscopy |
| TEM | transmission electron microscopy |
References
- Cosentino, M.J.; Cockett, A.T.K. Review article: Structure and function of the epididymis. Urol. Res. 1986, 14, 229–240. [Google Scholar] [CrossRef] [PubMed]
- LANZ, V. and Neuhaeuser, G. Morphometric analysis of the human epididymis. Zeitschrift fur Anatomie und Entwicklungsgeschichte 1964, 124, 126–152. [Google Scholar] [CrossRef]
- Turner, T.T.; Gleavy, J.L.; Harris, J.M. Fluid Movement in the Lumen of the Rat Epididymis: Effect of Vasectomy and Subsequent Vasovasostomy. J. Androl. 1990, 11, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.X.; Temple-Smith, P.; Wreford, N.G. Postnatal differentiation and development of the rat epididymis: A stereological study. Anat. Rec. 1994, 238, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Stoltenberg, M.; Therkildsen, P.; Andreasen, A.; Jensen, K.B.; Juhl, S.; Ernst, E.; Danscher, G. Computer-assisted visualization of the rat epididymis: a methodological study based on paraffin sections autometallographically stained for zinc ions. Histochem. J. 1998, 30, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.; Renfree, M.B. Wolffian Duct Development. Sex. Dev. 2014, 8, 273–280. [Google Scholar] [CrossRef]
- Branford, W.W. , Benson, G.V., Ma, L., Maas, R.L. and Potter, S.S. Characterization of Hoxa-10/Hoxa-11 transheterozygotes reveals functional redundancy and regulatory interactions. Developmental Biology 2000, 224, 373–387. [Google Scholar] [CrossRef]
- Snyder, E.M.; Small, C.L.; Bomgardner, D.; Xu, B.; Evanoff, R.; Griswold, M.D.; Hinton, B.T. Gene expression in the efferent ducts, epididymis, and vas deferens during embryonic development of the mouse. Dev. Dyn. 2010, 239, 2479–2491. [Google Scholar] [CrossRef]
- Westfalewicz, B.; Dietrich, M.; Mostek, A.; Partyka, A.; Bielas, W.; Niżański, W.; Ciereszko, A. Identification and functional analysis of bull (Bos taurus) cauda epididymal fluid proteome. J. Dairy Sci. 2017, 100, 6707–6719. [Google Scholar] [CrossRef]
- Rowley, M.J.; Teshima, F.; Heller, C.G. Duration of Transit of Spermatozoa through the Human Male Ductular System. Fertil. Steril. 1970, 21, 390–396. [Google Scholar] [CrossRef]
- Acott, T.S.; Carr, D.W. Inhibition of Bovine Spermatozoa by Caudal Epididymal Fluid: II. Interaction of pH and a Quiescence Factor 1. Biol. Reprod. 1984, 30, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Discutient Application to the Indurated Epididymis. Atlanta Med Surg J, 1859. 4(8): p. 515.
- Bryant, T. Case of Torsion of the Spermatic Cord, with Strangulation of the Epididymis and Testicle in an Incompletely Descended Organ. J. R. Soc. Med. -75. [CrossRef]
- Rockwell, F.W. Case of Removal of Both Testicles for Recurrent Carcinoma of the Epididymis. Annals of Surgery 1888, 8, 446. [Google Scholar] [CrossRef] [PubMed]
- Some Diseases of the Male Genital System: VIII. Tuberculous Disease of the Testis and Epididymis. Hospital (Lond 1886), 1908. 43(1120): p. 526.
- Griffiths, J. Observations on the Appendix of the Testicle, and on the Cysts of the Epididymis, the Vasa Efferentia, and the Rete Testis. . 1893, 28, 107–116. [Google Scholar] [PubMed]
- Watson, J.H. Some Observations on the Origin and Nature of the so-called Hydatids of Morgagni found in Men and Women, with Especial Reference to the Fate of the Müllerian Duct in the Epididymis. . 1902, 36, 147–61. [Google Scholar] [PubMed]
- Goglia, G. [Histochemical research on the ductus epididymis of the horse]. . 1954, 30, 1151–4. [Google Scholar] [PubMed]
- Goglia, G.; Magli, G. [Histochemical research on secretory processes of coni vasculosi and epididymis canal in the guinea pig]. . 1957, 33, 418–21. [Google Scholar]
- Montagna, W. SOME CYTOCHEMICAL OBSERVATIONS ON HUMAN TESTES AND EPIDIDYMIDES. Ann. New York Acad. Sci. 1952, 55, 629–642. [Google Scholar] [CrossRef]
- Allen, J.M.; Slater, J.J. A CYTOCHEMICAL STUDY OF GOLGI ASSOCIATED THIAMINE PYROPHOSPHATASE IN THE EPIDIDYMIS OF THE MOUSE. J. Histochem. Cytochem. 1961, 9, 418–423. [Google Scholar] [CrossRef]
- Allen, J.M. THE HISTOCHEMISTRY OF GLUCOSE-6-PHOSPHATASE IN THE EPIDIDYMIS OF THE MOUSE. J. Histochem. Cytochem. 1961, 9, 681–689. [Google Scholar] [CrossRef]
- Birnbaum, D.; Hall, T.; Lee, R. The Zinc Content of Rat Sperm Cells from Ejaculate, Vas, Epididymis and Testis..CONFERENCE NAME, LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; pp. 321–324.
- Gray, C.P. , Biorn, C. L. and Drinker, H.R. Tumors of the epididymis. The Journal of Urology 1961, 86, 620–624. [Google Scholar]
- Bresler, V.M. [Tumors of the interstitial tissue of the epididymis in mice induced by diethylstibesterol]. . 1960, 28–34. [Google Scholar]
- Bresler, V.M. Tumors of the interstitial tissue of the epididymis in mice induced by diethylstibesterol. Voprosy onkologii 1960, 6, 28–34. [Google Scholar]
- Naide, Y. , [Epididymitis]. Ryoikibetsu Shokogun Shirizu, 1999(25 Pt 3): p. 237-40.
- Jonté, G.; Holstein, A. On the Morphology of the Transitional Zones from the Rete Testis into the Ductuli Efferentes and from the Ductuli Efferentes into the Ductus Epididymidis. Investigations on the Human Testis and Epididymis. Andrologia 2009, 19, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Fouchécourt, S.; Métayer, S.; Locatelli, A.; Dacheux, F.; Dacheux, J.-L. Stallion Epididymal Fluid Proteome: Qualitative and Quantitative Characterization; Secretion and Dynamic Changes of Major Proteins1. Biol. Reprod. 2000, 62, 1790–1803. [Google Scholar] [CrossRef] [PubMed]
- Jury, J.A. , Perry, A.C. and Hall, L. Identification, sequence analysis and expression of transcripts encoding a putative metalloproteinase, eMDC II, in human and macaque epididymis. MHR: Basic science of reproductive medicine 1999, 5, 1127–1134. [Google Scholar] [PubMed]
- Salisbury, G.W.; Graves, C.N.; Nakabayashi, N.T.; Cragle, R.G. OBSERVATIONS ON THE AEROBIC METABOLISM OF BULL AND GOAT EPIDIDYMAL SPERMATOZOA. Reproduction 1963, 6, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, G.W.; Graves, C.N. SUBSTRATE-FREE EPIDIDYMAL-LIKE BOVINE SPERMATOZOA. Reproduction 1963, 6, 351–359. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, Y.; Sun, C.; Ge, S.; Tan, Y.; Shen, H.; Yang, P. A Multi-Omics Study of Human Testis and Epididymis. Molecules 2021, 26, 3345. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Belleannée, C.; Jones, R.; Labas, V.; Belghazi, M.; Guyonnet, B.; Druart, X.; Gatti, J.L.; Dacheux, F. Mammalian epididymal proteome. Mol. Cell. Endocrinol. 2009, 306, 45–50. [Google Scholar] [CrossRef]
- Wong, J.; Damdimopoulos, A.; Damdimopoulou, P.; Gasperoni, J.G.; Tran, S.C.; Grommen, S.V.; De Groef, B.; Dworkin, S. Transcriptome analysis of the epididymis from Plag1 deficient mice suggests dysregulation of sperm maturation and extracellular matrix genes. Dev. Dyn. 2020, 249, 1500–1513. [Google Scholar] [CrossRef]
- Lye, R.; Hinton, B.T. Technologies for the study of epididymal-specific genes. Mol. Cell. Endocrinol. 2004, 216, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Légaré, C.; Sullivan, R. Differential gene expression profiles of human efferent ducts and proximal epididymis. Andrology 2019, 8, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Domeniconi, R.F. , Souza, A.C.F., Xu, B., Washington, A.M. and Hinton, B.T. Is the epididymis a series of organs placed side by side? Biology of reproduction 2016, 95, 10–1. [Google Scholar] [CrossRef] [PubMed]
- Eddy, E.M. , Washburn, T.F., Bunch, D.O., Goulding, E.H., Gladen, B.C., Lubahn, D.B. and Korach, K.S. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef]
- Wu, C.; Wang, C.; Zhai, B.; Zhao, Y.; Zhao, Z.; Yuan, Z.; Fu, X.; Zhang, M. Study on the region-specific expression of epididymis mRNA in the rams. PLOS ONE 2021, 16, e0245933. [Google Scholar] [CrossRef]
- Jervis, K.M.; Robaire, B. Effects of caloric restriction on gene expression along the epididymis of the Brown Norway rat during aging. Exp. Gerontol. 2003, 38, 549–560. [Google Scholar] [CrossRef]
- Rinaldi, V.D. , Donnard, E., Gellatly, K., Rasmussen, M., Kucukural, A., Yukselen, O., Garber, M., Sharma, U. and Rando, O.J. An atlas of cell types in the mouse epididymis and vas deferens. elife 2020, 9, e55474. [Google Scholar] [CrossRef]
- Dharmat, R. , Kim, S., Li, Y. and Chen, R. Single-cell capture, RNA-seq, and transcriptome analysis from the neural retina. Retinal Development: Methods and Protocols 2020, 2092, 159–186. [Google Scholar]
- Lang, X. , Adjei, M., Wang, C., Chen, X., Li, C., Wang, P., Pan, M., Li, K., Shahzad, K. and Zhao, W. RNA-Seq reveals the functional specificity of epididymal caput, corpus, and cauda genes of cattleyak. Animal Science Journal 2022, 93, e13732. [Google Scholar] [CrossRef]
- Ikawa, M. , Nakanishi, T., Yamada, S., Wada, I., Kominami, K., Tanaka, H., Nozaki, M., Nishimune, Y. and Okabe, M. Calmegin is required for fertilin α/β heterodimerization and sperm fertility. Developmental biology 2001, 240, 254–261. [Google Scholar] [CrossRef]
- Nishimura, H.; Kim, E.; Nakanishi, T.; Baba, T. Possible Function of the ADAM1a/ADAM2 Fertilin Complex in the Appearance of ADAM3 on the Sperm Surface. J. Biol. Chem. 2004, 279, 34957–34962. [Google Scholar] [CrossRef] [PubMed]
- Cho, C. , O'Dell Bunch, D., Faure, J.E., Goulding, E.H., Eddy, E.M., Primakoff, P. and Myles, D.G.,. Fertilization defects in sperm from mice lacking fertilin β. Science 1998, 281, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Klinefelter, G.R.; Amann, R.P.; Hammerstedt, R.H. Culture of Principal Cells From the Rat Caput Epididymidis1. Biol. Reprod. 1982, 26, 885–901. [Google Scholar] [CrossRef]
