
Submitted:
03 May 2023
Posted:
04 May 2023
You are already at the latest version
Abstract
Anti-islet autoantibodies serve as key markers in immune-mediated type 1 diabetes (T1D) and slowly-progressive T1D (SPIDDM), also known as latent autoimmune diabetes in adults (LADA). Autoantibodies to insulin (IAA), glutamic acid decarboxylase (GADA), tyrosine phosphatase-like protein IA-2 (IA-2A), and zinc transporter 8 (ZnT8A) are currently employed in the diagnosis, pathological analysis, and prediction of T1D. GADA can also be detected in non-diabetic patients with autoimmune diseases other than T1D and may not necessarily reflect insulitis. Conversely, IA-2A and ZnT8A serve as surrogate markers of pancreatic β-cell destruction. Combinatorial analysis of these four anti-islet autoantibodies demonstrated that 93-96% of acute-onset T1D and SPIDDM cases were diagnosed as immune-mediated T1D, while the majority of fulminant T1D cases were autoantibody-negative. Evaluating the epitopes and immunoglobulin subclasses of anti-islet autoantibodies help distinguish between diabetes-associated and non-diabetes-associated autoantibodies and is valuable for predicting future insulin deficiency in SPIDDM (LADA) patients. Additionally, GADA in T1D patients with autoimmune thyroid disease reveals the polyclonal expansion of autoantibody epitopes and immunoglobulin subclasses. Recent advancements in anti-islet autoantibody assays include nonradioactive fluid-phase assays and the simultaneous determination of multiple biochemically defined autoantibodies. Developing a high-throughput assay for detecting epitope-specific or immunoglobulin isotype-specific autoantibodies will facilitate more accurate diagnosis and prediction of autoimmune disorders.
Keywords:
Enzyme-linked immunosorbent assay
; Epitope
; Glutamic acid decarboxylase
; Latent-autoimmune diabetes in adults
; Prediction
; Type 1 diabetes
1. Introduction
Diabetes mellitus is a chronic metabolic disorder characterized by hyperglycemia, and is classified into four categories: type 1 diabetes (T1D), type 2 diabetes (T2D), specific types of diabetes due to other causes, and gestational diabetes [1]. Patients with untreated or uncontrolled hyperglycemia over a prolonged period of time may develop microvascular and macrovascular complications, such as diabetic neuropathy, retinopathy, nephropathy, cardiovascular and cerebrovascular disease, and peripheral vascular disease. Consequently, early detection or prediction of diabetes onset and timely intervention with appropriate medications, alongside medical nutrition therapy and exercise, are crucial.
T1D is an organ-specific autoimmune disease characterized by pancreatic β-cell destruction, leading to absolute insulin deficiency. Evidence supporting the autoimmune basis of T1D includes: i) the presence of lymphocytic infiltration around and into the islets (termed insulitis), ii) the appearance of autoantibodies to multiple islet autoantigens, iii) the presence of both major histocompatibility complex (MHC)-linked and non-MHC-linked disease susceptibility genes, and iv) the increased propensity to develop multiple organ-specific autoimmune diseases [2].
The risk of developing T1D varies considerably based on country of residence and ethnicity, with Japan having one of the lowest incidence rates of T1D worldwide [3]. This variation may be attributed to differences in genetic background and environmental factors. In the current etiological classification of diabetes, T1D is divided into immune-mediated and idiopathic types, distinguished solely by the presence or absence of anti-islet autoantibodies in peripheral blood [1]. Furthermore, based on the rate of β-cell destruction, there are three T1D subtypes: fulminant T1D, acute-onset T1D, and slowly-progressive T1D (SPIDDM), also known as latent autoimmune diabetes in adults (LADA) [4]. Since anti-islet autoantibodies are known to appear before disease onset, they serve as important humoral immune markers for predicting and diagnosing T1D. This review article explores the significance and clinical utility of anti-islet autoantibodies in Japanese T1D patients.
2. History of Anti-Islet Autoantibody Discovery
Discovery of islet cell antibodies (ICA) as the first anti-islet autoantibodies in T1D was made by Bottazzo and coworkers in 1974 [5]. ICA detection involved using indirect immunofluorescence on frozen pancreatic tissue sections of human blood group O, which may recognize various autoantigens. In 1982, Baekkeskov and coworkers discovered autoantibodies against an islet protein with a molecular weight of 64,000 (64kDa antibody) using the immunoprecipitation method and 35S-methionine-labeled human islet cells [6]. Moreover, in 1983, Palmer and coworkers reported insulin autoantibody (IAA) in insulin naïve new-onset patients with T1D, measured by polyethylene glycol competitive assay using 125I- Tyr A14 human monoiodinated insulin [7].
To overcome the limitation of ICA assay, such as being time-consuming, requiring human pancreatic tissue, and yielding difficulty in obtaining quantitative results, extensive efforts have been made to identify target antigens against ICA using advanced molecular biological techniques. To date, more than 10 target antigens have been discovered. After identifying the 64kDa islet protein as glutamic acid decarboxylase (GAD) in 1990 [8], several autoantibodies (Figure 1) were discovered. Currently, in addition to IAA and GAD autoantibodies (GADA), tyrosine phosphatase-like protein IA-2 autoantibodies (IA-2A) [9] and zinc transporter 8 autoantibodies (ZnT8A) [10] are employed for the diagnosis, pathological analysis, and prediction of T1D. Additionally, detection methods have evolved from immunohistochemical staining used in ICA to the radioimmunoassay (RIA), enzyme-linked immunosorbent assay (ELISA), and electrochemiluminescence (ECL) assay using recombinant autoantigens.
3. Significance of Anti-Islet Autoantibodies in the Pathophysiology of T1D
The Juvenile Diabetes Research Foundation (JDRF), American Diabetes Association (ADA), and Endocrine Society recommend classifying the natural history of T1D into three stages [11] (Figure 2). In Stage 1, autoimmunity to pancreatic islet cells is induced by cytotoxic T cells, and the destruction of pancreatic β-cells (insulitis) begins, even though blood glucose levels remain within the normal range. Two or more IAA, GADA, IA-2A, and ZnT8A, emerge in this stage, with the 5-year and 10-year risks of T1D development being approximately 44% and 70%, respectively, while the lifetime risk approaches 100% [12]. However, it is currently understood that anti-islet autoantibodies do not directly cause pancreatic β-cell destruction; they arise as the result of β-cell destruction mediated by T-cells. In Stage 2, like Stage 1, individuals test positive for two or more anti-islet autoantibodies and β-cell destruction progresses, leading to glucose intolerance or dysglycemia. The 5-year risk of T1D development at this stage is approximately 75%, and the lifetime risk approaches 100% [13]. Stage 3 is marked by the manifestations of typical clinical symptoms and signs of T1D, including polyuria, polydipsia, weight loss, general fatigue, diabetic ketoacidosis, and others. In this stage, the amount of residual pancreatic β-cells decreases to 20-30% of normal levels. Since anti-islet autoantibodies appear in the peripheral blood during Stage 1 and Stage 2, they are used as a tool for predicting the onset of T1D. The number of positive autoantibodies is more important for prediction than the specific positive autoantibodies [14].
Additionally, a study involving first-degree relatives of patients with T1D reported that IAA or GADA appeared first, followed by IA-2A and ZnT8A just before the onset of diabetes [15]. We observed a similar sequential emergence of anti-islet autoantibodies in patients with T1D who developed the condition during interferon therapy [16]. Based on this evidence, IA- 2A and ZnT8A are considered surrogate markers for pancreatic β-cell destruction. This is supported by a report demonstrating epitope spreading of IA-2A during the preclinical stage of T1D [17].
Conversely, as shown in Table 1, GADA is detected not only in patients with acute-onset T1D and SPIDDM but also in some patients with fulminant T1D and non-diabetic patients with polyglandular autoimmune syndrome, autoimmune thyroid disease (AITD), and stiff-person syndrome. Therefore, unlike IA-2A and ZnT8A, it is presumed that GADA may not be a specific marker for pancreatic β-cell destruction. In our case, where GADA became re- elevated more than 10 years after the onset of T1D, anti-thyroid autoantibodies turned positive just before the re-elevation of GADA [18]. Therefore, it is important to note that GADA may be associated with thyroid autoimmunity rather than insulitis in some instances.
4. Role of Anti-Islet Autoantibodies in the Diagnosis of T1D
In this section, we discuss the key considerations when diagnosing T1D using anti-islet autoantibodies. First, because distinguishing IAA from insulin antibodies produced by exogenous insulin injection is challenging, we considered patients positive for IAA if antibodies to insulin were present in insulin-naïve patients or those within 2 weeks of initiating insulin therapy. Our study using sera obtained within 2 weeks after onset revealed that the prevalence of IAA, GADA, IA-2A, and ZnT8A in patients with acute-onset T1D was 58%, 80%, 66%, and 63%, respectively (Figure 3A). Similarly, the prevalence of these autoantibodies in SPIDDM patients was 41%, 83%, 27%, and 27%, respectively (Figure 3B). Thus, a combinatorial analysis of the four autoantibodies indicated that 96% of acute-onset T1D and 93% of SPIDDM were immune-mediated T1D, while approximately 5% were classified as idiopathic T1D [19]. These data are consistent with previous reports from studies conducted in countries with Caucasoid populations [10,20,21,22,23]. In contrast, the majority of patients with fulminant T1D were negative for anti-islet autoantibodies (Figure 3C) and are classified into idiopathic T1D, suggesting that the mechanism of β-cell destruction in fulminant T1D may differ from acute- onset T1D and SPIDDM.
In Japan, medical insurance guidelines instruct physicians to measure GADA first when T1D is suspected. However, about 10% of non-fulminant T1D patients are positive for other anti-islet autoantibodies without GADA. Therefore, measuring multiple autoantibodies is crucial for accurate diagnosis of immune-mediated T1D.
5. Epitopes for Anti-Islet Autoantibodies and Their Clinical Relevance
- 1)
- Insulin autoantibodies
Insulin is a peptide hormone converted from proinsulin in pancreatic β-cells. Proinsulin consists of an A-chain (21 amino acids), a B-chain (30 amino acids) and a C-peptide (31 amino acids), and mature insulin is generated after C-peptide is excised by a series of proteolytic cleavages. Fewer epitope analysis studies have been conducted on IAA compared to those of GADA and IA-2A. In an earlier study, the significance of amino acid A13 of the A-chain for the binding of IAA in T1D was demonstrated [23]. Furthermore, it was revealed that conformational epitope spanning amino acid residues A8-A13 on the A-chain and B1-B3 on the B-chain was the major binding site for IAA and was disease-associated. Another epitope analysis using a recombinant Fab of the insulin-specific monoclonal antibody reported that the IAA epitope was located in the A-chain residues A8-A10 [24,25]. However, radioimmunoassay for IAA does not distinguish between epitopes on the insulin molecule bound by IAA from insulin antibodies induced by exogenous insulin injection [26]. Therefore, Devendra and coworkers used the random phage-display peptide library and serum obtained from an IAA-positive T1D patient and an insulin-treated insulin autoimmune syndrome patient to examine the difference in epitopes bound by both antibodies [27]. As a result, they identified the two phagotopes, designated IAS-9 and IDD-10, which were able to discriminate between diabetes-associated and non-diabetes associated insulin antibodies. This suggests that phage display technology could potentially be exploited to develop an IAA-specific RIA.
