
Submitted:
06 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
Pufferfish (Tetraodontidae) inhabiting the Mediterranean Sea can represent an emerging public health risk due to the possible accumulation of marine neurotoxins (TTXs and STXs) in their tissues. In this study, the presence of pufferfish species in the Strait of the Sicily (Lampedusa Island, Italy) was investigated using a Citizen Science (CS) approach involving local fishermen. A sub-group of specimens collected were sent to the National reference Laboratory on Marine Biotoxins for de-termining the presence of TTXs using a validated HILIC-MS/MS method on fish tissue. The STXs presence was also screened in some specimens. Overall, 56 specimens, identified as Sphoeroides pachygaster (Müller &Troschel, 1848) were collected. Data on specimen total length, body weight, fishing method, catch area (with relative depth temperature and salinity) were analyzed and compared with the S. pachygaster records reported in Guardone et al. (2018), which were appro-priately updated until 2022. All the analysed tissues were found to be negative for both TTXs and STXs. The key role of the CS in monitoring potentially toxic marine species was proved. Outcomes from this study, the first investigating the toxicity of S. pachygaster in Italian waters, may provide useful data for the proper assessment of this emerging risk
Keywords:
marine toxins
; TTX
; Saxitoxins
; pufferfish
; not indigenous species
; Mediterranean
; risk assessment
1. Introduction
Numerous non-indigenous species (NIS), originally distributed at tropical latitudes, currently inhabit the Mediterranean Sea [1], mainly as a consequence of anthropogenic environmental modification, such in the case of the “Lessepsian” migration. Fish of the Tetraodontidae family (commonly known as pufferfish) are involved in this phenomenon, with at least eleven species already described in the Mediterranean waters [2]. Among them, the particularly invasive Silver pufferfish, Lagocephalus sceleratus (Gmelin, 1789), and other species with less invasive character, namely the Oceanic pufferfish, L. lagocephalus (Linnaeus, 1758), the Blunthead pufferfish, Sphoeroides pachygaster (Müller & Troschel, 1848) and the Guinean puffer, S. marmoratus (Lowe, 1838) were reported along the Italian coasts [2]. Pufferfish are considered the most common cause of Tetrodotoxin (TTX) poisoning worldwide [2,3]. The TTX is the most lethal marine toxin [4], and it can be accumulated in different pufferfish tissues via the food chain [5], with concentration usually dependent on the sex, habitat, and season [6,7,8]. Factually, the term TTX is used to identify different thermostable toxins and analogues [6,7,9], belonging to a structurally distinct group of guanidinium alkaloids [10], reason why we referred to TTXs from this point forward. TTXs are mainly produced by more than 30 genera of marine bacteria [3,11]. Such TTXs-producing bacteria have been found associated with different tissues of several aquatic species, including dinoflagellate [3]. Besides pufferfish, also gastropods, arthropods and cephalopods are reported as sources of poisoning [12]. The TTXs share a high affinity with another guanidinium alkaloids group, Saxitoxins (STXs), also known as Paralytic Shellfish Toxins (PSTs) [10,13], which are produced by marine dinoflagellates, such as Gymnodinium catenatum, Alexandrium spp. and Pyrodinium bahamense, and freshwater cyanobacteria such as Cylindrospermopsis spp. and Lyngbya spp. [14,15]. Given this structural affinity, TTXs and STXs share a similar mechanism of action: the eventual ingestion of seafood contaminated with TTXs/STXs leads in fact in both cases to the blockage of the action potential of voltage-gated Na+ channels along nerves, which inhibits generation impulse conduction, resulting in muscle paralysis [16].
Cases of TTXs/STXs human poisoning due to the unaware ingestion of several species of toxic pufferfish are reported worldwide [12,17,18]. With respect to the Mediterranean area, the reported human TTXs poisoning cases were mainly related to the unintentional consumption of L. sceleratus [3]. The first toxicity study on pufferfish caught in the Mediterranean Sea was performed in 2008 [19]. Factually, it seems that the presence of TTXs in pufferfish species inhabiting this area has been investigated since that year (Table 1).
Among the four species recorded along the Italian coasts (L. sceleratus, L. lagocephalus, S- pachygaster, S. marmoratus), TTXs were found in all specimens of L. sceleratus from different Mediterranean areas and in specimens of L. lagocephalus collected in the southwestern region, along the Tunisian coast (Table 1). Contrariwise, TTXs were never found in Mediterranean specimens of S. pachygaster [21,27,36,38] in Table 1]. However, poisoning cases related to the ingestion of Sphoerides spp. are reported in Centre-South America and Oceania [12]. With respect to STXs, no human poisoning due to the ingestion of pufferfish are reported to date in the Mediterranean area [2], although STXs accumulation in Mediterranean L. sceleratus was described [20]. Interestingly, neither the presence of TTXs nor STXs was investigated in pufferfish specimens collected along the Italian coasts (Table 1).
As concerns the presence of both these guanidinium neurotoxins in marine organisms different from pufferfish, TTXs were recently detected in different species of molluscs and gastropods collected from marine water surrounding European countries including UK, Greece, Netherlands, Portugal, Spain, Italy and France [3]. In addition, some studies have highlighted the presence of TTXs in bivalve molluscs collected in the Mediterranean Sea and in Italian waters [39]. Also, high concentration of both TTXs and STXs were found in in mussel from Sicily (Syracusa bay) in 2019 [40].
The potential spreading of TTXs and STXs and of organisms potentially accumulating such toxins in the Mediterranean waters represents an emerging risk for EU citizens, and unavoidably catches the attention of both the scientific community and the control authorities. Since this phenomenon is of large-scale, involving citizens could help in its monitoring [41]. The so-called “Citizien Science” (CS), intended as the science where the public actively contribute to research [42], can provide information that otherwise could not be obtained, and it is often applied to phenomena related to changing biodiversity [37]. Targeted campaigns and meetings have been already held to make the EU citizens aware of the TTXs poisoning risk linked to the presence of potentially toxic pufferfish in the Mediterranean Sea [43,44]. A CS approach was already used in our previous project entitled “Climate change and food safety: molecular, microbiological and toxicological analysis on toxic fish species in the Tyrrhenian Sea” funded by the Italian Ministry of Health [45]. In that context, a revision of pufferfish records in the Mediterranean Sea, and specifically along the Italian coasts, was proposed for the characterization of this emerging risk in an early stage of the process [2]. Unfortunately, no specimens were collected during that project.
In this study, the CS was again used by involving local fishermen with the aim to monitor the presence of pufferfish species in the Strait of the Sicily (Lampedusa Island). The involved fishermen collected specimens of S. pachygaster, while no specimens of the other three species inhabiting the Italian coasts were captured. Considering the scarcity of data as regards the presence of TTXs [21,27,36] (Table 1) in Mediterranean S. pachygaster, part of the specimens collected in this study was analysed for TTXs detection by the Marine Research Centre, the National Reference Laboratory for Marine Biotoxins (NRL) (Cesenatico, Italy). This study, other than depicting an in-depth view on the distribution of S. pachygaster in the investigated area, will provide new toxicological data for a proper TTXs risk assessment.
2. Materials and Methods
2.1. Citizen Science for the monitoring of pufferfish species
2.1.1. Organization of the monitoring activities
In January 2020, initial contacts were established with the competent authority of Lampedusa (Sicily, Italy) and the Fishermen association operating in the waters around the island, in order to enquire about their willingness to participate in a CS project for the monitoring of pufferfish species. The willingness was confirmed by both the Competent Authority and by 20 fishermen out of a total of 40 regularly registered at the municipality. Thus, an informative brochure was produced and distributed (Figure S1). The brochure, including pictures and morphological description of the 4 pufferfish species already recorded along the Italian coasts (L. lagocephalus, L. scerelatus, S. marmoratus and S. pachygaster), was intended to help fishermen in the species identification.
2.1.2. Project dissemination
In July 2020, a meeting was organized by Department of Veterinary Sciences of the University of Pisa and the municipality of Lampedusa to present the project and establish a collaborative relationship between interested parties, especially addressed to the collection of data and pufferfish specimens. The meeting was held at the Lampedusa Marine Protected Area headquarters. General information on the pufferfish species described in the Mediterranean Sea, with particular attention to those already reported along the Italian coast, and the related risks for consumers were provided. The meeting was attended by stakeholders belonging to the fishing sector of the marine area, including the Italian Institute for Environmental Protection and Research (I.S.P.R.A of Palermo), the Port authorities of Lampedusa, the Fishermen association of Lampedusa, the Local administration of Lampedusa and Linosa and the Local dive centers.
2.2. Pufferfish specimens’ collection and identification
2.2.1. Specimens’ collection
The fishermen were asked to photograph and weigh each collected specimen accidentally caught during routine fishing activities, and also instructed on how to preserve (preferably frozen) and transfer them to a collection point in the island. The fishermen were also asked to record the date of catch (month and year). Whole specimens (when possible), were transferred frozen to the FishLab (Department of Veterinary Sciences, University of Pisa). Once received, each specimen total length (from the apex of the snout to the caudal peduncle) was measured. Then, the specimen body weight (mean and range) and the specimen total length (mean and range) were calculated. The fishermen were also asked to collect additional information on the GPS coordinate of the approximate fishing site and the fishing method. Subsequently, the depth and the relative type of seabed for each GPS coordinate provided (corresponding to the catch point of a variable number of specimens) was assessed using the database ETOPO1 (National Geophysical data centre; update to the year 2009) [46]. In addition, the average salinity and the water temperature measured at bottom level was assessed using the database World Ocean Atlas 2018 (Ocean Climate Laboratory of the National Oceanografic data centre U.S) [47]. Then, mean and range of depth, salinity and temperature were calculated. Data on mean salinity and temperature are discussed in details in section 3.3. The average temperature and salinity are discussed in the results section of TTXs detection (3.3).
