
Submitted:
05 May 2023
Posted:
06 May 2023
You are already at the latest version
Abstract
Heavy metals and metalloids (HMs) are environmental pollutants, most notably cadmium, lead, arsenic, mercury, and chromium. When HMs accumulate to toxic levels in agricultural soils, these non-biodegradable elements adversely affect crop health and productivity. The toxicity of HMs on crops depends upon factors including crop type, growth condition, and developmental stage; nature of toxicity of the specific elements involved; soil physical and chemical properties; occurrence and bioavailability of HM ions in the soil solution; and soil rhizosphere chemistry. HMs can disrupt normal structure and function of cellular components and impede various metabolic and developmental processes. This review evaluates: (1) HM contamination in arable lands through agricultural practices, particularly due to chemical fertilizers, pesticides, livestock manures and compost, sewage-sludge based biosolids, and irrigation; (2) factors affecting the bioavailability of HM elements in the soil solution, and their absorption, translocation, and bioaccumulation in crop plants; (3) mechanisms by which HM elements directly interfere with the physiological, biochemical, and molecular processes in plants with particular emphasis on the generation of oxidative stress, inhibition of photosynthetic phosphorylation, enzyme/protein inactivation, genetic modifications, and hormonal deregulation, and indirectly through inhibition of soil microbial growth, proliferation, and diversity; and (4) visual symptoms of highly toxic non-essential HM elements in plants with an emphasis on crop plants. Finally, suggestions and recommendations are made to minimize crop losses from suspected HM contamination in agricultural soils.
Keywords:
Heavy metals
; arable lands
; agricultural practices
; soil binding models
; action mechanisms
; visual symptoms
; crop production
1. Introduction
Metals including potentially toxic elements are inorganic elements containing atomic densities (g.cm−3) several times higher than H2O (1.g.cm−3) and broadly classified into heavy and light metals, and semi-metals (Figure 1). Based on physical, physiological, and chemical properties, metals have been classified under several sub-groups, namely: transition metals: e.g., chromium (Cr), manganese (Mn), iron (Fe), cobalt (Co), nickel (Ni), copper (Cu), and molybdenum (Mo); post-transition metals: e.g., aluminum (Al), zinc (Zn), cadmium (Cd), mercury (Hg), and lead (Pb); alkaline earth metals: e.g., calcium (Ca), magnesium (Mg), beryllium (Be), and barium (Ba); alkali metals: e.g., lithium (Li), sodium (Na), potassium (K), and cesium (Cs); and metalloids, which are also referred to as semi-metals because of their metallic and non-metallic properties: e.g., boron (B), silicon (Si), arsenic (As), and antimony (Sb) [1].
Heavy metals and metalloids (HMs) are environmental pollutants. They are also agricultural soil contaminants because if present at elevated levels in the soil, HMs can negatively impact crop health and productivity [2,3]. HMs are recalcitrant to degradation, and if not taken up by plants or removed by leaching, they can accumulate in the soil and persist for long periods [4,5,6]. The elements that are frequently found to contaminate agricultural soils and cause toxic effects at elevated levels on plants include Cd, Pb, Cr, As, Hg, Ni, Cu and Zn [4,7]. Among them, Cd, Pb, As, Hg, and Cr are highly toxic and detrimental to plant health at almost all levels of contamination [8,9,10].
Several elements are classified as essential mineral nutrients for plant growth and productivity (Figure 1). Examples include Cu, Zn, Fe, Mn, Mo, Ni, Mg, Ca, and B. At relatively low concentrations, these elements can enhance specific cellular functions in plants including ion homeostasis, pigment biosynthesis, photosynthesis, respiration, enzyme activities, gene regulation, sugar metabolism, nitrogen fixation etc. [3,8,11]. However, when accumulated at concentrations above optimum, these same essential elements can adversely affect plant growth, development, and reproduction [2,3]. Conversely, if the concentration falls below certain threshold levels, they also produce mineral deficiency symptoms in plants [11].
HM contamination in agricultural soil is a global issue. The problem is more prominent in many developing countries partly because of rapid urbanization and industrialization, and perhaps partly due to lack of proper awareness about the toxic consequences of these elements not only to human health but also to crop health [12,13,14,15,16]. This review concentrates on the adverse effects of HMs on crop health.
2. Sources of HM contamination in arable lands
Agricultural soil is an important non-renewable natural resource. It can be contaminated with toxic HM elements such as Cd, Pb, Cr, As, Hg, Cu, Ni, Zn, Al, and several others due to natural causes as well as anthropogenic activities. Natural causes include, among many others, weathering of metal-bearing rocks by rainwater and atmospheric deposition. Anthropogenic activities include industrial activities (e.g., mining, leather tanning, textile, and petrol-chemical), disposal of metal containing wastes, vehicle exhausts, and agricultural practices [4,15,17,18,19,20]. However, irrespective of the source of contamination, continued addition of HMs to arable lands can result in soils that can be too toxic to support plant growth and productivity. The following subsections review primarily the contamination of HMs in farmlands through suspected agricultural practices as outlined in Figure 2.
2.1. Application of chemical fertilizers
Chemical fertilizers, particularly inorganic fertilizers, are a crucial input for crop production. Consequently, large quantities of inorganic fertilizers including nitrogen (N), phosphorus (P), potassium (K), and compound/mixed fertilizers are routinely added to agricultural lands to supply adequate quantities of these macronutrients. For instance, it was estimated that in 2019, more than 220 million tons of commercial fertilizers and liming materials were applied worldwide mostly to agricultural fields [21].
Among these fertilizers, P-fertilizers contain the highest level of HM contaminants [4,22,23,24]. For example, superphosphate fertilizers can contain Cd, Co, Cu, Pb, Zn, Cr and Ni as contaminants. In a study that assessed soil with and without P-fertilizer amendments, the concentration of Zn was higher not only in the amended soil but also in the plants grown in that soil [25]. Cadmium content in the soil has been shown to increase persistently due to application of P-fertilizers [12,23,26]. Cadmium is extremely toxic to plants because of its high solubility and mobility in soil solution. The concentration of Cd present as an impurity in several P-fertilizers evaluated in a study is shown in Table 1.
In addition to P-fertilizers, copper sulphate, iron sulphate, and zinc sulphate fertilizers can also contain HM contaminants including Pb [22,27,28]. A study reported from greenhouse experiments that repeated application of chemical fertilizers significantly increased the accumulation of several HM elements in the soil (Table 2). Sources of HM contamination in fertilizers include the raw materials used in the manufacture of inorganic fertilizers. For instance, phosphate-rock, also known as phosphorite is utilized in the production of P-fertilizers [29,30]. Over 90% of potash extracted from mines is used in the manufacture of K-fertilizer [31]. Based on the level of HM impurities, chemical fertilizers can be ranked as follows: P-fertilizers ≥ compound fertilizers> K-fertilizers> N-fertilizer [32,33].
2.2. Pesticide application
Pesticides play an important role in global agriculture. It has been estimated that without pesticides the world’s food production can be reduced close to ~40% [34]. Another study estimated a 78% loss of fruit production, 54% loss of vegetable production, and 32% loss of cereal production without pesticide use [35]. Pesticides such as insecticides, fungicides, rodenticides, nematicides, and herbicides are composed of either organic or inorganic compounds that are toxic to the targeted organisms. Analysis of these compounds shows that some of them contain HM elements either as active ingredients (Table 3) or as impurities in the formulations (Table 4).
Several fungicides and insecticides extensively used in the past in agricultural lands were shown to contain significant concentrations of HM elements in their active ingredients. Examples include Cu containing fungicides such as copper sulphate (Bordeaux mixture also referred to as Bordo® mix) and copper oxychloride; Pb containing insecticide such as lead arsenate; and Cu containing insecticide such as copper acetoarsenite. The commonly found HM elements in the active ingredients of pesticide products include Cu, As, Pb, Hg, Cr, Zn, Al, Li, Ba, B, and Ti (titanium) [36,37].
On the other hand, HM elements can also be present in pesticide products as impurities. For example, certain pesticide products used for pest control in Japan contained Cd, Hg, As, Cu, Zn, and Pb as contaminants [17]. Chemical analysis of several pesticides including 11 glyphosate-based herbicide formulations by utilizing inductively coupled plasma/optical emission spectrometry (ICP-OES) detected As, Cr, Co, Pb, and Ni as contaminants [38]. Similar analysis of several insecticides using ICP mass spectrometry detected Zn, Cu, Cr, Co, Pb and Tl (thallium) as contaminants [39]. It has been suggested that the HM elements contaminate pesticide products during manufacturing process, while some of them are intentionally added as nano pesticides for increased efficacy [38,40].
2.3. Application of livestock manures and compost
Livestock manures are organic fertilizers composed predominantly of poultry, cattle, and pig manures. Application of these manures and the compost made from them to farmlands is a common practice in agricultural crop production. However, these manures and compost contain high concentrations of HM elements such as Cu, Zn, Cd, Ni, Cr, As, Pb, and Hg as contaminants [20,26,41,42]. A study conducted to determine the concentration of HM elements in different livestock and poultry manures is presented in Table 5. The major sources of contamination of HM elements in the manures include the minerals supplied with the commercial feeds [41,42]. For example, supplementation of animal feeds with growth-promoting organic arsenical products was practiced for many years [43]. Some studies confirmed that Zn, Cu, As, and Cd were artificially added to commercial feeds to promote animal growth and improve disease resistance [44,45]. However, animals cannot digest these HM elements and discharge them through manures [46]. Because HMs are non-degradable elements, they are also not broken down during the composting process [47]. Thus, long-term repeated applications of manures and compost can result in the buildup of HM elements to toxic levels in agricultural soil [48,49] and can affect crop health and productivity.
2.4. Application of sewage-sludge based biosolids.
Sewage-sludge originated from municipal and industrial wastes can be highly contaminated with HM elements such as As, Cd, Cr, Cu, Pb, Hg, Ni, Mo, Zn, and others. Long term application of untreated sewage sludge in some developing countries has led to increased concentrations of HMs in the agricultural lands [14,20,50]. However, the biosolids generated from sewage-sludge processing plants can be typically low in HM contamination and can contain organic materials rich in nutrients, and be used as fertilizers [51,52]. When applied to arable lands, processed sewage-sludge can improve soil physical properties and crop productivity. Utilization of these byproducts for agricultural crop production is therefore a common practice in many countries. In the United States, about 3.0 million dry tons of biosolids are utilized annually for crop production [4]. The European community countries utilized >30% of processed sewage-sludge as a fertilizer in agricultural lands [53]. Australia incorporates over 175K tons of dry biosolid into agricultural soil [54]. In the United States, federal regulations limit concentrations of major elements (e.g., As, Cd, Cu, Pb, Hg, Ni, Se, and Zn) commonly found in biosolids for land application (Table 6), [52]. Although biosolids produced from sewage-sludge processing treatment are generally low in HM concentration, repeated applications of these products can result in the buildup of HM elements in agricultural soil and can negatively impact crop health and productivity.
2.5. Land Irrigation
Irrigation of agricultural lands with contaminated water from surface water bodies as well as groundwater sources is another route of HM contamination in agricultural soil. The above irrigation practices are more frequently followed in some developing countries [9,15,16,56,57]. A review of a large number of related articles published in a span of over two decades (1994 to 2019) that determined the HM contamination in surface water bodies throughout the world showed that the average content of Cr, Mn, Co, Ni, As and Cd exceeded the permissible limits as prescribed by WHO and United States EPA [9,20]. Studies conducted to determine HM concentration in irrigation water in several locations of the Gazipur district of Bangladesh, and the Gondar city of Ethiopia showed that in almost all of their tests the concentration of HMs exceeded the FAO (Food and Agriculture Organization) prescribed admissible levels (Table 7).
The causes for HM contamination in surface water are both natural and anthropogenic, which include but are not limited to atmospheric deposition; geological and biological weathering; and discharge of agricultural, municipal, domestic, and industrial wastes. HM elements such as Pb, Ni, Cr, Cd, As, Hg, Zn, Cu, and others from diverse sources are transported to surface water bodies and irrigation canals through runoff, and to the underground aquifer through vertical leaching with percolating rainwater [16,59,60,61]. Thus, irrigation of croplands using HM contaminated water not only affects growth and productivity of crops [62] but can also threaten soil quality. It should however be noted that the extent of crop damage will depend on the pH of the irrigation water and redox potential and water solubility of the contaminated HM elements.
3. Factors affecting HM interactions with crop plants.
Several plant, soil, and metal related factors can influence HM interaction with crop plants [57,63,64,65,66,67]. The crop related factors include crop type (species, variety, genotype); growth stage and growth condition; metabolic activities; and uptake, translocation, and bioaccumulation capabilities. The major soil related factors include soil pH, organic matter (OM) content, cation exchange capacity (CEC), rhizosphere chemistry, and microbial activity. The minor soil related factors may include soil texture, hydration level, aeration (compactness), and temperature. The HM-related factors include speciation (organic vs. inorganic form), oxidation state, concentration, solubility, mobility, bioavailability, and interaction with soil particles and with the essential (e.g., Mg, Ca, Zn) or non-essential (e.g., Cd, Pb, Hg) ionic species.
3.1. Plant responses to HM toxicity
3.1.1. Plant type, growth stage, and growth conditions
The above features can influence HM interaction with crop plants. A few examples are cited below. Bean (Phaseolus spp.) plants exhibited tolerance to Cu toxicity at the early stages of growth as indicated by their primary photochemistry of photosynthesis [2,68]. The tolerance of alfalfa (Medicago sativa) plants to Cd, Cu and Zn toxicities was shown to be positively correlated with the plant age [69]. Tobacco (Nicotiana tabacum) plants accumulated relatively high concentrations of HMs in the leaves [70]. Among many vegetable species tested, specific species in the Brassicaceae family accumulated highest amounts of Cr, although Cr translocation from root to shoot was extremely limited in almost all species tested [71,72]. Investigation of several African vegetable species showed that the matembele (Ipomoea batatas) plants had the highest HM content followed by mchicha (Amaranthus hybridus), eggplant (Solanum melongena), and bamia (Abelmoschus esculentus) [65]. Root and shoot tissues of winter wheat (Triticum aestivum) tolerated high concentrations of Cd and can be used as an indicator for Cd contamination in agricultural soils [73]. Cadmium absorption capabilities of different rice varieties differed under similar growing conditions as some Japonica rice varieties had lower Cd concentration than most Indica varieties, and certain African upland rice varieties had even lower Cd absorption capabilities than the Japonica varieties [17]. Variability in HM absorption capabilities among different plant species or varieties may be caused by differences in morphological, physiological, anatomical, and genetic characteristics. Responses of several crop species to HM toxicity in the soil/nutrient culture as reported in some studies are shown in Table 8.
3.1.2. Plant metabolic activities
Plants have diverse mechanisms to prevent harmful effects of HMs including binding of HMs with the cell wall, transporting of HMs to vacuolar compartments, and synthesizing of metal binding proteins such as cysteine-rich metallothionein and phytochelatins. These proteins perform metal ion homeostasis, chelation, sequestration, and detoxification of excess HM elements in plant cells [77,78,79,80]. Reduced glutathione (GSH), the amino acid derivative of glutamic acid, cysteine and glycine has a strong affinity for HM elements such as Cd, Cu, Hg, Pb, Zn, Ni and As and acts as a ligand to chelate HMs to alleviate toxic effects on plants. Depending on their binding affinities with the GSH, HMs can be ranked as follows: Cd> Pb> Zn> Hg> As> Cu [81,82,83]. Some proteins belonging to mitogen-activated protein kinase (MAPK) are stimulated under Cu or Cd accumulation. These Cu or Cd-induced MAPKs enhance translation of transporters for HM sequestration and removal from plant cells [84]. Based on proteomic and other analyses, a study suggested that hemp plants can acclimate to high levels of Pb toxicity by enhancing photosynthesis (primary photochemistry), cellular respiration, and intercellular N and C assimilation; preventing unfolded protein aggregation and degrading misfolded proteins; and increasing transmembrane ATP transport [85]. Plants also release chemical messengers such as ethylene and jasmonic acid when grown in soil containing high levels of HMs that reduce HM toxicity in plants [86,87]. However, further research is needed to better understand how these signaling molecules interact with HM toxicity in plants.
3.1.3. Uptake, translocation, and bioaccumulation in plants
HMs enter plant roots via passive (apoplastic) or active (symplastic) movements [88,89]. However, the degree of phytotoxicity depends not only on root absorption but also on the translocation to different parts and accumulation to toxic levels in plants. For instance, a study showed that after 20 days of treatment, the translocation of Hg was <2% in the leaf and <4% in the shoot as compared to total quantity (µg g−1 DW) absorbed by roots of tomato seedlings [90]. Metal transporters play important roles in the uptake, translocation, and detoxification (by moving to vacuoles) of HMs in plants [91].
There are also antagonistic or synergistic interactions between HM elements during absorption or translocation in plants. For example, the presence of Hg in the growth medium significantly reduced As accumulation in the roots indicating the antagonistic effects of Hg against As absorption. However, the effect was synergistic when As was translocated to shoot particularly at higher Hg concentrations [76]. Cadmium uptake was reduced in rice plants when Fe plaque formed around roots indicating antagonistic effects of Fe on Cd uptake [92]. Another study showed that the Cr and Pb concentration in the locally grown vegetable species at the HM contaminated sites in Dhaka, Bangladesh was respectively, 10 and ~2 times higher than the FAO/WHO prescribed permissible limits in plants [93]. Based on the observations in several vegetable species, some studies suggested that differences in HM toxicity in plants can be attributed to their uptake and translocation differences [93,94].