- 2)
- GAD autoantibodies
GAD is an enzyme that catalyzes a decarboxylation reaction to produce γ-aminobutyrate from L-glutamate. There are two isoforms of GAD, GAD65 (585 amino acids) and GAD67 (594 amino acids), both of which are abundant in the central nervous system and pancreatic islets. GADA is present in 70-80% of prediabetic relatives and new-onset patients with T1D. The major antigenic region of GADA has been determined using truncated peptide or chimeric proteins of GAD65 and GAD67 to maintain the conformational structure, as previous studies reported that GADA in patients with T1D recognizes the conformational structure of the GAD molecule [26]. Moreover, it has been reported that disease-associated GADA is directed against GAD65, and humoral immune reactivity to GAD67 is likely to be cross-reactive to GAD65 [27,28]. Figure 4A shows a schematic representation of the GAD65 and the localization of GADA epitopes in T1D. Previous studies have demonstrated that GADA in T1D patients recognizes disease-specific GAD65 epitopes, located at the middle and C-terminal regions of GAD65 [29,30]. However, these studies may have also detected epitopes that cross-react with the GAD67 autoantibody. To detect GAD65 autoantibody-specific epitopes, we performed competitive radioimmunoassay with recombinant GAD67 protein using a series of GAD65/GAD67 chimeric constructs, and identified the autoantibody epitopes in the N- terminal region (amino acids 1-244; N), the middle domain (amino acids 245-359; E1 and amino acids 360-442; E3), and the C-terminal region (amino acids 443-585; E2) of GAD65 [31].
Although GADA epitope specificities remain relatively stable after the clinical onset of T1D, it has been reported that in genetically predisposed subjects, GADA is initially generated against the middle and C-terminal regions of GAD65. Furthermore, the autoimmune response may undergo intramolecular epitope spreading towards epitopes on the N-terminus and further epitopes located in the middle [32].
- 3)
- IA-2 autoantibodies
IA-2 (also known as ICA512) is a transmembrane glycoprotein of the protein tyrosine phosphatase (PTP) family that colocalizes with IA-2β/phogrin and is expressed in insulin- secretory granule membranes in pancreatic β-cells [9,33,34]. As shown in Figure 4B, this protein is 979 amino acids long and comprises of a luminal domain (amino acids 27-576), transmembrane domain (amino acids 577-600), and cytoplasmic domain (amino acids 601-979). Although the PTP core sequence is found at amino acids 907-917 in the cytoplasmic domain, the expressed recombinant protein does not exhibit protein tyrosine phosphatase activity. Studies evaluating the biological properties of IA-2 have demonstrated its role as an important regulator of dense core vesicle number as well as glucose-induced and basal insulin secretion [35,36].
IA-2A is present in 60-70% of prediabetic relatives and new-onset patients with T1D. We and others have analyzed IA-2A epitopes recognized by diabetic sera using a series of IA-2 fragments or IA-2/phogrin chimeric proteins, and found that the major epitopes are localized in the cytoplasmic domain [37]. Approximately 95% of T1D patients and prediabetic relatives who are IA-2A positive recognize the PTP-like domain (amino acids 687-979), whereas only 5% of sera react with the luminal domain [37,38]. Furthermore, our binding and competition analysis using multiple IA-2/phogrin chimeric constructs demonstrated that a major unique epitope for IA-2A is localized to amino acids 762-887. A conformational epitope associated with the C-terminal 31 amino acids of IA-2 is recognized by one-third of sera, and a minor epitope is located on amino acids 601-762 of IA-2. Notably, intramolecular epitope spreading was found for relatives of T1D patients who later progressed to T1D. However, relatives who remained nondiabetic exhibited a decrease in the number of recognized epitopes. These studies are consistent with the hypothesis that IA-2 may be recognized as a consequence of β- cell destruction [17].
Another important epitope has been mapped in the juxta-membrane domain of IA-2 (amino acids 601-629; IA-2JM). Our data demonstrated that the age of disease onset in patients with IA-2JMA only was significantly higher than that in patients who reacted with the PTP-like domain, suggesting that autoantibody recognition of IA-2 epitopes in autoimmune diabetes is associated with the age of disease onset, which may reflect the intensity of the β-cell destruction process [39].
- 4)
- ZnT8 autoantibodies
ZnT8, a member of the zinc transporter family, is specifically expressed in the insulin- secretory granule membrane of pancreatic β-cells and is responsible for transportating zinc influx from the cytoplasm into insulin granules, and facilitating insulin hexamer formation [40,41]. Zinc ions transported into the insulin-secreting granule cavity by ZnT8 are used to stabilize insulin hexamers, and zinc ions ejected outside the cells during insulin secretion have an autocrine effect on β-cells, or alternatively, exert a paracrine action on α-cells. In 2007, Sladek and coworkers identified four loci containing variants that confer T2D risk through a genome-wide association study, including a non-synonymous polymorphism in the ZnT8 gene (SLC30A8), rs13266634 (C/T), which causes an R325W modification in the protein sequence [42]. In the same year, Hutton and coworkers discovered ZnT8 as a major autoantigen in T1D, and ZnT8A has been recognized as one of the four major anti-islet autoantibodies [10].
ZnT8A are present in 50-60% of prediabetic relatives and new-onset patients with T1D. As shown in Figure 4C, ZnT8 is a 369-amino acid polytopic transmembrane protein with cytoplasmic N- and C- terminal tails. It has been reported that ZnT8A recognizes 101 amino acids localized in the cytoplasmic C-terminal region. In particular, the amino acid residue 325 (R325W) defined by the SLC30A8 polymorphism is critical for humoral autoimmunity to this autoantigen, and binding of ZnT8A against two isotypes (ZnT8-325R, ZnT8-325W) depends on the patient's SLC30A8 genotype [43,44]. Consequently, heterozygotes with the CT genotype respond to both ZnT8-325R and ZnT8-325W, while CC and TT homozygotes respond exclusively to ZnT8-325R or ZnT8-325W, respectively. Thus, individuals respond to endogenous ZnT8 protein determined by their own genome, and therefore, the current ZnT8Aassay, therefore, uses a hybrid protein of two ZnT8 isotypes as antigens.
Furthermore, Wenzlau and coworkers identified that residues 332R, 333E, 336K, and 340K contribute to a conformational ZnT8A epitope independent of residue 325 by comparing human and mouse chimeric ZnT8 proteins [45], suggesting that this epitope may add to the diagnostic utility of measuring ZnT8A.
6. Prediction of Future Insulin Deficiency in Patients with SPIDDM (LADA)
SPIDDM (also known as LADA) is characterized by the presence of anti-islet autoantibodies and a gradual decline in insulin secretory capacity. The Immunology of Diabetes Society defined LADA as follows; 1) onset of diabetes >35 years, 2) positive test for at least one of the known anti-islet autoantibodies, and 3) requirement of insulin treatment >6 months after the diagnosis of diabetes [46]. LADA encompasses anti-islet autoantibody- positive diabetic patients in both insulin-dependent and non-insulin-dependent states, which is nearly identical to SPIDDM. According to the recently revised diagnostic criteria for SPIDDM, the interval from diabetes diagnosis to the requirement of insulin treatment is >3 months [47]. Additionally, patients with exhausted endogenous insulin secretion (Fasting C- peptide <0.6ng/ml) at the last observed time point are defined as “SPIDDM (definite)”. In contrast, anti-islet autoantibody-positive patients in non-insulin-dependent state are classified as "SPIDDM (probable)" (Table 2). Using this diagnostic criterion, measuring anti-islet autoantibodies other than GADA results in an approximately 3-fold increase in the incidence of SPIDDM among non-insulin-treated diabetic patients compared with measuring GADA alone (2.0-2.4% vs. 7-8%) [48,49,50]. Since this subtype of T1D is generally indistinguishable from T2D at the time of diagnosis, measuring anti-islet autoantibodies is crucial for early diagnosis and appropriate treatment of SPIDDM (LADA).
Anti-islet autoantibody positivity, especially ICA and GADA, is predictive for progression to a future insulin-dependent state after the diagnosis of diabetes. For example, the UKPDS (United Kingdom Prospective Diabetes Study) found that at least 50% of LADA patients required insulin treatment 6 years post-diagnosis [51]. However, not all SPIDDM (LADA) patients required insulin treatment, even after 10 years from diagnosis.
According to a nationwide survey [52] conducted by the Japan Diabetes Society, the predictors of progression to insulin dependent state include (1) age of onset ≤ 47 years, (2) period until GADA positive detection ≤ 5 years, (3) GADA titer (RIA method) ≥ 13.6 U/ml, and (4) fasting C-peptide ≤ 0.65 ng/mL. Additionally, the number of positive anti-islet autoantibodies and GADA epitope recognition are also important for prediction. To identify the predictive markers for early insulin requirement in non-insulin-dependent SPIDDM (probable), we evaluated IAA, IA-2A, and ZnT8A along with GADA-specific epitope recognition in 47 GADA-positive diabetic patients [49]. Among these patients, 38% had one or more of IAA, IA-2A, or ZnT8A and 15% had two or more of these autoantibodies. A high GADA titer (≥ 10U/mL), the presence of GADA-E1, and the presence of one or more among IAA, IA-2A, or ZnT8A at diagnosis marked the risk for early insulin therapy requirement. Furthermore, multiple anti-islet autoantibodies were the most relevant risk factor for the insulin requirement (odds ratio 13.77; 95% CI 2.77-68.45; P<0.001) in a multivariate logistic regression analysis. Therefore, measuring anti-islet autoantibodies other than GADA and ICA is essential for predicting the progression risk of SPIDDM (LADA) patients.
7. Type 1 Diabetes and Associated Autoimmune Diseases
Clinicians should be aware that complications arising from additional autoimmune disorders are more frequently observed in T1D patients [53], with the most common organ- specific autoimmune disease associated with T1D being autoimmune thyroid disease (AITD), such as Graves’ disease and Hashimoto’s thyroiditis, affecting more than 90% of people with T1D and autoimmune disorders [54,55]. Additionally, it has been reported that children, particularly girls, with T1D at an increased risk for developing other autoimmune diseases, with the prevalence of anti-thyroid autoantibodies at disease onset being about 20% [56]. Furthermore, the prevalence of anti-thyroid antibodies increases with age, and the presence of these antibodies at diagnosis of T1D is predictive of future thyroid disease [56]. Long-term follow-up suggests that up to 30% of patients with T1D develop AITD [57], and patients with thyroid autoimmunity are 18 times more likely to develop AITD than those without [58]. Therefore, to enable early diagnosis of AITD in children with T1D, the International Society for Pediatric and Adolescent Diabetes Consensus Clinical Guidelines recommend screening for thyroid function by analyzing circulating TSH at the time of diabetes diagnosis and every 2nd year thereafter in asymptomatic individuals without goiter, and with more frequent screening in patients with goiter present [59].
To characterize the T1D patients with coexisting AITD (referred to as autoimmune polyendocrine syndrome type 3 variant, APS3v), we analyzed their clinical characteristics compared to those without AITD [60]. Patients with APS3v demonstrated a significant female predominance, a slower and older age of onset for T1D, and a higher prevalence and level of GADA. Furthermore, among patients with Graves’ disease, 60% developed the disease before T1D, 30% had antecedent T1D, and 10% developed T1D and Graves’ disease simultaneously. The interval between the onset of T1D and Graves’ disease was <10 years in most cases but approached or exceeded 20 years in some instances.