2.2.2. Specimens’ identification
All the collected specimens (n=56, of which 42 in 2020 and 14 in 2021) were morphologically identified as S. pachygaster using the FAO morphological keys (FishBase.org) directly on the specimens (if received as whole) or on the photographs provided by the fishermen. The morphological identification was confirmed by molecular analysis conducted as described in Malloggi et al. [48] on 10 specimens randomly selected among those received. In particular, two distinct S. pachygaster haplotypes were found by analyzing the obtained sequences. One representative COI sequence of each haplotype was deposited on GenBank (Accession number will be provide as soon as released).
2.3. Collection of records of S. pachygaster in the Mediterranean and Italian waters
All the results obtained from the previous sections were compared to the available literature on the Mediterranean records of S. pachygaster. To do this, the records reported in the study of Guardone et al. [2] (from 1979 until 2016), were updated until 2022 in order to collect new records of S. pachygaster specimens, using the same bibliographic searching and inclusion criteria [2]. On the new records, the same data considered by Guardone et al. [2] was collected (Table 2).On the collected data, the following parameters (to be compared to our results) were calculated: 1) the overall number of specimens recorded (also classified per date of catch), 2) the total specimen length (mean and range calculated as in section 2.2.1), 3) the specimen body weight (mean and range calculated as in section 2.2.1), the specimen depth of the catch area (mean and range). The number of specimens used for the calculation of the aforesaid parameters was different due to the high variability in the data presentation observed in new records.
2.4. TTXs detection
Twenty specimens were selected for TTXs analysis among the 42 collected in 2020 (as in this year the sampling was complete, from February to September). All specimens heavier than the mean body weight calculated in Guardone et al. [2], corresponding to 1075.3 g, were used, considering that TTXs presence was often associated with higher fish sizes [20]. Since in 2020 only 7 specimens of the considered weight (defined as large specimens) were collected, other 13 smaller specimens were included to reach at least 20 specimens to be used for TTXs detection. The dates of catch of these 13 selected smaller specimens were distributed throughout the entire year, in order to have at least one specimen per month. The 5 organs (liver, gonads, skin, muscle and intestine) in which usually TTXs concentrate [18,59] were sampled for a total of 100 tissue samples. All the tissue samples, identified by the specimen code, were packaged separately, and sent to the NRL for the TTXs detection. The TTXs extraction and quantification were carried out using Hydrophilic Interaction Liquid Chromatography coupled to tandem Mass Spectrometry (HILIC-MS/MS) [60,61]. The method, which allows the identification and quantification of TTXs and PSTs in bivalve molluscs, was also validated on fish matrix as regards TTXs by the Italian NRL. Table SM1 reports UPLC gradient conditions, HILIC-MS/MS set of parameters employed and transitions for TTXs identification in accord with the method based on Electrospray ionization-mass spectrometry (ESI-MS), respectively. At least six-point calibration curve ranging from 0.2 µg/l to 400 µg/l was used (R2≥0,98) for the quantification of the toxin. The Limit of detection (LOD) was 3 µg/kg and the limit of quantification (LOQ) 10 µg/kg. TAB1 UPLC LC gradient condition (11 min run time). The concentration was calculated on the basis of extrapolation from the linear regression line (R2 ≥0.98) by applying the following formula:
3. Results and Discussion
3.1. Citizen science for monitoring the presence of pufferfish species in the Strait of the Sicily (Lampedusa Island)
The CS is an excellent method of collecting non-traditional data [62]. To date, there have been thousands of EU projects involving the CS, especially in the areas of life and natural sciences [63,64]. In 2015 it was estimated that 25% of the projects involving CS in the world concerned marine or coastal areas [65]. In fact, CS may allow to monitor biodiversity for large- scale (temporal and spatial) and to detect long-term changes of ecosystem [66]. In the context of marine organism invasions, CS can help to monitor the distribution, behavior, and dynamics of NIS and make the difference for the early detection, allowing to cover large geographical areas at a lower coast. Indeed, the presence of invasive species and their impacts on the marine ecosystem remain largely unrecorded due to logistical difficulties and the need to invest huge resources [67]. Encarnação et al. [68] performed a literature review to understand the effective contribution of CS for the detection and the monitoring of NIS across different ecosystem and highlighted that literature on the marine ecosystems was almost the double respect to the terrestrial ones in term of recorded NIS.
Kousteni et al. [66] investigated the literature records of NIS of fish and molluscs in the European marine waters provided by the CS. A total of 8513 records of NIS, corresponding to 68 species of Actinopterygii and 72 of Mollusca, were reported from 1905 to 2019. It is interesting to underline that the highest CS records among fish were those of the toxic invasive L. sceleratus (24.85%). This species shows in fact an unprecedented invasive character in terms of both abundances and geographical range [69], to be considered among the fastest expanding Lessepsian immigrants.
In the Mediterranean basin, such types of projects have been carried out to monitor coral reefs, gelatinous plankton, fish and NIS [67]. In our previous project [45], the dissemination activities were carried out using brochures, posters, reports, socials and newspaper articles. In addition, direct contacts with fishermen, divers and control authorities were established to create a network for the collection of data for a better assessment of the risk related to the spreading of toxic pufferfish. The project did not result in the collection of pufferfish specimens.
In the CS project ‘‘AlienFish’’, aimed at monitoring rare NIS of fish in Italian waters the pivotal role of the involvement of sea users and professional fishermen was highlighted. Therefore, in this study we decided to directly involve fishermen for monitoring the presence of pufferfish species in the area of interest. Interestingly when the informative brochure describing the 4 species recorded along the Italian coasts (see section 2.1.1) was distributed, some fishermen immediately recognized the species S. pachygaster. The other pufferfish species were instead not known. Specimens’ collection began immediately after the first contact in January 2020, demonstrating how the fishermen were aware of the regular presence of S. pachygaster in the water surrounding the Island. In fact, during infomal preliminary contact with fishermen, we were informed that this species is commonly fished as by-catch during their routine fishing. The 56 specimens analyzed in this study were collected by only five fishermen out of the twenty who confirmed their willingness in participating in this project (Table 3). With respect to the number of collected specimens, our study ranks only second after that by Ragonese et al. [70], dated over two decades ago, in which 403 specimens of S. pachygaster were collected in the Strait of Sicily (Italy). In both cases the collection was achieved thanks to the involvement of local fishermen.
3.2. Occurrence of S. pachygaster in the Mediterranean and Italian waters: data from our specimens’ collection and literature.
S. pachygaster is a circumglobally distributed species found in temperate and tropical waters [55]. Its presence has been reported worldwide including South Atlantic Ocean, Atlantic French waters, Irish waters, Canadian waters, Cape Verde islands until New Zealand waters [54]. However, most of the biogeographical information available online reports the presence of this species in the (eastern) Atlantic Ocean and around the Australian continent [54]. For this reason, the most plausible hypothesis for its expansion in the Mediterranean Sea is its entrance from the Atlantic Ocean (from the West African coastline) through the Strait of Gibraltar [71], even though a Lessepsian migration cannot be excluded given the circumglobal distribution of the species [2].
3.2.1. Overall specimens’ number and date of catch
According to the review of Guardone et al. [2], 716 specimens of S. pachygaster were recorded in the Mediterranean Sea from 1979 to July 2016. By analysing the 13 new records (Table 2), we observed that the overall number of Mediterranean S. pachygaster was 45 (from 2017 to 2022). By adding the 56 specimens collected in the present study to those reported from the literature ([2] and new records) 817 specimens were recorded in the Mediterranean waters from 1979 to 2022. Interestingly, 561 of them (68.7%) were caught in the Central Mediterranean (CMED) and 550 (98% of the CMED specimens) were caught in the Strait of Sicily (Italy).
As regards the 56 specimens collected in this study, 42 (75%) were caught in 2020 (February – September) and 14 (25%) in 2021 (January – May) (Table 3). The mean number of specimens collected per capture (n=16) was 3.3, with a range of 1- 5 specimens for all considered months, except for July 2020 in which 12 specimens were collected in a single capture. This data cannot be compared with those from the literature because in many cases catch ranges were given (e. g. “colleted from 2016 to 2020”), without specifying how many specimens per year were actually caught. However, extrapolating the number of specimens only for the period 2020 - 2021 (corresponding to the collection period of this study) of the new records, we found that five specimens were collected in 2020 and only one in 2021 (Table 2). Only the one in 2021 was recorded in the same catch area considered in this study (Strait of Messina, Italy). Thus, the records related to the specimens collected in this study represent the first 2020 record for S. pachygaster in Sicily. Respect to the months, the literature comparison did not allow to confirm an eventual seasonal catch pattern (with its peak a in July) since the months are often not detailed neither in Guardone et al. [2] nor in the new records.
3.2.2. Specimens’ total length and body weight
Despite specific instructions were provided to the fishermen not all the specimens were weighed on board. In addition, also considering the difficulty in logistics, some specimens were not stored in the correct way limiting the measurements. Totally, we were able to obtain both weight and length for 45 out of 56 specimens (80.3%). All the specimens from 2020 were weighed and measure while only three from 2021. In fact, considering that the project was intended to last only one year (2020), the instruction given to fishermen were not always followed during the captures in 2021. Moreover, only two out of the five fishermen continued to collect the specimens. The specimen total length and body weight of each specimen are reported in Table 3. The mean total length was 254 mm (range of 160- 450 mm), while the mean body weight was 635.1 g (range weight of 100- 2570 g). By analysing these data from new records, we observed that the specimen mean total length was 407.4 mm (n=5) with a range of 190-520 mm (n=17) while the mean body weight was 1022.2 g (n=9) with a range of 130- 2420 g (n=24). The mean total length, with relative ranges, found in the collected specimens and in new records are in line with those reported in Guardone et al. [2]. The mean body weight of the specimens collected in our study was instead lower than that of Guardone et al. [2] (despite being calculated on a similar number of specimens: n=45 and n=41 respectively) while the specimen mean total length of the update was higher but calculated on a very low number of subjects (n=5).