Nonetheless, the overall phytotoxicity of HMs depends on, in addition to other factors, how plants carry out physiological functions such as phytostabilization (immobilization of HMs in the soil that can reduce bioavailability), rhizofiltration (adsorption of HMs with plant roots in the rhizosphere), phytoextraction (uptake and translocation of HMs in plants), phytoaccumulation (accumulation of HMs inside plants in active forms), and phytovolatilization (release of absorbed HMs in the atmosphere as volatile forms) [95,96,97]. However, the prevalence and bioavailability of HMs in the soils are basic requirements for phytotoxic effects in plants.
3.2. Occurrence and bioavailability of HMs
The occurrence of HMs in agricultural soils depends largely on the factors discussed in Section 2. The bioavailability and extractable concentration however appear to be predominantly controlled by the solubility of HMs in the soil solution and the OM content in the soil. Although there may be exceptions, in general the solubility of HMs in the soil can be positively correlated with [H+] (acidity). For instance, the solubility of most HM ions is lower in the basic pH range whereas it is higher in the range of acidic pH [54,98,99]. It was shown that a one-unit decrease in pH value resulted in about two-fold more increase in the bioavailable concentration of certain elements such as Zn, Ni, Cd, Al, and Cu in the soil solution [100,101]. Agricultural soils can be expected to be more acidic particularly in the moderate to high rain fall areas due to loss of base forming ions (basic cations e.g., Ca2+, Mg2+, K+, and Na+) from the farmlands because of prolonged leaching. Furthermore, acidity can also buildup over time in the soil due to formation of inorganic acids such as phosphoric acid, sulfuric acid, and nitric acid in the soil due to oxidation of applied phosphorus, sulfur, and ammonium/nitrate fertilizers, respectively [102]. On the other hand, the OM content in the soil appears to have negative effect on HM uptake in plants perhaps due to chelation of HMs by forming metal-OM complexes [103,104]. The above suggests that HM bioavailability can be reduced in soil containing high OM content.
3.2.1. Hypothetical soil binding diagrams of HMs
While the exact mechanism(s) by which the soil pH and OMs control the bioavailability of HMs for plant uptake is (are) unclear, based on the above discussion we have formulated a hypothetical soil-binding model as displayed in Figure 3. It shows that in basic soils containing high OM content and low [H+], most metal cations can be tightly attached to the negatively charged soil particles and become less available or unavailable for plant absorption (Figure 3A). On the other hand, in acidic soil containing high [H+], these metal cations can either not able to compete with H+ to bind with the soil particles or be released from soil particles in the presence high [H+] (Figure 3C). These may consequently leave more HM elements in the soil solutions for plant absorption [105,106]. This diagrammatic model also suggests that soil OMs can play a crucial role in HM mobility and bioavailability for plant absorption. Since OMs make soil particles more negatively charged [107], most HM cations are attracted to, and perhaps bound tightly with the soil particles containing high OM (Figure 3A), becoming unavailable for plant absorption. On the contrary, soil particles with low OM content will have less net negative charges to bind with positively charged HM elements (Figure 3B), and this can result in the availability of some free HM ions in the soil solutions for plant absorption.
The hypothetical model presented above is consistent with previous studies which suggest that HMs can bind with organic molecules and become less biologically available for plant uptake [103,108]. The above model also explains the reason why the bioavailability of HMs applied in the form of manure and compost is lower than that of HMs applied in salt forms [109]. A greenhouse study reported that application of chemical fertilizers increased the accumulation and bioavailability of Cu, Ni, Pb, and Zn in the greenhouse soil, and suggested that these HM elements were the contaminants of applied inorganic fertilizers [24]. Long-term field research also showed that the metal concentration in plants grown in soil amended with salt-forms of HMs was higher than that in plants amended with an equivalent quantity of HMs in organic or compost form [110].
From the above discussion, it appears that inorganic forms of HMs are readily available for plant uptake, whereas the organic and compost forms require microbial decomposition and conversion to inorganic forms for plant absorption. However, care should be practiced in adding processed sewage sludge based organic biosoilds in farmlands because excessive applications can cause soil structural modifications; deficiency of Zn, Mn, and Fe in plants due to phosphorus over-loading in the soil; and a buildup of HM elements such as Cu, Pb, and Zn to toxic levels for plant growth [111]. Further research would be needed to clarify the role of OMs and pH on the binding interaction of different HM elements with soil particles.
3.2.2. HM precipitation in the soil—effect of pH
Other lines of research suggest that in basic soils some HMs can be precipitated due to transformation into insoluble forms such as oxides, hydroxides, sulfides, sulfates, phosphates, silicates, carbonates etc. becoming biologically unavailable for plant absorption, whereas in the acidic soil, they can remain in free cationic forms in the soil solution and are biologically available for plant uptake [112,113,114]. The pH dependence of HM precipitation can vary among different elements because in most cases it is dependent on the oxidation states of the specific elements involved and the type reactions taking place in the soil solutions [112,115]. In general, an element containing higher oxidation states is more acidic than an element with lower oxidation states e.g., Fe3+ salts are more acidic than Fe2+ salts.
It appears that the above reports are consistent with the hypothetical diagrams presented in Figure 3, which also suggest that acidic soil pH can increase the bioavailability of certain metal cations, whereas basic pH reduces their availability for plant uptake. It has therefore been a common practice in agricultural farmlands with acidic soil to apply lime that elevates soil pH and perhaps alleviates metal toxicity to plants by converting them to insoluble forms as shown by the equation below [116].
CaCO3 + H2O ➜ Ca2+ + 2HCO3- + 2OH-
Al+3 + 3OH- ➜ Al(OH)3 (insoluble)
H+ + OH- ➜ H2O
However, it is important to keep in mind that over-liming can change soil physical, chemical, and biological properties, resulting in a situation where plants can suffer most notably from deficiencies of mineral nutrients such as Fe, Mn, Cu and Zn [117,118]. This is because the alkaline pH not only reduces uptake of toxic HM elements but also the uptake of essential mineral elements in plants. It is also worth mentioning that metal hydroxides including Al(OH)3 can induce generation of ROS if sprayed to plants [119].
3.3. Rhizosphere chemistry and HM chelation
Plants’ rhizosphere chemistry plays an important role in HM-plant interactions. For instance, under normal growing conditions, roots secrete organic molecules such as amino acids (e.g., methionine, lysine, and histidine) and organic acids (e.g., oxalic acid, citric acid, malic acid, tartaric acid, and succinic acid) that can bind with HMs and convert them to non-toxic forms [120,121]. It was shown that under Cd stress conditions, the total amount of organic acids secreted by the roots of Cd-tolerant rice varieties was ≥2 times higher than that of Cd-sensitive varieties [122]. Roots of a non-crop plant species grown in nutrient solutions containing Pb, Zn, Cu, and Cd salts secreted oxalic and malic acids in the media that made the plants more tolerant to the toxicity of these elements, suggesting chelation of HMs by these organic molecules [123]. The root-secreted organic molecules also provide nutrient resources to rhizosphere microbial populations to generate metabolites that can bind with the HMs and prevent them from root absorption [124]. For instance, a wide range of beneficial as well as pathogenic bacterial and fungal populations produce organic acids such as gluconic acid, oxalic acid, acetic acid, and malic acid as natural chelating agents for HM detoxification [125].
4. Key mechanisms of plant growth inhibition by HMs
Plants absorb HMs by roots from the soil solutions in the form of ions and transport them to various subcellular compartments through a diverse set of ion channels and transporter proteins such as HM ATPase, ATP binding cassette transporter, and cation diffusion facilitator [121]. If HMs exert detrimental effects on root growth, it will affect water balance and mineral nutrient uptake and translocation to the above ground shoot, causing negative impact on plant growth, biomass accumulation, and productivity.
On the other hand, if the concentration of HMs exceeds certain limits in the plants, they will affect cellular ionic homeostasis across membranes; structure and function of cell organelles (chloroplasts, mitochondria, nucleus, and vacuoles), and macromolecules (carbohydrates, lipids, proteins, and nucleic acids); and physiological, biochemical, and molecular processes in plants [11,79,80,126,127,128,129,130,131]. For instance, elevated levels of HMs have been shown to negatively affect chloroplast fine structure, chlorophyll a/b ratios, biosynthesis of photosynthetic machinery, pigment composition in grana and stroma membranes, and the activities of catalytic enzymes and non-catalytic proteins associated with various metabolic and developmental processes. The following sub-sections focus on key mechanisms of plant growth inhibition by HMs.
4.1. Generation of oxidative stress
Plants respond to toxic levels of HMs by overproduction of reactive oxygen species (ROS) such as superoxide radical (O2-), hydrogen peroxide (H2O2), hydroxyl radical (OH-), and singlet oxygen (1O2) at several sites including mitochondria, chloroplasts, peroxisomes, and at the extracellular side of the plasma membrane [132,133,134]. Although the redox active (Cr, Cu, Mn, Fe) and non-redox active (Cd, Ni, Hg, Zn, Al) elements generate ROS by different mechanisms, the generated ROS induce oxidative stress in plants leading to a variety of damages to cellular macromolecules including lipids, proteins, and nucleic acids. Some of the key consequences of cellular oxidative damage include lipid peroxidation; protein carbonylation, chain oxidation, misfolding, and aggregation; and breaks in DNA double-strands [128,130,135,136,137,138,139,140,141,142].
While in some cases, ROS generated in plants due to biotic or abiotic stresses including HMs can provide protection against certain fungal diseases [119,143], plants develop an antioxidant defense system involving ROS scavenging enzymes such as superoxide dismutase, catalase, peroxidases, and glutathione reductase that can dissipate ROS [144] and can protect plants from oxidative damage. However, certain HMs can disrupt the activity of some enzymes involved in defense responses [138,145,146]. Thus, the information generated above suggests that the contamination of elevated levels of HMs in agricultural soils and accumulation at toxic levels in crop plants can affect crop health and productivity not only by inducing oxidative stress but also by disrupting the antioxidant defense system in plants. However, further research is needed to clearly understand the specific relationship between HM stress and the antioxidant responses in plants.
4.2. Inhibition of photosynthetic phosphorylation
In the light-dependent reactions of photosynthesis, most HMs interfere with primary photochemistry, resulting in the inhibition of photosynthetic electron transport and phosphorylation. These effects were demonstrated in isolated chloroplasts, thylakoid membranes, and photosystem II (PSII) submembrane fractions [128,147,148]. PSII mediated electron transport was suggested to be more affected by these elements compared to photosystem I (PSI) mediated electron transport. However, the actual mechanism of inhibition of PS II by HMs was unclear except that the oxygen-evolving complex (OEC) of PSII was suggested to be the probable target site [149]. By measuring the variable fluorescence of intact PSII membrane preparations in the presence of Pb2+ and other additives, it was suggested that the principal site of action of HMs was located on the water oxidizing side (WOS) of PSII [147]. The kinetics of variable fluorescence rise diminished as a function of Pb2+ concentration, suggesting that the electron transport on the WOS of PSII was inhibited by Pb2+.
The above observation was further verified by immunoblotting with antibodies recognizing three extrinsic polypeptides of molecular masses, 16, 23, and 33 kDa associated with oxygen evolution of PSII. This study showed that the tested HM elements (Pb2+ and Zn2+) selectively dissociated the above polypeptides from OEC [150]. It should be noted that these three polypeptides act as a shield to protect OEC from exogenous reductants in PSII submembrane preparations. Since depletion of these three extrinsic polypeptides from OEC by either HMs or detergent treatments can inactivate PSII [147,151,152,153], the generation of ATP, and the high-energy reducing agent, NADPH through noncyclic phosphorylation (to facilitate reduction of CO2 to carbohydrate) is also expected to be inhibited (Figure 4). However, it is possible that in the absence of fully functional non-cyclic phosphorylation, plants can utilize PSI mediated cyclic electron flow to partially generate ATP to continue CO2 fixation at a reduced rate [154]
4.3. Inactivation of enzyme activities
4.3.1. Inactivation of soil enzyme activities
Enzymes in soil originate mostly from microorganisms and plants. The activities of these enzymes are a sensitive bioindicator of soil physical and chemical properties including nutrient cycling. These enzymes carry out many catalytic processes in the soil including decomposition of OMs to release mineral nutrients for plants. Some oxidoreductase enzymes such as dehydrogenases, nitrate reductase, catalase and peroxidases are involved in the degradation of many organic contaminants in the soil [155].
HMs inhibit the activities of many soil-associated enzymes involved in the transformation of carbon, nitrogen, phosphorus, and sulfur [156,157]. For example, the activities of catalase, urease, invertase, and phosphatase were inhibited in the soil upon addition of Pb, Zn, and Cu resulting in the reduction of growth and grain yield of barley [156]. Another study reported that the activity of seven enzymes was significantly reduced in response to soil contamination of Pb, Zn, Cd, Cu and As, and the order of inhibition of these enzymes was ranked as follows: arylsulfatase > dehydrogenase > β-glucosidase > urease > acid phosphatase > alkaline phosphatase > catalase [158]. This study further noted that clay-content and soil-depth negatively impacted the HM inhibition of soil enzyme activities. HM-induced inhibition of enzyme activities in the soil can occur due to multiple reasons such as formation of HM-substrate complexes, interaction of HMs with the enzyme-substrate complexes, binding of HMs with the active sites of the enzymes, denaturation of enzyme proteins, and interference with the growth of microbial populations involved in the synthesis of soil-borne enzymes [159,160].
4.3.2. Inactivation of plant enzyme activities
HMs interfere with cellular metabolic and developmental processes by inactivation of numerous enzymes and proteins in plants by binding to their active sites and functional groups such as carboxyl, amino, carbonyl, and sulfhydryl groups [130,146,161]. For instance, certain HMs inhibit the activities of enzymes involved in carbohydrate and phosphorus metabolism in plants (e.g., ribulose-1,5-biphosphate carboxylase, rubisco; phosphoenolpyruvate carboxylase; phosphoribulokinase; aldolase; fructose-6-phosphate kinase; fructose-1,6-bisphosphatase; NADP+- glyceraldehyde-3-phosphate dehydrogenase; carbonic anhydrase; and phosphatases) through conformational modifications by binding to their functional side chains [162,163,164]. Because of their strong affinity for -SH group, some HMs inhibit photosynthetic and water channel proteins by disrupting the disulfide bonds responsible for their structure and activity [8,79,165,166,167]. Some HMs inhibit the folding of nascent proteins causing aggregation in living cells. Nickel and Cd can make proteins non-functional by structural modifications such as unfolding which is corrected by plant chaperone system [168]. Bivalent Zn can inactivate rubisco activity by replacing bivalent Mg from the active site [169]. Lead and Zn can inactivate the water oxidizing enzyme of PSII by depleting Mn from the tetra-Mn-complex along with 33kDa extrinsic polypeptide [150]. Thus, HM induced enzyme inactivation and protein denaturation can cause multiple disturbances in crop plants affecting growth and crop productivity [141].
4.4. Genetic modifications
4.4.1. Effects on DNA metabolism
HMs are genotoxic but perhaps not mutagenic as there has been no report suggesting that HMs can induce gene mutations in plants, although Cr6+ is believed to be mutagenic to mammalian cells [170]. It is to be noted that all mutagenic substances are genotoxic, but not all genotoxic substances are mutagenic [171]. As discussed earlier, HMs can damage DNA molecules through generation of ROS in both plants and animals. They can also impair DNA replication and repair by inactivating enzymes involved in these processes [172]. For instance, As inhibits Poly-(ADP-ribose) polymerase-1 in humans, which is involved in the process of DNA-breakage repair caused by oxidative stress [173,174]. HMs such as Cd, Hg, and Pb can exert genotoxic effects on plants causing various types of lesions in DNA molecules. Elevated levels of Cd and Pb induce significant breakages in DNA double-strand, causing genome instability in faba bean (Vicia faba). Soil contaminated with Hg, Pb, Cu, Cd, and Zn caused increased levels of chromosomal abnormalities such as bridges, laggards, stickiness, and fragmentation in chickpea (Cicer arietinum). Certain HMs can damage DNA molecules by binding to phosphate backbones or nucleobases causing cleavage of DNA molecules. Mercury can form covalent bonds with DNA molecules resulting in the induction of sister chromatid exchange, decrease in mitotic index, and increase in the frequency of chromosomal aberrations [175,176].
4.4.2. Effects on gene expression
HMs regulate up or down expressions of many genes including the members of metal ATPase (HMA2, HMA3, and HMA4), metal transporter (ZIPs, MTPs, NRAMPs, ABCs), signal transduction (MAPKs), and metabolism-related families [177,178]. They can affect gene expression through inactivation of transcription factors (TFs) by replacing metallic components from metal-containing TFs. For example, Cd toxicity to Zn-finger TF was significantly reduced in the presence of sufficiently high level of Zn2+, suggesting protective effect of Zn2+ against Cd toxicity to Zn finger TFs [179]. TFs of diverse families modulate plant responses to HM toxicity through positive or negative regulations of stress responsive genes [175,180]. For instance, barley plants overexpress dehydration related TFs to protect against toxic effects of Cd and Hg [10]. However, it was also shown that constitutive overexpression of some genes caused enhanced uptake of HMs in plants. Some genes expressed in response to metal exposure are encoded for proteins that perform membrane transport function for HM sequestration. Genetic modification of plants with such genes may be useful to enhance phytoremediation efforts in HM-contaminated soil. Plant cells also utilize various molecular mechanisms such as signal transduction, gene overexpression, RNA processing and transport, and post-translational modifications to counter the toxic effects of HMs and other stress factors in plants [10,181,182,183].