- Lee, G.Y.; A Kenny, P.; Lee, E.H.; Bissell, M.J. Three-dimensional culture models of normal and malignant breast epithelial cells. Nat. Methods 2007, 4, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Bunick, D.; Bahr, J.; Klinefelter, G.; Hess, R. Isolation and culture of epithelial cells from rat ductuli efferentes and initial segment epididymidis. Tissue Cell 1998, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, J.; St-Pierre, N.; Viger, R.S.; Hermo, L.; Cyr, D.G. Characterization of a Novel Rat Epididymal Cell Line to Study Epididymal Function. Endocrinology 2005, 146, 4710–4720. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Suzuki, K.; Matusik, R.J.; Obinata, M.; Orgebin-Crist, M.-C. Immortalized epididymal cell lines from transgenic mice overexpressing temperature-sensitive simian virus 40 large T-antigen gene. . 2002, 23, 854–69. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, J.; Ong, I.M.; Stanic, A.K. Single-cell technologies in reproductive immunology. Am. J. Reprod. Immunol. 2019, 82, e13157–e13157. [Google Scholar] [CrossRef]
- Shi, J.; Fok, K.L.; Dai, P.; Qiao, F.; Zhang, M.; Liu, H.; Sang, M.; Ye, M.; Liu, Y.; Zhou, Y.; et al. Spatio-temporal landscape of mouse epididymal cells and specific mitochondria-rich segments defined by large-scale single-cell RNA-seq. Cell Discov. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef]
- Johnston, D.S.; Jelinsky, S.A.; Bang, H.J.; DiCandeloro, P.; Wilson, E.; Kopf, G.S.; Turner, T.T. The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in the Epididymis1. Biol. Reprod. 2005, 73, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef] [PubMed]
- Jervis, K.M.; Robaire, B. Dynamic Changes in Gene Expression along the Rat Epididymis1. Biol. Reprod. 2001, 65, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, T.R.; Griswold, M.D. Androgen-Regulated Genes in the Murine Epididymis1. Biol. Reprod. 2004, 71, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhai, X.; Ma, Y.; Zhang, T.; Wang, Z.; Chong, T. Anatomic characteristics of epididymis based on histology, proteomic, and 3D reconstruction. Andrology 2020, 8, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, J.; Belghazi, M.; Lanson, Y.; Dacheux, F. Human epididymal secretome and proteome. Mol. Cell. Endocrinol. 2006, 250, 36–42. [Google Scholar] [CrossRef] [PubMed]
- I Adamali, H.; Somani, I.H.; Huang, J.Q.; Mahuran, D.; A Gravel, R.; Trasler, J.M.; Hermo, L. I. Abnormalities in cells of the testis, efferent ducts, and epididymis in juvenile and adult mice with beta-hexosaminidase A and B deficiency. . 1999, 20, 779–802. [Google Scholar]
- Jelinsky, S.A.; Turner, T.T.; Bang, H.J.; Finger, J.N.; Solarz, M.K.; Wilson, E.; Brown, E.L.; Kopf, G.S.; Johnston, D.S. The Rat Epididymal Transcriptome: Comparison of Segmental Gene Expression in the Rat and Mouse Epididymides1. Biol. Reprod. 2007, 76, 561–570. [Google Scholar] [CrossRef]
- Harris, A.; Browne, J.; Leir, S.-H.; Eggener, S. Region-specific microRNA signatures in the human epididymis. Asian J. Androl. 2018, 20, 539. [Google Scholar] [CrossRef]
- Browne, J.A.; Yang, R.; Leir, S.-H.; Eggener, S.E.; Harris, A. Expression profiles of human epididymis epithelial cells reveal the functional diversity of caput, corpus and cauda regions. Mol. Hum. Reprod. 2015, 22, 69–82. [Google Scholar] [CrossRef]
- Sipilä, P.; Pujianto, D.A.; Shariatmadari, R.; Nikkilä, J.; Lehtoranta, M.; Huhtaniemi, I.T.; Poutanen, M. Differential Endocrine Regulation of Genes Enriched in Initial Segment and Distal Caput of the Mouse Epididymis as Revealed by Genome-Wide Expression Profiling1. Biol. Reprod. 2006, 75, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh, M.; Robaire, B. Identification of Early Response Genes and Pathway Activated by Androgens in the Initial Segment and Caput Regions of the Regressed Rat Epididymis. Endocrinology 2010, 151, 4504–4514. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.P.B.; Lopes, T.N.; da Silva, A.F.T.; Santi, L.; Beys-Da-Silva, W.O.; Yates, J.R.; Bustamante-Filho, I.C. Changes in porcine cauda epididymal fluid proteome by disrupting the HPT axis: Unveiling potential mechanisms of male infertility. Mol. Reprod. Dev. 2020, 87, 952–965. [Google Scholar] [CrossRef] [PubMed]
- Kunkitti, P.; Bergqvist, A.-S.; Sjunnesson, Y.; Axnér, E. The ability of feline spermatozoa in different epididymal regions to undergo capacitation and acrosome reaction. Anim. Reprod. Sci. 2015, 161, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Kunkitti, P.; Axnér, E.; Bergqvist, A.-S.; Sjunnesson, Y. In vitro fertilization using frozen-thawed feline epididymal spermatozoa from corpus and cauda regions. Theriogenology 2016, 86, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Kunkitti, P.; Bergqvist, A.-S.; Sjunnesson, Y.; Johannisson, A.; Axnér, E. The tolerance of feline corpus and cauda spermatozoa to cryostress. Theriogenology 2015, 85, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Esponda, P.; Bedford, J.M. The influence of body temperature and castration on the protein composition of fluid in the rat cauda epididymidis. Reproduction 1986, 78, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Regalado, F.; Esponda, P.; Nieto, A. Temperature and androgens regulate the biosynthesis of secretory proteins from rabbit cauda epididymidis. Mol. Reprod. Dev. 1993, 36, 448–453. [Google Scholar] [CrossRef]
- Wong, P.Y.D.; Au, C.L.; Bedford, J.M. Biology of the Scrotum. II. Suppression by Abdominal Temperature of Transepithelial Ion and Water Transport in the Cauda Epididymidis. Biol. Reprod. 1982, 26, 683–689. [Google Scholar] [CrossRef]
- Foldesy, R.G.; Bedford, J.M. Biology of the Scrotum. I. Temperature and Androgen as Determinants of the Sperm Storage Capacity of the Rat Cauda Epididymidis. Biol. Reprod. 1982, 26, 673–682. [Google Scholar] [CrossRef]
- Bedford, J.M. Human Spermatozoa and Temperature: The Elephant in the Room. Biol. Reprod. 2015, 93, 97. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, S. Are the basal cells of the mammalian epididymis still an enigma? Reproduction, Fertility and Development 2014, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.L.; Flickinger, C.J. Development of cell types and of regional differences in the postnatal rat epididymis. Am. J. Anat. 1979, 154, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, J.; Gregory, M.; Pinel, L.; Cyr, D.G. Differential gene expression and hallmarks of stemness in epithelial cells of the developing rat epididymis. Cell Tissue Res. 2022, 389, 327–349. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.S. Isolation and Cell Culture of the Epithelial Cells of Cauda Epididymidis of the Bull1. Biol. Reprod. 1985, 33, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Bongso, A. and Trounson, A. Andrology: Evaluation of motility, fertilizing ability and embryonic development of murine epididymal sperm after coculture with epididymal epithelium. Human reproduction 1996, 11, 1451–1456. [Google Scholar] [CrossRef]
- Moore, H.D.M.; Hartman, T.D. In-vitro development of the fertilizing ability of hamster epididymal spermatozoa after co-culture with epithelium from the proximal cauda epididymidis. Reproduction 1986, 78, 347–352. [Google Scholar] [CrossRef]
- Orgebin-Crist, M.C. , Jonas-Davies, J., Storey, P. and Olson, G.E. Effect of d-valine and cytosine arabinoside on [3 H] thymidine incorporation in rat and rabbit epididymal epithelial cell cultures. In vitro 1984, 20, 45–52. [Google Scholar] [CrossRef]
- Castellón, E.; Huidobro, C. Androgen regulation of glycosidase secretion in epithelial cell cultures from human epididymis. Hum. Reprod. 1999, 14, 1522–1527. [Google Scholar] [CrossRef]
- Higgins, S.J. , Young, P. and Cunha, G.R. Induction of functional cytodifferentiation in the epithelium of tissue recombinants: II. Instructive induction of Wolffian duct epithelia by neonatal seminal vesicle mesenchyme. Development 1989, 106, 235–250. [Google Scholar] [CrossRef]
- De Gendt, K.; Verhoeven, G. Tissue- and cell-specific functions of the androgen receptor revealed through conditional knockout models in mice. Mol. Cell. Endocrinol. 2012, 352, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Hinton, B.T.; Palladino, M.A. Epididymal epithelium: Its contribution to the formation of a luminal fluid microenvironment. Microsc. Res. Tech. 1995, 30, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.-L.; Li, S.; Huang, J.-H.; Yang, D.-L.; Zhang, G.; Chen, S.-L.; Ruan, Y.-C.; Ye, K.-N.; Cheng, C.H.K.; Zhou, W.-L. Sodium Coupled Bicarbonate Influx Regulates Intracellular and Apical pH in Cultured Rat Caput Epididymal Epithelium. PLOS ONE 2011, 6, e22283. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, G.A. New insights into epididymal biology and function. Hum. Reprod. Updat. 2008, 15, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Shum, W.W.C.; Da Silva, N.; Brown, D.; Breton, S. Regulation of luminal acidification in the male reproductive tract via cell–cell crosstalk. J. Exp. Biol. 2009, 212, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Leir, S.-H.; Yin, S.; Kerschner, J.L.; Cosme, W.; Harris, A. An atlas of human proximal epididymis reveals cell-specific functions and distinct roles for CFTR. Life Sci. Alliance 2020, 3, e202000744. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A. , Shur, B.D. and Hess, R.A. Estrogen, efferent ductules, and the epididymis. Biology of reproduction 2011, 84, 207–217. [Google Scholar] [CrossRef]