3.2.3. Additional information on catch areas and fishing method
The catch area of the specimens collected in this study was distributed in a range from 18 nautical miles eastward to 12 nautical miles westward from the Island of Lampedusa. Overall, 16 GPS coordinates corresponding to the fishing areas were provided by fisherman (Figure 1). As it can be seen in Figure 1 the area around Lampedusa is not extensive and all the five fishermen tend to follow similar routes. Since all the specimens were caught using trawls, each GPS coordinate represents an indicative point of the route travelled during the day’s outing. In fact, fishermen normally sound the bottom of small areas of sea during the day. These 16 GPS coordinates can be grouped into six distinct areas (from A to F) (Figure 1) in which captures were concentrated. Each area is associated to one to six GPS coordinates, corresponding to the number of capture (Figure 1). The number of specimens caught in each area is similar, except in areas A (n=16) and E (n=25) where the most part of catches occurred. The Fisherman 5 was the one who caught most of the specimens (n=31; 55.3%), followed by Fisherman 1 (n=10; 17.8%); Fisherman 2 (n=7;12.6%), Fisherman 4 (n=5; 8.9%), Fisherman 3 (n=3; 5.4%). Details on fishermen, captures and specimens collected per area are given in Table S2.
The area of competence of the Lampedusa fishermen, characterized by highly irregular bottom bathymetry where very great depths are reached [72], represents the ideal habitat for S. pachygaster. This species is in fact benthopelagic usually living between 50 and 250 m [73], but also up to 480 m depth [74]. In this study, the specimen mean catch depth, calculated using the 16 GPS coordinates, was 96.8 m, with a range of 16-170 m (Table S3). However, the most part of specimens (n=33; 58.9%) were caught at depth over than 120 m. The specimen mean depth of catch observed in new records was higher (181 m) with a range from 10 to 400 m (Table 2). However, not considering the record from Hussein et al. [54], related to the catch one specimens by the coast, the range varies from 120 to 400. Overall, the comparison of the analysis of specimen’s depth of catches from this study and from the new records are in line with Akbora et al. [31] who reported that this species is mostly fished at depths greater than 100 m, and consistent with what was reported in Guardone et al. [2] (mean depth 149.3 m).
As regards the type of seabed, in this study it corresponds to medium sand or sand (n=7; 43.8%) and fine sand or silty sand (n=9; 56.2%) in deeper waters (Table S3). The mean depth of catch calculated for the two types o seabed was 54.7 m and 129.4 m, respectively. Respect to the new records only one reported the type of seabed (soft bottom) without specifying the depth (Table 2).
Because of its natural habits, S. pachygaster is normally a by-catch of semi-industrial fisheries, purse seines bottom trawls, gillnets, trammel-nets, bottom long-lines and drift longline [54]. According to some authors, this species could be considered invasive because it could constitute an economic burden not to be underestimated for fishermen that have to discard such a large number of specimens from their gear [75]. It is interesting to underline that Strait of Sicily was selected precisely by Ragonese et al. [70] because there were one of the largest trawling fleets. Also, today bottom trawling is the most significant fishing activity in this area [76], which, turns out to be, as already mentioned, the fishing method most frequently involved during the catch of S. pachygaster. These fishing methods are in line with the seabed types found in this study and those reported by Guardone et al. [2] (muddy or muddy/sandy). Although in Guardone et al. [2] the fishing method was categorised without specifying the type of net used, 100% of S. pachygaster were caught by commercial fishing. In fact, trawls can only be used with a sandy seabed. As regards the 45 specimens described in the new records, 44 (97.8%) were caught while only one specimen was found stranded on the beach (Table 2). Among those caught, the most used fishing method, reported for 28 (63.6%) specimens, was in fact trawling (n=22; 78.6%) followed by longline (n=3; 10.7%), trammel-net (n=2; 7.1%) and only in case by a fisherman 300 m from the coast (3.6%) (Table 2).
Overall, the results of this study support what is reported in the literature, confirming that since 1990s the area of the Strait of Sicily hosted an established population. Some authors have suggested that this species arrived here prior to the date of the first Mediterranean record [73]. Moreover, the wide size range and the numerical consistent found in this study, as in that of Ragonese et al. [70], indicates that a complete sexual cycle exists supporting the fact that it can be considered stable in these waters. Therefore, S. pachygaster could be considered an established or naturalized population according to the definitions reported in Zenetos et al. [77].
3.3. TTXs detection
While no limit is set for TTXs and pufferfish should not be commercialized, the Regulation (EC) No 853/2004 sets a maximum PSTs concentrations of 800 mg STX eq./kg in bivalve molluscs, echinoderms, tunicates and marine gastropods [78]. This is also the maximum permit level in US and Canada, while in Japan the limit is higher (2 mg STX eq./kg) [39,79]. In a scientific opinion, the EFSA Panel on Contaminants in the Food Chain recognized the need for further data on the acute oral toxicity of TTX [9]. In addition, the EFSA Panel suggested that, since TTXs and STXs had similar modes of action and their toxicity is probably additive, they could perhaps be combined to yield one health-based guideline value [9]. For this reason, it would be much accurate evaluate them together.
In our study, for the first time, 20 specimens of S. pachygaster collected along the Italian coasts were analyzed for the presence of TTXs (Table 3). In addition, 50 tissue samples belonging to ten specimens (among those analyzed for TTXs) (Table 3) were randomly analyzed for PSTs (including STXs) detection to use as screening. In fact, despite the method used in this study was not fully validated for the presence or absence of PSTs in fish tissue, EFSA recommends assessing the presence of both these guanidinium neurotoxins [9]. In addition, Mediterranean S. pachygaster was never investigated for the presence of STXs, although poisoning cases associated with specimens belonging to the genus Sphoerodies were reported in North America, although rather dated [17,18]. All the 20 specimens analyzed for TTXs, of which 7 (35%) above the mean body weight reported in Guardone et al. [2] and 13 (65%) smaller, resulted negative. In fact, TTXs concentrations found were below the LOQ in each tissue analysed (<10 µg/kg). Also, the 10 specimens randomly analyzed (three large and seven small) for STX detection, resulted negative.
Data regarding the toxicity of S. pachygaster is discordant. For instance, this species is considered as non-toxic along Japanese coast [80,81,82]. Also, Amano et al. [83] reported that all the 4 specimens caught in the East China Sea in 2009 were found as not containing TTX in any examined tissue. Interestingly, S. pachygaster was the only TTX negative species among the five analysed in that study [83]. Contrariwise, S. pachygaster of pacific waters was considered as weakly toxic [5]. In the Mediterranean, it was instead reported as non-toxic [38]. Factually, genes for homologous TTXs and STXs binding protein were detected in S. pachygaster, as observed in other species which are known as toxic [84]. In the study of Nagashima et al. [85], liver tissue slices of non- toxic species of pufferfish incubated with TTX accumulated the toxin after 8 h, regardless of the toxicity of the species. In addition, poisoning cases related to ingestion of Sphoerides spp. (including S. testudineus and S. maculatus) are reported in Centre- South America and Oceania [12], suggesting that species belonging to this genus can accumulate toxins. Furthermore, TTXs was found in two Sphoeroides species (S. annulatus and S. lobatus) caught near the Mexican Peninsula of Baja California [86]. Despite human poisonings due to pufferfish ingestion is usually recollected to TTXs, we should take into consideration that in the large majority of the cases, no laboratory analysis was carried out to confirm the toxin involved and the TTX intoxication was clinically diagnosed on the basis of the anamnesis and of the symptoms [12]. In fact, the occurrence of TTXs in some pufferfish species has been reported together with STXs [87]. For example, STX-positive (not TTX) pufferfish belonging to the genus Sphoeroides have been recorded since 2002 in the USA [88]. Worldwide, STXs has been found along with TTXs in several marine and freshwater pufferfish species in areas of the Far East, including Japan, Philippines and Thailand [18]. Several species belonging to the Sphoeroides genus were recently found containing STXs in American waters [18,89]. Thereofore, it would be more accurate to say that S. pachygaster could accumulate TTXs and STXs in particular conditions [55]. Actually, only 3 studies have investigated the presence of TTXs in specimens of S. pachygaster caught in Mediterranean basin [21,27,36] and no one has investigated STX in this species (Table 1). Reverté et al. [21] analyzed two pufferfish, of which one S. pachygaster caught in Spain, and they resulted negative to TTXs. Rambla-Alegrè et al. [27] evaluated the presence of TTXs in 5 specimens of S. pachygaster (on a total of 20 specimens of 3 different species of pufferfish) caught along the Mediterranean coast of Spain, and all were found as non-containing TTXs. Recently, Ulman et al. [36] reported the absence of TTXs in one S. pachygaster specimen caught in the Eastern Mediterranean Sea (EMED). To date, the exact biosynthetic pathways and genes implicated in the TTX production are still unclear [90]. However, the available information, suggest that marine organisms, like pufferfish, acquire TTXs from other toxic organisms bearing bioconcentrated TTXs through the food chain, with marine bacteria being at the start [5]. Some studies have in fact demonstrated that wild toxic pufferfish species bred under controlled conditions and artificially fed with diets do not