4.5. Hormonal deregulation
Among the plant hormones, auxins, cytokinins, gibberellins, abscisic acid (ABA), and ethylene are predominantly involved in growth regulations. However, ABA and ethylene also participate in stress responses along with defense hormones, salicylic acid (SA), jasmonic acids (JA), and brassinosteroids (BSs) [184,185]. Several investigations involving different crop species demonstrated that exogenous application of some hormones can partially alleviate the toxic effects of certain HMs on selected plant growth parameters, suggesting that HMs might exert negative impact on endogenous hormone levels in plants [186,187]. Several examples are cited below.
Exogenous application of kinetin (a cytokinin) was shown to reduce the inhibitory effects of Cd on several physiological parameters of pea plant, Pisum sativum (Table 9), suggesting that kinetin might have alleviated the toxic effect of Cd [188]. Exogenous GA3 reduced the inhibitory effects of Cd and Pb on soluble protein contents in both broad bean (Vicia faba) and lupin (Lupinus albus), suggesting that GA3 might have relieved the toxic effects of Cd and Pb on these vegetable species [189]. The auxin-induced alleviation of Cd toxicity in Arabidopsis thaliana was suggested to be due to increase in the level of hemicellulose content in the cell that fixed Cd within the cell wall and lessened Cd translocation from root to shoot [190]. Wheat plants treated with Cd exhibited a significant reduction in growth, pigment content, and the activities of antioxidant enzymes (superoxide dismutase, catalase, and peroxidase); however, pretreatment of these plants with indole acidic acid (IAA) or SA remarkably reduced Cd toxicity. These observations suggest that IAA and/or SA enhanced the antioxidant defense activities in Cd stressed wheat [191]. Treatment of tomato plants with BSs partially alleviated the toxic effect of Cd on growth and photosynthetic activity, which was suggested to be due to BSs-induced improvement of antioxidant activity in plants [192]. Based on these and similar observations in other studies, it can be assumed that the defense hormones (SA and BSs) can stimulate antioxidant defense system in plants in conjunction with other growth hormones (auxins and gibberellin) when exposed to HM toxicity [186,193,194].
In contrast, HMs can also influence the level of ABA, the negative growth regulator in plants. For example, numerous studies have shown that the level of endogenous ABA is elevated in the tissues of different plant species when exposed to the toxic levels of HMs such as Cd, Hg, Cu, Zn, Pb, and Ni [187,195]. Molecular analysis of plant tissues exposed to HMs showed strong expression of ABA biosynthesis genes and up-regulation of several ABA signaling genes [187]. Based on the above reports, it can be assumed that ABA perhaps coordinates protection against HM toxicity in plants. However, a study conducted utilizing ABA-deficient and ABA-sensitive mutants failed to establish such relationships at least at early stages of plant growth [196]. Thus, it is unclear as to how HMs and ABA interact with each other in plants although it has been established that ABA strongly reduced the phytotoxicity of HMs in plants [197,198]. Because ABA acts as a negative regulator of plant growth, it might be possible that ABA-induced inhibition of plant growth restricts HM translocation in plants.
While the exact mechanism of interactions between HMs and different growth and defense hormones is not fully understood, based on the reports presented above it can be assumed that plant hormones can modify HM toxicity in plants. Further research can help better establish the links between hormone signaling pathways and metal-binding ligands in plants.
4.6. Inhibition of soil microorganisms
Beneficial soil microorganisms including bacteria, fungi, actinomycetes and several others are indispensable components for crop productivity. They contribute to soil fertility and crop health in many different ways such as releasing nutrients from organic matters, recycling plant nutrients, and fixing atmospheric nitrogen to facilitate plant uptake; producing hormones, enzymes, and secondary metabolites to promote plant growth; degrading pesticides and other pollutants in the soil; controlling soil-borne pathogens by colonizing around plant roots to form physical barrier; producing antibiotics to inhibit pathogenic microbes; and improving soil physical structure to sustain agroecosystem [199,200].
Although at low concentrations, certain HMs can stimulate growth, at elevated levels they severely inhibit growth, proliferation, and diversity of soil microbial populations involved in beneficial activities stated above, thus indirectly affecting the crop health and productivity. Previous studies indicated that microorganisms are in general more sensitive to HM toxicity than other living organisms including plants growing in the same edaphic environment. However, the degree of toxicity of HMs to different microbial groups can also vary because it is dependent on inherent toxicity of the HM elements involved and their bioavailability in the soil [103,201,202,203,204]. Furthermore, some plant growth promoting bacterial species (e.g., Pseudomonas, Arthrobacter, Rhodococcus, Mesorhizobium, Agrobacterium, Bacillus, Azoarcus, Azospirillum, Azotobacter, Burkholderia, Klebsiella, Alcaligenes, Serratia, Rhizobium, and Enterobacter) are naturally tolerant to high concentrations of HMs for which they can be used in bioremediation of HM contaminated soils provided the soil conditions are favorable for their growth and proliferation [205,206].
HMs negatively affect soil microbial populations through enzyme/protein denaturation and destruction of cell membrane integrity. Many studies reported that HMs impair substrate utilization in enzyme-catalyzed reactions particularly during microbial respiration. For instance, when microbial growth media were amended with Zn, Cu, and Pb, the evolution of CO2 was significantly reduced; however, when the media were supplied with adequate source of organic carbon, the negative effects on respiration was substantially reduced, confirming the effects on microbial respiration [203,207]. Despite some conflicting reports, the microbiological characteristics such as basal soil respiration; activities of dehydrogenase enzymes, which are inactive outside microbial cells; and quantification of phospholipid fatty acid (PLFA) molecule, which is decomposed upon microbial cell death can be used as sensitive indicators to approximately determine HM contamination in agricultural soils [208]. However, for precise determination of microbial diversity in HM contaminated soils, various molecular techniques including restriction fragment length polymorphism (RFLP) and sequence analysis of microbial genetic constituents such as 16S and 18SrRNA can be carried out [209].
5. Visual toxicity symptoms of HMs in plants
Numerous studies have shown that at elevated levels HMs induce oxidative stress, raise endogenous ABA levels, and interfere with many physiological and metabolic processes causing various growth abnormalities in plants including crop plants [3,187,210]. Because these abnormalities often resemble nutrient deficiencies as well as damages caused by diseases or pesticides [20,130,211,212,213,214,215], here we listed toxicity symptoms of non-essential HM elements that do not produce deficiency symptoms in plants (Table 10). Some of the common visible symptoms of these HM elements include chlorosis, inhibition of seed germination, stunting of root and shoot growth, reduction of biomass accumulation and yield, and occasional death of plants. Based on degree of plant toxicity, these HMs can be tentatively ranked as follows: Cd> As> Pb> Hg> Cr [216].
6. Conclusions and perspectives
HMs are non-decomposable elements. Their contamination in agricultural soil is therefore a major threat to sustainable crop production in agriculture worldwide. The toxic effects of HMs on crop plants would become visible when their concentrations exceed threshold limits in the soil and tolerance levels inside crop plants. The bioavailability of HM elements toward crop uptake generally increases with acidic pH, lower OMs, and lower CEC of the soil. The HM induced reduction in plant growth and crop productivity can be attributed directly to, among others, the generation of oxidative stress; perturbation of ion homeostasis and water balance; decrease in mineral nutrient uptake and assimilation; reduction in photosynthetic rate; inhibition of enzyme activities; hormonal deregulations; and indirectly to the inhibition of beneficial microbial growth and proliferation in the soil.
However, plants have three major lines of defense against HM toxicity. The first line of defense involves prevention of HMs from entering the plants by forming extracellular complexes with root secreted organic molecules (e.g., amino acids, oxalic acid, citric acid, malic acid, tartaric acid, and succinic acid) or with root secreted secondary metabolites (e.g., flavonoids, phenolics, alkaloids, and other S- and N-containing compounds), or with microbial metabolites at the rhizosphere. The second line of defense includes chelation of HMs with carboxyl, hydroxyl, amino, and aldehyde groups of cellulose, hemicellulose, pectin, and proteins and compartmentalization inside cell vacuoles or osmotic adjustment of HMs by soluble sugars and proteins in plants. The third line of defense involves detoxification of generated ROS by antioxidant defense system in plants. Thus, if plants fail to properly execute the above defense barriers against HMs or if HMs could overcome these barriers, then the latter can greatly harm growth and productivity of plants including crop species.
To minimize crop injury from suspected HM buildup in agricultural soil, it is important that producers keep good records of application of pesticides, chemical fertilizers, livestock manures and associated composts, and sewage-sludge based biosoilds, as well as farmland irrigation. They should follow proper tillage practices (e.g., conservation tillage can increase soil acidity [243]) and crop rotation, maintain good OM levels in the soil, and adhere to judicial use of chemical fertilizers and pesticides in the farmlands. Since the growth of microbial populations is considered as one of the most sensitive indicators of monitoring metal toxicity determination, growers can test their soil samples by accredited commercial microbiological laboratories to determine microbial growth, diversity, and biomass. Because certain soil related factors, particularly pH, OM content, and CEC stated above can influence HM solubility and bioavailability for plant uptake, producers can also test their soil samples by certified chemical laboratories if crop injury is suspected from HM toxicity. However, prior to performing the above expensive tests, it is advisable to conduct a simple soil bioassay test utilizing HM sensitive crop cultivars following a similar procedure described in [215]. If HM contamination is confirmed from the afore-mentioned tests, it is important to follow proper remediation procedures.
For research-community engaged in molecular agriculture, it is important to develop HM tolerant crop cultivars by genetic manipulations of endogenous metal binding genes and the genes of antioxidant defense system of the target crop species. In addition, development of microbial biosensors for rapid detection of contaminated soil and the degree of contamination is also important for sustainable soil health and improved crop production. To facilitate research in the above fields, availability of funding is crucial.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Maximum allowable concentrations of HM elements in agricultural soil in several countries. Table S2: HM contamination in the soil of arable lands through suspected agricultural practices. Table S3: A summary of concentration fold differences of several HM elements in the edible plant parts of a number crop and vegetable species reported in some studies. Table S4: Maximum allowable limits of certain HM elements in food plants and plant-derived food commodities recommended by Codex Alimentarius commission. References [244,245,246,247] are cited in Supplementary Materials.
Conflicts of Interest
Authors declare no conflicts of interest.
References
- Pourret, O. , Hursthouse A. It’s Time to Replace the Term “Heavy Metals” with “Potentially Toxic Elements” When Reporting Environmental Research. Int. J. Environ. Res. Public Health, 2019, 16, 4446–4451. [Google Scholar]
- Maksymiec, W. Signaling responses in plants to heavy metal stress. Acta Physiol Plant. 2007, 29: 177–187.
- Shahid M., Khalid S., Abbas G., Shahid N., Nadeem M., Sabir M., Aslam M., Dumat C. Heavy Metal Stress and Crop Productivity. 2015. Hakeem, HR (ed.), Crop Prdo. Global Envi. Issues, Springer Inter. Publishing, Switzerland.
- Wuana, R.A. , Okieimen F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. Intern. Sch. Res. Net. Ecol 2011, 402647, 1-20.
- Ghori N.H., Ghori T., Hayat M.Q., Imadi S.R., Gul A., Altay V., Ozturk M. Heavy metal stress and responses in plants. Int. J. Envn. Sci. Tech. 2019. 16, 1807-1828.
- Ali, H. , Khan E., Ilahi I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. 2019. J. Chem 6730305, 1-14.
- Tóth, G. , Hermann T. , Da Silva M.R., Montanarella C. Heavy metals in agricultural soils of the European Union with implications for food safety. Envirn. Intern. 2016, 88, 299–309. [Google Scholar]
- Tiwari, S. , Lata C. Heavy Metal Stress, Signaling, and Tolerance Due to Plant-Associated Microbes: An Overview. 2018. CSIR-National Botanical Research Institute, Lucknow, India.
- Rai, P.K. , Leeb S.S., Zhangc, Tsangd UF, Kime KH. Heavy metals in food crops: Health risks, fate, mechanisms, and management. 2019. Environ. Inter. 2019 125, 365–385.
- Singh, S. , Parihar, P., Singh, R., Singh, V.P., and Prasad, SM. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016. 6, 1143.
- Bashir K, Rasheed S, Kobayashi T, Seki M, Nishizawa NK. Regulating Subcellular Metal Homeostasis: The Key to Crop Improvement. 2016. Front. Plant Sci. 2016. 7, 1192, 1-9.
- Järup, L. Hazards of heavy metal contamination. British Medical Bulletin. 2003. 68, 167–182. [CrossRef]
- Sharma RK, Agrawal M, Marshall, FM. Atmospheric deposition of heavy metals (Cu, Zn, Cd and Pb) in Varanasi City, India. Environ. Monit. Assess 2008. 142, 269–278.
- Kumar, K. , Singh D.P., Barman S.C., Kumar N. Heavy Metal and Their Regulation in Plant System: An Overview. Chapter 2, Singh et al. (eds.), Plant Responses to Xenobiotics, Springer. Nature Singapore Pte Ltd. 2016. 19-38.
- Hasnine, M.T. , Huda M.E., Khatun R., Saadat A.H.M., Ahasan M., Akter S., Uddin M.F., Monika A.N., Rahman M.A., Ohiduzzaman M. Heavy Metal Contamination in Agric. Soil at DEPZA, Bangladesh. Envir. Ecol. Res. 2017. 5, 510-516.
- Ahmed, M. , Matsumoto M., Ozaki A., Thinh N.V., Kurosawa K. Heavy metal Contamination of Irrigation Water, Soil, and Vegetables and the Difference between Dry and Wet Seasons Near a Multi-Industry Zone in Bangladesh. Water 2019. 11, 583, 1-12.
- Arao T., Ishikawa S., Murakami M., Abe K., Maejima Y., Makino T. Heavy metal contamination of agricultural soil and countermeasures in Japan. Paddy Water Environ. 2010. 8, 247–257.
- Gebreyesus, S.T. Heavy Metals in Contaminated Soil: Sources & Washing through Chemical Extractants. Am. Sci. Res. J. Eng. Tech. Sci. (ASRJETS). 2014. 10, 54–60.
- Obinnaa, I.B. , and Ebere E.C. Water pollution by heavy metal and organic pollutants: Brief review of sources, effects, and progress on remediation with aquatic plants. Anal. Meth. Envirn. Chem. J. 2019,2, 25-38.
- Kumar, V. , Singh J., Kumar P. Heavy metals accumulation in crop plants: Sources, response mechanisms, stress tolerance and their effects. In: Kumar, V., Kumar, R., Singh, J. and Kumar, P. (eds) Contaminants in Agriculture and Environment: Health Risks and Remediation. Agro Environ Media, Haridwar, India, 2019. 1, 38-57.
- Ritche, H. , Roser M., Rosado P. Fertilizer consumption, 1961 to 2019. World in data. Source: Food and Agriculture Organization of the United Nations via the United States Department for Agriculture (USDA); 2022 (OurWorldInData.org). https://ourworldindata.org/fertilizers.
- Gimeno-Garcíaa, E.; Andreua, V.; Boluda, R. Heavy metals incidence in the application of inorganic fertilizers and pesticides to rice farming soils. Environ. Pollution. 1996. 92, 19–25.
- Mar, S.S. , Okazaki M., Motobayashi T. The influence of phosphate fertilizer application levels and cultivars on cadmium uptake by Komatsuna (Brassica rapa L. var. perviridis). Soil Sci. Plant Nutr. 2012. 58, 492—502.
- Wei, B. , Yu J. , Cao Z., Meng M., Yang L., Chen Q. The Availability and Accumulation of Heavy Metals in Greenhouse Soils Associated with Intensive Fertilizer Application. Int. J. Environ. Res. Public Health. 2020, 17, 5359. [Google Scholar]
- Thomas, E.Y. , Omueti J.A.I. Ogundayomi O. The Effect of Phosphate Fertilizer on Heavy Metal in soils and Amaranthus caudatus. Agric. Biol. J. N. Am., 2012. 3: 145-149.
- Zhuang, Z. , Mu H., Fu P., Wan Y., Yu Y., Wang Q., Li H. Accumulation of potentially toxic elements in agricultural soil and scenario analysis of Cd inputs by fertilization: A case study in Quzhou county. J. Environ. Mgmt. 2020, 269, 110797.
- Satarug, S. , Baker J.R., Urbenjapol S., Haswell-Elkins M., Reilly P.E.B., Williams D.J., Moore M.R. A global perspective on cadmium pollution and toxicity in non-occupationally exposed populations. Toxicol. Letters. 2003. 137, 65 – 83.
- Atafar, Z. , Mesdaghinia A., Nouri J., Homaee M., Yunesian M., Moghaddam M., Mahvi A.H. Effect of fertilizer application on soil heavy metal conc. Environ Monit Assess. 2010.160: 83–89.
- Al-Shawi, A.W. , Dahl R. The determination of cadmium and six other heavy metals in nitrate/phosphate fertilizer solution by ion chromatography. Anal. Chim. Acta, 1999. 391, 35–42.
- Samreen, S. , Kausar, S. Phosphorus Fertilizer: The Original and Commercial Sources. In: Phosphorus. 2019. Ed. Zhang T. InterTech Open. https://www.intechopen.com/chapters/64614.
- Paz, C.G. et al., Fertilizer Raw Materials. In: Chesworth, W. (eds) Encyclopedia of Soil Science. Encyclopedia of Earth Sciences Series. 2008. Springer, Dordrecht. [CrossRef]
- Luo L., Maa Y., Zhang S., Wei D., Zhu Y-G. An inventory of trace element inputs to agricultural soils in China. J. Environ. Mgmt. 2009. 90, 2524–2530.
- McNalley, P. Heavy Metals in Fertilizers. Data collection: MNDA. EH: Minnesota Dept. Health. 2020. https://www.health.state.mn.us/communities/environment/risk/studies/metals.html.