The prevalence of GADA in nondiabetic patients with ATID is 8-10% [61,62,63], and it has been reported that GADA positivity is associated with a decreased insulin secretion capacity [64], suggesting that GADA positivity could be a marker of subclinical insulitis. Furthermore, the incidence of T1D/SPIDDM in anti-islet autoantibody-positive patients with Graves' disease has been reported to be 2.5 times higher than in autoantibody-negative patients [65]. The exact reasons why the level of GADA in T1D patients with AITD is extremely high compared to patients without AITD remain unclear. Previous studies have reported that GAD65 is expressed in the thyroid gland [66,67], and T1D patients with AITD demonstrated higher polyclonality in GADA epitope recognition and IgG subclasses compared to patients without AITD (Figure 5). Forty-eight percent of T1D patients with AITD recognized GAD65 N- terminal region, middle domain, and C-terminal region epitopes, while the same was true of only 9% of those without AITD (P<0.005). Additionally, the prevalence of patients with two or more IgG subclasses was significantly higher in T1D patients with AITD than those without AITD (57% vs. 17%, P<0.001). Therefore, the overproduction of GADA in T1D patients with AITD might be attributable to the activation of polyclonal B-lymphocyte response by GAD in the thyroid gland [61].
8. Recent Advances in Anti-Islet Autoantibody Assay
As described in the previous section, ICA is detected by immunohistochemistry using indirect immunofluorescence or immunoenzymatic techniques on frozen sections of human blood group O pancreas. Achieving concordance of results obtained from different laboratories is essential for comparing studies from various centers. To address this issue, a series international workshops on the standardization of ICA was initiated in 1985 under the auspices of the Immunology and Diabetes Workshop (currently Immunology and Diabetes Society). However, wide variations in results persisted despite using the same experimental protocol [68]. Therefore, subsequent workshops employed standard curves constructed from a reference serum (JDRF standard), which led to a decrease in interlaboratory variation [69].
Following the discovery of GAD as a major target antigen against ICA, numerous recombinant anti-islet autoantibody assays, including radioligand binding assays (RBA), RIA, and ELISA, have been developed. With these assays, the reported prevalence of anti-islet autoantibodies such as GADA, in patients with recently diagnosed T1D ranged widely from 25% [70] to 80% [71,72]. To standardize the recombinant anti-islet autoantibody assays, the first GADA workshop was held in 1993. In contrast to the ICA workshop, considerable concordance in ranking GADA levels in 16 different samples was observed among various assay formats [73]. In the second GADA workshop, which used a large number of T1D and control sera, the RBA format demonstrated greater sensitivity than conventional ELISA [74]. Solid-phase ELISA formats can detect insulin-binding antibodies in patients receiving insulin injections. However, these assays fail to detect IAA associated with disease risk., likely due to the high apparent affinity (1010) and extremely low capacity (10-12) of prediabetes autoantibodies [23]. Furthermore, the widely used fluid-phase RBA has proved challenging for many laboratories to implement. In response to the need for improved anti-islet autoantibody assays, a nonradioactive fluid-phase assay has been developed that offers high sensitivity and specificity. This assay employs a modified ELISA format, which is based on the autoantibodies’ ability to form a bridge between recombinant autoantigens (such as GAD, IA-2, and ZnT8) coated on the ELISA plate and biotin-labeled corresponding autoantigens, allowing for the detection of autoantibody-bound antigens rather than immunoglobulins themselves [75]. Using the same assay format, an anti-islet autoantibody assay has recently been developed that allows for the simultaneous measurement of multiple autoantibodies in a single well (3 Screen ICA ELISA) [76]. Although this assay cannot distinguish which of the three autoantibodies are present, it may serve as a useful screening test for T1D and facilitate the efficient diagnosis of immune-mediated T1D. Other sensitive high-throughput assays effective for large-scale screening include a multiplex ECL assay and a multiplex agglutination- PCR autoantibody assay that combines all four biochemically defined anti-islet autoantibodies [77,78]. These innovations coming from around the globe will enhance our ability to identify high-risk individuals for T1D more accurately and efficiently at an early stage, promoting the advancement of early interventions for the benefit of public health.
9. Conclusions
This article focused on reviewing the current understanding of anti-islet autoantibodies in T1D.The most important goal of diabetes management is to improve the quality of life and extend the lifespan by preventing micro- and macro-vascular diseases. To achieve this goal, it is crucial to maintain good glycemic control and residual β-cell function. Over the past 30 years, while numerous clinical trials regarding primary prevention have been conducted, to date, no treatment or agent has been proven effective in preventing T1D in susceptible individuals. However, several therapeutic agents have demonstrated usefulness in delaying progression from stage 2 to stage 3 T1D while also preserving β-cell function in stage 3. These agents include abatacept (anti-CTLA-4), alefacept (a fusion protein of soluble lymphocyte function antigen with Fc fragments of IgG1), rituximab (anti-CD20), and anti-thymocyte globulin, which preserve C-peptide production compared with placebo in stage 3 T1D [79,80,81,82]. Moreover, teplizumab (anti-CD3) has been shown to delay progression from stage 2 to stage 3 T1D by a median of 3 years [83,84]. Based on these results, in November 2022, the U.S. Food and Drug Administration approved teplizumab as the first drug to delay progression from stage 2 to stage 3 T1D in adults and children > 8 years. Since the number of positive anti-islet autoantibodies was primarily used for selecting high-risk individuals in these studies, the ability to assess multiple autoantibodies in a single test should prove valuable for future interventional trials.
The clinical utilities of anti-islet autoantibodies in patients with diabetes include diagnosis (immune-mediated or idiopathic), prediction (progressor or non-progressor) and understanding of pathophysiology (insulitis-specific or nonspecific phenomenon) (Figure 6). Since the autoantibody level of anti-islet autoantibodies decreases with disease duration and can become negative, it is essential to measure them early in the onset of T1D for accurate diagnosis. SPIDDM or LADA is often indistinguishable from T2D; therefore, earlier measurement of anti- islet autoantibodies is of great clinical importance for early diagnosis and appropriate treatment. In addition to the anti-islet autoantibody profiles, age of onset and genetic risk score should also be considered for risk triage. Furthermore, the development of a high-throughput assay to detect epitope-specific or immunoglobulin isotype-specific autoantibodies should warrant accurate diagnosis and prediction of autoimmune disorders.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
List of Abbreviations
| ADA | American Diabetes Association |
| AITD | autoimmune thyroid disease |
| APS3v | autoimmune polyendocrine syndrome type 3 variant |
| ECL | electrochemiluminescence |
| ELISA | enzyme-linked immunosorbent assay |
| GAD | glutamic acid decarboxylase |
| IA-2 | tyrosine phosphatase-like protein IA-2 |
| IAA | insulin autoantibodies |
| ICA | islet cell antibodies |
| JDRF | Juvenile Diabetes Research Foundation |
| JM | juxta-membrane |
| LADA | latent-autoimmune diabetes in adults |
| MHC | major histocompatibility complex |
| PTP | protein tyrosine phosphatase |
| RBA | radio-ligand binding assay |
| RIA | radioimmunoassay |
| SPIDDM | slowly-progressive type 1 diabetes |
| T1D | type 1 diabetes |
| T2D | type 2 diabetes |
| UKPDS | United Kingdom Prospective Diabetes Study |
| ZnT8 | zinc transporter 8 |
References
- ElSayed, N.A.; Aleppo, G.; Aroda, V.R.; Bannuru, R.R.; Brown, F.M.; Bruemmer, D.; Collins, B.S.; Gaglia, J.L.; Hilliard, M.E.; Isaacs, D.; et al. 2. Classification and Diagnosis of Diabetes: Standards of Care in Diabetes—2023. Diabetes Care 2022, 46, S19–S40. [Google Scholar] [CrossRef]
- Roep, B.O.; Thomaidou, S.; van Tienhoven, R.; Zaldumbide, A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat. Rev. Endocrinol. 2020, 17, 150–161. [Google Scholar] [CrossRef]
- Karvonen, M.; Viik-Kajander, M.; Moltchanova, E.; Libman, I.; LaPorte, R.; Tuomilehto, J. Incidence of childhood type 1 diabetes worldwide. Diabetes Mondiale (DiaMond) Project Group. . Diabetes Care 2000, 23, 1516–1526. [Google Scholar] [CrossRef]
- Kawasaki, E.; Matsuura, N.; Eguchi, K. Type 1 diabetes in Japan. Diabetologia 2006, 49, 828–836. [Google Scholar] [CrossRef]
- Bottazzo, G.F.; Florin-Christensen, A.; Doniach, D. Islet-cell antibodies in diabetes mellitus with autoimmune polyendocrine deficiencies. Lancet 1974, 2, 1279–1283. [Google Scholar] [CrossRef]
- Baekkeskov, S.; Nielsen, J.H.; Marner, B.; Bilde, T.; Ludvigsson, J.; Lernmark, A. Autoantibodies in newly diagnosed diabetic children immunoprecipitate human pancreatic islet cell proteins. Nature 1982, 298, 167–169. [Google Scholar] [CrossRef]
- Palmer, J.P.; Asplin, C.M.; Clemons, P.; Lyen, K.; Tatpati, O.; Raghu, P.K.; Paquette, T.L. Insulin Antibodies in Insulin-Dependent Diabetics Before Insulin Treatment. Science 1983, 222, 1337–1339. [Google Scholar] [CrossRef]
- Baekkeskov, S.; Aanstoot, H.-J.; Christgai, S.; Reetz, A.; Solimena, M.; Cascalho, M.; Folli, F.; Richter-Olesen, H.; Camilli, P.-D. Identification of the 64K autoantigen in insulin-dependent diabetes as the GABA-synthesizing enzyme glutamic acid decarboxylase. Nature 1990, 347, 151–156. [Google Scholar] [CrossRef]
- Lan, M.S.; Lu, J.; Goto, Y.; Notkins, A.L. Molecular cloning and identification of a receptor- type protein tyrosine phosphatase, IA-2, from human insulinoma. DNA Cell Biol. 1994, 13, 505–14. [Google Scholar] [CrossRef]
- Wenzlau, J.M.; Juhl, K.; Yu, L.; Moua, O.; Sarkar, S.A.; Gottlieb, P.; Rewers, M.; Eisenbarth, G.S.; Jensen, J.; Davidson, H.W.; et al. The cation efflux transporter ZnT8 (Slc30A8) is a major autoantigen in human type 1 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 17040–17045. [Google Scholar] [CrossRef]
- Karvonen, M.; Viik-Kajander, M.; Moltchanova, E.; Libman, I.; LaPorte, R.; Tuomilehto, J. Incidence of childhood type 1 diabetes worldwide. Diabetes Mondiale (DiaMond) Project Group. Diabetes Care 2000, 23, 1516–1526. [Google Scholar] [CrossRef]