- Hermo, L. , Oka, R. and Morales, C.R. Secretion and endocytosis in the male reproductive tract: a role in sperm maturation. International review of cytology 1994, 154, 105–189. [Google Scholar]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef]
- Cyr, D.G.; Robaire, B.; Hermo, L. Structure and turnover of junctional complexes between principal cells of the rat epididymis. Microsc. Res. Tech. 1995, 30, 54–66. [Google Scholar] [CrossRef]
- Cyr, D.G.; Gregory, M.; Dubé. ; Dufresne, J.; Chan, P.T.K.; Hermo, L. Orchestration of occludins, claudins, catenins and cadherins as players involved in maintenance of the blood-epididymal barrier in animals and humans. Asian J. Androl. 2007, 9, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Y.; Zhang, B.L.; Gao, D.Y.; Li, Q.; Xu, X.Y.; Shum, W. Epididymal epithelial degeneration and lipid metabolism impairment account for male infertility in occludin knockout mice. Front. Endocrinol. 2022, 13, 1069319. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, S.; Romanello, M.G.; Domeneghini, C. Morphological examination of epididymal epithelium in the mule (E. hinnus) in comparison with parental species (E. asinus and E. caballus).. 1991, 6, 325–37. [Google Scholar]
- Schön, J.; Blottner, S. Seasonal variations in the epididymis of the roe deer (Capreolus capreolus). Anim. Reprod. Sci. 2009, 111, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.H.; Nashan, D.; Sorg, C.; Oberpenning, F.; Schulze, H.; Nieschlag, E.; Cooper, T.G. Basal Cells of the Human Epididymis—Antigenic and Ultrastructural Similarities to Tissue-Fixed Macrophages1. Biol. Reprod. 1994, 50, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Seiler, P.; Wenzel, I.; Wagenfeld, A.; Yeung, C.H.; Nieschlag, E.; Cooper, T.G. The appearance of basal cells in the developing murine epididymis and their temporal expression of macrophage antigens. Int. J. Androl. 1998, 21, 217–226. [Google Scholar] [CrossRef]
- Shum, W.W.C.; Da Silva, N.; McKee, M.; Smith, P.J.; Brown, D.; Breton, S. Transepithelial Projections from Basal Cells Are Luminal Sensors in Pseudostratified Epithelia. Cell 2008, 135, 1108–1117. [Google Scholar] [CrossRef]
- Mou, H.; Vinarsky, V.; Tata, P.R.; Brazauskas, K.; Choi, S.; Crooke, A.K.; Zhang, B.; Solomon, G.M.; Turner, B.; Bihler, H.; et al. Dual SMAD Signaling Inhibition Enables Long-Term Expansion of Diverse Epithelial Basal Cells. Cell Stem Cell 2016, 19, 217–231. [Google Scholar] [CrossRef]
- Mandon, M.; Hermo, L.; Cyr, D.G. Isolated Rat Epididymal Basal Cells Share Common Properties with Adult Stem Cells1. Biol. Reprod. 2015, 93, 115. [Google Scholar] [CrossRef]
- Cyr, D.G.; Pinel, L. Emerging organoid models to study the epididymis in male reproductive toxicology. Reprod. Toxicol. 2022, 112, 88–99. [Google Scholar] [CrossRef]
- Yalcin-Ozuysal. ; Fiche, M.; Guitierrez, M.; Wagner, K.-U.; Raffoul, W.; Brisken, C. Antagonistic roles of Notch and p63 in controlling mammary epithelial cell fates. Cell Death Differ. 2010, 17, 1600–1612. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Acebron, S.P.; Herbst, J.; Hatiboglu, G.; Niehrs, C. Post-transcriptional Wnt Signaling Governs Epididymal Sperm Maturation. Cell 2015, 163, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Boyer, A.; Yeh, J.R.; Zhang, X.; Paquet, M.; Gaudin, A.; Nagano, M.C.; Boerboom, D. CTNNB1 Signaling in Sertoli Cells Downregulates Spermatogonial Stem Cell Activity via WNT4. PLOS ONE 2012, 7, e29764. [Google Scholar] [CrossRef] [PubMed]
- Girardet, L.; Cyr, D.G.; Belleannée, C. Arl13b controls basal cell stemness properties and Hedgehog signaling in the mouse epididymis. Cell. Mol. Life Sci. 2022, 79, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Zhang, L.; Yang, L.; Yang, P.; Ullah, S.; Zhang, Q.; Chen, Q. Ultrastructure of epididymal epithelium and its interaction with the sperm in the soft-shelled turtle Pelodiscus sinensis. Micron 2013, 54-55, 65–74. [Google Scholar] [CrossRef]
- Robaire, B.; Viger, R.S. Regulation of Epididymal Epithelial Cell Functions1. Biol. Reprod. 1995, 52, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Douglass, J.A.M.E.S. , Garrett, S.H. and Garrett, J.E. Differential patterns of regulated gene expression in the adult rat epididymis. Annals of the New York Academy of Sciences 1991, 637, 384–398. [Google Scholar] [CrossRef]
- Hermo, L.; Dworkin, J.; Oko, R. Role of epithelial clear cells of the rat epididymis in the disposal of the contents of cytoplasmic droplets detached from spermatozoa. Am. J. Anat. 1988, 183, 107–124. [Google Scholar] [CrossRef]
- Hermo, L.; Oko, R.; Robaire, B. Epithelial cells of the epididymis show regional variations with respect to the secretion or endocytosis of immobilin as revealed by light and electron microscope immunocytochemistry. Anat. Rec. 1992, 232, 202–220. [Google Scholar] [CrossRef]
- Hamilton, D.W.; Olson, G.E.; Cooper, T.G. Regional variation in the surface morphology of the epithelium of the rat ductuli efferentes, ductus epididymidis and vas deferens. Anat. Rec. 1977, 188, 13–27. [Google Scholar] [CrossRef]
- Moore, H.D.M.; Bedford, J.M. Short-term effects of androgen withdrawal on the structure of different epithelial cells in the rat epididymis. Anat. Rec. 1979, 193, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Belleannée, C.; Da Silva, N.; Shum, W.W.; Brown, D.; Breton, S.; Roy, J.W.; Hill, E.; Ruan, Y.C.; Vedovelli, L.; Păunescu, T.G.; et al. Role of purinergic signaling pathways in V-ATPase recruitment to apical membrane of acidifying epididymal clear cells. Am. J. Physiol. Physiol. 2010, 298, C817–C830. [Google Scholar] [CrossRef]
- Kujala, M.; Hihnala, S.; Tienari, J.; Kaunisto, K.; Hästbacka, J.; Holmberg, C.; Kere, J.; Höglund, P. Expression of ion transport-associated proteins in human efferent and epididymal ducts. Reproduction 2007, 133, 775–784. [Google Scholar] [CrossRef]
- Da Silva, N.; Shum, W.W.C.; Breton, S. Regulation of vacuolar proton pumping ATPase-dependent luminal acidification in the epididymis. Asian J. Androl. 2007, 9, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.-H.; Leung, G.P.; Leung, M.C.; Shum, W.W.; Zhou, W.-L.; Wong, P.Y. Cell–cell Interaction Underlies Formation of Fluid in the Male Reproductive Tract of the Rat. J. Gen. Physiol. 2005, 125, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R. , Legare, C., Thabet, M. and Thimon, V. Gene expression in the epididymis of normal and vasectomized men: what can we learn about human sperm maturation? Journal of andrology 2011, 32, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.S.; Turner, T.T.; Finger, J.N.; Owtscharuk, T.L.; Kopf, G.S.; Jelinsky, S.A. Identification of epididymis-specific transcripts in the mouse and rat by transcriptional profiling. Asian J. Androl. 2007, 9, 522–527. [Google Scholar] [CrossRef]
- Xie, S.; Zhu, Y.; Ma, L.; Lu, Y.; Zhou, J.; Gui, Y.; Cao, L. Genome-wide profiling of gene expression in the epididymis of alpha-chlorohydrin-induced infertile rats using an oligonucleotide microarray. Reprod. Biol. Endocrinol. 2010, 8, 37–37. [Google Scholar] [CrossRef]
- Krutskikh, A.; Poliandri, A.; Dacheux, J.L.; Poutanen, M.; Huhtaniemi, I.; Cabrera-Sharp, V. Epididymal protein Rnase10 is required for post-testicular sperm maturation and male fertility. FASEB J. 2012, 26, 4198–4209. [Google Scholar] [CrossRef]
- Devor, E.J.; Moffat-Wilson, K.A.; Galbraith, J.J. LOC 390443 (RNase 9) on Chromosome 14q11.2 Is Related to the RNase A Superfamily and Contains a Unique Amino-Terminal Preproteinlike Sequence. Hum. Biol. 2004, 76, 921–935. [Google Scholar] [CrossRef]
- Castella, S.; Fouchécourt, S.; Teixeira-Gomes, A.P.; Vinh, J.; Belghazi, M.; Dacheux, F.; Dacheux, J.-L. Identification of a Member of a New RNase A Family Specifically Secreted by Epididymal Caput Epithelium1. Biol. Reprod. 2004, 70, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.A. , Cai, Y., Sang, Y., Blecha, F. and Zhang, G. Cross-species analysis of the mammalian β-defensin gene family: presence of syntenic gene clusters and preferential expression in the male reproductive tract. Physiological genomics 2005, 23, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Semple, C.A. , Gautier, P., Taylor, K. and Dorin, J.R. The changing of the guard: Molecular diversity and rapid evolution of β-defensins. Molecular diversity 2006, 10, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Semple, C.A. , Gautier, P., Taylor, K. and Dorin, J.R. The changing of the guard: Molecular diversity and rapid evolution of β-defensins. Molecular diversity 2006, 10, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Pazgier, M. , Hoover, D.M., Yang, D., Lu, W. and Lubkowski, J. Human β-defensins. Cellular and Molecular Life Sciences CMLS 2006, 63, 1294–1313. [Google Scholar] [CrossRef] [PubMed]
- Schutte, B.C. , Mitros, J.P., Bartlett, J.A., Walters, J.D., Jia, H.P., Welsh, M.J., Casavant, T.L. and McCray Jr, P.B. Discovery of five conserved β-defensin gene clusters using a computational search strategy. Proceedings of the National Academy of Sciences 2002, 99, 2129–2133. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Ouchi, Y. Antimicrobial peptide defensin: Identification of novel isoforms and the characterization of their physiological roles and their significance in the pathogenesis of diseases. Proc. Jpn. Acad. Ser. B 2012, 88, 152–166. [Google Scholar] [CrossRef]