- Oerke, W.C. Crop losses to pests. J. Agric. Sci. 2006. 144, 31–43. [CrossRef]
- Tudi, M. , Ruan H. D., LiWang L.L., Sadler R., Connell D., Chu C., Phung D.T. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health. 2021, 18, 1112. [Google Scholar]
- PPDB (Pesticides Properties Data Base). 2007. International Union of Pure and applied Chemistry, UPAC. http://sitem.herts.ac.uk/aeru/ppdb/en/atoz.htm.
- Lewis K.A., Tzilivakis J., Warner D, Green A. An international database for pesticide risk assessments and management. J. Human and Ecol. Risk Assess. 2016. 22, 1050–1064.
- Defarge, N. , de-Vendômois S., Séralinia GE. Toxicity of formulants and heavy metals in glyphosate-based herbicides and other pesticides. Toxicol. Reports. 2018. 5, 156–163.
- Alnuwaiser, M.A. An Analytical Survey of Trace Heavy Elements in Insecticides. Intern. J. Anal. Chem. 2019. 815 0793, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, P. , Kumar D.K., Yadav A., Yadav K. Nanobiotechnology and its application in agriculture and food production. Nanotech. Food, Agric. Envirn. 2020. 105-134.
- Wang H., Dong Y., Yang Y., Toor G.S., Zhang X. Changes in heavy metal contents in animal feeds and manures in an intensive animal production region of China. J. Environ. Sci. 2013, 25, 2435–2442.
- Liu, W.R. , Zenga D., She L., Su W.X., He D.C., Wua G.Y., Ma X.R., Jiang S., Jiang C.J., Ying G.G. Comparisons of pollution characteristics, emission situations, and mass loads for heavy metals in the manures of different livestock and poultry in China. Sci. Total Environ. 2020.734, 139023.
- NAS (National Academy of Science). Arsenic: Medical and Biologic Effects of Environmental Pollutants. 5. Biologic Effects of Arsenic on Plants and Animals. 1977. 5, 1-332. Washington (DC): National Academies Press, US. https://www.ncbi.nlm.nih.gov/books/NBK231025/. 2310.
- Hu Y., Zhang W., Chen G., Cheng H., Tao S. Public health risk of trace metals in fresh chicken meat products on the food markets of a major production region in southern China. Environ. Pollut. 2018, 234, 667–676.
- Zhang F., Yanxia Li., Yang M., Wei L. Content of Heavy Metals in Animal Feeds and Manures from Farms of Different Scales in Northeast China. Int. J. Environ. Res. Public Health. 2012, 9, 2658–2668.
- Jensen J., Larsen M. M., Bak J. National monitoring study in Denmark finds increased and critical levels of copper and zinc in arable soils fertilized with pig slurry. Environ. Pollut. 2016, 214, 334–340.
- Lopes, C. , Herva M., Franco-Uría A., Roca E. Inventory of heavy metal content in organic waste applied as fertilizer in agriculture: evaluating the risk of transfer into the food chain. Environ. Sci. Pollut. Res. 2011; 18, 918e939. [Google Scholar]
- Zhao Y., Yan Z., Qin J. Effects of long-term cattle manure application on soil properties and soil heavy metals in corn seed production in NW China. Environ Sci. Pol. Res. 2014. 21, 7586–7595.
- Yang X.P., Li Q., Tang Z., Zhang W.W., Yu G.H., Shen Q.R., Zhao F.J. Heavy metal conc. and arsenic speciation in animal manure composts in China. Waste Mgmt. 2017. 64, 333–339.
- Zarcinas, B.A. , Ishak C.E., McLaughlin M.J., Cozens G. Heavy metals in soils and crops in southeast Asia. 1. Peninsular Malaysia. Environ. Geochem. Health. 2004. 26: 343–357.
- MDEQ bulletin. What are biosolids, how are they used, and are they safe? Water Resources Division, Michigan Department of Environmental Quality. 2014. 22014. https://www.michigan.gov/-/media/Project/Websites/egle/Documents/Programs/WRD/Biosolids/biosolids-what-how-safe.
- Kissel, K.E. , Rodriguez L., Paz J., Mitchell C., Patrick S., Zhang H., Fielder K., Morris L. Metal Conc. Standards for Land Application of Biosolids and Other By-Products in Georgia. Bulletin. 2017. 1353, 1-4 (Source: EPA: 40 CFR, Part 503).
- Silveira, M.L.A. , Alleoni L.R.F., Guilherme L.R.G. Biosolids and heavy metals in soils. Sci. Agric. 2003. 60, 64–111, 2003.
- McLaughlin, M.J. , Hamon R.E., McLaren R.G., Speir T.W., Rogers S.L. A bioavailability-based rationale for controlling metal and metalloid contamination of agric.land in Australia and New Zealand. Aust. J. Soil Res. 2000. 38, 1037–1086.
- EPA (Environmental Protection Agency). A Plain English Guide to the EPA Part 503 Biosolids Rule. 2023. EPA 40 CFR Part 503. https://www.epa.gov/biosolids/plain-english-guide-epa-part-503-biosolids-rule.
- Hossain, M.F. Arsenic contamination in Bangladesh. Agric. Ecosys. Environ. 2006. 113, 1–16.
- Hussain, M.M. , Hina A., Saeed A., Sabahat S., Jannat F., Aslam M. Impact of heavy metals on plants and animals in relation to sewage water: A Review. Sci. Tech. Dev. 2017. 36, 215-226.
- Berihun, B.T. , Amare D.E., Raju R.P., Ayele D.T., Dagne H. Determination of the Level of Metallic Contamination in Irrigation Vegetables, the Soil, and the Water in Gondar City, Ethiopia. Nutr. Dietary Suppl. 2021. 13, 1–7.
- Rai P.K., Tripathi B.D. Heavy metals in industrial wastewater, soil and vegetables in Lohta village, India. Toxic. Environ. Chem. 2007, 90, 247–257.
- Mohankumar, K. , Hariharan V., Rao N.P. Heavy Metal Contamination in Groundwater around Industrial Estate vs. Residential Areas in Coimbatore, India. J. Clinical Diag. Res. 2016. 10, BC05-BC07.
- Huq E.M., Su C., Lia J,, Sarven M.S. Arsenic enrichment and mobilization in the Holocene alluvial aquifers of Prayagpur of Southwestern Bangladesh. Intern. Biod. Biodeg. 2018, 128, 186–194.
- Ashfaque F., Inam A., Sahay S., Iqbal S. Influence of Heavy Metal Toxicity on Plant Growth, Metabolism and Its Alleviation by Phytoremediation - A Promising Technology. J. Agric. Ecol. Res. Inter. 2016. 6, 1-19 JAERI.23543.
- Rieuwerts, J.S. , Thornton I., Farago M.E., Ashmore M.R. Factors influencing metal bioavailability in soils: preliminary investigations for the development of a critical loads approach for metals. Chem. Spec. Bioavail. 1998. 10, 61-75.
- Jung, M.C. Heavy metal concentrations in soils and factors affecting metal uptake by plants in the vicinity of a Korean Cu-W Mine. Sensors. 2008. 8, 2413–2423. [CrossRef]
- Kacholi, D.S. , Sahu M. Levels and health risk assessment of heavy metals in soil, water, and vegetables of Dar es Salaam, Tanzania. J. Chem. 2018. 1402674.
- Njoku, K.L. , Nwani S. O. Phytoremediation of heavy metals contaminated soil from mechanic workshop and dumpsite using Amaranthus spinosus. Scien. African. 2022. 17, e01278.
- Ahn, Y. , Yun H. S., Pandi K., Park S., Ji M., Choi J. Heavy metal speciation with prediction model for heavy metal mobility and risk assessment in mine-affected soils. Environ. Sci. Poll. Res. Intern. 2020. 27, 3213– 3223.
- Maksymiec, W. , Baszynski T. Chlorophyll fluorescence in primary leaves of excess Cu-treated runner bean plants depends on their growth stages and the duration of Cu action. J Plant Physiol. 1996. 149, 196–200.
- Peralta-Videaa J.R., de la Rosaa G., Gonzalez J.H., Gardea-Torresdey J.L. Effects of the growth stage on the heavy metal tolerance of alfalfa plants. Adv. Environ. Res. 2004. 8, 679-685.
- Regassa, G. , Chandravanshi B. S. Levels of heavy metals in the raw and processed Ethiopian tobacco leaves. Springer Plus. 2016. 5, 232.
- Zayed, A. , Mel Lytle C., Qian J-H., Terry N. Chromium accumulation, translocation and chemical speciation in vegetable crops. Planta. 1998. 206: 293-299.
- Memon, A.R. Heavy Metal–Induced Gene Expression in Plants. In: Naeem, M., Ansari, A., Gill, S. (eds) Contaminants in Agriculture. 2020. Springer, Cham. [CrossRef]
- Shumaker, K.L. , Begonia G. Heavy Metal Uptake, Translocation, and Bioaccumulation Studies of Triticum aestivum Cultivated in Contaminated Dredged Materials. Int. J. Environ. Res. Public Health. 2005, 2, 293–298. [Google Scholar] [PubMed]
- Alaboudi, K.A. , Ahmed B., Brodie G. Phytoremediation of Pb and Cd contaminated soils by using sunflower (Helianthus annuus) plant. Annals Agric. Sci. 2018. 93, 123-127.
- Cu, N.X. Effect of Heavy Metals on Plant Growth and Ability to Use Fertilizing Substances to Reduce Heavy Metal Accumulation by Brassica Juncea. Global J. Sci. Front. Res. 2015. D Agric. Vet. 15, 1.0.
- Du, X. , Zhu Y-G., Liu Y-J., Zhao X-S. Uptake of mercury (Hg) by seedlings of rice (Oryza sativa L.) grown in solution culture and interactions with arsenate uptake. Environ. Exp. Bot. 2005. 54, 1–7.
- Satofuka, H.T. , Fukui M., Takagi H., Imanaka A.T. (2001). Metal-binding properties of phytochelatin-related peptides. J Inorg Biochem. 2001. 86, 595-602.
- Cobbett, C. , Goldsbrough P. (2002). Phytochelatins and metallothionines: Roles in Heavy Metal Detoxification and Homeostasis. Ann. Review Plant Biol. 2002. 53, 159.
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. SA J. Bot. 2010. 76, 167–179. [CrossRef]
- Hasan, M.D. , Cheng Y., Kanwar M.K., Chu X-Y., Ahammed G.J., Qi Z-U. Responses of Plant Proteins to Heavy Metal Stress—A Review. 2017. Front. Plant Sci. 8, 1492.
- Grill, E. , Winnacker E.L., Zenk M.H.. Phytochelatins: The Principal Heavy Metal Complexing Peptides of Higher Plants. Science. 1985. 230, 674-676.
- Gril, E. , Winnacker E.L., Zenk M.H. Phytochelatins, a class of heavy-metal-binding peptides from plants, are functionally analogous to metallothioneins. Proc. Natl. Acad. Sci. 1987. 84, 439-443.
- Pal, R. , Rai J.P.N. Phytochelatins: Peptides Involved in Heavy Metal Detoxification. Appl. Biochem. Biotechnol. 2010. 160, 945–963.
- Jonak, J. , Nakagami H. , Hirt H. Heavy Metal Stress. Activation of Distinct Mitogen-Activated Protein Kinase Pathways by Copper and Cadmium. Plant Physiol. 2004, 136, 3276–3283. [Google Scholar] [PubMed]
- Xia, C. , Hong L., Yang Y., Yanping X., Xing H., Gang D. Protein Changes in Response to Lead Stress of Lead-Tolerant and Lead-Sensitive Industrial Hemp Using SWATH Technology. Genes. 2019. 10, 396.
- Thao, N.P. , Khan M. Q.R., Thu N.B.A., Hoang X.L.T., Asgher M., Khan N.A., Tran L.S.P. Role of Ethylene and Its Cross Talk with Other Signaling Molecules in Plant Responses to Heavy Metal Stress. Plant Physiol. 2015, 169, 73–84. [Google Scholar]
- Maksymiec, W. , Wianowska D. , Dawidowicz A.L., Radkiewicz S., Mardarowicz M., Krupa Z. The level of jasmonic acid in Arabidopsis thaliana and Phaseolus coccineus plants under heavy metal stress. J. Plant Physiol. 2005, 162, 1338–1346. [Google Scholar]
- Amari T., Ghnaya T., Abdelly C. Nickel, cadmium and lead phytotoxicity and potential of halophytic plants in heavy metal extraction. 2017, SA J. Bot. 111, 99–110.
- Yan, A. , Wang Y. , Tan S.N., Yusof L.K.M., Ghosh S., Chen1 Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 359. [Google Scholar]
- Cho U-H. , Park J-O. Mercury-induced oxidative stress in tomato seedlings. Plant Sci. 2000, 156, 1–9.
- Jin, F. , Wang C. , Lin H.J., Shen Y.O., Zhang Z.M, Zhao M.J., Pan G.T. Heavy metal-transport proteins in plants: a review. NCBI. 2010, 21, 1875–82. [Google Scholar]
- Siddique, A.B. , Rahman M. M., Islam M.R., Naidu R. Varietal variation and formation of iron plaques on cadmium accumulation in rice seedling. Environ. Adv. 2021, 5, 100075. [Google Scholar]
- Islam, M.D. , Hasan M. M., Rahaman A., Haque P., Islam M.S., Rahman M.M. Translocation and bioaccumulation of trace metals from industrial effluent to locally grown vegetables and assessment of human health risk in Bangladesh. SN Applied Sci. 2020, 2, 1315. [Google Scholar]
- Bigdeli, M. , Seilsepour M. Investigation of Metals Accumulation in Some Vegetables Irrigated with Waste Water in Shahre Rey-Iran and Toxicological Implications. Am.-Eur. J. Agric. Environ. Sci., 2008, 4, 86–92. [Google Scholar]
- Kafle, A. , Timilsina A. , Gautam A., Adhikari K., Bhattarai A., Aryal N. Phytoremediation: Mechanisms, plant selection and enhancement by natural and synthetic agents. Environ. Adv. 2022, 8, 100203. [Google Scholar]
- Chen T-B. , Zheng Y-M., Lei M., Huang Z-C., Wu H-T., Chen H., Fan K-K., Yu K., Wu X., Tian Q-Z. Assessment of heavy metal pollution in surface soils of urban parks in Beijing, China. Chem. 2005, 60, 542–551.
- Islam, M.S. , Ahmed M. K., Raknuzzaman M., Mamun MHA., Islam M.K. Heavy metal pollution in surface water and sediment: A preliminary assessment of an urban river in a developing country. Ecol. Indic. 2015, 48, 282–291. [Google Scholar]
- Blake, L. , Goulding K. W.T. Effects of atmospheric deposition, soil pH and acidification on heavy metal contents in soils and vegetation of semi-natural ecosystems at Rothamsted Experimental Station, UK. Plant Soil. 2002, 240, 235–251. [Google Scholar]
- Soil Quality Indicators: pH - Soil Quality Information bulletin. USDA Natural Resources Conservation Service. 2011.
- Chuan, M.C. , Shu G. Y., Liu J.C. Solubility of heavy metals in a contaminated soil: Effects of redox potential and pH. Water Air Soil Pollut. 1996, 90, 543–556. [Google Scholar]
- Sanders, J.R. , McGrath S. P., Adams T.M. Zinc, copper and nickel concentrations in ryegrass grown on sludge-contaminated soils of different pH. J. Sci. Food Agric. 1986, 37, 961–968. [Google Scholar]
- McLaughlin, M. Fertilizers and Soil Acidity. Fert. Tech. Res. bulletin. 2009. The Univ. of Adelaide, Australia.
- Giller, K.E. , Witter E. , Mcgarth S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar]
- Quenea, K. , Lamy, I. , Winterton, P., Bermond, A., Dumat, C. (2009). Interactions between metals and soil organic matter in various particle size fractions of soil contaminated with waste water. Geoderma. 2009, 1, 217–223. [Google Scholar]
- Gray, C.W. , Mclaren R. G. (2006). Soil factors affecting heavy metal solubility in some New Zealand soils. Water Air & Soil Pollution. 2006, 175, 3–14. [Google Scholar]
- McCauley, A. , Jones, C., and Olson-Rutz, K. Soil pH and organic matter. Nutrient management module No. 4449-8, 2017, 1-16.
- Reijonen, I. , Metzler. , Hartikainen H. Impact of soil pH and organic matter on the chemical bioavailability of vanadium species: The underlying basis for risk assessment. Environ. Pollution. 2016, 210, 371–379. [Google Scholar]
- Angelova, V. , Ivanova R., Pevicharova G., Ivanov K. Effect of organic amendments on heavy metals uptake by potato plants. 19th World Congress of Soil Science. 2010, Symposium 3.5.1. Heavy metal contaminated soil. 84-87.
- Cunningham, J.D. , Keeney, D. R., Ryan, J.A. (Phytotoxicity and Uptake of Metals Added to Soils as Inorganic Salts or in Sewage Sludge. J. Environ. Qual., 1975, 4, 455–460. [Google Scholar]
- Giordano, P.M. , Mays, D.A. Effect of land disposal applications of municipal wastes on crop yields and heavy metal uptake. Environ. Protect. Technol. Ser. 1977, EPA-600/2-77-014. USEPA, Washington, DC.
- McBride, M.B. Long-Term Biosolids Application on Land: Beneficial Recycling of Nutrients or Eutrophication of Agroecosystems? Soil Syst. 2022, 6, 1–15. [Google Scholar] [CrossRef]
- Gadd, G.M. , Griffiths A. J. Microorganisms and Heavy Metal Toxicity. Microbial Ecol. 1978, 4, 303–317. [Google Scholar]
- Ayres, D.M., Davis, A.P., Gietka, P.M. Removing Heavy Metals from Wastewater. Engineering Research Center Report. Univ. of Maryland. 1994, 1-21.
