
Submitted:
05 May 2023
Posted:
06 May 2023
You are already at the latest version
Abstract
Vessel co-option correlates with resistance against anti-angiogenic agents and chemotherapy in colorectal cancer liver metastasis (CRCLM). We previously identified higher intensity of neutrophils in the tumour microenvironment of vessel co-opting CRCLM lesions compared to their angiogenic counterparts. Herein, we demonstrated that over 50% of the neutrophils in vessel co-opting lesions are expressing pro-apoptotic markers including cleaved caspase-3 and poly (ADP-ribose) polymerase-1 (PARP-1). Our previous publications suggested upregulation of transforming growth factor-beta (TGFβ1) in the microenvironment of vessel co-option CRCLM. Therefore, we examined the effect of TGFβ1 on the expression of cleaved caspase-3 and PARP-1 in neutrophils in vitro. Significantly, we noticed the upregulation of pro-apoptotic markers upon exposure to TGFβ1. This finding might pave the way to determine the role of neutrophils in developing vessel co-option in CRCLM in the future.
Keywords:
CRCLM
; vessel co-option
; angiogenesis
; neutrophil
; TGFβ1
; apoptosis
1. Introduction
Colorectal cancer (CRC) is the world’s third most deadly cancer, and 50% of CRC patients develop metastasis to the liver [1]. Colorectal cancer liver metastasis (CRCLM) presents in two main histopathological growth patterns (HGPs), and they have been identified as desmoplastic HGP (DHGP) and replacement HGP (RHGP) [2]. In DHGP lesions, a desmoplastic stromal ring separates the cancer cells from normal liver parenchyma. However, the cancer cells in RHGP lesions infiltrate through normal liver parenchyma and replace the hepatocytes near the tumour periphery [2]. Beyond the histology, the DHGP and RHGP tumours exist many differences in vascularization and the immune landscape [3]. In this context, the DHGP tumour are characterized by sprouting angiogenesis to obtain blood supply, while the RHGP tumours utilize vessel co-option vascularization [2]. In vessel co-option, the cancer cells hijack the pre-existing blood vessels to obtain blood supply and are not responsive to anti-angiogenic therapy [2,4]. Therefore, the DHGP tumours are known as angiogenic tumours, whereas the RHGP tumours are known as vessel co-opting tumours [2,5].
Neutrophils are the most abundant innate immune cells [6] and have multifaceted roles in cancer [7,8]. They can promote tumour cell proliferation and stimulate angiogenesis, also playing an essential role in matrix remodeling [7,9]. We previously reported higher levels of neutrophils in RHGP tumours compared to their angiogenic counterparts [10]. Moreover, we found runt related transcription factor-1 (RUNX1) mediates neutrophil infiltration in CRCLM through angiopoietin-1 (Ang1) [11]. Importantly, the neutrophils in RHGP lesions were discovered to express high levels of lysyl oxidase-like 4 (LOXL4) protein, which is mediated by transforming growth factor beta-1 (TGFβ1) [10].
LOXL4 is primarily involved in extracellular matrix remodeling, and its expression has been found to be elevated in different types of cancer including hepatocellular carcinoma (HCC) [12] and head and neck squamous cell carcinoma (HNSCC) [13]. The upregulation of LOXL4 in tumour is linked to cancer cell migration and invasion, and this function of LOXL4 is mediated by the activated FAK/SRC pathway [14]. The upregulation of LOXL4 facilitates immune evasion by cancer cells that lead to the establishment of an immunosuppressive microenvironment in various cancers [15]. In this context, Tan et al. [16] have suggested that the injection of LOXL4 induced intratumoral infiltration of macrophages and accelerated tumour growth in HCC. Additionally, exposure of macrophages to LOXL4 invoked their immunosuppressive phenotype and induced the expression of programmed death ligand 1 (PD-L1), which plays a key role in the suppression of CD8+ T cells [16]. A recent study by Görögh et al. [17] demonstrated the therapeutic effect of anti-LOXL4 monoclonal antibody in HNSCC xenografts. Accordingly, the injection of anti-LOXL4 monoclonal antibody into inoculated mice with HNSCC resulted in extensive tumour destruction [17].
The role of LOXL4 in tumour vascularization is poorly studied and requires further investigation. Opposite to our previous study [10], the majority of studies suggested a positive correlation between the upregulation of LOXL4 in the tumour microenvironment and tumour angiogenesis. In this context, the upregulation of LOXL4 in keratocystic odontogenic tumour (KCOT) stromal tissue and primary KCOT stromal fibroblasts promoted tumour angiogenesis [18]. Likewise, the overexpression of LOXL4 induced angiogenesis in HCC [16].
Due to neutrophils’ abundant recruitment to the site of pathology, neutrophils undergo programmed cell death to regulate the toxic neutrophil products to the surrounding tissue [19]. Interestingly, the released DNA strands with neutrophil proteins promote cancer cell proliferation, adhesion and invasion [20]. However, the role of apoptotic neutrophils in tumour vascularization is unclear.