- Ribeiro, C.M. , Silva, E.J., Hinton, B.T. and Avellar, M.C.W. β-defensins and the epididymis: contrasting influences of prenatal, postnatal, and adult scenarios. Asian journal of andrology 2016, 18, 323. [Google Scholar]
- Hu, S.-G.; Zou, M.; Yao, G.-X.; Ma, W.-B.; Zhu, Q.-L.; Li, X.-Q.; Chen, Z.-J.; Sun, Y. Androgenic regulation of beta-defensins in the mouse epididymis. Reprod. Biol. Endocrinol. 2014, 12, 76–76. [Google Scholar] [CrossRef]
- Jalkanen, J.; Huhtaniemi, I.; Poutanen, M. Discovery and characterization of new epididymis-specific beta-defensins in mice. Biochim. et Biophys. Acta (BBA) - Gene Struct. Expr. 2005, 1730, 22–30. [Google Scholar] [CrossRef]
- Pujianto, D.A.; Muliawati, D.; Rizki, M.D.; Parisudha, A.; Hardiyanto, L. Mouse defensin beta 20 (Defb20) is expressed specifically in the caput region of the epididymis and regulated by androgen and testicular factors. Reprod. Biol. 2020, 20, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Ni, M.; Xie, S.; Zhang, Y.; Zhang, C.; Ni, Z.; Chu, C.; Wu, L.; Zhou, Y.; Zhang, Y. DICER1 regulates antibacterial function of epididymis by modulating transcription of β-defensins. J. Mol. Cell Biol. 2018, 11, 408–420. [Google Scholar] [CrossRef]
- Radhakrishnan, Y. , Hamil, K.G., Yenugu, S., Young, S.L., French, F.S. and Hall, S.H. Identification, characterization, and evolution of a primate β-defensin gene cluster. Genes & Immunity 2005, 6, 203–210. [Google Scholar]
- Rodríguez-Jiménez, F.J. , Krause, A., Schulz, S., Forssmann, W.G., Conejo-Garcia, J.R., Schreeb, R. and Motzkus, D. Distribution of new human β-defensin genes clustered on chromosome 20 in functionally different segments of epididymis. Genomics 2003, 81, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. , Diao, H., Ni, Z., Hu, S., Yu, H. and Zhang, Y. The epididymis-specific antimicrobial peptide β-defensin 15 is required for sperm motility and male fertility in the rat (Rattus norvegicus). Cellular and Molecular Life Sciences 2011, 68, 697–708. [Google Scholar] [CrossRef]
- Björkgren, I.; Alvarez, L.; Blank, N.; Balbach, M.; Turunen, H.; Laajala, T.D.; Toivanen, J.; Krutskikh, A.; Wahlberg, N.; Huhtaniemi, I.; et al. Targeted inactivation of the mouse epididymal beta-defensin 41 alters sperm flagellar beat pattern and zona pellucida binding. Mol. Cell. Endocrinol. 2016, 427, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Dubé, E.; Hermo, L.; Chan, P.T.; Cyr, D.G. Alterations in Gene Expression in the Caput Epididymides of Nonobstructive Azoospermic Men1. Biol. Reprod. 2008, 78, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W.; et al. A Common Mutation in the Defensin DEFB126 Causes Impaired Sperm Function and Subfertility. Sci. Transl. Med. 2011, 3, 92ra65–92ra65. [Google Scholar] [CrossRef]
- Aram, R.; Chan, P.T.K.; Cyr, D.G. Beta-defensin126 is correlated with sperm motility in fertile and infertile men†. Biol. Reprod. 2019, 102, 92–101. [Google Scholar] [CrossRef]
- Tollner, T.L.; Yudin, A.I.; Treece, C.A.; Overstreet, J.W.; Cherr, G.N. Macaque sperm coating protein DEFB126 facilitates sperm penetration of cervical mucus. Hum. Reprod. 2008, 23, 2523–2534. [Google Scholar] [CrossRef]
- Charkoftaki, G. , Wang, Y., McAndrews, M., Bruford, E.A., Thompson, D.C., Vasiliou, V. and Nebert, D.W. Update on the human and mouse lipocalin (LCN) gene family, including evidence the mouse Mup cluster is result of an “evolutionary bloom”. Human genomics 2019, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hamil, K.G.; Liu, Q.; Sivashanmugam, P.; Anbalagan, M.; Yenugu, S.; Soundararajan, R.; Grossman, G.; Rao, A.; E Birse, C.; Ruben, S.M.; et al. LCN6, a novel human epididymal lipocalin. Reprod. Biol. Endocrinol. 2003, 1, 112–112. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Kaur, S.; Guha, S.; Batra, S.K. The multifaceted roles of neutrophil gelatinase associated lipocalin (NGAL) in inflammation and cancer. Biochim. Biophys. Acta BBA Rev. Cancer 2012, 1826, 129–169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y. and Rui, L. Lipocalin 13 regulation of glucose and lipid metabolism in obesity. Vitamins & Hormones 2013, 91, 369–383. [Google Scholar]
- Åkerstrom, B.; Flower, D.R.; Salier, J.-P. Lipocalins: unity in diversity. Biochim. et Biophys. Acta (BBA) - Protein Struct. Mol. Enzym. 2000, 1482, 1–8. [Google Scholar] [CrossRef]
- Fouchécourt, S.; Lareyre, J.-J.; Chaurand, P.; DaGue, B.B.; Suzuki, K.; Ong, D.E.; Olson, G.E.; Matusik, R.J.; Caprioli, R.M.; Orgebin-Crist, M.-C. Identification, Immunolocalization, Regulation, and Postnatal Development of the Lipocalin EP17 (Epididymal Protein of 17 Kilodaltons) in the Mouse and Rat Epididymis. Endocrinology 2003, 144, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Lareyre, J.-J.; Sánchez, D.; Gutierrez, G.; Araki, Y.; Matusik, R.J.; Orgebin-Crist, M.-C. Molecular evolution of epididymal lipocalin genes localized on mouse chromosome 2. Gene 2004, 339, 49–59. [Google Scholar] [CrossRef]
- Chu, S.-T.; Lee, Y.-C.; Nein, K.-M.; Chen, Y.-H. Expression, immunolocalization and sperm-association of a protein derived from 24p3 gene in mouse epididymis. Mol. Reprod. Dev. 2000, 57, 26–36. [Google Scholar] [CrossRef]
- Garrett, S.H.; Garrett, J.E.; Douglass, J. In situ histochemical analysis of region-specific gene expression in the adult rat epididymis. Mol. Reprod. Dev. 1991, 30, 1–17. [Google Scholar] [CrossRef]
- Rankin, T.L.; Tsuruta, K.J.; Holland, M.K.; Griswold, M.D.; Orgebin-Crist, M.-C. Isolation, Immunolocalization, and Sperm-Association of Three Proteins of 18, 25, and 29 Kilodaltons Secreted by the Mouse Epididymis1. Biol. Reprod. 1992, 46, 747–766. [Google Scholar] [CrossRef]
- Ong, D.E.; Newcomer, M.E.; Lareyre, J.-J.; Orgebin-Crist, M.-C. Epididymal retinoic acid-binding protein. Biochim. et Biophys. Acta (BBA) - Protein Struct. Mol. Enzym. 2000, 1482, 209–217. [Google Scholar] [CrossRef]
- Tone, M.; Nolan, K.F.; Walsh, L.A.; Tone, Y.; Thompson, S.A.; Waldmann, H. Structure and chromosomal location of mouse and human CD52 genes. Biochim. et Biophys. Acta (BBA) - Gene Struct. Expr. 1999, 1446, 334–340. [Google Scholar] [CrossRef]
- Pujianto, D.A.; Permatasari, S. Mouse CD52 is predominantly expressed in the cauda epididymis, regulated by androgen and lumicrine factors. J. Hum. Reprod. Sci. 2021, 14, 350–355. [Google Scholar] [CrossRef]
- Ernesto, J.I.; Muñoz, M.W.; Battistone, M.A.; Vasen, G.; Martínez-López, P.; Orta, G.; Figueiras-Fierro, D.; De la Vega-Beltran, J.L.; Moreno, I.A.; Guidobaldi, H.A.; et al. CRISP1 as a novel CatSper regulator that modulates sperm motility and orientation during fertilization. J. Cell Biol. 2015, 210, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Da Ros, V.G.; Maldera, J.A.; Willis, W.D.; Cohen, D.J.; Goulding, E.H.; Gelman, D.M.; Rubinstein, M.; Eddy, E.M.; Cuasnicu, P.S. Impaired sperm fertilizing ability in mice lacking Cysteine-RIch Secretory Protein 1 (CRISP1). Dev. Biol. 2008, 320, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.W.; Battistone, M.A.; Carvajal, G.; Maldera, J.A.; Curci, L.; Torres, P.; Lombardo, D.; Pignataro, O.P.; Da Ros, V.G.; Cuasnicu, P.S.; et al. Influence of the genetic background on the reproductive phenotype of mice lacking Cysteine-Rich Secretory Protein 1 (CRISP1). Biol. Reprod. 2018, 99, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Haendler, B.E.R.N.A.R.D. , Krätzschmar, J., Theuring, F.R.A.N.Z. and Schleuning, W.D. Transcripts for cysteine-rich secretory protein-1 (CRISP-1; DE/AEG) and the novel related CRISP-3 are expressed under androgen control in the mouse salivary gland. Endocrinology 1993, 133, 192–198. [Google Scholar] [CrossRef]
- Jalkanen, J.; Huhtaniemi, I.; Poutanen, M. Mouse Cysteine-Rich Secretory Protein 4 (CRISP4): A Member of the Crisp Family Exclusively Expressed in the Epididymis in an Androgen-Dependent Manner1. Biol. Reprod. 2005, 72, 1268–1274. [Google Scholar] [CrossRef]
- Cohen, D.J.; Maldera, J.A.; Vasen, G.; Ernesto, J.I.; Muñoz, M.W.; Battistone, M.A.; Cuasnicú, P.S. Epididymal Protein CRISP1 Plays Different Roles During the Fertilization Process. J. Androl. 2011, 32, 672–678. [Google Scholar] [CrossRef]
- Carvajal, G.; Brukman, N.G.; Muñoz, M.W.; Battistone, M.A.; Guazzone, V.A.; Ikawa, M.; Haruhiko, M.; Lustig, L.; Breton, S.; Cuasnicu, P.S. Impaired male fertility and abnormal epididymal epithelium differentiation in mice lacking CRISP1 and CRISP4. Sci. Rep. 2018, 8, 17531. [Google Scholar] [CrossRef]
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: the evolving tale of aquaporin biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, N.; Silberstein, C.; Beaulieu, V.; Piétrement, C.; Van Hoek, A.N.; Brown, D.; Breton, S. Postnatal Expression of Aquaporins in Epithelial Cells of the Rat Epididymis1. Biol. Reprod. 2006, 74, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.Y.; Li, Y.; Bi, Z.W.; Yu, H.M.; Li, X.J. Expression and Immunohistochemical Localization of Aquaporin-1 in Male Reproductive Organs of the Mouse. Anat. Histol. Embryol. 2007, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Squillacioti, C.; Mirabella, N.; Liguori, G.; Germano, G.; Pelagalli, A. Aquaporins Are Differentially Regulated in Canine Cryptorchid Efferent Ductules and Epididymis. Animals 2021, 11, 1539. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Lee, B.; Kim, J.; Kim, J.; Hong, S.H.; Kim, D.; Choi, S.; Cho, B.-N.; Cho, C. Expressional and functional analyses of epididymal SPINKs in mice. Gene Expr. Patterns 2019, 31, 18–25. [Google Scholar] [CrossRef]
- Jalkanen, J.; Kotimäki, M.; Huhtaniemi, I.; Poutanen, M. Novel epididymal protease inhibitors with Kazal or WAP family domain. Biochem. Biophys. Res. Commun. 2006, 349, 245–254. [Google Scholar] [CrossRef]