- Prokkola, H. , Nurmesniemi, E-T. , Lassi, U. Removal of Metals by Sulphide Precipitation Using Na2S and HS Solution. Chem Eng., 2020, 4, 51, 1–10. [Google Scholar]
- Król, K. , Mizerna K. , Bożym M. An assessment of pH-dependent release and mobility of heavy metals from metallurgical slag. J. Hazard. Materials. 2020, 384, 121502. [Google Scholar]
- Gatiboni, L. , Hardy D. Soil Acidity and Liming for Agricultural Soils. SoilFacts. 2022 AG-439-50.
- Moreira, S.G. , Prochnow L. G., Pauletti V., Silva M.M., Kiehl J.C., Silva G.M.C. Effect of liming on micronutrient availability to soybean grown in soil under different lengths of time under no tillage. Acta Scient. Agro., 2017, 39, 89–97. [Google Scholar]
- Olego, M.A. , Quiroga M. J., Sánchez-García M., Cuesta M., Cara-Jiménez J., Garzón-Jimeno J.E. Effects of overliming on the nutritional status of grapevines with special reference to micronutrient content. OENO One. 2021, 1, 57–73. [Google Scholar]
- Zhang B., Wang, X.Q., Li X., Ni Y.Q., Li H.Y. Aluminum uptake and disease resistance in Nicotiana rustica leaves. Ecotoxicol. Environ. Safety. 2010 73, 655–663.
- Sharma, P. , Dubey, R. S. Toxic heavy metals in plants. Braz. J. Plant Physiol. 2005, 17, 1677–9452. [Google Scholar]
- Yu, G. , Ma J. , Jiang M., Li J., Gao J., Qiao S., Zhao Z. (2019). The Mechanism of Plant Resistance to Heavy Metal. IOP Conference Series: Earth Environ. Sci. 2019, 310, 052004 (GBEM2019). [Google Scholar]
- Fu, H. , Yu H. , Li T., Zhang X. Influence of cadmium stress on root exudates of high cadmium accumulating rice line (Oryza sativa L.). Ecotox. Environ. Safety. 2017, 150, 168. [Google Scholar]
- Chen, J. , Shafi M., Wang Y., Wu L., Ye Z., Liu C., Zhong B., Guo H., He L., Liu D. Organic acid compounds in root exudation of Moso Bamboo (Phyllostachys pubescens ) and its bioactivity as affected by heavy metals. J. Environ Sci Pollut. Res Int, 2016. 23, 20977-20984.
- Caracciolo, A.B. , Terenzi V. Rhizosphere Microbial Communities and Heavy Metals. Microorganisms. 2021, 9, 1462. [Google Scholar]
- Gajewska, J. , Floryszak-Wieczorek J. , Sobieszczuk-Nowicka E., Mattoo A., Arasimowicz-Jelonek M. Fungal and oomycete pathogens and heavy metals: an inglorious couple in the environment. IMA Fungus. 2022, 13, 6. [Google Scholar] [PubMed]
- Hossain, Z. , Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front Plant Sci. 2013, 3, 310. [Google Scholar] [PubMed]
- Argüello, J.M. , Raimunda, D., González-Guerrero, M. Metal Transport across Biomembranes: Emerging Models for a Distinct Chemistry. J. Biol. Chem. 2012. 287, 13510–13517.
- Aggarwal, A. , Sharma, L., Tripathi, B.N., Munjal, A.K., Baunthiyal, M., and Sharma, V. Metal Toxicity and Photosynthesis. In: Photosynthesis: Overviews on Recent Progress & Future Perspective. Chapter 16. 2014, pp. 229–236.
- Xu, H. , Martinoia, E. , and Szabo, I. Organellar Channels and Transporters. Cell Calcium. 2015, 58, 1–10. [Google Scholar] [PubMed]
- Asati, A. , Pichhode M. , Nikhil K. Effect of Heavy Metals on Plants. Inter. J. Appl. Inno. Eng. Mgmt. 2016, 5, 56–66. [Google Scholar]
- Caroli, M.D. , Furini, A. , DalCorso, G., Rojas, M., Sansebastiano, G.P.D. Endomembrane Reorganization Induced by Heavy Metals. Plants. 2020, 9, 482, 1–20. [Google Scholar]
- Bernia, R. , Luyckxc, M. , Xu, X., Legay, S., Sergeant, K., Hausman, J-F. Lutts, S., Caia, S., Guerriero, G. Reactive oxygen species and heavy metal stress in plants. Environ. Expt. Bot. 2019, 161, 98–106. [Google Scholar]
- Tamás, L. , Mistrík, I. , Zelinová, V., 2017. Heavy metal-induced reactive oxygen species and cell death in barley root tip. Environ. Exp. Bot. 2017, 140, 34–40. [Google Scholar]
- Smeets, K. , Cuypers A. , Lambrechts A., Semane B., Hoet P., Van Laere A., Vangronsveld J. Induction of oxidative stress and antioxidative mechanisms in Phaseolus vulgaris after Cd application. Plant Physiol. Biochem. 2005, 43, 437–444. [Google Scholar]
- van Assche, F. , Clijsters H. Effects of metals on enzyme activity in plants. Plant Cell Environ. 1990, 13, 195–206. [Google Scholar]
- Rucińiska-Sobkowiak, R. Oxidative stress in plants exposed to heavy metals. Postepy Biochem. 2010, 56: 191-200.
- Gjorgieva, D., Panovska, T.K., Ruskovska, T., BaIeva, K., Stafilov T. Influence of Heavy Metal Stress on Antioxidant Status and DNA Damage in Urtica dioica. BioMed Res. Intern. 2013, 276417, 1-6.
- Panda, S. , Chaudhury, I. , Khan, M. Heavy Metals Induce Lipid Peroxidation and Affect Antioxidants in Wheat Leaves. Biol. Plant. 2003, 46, 289–294. [Google Scholar]
- Gichner, T. , Patková Z. , Száková J,, Demnerová K. Toxicity and DNA damage in tobacco and potato plants growing on soil polluted with heavy metals. Ecotoxicol. Environ. Safety. 2006, 65, 420–426. [Google Scholar] [PubMed]
- Tamás, M.J. , Sharma, S. K., Ibstedt, S., Jacobson, S., Christen, P. 2014. Heavy Metals and Metalloids as a Cause for Protein Misfolding and Aggregation. Biomole. 2014, 4, 252–267. [Google Scholar]
- Kalaivanan, D. , Ganeshamurthy, A.N. Mechanisms of Heavy Metal Toxicity in Plants. Chapter In: Rao et al. (eds.), Abiotic Stress Physiology of Horticultural Crops. 2016, Springer, India.
- Salas-Moreno, M. , Contreras-Puentes, N. , Rodríguez-Cavallo, E., Jorrín-Novo, J., Marrugo-Negrete, J., Méndez-Cuadro, D. Protein Carbonylation as a Biomarker of Cd and Pb, Damage in Paspalum fasciculatum. Plants. 2019, 8, 513. [Google Scholar]
- Lehmann S, Serrano M. , L’Haridon F., Tjamos S.E., Metraux J-F. Reactive oxygen species and plant resistance to fungal pathogens. Phytochem. 2015, 112, 54–62.
- Dumanovic, J. , Nepovimova E., Natic M., Kuc K., Jac’evic V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 552969.
- Sai Kachout, S. , Ben Mansoura, A. , Leclerc, J.C., Mechergui, R., Rejeb. M.N., Ouerghi, Z. Effects of heavy metals on antioxidant activities of Atriplex hortenesis and A. rosea. J. Environ. Agric. Food Chem. 2010, 9, 444–457. [Google Scholar]
- Azevedo A, Rodriguez E. (2012). Phytotoxicity of Mercury in Plants: A Review. J. Bot. Article ID 848614, 1-6.
- Rashid, A., Popovic, R. Protective role of CaCl2 against Pb2+ inhibition in photosystem II. FEBS Lett. 1990, 271, 181-184.
- Rashid, A. , Bernier, M. , Pazdernick, L., Carpentier, R. Interaction of zinc with the donor side of photosystem II. Photosynth. Res. 1991, 30, 123–130. [Google Scholar]
- van Assche, F. , Clijsters, H. Multiple Effects of Heavy Metal Toxicity on Photosynthesis. In: Marcelle, R., Clijsters, H., van Poucke, M. (eds) Effects of Stress on Photosynthesis. Adv. Agric. Biotech. 1983, 3. Springer, Dordrecht.
- Rashid, A. , Camm, E. , Ekramoddoullah, A.K.M. Molecular mechanism of action of Pb++ and Zn++ on water oxidizing complex of photosystem II. FEBS Lett. 1994, 350, 296–298. [Google Scholar]
- Rashid, A. Carpentier, R. CaCl2 inhibition of H2O2 electron donation to PS II in submembrane preparations depleted in extrinsic polypeptides. FEBS Lett. 1989, 258, 331–334. [Google Scholar]
- Rashid, A. , Carpentier, R. The 16 and 23 kDa extrinsic polypeptides and the associated Ca++ and Cl- modify atrazine interaction with photosystem II core complex. Photosynth. Res. 1990, 24, 221–227. [Google Scholar] [PubMed]
- Rashid, A. Popovic, R. Electron donation to photosystem II by DPC is inhibited by both the endogenous Mn-complex and by exogenous Mn ions. Biochem. Cell Biol. 1995, 73, 241–24. [Google Scholar] [PubMed]
- Klob, W. , Kandler O. , Tanner W. The Role of Cyclic Photophosphorylation in Vivo. Plant Physiol. 1973, 51, 825–827. [Google Scholar]
- Rao, S.M. , Grover M., Kundu S., Desa S. Soil Enzymes. In book: Encyclopedia of Soil Science. 2017, Third Edition Chapter: Soil Enzymes. Publisher: Taylor & Francis. All rights reserved.
- Belyaeva, O.N. , Haynes R.J., Birukova O.A. Barley yield and soil microbial and enzyme activities as affected by contamination of two soils with lead, zinc or copper. Biol. Fert. Soils. 2005, 41: 85–94.
- Jaworska, H. , Lemanowicz J. Heavy metal contents and enzymatic activity in soils exposed to the impact of road traffic. Sci. Reports. 2019, 9, 19981. [Google Scholar]
- Aponte, A.N. , Meli P. , Butler B., Paolin J., Matus F.J., Merino K., Cornejo K., Kuzyakov Y. Meta-analysis of heavy metal effects on soil enzyme activities. Sci. Total Environ. 2020, 737, 139744. [Google Scholar]
- Dick, R.P. Soil enzyme activities as integrative indicators of soil health. In: Pankhurst CE, Doube BM, Gupta VVSR (eds) Biological indicators of soil health. CAB, Wallingford. 1997, pp 121–156.
- Yeboah, J. , Shi, G. , Shi, W. Effect of Heavy Metal Contamination on Soil Enzymes Activities. J. Geosci. Environ. Prot. 2021, 9, 135–154. [Google Scholar]
- Ajsuvakova, O.P. , Tinkova, A., Aschner M., Rochae J.B.T., Michalkef B., Skalnayaa M.G., Skalny A.V., Butnariug M., Dadari M., Saracg J., Aaseth J., Bjørklundk G. Sulfhydryl groups as targets of Hg toxicity. Coord Chem Rev. 2020, 15; 417.
- Siborova, M. (1988). Cd2+ ions affect the quaternary structure of ribulose-1,5-bisphosphate carboxylase from barley leaves. Biochem. Physiol. 1988, 183, 371–378. [Google Scholar]
- Bielen, A. , Remans T. , Vangronsveld J., Cuypers A. The influence of metal stress on the availability and redox state of ascorbate, and possible interference with its cellular functions. Intern. J. Mol. Sci. 2013, 14, 6382–6413. [Google Scholar]
- Hossain, M.A. , Piyatida, P., da Silva, J.A.T., Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 872875.
- De Filippis, L.F. , Pallaghy C.K. Heavy metals: Sources and biological effects. In: LC Rai, JP Gaur, CJ Soeder (eds.), Algae and Water Pollution. E. Schweizerbart’sche, Verlagsbuchhandlung, Stuttgart. 1994, 31-77.
- Hampp, R. , Ziegler H. , Ziegler I. Influence of Lead Ions on the Activity of Enzymes of the Reductive Pentose Phosphate Pathway. Biochem. Physiol. Pflanzen. 1973, 164, 588–595. [Google Scholar]
- Ernst, W.H.O. Biochemical aspects of cadmium in plants. In: Nriagu J.O. (ed) Cadmium in the environment, part 1. 1980, 639-653. New York: Wiley & Sons.
- Jain, S. , Muneerb M. , Guerriero G., Liu S., Vishwakarma K., Chauhan D., Dubey N., Tripathi D.K., Sharma S. Tracing the role of plant proteins in the response to metal toxicity: a comprehensive review. Plant Signal. Behav. 2018, 13, e1507401. [Google Scholar]
- Tsonev, T. , Lidon, F. J.C. Zinc in plants - An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Hamilton, J.W. , Kaltreider R. C., Bajenova O.V., lhnat M.A., McCaffrey J., Turpie B.W., Rowell E.E., Oh J., Nemeth M.J., Pesce C.A., Lariviere R. Molecular Basis for Effects of Carcinogenic Heavy Metals on Inducible Gene Expression. Environ. Health Persp. 1998, 106, Suppl 4. [Google Scholar]
- Badr, A. , El-Shazly H.H., Mohamed H.I. Plant Responses to Induced Genotoxicity and Oxidative Stress by Chemicals. Chapter, 2021. [CrossRef]
- Morales, M.E. , Rebecca J. , Derbes S., Catherine M.A., Ortego JC., Stark J., Deininger P.L., Roy-Engel A.S.M. Heavy Metal Exposure Influences Double Strand Break DNA Repair Outcomes. PLoS One. 2016, 11, e0151367. [Google Scholar]
- Ding, S. , Ma C. , Shi W., Liu W., Lu W., Liu O., Luo Z.B. Exogenous glutathione enhances cadmium accumulation and alleviates its toxicity in Populus × canescens. J. Tree Physiol. 2017, 37, 1697–1712. [Google Scholar]
- Qin, X.J. , Liu W. , Li Y.N., Sun X., Hai C.X., Hudson L.G., Liu K.J. Poly(ADP-ribose) polymerase-1 inhibition by arsenite promotes the survival of cells with unrepaired DNA lesions induced by UV exposure. Toxicol Sci. 2012, 127, 120–129. [Google Scholar] [PubMed]
- Dutta, S. , Mitra, M. , Agarwal, P., Mahapatra, K., Sett, D.D.U., and Roy, S. Oxidative and genotoxic damages in plants in response to heavy metal stress and maintenance of genome stability. Plant Signal. Behav. 2018, 13, e1460048. [Google Scholar]
- Pourrut, B. , Jean, S. , Silvestre, J., and Pinelli, E. Lead-induced DNA damage in Vicia faba root cells: potential involvement of oxidative stress. Mutat. Res. 2011, 726, 123–128. [Google Scholar]
- Memon, A.R. , Zahirovic E. Genomics and transcriptomics analysis of Cu accumulator plant Brassica nigra L. J. Appl. Biol. Sci. 2014, 8, 1–8. [Google Scholar]
- Verbruggen, N. , Juraniec, M. , Baliardini, C., Meyer, C.-L. Tolerance to cadmium in plants: The special case of hyperaccumulators. BioMetals. 2013, 26, 633–638. [Google Scholar]
- Geng, A. , Wang X. , Wu L, Wang F., Wu Z., Yang H., Chen Y., Wen D., Liu X. Silicon improves growth and alleviates oxidative stress in rice seedlings (Oryza sativa L.) by strengthening antioxidant defense and enhancing protein metabolism under arsanilic acid exposure. Ecotoxicol. Environ. Safety. 2018, 158, 266–273. [Google Scholar]
- Gwozdz, E.A. , Przymusinski R. , Rucinska R., Deckert J. Plant cell responses to heavy metals: Molecular and physiological aspects. Acta Physiol. Plant. 1997, 19, 459–65. [Google Scholar]
- Rashid, A. Defense responses of plant cell wall non-catalytic proteins against pathogens. Physiol. Mol. Plant Pathol. 2016, 94, 38–46. [Google Scholar] [CrossRef]
- Tamás, L. , Mistrík, I. , Huttová, J., Halusková, L., Valentovicová, K., Zelinová, V. Role of reactive oxygen species-generating enzymes and H2O2 during Cd, Hg and osmotic stresses in barley root tip. Planta. 2010, 231, 221–231. [Google Scholar]
- Kuriakose, S.V. , Prasad, M. N.V. Cadmium stress affects seed Germination and seedling growth in Sorghum bicolor (L.) by changing the activities of hydrolyzing enzymes. Plant Growth Regul. 2008, 54, 143–156. [Google Scholar]
- Asami, T. , Nakagawa Y. Preface to the Special Issue: Brief review of plant hormones and their utilization in agriculture. J. Pestic Sci. 2018, 43, 154–158. [Google Scholar]
- Verma, V. , Ravindran P. , Kumar P.P. Plant hormone-mediated regulation of stress responses. Review. BMC Plant Biol. 2016, 16, 86. [Google Scholar]
- Sharma, A. , Sidhu G. P.S., Araniti F., Bali A.S., Shahzad B., Tripathi D.K., Brestic M., Skalicky M., Landi M. The Role of Salicylic Acid in Plants Exposed to Heavy Metals. Molecules. 2020, 25, 540. [Google Scholar]
- Bücker-Neto, L. Paiva, A.L.S., Machado, R.D., Arenhart, R.A., Margis-Pinheiro, M. (2017). Interactions between plant hormones and heavy metals responses. Genet. Mol. Biol. 2027. 40, 373-386.