In this manuscript, we examined the expression levels of pro-apoptotic markers [12], cleaved caspase-3 and cleaved poly (ADP-ribose) polymerase-1 (PARP-1) [21,22], in the neutrophils of CRCLM specimens. Our data suggested the upregulation of pro-apoptotic markers in the majority of neutrophils in vessel co-opting lesions compared to their angiogenic counterparts. Moreover, our in vitro data suggest that this process is possibly facilitated by TGFβ1.
2. Materials and methods
2.1. Patient samples
The blood and specimens of CRCLM patients were obtained through the MUHC-Research Institute Liver Disease Biobank from patients who underwent surgical resection at the Royal Victoria Hospital in Montreal. Informed consent was obtained from all patients, and ethical approval was granted by the McGill University Health Centre Research Ethics Board (protocol #SDR-11-066).
2.2. Neutrophil isolation
Human neutrophils were isolated from the peripheral blood of CRCLM patients following the protocol published in our previous publication [10]. After collection from a donor, whole blood was centrifuged at 2000g for 15min at 21°C, after which the plasma was carefully aspirated and replaced with 1× PBS (Wisent Inc., St-Bruno, QC, Canada, #311-010-CL). The cells were separated using dextran sedimentation, laid over lymphocyte separation media, and spun down. The resulting pellet was then subjected to hypotonic lysis with water. The purity of the isolated neutrophils was ascertained by trypan blue staining based on their morphological appearance, which contained less than 0.5% monocytes or lymphocytes. The purified neutrophils were resuspended in RPMI (Wisent Inc., St-Bruno, QC, Canada, #350-007-CL) supplemented with 5% serum at a final concentration of 1x106 cells/ml.
2.3. Neutrophil co-culturing and stimulation
After isolation, the neutrophils were cultured either independently or co-cultured with cancer cells and hepatocytes following the protocols in our previous publications [10]. Briefly, we resuspended 1x106 neutrophils in 1mL of RPMI (Wisent Inc., St-Bruno, QC, Canada, #350-007-CL) supplemented with 5% FBS (Wisent Inc., St-Bruno, QC, Canada, #085-150). The cells were then placed on coverslips mounted in 6 well companion plates (Falcon #353502) and incubated for 1h at 37°C. Then, the media was aspirated and replaced by RPMI supplemented with 1% FBS. The inserts (Falcon, #353090), containing a mixture of cancer cells and hepatocytes, were also added to the cultured neutrophils and incubated for 20 hours at 37°C.
The stimulation of neutrophils with recombinant TGFβ1 protein (Peprotech, Cranbury, NJ, USA, #100-21) was performed following the protocol that was published in the previous publications[10]. After isolation, the neutrophils (1x106 cells/ml) were placed on coverslips mounted in 6 well plates and incubated in RPMI supplemented with 5% FBS for 1h at 37°C. Then, the media was removed and replaced with RPMI supplemented with 1% FBS and 100pm recombinant TGFβ1 protein, and incubated for 20 hours at 37°C.
2.4. Immunofluorescence staining
Formalin-fixed paraffin-embedded (FFPE) human CRCLM blocks were deparaffinized with xylene followed by hydration with graded concentrations of ethanol and then with distilled water. The blocks were subjected to antigen retrieval (pH = 6.0) for 20 minutes and washed with PBS followed by inhibiting endogenous peroxidase using dual endogenous enzyme blocker (Dako, #S2003). The sections were then blocked with 1% goat serum for 1 hour followed by incubation with the indicated primary antibody prepared in 1% goat serum overnight at 4°C. Next day, the sections were incubated with secondary antibody 1:1000 (Alexa Flour 594 goat anti-rabbit IgG and Alexa Flour 488 goat anti-mouse IgG (Invitrogen #A11037 and #A10680 respectively)) at room temperature for 1 hour followed by washing with PBST thrice. The sections were incubated with DAPI (Thermo Fisher Scientific, #D1306, diluted in PBS 1:1000) for 10 minutes at room temperature. Then, 1-2 drops of ProLong Gold Antifade Mountant (Thermo Scientific, Saint-Laurent, QC, Canada, #P36934) were added to each section and covered with cover glasses.
We stained the cultured neutrophils following the protocol in the previous publication [10]. Briefly, the neutrophils were fixed by incubating with 4% paraformaldehyde (Biolegend, #420801) for 10 minutes. The neutrophils were washed with PBS, and permeabilized with 0.1% Triton X-100 (Bio-Rad, #161-0407). The cells then washed with PBS thrice and blocked with 5% BSA (GE Healthcare Life Science, #SH30574.02) for 1 hour. The neutrophils were then incubated with primary antibodies at 4 °C overnight. The next day, the cells were washed with PBS and incubated with secondary antibodies for 1 hour in the dark. After incubation, the cells were washed thrice with PBS and incubation with DAPI (Thermo Fisher Scientific, D1306, dilution 1:1000 in PBS) for 10 min followed by washing with PBST. The cover coverslips were mounted using ProLong Gold Antifade Mountant (Thermo Fisher Scientific, P36934). Slides were visualized using a Zeiss LSM780 confocal microscope system.