- Ma, L.; Yu, H.; Ni, Z.; Hu, S.; Ma, W.; Chu, C.; Liu, Q.; Zhang, Y. Spink13, an Epididymis-specific Gene of the Kazal-type Serine Protease Inhibitor (SPINK) Family, Is Essential for the Acrosomal Integrity and Male Fertility. J. Biol. Chem. 2013, 288, 10154–10165. [Google Scholar] [CrossRef]
- Mangiola, S.; A Doyle, M.; Papenfuss, A.T. Interfacing Seurat with the R tidy universe. Bioinformatics 2021, 37, 4100–4107. [Google Scholar] [CrossRef]
- Satija, R.; A Farrell, J.; Gennert, D.; Schier, A.F.; Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 2015, 33, 495–502. [Google Scholar] [CrossRef]
- Zhang, J.-S.; Liu, Q.; Li, Y.-M.; Hall, S.H.; French, F.S.; Zhang, Y.-L. Genome-wide profiling of segmental-regulated transcriptomes in human epididymis using oligo microarray. Mol. Cell. Endocrinol. 2006, 250, 169–177. [Google Scholar] [CrossRef]
- Suzuki, K.; Yu, X.; Chaurand, P.; Araki, Y.; Lareyre, J.-J.; Caprioli, R.M.; Orgebin-Crist, M.-C.; Matusik, R.J. Epididymis-specific lipocalin promoters. Asian J. Androl. 2007, 9, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, J.; Yamanaka, H.; Taguchi, I.; Shida, K. Free Amino Acids in the Caput and the Cauda Epididymis of Adult Rats. Endocrinol. Jpn. 1976, 23, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, J.-L.; Castella, S.; Gatti, J.L.; Dacheux, F. Epididymal cell secretory activities and the role of proteins in boar sperm maturation. Theriogenology 2005, 63, 319–341. [Google Scholar] [CrossRef]
- Axnér, E.; Linde-Forsberg, C.; Einarsson, S. Morphology and motility of spermatozoa from different regions of the epididymal duct in the domestic cat. Theriogenology 1999, 52, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, T.; Wada, M.; Anzai, M.; Hori, T. Artificial Insemination with Frozen Epididymal Sperm in Cats. J. Veter- Med Sci. 2003, 65, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Toyonaga, M.; Sato, Y.; Sasaki, A.; Kaihara, A.; Tsutsui, T. Artificial insemination with cryopreserved sperm from feline epididymides stored at 4 °C. Theriogenology 2011, 76, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H. , Martinez, E.A., Calvete, J.J., Pena Vega, F.J. and Roca, J. Seminal plasma: relevant for fertility? International Journal of Molecular Sciences 2021, 22, 4368. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Kvist, U.; Saravia, F.; Wallgren, M.; Johannissono, A.; Sanz, L.; Pena, F.; Martinez, E.; Roca, J.; Vazquez, J.; et al. The physiological roles of the boar ejaculate. 2020, 18, 1–21. [CrossRef]
- E Lake, P. The male reproductive tract of the fowl. . 1957, 91, 116–29. [Google Scholar]
- Lee, J.Y.; Dada, R.; Sabanegh, E.; Carpi, A.; Agarwal, A. Role of Genetics in Azoospermia. Urology 2011, 77, 598–601. [Google Scholar] [CrossRef]
- Maduro, M.R.; Lamb, D.J. Understanding new genetics of male infertility. . 2002, 168, 2197–205. [Google Scholar] [CrossRef]
- O'Hara, L.; Welsh, M.; Saunders, P.T.; Smith, L.B. Androgen Receptor Expression in the Caput Epididymal Epithelium Is Essential for Development of the Initial Segment and Epididymal Spermatozoa Transit. Endocrinology 2010, 152, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S. , Jeremias, J., Bongiovanni, A.M. and Munoz, M.G. Immune regulation in the male genital tract. Infectious diseases in obstetrics and gynecology 1996, 4, 131–135. [Google Scholar] [CrossRef]
- Wagenlehner, F.; Pilatz, A.; Linn, T.; Diemer, T.; Schuppe, H.C.; Schagdarsurengin, U.; Hossain, H.; Meinhardt, A.; Ellem, S.; Risbridger, G.; et al. Prostatitis and andrological implications. . 2013, 65. [Google Scholar]
- Aitken, R.; Smith, T.; Jobling, M.; Baker, M.; De Iuliis, G. Oxidative stress and male reproductive health. Asian J. Androl. 2014, 16, 31–8. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, N.B. , Dufaure, I., Lareyre, J.J., Rigaudiere, N., Mattei, M.G. and Dufaure, J.P. Structural organization and regulation of the gene for the androgen-dependent glutathione peroxidase-like protein specific to the mouse epididymis. Molecular Endocrinology 1993, 7, 258–272. [Google Scholar] [PubMed]
- Drevet, J.R. The antioxidant glutathione peroxidase family and spermatozoa: A complex story. Mol. Cell. Endocrinol. 2006, 250, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Wijayarathna, R.; Hedger, M.P. Activins, follistatin and immunoregulation in the epididymis. Andrology 2019, 7, 703–711. [Google Scholar] [CrossRef]
- Jones, R.C.; Dacheux, J.-L.; Nixon, B.; Ecroyd, H.W. Role of the epididymis in sperm competition. Asian J. Androl. 2007, 9, 493–499. [Google Scholar] [CrossRef]
- Elbashir, S.; Magdi, Y.; Rashed, A.; Henkel, R.; Agarwal, A. Epididymal contribution to male infertility: An overlooked problem. Andrologia 2020, 53, e13721. [Google Scholar] [CrossRef]
- Lemaire, R. [Contributions to the histochemical study of nuclear inclusions in the dog epididymis]. . 1960, 154, 460–3. [Google Scholar]
- Shorluian, P.M. [A case of primary cancer of the epididymis]. . 1960, 25. [Google Scholar]
- Krylov, V.S.; Borovikov, A.M. Microsurgical method of reuniting ductus epididymis. Fertil. Steril. 1984, 41, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Goyal, H.; Vig, M.M. Histochemical activity of alkaline phosphatase and acid phosphatase in the epididymis of mature intact and androgen-deprived bulls. . 1984, 45, 444–50. [Google Scholar] [PubMed]
- BRIZ, M.D. , BONET, S., PINART, B., EGOZCUE, J. and CAMPS, R. Comparative study of boar sperm coming from the caput, corpus, and cauda regions of the epididymis. Journal of andrology 1995, 16, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Setty, B.S.; Jehan, Q. Functional maturation of the epididymis in the rat. Reproduction 1977, 49, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yao, T.; Weng, X.; Yao, R.; Li, W.; Xie, L.; Yue, X.; Li, F. Antioxidant properties and transcriptome of cauda epididymis with different levels of fertility in Hu lambs. Theriogenology 2022, 182, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ahmed, S.; Ahmed, S.; Yangliu, Y.; Wang, H.; Cai, X. Analysis of long non-coding RNAs in epididymis of cattleyak associated with male infertility. Theriogenology 2020, 160, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Cyr, D.G.; Dufresne, J.; Gregory, M. Cellular junctions in the epididymis, a critical parameter for understanding male reproductive toxicology. Reprod. Toxicol. 2018, 81, 207–219. [Google Scholar] [CrossRef]
- Belleannée, C.; Thimon, V.; Sullivan, R. Region-specific gene expression in the epididymis. Cell Tissue Res. 2012, 349, 717–731. [Google Scholar] [CrossRef]
- Zhao, W.; Solangi, T.H.; Wu, Y.; Yang, X.; Xu, C.; Wang, H.; Zheng, X.; Cai, X.; Zhu, J. Comparative rna-seq analysis of region-specific miRNA expression in the epididymis of cattleyak. Reprod. Domest. Anim. 2021, 56, 555–576. [Google Scholar] [CrossRef]
- Wu, C.; Wang, C.; Zhai, B.; Zhao, Y.; Zhao, Z.; Yuan, Z.; Zhang, M.; Tian, K.; Fu, X. Study of microRNA Expression Profile in Different Regions of Ram Epididymis. Reprod. Domest. Anim. 2021, 56, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Carnett, J.B.; Levi, J.V.; E Pennington, M. The surgical treatment of sterility due to obstruction at the epididymis; together with a study of the morphology of human spermatozoa. . 1902, 15, 2–15. [Google Scholar]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Légaré, C.; Lamontagne-Proulx, J.; Breton, S.; Soulet, D. Revisiting structure/functions of the human epididymis. Andrology 2019, 7, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Abou-Haïla, A.; Fain-Maurel, M.-A. Regional differences of the proximal part of mouse epididymis: Morphological and histochemical characterization. Anat. Rec. 1984, 209, 197–208. [Google Scholar] [CrossRef]
- Kirchhoff, C. The dog as a model to study human epididymal function at a molecular level. Mol. Hum. Reprod. 2002, 8, 695–701. [Google Scholar] [CrossRef]
- Chen, H.; Alves, M.B.R.; Belleannée, C. Contribution of epididymal epithelial cell functions to sperm epigenetic changes and the health of progeny. Hum. Reprod. Updat. 2021, 28, 51–66. [Google Scholar] [CrossRef]
- Robaire, B.; Hamzeh, M. Androgen Action in the Epididymis. J. Androl. 2011, 32, 592–599. [Google Scholar] [CrossRef]
- Turner, T.T. , Bomgardner, D., Jacobs, J.P. and Nguyen, Q.A.T. Association of segmentation of the epididymal interstitium with segmented tubule function in rats and mice. REPRODUCTION-CAMBRIDGE- 2003, 125, 871–878. [Google Scholar] [CrossRef]
- Guyonnet, B.; Marot, G.; Dacheux, J.-L.; Mercat, M.-J.; Schwob, S.; Jaffrézic, F.; Gatti, J.-L. The adult boar testicular and epididymal transcriptomes. BMC Genom. 2009, 10, 369–369. [Google Scholar] [CrossRef]
Figure 1.
Paper counts published annually on epididymis. Search for epididymis keywords in PubMed. The number of articles published each year is shown in the line graph. From 1960 to 2022, articles about epididymis published in PubMed every year. More and more articles are published every year, and the trend is rising in a straight line.
Figure 1.
Paper counts published annually on epididymis. Search for epididymis keywords in PubMed. The number of articles published each year is shown in the line graph. From 1960 to 2022, articles about epididymis published in PubMed every year. More and more articles are published every year, and the trend is rising in a straight line.
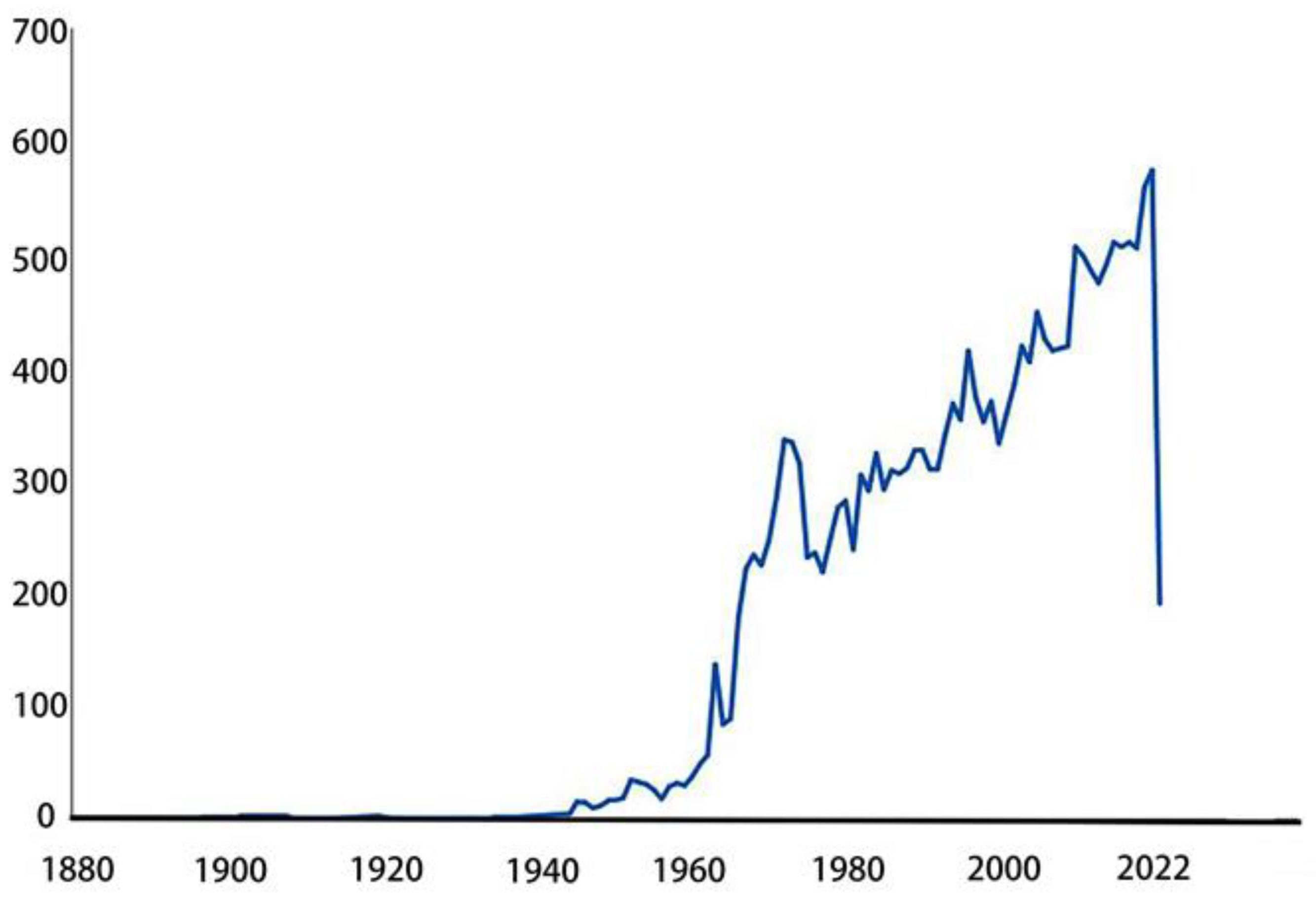
Figure 2.