present toxin while, such non-toxic pufferfish become toxic when fed with TTX-containing diets [5]. A possible explanation could be that marine organisms consume microalgae (dinoflagellates), which host TTXs-producing symbiotic bacteria, and accumulate them along the trophic chain [3]. This theory, involving microalgae in bacterial symbiosis, is supported by the fact that significant concentrations of TTXs have been detected in dinoflagellate cell cultures [91]. Another study in which TTXs was found in bivalve molluscs collected in the Gulf of Syracuse (Sicily, Italy) supports this hypothesis. In fact, the highest abundance of TTXs in dinoflagellates was found at the same time as toxins were found in mussels. A possible correlation between the presence of TTXs in molluscs and species of dinoflagellates was also hypothesized in a study conducted in the Aegean Sea (Greece) [92]. This demonstrates how phytoplankton could play a key role in TTXs transfer [40]. Whereas TTXs in pufferfish is mostly of bacterial origin, STXs originates from microalgae, principally from dinoflagellates in marine species and from cyanobacteria in freshwater species [15]. In the case that pufferfish consume crustaceans and molluscs exposed to dinoflagellate blooms, they may turn in vectors for STXs [18]. In fact, it has been assumed that Japanese Takifugu found with high concentration had eaten bivalves exposed to the toxic dinoflagellate [93]. However, questions about the origin of STXs arise from the study of Pinto et al. [59] in which, despite pufferfish were found positive for STXs, the PSTs were never found in marine organisms from Madeira Island. Neither PSTs-producing dinoflagellates, such as Gymnodinium catenatum and Alexandrium spp. are known in this area [94]. However, we have to consider that also some xanthid crabs, horseshoe crabs and marine snails may possess PSTs, not associated with dinoflagellates. For instance, some authors reported that pufferfish and bivalves could acquire TTXs by consuming toxic planoceri flatworms [95]. In addition, because the skin and mucus of all toxic pufferfish species routinely contained the highest levels of STXs respect to other tissue, this suggests that, in the absence of toxic food items, STXs could be taken up via an undetermined pathway. Despite this, the main source of TTXs and STXs remains bivalve molluscs [96]. As already mentioned, TTXs production in dinoflagellate species may be due to its relationship with symbiotic bacteria, which are able to produce toxic compounds when environmental factors are favorable. Water temperature, salinity and depth are among the environmental factors that play a key role in favoring the occurrence of TTXs in marine organisms. In this study, the water mean temperature and salinity at point of catch was 15.8 °C (range 15.3 – 17.6 °C) and 36.8‰ (range 36- 38‰), respectively (temperature and salinity of which GPS coordinates are reported in Table S3). According to the available literature, TTXs were typically present in shellfish collected at temperatures of 15°C [90], even though Turner et al. [61] did not correlate TTXs occurrence and water temperature. Bordin et al. [97] reported TTXs in shellfish collected in Northern Adriatic Sea (Italy), in May when the water temperature ranged between 19°C and 21°C, while Vlamis et al. [92] found TTX-positive mussels during the same period, but they did not report the water temperature. In Leão et al. [98] low concentration of TTXs in shellfish, collected in Spain, were found in samples collected in July from an intertidal area with a water temperature of 17°C. It seems that abiotic factors could stimulate TTXs production in bacteria, such as Vibrio [61] and bacterial counts seemed to be influenced by temperature and salinity, in particular higher counts occurred in warmer months and in water with lower salinity values [99]. As regards water depth shellfish resulted positive to TTXs were collected in inter-tidal or water where salinity level was lower than open marine waters [79]. Also, Dell’Aversano et al. [40] highlighted how warm water, low water depth and strong solar radiation are potential critical factors for TTXs accumulation. In addition to TTXs contamination with a level up to 6.4 mg TTX eq/Kg, they found significant level of contamination from PST in mussels collected in Syracuse Bay (Sicily) during spring- summer of 2015, 2016 and 2017, demonstrating how TTXs levels can also be associated with STXs levels in mussels. Bivalve molluscs were collected at 10 sites in concomitance with a mixed bloom of Alexandrium spp producer of PST [40]. In the study of Bacchiocchi et al. [95], bivalve molluscs with the highest toxin concentrations came from shallow waters (20-40 cm) with high temperatures (20-40 °C) and salinity (from 34.3 to 36) suggesting that environmental factors such as strong solar radiation and relatively high water temperature may favor TTX accumulation in mussels.
All these factors could explain the TTXs and STXs occurrence in bivalve molluscs or pufferfish collected from different geographical areas. In fact, other studies reported the co-occurence of TTXs and PSTs in bilvalves and gastropods [95,100,101]. Therefore, the different toxicity pattern of S. pachygaster and L. sceleratus could be related to their habitat depth despite both feed on molluscs, shrimps and small teleosts. In fact, as already detailed in section 3.2.2. S. pachygaster usually lives over 100 m of depth while L sceleratus is mostly found in shallow waters (mean 24.3 m) [2]. Factually, TTXs have not been found in the Mediterranean at least until 2003 [59]; probably, the propagation of TTXs-producing bacteria in the Mediterranean waters has been favoured by climatic changes and transports through the Suez Canal [19]. Therefore, we cannot exclude that other non-toxic Mediterranean species, such as S. pachygaster, may accumulate the TTX through the trophic chain.
4. Conclusions
This is the first survey aimed at evaluating the toxicity of S. pachygaster specimens collected along the Italian coasts (Strait of Sicily). With 56 collected specimens in 2020-2021, this sampling is the largest after another performed over 15 years ago. The specimens’ collection was possible thanks to the involvement of local fishermen, especially trained for the identification of pufferfish species inhabiting the Italian coasts, confirming the key role of the CS in the monitoring on marine NIS. All the analysed specimens were found to be negative for the presence of TTXs, and part of them, also analyzed for STXs as screening, also resulted negative. However, despite the assumed non-toxicity of S. pachygaster in the study area, further studies should be conducted, especially in the light of climate change that, affecting water temperature, could influence the production of toxins by bacteria and dinoflagellates. Overall this study, other than proving that S. pachygaster is currently well established in the Strait of Sicily provide new data that are essential for a proper assessment of this emerging risk.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: informative brochure distributed to the fishermen; including pictures and morphological description of the 4 pufferfish species already recorded along the Italian coasts (Lagocephalus lagocephalus, Lagocephalus scerelatus, Sphoeroides marmoratus and Sphoeroides pachygaster); Table S1: HILIC-MS/MS set of employed parameters; Table S2: Details of captures, fishermen (Fm) and specimens collected per area; Table S3: Fisherman (Fm), catch date, number of specimens caught, GPS coordinates with relative depth, type of seabed, temperature and salinity.
Author Contributions
Conceptualization, A.A.; L. Gu; L. Ga; data curation C.M; formal analysis, C.M, B. R. S. D..; funding acquisition, A. A. investigation B. R.; L. Gu, S. D; methodology A.A.; A. G. project administration, A.A; supervision, A.A.; writing—original draft, C.M, A.G and A.A; writing—review & editing A.G and A.A, .
Funding
this research was financed with funds granted from the University of Pisa.
Data Availability Statement
Data is included in the manuscript and available in supplementary tables attached to the text; further data is also available at the authors on request.
Acknowledgments
We thank Dr. Rizzo for the supporting in the project organization and all the fishermen of Lampedusa for their help in collecting the samples.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Tonon, L. A. C.; de Azevedo, G. P.; Monteiro, A. F.; Bernardi, D. I.; Gubiani, J. R., Ioca, L. P.; Mattsson, H.K.; Moreira, A. P. B.; Gomes, A. F.; Pires, O. R. jr.; Pedrosa, C. S. G.; Souza, L. R.Q.; Rehen. S. K.; Thompson, C. C.; Thompson, F. L.; Berlinck, R. G. New tetrodotoxin analogs in Brazilian pufferfishes tissues and microbiome. Chemosphere 2020, 242, 125211. [CrossRef]
- Guardone, L.; Gasperetti, L.; Maneschi, A.; Ricci, E.; Susini, F.; Guidi, A.; Armani, A. Toxic invasive pufferfish (Tetraodontidae family) along Italian coasts: Assessment of an emerging public health risk. Food Control 2018,91, 330-338. [CrossRef]
- Katikou, P.; Gokbulut, C.; Kosker, A. R.; Campàs, M.; Ozogul, F. An updated review of tetrodotoxin and its peculiarities. Mar. drugs 2022, 20(1), 47. [CrossRef]
- Fernández-Ortega, J. F.; Morales-de los Santos, J. M.; Herrera-Gutiérrez, M. E.; Fernández-Sánchez, V.; Loureo, P. R.; Rancaño, A. A.; Téllez-Andrade, A. Seafood intoxication by tetrodotoxin: First case in Europe. J Emerg. Med. 2010; 39(5), 612-617. [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin–distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. drugs 2008, 6(2), 220-242. [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6(2), 693-755. [CrossRef]
- Jal, S.; Khora, S.S. An overview on the origin and production of tetrodotoxin, a potent neurotoxin. J. Appl. Microbiol. 2015, 119, 907–916. [CrossRef]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H., Doi, H. et al. Contrasting toxin selectivity between the marine pufferfish Takifugu pardalis and the freshwater pufferfish Pao suvattii. Toxins 2019, 11(8), 470. [CrossRef]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain); Knutsen, H.K.; Alexander, J.Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Scientific opinion on the risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues inmarine bivalves and gastropods. EFSA J. 2017, 15, 4752.