- Al-Hakimi, A.M.A. Modification of cadmium toxicity in pea seedlings by kinetin. Plant Soil Environ. 2007, 53:129-135.
- El-Monem, A.M.S. , Farghal I. I., Sofy M.R. Role of gibberellic acid in abolishing the detrimental effects of Cd and Pb on broad bean and lupin plants. Res. J. Agric Biol Sci. 2009, 5, 6–13. [Google Scholar]
- Zhu, X.F. , Wang Z. W., Dong F., Lei G.J., Shi Y.Z., Li G.X., Zheng S.J. Exogenous auxin alleviates cadmium toxicity in Arabidopsis thaliana by stimulating synthesis of hemicellulose 1 and increasing the cadmium fixation capacity of root cell walls. J. Hazard. Mater. 2013, 263, 98–403. [Google Scholar]
- Agami, R.A. , Mohamed G. F. Exogenous treatment with indole-3-acetic acid and salicylic acid alleviates cadmium toxicity in wheat seedlings. Ecotox. Environ. Safety. 2013, 94, 164–171. [Google Scholar]
- Ahammed, G.J. , Choudhury, S. P., Chen, S., Xia, X., Shi, K., Zhou, Y., and Yu, J. Role of brassinosteroids in alleviation of phenanthrene–cadmium co-contamination-induced photosynthetic inhibition and oxidative stress in tomato. J. Expt. Bot. 2013, 64, 199–213. [Google Scholar]
- Hayat, S. , Ali B. , Hasan S.A., Ahmad A. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ Exp Bot. 2007, 60, 33–41. [Google Scholar]
- Hasan, S.A. , Hayat, S. , Ahmad, A. Brassinosteroids protect photosynthetic machinery against the cadmium induced oxidative stress in two tomato cultivars. Chem. 2011, 84, 1446–1451. [Google Scholar]
- Vishwakarma, K. , Upadhyay N. , Kumar N., Yadav G., Singh J., Mishra R.K., Kumar V., Verma R., Upadhyay R.G., Pandey M., Sharma S. Abscisic Acid Signaling and Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2017, 8, 161. [Google Scholar]
- Sharma, S. , Kumar V. Responses of wild type and abscisic acid mutants of Arabidopsis thaliana to cadmium. J. Plant Physiol. 2002, 159, 1323–1327. [Google Scholar]
- Hu, B. , Deng F. , Chen G., Chen X., Gao W., Long L., Xia L., Chen Z-H. Evolution of Abscisic Acid Signaling for Stress Responses to Toxic Metals and Metalloids. Front. Plant Sci. 2020, 11, 909. [Google Scholar]
- Nguyen, T.Q. , Sesin V. , Kisiala A., Emery R.J.N. Phytohormonal Roles in Plant Responses to Heavy Metal Stress: Implications for Using Macrophytes in Phytoremediation of Aquatic Ecosystems. Environ. Toxicol. Chem. 2021, 40, 7–22. [Google Scholar]
- Mohamed, H.B. , Sofy M.R., Almoneafy A.A., Abdelhamid M.T., Basit A., Sofy A.R., Lone R., El-Enain M.M.A. Role of Microorganisms in Managing Soil Fertility and Plant Nutrition in Sustainable Agriculture. Chapter 4; 2021. In: H. I. Mohamed et al. (eds.), PGP Microbes for Sustainable Biotic and Abiotic Stress Mgmt. Springer Nature; Switzerland AG.
- Shah, K.K. , Tripathi S. , Shrestha T.J., Modi B., Paudel N., Das B.D. Role of soil microbes in sustainable crop production and soil health: A review. Agric. Sci. Technol. 2021, 13, 109–118. [Google Scholar]
- Gikas, P. Kinetic responses of activated sludge to individual and joint nickel (Ni(II)) and cobalt (Co(II)) - an isobolographic approach. J. Hazard. Materials. 2007, 143, 246–256. [Google Scholar] [CrossRef]
- Gikas, P. Single and combined effects of nickel (Ni(II)) and cobalt (Co(II)) ions on activated sludge and on other aerobic microorganisms: J. Hazard. Materials. 2008, 159, 187–203. [Google Scholar] [CrossRef]
- Xie, Y. , Fan J. , Zhu W., Amombo E., Lou Y., Chen L., Fu J. Effect of Heavy Metals Pollution on Soil Microbial Diversity and Bermudagrass Genetic Variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar]
- Abdu, N. , Abdullahi A. A., Abdulkadir A. Heavy metals and soil microbes. Environ Chem Lett. 2017, 15, 65–84. [Google Scholar]
- Wang, Y. , Narayanan M. , Shi X., Chen X., Li Z., Natarajan D., Ma Y. Plant growth-promoting bacteria in metal contaminated soil: Current perspectives on remediation mechanisms. Front. Microbiol. 2022, 13, 966226. [Google Scholar]
- González Henao, S. , Ghneim-Herrera T. (2021) Heavy Metals in Soils and the Remediation Potential of Bacteria Associated With the Plant Microbiome. Front. Environ. Sci. 2021, 9, 604216. [Google Scholar]
- Nwachukwu, O.I. , Pulford, I. D. Microbial respiration as an indication of metal toxicity in contaminated organic materials and soil. J. Hazard. Materials. 2011, 185, 1140–1147. [Google Scholar]
- Kizilkaya, R. , Akin, T. , Bayrakli, B., Salam, M. Microbiological characteristics of soils contaminated with heavy metals, Eur. J. Soil Biol. 2004, 40, 95–102. [Google Scholar]
- Chu, D. Effects of heavy metals on soil microbial community. Earth Environ. Sci. 2018, 113, 012009; IOP Publishing, IOP Conf Series. [Google Scholar] [CrossRef]
- Gill, S.S. , Tuteja, N. Reactive oxygen species and antioxidant Machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar]
- Chibuike, G. U. , and Obiora, S. C. Heavy metal polluted soils: effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 752708.
- Koike, S.T. , Subbara K. V., Davis R.M., Turini T.A. (2003). Vegetable diseases caused by plant pathogens. ANR Pub. 2003, 8099, 1–13. [Google Scholar]
- Sanogo, S. Microbial clicks and combobulation: integrating microbes without passports and visas for managing soilborne diseases. Arch. Phytopath. Plant Prot. 2020, 53, 691–697. [Google Scholar] [CrossRef]
- Rashid, A. , Hwang S. F., Ahmed H.U., Turnbull1 G.D., Strelkov S.F. Gossen B.D. Effects of soil-borne Rhizoctonia solani on canola seedlings after application of glyphosate herbicide. Canadian J. Plant Sci. 2013, 93, 97–107. [Google Scholar]
- Rashid, A. , Sharma P., Evans, I. Plant bioassay techniques for detecting and identifying herbicide residues in soil. Agri-Facts: Practical information for Alberta’s Agriculture Industry. Agdex # 609-1, 2001. Edmonton, Alberta, Canada.
- Yamada, M. , Malambane G. , Yamada S., Suharsono S., Tsujimoto H., Moseki B., Akashi K. Differential physiological responses and tolerance to potentially toxic elements in biodiesel tree Jatropha curcas. Sci. Reports. 2018, 8, 1635. [Google Scholar]
- Gill, S.S. , Hasanuzzaman, M. , Nahar, K., Macovei A., Tuteja N. Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physiol. Biochem. 2013, 63, 254–261. [Google Scholar] [PubMed]
- López-Millán, A.F. , Sagardoy R. , Solanas M., Abadía A., Abadía J. Cadmium toxicity in tomato (Lycopersicon esculentum) plants grown in hydroponics. Environ. Expt. Bot. 2009, 65, 376–385. [Google Scholar]
- DalCorso, G. , Farinati S. , Furini A. Regulatory networks of Cd stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar]
- Sun, S. , Li M. , Zuo J., Jiang W., Liu, D. Cadmium effects on mineral accumulation, antioxidant defense system and gas exchange in cucumber. Z. Agric. 2016, 102, 193–200. [Google Scholar]
- Rizwan, M. , Ali, S. , Adrees, M., Rizvi, H., Zia-ur-Rehman, M., Hannan, F., Qayyum, M.F., Hafeez, F., Ok, Y.S. Cadmium stress in rice: toxic effects, tolerance mechanisms, and mgmt: a review. Env. Sci. Poll. Res. 2016, 23, 17859–17879. [Google Scholar]
- Ullah, I. , Mateen A. , Ahmad A., Munir I., Iqbal A., Alghamdi K.M.S., Al-Solami H.M., Siddiqui M.F. Heavy metal ATPase genes expression induced by endophytic bacteria, AI001, and AI002” mediate cadmium translocation and phytoremediation. Environ. Poll. 2022, 293, 118508. [Google Scholar]
- Haider, F.U. , Liqun, C. , Coulter, J.A., Alam S., Cheema A.S., Wu J., Zhang R., Wenjun M., Farooq M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotox. Environ. Safety. 2021, 211, 111887. [Google Scholar]
- Nagajyoti, P.C. , Lee, K. D., Sreekanth T.M.V. Heavy metals, occurrence, and toxicity for plants: a review. Environ Chem. Lett. 2010, 8, 199–216. [Google Scholar]
- Glinska, S. , Michlewska, S. , Gapinska, M., Seliger, P., Bartosiewicz, R. The effect of EDTA and EDDS on lead uptake and localization in hydroponically grown Pisum sativum L. Acta Physiol Plant. 2014, 36, 399–408. [Google Scholar]
- Aslam, M. , Aslam A. , Sheraz M., Ali B., Ulhassan Z., Najeeb U., Zhou Z., Gill R.A. Lead Toxicity in Cereals: Mechanistic Insight Into Toxicity, Mode of Action, and Management. Front. Plant Sci. 2021, 11, 587785. [Google Scholar]
- Gautam, M. , Sengar R. S., Chaudhary R., Sengar K., Garg S. Possible cause of inhibition of seed germination in rice by heavy metals Pb and Hg. Toxicol. Environ. Chem. 2010, 92, 1111–1119. [Google Scholar]
- Hossain, M.A. , Uddin, M. N., Sarwar, A.K.M.G. Toxicity of arsenic on germination and seedling growth of rice. J. Bang. Soc. Agric. Sci. Technol. 2007, 4, 153–156. [Google Scholar]
- Abbas, G. , Murtaza B. , Bibi I., Shahid M., Niazi N.K., Khan, M.A., Amjad A., Hussain M., Natasha. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. Int. J. Environ. Pub. Health Res. 2018, 15, 59, 1–45. [Google Scholar]
- Imran, M.A. , Sajid, Z. A., Chaudhry, M.N.. Arsenic (As) Toxicity to Germination and Vegetative Growth of Sunflower (Helianthus annuus L.). Pol. J. Environ. Stud. 2015, 24, 1993–2002. [Google Scholar]
- Das, I. , Ghosh K. , Das D., Das S.K. Assessment of arsenic toxicity in rice plants in areas of West Bengal. Chem. Spec. Bioavail. 2013, 25, 201–108. [Google Scholar]
- Huq, S.M.I. , Parvin K. , Rahman R., Joardar J.C. Response of cowpea (Vigna sinensis to Arsenic. Canadian J. Pure. Appl. Sci. 2009, 3, 897–902. [Google Scholar]
- Patra, M. , Sharma, M. Mercury Toxicity in Plants. Bot. Review. 2000, 66, 379–422. [Google Scholar]
- Cargnelutti, D. , Tabaldi L. A., Spanevello R.M., Jucoski G.O., Battisti V., Redin M., Linares C.E.B., Dressler V.L., Flores M.M., Nicoloso F.T., Morsch V.M., Schetinger, M.R.C. Mercury toxicity induces oxidative stress in growing cucumber seedlings. Chem. 2006, 65, 999–1006. [Google Scholar]
- Marrugo, J. , Hernández J. , Pinedo-Hernández J., Montes E, Sergi D. Mercury uptake and effects on growth in Jatropha curcas. J. Environ. Sci. 2016, 48, 120–125. [Google Scholar]
- Singh, H.P. , Mahajan P. , Kaur S., Batish D.R., Kohli R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar]
- Amin, H. , Arain B. A., Amin F., Surhio M.A. Phytotoxicity of Chromium on Germination, Growth and Biochemical Attributes of Hibiscus esculentus L. Am. J. Plant Sci. 2013, 4, 2431–2439. [Google Scholar]
- Samantaray, S. , Rout G. R., Das P. Role of Cr on plant growth and metabolism. Acta Physiol. Plant. 1998, 20, 201–212. [Google Scholar]
- Stambulska, U. , Bayliak M.M., Lushchak V.I. Chromium(VI) Toxicity in Legume Plants: Modulation Effects of Rhizobial Symbiosis. BioMed Res. Int. 2018, 803121.
- Shankar, S. , Shanker, U., Shikha. Arsenic contamination of groundwater: A Review of sources, prevalence, health risks, and strategies for mitigation. Sci. World J. 2014, 304524.
- Oliveira, H. Chromium as an Environmental Pollutant: Insights on Induced Plant Toxicity. J. Bot. 2012, 375843. [Google Scholar] [CrossRef]
- Nikolaou, K.E. , Chatzistathis T. , Theocharis S., Argiriou A., Koundouras S., Zioziou E. Effects of Chromium Toxicity on Physiological Performance and Nutrient Uptake in Two Grapevine Cultivars (Vitis vinifera L.) Growing on Own Roots or Grafted onto Different Rootstocks. Hort. 2022, 8, 493. [Google Scholar]
- Yuan, J. , Yan L. , Li G., Sadiq M., Rahim N., Wu J., Ma W., Xu G., Du M. Effects of conservation tillage strategies on soil physicochemical indicators and N2O emission under spring wheat monocropping systems. Sci. Rep. 2022, 12, 7066. [Google Scholar]
- He, Z. , Shentu J. , Yang X., Baligar V.C., Zhang T., Stoffella P.J. Heavy Metal Contamination of Soils: Sources, Indicators, and Assessment. J. Environ. Indicators. 2015, 9, 17–18. [Google Scholar]
- Cai, L. , Xu Z. , Ren M., Guo Q., Hua X., Hua G., Wand H., Peng P. Source identification of eight hazardous heavy metals in agricultural soils of Huizhou, Guangdong Province, China. Ecotox. Environ. Safety. 2012, 78, 2–8. [Google Scholar]
- Li, R. , Wu H. , Ding J., Fu, W., Gan L., Li Y. Mercury pollution in vegetables, grains and soils from areas surrounding coal-fired power plants. Sci. Reports. 2017, 7, 46545. [Google Scholar]
- Codex Alimentarius Commission. FAO- WHO. CF/14 INF/1; 2021.
Figure 1.
Classification of metallic and non-metallic elements, frequently found in agricultural soils. Mg (magnesium), Ca (calcium), Fe (iron), B (boron), Mn (manganese), Zn (zinc), Mo (molybdenum), Cu (copper), Pb (lead), Ni (nickel), Cr (chromium), As (arsenic), Hg (mercury), Cd (cadmium), Al (aluminum), Li (lithium), K (potassium), Na (sodium), Si (silicon).
Figure 1.
Classification of metallic and non-metallic elements, frequently found in agricultural soils. Mg (magnesium), Ca (calcium), Fe (iron), B (boron), Mn (manganese), Zn (zinc), Mo (molybdenum), Cu (copper), Pb (lead), Ni (nickel), Cr (chromium), As (arsenic), Hg (mercury), Cd (cadmium), Al (aluminum), Li (lithium), K (potassium), Na (sodium), Si (silicon).
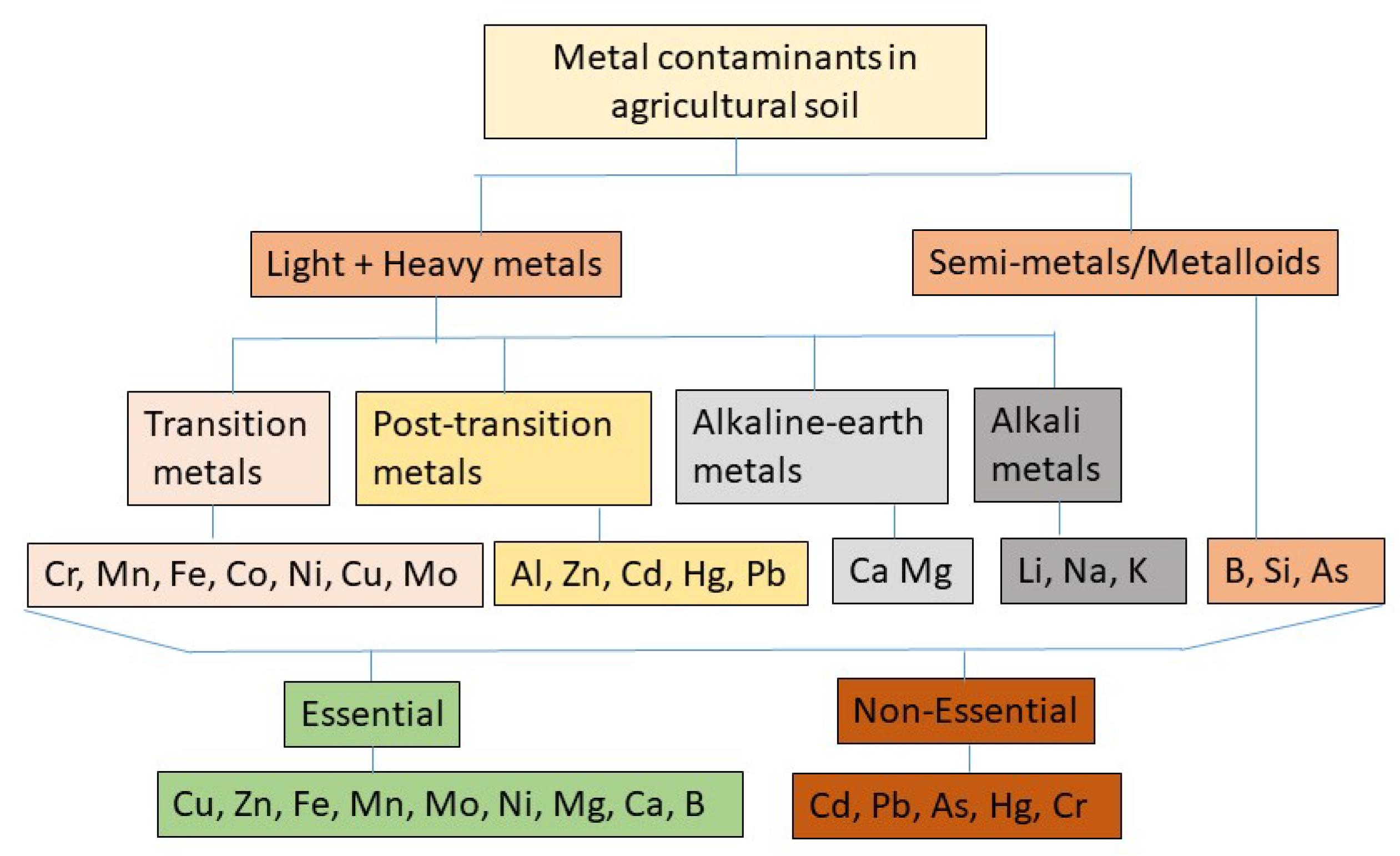
Figure 2.