The following primary antibodies were used: HSA (Santa Cruz, #SC5893, dilution 1:300), cleaved caspase-3 (Cell signaling, # 9664S, dilution 1:100), cleaved PARP-1 (Cell signaling, # 5625S, dilution 1:50), elastase (R&D Systems, #MAB91671, dilution 1:250), and annexin-v (R&D Systems, #4830-01-K, dilution 1:250). The secondary antibodies used were: Alexa Fluor 488-conjugated goat anti-mouse (Invitrogen, A-10680; dilution 1:500), Alexa Fluor 594-conjugated goat anti-rabbit (Invitrogen, A-11037; dilution 1:500), and Alexa Fluor 647-conjugated goat anti-mouse (Invitrogen, A-32728; dilution 1:500).
The number of neutrophils expressing pro-apoptotic markers was calculated manually. The intensity of the positive signals was quantified with ImageJ (NIH, Bethesda, MD) software. The average pixel intensity was analyzed by measuring three areas for each sample that randomly selected.
2.5. Statistical analysis
GraphPad Prism software version 9.5.0 (GraphPad Software, La Jolla, CA, USA) was used for statistical analyses. Data as standard deviation. Statistical significance between the two groups was analyzed using unpaired Student’s t-test. p-values of <0.05 were considered to be significant
3. Results
3.2. Vessel co-opting tumours comprise a higher number of apoptotic neutrophils compared to angiogenic lesions
Previous investigations suggested that tumour-associated neutrophils (TANs) promote tumour angiogenesis [23]. Additionally, it has been shown that the lower levels of apoptosis in TANs positively correlate with improved tumour angiogenesis [24]. Since vessel co-opting CRCLM lesions are characterized by lacking angiogenic vascularization [2], we postulated a higher number of apoptotic TANs in vessel co-oping CRCLM lesions in comparison to angiogenic tumours. To test this, we performed immunofluorescence staining for chemonaïve CRCLM specimens (n=3 RHGP; n=3 DHGP) using cleaved caspase-3 or cleaved PARP-1, pro-apoptosis biomarkers [21], and elastase, neutrophil biomarker [10], antibodies. As shown in Figure 1, we observed the expression of cleaved caspase-3 and cleaved PARP-1 in over 50% of the neutrophils in vessel co-opting lesions, while none of the neutrophils in angiogenic lesions expressed the pro-apoptotic markers. Interestingly, we also noticed an inverse association between the expression levels of elastase and pro-apoptotic markers in neutrophils. This correlation requires further investigation in future. Taken together, our data indicate that the majority of neutrophils in vessel co-opting CRCLM are apoptotic compared to their counterparts in angiogenic CRCLM lesions.
3.3. TGFβ1 mediates apoptosis in CRCLM tumours
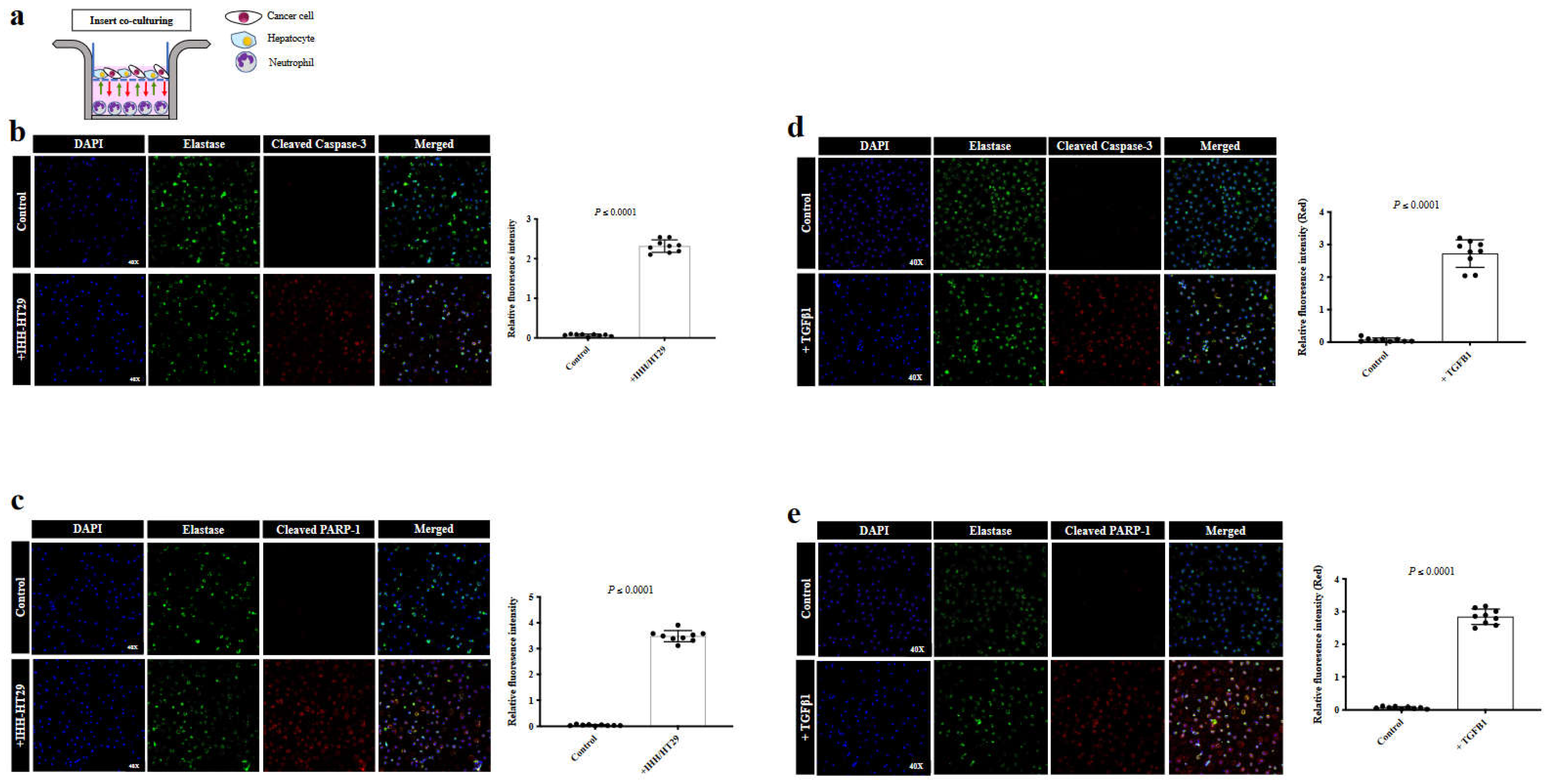
The cancer cells in vessel co-opting CRCLM lesions are characterized by their direct contact with hepatocytes [2,25]. Therefore, we conducted insert co-culturing for freshly isolated neutrophils from CRCLM patients with a mixture of human hepatocytes (IHH) and colorectal cancer (HT29) cells (Figure 2a) followed by immunofluorescence staining using a neutrophil marker (elastase) [10] and pro-apoptotic marker (cleaved caspase-3 and PARP-1) [21]. Importantly, we found a significant increase in the expression levels of cleaved caspase-3 (Figure 2b) and cleaved PARP-1 (Figure 2c) in the neutrophils upon their co-culturing with hepatocyte/cancer cells mixture.