- Cembella, A.D.; Durán-Riveroll, L.M. Chapter One—Marine guanidinium neurotoxins: Biogenic origins and interactions, biosynthesis and pharmacology. In Advances in Neurotoxicology; Novelli, A., Fernández-Sánchez, M.T., Aschner, M., Costa, L.G., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 1–47. [CrossRef]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins 2017, 9, 166. [CrossRef]
- Guardone, L.; Maneschi, A.; Meucci, V.; Gasperetti, L.; Nucera, D.; Armani, A. A global retrospective study on human cases of tetrodotoxin (TTX) poisoning after seafood consumption. Food Rev. Int. 2020, 36(7), 645-667. [CrossRef]
- Gao, Y.; Yu, R. C.; Murray, S. A.; Chen, J. H.; Kang, Z. J.; Zhang, Q. C.; Kong, F.Z.; Zhou, M. J. High specificity of a quantitative PCR assay targeting a saxitoxin gene for monitoring toxic algae associated with paralytic shellfish toxins in the Yellow Sea. Appl. Environ. Microbiol. 2015, 81(20), 6973-6981. [CrossRef]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [CrossRef]
- Raposo, M. I.; Gomes, M. T. S.; Botelho, M. J.; Rudnitskaya, A. Paralytic shellfish toxins (PST)-transforming enzymes: A review. Toxins 2020, 12(5), 344. [CrossRef]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, Function and Pharmacology of Voltage-gated Sodium Channels. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362(6), 453–479. [CrossRef]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M.; Abbott, J.P.; Flewelling, L.J.; Richardson, R.W.; Dickey, R.W.; Jester, E.L.E.; Etheridge, S.M.; Deeds, J.R.; Van Dolah, F.M.; Leighfield, T.A.; Zou, Y.; Beaudry, C.G.; Benner, R.A.; Rogers, P.L.; Scott, P.S.; Kawabata, K.; Wolny, J.L.; Steidinger, K.A. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect 2006 114, 1502–1507. [CrossRef]
- Abbott, J. P.; Flewelling, L. J.; Landsberg, J. H. Saxitoxin monitoring in three species of Florida puffer fish.Harmful Algae 2009, 8(2), 343-348. [CrossRef]
- udi, M.; Abdelmouleh, A., Jamoussi, K., Kammoun, A.; El Feki, A. Hematological toxicity associated with tissue extract from poisonous fish Lagocephalus lagocephalus—Influence on erythrocyte function in wistar rats. J Food Sci. 2008, 73(7), H155-H159. [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi; A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54(1), 50-55. [CrossRef]
- Reverté, L.; De La Iglesia, P.; Del Río, V.; Campbell, K.; Elliott, C.T.; Kawatsu, K.; Katikou, P.; Diogène, J.; Campàs, M. Detection of Tetrodotoxins in Puffer Fish by a Self-Assembled Monolayer-Based Immunoassay and Comparison with Surface Plasmon Resonance, LC-MS/MS, and Mouse Bioassay. Anal. Chem. 2015, 87, 10839–10847. [CrossRef]
- Bane, V.; Brosnan, B., Barnes, P.; Lehane, M.; Furey, A. High-resolution mass spectrometry analysis of tetrodotoxin (TTX) and its analogues in puffer fish and shellfish. Food Addit. Contam. Part A 2016, 33(9), 1468-1489. [CrossRef]
- Kırımer, N.; Göger, F.; Kürkçüoğlu, M.; Özbek, E. Ö.; Coban, B.; Balcıoğlu, E. B.; Öztürk, B.; Güven, K. C.. Tetrodotoxin and fatty acids contents of Lagocephalus sceleratus (Gmelin, 1789) collected in Antalya, Turkey, by MS/MS and GC/MS analyses. J. Black Sea/Medit. Environ 2016, 22(3), 278-288.
- Kosker, A. R.; Özogul, F.; Durmus, M.; Ucar, Y.; Ayas, D.; Regenstein, J. M.; Özogul, Y. Tetrodotoxin levels in pufferfish (Lagocephalus sceleratus) caught in the Northeastern Mediterranean Sea. Food chem. 2016, 210, 332-337. [CrossRef]
- Acar, C.; Ishizaki, S.; Nagashima, Y. Toxicity of the Lessepsian pufferfish Lagocephalus sceleratus from eastern Mediterranean coasts of Turkey and species identification by rapid PCR amplification. Eur. Food Res.Technol. 2017, 243, 49-57. [CrossRef]
- Alabssawy, A. N. Antimicrobial activity of Tetrodotoxin extracted from liver, skin and muscles of puffer fish, Lagocephalus sceleratus inhabiting Mediterranean Sea, Egypt. Int. J. Cancer Biomed. Res. 2017, 1(1), 2-10. [CrossRef]
- Rambla-Alegre, M.; Reverté, L.; Del Río, V.; de la Iglesia, P.; Palacios, O.; Flores, C.; Elliott, C. T.; Maillaude, C.; Boundy, M. J.; Harwoodf, D. T.; et al. Evaluation of Tetrodotoxins in Puffer Fish Caught along the Mediterranean Coast of Spain. Toxin Profile of Lagocephalus Sceleratus. Environ. Res. 2017, 158, 1–6. [CrossRef]
- Kosker, A. R.; Özogul, F.; Durmus, M.; Ucar, Y.; Ayas, D.; Šimat, V.; Özogul, Y. First report on TTX levels of the yellow spotted pufferfish (Torquigener flavimaculosus) in the Mediterranean Sea. Toxicon 2018, 148, 101-106. [CrossRef]
- Leonardo, S.; Kiparissis, S.; Rambla-Alegre, M.; Almarza, S.; Roque, A., Andree, K. B.; Christidis, A.; Flores, C.; Caixach, J., Campbell, K. et al. Detection of tetrodotoxins in juvenile pufferfish Lagocephalus sceleratus (Gmelin, 1789) from the North Aegean Sea (Greece) by an electrochemical magnetic bead-based immunosensing tool. Food chemistry 2019, 290, 255-262. [CrossRef]
- Kosker, A. R.; Özogul, F., Ayas, D.; Durmus, M.; Ucar, Y.; Regenstein, J. M.; Özogul, Y. Tetrodotoxin levels of three pufferfish species (Lagocephalus sp.) caught in the North-Eastern Mediterranean sea. Chemosphere 2019, 219, 95-99. [CrossRef]
- Akbora, H.D.; Kunter, ˙I.; Erçetin, T.; Elagöz, A.M.; Çiçek, B.A. Determination of tetrodotoxin (TTX) levels in various tissues of the silver cheeked puffer fish (Lagocephalus sceleratus (Gmelin, 1789)) in Northern Cyprus Sea (Eastern Mediterranean). Toxicon 2020, 175, 1–6. [CrossRef]
- Ujević, I.; Roje-Busatto, R.; Dragičević, B.; Dulčić, J. Tetrodotoxin in invasive silver-cheeked toadfish lagocephalus sceleratus (Gmelin, 1789) in the Adriatic Sea. The Montenegrin Adriatic Coast: Mar. Chem. Poll. 2021, 141-149. [CrossRef]
- Christidis, G.; Mandalakis, M.; Anastasiou, T. I.; Tserpes, G.; Peristeraki, P.; Somarakis, S. Keeping Lagocephalus sceleratus off the Table: Sources of Variation in the Quantity of TTX, TTX Analogues, and Risk of Tetrodotoxication. Toxins 2021, 13(12), 896. [CrossRef]
- Farrag, M. M. Toxicity pattern of pufferfish Lagocephalus sceleratus (Gmelin, 1789), Mediterranean Sea, Egypt: Awareness and food safety. Aqua. Aquar. Conserv. Legis. 2022, 15(2), 941-962.
- Hassoun, A. E. R.; Ujević, I.; Jemaa, S.; Roje-Busatto, R.; Mahfouz, C.; Fakhri, M.; Nazlić, N. Concentrations of tetrodotoxin (TTX) and its analogue 4, 9-anhydro TTX in different tissues of the silver-cheeked pufferfish (Lagocephalus sceleratus, Gmelin, 1789) caught in the South-Eastern Mediterranean Sea, Lebanon. Toxins 2022, 14(2), 123. [CrossRef]
- Ulman, A.; Tekin, H.; Aysal, A. I.; Akbora. H.D.; Çiçek, B. A. Contribution to puffer toxicity: TTX analysis of three pufferfish species in the eastern mediterranean. In Abstract and Proceeding book of 2nd International symposium on Pufferfish/Lionfish. Turan, C. (Eds), Published by Natural and Engineering Sciences, Iskenderun, Turkey, 20-22 May 2022, pp 11.
- Zenetos A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; Garcia Raso, J.; Cinar, M.; Almogi-Labin, A.; Ates, A.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13(2), 328–352. [CrossRef]
- Ragonese, S.; Morara, U. Scientific note evidence of short-term discard mortality of trawled Sphoeroides pachygaster (Osteichthyes, Tetraodontidae) off the southern coast of Sicily (Central Mediterranean Sea). Pan-Am. J. Aquat. Sci. 2012, 7, 73-76.
- Antonelli, P.; Salerno, B.; Bordin, P.; Peruzzo, A.; Orsini, M.; Arcangeli, G.; Barco, L.; Losasso, C. Tetrodotoxin in live bivalve mollusks from Europe: Is it to be considered an emerging concern for food safety? Compr. Rev. Food Sci. and Food Saf., 2022 21(1), 719-737. [CrossRef]
- Dell’Aversano, C.; Tartaglione, L.; Polito, G.; Dean, K.; Giacobbe, M.; Casabianca, S.; Capellacci, S.; Penna, A.; Turner, A.D. First detection of tetrodotoxin and high levels of paralytic shellfish poisoning toxins in shellfish from Sicily (Italy) by three different analytical methods. Chemosphere 2019, 215, 881–895. [CrossRef]
- Poursanidis, D.; Zenetos, A. The role played by citizen scientists in monitoring marine alien species in Greece. Cah. Biol. Mar. 2013, 54, 419-426.
- Vohland, K.; Land-Zandstra, A.; Ceccaroni, L.; Lemmens, R.; Perelló, J.; Ponti, M.; Samson, R.; Wagenknecht, K. The Science of Citizen science. Gewerbestrasse: Springer 2021. [CrossRef]
- Directorate-General for Maritime Affairs and Fisheries, 2022. Dealing with a wicked guest, with some EU help. https://oceans-and-fisheries.ec.europa.eu/news/dealing-wicked-guest-some-eu-help-2022-06-02_enISPRA, 2016. Attenzione al pesce palla maculato è tossico e non va mangiato! https://www.isprambiente.gov.it/files/comunicati-stampa/2015/Locandina_pesce_palla.pdf.
- Guidi, A.; Armani, A.; Guardone, L.; Guarducci, M.; Gasperetti, L.; Susini, F.; Longo, M.; Maneschi, A., Dadovich, N. Climate change and food safety: Citizen science for monitoring the presence of toxic alien fish species in Italian waters. J. Bio. Res. 2016, (Italy), 89(1S), 13.
- Amante, C. and B. W. Eakins, ETOPO1 1 Arc-Minute Global Relief Model: Procedures, Data Sources and Analysis. NOAA Technical Memorandum NESDIS NGDC-24, 19 pp, March 2009.