Paper counts published annually on region specific-gene expresion in epididymis. Search for epididymis region specific gene expression keywords in PubMed. The number of articles published each year is shown in the line graph. It is found that relevant articles only appeared in 1983. From 1983 to 2022, the trend first rose and then declined.
Figure 2.
Paper counts published annually on region specific-gene expresion in epididymis. Search for epididymis region specific gene expression keywords in PubMed. The number of articles published each year is shown in the line graph. It is found that relevant articles only appeared in 1983. From 1983 to 2022, the trend first rose and then declined.
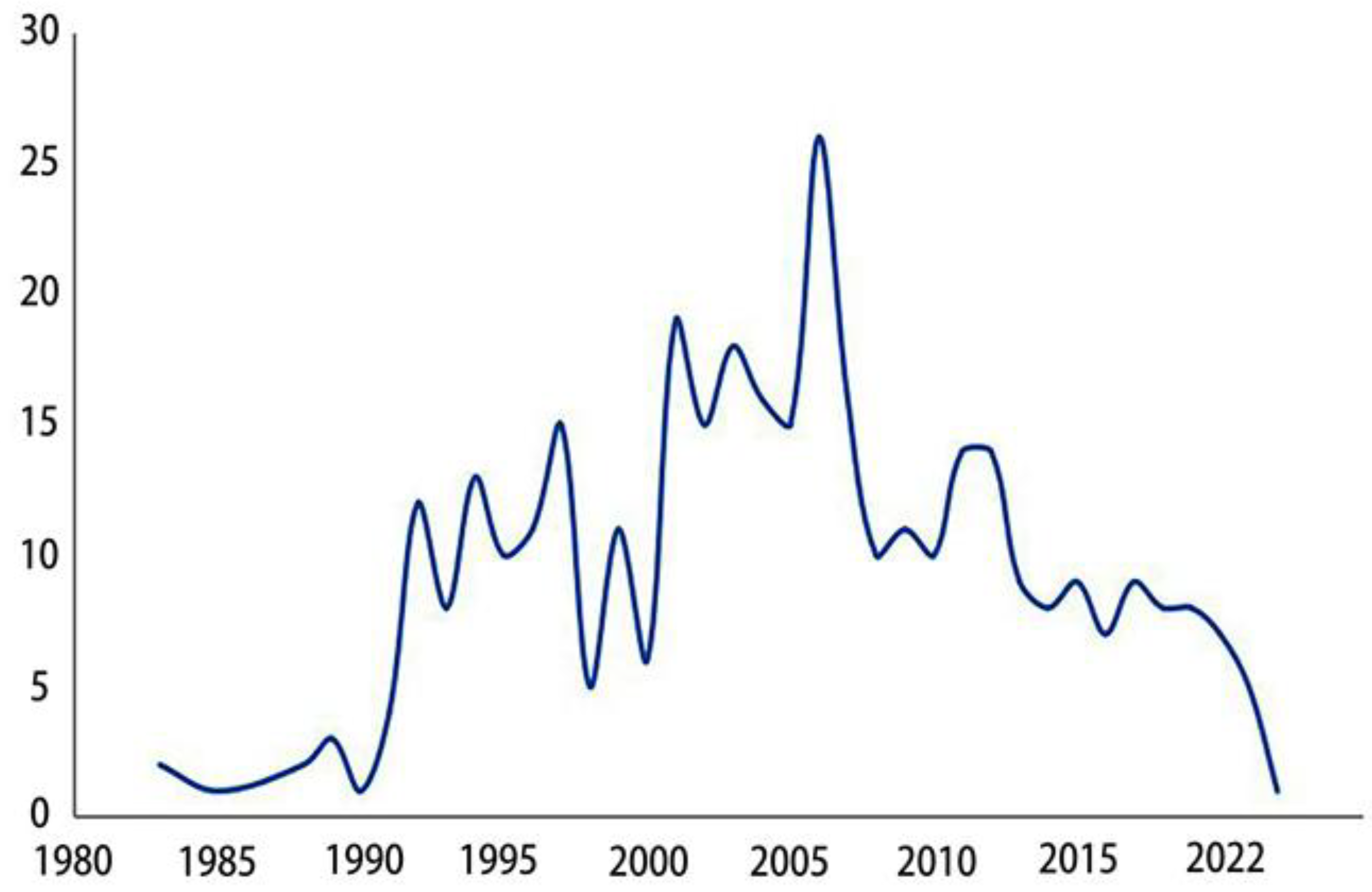
Figure 3.
Anatomy of the epididymis. The epididymis is attached to the testis. From top to bottom, it is the initial segment, the caput of the epididymis, the corpus of the epididymis, and the cauda of the epididymis.
Figure 3.
Anatomy of the epididymis. The epididymis is attached to the testis. From top to bottom, it is the initial segment, the caput of the epididymis, the corpus of the epididymis, and the cauda of the epididymis.
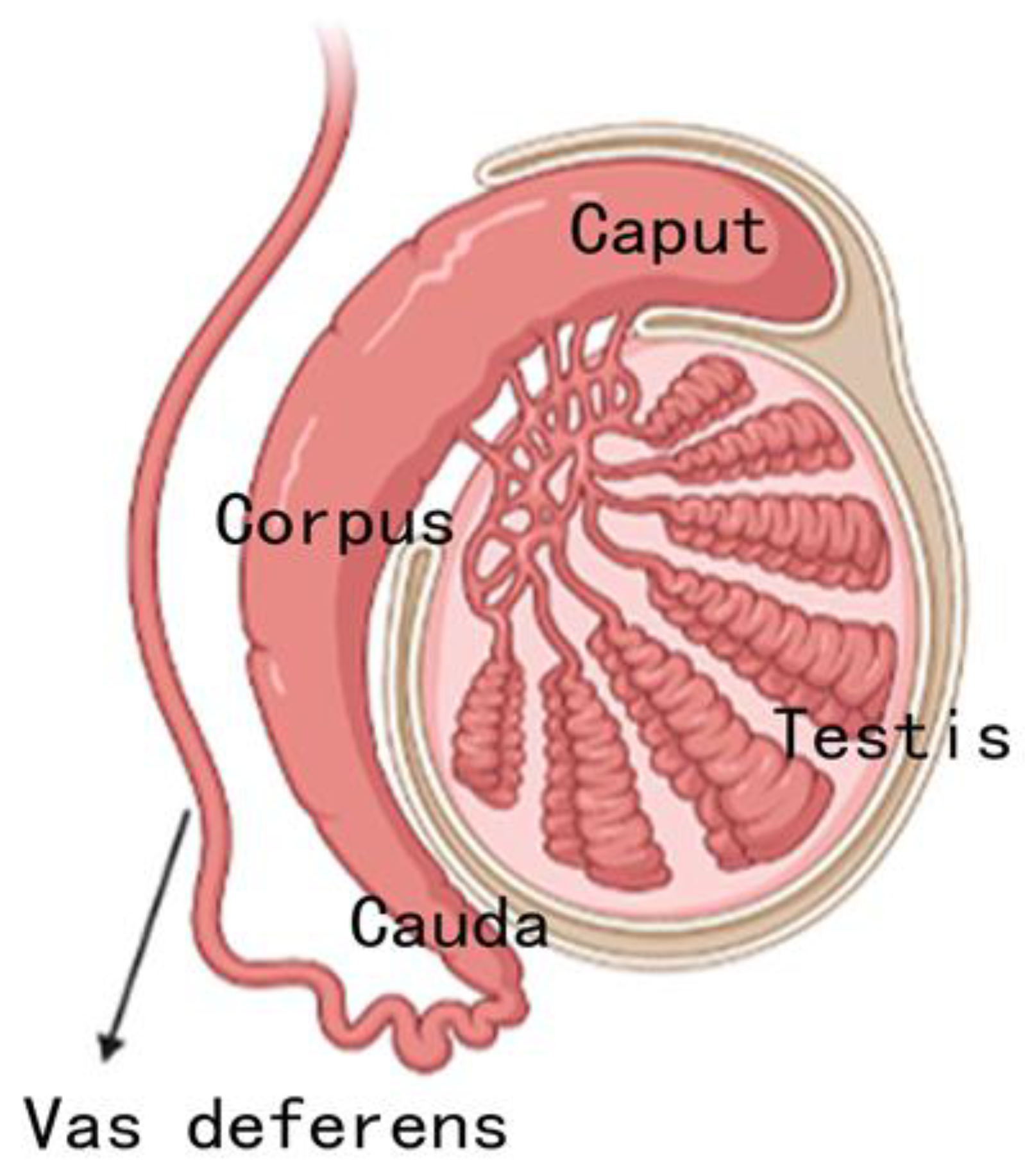
Figure 4.
Single-cell cluster, clustered by t-SNE and annotated according to the four anatomical regions (caput, corpus, cauda and vas deferens). The data of epididymal caput, carpus, cauda and vas deferens were analyzed jointly. This is the t-SNE cluster diagram of four anatomical regions. Each color represents a cluster of cells in an anatomical region. The red dot represents the epididymis caput cells; The blue dot represents the epididymis corpus cells; The green dot represents the epididymis cauda cells; The purple dot represents the epididymis vas deference cells.
Figure 4.
Single-cell cluster, clustered by t-SNE and annotated according to the four anatomical regions (caput, corpus, cauda and vas deferens). The data of epididymal caput, carpus, cauda and vas deferens were analyzed jointly. This is the t-SNE cluster diagram of four anatomical regions. Each color represents a cluster of cells in an anatomical region. The red dot represents the epididymis caput cells; The blue dot represents the epididymis corpus cells; The green dot represents the epididymis cauda cells; The purple dot represents the epididymis vas deference cells.