- Ziegler, A.G.; Rewers, M.; Simell, O.; Simell, T.; Lempainen, J.; Steck, A.; Winkler, C.; Ilonen, J.; Veijola, R.; Knip, M.; et al. Seroconversion to Multiple Islet Autoantibodies and Risk of Progression to Diabetes in Children. JAMA 2013, 309, 2473–2479. [Google Scholar] [CrossRef]
- Krischer, J.P.; the Type 1 Diabetes TrialNet Study Group. The use of intermediate endpoints in the design of type 1 diabetes prevention trials. Diabetologia 2013, 56, 1919–1924. [Google Scholar] [CrossRef]
- Verge, C.F.; Gianani, R.; Kawasaki, E.; Yu, L.; Pietropaolo, M.; Jackson, R.A.; Chase, H.P.; Eisenbarth, G.S. Prediction of type 1 diabetes in first-degree relatives using a combination of insulin, GAD, and ICA512bdc/IA-2 autoantibodies. Diabetes 1996, 45, 926–933. [Google Scholar] [CrossRef]
- Yu, L.; Rewers, M.; Gianani, R.; Kawasaki, E.; Zhang, Y.; Verge, C.; Chase, P.; Klingensmith, G.; Erlich, H.; Norris, J.; et al. Antiislet autoantibodies usually develop sequentially rather than simultaneously. J. Clin. Endocrinol. Metab. 1996, 81, 4264–4267. [Google Scholar] [CrossRef]
- Nakamura, K.; Kawasaki, E.; Abiru, N.; Jo, O.; Fukushima, K.; Satoh, T.; Kuriya, G.; Kobayashi, M.; Kuwahara, H.; Yamasaki, H.; et al. Trajectories of anti-islet autoantibodies before development of type 1 diabetes in interferon-treated hepatitis C patients. Case reports and a literature review. Endocr. J. 2010, 57, 947–951. [Google Scholar] [CrossRef]
- Kawasaki, E.; Yu, L.; Rewers, M.J.; Hutton, J.C.; Eisenbarth, G.S. Definition of multiple ICA512/phogrin autoantibody epitopes and detection of intramolecular epitope spreading in relatives of patients with type 1 diabetes. Diabetes 1998, 47, 733–742. [Google Scholar] [CrossRef]
- Kawasaki, E.; Yasui, J.-I.; Tsurumaru, M.; Takashima, H.; Ikeoka, T.; Mori, F.; Akazawa, S.; Ueki, I.; Kobayashi, M.; Kuwahara, H.; et al. Sequential elevation of autoantibodies to thyroglobulin and glutamic acid decarboxylase in type 1 diabetes. World J. Diabetes 2013, 4, 227–230. [Google Scholar] [CrossRef]
- Kawasaki, E.; Nakamura, K.; Kuriya, G.; Satoh, T.; Kobayashi, M.; Kuwahara, H.; Abiru, N.; Yamasaki, H.; Matsuura, N.; Miura, J.; et al. Differences in the humoral autoreactivity to zinc transporter 8 between childhood- and adult-onset type 1 diabetes in Japanese patients. Clin. Immunol. 2011, 138, 146–153. [Google Scholar] [CrossRef]
- Achenbach, P.; Lampasona, V.; Landherr, U.; Koczwara, K.; Krause, S.; Grallert, H.; Winkler, C.; Pflüger, M.; Illig, T.; Bonifacio, E.; et al. Autoantibodies to zinc transporter 8 and SLC30A8 genotype stratify type 1 diabetes risk. Diabetologia 2009, 52, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Wenzlau, J.M.; Frisch, L.M.; Hutton, J.C.; Fain, P.R.; Davidson, H.W. Changes in Zinc Transporter 8 Autoantibodies Following Type 1 Diabetes Onset: The Type 1 Diabetes Genetics Consortium Autoantibody Workshop. Diabetes Care 2015, 38, S14–S20. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, I.; Weets, I.; Asanghanwa, M.; Ruige, J.; Van Gaal, L.; Mathieu, C.; Keymeulen, B.; Lampasona, V.; Wenzlau, J.M.; Hutton, J.C.; et al. Contribution of Antibodies Against IA-2β and Zinc Transporter 8 to Classification of Diabetes Diagnosed Under 40 Years of Age. Diabetes Care 2011, 34, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Castaño, L.; Ziegler, A.G.; Ziegler, R.; Shoelson, S.; Eisenbarth, G.S. Characterization of insulin autoantibodies in relatives of patients with type I diabetes. Diabetes 1993, 42, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Padoa, C.J.; Crowther, N.J.; Thomas, J.W.; Hall, T.R.; Bekris, L.M.; Torn, C.; Landin-Olsson, M.; Ortqvist, E.; Palmer, J.P.; Lernmark, A.; et al. Epitope analysis of insulin autoantibodies using recombinant Fab. Clin. Exp. Immunol. 2005, 140, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.R.; Thomas, J.W.; Padoa, C.J.; Torn, C.; Landin-Olsson, M.; Ortqvist, E.; Hampe, C.S. Longitudinal epitope analysis of insulin-binding antibodies in type 1 diabetes. Clin. Exp. Immunol. 2006, 146, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tree, T.I.; Morgenthaler, N.G.; Duhindan, N.; Hicks, K.E.; Madec, A.M.; Scherbaum, W.A.; Banga, J.P. Two amino acids in glutamic acid decarboxylase act in concert for maintenance of conformational determinants recognised by type 1 diabetic autoantibodies. Diabetologia 2000, 43, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.; Yano, M.; Abiru, N.; Akazawa, S.; Nagataki, S. Detection of recombinant GAD65 and GAD67 antibodies using a simple radioimmunoassay. Diabetes Res. Clin. Pr. 1996, 32, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Jayakrishnan, B.; Hoke, D.E.; Langendorf, C.G.; Buckle, A.M.; Rowley, M.J. An Analysis of the Cross-Reactivity of Autoantibodies to GAD65 and GAD67 in Diabetes. PLOS ONE 2011, 6, e18411. [Google Scholar] [CrossRef]
- Daw, K.; Powers, A.C. Two distinct glutamic acid decarboxylase auto-antibody specificities in IDDM target different epitopes. Diabetes 1995, 44, 216–220. [Google Scholar] [CrossRef]
- Hampe, C.S.; Hammerle, L.P.; Bekris, L.; Örtqvist, E.; Kockum, I.; Rolandsson, O.; Landin-Olsson, M.; Törn, C.; Persson, B.; Lernmark, A. Recognition of Glutamic Acid Decarboxylase (GAD) by Autoantibodies from Different GAD Antibody-Positive Phenotypes1. J. Clin. Endocrinol. Metab. 2000, 85, 4671–4679. [Google Scholar] [CrossRef]
- Kawasaki, E.; Abiru, N.; Ide, A.; Sun, F.; Fukushima, T.; Takahashi, R.; Kuwahara, H.; Fujita, N.; Kita, A.; Oshima, K.; et al. Epitope analysis of GAD65 autoantibodies in Japanese patients with autoimmune diabetes. Ann. New York Acad. Sci. 2003, 1005, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, M.; Banga, J.P.; Madec, A.M.; Binder, K.A.; Strebelow, M.; Rjasanowski, I.; Wassmuth, R.; Gilliam, L.K.; Luo, D.; Hampe, C.S. Dynamic changes of GAD65 autoantibody epitope specificities in individuals at risk of developing type 1 diabetes. Diabetologia 2005, 48, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Rabin, D.U.; Pleasic, S.M.; Palmer-Crocker, R.; Shapiro, J.A. Cloning and expression of IDDM- specific human autoantigens. Diabetes 1992, 41, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.; Hutton, J.C.; Eisenbarth, G.S. Molecular Cloning and Characterization of the Human Transmembrane Protein Tyrosine Phosphatase Homologue, Phogrin, an Autoantigen of Type 1 Diabetes. Biochem. Biophys. Res. Commun. 1996, 227, 440–447. [Google Scholar] [CrossRef]
- Harashima, S.-I.; Clark, A.; Christie, M.R.; Notkins, A.L. The dense core transmembrane vesicle protein IA-2 is a regulator of vesicle number and insulin secretion. Proc. Natl. Acad. Sci. 2005, 102, 8704–8709. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Hirai, H.; Zhang, G.; Zhang, M.; Takahashi, N.; Kasai, H.; Satin, L.S.; Leapman, R.D.; Notkins, A.L. Deletion of Ia-2 and/or Ia-2β in mice decreases insulin secretion by reducing the number of dense core vesicles. Diabetologia 2011, 54, 2347–2357. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.; Yu, L.; Gianani, R.; Verge, C.F.; Babu, S.; Bonifacio, E.; Eisenbarth, G.S. Evaluation of islet cell antigen (ICA) 512/IA-2 autoantibody radioassays using overlapping ICA512/IA-2 constructs. J Clin Endocrinol Metab. 1997, 82, 375–380. [Google Scholar] [CrossRef]
- Acevedo-Calado, M.; James, E.A.; Morran, M.P.; Pietropaolo, S.L.; Ouyang, Q.; Arribas-Layton, D.; Songini, M.; Liguori, M.; Casu, A.; Auchus, R.J.; et al. Identification of Unique Antigenic Determinants in the Amino Terminus of IA-2 (ICA512) in Childhood and Adult Autoimmune Diabetes: New Biomarker Development. Diabetes Care 2017, 40, 561–568. [Google Scholar] [CrossRef]
- Kawasaki, E.; Sera, Y.; Fujita, N.; Yamauchi, M.; Ozaki, M.; Abe, T.; Yamakawa, K.; Uotani, S.; Takino, H.; Yamasaki, H.; Yamaguchi, Y.; Uchigata, Y.; Matsuura, N.; Eguchi, K. Association between IA-2 autoantibody epitope specificities and age of onset in Japanese patients with autoimmune diabetes. J Autoimmun. 2001, 17, 323–331. [Google Scholar] [CrossRef]
- Dwivedi, O.P.; Lehtovirta, M.; Hastoy, B.; Chandra, V.; Krentz, N.A.J.; Kleiner, S.; Jain, D.; Richard, A.-M.; Abaitua, F.; Beer, N.L.; et al. Loss of ZnT8 function protects against diabetes by enhanced insulin secretion. Nat. Genet. 2019, 51, 1596–1606. [Google Scholar] [CrossRef]
- Nicolson, T.J.; Bellomo, E.A.; Wijesekara, N.; Loder, M.K.; Baldwin, J.M.; Gyulkhandanyan, A.V.; Koshkin, V.; Tarasov, A.I.; Carzaniga, R.; Kronenberger, K.; et al. Insulin Storage and Glucose Homeostasis in Mice Null for the Granule Zinc Transporter ZnT8 and Studies of the Type 2 Diabetes–Associated Variants. Diabetes 2009, 58, 2070–2083. [Google Scholar] [CrossRef] [PubMed]
- Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Shen, L.; Serre, D.; Boutin, P.; Vincent, D.; Belisle, A.; Hadjadj, S.; et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature 2007, 445, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Wenzlau, J.M.; Liu, Y.; Yu, L.; Moua, O.; Fowler, K.T.; Rangasamy, S.; Walters, J.; Eisenbarth, G.S.; Davidson, H.W.; Hutton, J.C. A Common Nonsynonymous Single Nucleotide Polymorphism in the SLC30A8 Gene Determines ZnT8 Autoantibody Specificity in Type 1 Diabetes. Diabetes 2008, 57, 2693–2697. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.; Uga, M.; Nakamura, K.; Kuriya, G.; Satoh, T.; Fujishima, K.; Ozaki, M.; Abiru, N.; Yamasaki, H.; Wenzlau, J.M.; Davidson, H.W.; Hutton, J.C.; Eguchi, K. Association between anti- ZnT8 autoantibody specificities and SLC30A8 Arg325Trp variant in Japanese patients with type 1 diabetes. Diabetologia. 2008, 51, 2299–2302. [Google Scholar] [CrossRef] [PubMed]
- Wenzlau, J.M.; Frisch, L.M.; Hutton, J.C.; Davidson, H.W. Mapping of conformationalautoantibody epitopes in ZnT8. Diabetes Metab Res Rev. 2011, 27, 883–886. [Google Scholar] [CrossRef]
- Buzzetti, R.; Tuomi, T.; Mauricio, D.; Pietropaolo, M.; Zhou, Z.; Pozzilli, P.; Leslie, R.D. Management of Latent Autoimmune Diabetes in Adults: A Consensus Statement From an International Expert Panel. Diabetes 2020, 69, 2037–2047. [Google Scholar] [CrossRef]
- Shimada, A.; Kawasaki, E.; Abiru, N.; Awata, T.; Oikawa, Y.; Osawa, H.; Kajio, H.; Ozawa, J.; Takahashi, K.; Chujo, D.; Noso, S.; Fukui, T.; Miura, J.; Yasuda, K.; Yasuda, H.; Imagawa, A.; Ikegami, H. New diagnostic criteria (2023) of slowly progressive type 1 diabetes (SPIDDM) -Report from Committee of type 1 diabetes in Japan Diabetes Society-. J Japan Diab Soc. in press (in Japanese)
- Kasuga, A.; Maruyama, T.; Nakamoto, S.; Ozawa, Y.; Suzuki, Y.; Saruta, T. High-Titer Autoantibodies against Glutamic Acid Decarboxylase Plus Autoantibodies against Insulin and IA-2 Predicts Insulin Requirement in Adult Diabetic Patients. J. Autoimmun. 1999, 12, 131–135. [Google Scholar] [CrossRef]