A flow chart showing the contamination of HMs in farmlands through agricultural practices, their absorption and translocation in crop plants, and the key mechanisms of action in plants.
Figure 2.
A flow chart showing the contamination of HMs in farmlands through agricultural practices, their absorption and translocation in crop plants, and the key mechanisms of action in plants.
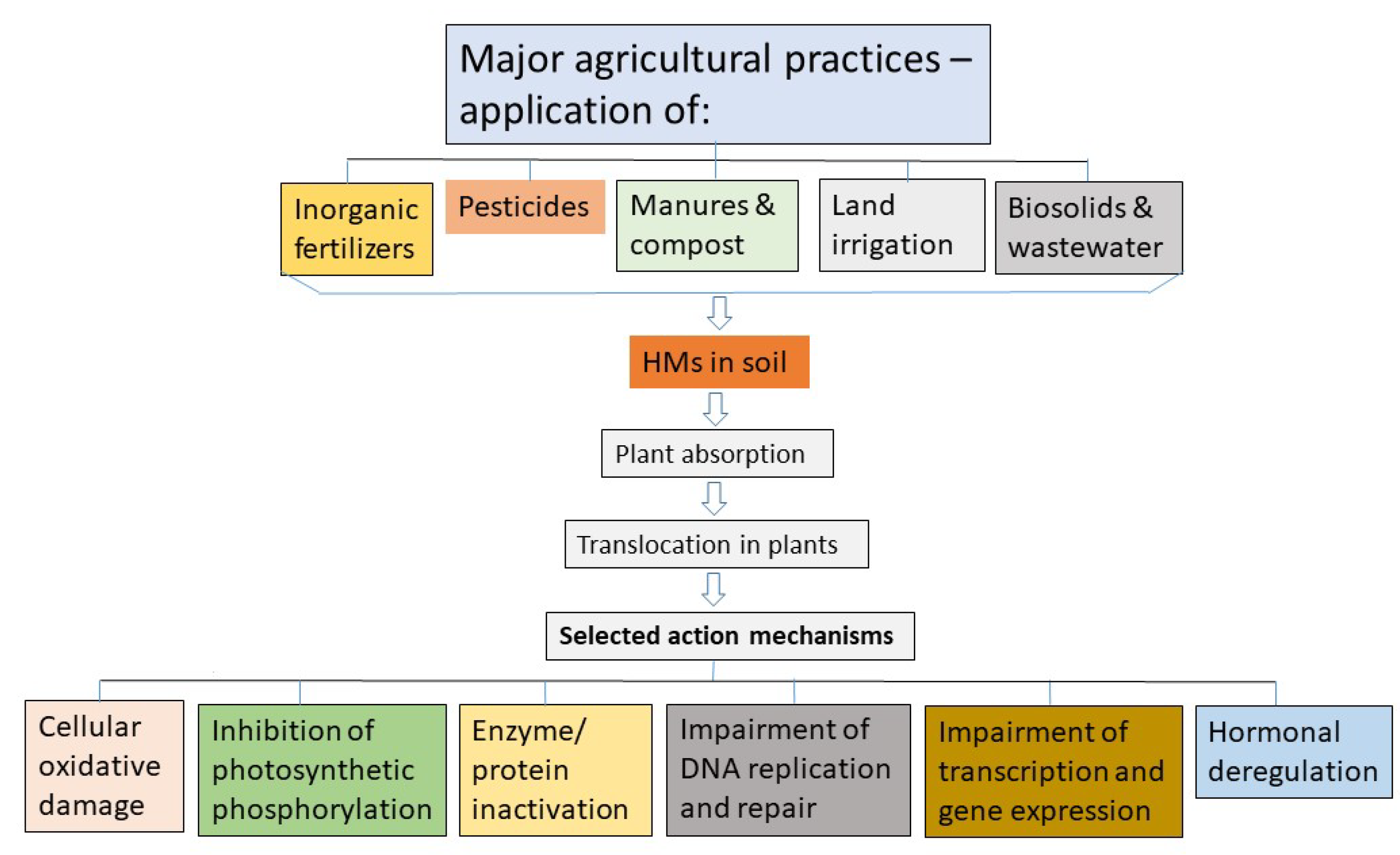
Figure 3.
Hypothetical structural diagrams showing binding interactions of HM elements (e.g., Cd) with soil particles. A (basic soil): Cd2+ molecules held tightly with the soil the particle in the presence of high OM content by electrostatic forces (EFs); B (neutral soil): some Cd2+ molecules are available in free cationic form in the soil due to low OM content and low EFs; C (acidic soil): Cd2+ molecules are available in free cationic form in the soil containing high [H+].
Figure 3.
Hypothetical structural diagrams showing binding interactions of HM elements (e.g., Cd) with soil particles. A (basic soil): Cd2+ molecules held tightly with the soil the particle in the presence of high OM content by electrostatic forces (EFs); B (neutral soil): some Cd2+ molecules are available in free cationic form in the soil due to low OM content and low EFs; C (acidic soil): Cd2+ molecules are available in free cationic form in the soil containing high [H+].
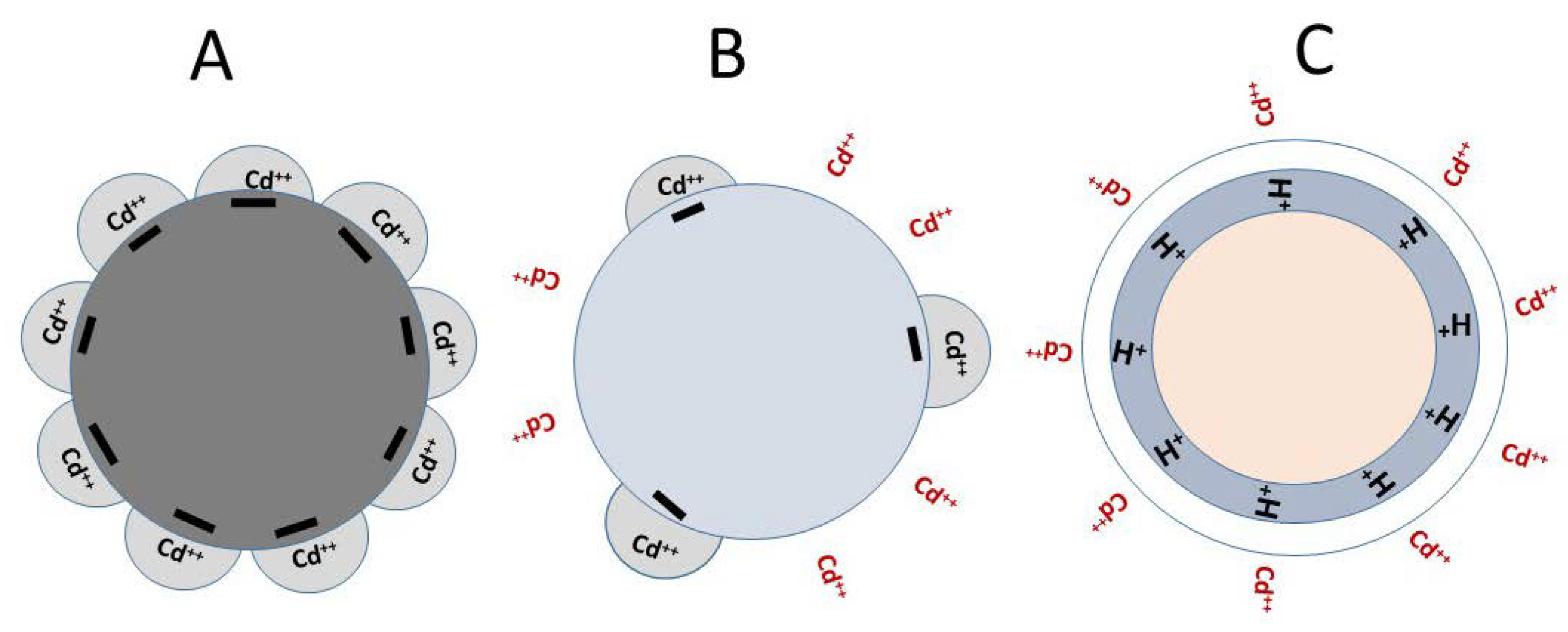
Figure 4.
Diagrammatic drawing of a portion of thylakoid membrane showing stroma and lumen sides, locations of photosystem II (PSII) and photosystem I (PSI) reaction centers (RCs), and oxygen evolving complex (OEC). A: Generation of ATP and NADPH through photosynthetic electron transport in the absence of Pb2+; B: Depletion of extrinsic polypeptide shield from OEC of PSII-RC and inhibition of phosphorylation and NADP reduction in the presence Pb2+.
Figure 4.
Diagrammatic drawing of a portion of thylakoid membrane showing stroma and lumen sides, locations of photosystem II (PSII) and photosystem I (PSI) reaction centers (RCs), and oxygen evolving complex (OEC). A: Generation of ATP and NADPH through photosynthetic electron transport in the absence of Pb2+; B: Depletion of extrinsic polypeptide shield from OEC of PSII-RC and inhibition of phosphorylation and NADP reduction in the presence Pb2+.
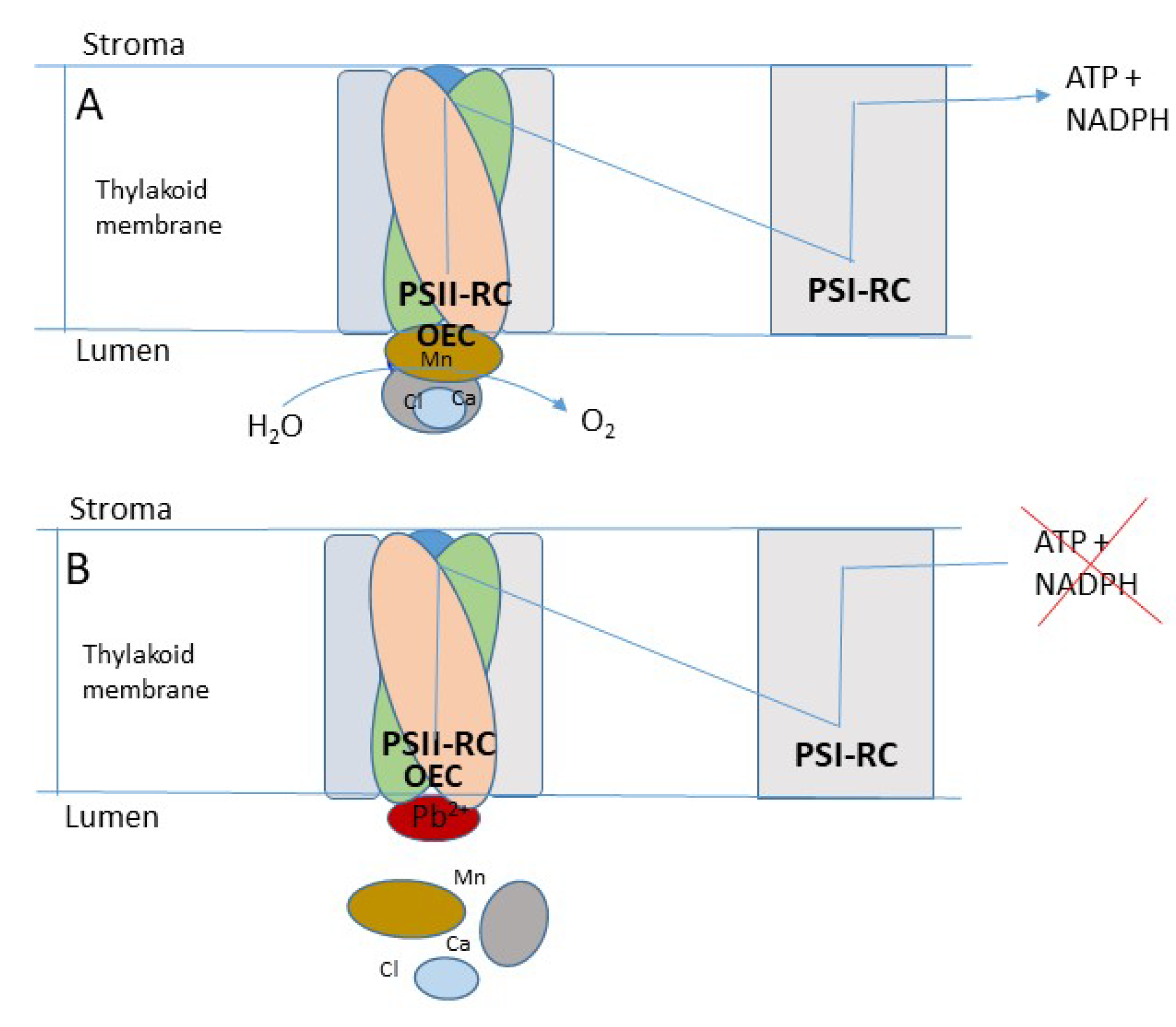

Table 1.
Cadmium concentration in several phosphate fertilizers. Source: [27].
Table 1.
Cadmium concentration in several phosphate fertilizers. Source: [27].
| Fertilizers | Cadmium content (mg Kg−1) | |
|---|---|---|
| Based on product | Based on P content | |
| Complete fertilizer | 23–29 | 418–527 |
| Single superphosphate | 16–26 | 186–302 |
| Superphosphate | 13–34 | 151–395 |
| Rock phosphate | 7.2–47 | 54–303 |
| High analysis fertilizer | <0.6–5.6 | 15–118 |
| Double superphosphate | <0.6–12 | <3.6–72 |
| Triple superphosphate | 0.8–7.0 | 24–35 |
| Mono-ammonium phosphate | 1.8–8.1 | 12–37 |
| Di-ammonium phosphate | 4.3–6.6 | 22–28 |
Table 2.
Heavy metal concentrations in greenhouse soil because of repeated application of inorganic fertilizers. Source: [24].
Table 2.
Heavy metal concentrations in greenhouse soil because of repeated application of inorganic fertilizers. Source: [24].
| Elements | MAX | MIN | Fold difference |
|---|---|---|---|
| mg Kg−1 soil | |||
| Cadmium | 0.65 | 0.06 | 10.8 |
| Copper | 171.5 | 21.0 | 8.2 |
| Nickel | 36.9 | 28.7 | 1.3 |
| Lead | 38.0 | 20.5 | 1.9 |
| Zinc | 433 | 71.9 | 6 |
| Chemical name | Formula | HM elements |
|---|---|---|
| Insecticides | ||
| Aluminium phosphide | AIP | Al |
| Aluminium silicate | Al₂Si₂O₇ | Al |
| Arsenic acid | H₃AsO₄ | As |
| Copper acetoarsenite | C₄H₆As₆Cu₄O₁₆ | As, Cu |
| Copper oxide | Cu₂O | Cu |
| Copper carbonate | CH₂Cu₂O | Cu |
| Copper naphthenate | C₂₂H₁₄CuO₄ | Cu |
| Lead arsenate | AsHO₄Pb | As, Pb |
| Lithium perfluorooctane sulfonate | C₈F₁₇LiO₃S | Li |
| Sodium meta-arsenite | NaAsO2 | As |
| Fungicide | ||
| Copper oxide | CuO | Cu |
| Copper bis(3-phenylsalicylate) | C₂₆H₁₈CuO₆ | Cu |
| Copper abietate | C₄0H₅₈CuO₄ | Cu |
| Copper acetate | Cu₂(CH₃COO)₄ | Cu |
| Copper carbonate | CH₂Cu₂O | Cu |
| Copper chloride | CuCl₂ | Cu |
| Copper hydroxide | H₂O₂Cu | Cu |
| Copper naphthenate | C₂₂H₁₄CuO₄ | Cu |
| Copper oxychloride | (ClCu₂H₃O₃)₂ | Cu |
| Copper sulphate | CuSO4-Ca(OH)2 | Cu |
| Mercuric oxide | HgO | Hg |
| Mercurous chloride | Hg₂Cl₂ | Hg |
| Methoxyethylmercury chloride | C₃H₇ClHgO | Hg |
| Methoxyethylmercury acetate | C₅H₁0HgO₃ | Hg |
| Phenyl mercuric acetate | C₈H₈O₂Hg | Hg |
| Phenylmercury chloride | C₆H₅ClHg | Hg |
| Phenylmercury nitrate | C₆H₅HgNO₃ | Hg |
| Sodium arsenite | NaAsO₂ | As |
| Zinc borate | ZnB3O4(OH)3 | Zn, B |
| Zinc oxide | ZnO | Zn |
| Zineb | C₄H₆N₂S₄Zn | Zn |
| Herbicides | ||
| Arsenic acid | H₃AsO₄ | As |
| Calcium arsenate | As₂Ca₃O₈ | As |
| Sodium arsenite | NaAsO₂ | As |
| Cacodylic acid | (CH3)2AsO(OH) | As |
| Rodenticides | ||
| Barium carbonate | BaCO3 | Ba |
| Zinc phosphide | Zn3P2 | Zn |
| Thallium sulfate | Tl2SO4 | Tl |
| Defoliants | ||
| Sodium dichromate | Na2Cr2O7 | Cr |
| Zinc chloride | ZnCl2 | Zn |
| Mercuric chloride | HgCl2 | Hg |
Table 4.