- Boyer, Tim P.; García, Hernán E.; Locarnini, Ricardo A.; Zweng, Melissa M.; Mishonov, Alexey V.; Reagan, James R.; Weathers, Katharine A.; Baranova, Olga K.; Paver, Christopher R.; Seidov, Dan; Smolyar, Igor V. (2018). World Ocean Atlas 2018. [indicate subset used]. NOAA National Centers for Environmental Information. Dataset. https://www.ncei.noaa.gov/archive/accession/NCEI-WOA18. Accessed [5 may 2023].
- Malloggi, C.; Tinacci, L.; Giusti, A.; Galli F. Dall'Ara, S.; Marconi, P.; Gasperetti, L.; Armani, A. Accidental discovery of a Tetraodontidae (Sphoeroides marmoratus) within a cuttlefish (Sepia officinalis) bought in a fish shop in Italy: Risk assessment associated with the presence of Tetrodotoxin- Ital. J. Food. Saf. 2023, Accepted in press.
- Cammilleri, G.; Graci, S.; Buscemi, M. D.; Collura, R.; Costa, A.; Dico, G. M. L., Cumbo, V. New report of Anikasis larvae from Blunthead Puffer, Sphoeroides pachygaster caught off Strait of Sicily. Biodivers. J. 2019, 10(4), 475-478. [CrossRef]
- Carlucci, R.; Mentino, D.; Semeraro, D.; Ricci, P.; Sion, L.; Scillitani, G. Comparative histochemical analysis of intestinal glycoconjugates in the blunthead pufferfish Sphoeroides pachygaster and grey triggerfish Balistes capriscus (Teleostei: Tetraodontiformes). J. Fish Biol. 2019, 94(1), 122-131. [CrossRef]
- Ragheb, E.; Rizkalla, S. I; Analyses of the non-target catch from the Egyptian Mediterranean trawlers, off Port Said. Egypt. J. Aquat. Res. 2019, 45(3), 239-246. [CrossRef]
- Vella, A.; Vella, N.; Karakulak, F. S.; Oray, I. DNA barcoding of Tetraodontidae species from the Mediterranean Sea: Filling knowledge gaps for improved taxonomic accuracy. Genet. Aquat. Org. 2017, 1(2), 61-69. [CrossRef]
- Gerovasileiou, V., Akel, E. S. H. K., Akyol, O. K. A. N.; Alongi, G.; Azevedo, F.; Babali, N et al. New New Mediterranean Biodiversity Records (July 2017). Mediterr. Mar. Sci. 2017, 18/2, 355-384. [CrossRef]
- Hussein, K. B.; Bensahla-Talet, L.; Chakouri, A. A review on the occurrence of the Blunthead puffer, Sphoeroides pachygaster (Müller & Troschel, 1848) in the Mediterranean with a new occurrence from Oran Bay (Western Algeria). Aquat. Scie. Eng. 2021,36(2), 89-94. [CrossRef]
- Akbora, H. D.; Snape, R.; Deniz, A. Y. A. S.; Çiçek, B. A. The first substantiated record of blunthead puffer Sphoeroides pachygaster (Müller and Troschel, 1848), from the coast of northern Cyprus (eastern Mediterranean). Mar. Sci. Tech. Bull. 2021, 10(1): 1-7. [CrossRef]
- Kizilkaya, İ. T.; Akyol, O. Additional Record of Sphoeroides pachygaster (Tetraodontidae) in the Aegean Sea (Fethiye, Turkey). COMU-JMSF 2020, 3(2), 136-139. [CrossRef]
- Gazzetta del Sud online- Messina. Available online: https://messina.gazzettadelsud.it/foto/cronaca/2021/08/20/pesce-palla-velenoso-ritrovato-gia-morto-nelle-acque-dello-stretto-foto-cc52a465-ea70-456c-bade-f04180224512/2/ (Accessed 5 may 2023).
- Karakuş, M.; İsmail Dal, Coşkun M. A.The first record of age findings of sphoeroides pachygaster (muller and troschel, 1848) in the eastern mediterranean sea. In Abstract and Proceeding book of 2nd International symposium on Pufferfish/Lionfish. Turan, C. (Eds), Published by Natural and Engineering Sciences, Iskenderun, Turkey, 20-22 May2022., pp 12.
- Pinto, E. P.; Rodrigues, S. M.; Gouveia, N.; Timóteo, V.; Costa, P. R. Tetrodotoxin and saxitoxin in two native species of puffer fish, Sphoeroides marmoratus and Lagocephalus lagocephalus, from NE Atlantic Ocean (Madeira Island, Portugal). Mar. Environ. Res. 2019, 151, 104780. [CrossRef]
- Boundy, M. J.; Selwood, A. I.; Harwood, D. T.; McNabb, P. S.; Turner, A. D. Development of a sensitive and selective liquid chromatography–mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1-12. [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the Pufferfish Toxin. Tetrodotoxin in European Bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [CrossRef]
- Eitzel, M.V.; Cappadonna, J.L.; Santos-Lang, C.; Duerr, R.E.; Virapongse, A.;West, S.E.; Kyba CC, M.; Bowser, A.; Cooper, C.B.;Sforzi, A.; et al. Citizen Science Terminology Matters: Exploring Key Terms. Citiz. Sci. Theory Pract. 2017, 2, 1–20. [CrossRef]
- European Commission. An Inventory of Citizen Science Activities for Environmental Policies. 2018 Brussels: European Commission.
- Science Europe (2018). Science Europe Briefing Paper on Citizen Science: D/2018/13.324/2. Brussels: Science Europe.
- Theobald, E. J.; Ettinger, A. K., Burgess, H. K.; DeBey, L. B., Schmidt, N. R.; Froehlich, H. E.; Wagner, C.; HilleRisLambers, J.; Tewksbury, J.; Harsch, M.A.; Parrish, J. K. Global change and local solutions: Tapping the unrealized potential of citizen science for biodiversity research. Biol. Conserv. 2015,181, 236-244. [CrossRef]
- Kousteni, V.; Tsiamis, K.; Gervasini, E.; Zenetos, A.; Karachle, P. K.; Cardoso, A. C. Citizen scientists contributing to alien species detection: The case of fishes and mollusks in European marine waters. Ecosphere,2022, 13(1), e03875. [CrossRef]
- Clements, K. R.; Karp, P.; Harris, H. E.; Ali, F.; Candelmo, A.; Rodríguez, S. J.; Balcàzar- Escalera, C.; Fogg, A. Q.; Green, S. J.; Solomon, J. N. The Role of Citizen Science in the Research and Management of Invasive Lionfish across the Western Atlantic. Diversity, 2021, 13(12), 673. [CrossRef]
- Encarnação, J.; Teodósio, M.A.; Morais, P. Citizen Science and Biological Invasions: A Review. Front. Environ. Sci. 2021, 8, 602980. [CrossRef]
- Nader, M. R., Indary, S., & Boustany, L. The puffer fish Lagocephalus sceleratus (Gmelin, 1789) in the Eastern Mediterranean. EastMed Technical Documents (FAO) 2012, GCP/INT/041/EC–GRE–ITA; FAO: Rome, Italy.
- Ragonese, S.; Jereb, P.; Morara, U. Morphometric relationships of Sphoeroides pachygaster (Pisces - Tetraodontidae) of the Strait of Sicily (Mediterranean Sea). Cah. de Bio. Mar. 1997, 38, 283e289.
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A. C. Building the European Alien Species Information Network (EASIN): A novel approach for the exploration of distributed alien species data. BioInvasions Records 2012, 1, 235–245. [CrossRef]
- Consoli, P.; Esposito, V.; Battaglia, P.; Altobelli, C.; Perzia, P.; Romeo, T. et al. Fish distribution and habitat complexity on banks of the Strait of Sicily (Central Mediterranean Sea) from remotely-operated vehicle (ROV) explorations. PLoS ONE 2016, 11(12), e0167809. [CrossRef]
- Ragonese, S.; Rivas, G.; Jereb, P. Spreading of the puffer Sphoeroides pachygaster Günther, 1870) (Pisces: Tetraodontidae) in the Sicilian channel. Is it an “exploding” population? Rapp. Comm. Int. Mer. 1992, 33, 308.
- Matsuura, K.; Tyler, J. C. Tetraodontiform fishes, mostly from deep waters, of New Caledonia. p.173-208. In Séret B. (ed.) Paris, France, 1997. Résultats des Campagnes MUSORSTOM, vol. 17. no.9. Mem. Mus. Natl. Hist. Nat. MNHN (Mémoires du Muséum National d’Histoire Naturelle. Zoologie), 174, 215 p 174.
- Golani, D.; Orsi- Relini, L.; Massutí, E.; Quignard, J. CIESM Atlas of Exotic Species in the Mediterranean. Vol. 1. Fishes. D 2003Sep.3067(3):377-8. Available at: https://scientiamarina.revistas.csic.es/index.php/scientiamarina/article/view/489. 8Accessed 5 may 2023).
- Jarboui, O.; Ceriola, L.; Fiorentino, F. Current fisheries management in the Strait of Sicily and progress towards an ecosystem approach. Fisheries and Aquaculture Technical Paper (FAO) 2022, 681,147-162. FAO: Rome, Italy.
- Zenetos, A.; Çinar, M. E.; Pancucci-Papadopoulou, M. A.; Harmelin, J. G.; Furnari, G.; Andaloro, F.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Med. mar. sci. 2005, 6(2), 63-118. [CrossRef]
- Regulation (EC) No 853/2004. Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. OJ L 139, 30.4.2004, 55–205.
- Turner, A. D.; Hatfield, R. G.; Maskrey, B. H.; Algoet, M.; & Lawrence, J. F. Evaluation of the new European Union reference method for paralytic shellfish toxins in shellfish: A review of twelve years regulatory monitoring using pre-column oxidation LC-FLD. TrAC 2019, 113, 124-139. [CrossRef]
- Masuda, H.; K. Amaoka, C.; Araga, T.; Uyeno T.; Yoshino, T. The fishes of the Japanese Archipelago. Tokai University Press, Tokyo, Japan, 1984, 1, 437.