Based on our recent publications [21,26], TGFβ1 significantly upregulated in the microenvironment of vessel co-option CRCLM tumours compared to their angiogenic counterparts. Accordingly, the expression of TGFβ1 is promoted by direct interaction between cancer cells and hepatocytes [26,27]. Therefore, we asked whether TGFβ1 involved in the induction of neutrophil apoptosis by co-cultured mixed cancer cells and hepatocytes. To answer this question, we exposed freshly isolated neutrophils from CRCLM patients to 100 picomolar of recombinant TGFβ1 for 20 hours followed by immunofluorescence staining using elastase, cleaved caspase-3 and cleaved PARP-1 antibodies. Indeed, the expression levels of cleaved caspase-3 (Figure 2d) and cleaved PARP-1 (Figure 2e) were significantly elevated in neutrophils upon exposure to TGFβ1.
We previously reported that TGFβ1 stimulates the expression of LOXL4 in the neutrophils of CRCLM tumours [10]. Since exposing neutrophils to TGFβ1 induces the expression LOXL4 [10] and pro-apoptotic markers (Figure 2d,e), we decided to examine the colocalization between apoptotic marker, annexin-v [22], and LOXL4 in vessel co-opting CRCLM sections. Interestingly, we noticed a colocalization of LOXL4 and annexin-v in the neutrophils (Supplementary Figure S1), indicating a positive correlation between the expression of LOXL4 and apoptosis in neutrophils. Indeed, further studies are required to confirm these results, as well as to determine the functional effect of LOXL4 in the development vessel co-option in CRCLM. Altogether, these data indicate TGFβ1 as a mediator of neutrophil apoptosis in CRCLM.
4. Discussion
The role of neutrophils in tumorigenesis is highly controversial. Some studies suggest apoptotic TANs as potent antitumour effector cells [28]. However, other studies suggest a positive correlation between TANs and tumour progression [29,30]. Various studies showed that high levels of intra-tumoural neutrophils are associated with unfavourable survival in diverse solid tumours, including breast and colorectal cancer [31,32]. Furthermore, it has been reported that neutropenia, a deficiency of neutrophils, is beneficial to survival in cancer patients undergoing chemotherapy [33].
The function of TANs in angiogenesis is also quite divisive. Various studies suggested a positive association between TANs and angiogenesis [34]. In this context, it has been proposed that TANs induce tumour angiogenesis through various proteins, including matrix metallopeptidase 9 (MMP-9), vascular endothelial growth factor (VEGF) and Bv8 (prokineticin) [35,36]. In contrast, our recent publications suggested the inverse correlation between TANs and angiogenesis in CRCLM [10,11]. Importantly, Loffredo et al. [37] have demonstrated releasing of antiangiogenic isoform of VEGF-A165b by neutrophils ex vivo.