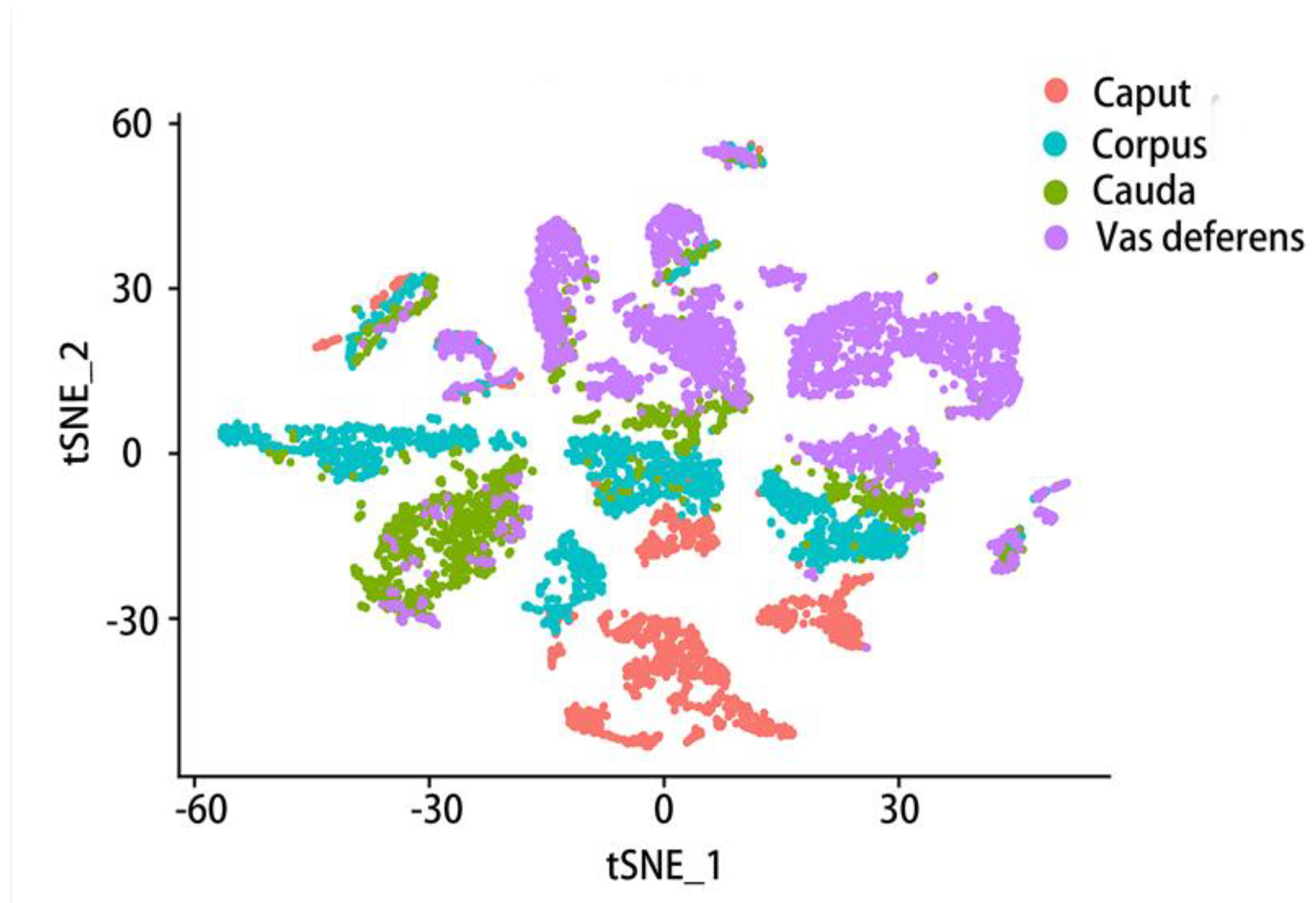
Figure 5.
Reclustering of extracted basal cells, visualized by t-SNE and annotated according to the four anatomical regions. This figure shows the t-SNE cluster diagram of basal cells in the caput, carpus, cauda and vas deferens of epididymis regions. Each color represents a cluster of cells in an anatomical region. The red dots represent the epididymis caput basal cells; The blue dots represent the epididymis corpus basal cells; The green dots represent the epididymis cauda basal cells; The purple dots represent the epididymis vas deference basal cells.
Figure 5.
Reclustering of extracted basal cells, visualized by t-SNE and annotated according to the four anatomical regions. This figure shows the t-SNE cluster diagram of basal cells in the caput, carpus, cauda and vas deferens of epididymis regions. Each color represents a cluster of cells in an anatomical region. The red dots represent the epididymis caput basal cells; The blue dots represent the epididymis corpus basal cells; The green dots represent the epididymis cauda basal cells; The purple dots represent the epididymis vas deference basal cells.
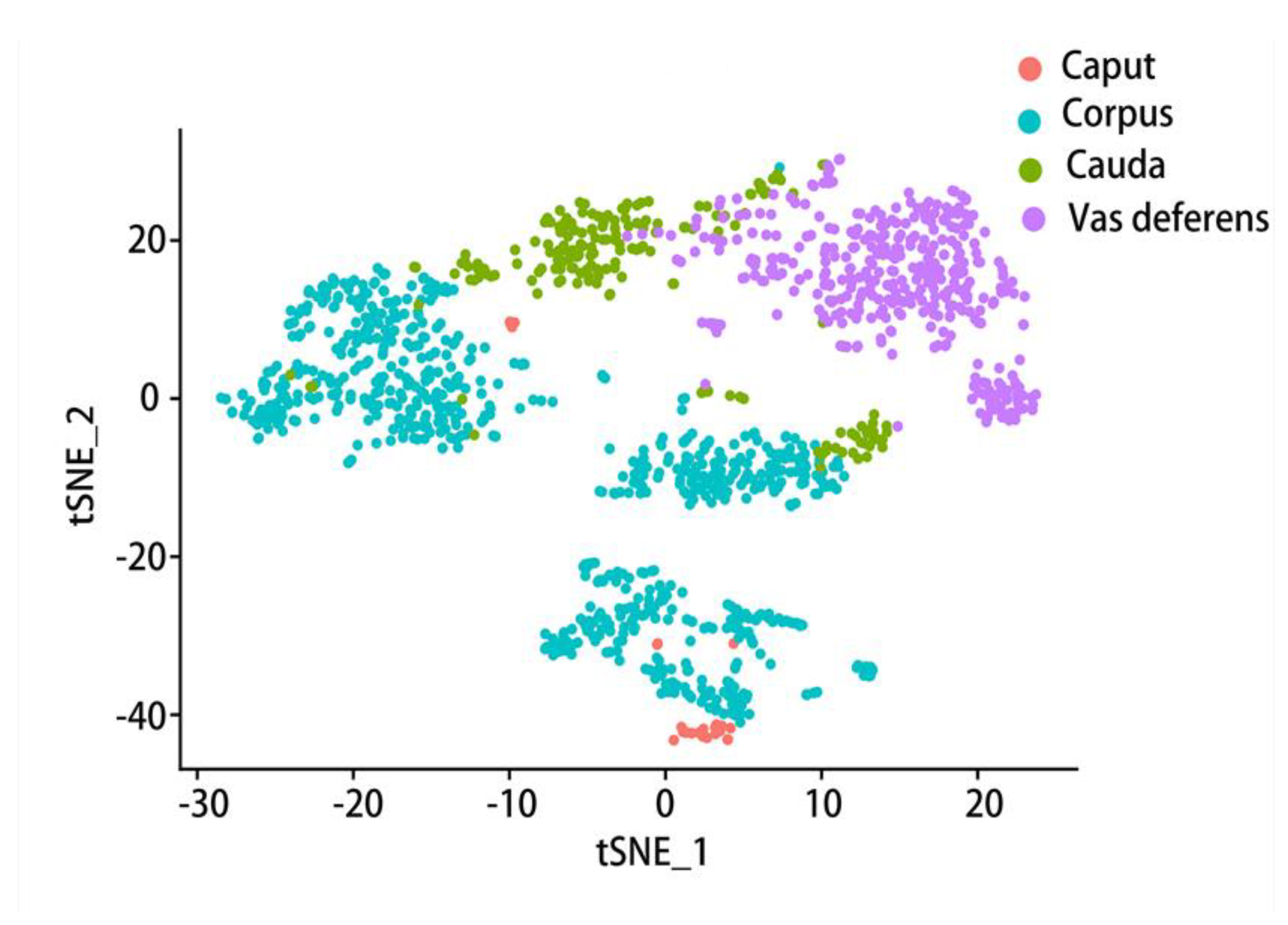
Figure 6.
Reclustering of extracted principal cells, visualized by t-SNE and annotated according to the four anatomical regions. This figure shows the t-SNE cluster diagram of principal cells in the caput, carpus, cauda and vas deferens of epididymis regions. Each color represents a cluster of cells in an anatomical region. The red dots represent the epididymis caput principal cells; The blue dots represent the epididymis corpus principal cells; The green dots represent the epididymis cauda principal cells; The purple dots represent the epididymis vas deference principal cells.
Figure 6.
Reclustering of extracted principal cells, visualized by t-SNE and annotated according to the four anatomical regions. This figure shows the t-SNE cluster diagram of principal cells in the caput, carpus, cauda and vas deferens of epididymis regions. Each color represents a cluster of cells in an anatomical region. The red dots represent the epididymis caput principal cells; The blue dots represent the epididymis corpus principal cells; The green dots represent the epididymis cauda principal cells; The purple dots represent the epididymis vas deference principal cells.
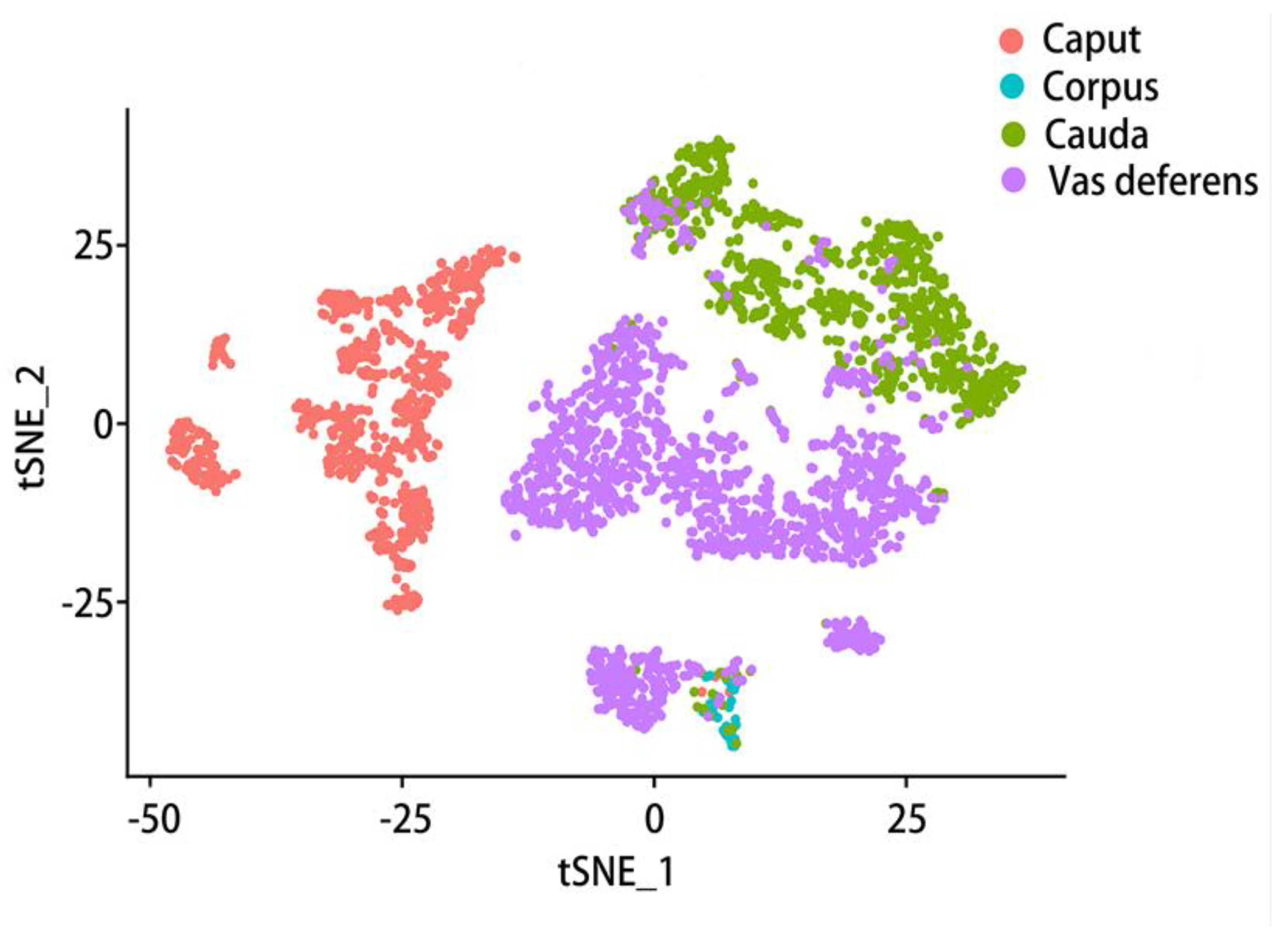
Figure 7.