- Tani, T. Nihonsan fugu no chudokugakutekikenkyu (Toxicological studies on Japanese puffer). Tokyo, Teikokutosho, 1945, 15-27.
- Jeong, D. Y.; Kim, D. S.; Lee, M. J.; Kim, S. R.; Byun, D. S.; Kim, H.D.; Park, Y. H. Toxicity of several puffers collected at a fish market of Pusan, Korea. Korean J. Fish. Aquat. Sci. 1994, 27(6): 682-689.
- Amano, M.; Takatani, T.; Sakayauchi, F.; Oi, R.; Sakakura, Y. The brain of the wild toxic marine pufferfishes accumulates tetrodotoxin. Toxicon 2022, 218, 1-7. [CrossRef]
- Hashiguchi, Y., Lee, J.M., Shiraishi, M., Komatsu, S., Miki, S., Shimasaki, Y.,Mochioka, N., Kusakabe, T., Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103e1118. [CrossRef]
- Nagashima, Y.; Ohta, A.; Yin, X.; Ishizaki, S.; Matsumoto, T.; Doi, H.; Ishibashi, T. Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish. Mar. Drugs 2018 Jan 8;16(1):17. [CrossRef]
- Ochoa, J. L.; Sánchez-Paz, A.; Cruz-Villacorta, A.; Nunez-Vázquez, E.; Sierra-Beltrán, A. Toxic Events in the Northwest Pacific Coastline of Mexico during 1992–1995: Origin and Impact. In Asia-Pacific Conference on Science and Management of Coastal Environment; Springer: Dordrecht, 1997; 195–200. [CrossRef]
- Shiu, Y. C.; Lu, Y. H.; Tsai, Y. H.; Chen, S. K.; Hwang, D. F. Occurrence of Tetrodotoxin in the Causative Gastropod Polinices Didyma and Another Gastropod Natica Lineata Collected from Western Taiwan. J. Food Drug Anal. 2003, 11(2), 159–163. [CrossRef]
- Quilliam, M.; Wechsler, D.; Marcus, S.; Ruck, B.; Wekell, M.; Hawryluk, T. Detection and identification of paralytic shellfish poisoning toxins in Florida pufferfish responsible for incidents of neurologic illness. Harmful algae 2002, 116-118.
- Deeds, J. R.; White, K. D.; Etheridge, S. M.; Landsberg, J. H. Concentrations of Saxitoxin and Tetrodotoxin in Three Species of Puffers from the Indian River Lagoon, Florida, the Location for Multiple Cases of Saxitoxin Puffer Poisoning from 2002 to 2004. Trans. Am. Fish. Soc. 2008, 137(5), 1317–1326. [CrossRef]
- Katikou, P. Public health risks associated with tetrodotoxin and its analogues in European waters: Recent advances after the EFSA scientific opinion. Toxins 2019, 11, 240. [CrossRef]
- Hwang, D.F.; Noguchi, T. Tetrodotoxin poisoning. Adv. Food Nutr. Res. 2007, 52, 141–236.
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [CrossRef]
- Nakamura, M.; Oshima, Y.; Yasumoto, T. Occurrence of saxitoxin in puffer fish. Toxicon 22 1984, 381–385. [CrossRef]
- Kaufmann, M.J.; Santos, F.; Maranhão, M.; Checklist of nanno- and microphytoplankton off Madeira Island (Northeast Atlantic) with some historical notes. Nova Hedwig. 2015, 101, 205–232. [CrossRef]
- Bacchiocchi, S.; Campacci, D.; Siracusa, M.; Dubbini, A.; Accoroni, S.; Romagnoli, T. et al. A Hotspot of TTX Contamination in the Adriatic Sea: Study on the Origin and Causative Factors. Mar. Drugs 2023, 21(1), 8. [CrossRef]
- Finch, S. C.; Boundy, M. J.; Harwood, D. T. The acute toxicity of tetrodotoxin and tetrodotoxin–saxitoxin mixtures to mice by various routes of administration. Toxins 2018, 10(11), 423. [CrossRef]
- Bordin, P.; Dall’Ara, S.; Tartaglione, L.; Antonelli, P.; Calfapietra, A.; Varriale, F.; Guiatti, D.; Milandri, A.; Dell’Aversano, C.; Arcangeli, G.; et al. First Occurrence of Tetrodotoxins in Bivalve Mollusks from Northern Adriatic Sea (Italy). Food Control 2021, 120, 107510. [CrossRef]
- Leão, J. M.; Lozano-Leon, A.; Giráldez, J.; Vilariño, Ó.; Gago-Martínez, A. Preliminary results on the evaluation of the occurrence of tetrodotoxin associated to marine Vibrio spp. in bivalves from the Galician Rias (Northwest of Spain). Marine Drugs 2018, 16(3), 81.
- Turner, J.W., Good, B., Cole, D., & Lipp, E. K. (2009). Plankton composition and environmental factors contribute to Vibrio seasonality. ISME Journal 2009, 3(9), 1082–1092. [CrossRef]
- Boundy, M. J.; & Harwood, D. T. Review of literature to help identify risks associated with Tetrodotoxin in seafood, including bivalve molluscs. Prepared for MPI. Cawthron Institute Report 2020, NO 2986A, 46.
- Jen, H. C.; Nguyen, A.-T.; Wu, Y J.; Hoang, T.; Arakawa,O.; Lin,W. F.; Hwang,D. F. Tetrodotoxin and paralytic shellfish poisons in gastropod species from Vietnam analyzed by high-performance liquid chromatography and liquid chromatography-tandem mass spectrometry. J. Food Drug Anal. 2014, 22(2), 178–188. [CrossRef]
Figure 1.
GPS coordinates of specimens captures with relative areas (A, B, C, D, E, F) in which captures were concentrated and fishermen.
Figure 1.
GPS coordinates of specimens captures with relative areas (A, B, C, D, E, F) in which captures were concentrated and fishermen.
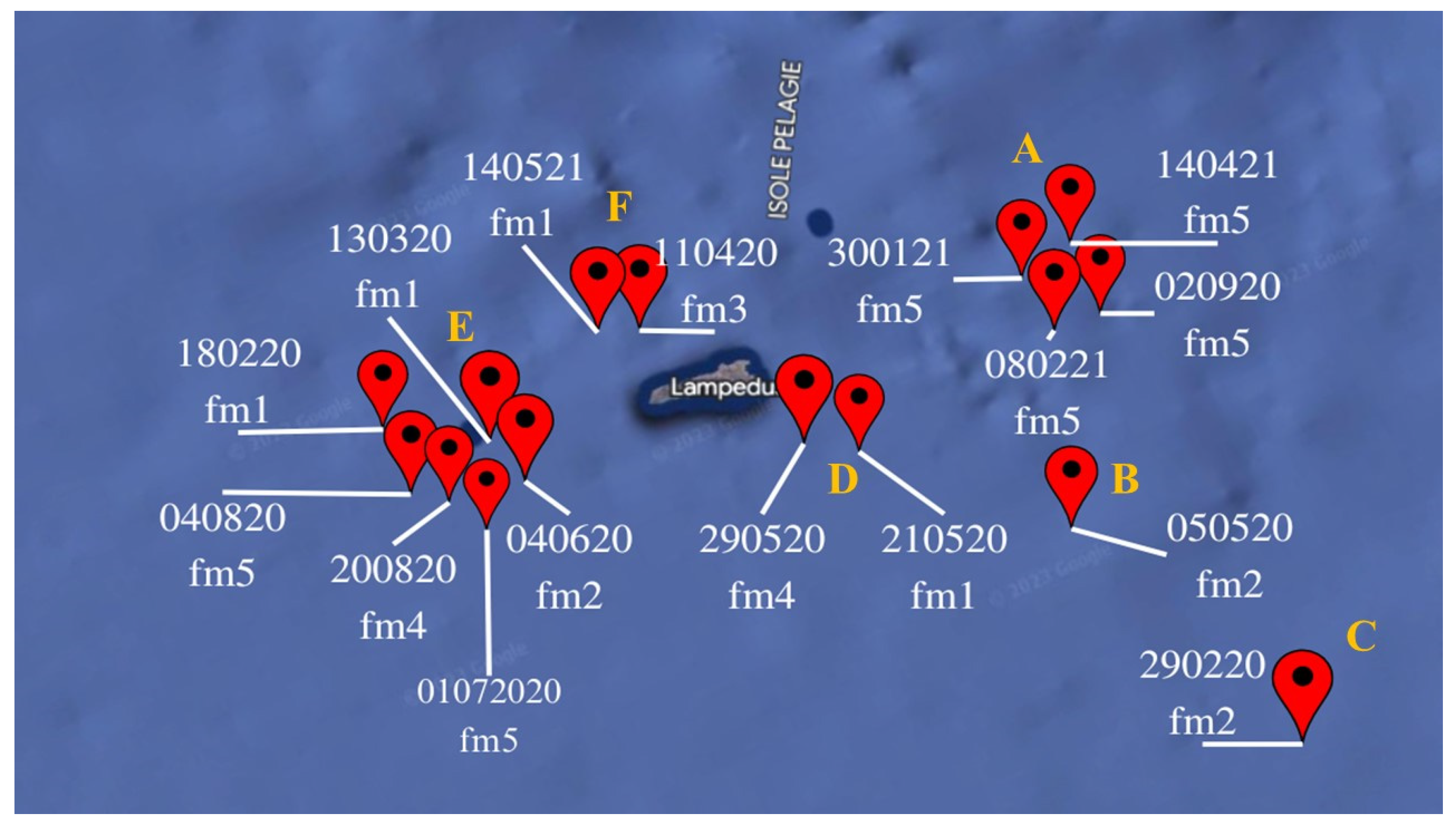

Table 1.