In the current study, we observed a higher number of apoptotic TANs in vessel co-opting CRCLM specimens compared to their angiogenic counterparts. Vessel co-opting CRCLM lesions are characterized by direct interaction between cancer cells and hepatocytes, resulting in overexpression of various proteins in the tumour microenvironment, such as TGFβ1 [21,38]. Moreover, our previous publications [21,26] also showed higher levels of TGFβ1 in the conditioned media upon co-culturing cancer cells with hepatocytes. Herein, we showed overexpression of pro-apoptotic markers in the neutrophils that co-cultured with mixed cancer cells and hepatocytes, and it is possibly incited by TGFβ1. In agreement with our data, TGFβ1 has been reported to promote neutrophil apoptosis indirectly through the IL-6/LPS pathway [39]. Thus, further investigations are required to determine the mechanistic pathways underline TGFβ1-dependent neutrophil apoptosis in CRCLM.
The role of apoptotic neutrophils in the vascularization of CRCLM tumours is largely unknown. Since our data demonstrated a positive correlation between the expression of LOXL4 and pro-apoptotic markers in neutrophils, we postulated that the presence of apoptotic neutrophils indirectly induces vessel co-option in CRCLM tumours through LOXL4. Various studies suggested LOXL4 as a stimulator of cancer cell motility in various cancers [16,18,40,41]. Of note, our previous publications demonstrated that the upregulation of cancer cell motility significantly increases the development of vessel co-option in CRCLM [2,26]. Therefore, we hypothesize that the apoptotic neutrophils promote vessel co-option in CRCLM through the induction of cancer cell motility, and LOXL4 plays a pivotal role in this process. Indeed, further investigations are needed to confirm this hypothesis.
In conclusion, our results suggest a higher number of apoptotic neutrophils in vessel co-opting CRCLM lesions compared to their angiogenic counterparts. The function of apoptotic neutrophils in CRCLM vascularization warrants further investigation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.R., A.L. and P.M.; methodology, M.R., M.T and J.B.; resources, D.K., S.P.; data curation, M.R.; writing, M.R.; review and editing, M.R., and P.M; funding acquisition, P.M.
Acknowledgments
The authors would like to acknowledge the support that provided by Dana Massaro and Ken Verdoni Liver Metastases Research Fellowship. We also thank all those patients who participated in this study and whose samples were pivotal to this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ibrahim, N.; Lazaris, A.; Rada, M.; Petrillo, S.; Huck, L.; Hussain, S.; Ouladan, S.; Gao, Z.; Gregorieff, A.; Essalmani, R.; et al. Angiopoietin1 Deficiency in Hepatocytes A Ff Ects the Growth of Colorectal Cancer Liver. Cancers (Basel) 2020, 12, 1–18. [Google Scholar]
- Frentzas, S.; Simoneau, E.; Bridgeman, V.L.; Vermeulen, P.B.; Foo, S.; Kostaras, E.; Nathan, M.R.; Wotherspoon, A.; Gao, Z.H.; Shi, Y.; et al. Vessel Co-Option Mediates Resistance to Anti-Angiogenic Therapy in Liver Metastases. Nat Med 2016, 22, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Mayer, T.Z.; Kim, D.H.; Rada, M.; Petrillo, S.; Lazaris, A.; Metrakos, P. Role of Innate Immune Cells in the Development of Vessel Co-Opting CRC Liver Metastases. Cancer Res 2020, 80. [Google Scholar] [CrossRef]
- Kuczynski, E.A.; Vermeulen, P.B.; Pezzella, F.; Kerbel, R.S.; Reynolds, A.R. Vessel Co-Option in Cancer. Nat Rev Clin Oncol 2019, 16, 469–493. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Kapelanski-Lamoureux, A.; Tsamchoe, M.; Petrillo, S.; Lazaris, A.; Metrakos, P. Angiopoietin-1 Upregulates Cancer Cell Motility in Colorectal Cancer Liver Metastases through Actin-Related Protein 2/3. Cancers (Basel) 2022, 14, 2540. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front Immunol 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Dong, L.; Cheng, L. Neutrophils in Cancer Carcinogenesis and Metastasis. J Hematol Oncol 2021, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, C.C.; Malanchi, I. Neutrophils in Cancer: Heterogeneous and Multifaceted. Nat Rev Immunol 2022, 22, 173–187. [Google Scholar] [CrossRef]
- Mollinedo, F. Neutrophil Degranulation, Plasticity, and Cancer Metastasis. Trends Immunol 2019, 40, 228–242. [Google Scholar] [CrossRef]