Expression of specific genes across all basal cells. Those genes were selected and looked at their epididymal distribution in different parts respectively. A: Lcn8 is highly expressed in epididymis caput basal cells. B: Lcn5 is highly expressed in epididymis caput and corpus basal cells. C: Defb28 is highly expressed in corpus, cauda and vas deferens of epididymis basal cells. D: Spag11b is highly expressed in epididymis caput basal cells.
Figure 7.
Expression of specific genes across all basal cells. Those genes were selected and looked at their epididymal distribution in different parts respectively. A: Lcn8 is highly expressed in epididymis caput basal cells. B: Lcn5 is highly expressed in epididymis caput and corpus basal cells. C: Defb28 is highly expressed in corpus, cauda and vas deferens of epididymis basal cells. D: Spag11b is highly expressed in epididymis caput basal cells.
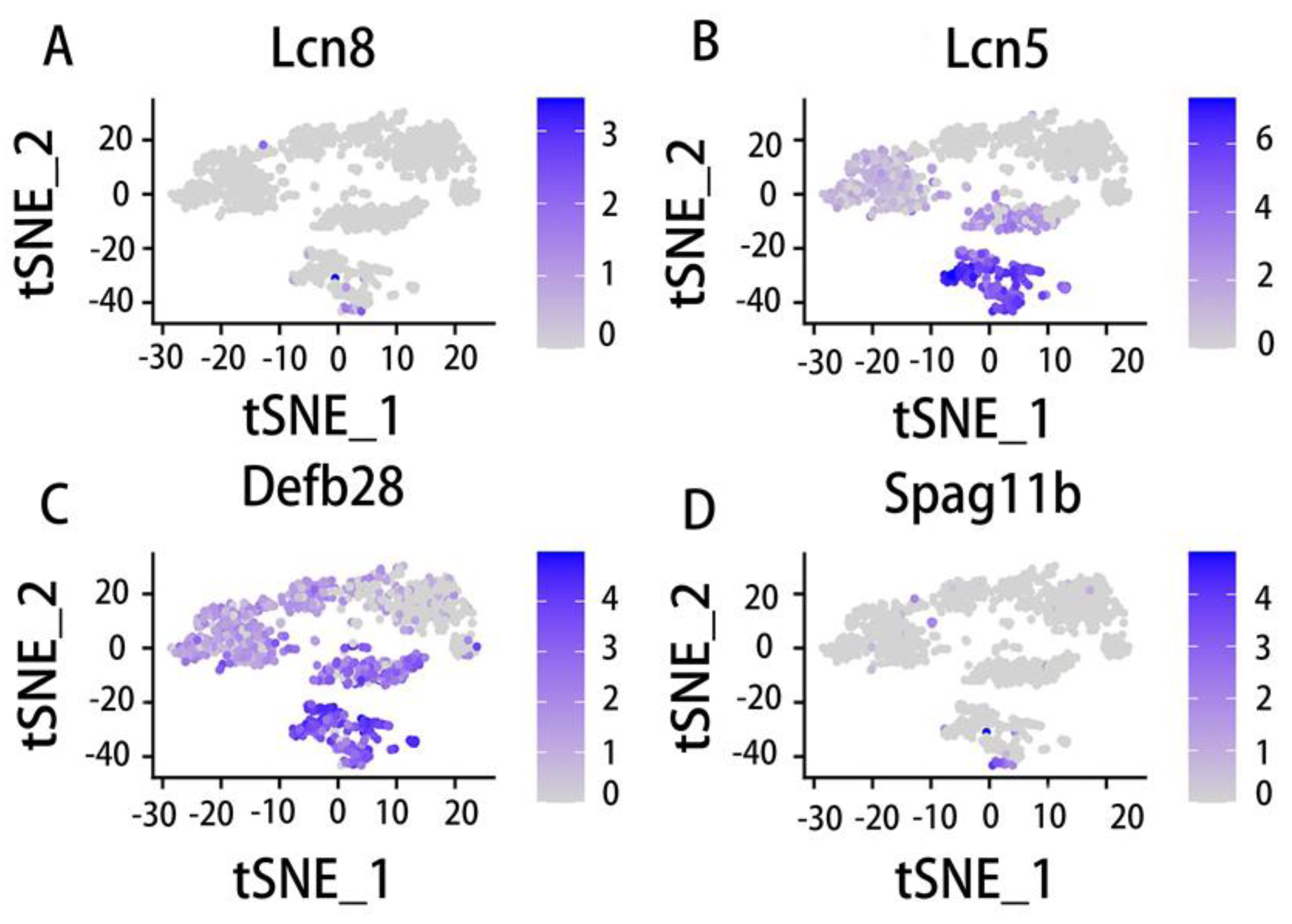
Figure 8.
Expression of specific genes across all principal cells. Those genes were selected and looked at their epididymal distribution in different parts respectively. A: Lcn8 is highly expressed in epididymis caput principal cells. B: Lcn5 is highly expressed in epididymis caput principal cells. C: Defb28 is highly expressed in epididymis cauda principal cells. D: Spag11b is highly expressed in epididymis caput principal cells.
Figure 8.
Expression of specific genes across all principal cells. Those genes were selected and looked at their epididymal distribution in different parts respectively. A: Lcn8 is highly expressed in epididymis caput principal cells. B: Lcn5 is highly expressed in epididymis caput principal cells. C: Defb28 is highly expressed in epididymis cauda principal cells. D: Spag11b is highly expressed in epididymis caput principal cells.
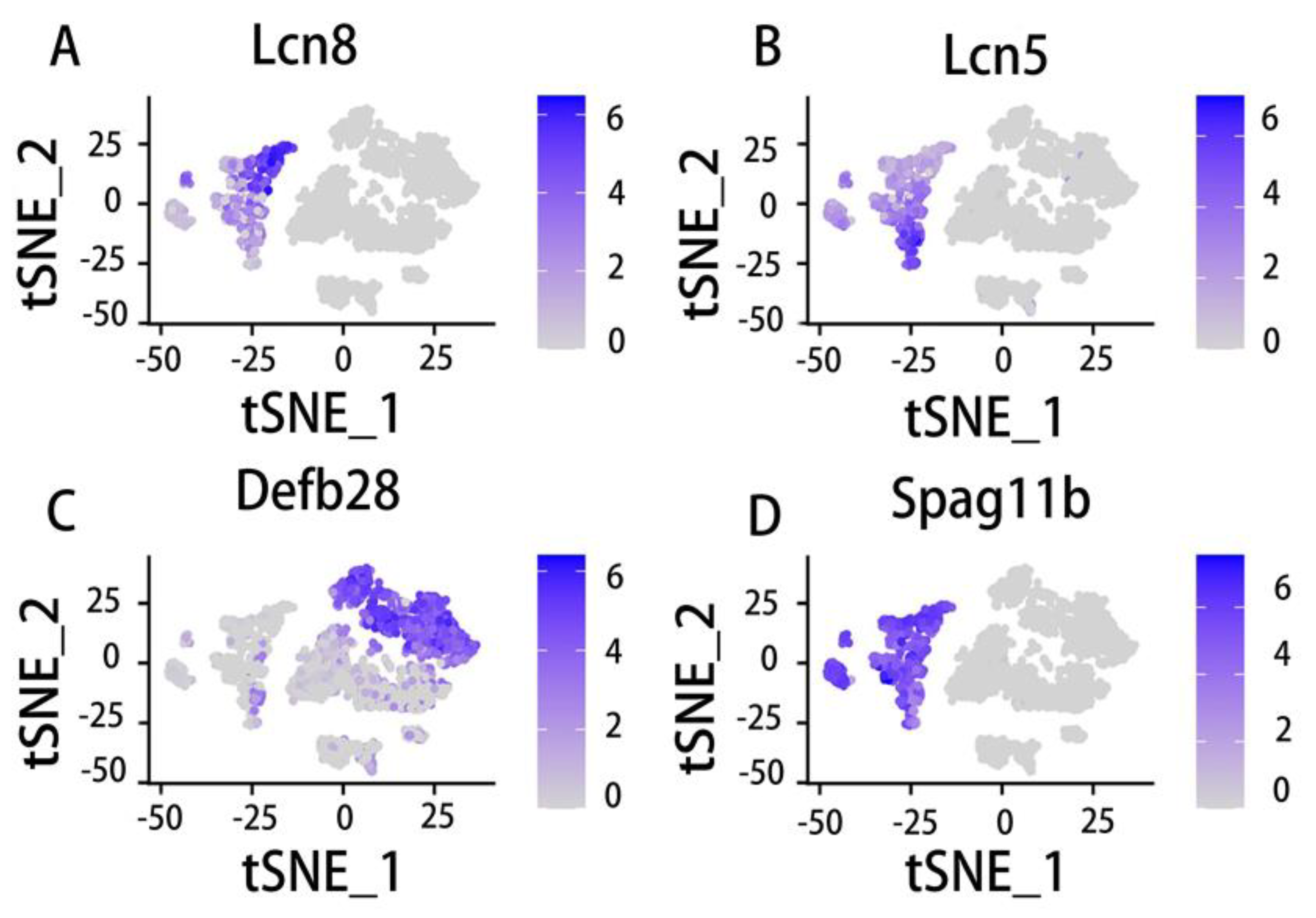

Table 1.
Representative articles on the application of new technologies in the field of epididymis research.
Table 1.
Representative articles on the application of new technologies in the field of epididymis research.
| Annual Range | Representative Article | New Technology | Findings |
|---|---|---|---|
| 1996-2000 | Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility | Gene-knockout | To explore the effect of specific genes on epididymal function |
| 2000-2001 | Stallion epididymal fluid proteome: qualitative and quantitative characterization; secretion and dynamic changes of major proteins | Proteomics | Understand epididymal mRNA expression and protein secretion |
| 2001-2005 | Dynamic changes in gene expression along the rat epididymis | Microarray technology | Obtain differential expression profiles of epididymis multiple genes. |
| 2005-2020 | The mouse epididymal transcriptome: transcriptional profiling of segmental gene expression in the epididymis | Transcriptome sequencing technology | It aims to improve the integrity of epididymal transcriptome by using whole genome array, and provide higher sensitivity by studying fragments in each region |
| 2020-2022 | An atlas of cell types in the mouseepididymis and vas deferens | Single-cell RNA-Seq | A bird's-eye view of the cell composition of epididymis was conducted to determine the new biological characteristics of epididymal cells. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