- Takino, H.; Yamasaki, H.; Abiru, N.; Sera, Y.; Abe, T.; Kawasaki, E.; Yamaguchi, Y.; Eguchi, K.; Kanazawa, Y.; Nagataki, S. Antibodies to GAD in Japanese patients classified as Type 2 diabetes at diagnosis. High titre of GADAb is a predictive marker for early insulin treatment--report of west Japan (Kyushu, Yamaguchi, Osaka) study for GAD Ab(+) diabetes. Diabet Med 2002, 19, 730–734. [Google Scholar] [CrossRef]
- Kawasaki, E.; Nakamura, K.; Kuriya, G.; Satoh, T.; Kuwahara, H.; Kobayashi, M.; Abiru, N.; Yamasaki, H.; Eguchi, K. Autoantibodies to Insulin, Insulinoma-Associated Antigen-2, and Zinc Transporter 8 Improve the Prediction of Early Insulin Requirement in Adult-Onset Autoimmune Diabetes. J. Clin. Endocrinol. Metab. 2010, 95, 707–713. [Google Scholar] [CrossRef]
- Turner, R.; Stratton, I.; Horton, V.; Manley, S.; Zimmet, P.; Mackay, I.R.; Shattock, M.; Bottazzo, G.F.; Holman, R. UKPDS 25: autoantibodies to islet-cell cytoplasm and glutamic acid decarboxylase for prediction of insulin requirement in type 2 diabetes. Lancet 1997, 350, 1288–1293. [Google Scholar] [CrossRef]
- Yasui, J.; Kawasaki, E.; Tanaka, S.; Awata, T.; Ikegami, H.; Imagawa, A.; Uchigata, Y.; Osawa, H.; Kajio, H.; Kawabata, Y.; et al. Clinical and Genetic Characteristics of Non-Insulin-Requiring Glutamic Acid Decarboxylase (GAD) Autoantibody-Positive Diabetes: A Nationwide Survey in Japan. PLOS ONE 2016, 11, e0155643–e0155643. [Google Scholar] [CrossRef]
- Nederstigt, C.; Uitbeijerse, B.S.; Janssen, L.G.M.; Corssmit, E.P.M.; de Koning, E.J.P.; Dekkers, O.M. Associated auto-immune disease in type 1 diabetes patients: a systematic review and meta-analysis. Eur. J. Endocrinol. 2019, 180, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Kakleas, K.; Soldatou, A.; Karachaliou, F.; Karavanaki, K. Associated autoimmune diseases in children and adolescents with type 1 diabetes mellitus (T1DM). Autoimmun. Rev. 2015, 14, 781–797. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E. Type 1 diabetes and autoimmunity. Clin Pediatr Endocrinol 2014, 23, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kordonouri, O.; Klinghammer, A.; Lang, E.B.; Grüters-Kieslich, A.; Grabert, M.; Holl, R.W. Thyroid autoimmunity in children and adolescents with type 1 diabetes: a multicenter survey. Diabetes Care 2002, 25, 1346–1350. [Google Scholar] [CrossRef] [PubMed]
- Umpierrez, G.E.; Latif, K.A.; Murphy, M.B.; Lambeth, H.C.; Stentz, F.; Bush, A.; Kitabchi, A.E. Thyroid dysfunction in patients with type 1 diabetes: a longitudinal study. Diabetes Care 2003, 26, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Glastras, S.J.; Craig, M.E.; Verge, C.F.; Chan, A.K.; Cusumano, J.M.; Donaghue, K.C. The Role of Autoimmunity at Diagnosis of Type 1 Diabetes in the Development of Thyroid and Celiac Disease and Microvascular Complications. Diabetes Care 2005, 28, 2170–2175. [Google Scholar] [CrossRef]
- Kordonouri, O.; Klingensmith, G.; Knip, M.; Holl, R.W.; Aanstoot, H.-J.; Menon, P.S.; E Craig, M. Other complications and diabetes-associated conditions in children and adolescents. Pediatr. Diabetes 2014, 15, 270–278. [Google Scholar] [CrossRef]
- Horie, I.; Kawasaki, E.; Ando, T.; Kuwahara, H.; Abiru, N.; Usa, T.; Yamasaki, H.; Ejima, E.; Kawakami, A. Clinical and Genetic Characteristics of Autoimmune Polyglandular Syndrome Type 3 Variant in the Japanese Population. J. Clin. Endocrinol. Metab. 2012, 97, E1043–E1050. [Google Scholar] [CrossRef]
- Kawasaki, E.; Abiru, N.; Yano, M.; Uotani, S.; Matsumoto, K.; Matsuo, H.; Yamasaki, H.; Yamamoto, H.; Yamaguchi, Y.; Akazawa, S.; et al. Autoantibodies to Glutamic Acid Decarboxylase in Patients with Autoimmune Thyroid Disease: Relation to Competitive Insulin Autoantibodies. J. Autoimmun. 1995, 8, 633–643. [Google Scholar] [CrossRef]
- Balakhadze, M.; Giorgadze, E.; Lomidze, M. The frequency of Langerhans islets β-cells autoantibodies (Anti-GAD) in Georgian children and adolescents with chronic autoimmune thyroiditis. Int J Endocrinol. 2016, 2016, 6597091. [Google Scholar] [CrossRef] [PubMed]
- Maugendre, D.; Verite, F.; Guilhem, I.; Genetet, B.; Allannic, H.; Delamaire, M. Anti-pancreatic autoimmunity and Graves' disease: study of a cohort of 600 Caucasian patients. Eur. J. Endocrinol. 1997, 137, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Lethagen, Å.L.; Ericsson, U.B.; Hallengren, B.; Groop, L.; Tuomi, T. Glutamic acid decarboxylase antibody positivity is associated with an impaired insulin response to glucose and arginine in nondiabetic patients with autoimmune thyroiditis. J Clin Endocrinol Metab. 2002, 87, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Jonsdottir, B.; Jönsson, I.; Lantz, M. Prevalence of diabetes and presence of autoantibodies against zinc transporter 8 and glutamic decarboxylase at diagnosis and at follow up of Graves’ disease. Endocrine 2019, 64, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Erdö, S.L.; Wolff, J.R. γ-aminobutyric acid outside the mammalian brain. J Neurochem. 1990, 54, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.R.; Brown, T.J.; Cassidy, D. Binding of antibodies in sera from Type 1 (insulin- dependent) diabetic patients to glutamate decarboxylase from rat tissues. Evidence for antigenic and non-antigenic forms of the enzyme. Diabetologia 1992, 35, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Bottazzo, G.F.; Gleichmann, H. Immunology and Diabetes Workshops: report of the first international workshop on the standardisation of cytoplasmic islet cell antibodies. Diabetologia 1986, 29, 125–126. [Google Scholar] [CrossRef]
- Bonifacio, E.; Dawkins, R.L.; Lernmark, Å. Immunology and Diabetes Workshops: Report of the Second International Workshop on the Standardisation of Cytoplasmic Islet Cell Antibodies. Diabetologia 1987, 30, 273–273. [Google Scholar] [CrossRef]
- Martino, G.V.; Tappaz, M.L.; Braghi, S.; Dozio, N.; Canal, N.; Pozza, G.; Bottazzo, G.F.; Grimaldi, L.M.; Bosi, E. Autoantibodies to glutamic acid decarboxylase (GAD) detected by an immuno-trapping enzyme activity assay: Relation to insulin-dependent diabetes mellitus and islet cell antibodies. J. Autoimmun. 1991, 4, 915–923. [Google Scholar] [CrossRef]
- Rowley, M.J.; Mackay, I.R.; Chen, Q.Y.; Knowles, W.J.; Zimmet, P.Z. Antibodies to glutamic acid decarboxylase discriminate major types of diabetes mellitus. Diabetes. 1992, 41, 548–551. [Google Scholar] [CrossRef]
- Kawasaki, E.; Takino, H.; Yano, M.; Uotani, S.; Matsumoto, K.; Takao, Y.; Yamaguchi, Y.; Akazawa, S.; Nagataki, S. Autoantibodies to glutamic acid decarboxylase in patients with IDDM and autoimmune thyroid disease. Diabetes. 1994, 43, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Schmidli, R.S.; Colman, P.G.; Bonifacio, E.; Bottazzo, G.F.; Harrison, L.C. High level of concordance between assays for glutamic acid decarboxylase antibodies. The first international glutamic acid decarboxylase antibody workshop. Diabetes 1994, 43, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Schmidli, R.S.; Colman, P.G.; Bonifacio, E. Disease sensitivity and specificity of 52 assays for glutamic acid decarboxylase antibodies. The second international GADAb Workshop. Diabetes 1995, 44, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.; Tanaka, M.; Miwa, M.; Abiru, N.; Kawakami, A. Novel enzyme-linked immunosorbent assay for bivalent ZnT8 autoantibodies. Acta Diabetol. 2013, 51, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Raab, J.; Haupt, F.; Scholz, M.; Matzke, C.; Warncke, K.; Lange, K.; Assfalg, R.; Weininger, K.; Wittich, S.; Löbner, S.; et al. Capillary blood islet autoantibody screening for identifying pre-type 1 diabetes in the general population: design and initial results of the Fr1da study. BMJ Open 2016, 6, e011144. [Google Scholar] [CrossRef] [PubMed]
- Lind, A.; Cortez, F.d.J.; Ramelius, A.; Bennet, R.; Robinson, P.V.; Seftel, D.; Gebhart, D.; Tandel, D.; Maziarz, M.; Agardh, D.; et al. Multiplex agglutination-PCR (ADAP) autoantibody assays compared to radiobinding autoantibodies in type 1 diabetes and celiac disease. J. Immunol. Methods 2022, 506, 113265. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Yu, L. Effective assay technologies fit for large-scale population screening of type 1 diabetes. Front. Clin. Diabetes Heal. 2023, 3, 1034698. [Google Scholar] [CrossRef]
- Orban, T.; Bundy, B.; Becker, D.J.; DiMeglio, L.A.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Greenbaum, C.J.; Marks, J.B.; Monzavi, R.; et al. Costimulation Modulation With Abatacept in Patients With Recent-Onset Type 1 Diabetes: Follow-up 1 Year After Cessation of Treatment. Diabetes Care 2014, 37, 1069–1075. [Google Scholar] [CrossRef]
- Rigby, M.R.; Harris, K.M.; Pinckney, A.; DiMeglio, L.A.; Rendell, M.S.; Felner, E.I.; Dostou, J.M.; Gitelman, S.E.; Griffin, K.J.; Tsalikian, E.; et al. Alefacept provides sustained clinical and immunological effects in new-onset type 1 diabetes patients. J. Clin. Investig. 2015, 125, 3285–3296. [Google Scholar] [CrossRef]
- Pescovitz, M.D.; Greenbaum, C.J.; Krause-Steinrauf, H.; Becker, D.J.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Marks, J.B.; McGee, P.F.; Moran, A.M.; Raskin, P.; Rodriguez, H.; Schatz, D.A.; Wherrett, D.; Wilson, D.M.; Lachin, J.M.; Skyler, J.S. Rituximab, B-lymphocyte depletion, and preservation of β-cell function. N Engl J Med 2009, 361, 2143–52. [Google Scholar] [CrossRef]
- Haller, M.J.; Long, S.A.; Blanchfield, J.L.; Schatz, D.A.; Skyler, J.S.; Krischer, J.P.; Bundy, B.N.; Geyer, S.M.; Warnock, M.V.; Miller, J.L.; et al. Low-Dose Anti-Thymocyte Globulin Preserves C-Peptide, Reduces HbA1c, and Increases Regulatory to Conventional T-Cell Ratios in New-Onset Type 1 Diabetes: Two-Year Clinical Trial Data. Diabetes 2019, 68, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Herold, K.C.; Bundy, B.N.; Long, S.A.; Bluestone, J.A.; DiMeglio, L.A.; Dufort, M.J.; Gitelman, S.E.; Gottlieb, P.A.; Krischer, J.P.; Linsley, P.S.; Marks, J.B.; Moore, W.; Moran, A.; Rodriguez, H.; Russell, W.E.; Schatz, D.; Skyler, J.S.; Tsalikian, E.; Wherrett, D.K.; Ziegler, A.G.; Greenbaum, C.J.; et al. An anti-CD3 antibody, Teplizumab, in relatives at risk for type 1 diabetes. N Engl J Med 2019, 381, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Sims, E.K.; Bundy, B.N.; Stier, K.; Serti, E.; Lim, N.; Long, S.A.; Geyer, S.M.; Moran, A.; Greenbaum, C.J.; Evans-Molina, C.; et al. Teplizumab improves and stabilizes beta cell function in antibody-positive high-risk individuals. Sci. Transl. Med. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chronology of anti-islet autoantibody discovery. The history of anti-islet autoantibody discovery began in 1974 with the discovery of ICA. Currently, four biochemically define autoantibodies (IAA, GADA, IA-2A, and ZnT8A) are used for prediction and diagnose of T1D.