- Palmieri, V.; Lazaris, A.; Mayer, T.Z.; Petrillo, S.K.; Alamri, H.; Rada, M.; Jarrouj, G.; Park, W.; Gao, Z.; McDonald, P.P.; et al. Neutrophils Expressing Lysyl Oxidase-like 4 Protein Are Present in Colorectal Cancer Liver Metastases Resistant to Anti-angiogenic Therapy. J Pathol 2020, 251, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Hassan, N.; Lazaris, A.; Metrakos, P. The Molecular Mechanisms Underlying Neutrophil Infiltration in Vessel Co-Opting Colorectal Cancer Liver Metastases. Front Oncol 2022. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Y.; Zhang, X.; Feng, M.; Ma, J.; Li, J.; Yang, X.; Fang, F. Exosome-Mediated Secretion of LOXL4 Promotes Hepatocellular Carcinoma Cell Invasion and Metastasis. Mol Cancer 2019, 18, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and Neck Squamous Cell Carcinoma. Nat Rev Dis Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Yao, F.; Huang, C. Lysyl Oxidase Family Proteins: Prospective Therapeutic Targets in Cancer. Int J Mol Sci 2022, 23, 12270. [Google Scholar] [CrossRef]
- Sun, C.; Ma, S.; Chen, Y.; Kim, N.H.; Kailas, S.; Wang, Y.; Gu, W.; Chen, Y.; Tuason, J.P.W.; Bhan, C.; et al. Diagnostic Value, Prognostic Value, and Immune Infiltration of LOX Family Members in Liver Cancer: Bioinformatic Analysis. Front Oncol 2022, 12. [Google Scholar] [CrossRef]
- Tan, H.-Y.; Wang, N.; Zhang, C.; Chan, Y.-T.; Yuen, M.-F.; Feng, Y. Lysyl Oxidase-Like 4 Fosters an Immunosuppressive Microenvironment During Hepatocarcinogenesis. Hepatology 2021, 73, 2326–2341. [Google Scholar] [CrossRef]
- Görögh, T.; Quabius, E.S.; Heidebrecht, H.; Nagy, A.; Muffels, T.; Haag, J.; Ambrosch, P.; Hoffmann, M. Lysyl Oxidase Like-4 Monoclonal Antibody Demonstrates Therapeutic Effect against Head and Neck Squamous Cell Carcinoma Cells and Xenografts. Int J Cancer 2016, 138, 2529–2538. [Google Scholar] [CrossRef]
- Jiang, W.-P.; Sima, Z.-H.; Wang, H.-C.; Zhang, J.-Y.; Sun, L.-S.; Chen, F.; Li, T.-J. Identification of the Involvement of LOXL4 in Generation of Keratocystic Odontogenic Tumors by RNA-Seq Analysis. Int J Oral Sci 2014, 6, 31–38. [Google Scholar] [CrossRef]
- Fox, S.; Leitch, A.E.; Duffin, R.; Haslett, C.; Rossi, A.G. Neutrophil Apoptosis: Relevance to the Innate Immune Response and Inflammatory Disease. J Innate Immun 2010, 2, 216–227. [Google Scholar] [CrossRef]
- Brostjan, C.; Oehler, R. The Role of Neutrophil Death in Chronic Inflammation and Cancer. Cell Death Discov 2020, 6. [Google Scholar] [CrossRef]
- Rada, M.; Tsamchoe, M.; Kapelanski-lamoureux, A.; Hassan, N.; Bloom, J.; Petrillo, S.; Kim, D.H.; Lazaris, A.; Metrakos, P. Cancer Cells Promote Phenotypic Alterations in Hepatocytes at the Edge of Cancer Cell Nests to Facilitate Vessel Co-Option Establishment in Colorectal Cancer Liver Metastases. 2022, 1–19.
- Althubiti, M.; Rada, M.; Samuel, J.; Escorsa, J.M.; Najeeb, H.; Lee, K.-G.; Lam, K.-P.; Jones, G.D.D.; Barlev, N.A.; Macip, S. BTK Modulates P53 Activity to Enhance Apoptotic and Senescent Responses. Cancer Res 2016, 76. [Google Scholar] [CrossRef] [PubMed]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front Oncol 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Andzinski, L.; Wu, C.F.; Lienenklaus, S.; Kröger, A.; Weiss, S.; Jablonska, J. Delayed Apoptosis of Tumor Associated Neutrophils in the Absence of Endogenous IFN-β. Int J Cancer 2015, 136, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Lazaris, A.; Kapelanski-Lamoureux, A.; Mayer, T.Z.; Metrakos, P. Tumor Microenvironment Conditions That Favor Vessel Co-Option in Colorectal Cancer Liver Metastases: A Theoretical Model. Semin Cancer Biol 2021, 71, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Kapelanski-Lamoureux, A.; Petrillo, S.; Tabariès, S.; Siegel, P.; Reynolds, A.R.; Lazaris, A.; Metrakos, P. Runt Related Transcription Factor-1 Plays a Central Role in Vessel Co-Option of Colorectal Cancer Liver Metastases. Commun Biol 2021, 4, 1–15. [Google Scholar] [CrossRef]
- Rada, M.; Tsamchoe, M.; Kapelanski-lamoureux, A.; Hassan, N.; Bloom, J.; Petrillo, S.; Kim, D.H.; Lazaris, A.; Metrakos, P. Cancer Cells Promote Phenotypic Alterations in Hepatocytes at the Edge of Cancer Cell Nests to Facilitate Vessel Co-Option Establishment in Colorectal Cancer Liver Metastases. 2022, 1–19.