Available studies analysing the presence of TTXs in different pufferfish species collected in the Mediterranean Sea (only the original studies related to the listed specimens are reported). The number of the analysed specimens, the catch area and the results of TTXs analysis are reported. The catch area is categorised according Zenetos et al. [37] as follows (i) Western Mediterranean Sea (WMED); (ii) Central Mediterranean Sea (CMED); (iii) Adriatic Sea (ADRIA); and (iv) Eastern Mediterranean Sea (EMED).
Table 1.
Available studies analysing the presence of TTXs in different pufferfish species collected in the Mediterranean Sea (only the original studies related to the listed specimens are reported). The number of the analysed specimens, the catch area and the results of TTXs analysis are reported. The catch area is categorised according Zenetos et al. [37] as follows (i) Western Mediterranean Sea (WMED); (ii) Central Mediterranean Sea (CMED); (iii) Adriatic Sea (ADRIA); and (iv) Eastern Mediterranean Sea (EMED).
| Specie | N° of specimens | Catch area | TTX results | Reference |
|---|---|---|---|---|
| L. lagocephalus | 10 | WMED-Tunisia, Gabes region | positive | Saoudi et al. [19] |
| L. sceleratus | 6 | EMED-Aegean Sea | positive | Katikou et al. [20] |
| S.pachygaster | 1 | WMED-Spain | negative | Revertè et al. [21] |
| L.sceleratus | 1 | EMED-Greece | positive | |
| L sceleratus | 1 | EMED-Aegean Sea | positive | Bane et al. [22] |
| L. sceleratus | 5 | EMED- Turkey | positive | Kırımer et al. [23] |
| L. sceleratus | 16 | EMED-Mersin Bay | positive | Kosker et al. [24] |
| L. sceleratus | 20 | EMED- Turkey | positive | Acar et al. [25] |
| L. sceleratus | 20 | EMED-Egypt | positive | Alabssawy [26] |
| L. lagocephalus | 14 | WMED-Spain | negative | Rambla-Alegre et al. [27] |
| L. sceleratus | 1 | positive | ||
| S. pachygaster | 5 | negative | ||
| Torquigener flavimaculosus | 16 | EMED-Mersin Bay | positive | Kosker et al. [28] |
| L. sceleratus | 2 | EMED-Aegean Sea | positive | Leonardo et al. [29] |
| L. sceleratus | 40 | EMED-Mersin Bay | positive | Kosker et al. [30] |
| Lagocephalus spadiceus | 40 | EMED-Mersin Bay | negative | |
| Lagocephalus suezensis | 40 | EMED-Mersin Bay | positive | |
| L. sceleratus | 16 | EMED-Cyprus | positive | Akbora et al. [31] |
| L. sceleratus | 1 | ADRIA | positive | Ujević et al. [32] |
| L. sceleratus | 83 | EMED- Crete | positive | Christidis et al. [33] |
| L. sceleratus | 112 | EMED- Egypt | positive | Farrag et al. [34] |
| L. sceleratus | 3 | EMED-Lebanon | positive | Hassoun et al. [35] |
| Lagocephalus guentheri | 1 | EMED | negative | Ulman et al. [36] |
| L. suezensis | 1 | Positive | ||
| S. pachygaster | 1 | Negative |
Table 2.
New records of Sphoeroides pachygaster collected in the Mediterranean Sea with relative analysed data. The catch is categorised as in Table 1.
Table 2.
New records of Sphoeroides pachygaster collected in the Mediterranean Sea with relative analysed data. The catch is categorised as in Table 1.
| Date of catch | Catch area | N of specimens | Reference | Type of record | Fishing method | Total Lenght (mm) |
Body Weight (g) |
Depth (m) |
Type of seabed |
|---|---|---|---|---|---|---|---|---|---|
| 2012-2015 | CMED- Strait of Sicily |
7 | Cammilleri et al. [49] | Caught | Trawl | - | 130-1750 | - | - |
| 2014-2016 | WMED- Spain | 5 | Rambla- Alegre et al. [27] | Caught | - | - | - | - | - |
| 2015-2016 | CMED- Ionian Sea | 5 | Carlucci et al. [50] | Caught | Trawl | - | - | - | - |
| 2016- 2017 | EMED - Egypt | 4 | Ragheb et al. [51] | Caught | Trawl | - | - | - | - |
| NR | CMED - Malta | 9 | Vella et al. [52] | Caught | - | 266-465 | 385-2275 | - | - |
| April 2017 | EMED - Lebanon | 3 | Gerovasileiou et al. [53] | Caught | Long line | 344-417 | - | - | Soft |
| 2020 | WMED - Algerian coast | 2 | Hussein et al. [54] | Caught | Fisherman Trawl |
360.6 409.5 |
2250 2420 |
10 120 |
- |
| March 2020 |
EMED - Northen Cyprus | 1 | Akbora et al. [55] | Caught | Trammel- net | 520 | 1200 | 250 | - |
| May 2020 | EMED - Turkey | 1 | Erguden et al. [reprted in 56] | Caught | - | 490 | - | 400 | - |
| July 2020 | EMED - Turkey | 1 | Kizilkaya and Akyol [56] | Caught | Trammel-net | 257 | - | 125 | - |
| August 2021 | CMED - Strait of Messina | 1 | [57] | stranded | - | - | - | - | - |
| March 2022 | EMED - Turkey | 5 | Karakuş et al. [58] | caught | Trawl | 190-410 | 10-1405 | 140-240 | - |
| NR | EMED - Turkey | 1 | Ulman et al. [36] | caught | - | - | - | - | - |
Table 3.
Number of collected specimens divided per catch date (and relative fisherman - Fm) with relative body weight (gr) and total lenght (cm), size (S= small; L=large) and type of analysis performed (TTXs/STXs).
Table 3.
Number of collected specimens divided per catch date (and relative fisherman - Fm) with relative body weight (gr) and total lenght (cm), size (S= small; L=large) and type of analysis performed (TTXs/STXs).
| Catch date (Fishermam) |
N of specimens | Specimen code |
Body Weight (gr) | Total Lenght (cm) | Size | Toxin Analysis |
|---|---|---|---|---|---|---|
| 180220 (Fm1) |
1 | P01 | 196 | 175 | S | TTXs |
| 290220 (Fm2) |
2 | P02(A) | 199 | 220 | S | TTXs STXs |
| P02(B) | 215 | 190 | S | - | ||
| 130320 (Fm1) |
3 | P03(A) | 670 | 270 | S | TTXs |
| P03(B) | 122 | 190 | S | TTXs STXs |
||
| P03(C) | 100 | 160 | S | - | ||
| 110420 (Fm3) |
3 | P04(A) | 687 | 260 | S | TTXs STXs |
| P04(B) | 170 | 117 | S | - | ||
| P04(C) | 160 | 131 | S | - | ||
| 50520 (Fm2) |
2 | P05(A) | 743 | 260 | S | TTXs |
| P05(B) | 264 | 220 | S | TTXs STXs |
||
| 210520 (Fm1) |
3 | P06(A) | 2570 | 420 | L | TTXs |
| P06(B) | 136 | 190 | S | - | ||
| P06(C) | 202 | 210 | S | - | ||
| 210520 (Fm4) |
1 | P07(A) | 841 | 360 | S | TTXs STXs |
| 290520 (Fm2) |
2 | P08(A) | 1726 | 430 | L | TTXs |
| P08(B) | 394 | 250 | S | TTXs STXs |
||
| 40620 (Fm5) |
3 | P09(A) | 207 | 210 | S | TTXs |
| P09(B) | 179 | 190 | S | - | ||
| P09(C) | 169 | 200 | S | - | ||
| 10720 (Fm5) |
12 | P10(A) | 349 | 220 | S | TTXs STXs |
| P10(B) | 199 | 210 | S | - | ||
| P10(C) | 163 | 190 | S | - | ||
| P10(D) | 296 | 235 | S | - | ||
| P10(E) | 202 | 200 | S | - | ||
| P10(F) | 378 | 250 | S | - | ||
| P10(G) | 574 | 260 | S | - | ||
| P10(H) | 290 | 220 | S | - | ||
| P10(I) | 205 | 210 | S | - | ||
| P10(J) | 296 | 220 | S | - | ||
| P10(K) | 523 | 270 | S | - | ||
| P10(L) | 496 | 270 | S | - | ||
| 40820 (Fm4) |
3 | P11(A) | 1655 | 360 | L | TTXs |
| P11(B) | 523 | 270 | S | TTXs STXs |
||
| P11(C) | 496 | 270 | S | - | ||
| 200820 (Fm5) |
3 | P12(A) | 2145 | 445 | L | TTXs STXs |
| P12(B) | 271 | 220 | S | TTXs STXs |
||
| P12(C) | 203 | 200 | S | - | ||
| 20920 (Fm4) |
4 | P13(A) | 264 | 230 | S | - |
| P13(B) | 2280 | 450 | L | TTXs STXs |
||
| P13(C) | 1850 | 450 | L | TTXs STXs |
||
| P13(D) | 1640 | 420 | L | TTXs STXs |
||
| 300121 (Fm5) |
5 | P14(A) | ND | ND | - | - |
| P14(B) | ND | ND | - | - | ||
| P14(C) | ND | ND | - | - | ||
| P14(D) | ND | ND | - | - | ||
| P14(E) | ND | ND | - | - | ||
| 080221 (Fm5) |
5 | P15(A) | 860 | 360 | S | - |
| P15(B) | ND | ND | - | - | ||
| P15(C) | ND | ND | - | - | ||
| P15(D) | ND | ND | - | - | ||
| P15(E) | ND | ND | - | - | ||
| 140421 (Fm5) |
2 | P16(A) | ND | ND | - | - |
| P16(B) | ND | ND | - | - | ||
| 140521 (Fm1) |
2 | P17(A) | 1148 | 410 | L | - |
| P17(B) | 1407 | 430 | L | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