Figure 1.
Chronology of anti-islet autoantibody discovery. The history of anti-islet autoantibody discovery began in 1974 with the discovery of ICA. Currently, four biochemically define autoantibodies (IAA, GADA, IA-2A, and ZnT8A) are used for prediction and diagnose of T1D.
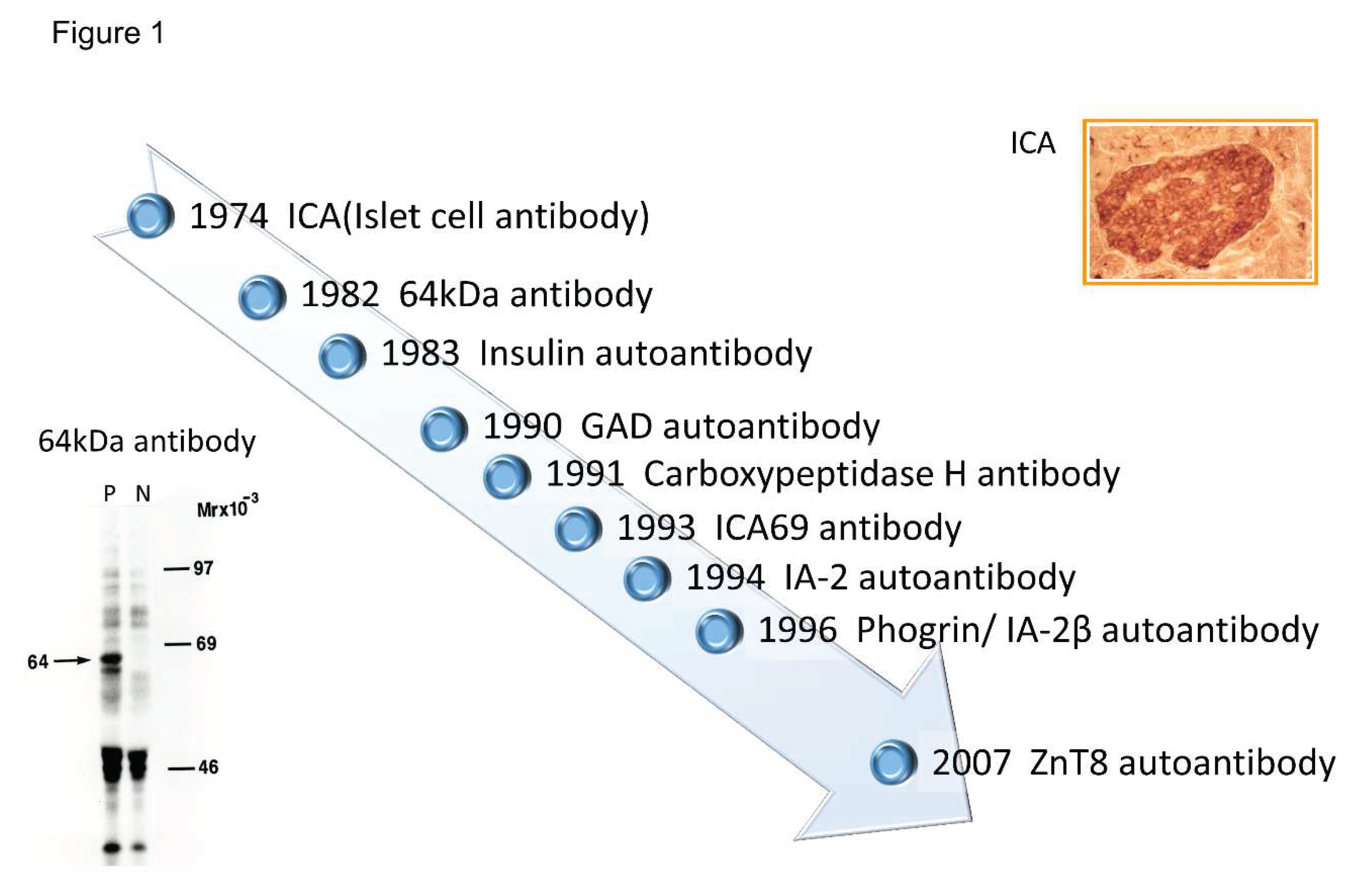
Figure 2.
Three stages of natural history of T1D. In Stage 1, autoimmunity to pancreatic islet is induced by cytotoxic T cells, and destruction of pancreatic β cells (insulitis) begins, although blood glucose levels are still within normal range. Two or more IAA, GADA, IA-2A, and ZnT8A emerges in this stage. In Stage 2, like Stage 1, individuals test two or more anti-islet autoantibodies and destruction of β-cell progresses to the development of glucose intolerance or dysglycemia. Stage 3 represents manifestations of the typical clinical symptoms and signs of T1D, including polyuria, polydipsia, weight loss, general fatigue, diabetic ketoacidosis, and others.
Figure 2.
Three stages of natural history of T1D. In Stage 1, autoimmunity to pancreatic islet is induced by cytotoxic T cells, and destruction of pancreatic β cells (insulitis) begins, although blood glucose levels are still within normal range. Two or more IAA, GADA, IA-2A, and ZnT8A emerges in this stage. In Stage 2, like Stage 1, individuals test two or more anti-islet autoantibodies and destruction of β-cell progresses to the development of glucose intolerance or dysglycemia. Stage 3 represents manifestations of the typical clinical symptoms and signs of T1D, including polyuria, polydipsia, weight loss, general fatigue, diabetic ketoacidosis, and others.
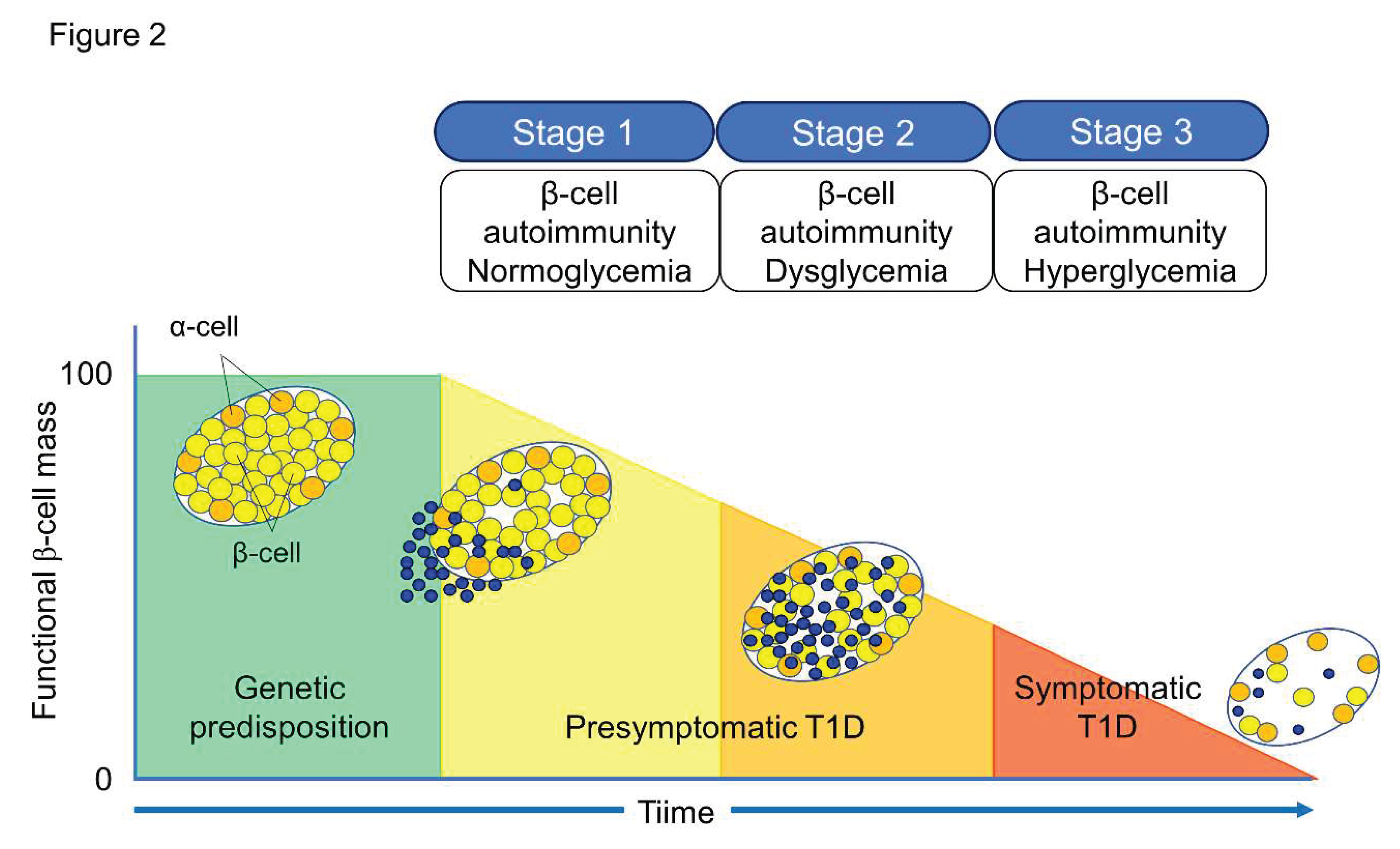
Figure 3.