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J Immunol Res 2015, 2015. [Google Scholar] [CrossRef]
- Granot, Z. Neutrophils as a Therapeutic Target in Cancer. Front Immunol 2019, 10, 1710. [Google Scholar] [CrossRef]
- Sionov, R.V.; Fridlender, Z.G.; Granot, Z. The Multifaceted Roles Neutrophils Play in the Tumor Microenvironment. Cancer Microenvironment 2015, 8, 125–158. [Google Scholar] [CrossRef]
- Shen, M.; Hu, P.; Donskov, F.; Wang, G.; Liu, Q.; Du, J. Tumor-Associated Neutrophils as a New Prognostic Factor in Cancer: A Systematic Review and Meta-Analysis. PLoS One 2014, 9, 1–10. [Google Scholar] [CrossRef]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S. v.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The Prognostic Landscape of Genes and Infiltrating Immune Cells across Human Cancers. Nat Med 2015, 21, 938–945. [Google Scholar] [CrossRef]
- Shitara, K.; Matsuo, K.; Oze, I.; Mizota, A.; Kondo, C.; Nomura, M.; Yokota, T.; Takahari, D.; Ura, T.; Muro, K. Meta-Analysis of Neutropenia or Leukopenia as a Prognostic Factor in Patients with Malignant Disease Undergoing Chemotherapy. Cancer Chemother Pharmacol 2011, 68, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, X.H.F. Tumor-Associated Neutrophils and Macrophages—Heterogenous but Not Chaotic. Front Immunol 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Zajac, E.; Juncker-Jensen, A.; Kupriyanova, T.A.; Welter, L.; Quigley, J.P. Tissue-Infiltrating Neutrophils Constitute the Major In Vivo Source of Angiogenesis-Inducing MMP-9 in the Tumor Microenvironment. Neoplasia (United States) 2014, 16, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Grecian, R.; Whyte, M.K.B.; Walmsley, S.R. The Role of Neutrophils in Cancer. Br Med Bull 2018, 128, 5–14. [Google Scholar] [CrossRef]
- Loffredo, S.; Borriello, F.; Iannone, R.; Ferrara, A.L.; Galdiero, M.R.; Gigantino, V.; Esposito, P.; Varricchi, G.; Lambeau, G.; Cassatella, M.A.; et al. Group V Secreted Phospholipase A2 Induces the Release of Proangiogenic and Antiangiogenic Factors by Human Neutrophils. Front Immunol 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Rada, M.; Kapelanski-Lamoureux, A.; Zlotnik, O.; Petrillo, S.; Lazaris, A.; Metrakos, P. Disruption of Integrin Alpha-5/Beta-1-Dependent Transforming Growth Factor Beta-1 Signaling Pathway Attenuates Vessel Co-Option in Colorectal Cancer Liver Metastases. bioRxiv 2022, 2003–2005. [Google Scholar] [CrossRef]
- Ganeshan, K.; Johnston, L.K.; Bryce, P.J. TGF-Β1 Limits the Onset of Innate Lung Inflammation by Promoting Mast Cell–Derived IL-6. The Journal of Immunology 2013, 190, 5731–5738. [Google Scholar] [CrossRef]
- Setargew, Y.F.I.; Wyllie, K.; Grant, R.D.; Chitty, J.L.; Cox, T.R. Targeting Lysyl Oxidase Family Meditated Matrix Cross-linking as an Anti-stromal Therapy in Solid Tumours. Cancers (Basel) 2021, 13, 1–26. [Google Scholar] [CrossRef]
- Lin, H.Y.; Li, C.J.; Yang, Y.L.; Huang, Y.H.; Hsiau, Y.T.; Chu, P.Y. Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. Int J Mol Sci 2020, 21, 1–23. [Google Scholar] [CrossRef]
Figure 1.
Tumor associated neutrophils in vessel co-opting CRCLM lesions expressing pro-apoptotic markers. Immunofluorescence staining of chemonaïve CRCLM specimens showing neutrophil biomarker, elastase (green), and cleaved caspase-3 (red) (a) or cleaved PARP-1 (red) (b). The right panels represent the percentage of the neutrophils expressing cleaved caspase-3 (red) or cleaved PARP-1. T: Tumour; L: Liver parenchyma; D: Desmoplastic ring.
Figure 1.
Tumor associated neutrophils in vessel co-opting CRCLM lesions expressing pro-apoptotic markers. Immunofluorescence staining of chemonaïve CRCLM specimens showing neutrophil biomarker, elastase (green), and cleaved caspase-3 (red) (a) or cleaved PARP-1 (red) (b). The right panels represent the percentage of the neutrophils expressing cleaved caspase-3 (red) or cleaved PARP-1. T: Tumour; L: Liver parenchyma; D: Desmoplastic ring.