Pesticide products containing HM elements as impurities.
| Trade name | Technical name | Metal impurities (ppb)* |
|---|---|---|
| Insecticides | Defarge et al., [38] | |
| Pyrinex® | Chorpyriphos | As (390), Cr (800), Ni (1200) |
| Polysect® | Acetamipride | Ni (50) |
| Fungicides | Defarge et al., [38] | |
| Eyetak® | Prochloraze | As (200), Co (90), Cr (200), Ni (190), Pb (12) |
| Folpan® | Folpet | As (260), Cr (2000), Ni (1200) |
| Maronee® | Tebuconazole | As (90), Co (50), Cr (100) |
| Opus® | Epoxiconazole | Cr (90), Ni (60) |
| Pictor® | Boscalid | As (300), Co (275), Cr (1000), Ni (600) |
| Teldor® | Fenhexamid | As (575), Cr (800), Ni (800) |
| Herbicides | Defarge et al., [38] | |
| R 3+® | Glyphosate based formulations | As (375), Co (50), Cr (175), Ni (20) |
| R Bioforce® | As (260), Cr (200), Ni (120) | |
| R Express® | As (60) | |
| R GT+® | As (450), Co (150), Cr (100), Ni (50), Pb (10) | |
| R WeatherMax® | As (500), Cr (100), Ni (20), Pb (10) | |
| Bayer GC® | As (75), Co (60), Cr, (110) Ni (20) | |
| Clinic EV® | As (400), Co (90), Cr (150), Ni (20) | |
| Glyfos® | As (200), Cr (1100), Ni (50), Pb (30) | |
| Glyphogan® | As (320), Co (125, Cr (100), Ni (40) | |
| Pavaprop-G® | Cr (110), N (190) | |
| Radical Tech+® | As (270), Co (70), Cr (50), Ni (50) | |
| Lonpar® | 2,4-D | As (160), Cr (150), Ni (180) |
| Matin® | Isoproturon | As (100), Cr (100), Ni (30), Pb (25) |
| Starane® | Fluoroxypyr | As (75), Cr (250), Ni (100), Pb (100) |
| Insecticides | Alnuwaiser [39] | |
| Sniper® | Fipronil | Zn (506), Cu (423), Cr (746), Co (275), Pb (88) |
| CyperCel® | Cypermethrin | Zn (2389), Cu (669), Cr (373), Co (18), Pb (807) |
| CyperSafe® | Cypermethrin | Zn (968), Cu (464), Cr (10), Co (6), Pb (119) |
| Scope 60® | Asaybrmthrin | Zn (527), Cu (539), Cr (437), Co (23), Pb (39) |
| Brodor® | Permethrin | Zn (10), Cr (16), Pb (186) |
| Clash® | Acephate + Buprofezin | Zn (1078), Cr (73), Co (39), Pb (1316) |
| Acefed® | Mithomail | Cu (19), Cr (48), Co (4), Pb (121) |
| Lanid® | - | Cu (128), Cr (60), Pb (98) |
| Probalt® | - | Cu (179), Cr (85), Co (25), Pb (46) |
| Nourcam® | - | - |
| Madar® | - | Zn (10), Cu (66), Cr (16), Co (10), |
| PifPaf® | - | Cu (110), Co (5), Thallium, Tl (19), Pb (12) |
| Paygon® | - | Zn (52), Tl (15), Pb (19) |
*European Union (EU)/World Health Organization (WHO) prescribed permissible levels (ppb) in water: As (10), Cr (50), Ni (20/70), Pb (10), Co (NA).
Table 5.
A case study showing maximum (MAX) vs. minimum (MIN) concentration (mg Kg−1 dry weight) of HM elements in different livestock and poultry manures, and their fold differences (FD). Source: [42].
Table 5.
A case study showing maximum (MAX) vs. minimum (MIN) concentration (mg Kg−1 dry weight) of HM elements in different livestock and poultry manures, and their fold differences (FD). Source: [42].
| Source | Level | Zn | Cu | Pb | Cd | Cr | Hg | As | Ni |
|---|---|---|---|---|---|---|---|---|---|
| Pig | MAX | 4639 | 1288 | 23 | 60 | 85 | 0.3 | 89 | 19 |
| MIN | 100 | 73 | 0.3 | 0.04 | 3.5 | 0.0 | 0.01 | 4.7 | |
| FD | 46 | 18 | 77 | 1500 | 24 | - | 8900 | 4.0 | |
| Chicken | MAX | 578 | 314 | 33 | 4.1 | 251 | 0.5 | 23 | 39 |
| MIN | 166 | 18 | 3.0 | 0.03 | 4.0 | 0.02 | 0.05 | 5.2 | |
| FD | 3.5 | 17 | 11 | 137 | 63 | 25 | 460 | 7.5 | |
| Duck | MAX | 682 | 199 | 41 | 2.5 | 64 | 0.07 | 6.8 | 16 |
| MIN | 98 | 35 | 4.5 | 0.3 | 7.0 | 0.03 | 0.01 | 8.4 | |
| FD | 7.0 | 5.7 | 9.1 | 8.3 | 9.1 | 2.3 | 680 | 1.9 | |
| Poultry | MAX | 682 | 314 | 41 | 4.1 | 251 | 0.5 | 23 | 39 |
| MIN | 77 | 15 | 2.0 | 0.03 | 2.5 | 0.02 | 0.01 | 5.2 | |
| FD | 8.9 | 21 | 21 | 137 | 100 | 25 | 2300 | 7.5 | |
| Cattle | MAX | 816 | 174 | 32 | 3.4 | 79 | 0.6 | 6.3 | 19 |
| MIN | 49 | 12 | 1.6 | 0.04 | 0.8 | 0.02 | 0.01 | 4.2 | |
| FD | 17 | 15 | 20 | 85 | 99 | 30 | 630 | 4.5 | |
| Sheep | MAX | 431 | 215 | 20 | 1.4 | 22 | 2.4 | 2.6 | 12 |
| MIN | 42 | 8.4 | 1.7 | 0.3 | 8.0 | 0.2 | 0.6 | 1.2 | |
| FD | 10 | 26 | 12 | 4.7 | 2.8 | 12 | 4.3 | 10 |
Table 6.
Regulatory limits for HM elements, commonly found in applied biosolids in the USA. Source: [55].
Table 6.
Regulatory limits for HM elements, commonly found in applied biosolids in the USA. Source: [55].
| Elements | Maximum permissible level (mg Kg−1) | Cumulative loading rate (Kg ha−1) | Monthly average concentrations (mg Kg−1) | Annual loading rate (Kg ha−1) |
|---|---|---|---|---|
| As | 75 | 41 | 41 | 2.0 |
| Cd | 85 | 39 | 39 | 1.9 |
| Cr | 3000 | 3000 | 1200 | 150 |
| Cu | 4300 | 1500 | 1500 | 75 |
| Pb | 840 | 300 | 300 | 15 |
| Hg | 57 | 17 | 17 | 0.9 |
| Ni | 420 | 420 | 420 | 21 |
| Se | 100 | 100 | 36 | 5.0 |
| Zn | 7500 | 2800 | 2800 | 140 |
Table 7.
HM concentrations in irrigation water reported in some studies as compared to FAO approved maximum permissible limits.
Table 7.
HM concentrations in irrigation water reported in some studies as compared to FAO approved maximum permissible limits.
| Elements | Cr | Cu | Zn | As | Cd | Pb | Ni | References |
|---|---|---|---|---|---|---|---|---|
| mg L−1 | ||||||||
| Max level (FD)* |
2.13 (21.3) |
4.62 (23.1) |
15.20 (7.6) |
0.52 (5.2) |
0.02 (2.0) |
1.15 (0.2) |
- | Ahmed et al., [16] |
| Max level (FD) |
0.94 (9.4) |
0.61 (3.1) |
0.86 (0.4) |
- |
0.04 (4.0) |
0.19 (0.04) |
0.12 (0.6) |
Berihun et al., [58] |
| FAO limit | 0.10 | 0.20 | 2.00 | 0.10 | 0.01 | 5.00 | 0.20 | |
*Numbers in parentheses represent concentration FDs, calculated based on average concentration obtained divided by FAO limits.
Table 8.
Responses of crop plants to HM toxicity with respect to certain growth parameters.
| Crop Species |
HMs elements |
Conc. (mg Kg−1 soil) |
Height (cm) | Dry wt. (g) | Yield | Refs | ||
| Shoot | Root | Shoot | Root | g/plot | Alaboudi et al., [74]* | |||
| Helianthus annuus |
Pb | 0 | 53 | 34 | 4.3 | 2.5 | - | |
| 80 | 28 | 10 | 2.6 | 1.7 | - | |||
| 200 | 15 | 2 | 1.5 | 0.8 | - | |||
| Cd | 0 | 53 | 34 | 3.9 | 2.4 | - | ||
| 80 | 16 | 6 | 1.1 | 0.6 | - | |||
| 200 | 9 | 1.7 | 0.8 | 0.2 | - | |||
| Brassica Juncea |
Cu | 0 | 20 | - | - | - | 70 | Cu [75]** |
| 100 | 15 | - | - | - | 50 | |||
| 200 | 13 | - | - | - | 33 | |||
| Pb | 0 | 20 | - | - | - | 70 | ||
| 100 | 13 | - | - | - | 35 | |||
| 200 | 11 | - | - | - | 31 | |||
| Zn | 0 | 20 | - | - | 70 | |||
| 300 | 17 | - | - | 64 | ||||
| 500 | 16 | - | - | 44 | ||||
| Oryza Sativa |
Hg | mg L−1 nutrient solution | Pot−1 | Pot−1 | Du et al., [76]*** | |||
| 0 | - | - | 0.52 | 0.19 | - | |||
| 0.5 | - | - | 0.28 | 0.17 | - | |||
| 1.0 | - | - | 0.22 | 0.12 | - | |||
| 2.5 | - | - | 0.17 | 0.08 | - | |||
*Analytical grade HM salts dissolved in distilled water were mixed with air-dried loamy sand soil (pH >6.25; EC ~0.27 dSm−1; OM ~10.23%; HM status = under detection limits). **HMs in the form of CuSO4, Pb(NO3)2, and ZnSO4 were mixed with alluvial soil (pH 6.15; OM ~2.44%; CEC 23.46 Cmol/kg; HM status (mg Kg−1 soil) = ~21.3; Pb ~57.2; Zn ~7.2). *** Different concentrations of HgCl2 were mixed with nutrient solution (pH 5.5).
Table 9.
Partial alleviation of toxic effects of cadmium on some physiological growth parameters in pea plant (Pisum sativum) in the presence of a cytokinin hormone. Source: [188].
Table 9.
Partial alleviation of toxic effects of cadmium on some physiological growth parameters in pea plant (Pisum sativum) in the presence of a cytokinin hormone. Source: [188].
| Physiological parameters* | Kinetin (µM) | Cadmium (µM) | ||
|---|---|---|---|---|
| 0 | 25 | 50 | ||
| Chla/Chlb content (%) | 0 20 |
100 114/112 |
73/72 158/149 |
48/44 139/129 |
| Photosynthesis rate (%) | 0 20 |
100 124 |
88 146 |
42 80 |
| Soluble sugars content (%) | 0 20 |
100 180 |
67 226 |
49 249 |
| Soluble proteins content (%) | 0 20 |
100 72 |
119 61 |
133 50 |
| Amino acid content (%) | 0 20 |
100 132 |
73 179 |
65 200 |
* Control treatments as 100%.
Table 10.
Visual toxicity symptoms of HMs with no inherent symptoms of deficiency in plants.
| Toxicity symptoms observed in some studies involving multiple plant/crop species | References for review |
|---|---|
| Cadmium (Cd2+): solubility in water—high; mobility in soil colloids—high; bioavailability in soil—high; translocability in plants—high, toxicity in plants—highly lethal | |
| Chlorosis, wilting, leaf epinasty, stunting of plant growth. | Gill et al., [217] |
| Growth inhibition, leaf chlorosis and necrosis, and root browning. | López-Millán et al., [218] |
| Leaf rolling and chlorosis | DalCorso et al., [219] |
| Leaf chlorosis with green coloration around veins; leaf rolling, and growth stunting. | Sun et al., [220] |
| Reduction in seed germination, plant growth, biomass accumulation, and crop quality. | Rizwan et al., [221] |
| Chlorosis on newly expanded leaves, root growth inhibition. | Yamada et al., [216] |
| Growth stunting, chlorosis, root tip browning, and plant death. | Kumar et al., [20] |
| Stunting of growth, leaf chlorosis, reduction in fresh and dry biomass, plant death. | Ullah et al., [222] |
| Reduction in seed germination, shoot and root growth. | Chibuike and Obiora, [211] |
| Chlorosis, growth stunting, and necrosis. | Haider et al., [223] |
| Stunting of plant growth and blackening of roots. | Sharma and Dubey, [120] |
| Lead (Pb2+): solubility in water—low; mobility in soil colloids—poor; bioavailability in soil—limited; translocability in plants—restricted; toxicity in plants—moderately lethal | |
| Inhibition of seed germination, early seedling growth, root and stem elongation, and leaf expansion. | Nagajyoti et al., [224] |
| Inhibition of seedling growth and secondary root growth. | Glinska et al., [225] |
| Inhibition of seed germination, seedling growth, root and stem elongation, and leaf expansion. | Asati et al., [130] |
| Inhibition of seed germination, seedling development, root elongation. | Tiwari and Lata, [8] |
| Stunting of shoot and root growth. | Kumar et al., [20] |
| Plant growth inhibition. | Xia et al., [85] |
| Inhibition of seed germination, root and shoot biomass, root elongation, and cell death. | Aslam et al., [226] |
| Inhibition of germination and seedling growth. | Gautam et al., [227] |
| Inhibition of seed germination, seedling height, number of roots per plant, and dry matter production. | Hossain et al., [228] |
| Arsenic (As3+ As5+): solubility in water—high; mobility in soil colloids—high; bioavailability in soil—high; translocability in plants—high, toxicity in plants—lethal | |
| Reduction in leaf and root growth, wilting and violet coloration of leaves. | Abbas et al., [229] |
| Reduction in shoot and root length, and number of leaves per plant. | Imran et al., [230] |
| Reddening of tips, blades, margins, and midribs followed by yellowing of entire leaves. | Das et al., [231] |
| Shortening of plant height, premature shedding of leaves, and reduction in the number and size of nodules. | Huq et al., [232] |
| Reduction in leaf area, leaf fresh weight, fruit yield, seed germination, seedling height, and dry matter production; stunting of growth; chlorosis and wilting. | Asati et al., [130] |
| Reduction in seed germination, seedling height, leaf area, dry matter production, crop yield; chlorosis and wilting. | Chibuike and Obiora, [211] |
| Mercury (Hg2+): solubility in water—low; mobility in soil colloids—low; bioavailability in soil—moderate; translocability in plants—limited, toxicity in plants—moderately lethal | |
| Abnormal germination, hypertrophy of root and coleoptile, inhibition of seedling growth. | Patra and Sharma, [233] |
| Reduction in germination, plant height, tiller and panicle production, biomass accumulation, and yield; chlorosis. | Chibuike and Obiora, [211] |
| Decrease in both root and shoot biomass. | Du et al., [76] |
| Inhibition of seed germination, shoot and root length, and fresh and dry matter production. | Gautam et al., [227] |
| Reduction in seed germination, embryo growth, primary root elongation. | Azevedo and Rodriguez, [146] |
| Inhibition of root and shoot biomass. | Cargnelutti et al., [234] |
| Inhibition of plant growth, biomass production, and leaf area. | Marrugo-Negrere et al., [235] |
| Inhibition of germination, seedling growth and development, biomass accumulation; leaf chlorosis and necrosis. | Singh et al., [236] |
| Chromium (Cr3+ Cr6+): solubility in water—moderate; mobility in soil colloids—moderate; bioavailability in soil—moderate; translocability in plants—restricted; toxicity in plants—moderately lethal | |
| Inhibition of root and plant growth; leaf chlorosis. | Sharma et al., [186] |
| Inhibition of seed germination, root and shoot growth; reduction of plant biomass. | Asati et al., [130] |
| Inhibition of seed germination, seedling and plant growth. | Amin et al., [237] |
| Growth retardation, root discoloration. | Yamada et al., [216] |
| Chlorosis, necrosis; reduction in dry wight, nodulation, crop yield; inhibition of plant growth, root length | Samantaray et al., [238] |
| Inhibition of seed germination and seedling development and reduction of plant biomass and crop yield. | Stambulska et al., [239] |
| Inhibition of germination, root and shoot growth, dry matter production, and yield. | Shanker et al., [240] |
| Decrease in seed germination, reduction in growth, and yield. | Oliveira, [241] |
| Leaf interveinal chlorosis and root browning. | Nikolaou et al., [242] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