Combinatorial analysis of anti-islet autoantibodies in patients with acute-onset T1D (A), SPIDDM (B), and fulminant T1D (C).
Figure 3.
Combinatorial analysis of anti-islet autoantibodies in patients with acute-onset T1D (A), SPIDDM (B), and fulminant T1D (C).
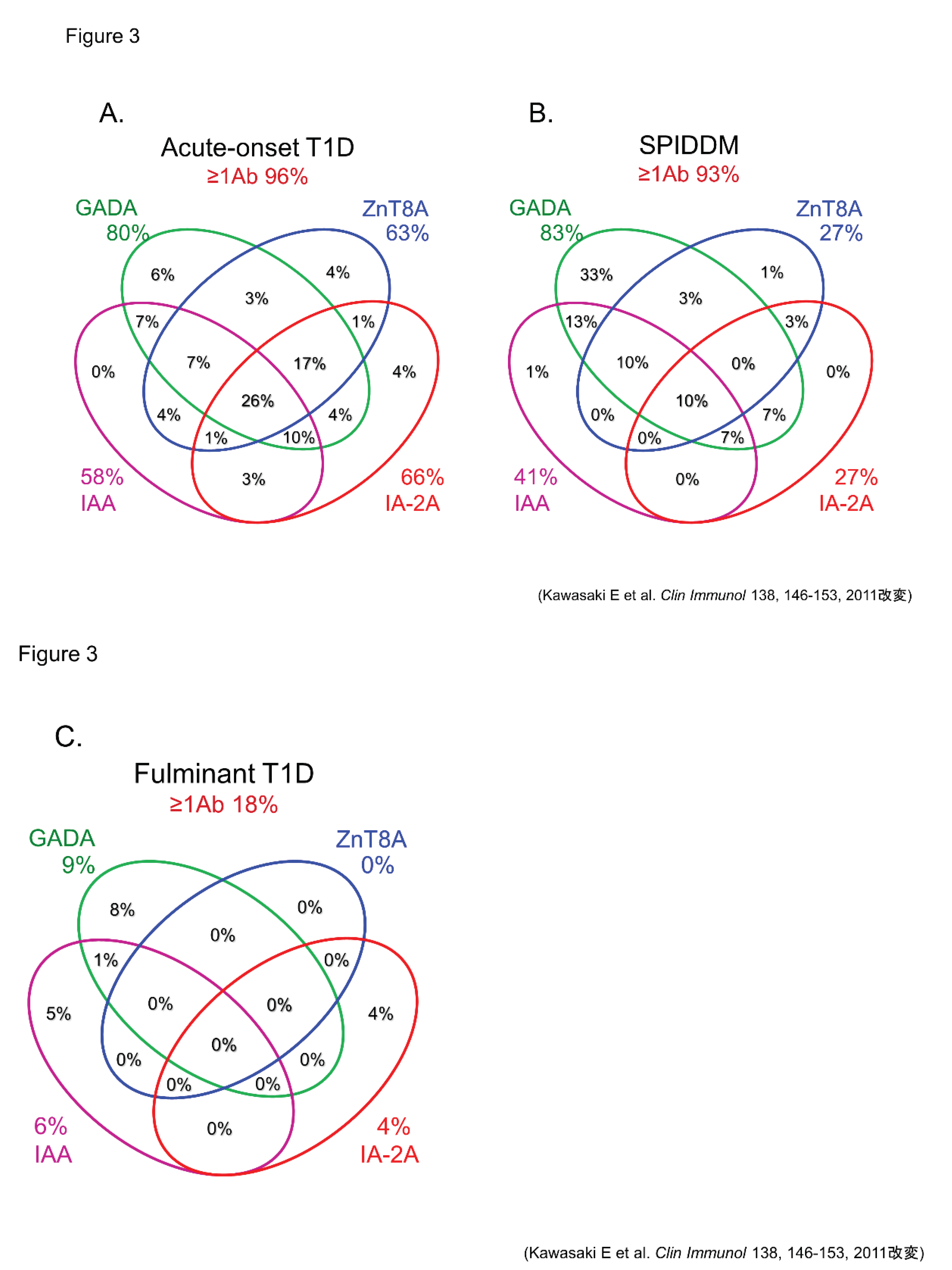
Figure 4.
Illustration of antigenic epitopes recognized by T1D sera in GAD65 (A), IA-2 (B), and ZnT8 (C) proteins.
Figure 4.
Illustration of antigenic epitopes recognized by T1D sera in GAD65 (A), IA-2 (B), and ZnT8 (C) proteins.
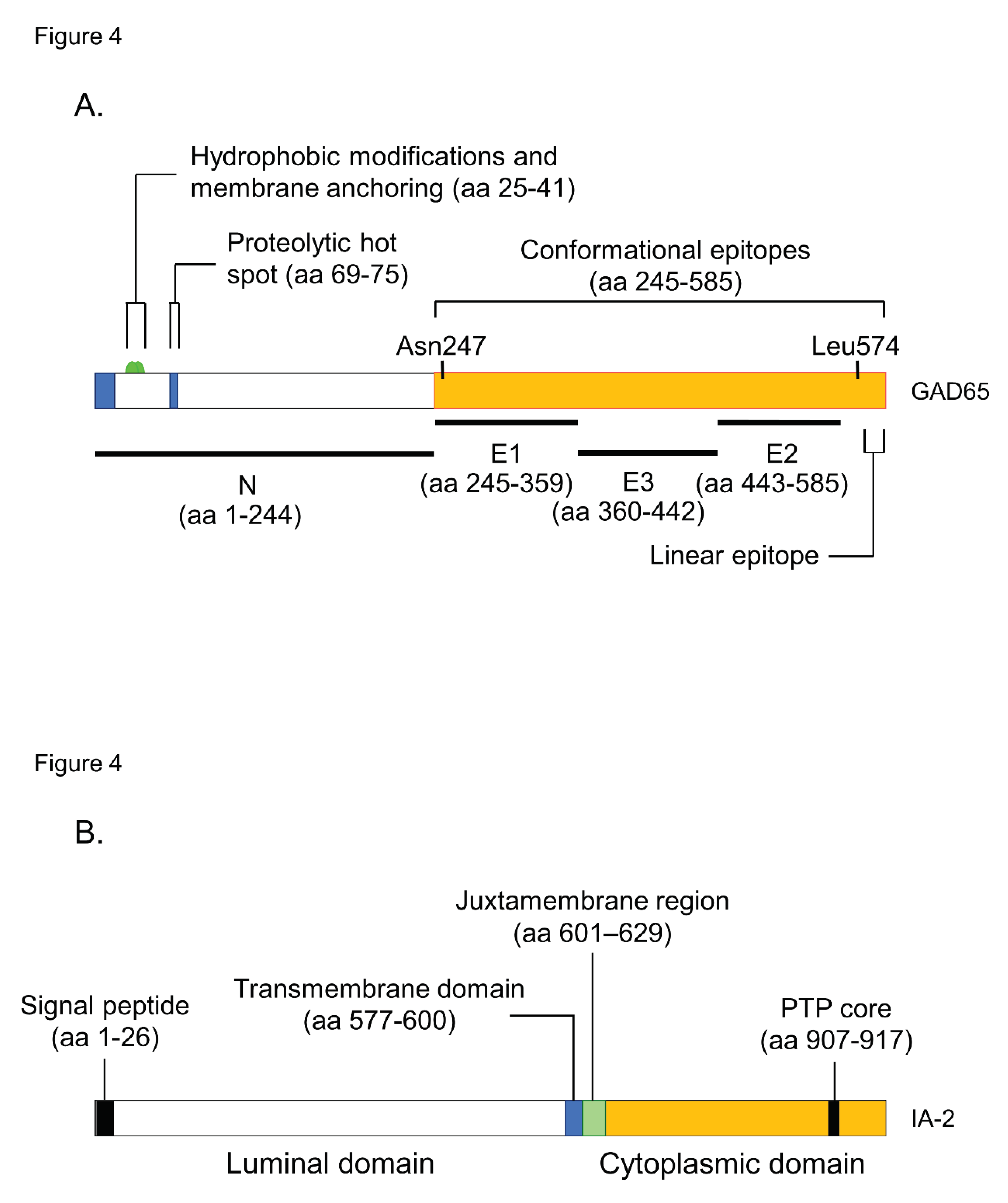
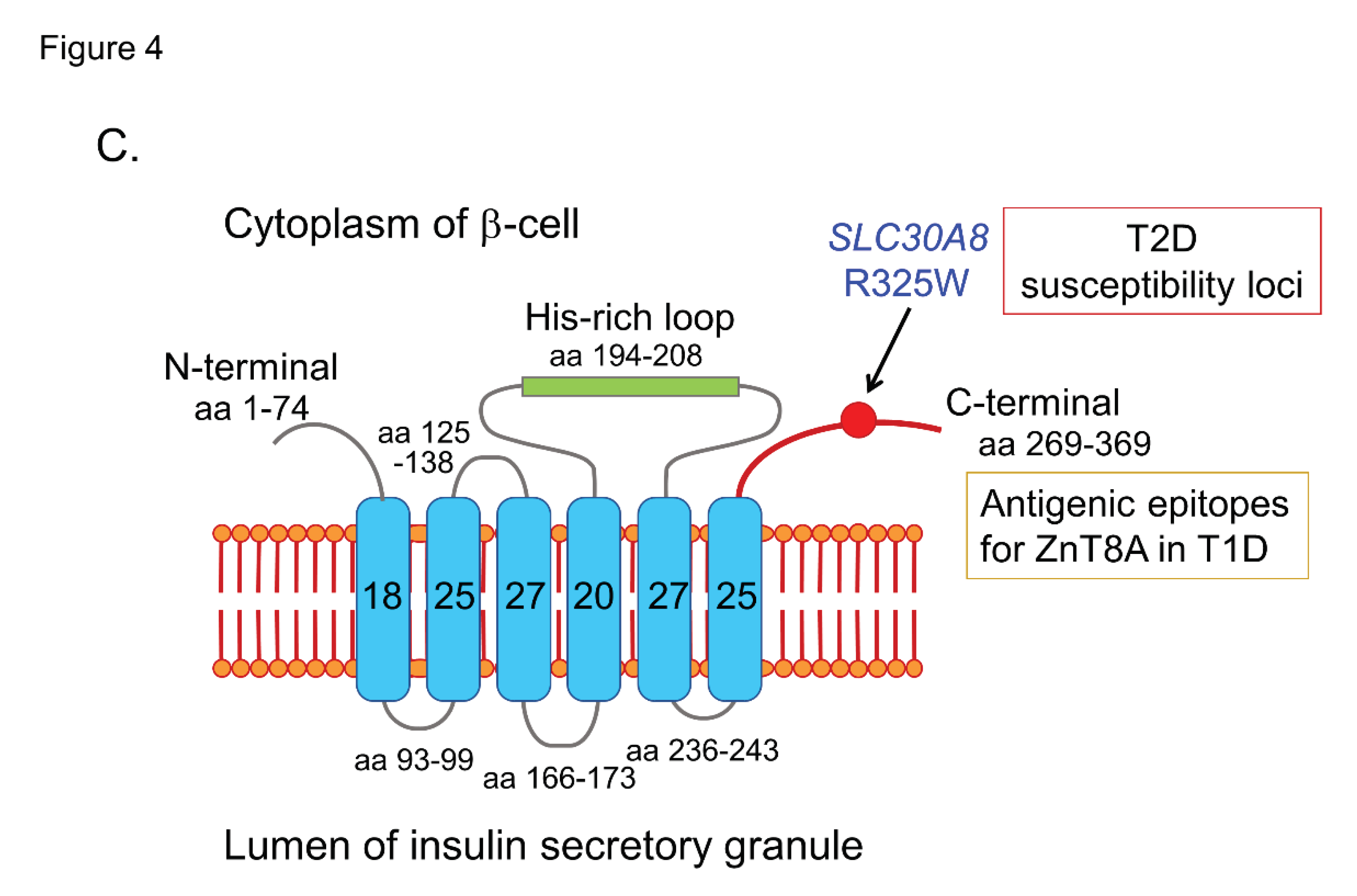
Figure 5.