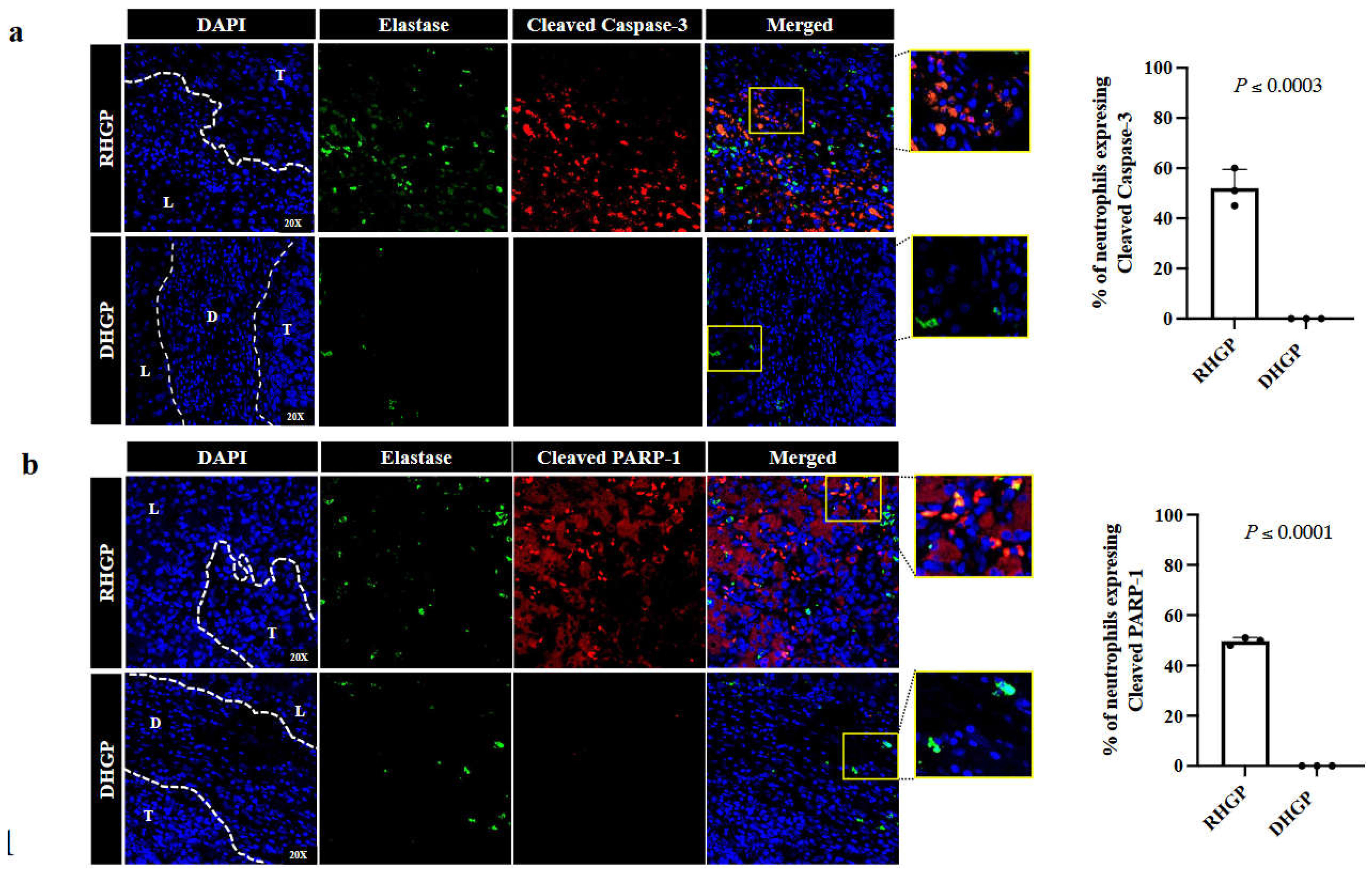
Figure 2.
TGFβ1 promotes apoptosis in neutrophils in vitro. a. Schematic of experimental design. (b and c). Immunofluorescence staining of neutrophils co-cultured with IHH hepatocytes and/or HT29 cancer cell lines using elastase (green) and cleaved caspase-3 (red) or PARP-1 (red) antibodies, respectively. The right panel shows the quantification of positive pixels. Average pixel intensity was measured from three randomly selected areas for each sample. (d and e). Represent the expression of cleaved caspase-3 or cleaved PARP-1 in the control neutrophils and those that were exposed to recombinant TGFβ1 for 20 hours. The right panel shows the quantification of positive pixels. Average pixel intensity was measured from three randomly selected areas for each sample. Results are representative of 3 independent experiments.
Figure 2.
TGFβ1 promotes apoptosis in neutrophils in vitro. a. Schematic of experimental design. (b and c). Immunofluorescence staining of neutrophils co-cultured with IHH hepatocytes and/or HT29 cancer cell lines using elastase (green) and cleaved caspase-3 (red) or PARP-1 (red) antibodies, respectively. The right panel shows the quantification of positive pixels. Average pixel intensity was measured from three randomly selected areas for each sample. (d and e). Represent the expression of cleaved caspase-3 or cleaved PARP-1 in the control neutrophils and those that were exposed to recombinant TGFβ1 for 20 hours. The right panel shows the quantification of positive pixels. Average pixel intensity was measured from three randomly selected areas for each sample. Results are representative of 3 independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