Comparison of GADA epitopes (top) and IgG subclasses (bottom) in T1D patients without and with autoimmune thyroid disease Left panel: T1D without AITD; Right panel: T1D with AITD AITD, autoimmune thyroid disease.
Figure 5.
Comparison of GADA epitopes (top) and IgG subclasses (bottom) in T1D patients without and with autoimmune thyroid disease Left panel: T1D without AITD; Right panel: T1D with AITD AITD, autoimmune thyroid disease.
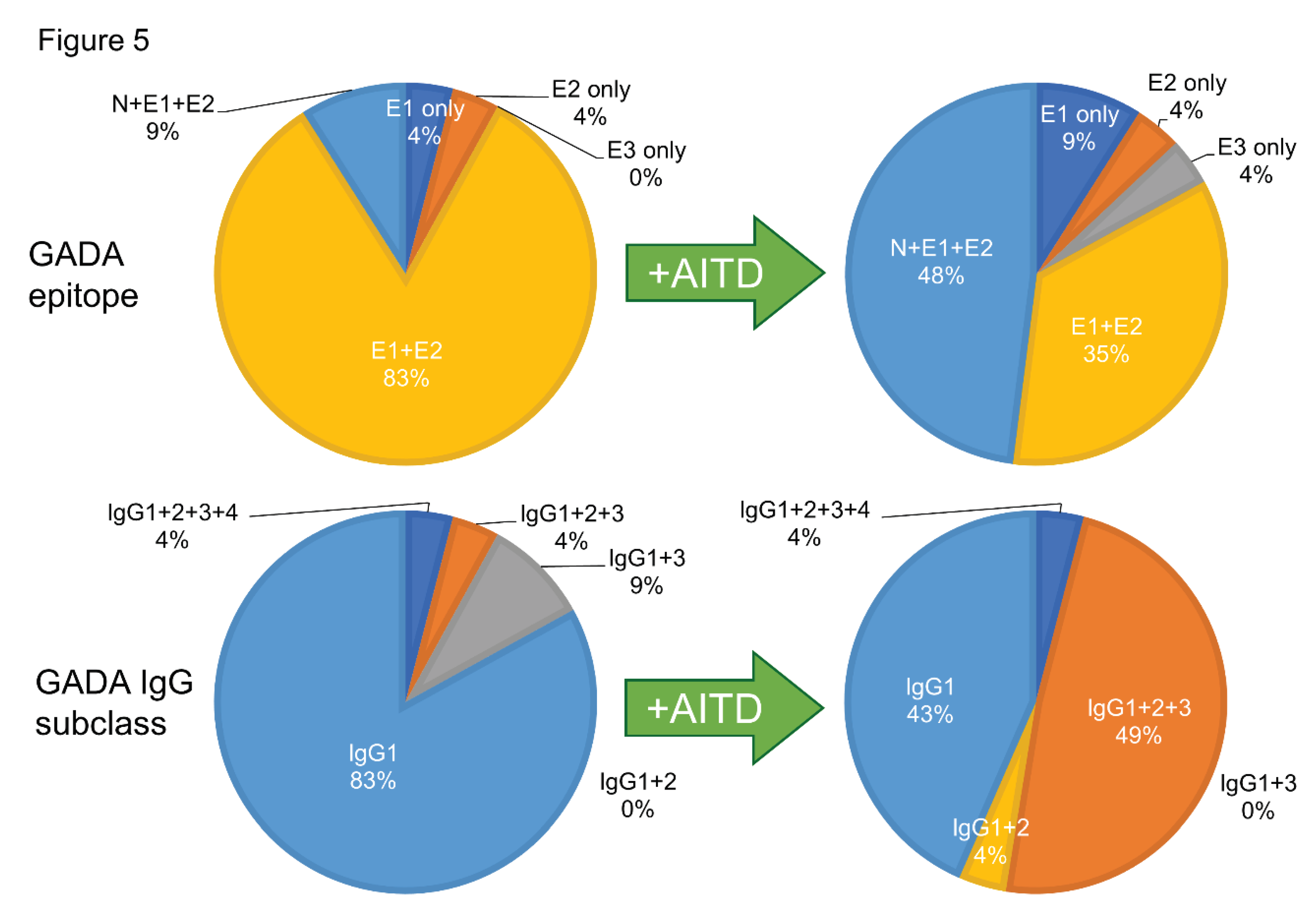
Figure 6.
Clinical utilities of anti-islet autoantibodies in patients with diabetes.
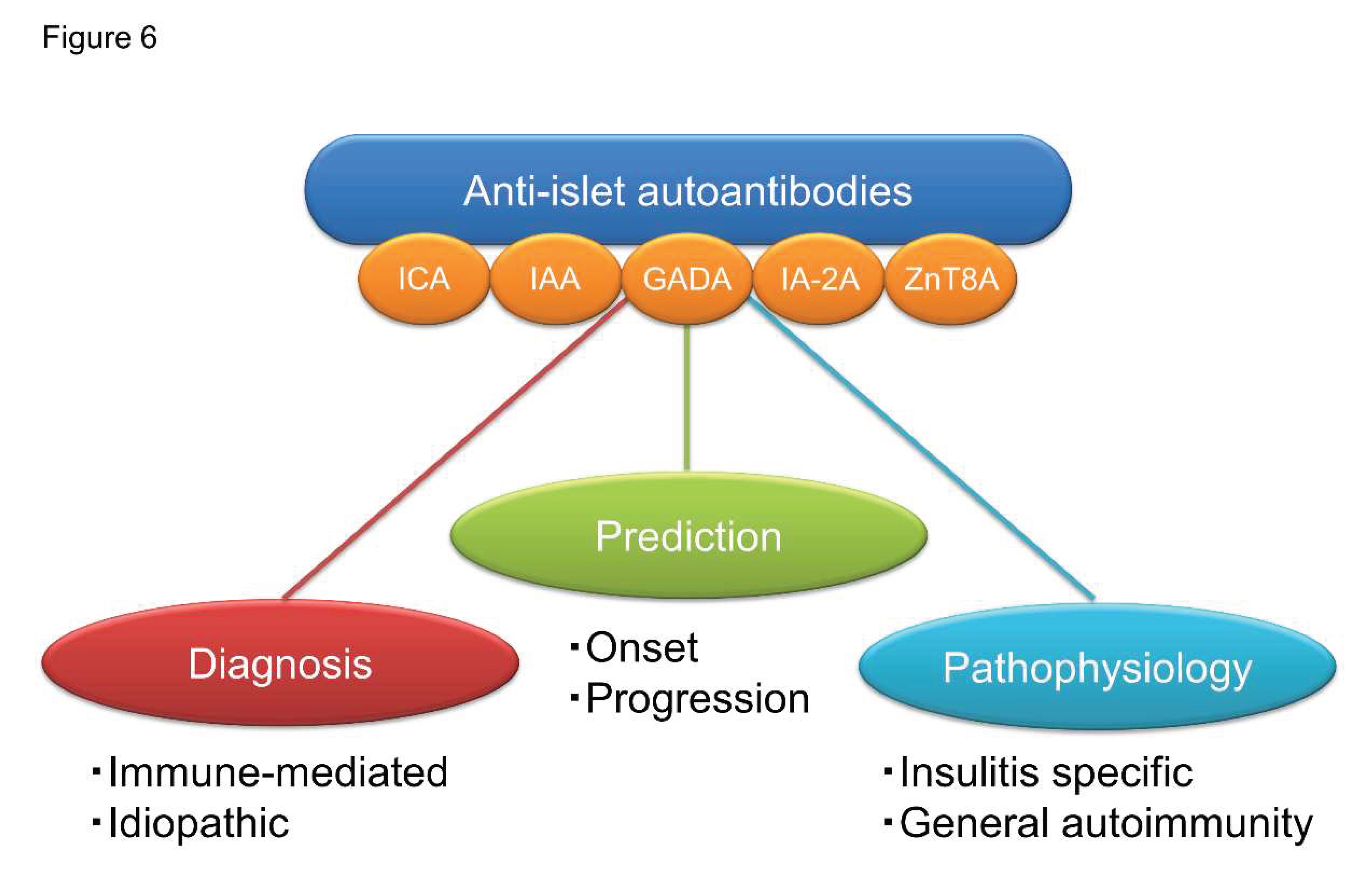

Table 1.
Disease specificity of GAD autoantibodies.
| Subject | Prevalence |
| Healthy control | <1% |
| Acute-onset type 1 diabetes (at onset) | 60~80% |
| Fulminant type 1 diabetes | 5~9% |
| LADA (SPIDDM) | 100% |
| Type 2 diabetes (diet/OHA) | 4~5% |
| Polyglandular autoimmune syndrome, type 1 | 30~40% |
| Polyglandular autoimmune syndrome, type 2 | 30~50% |
| Autoimmune thyroid disease | 6~8% |
| Stiff-person syndrome | 60~70% |
LADA, latent-autoimmune diabetes in adults; SPIDDM, slowly progressive insulin-dependent diabetes; OHA, oral hypoglycemic agents.
Table 2.
Diagnostic criteria (2023) of slowly progressive type 1 diabetes (SPIDDM).
| Required item: |
| 1) the presence of anti-islet autoantibodies at some time point during the diseasecourse a) |
| 2) the absence of ketosis or ketoacidosis at the diagnosis of diabetes and the unnecessity for insulin treatment to correct hyperglycemia immediately afterdiagnosis in principle |
| 3) gradual decrease of insulin secretion overtime, requirement of insulin treatment more than 3 months b) after diagnosis of diabetes, and exhausted endogenous insulin secretion (fasting serum C-peptide immunoreactivity < 0.6 ng/ml) at last observedtime point |
| Judgement: |
| when the case fulfills the criteria all of the three described above (1), 2), and 3)), the case is diagnosed with “slowly progressive type 1 diabetes (definite)”when the case fulfills the criteria only 1) and 2), but not 3), the case is diagnosed with“slowly progressive type 1 diabetes (probable)” |
| a) Anti-islet autoantibodies include glutamic acid decarboxylase (GAD) autoantibody, insulinoma-associated antigen-2 (IA-2) autoantibody, islet cell antibody (ICA), zinc transporter 8 (ZnT8) autoantibody or insulin autoantibody (IAA). The measurementof IAA should be performed before starting insulin treatment. |
| b) more than 6 months in typical case |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



