
Submitted:
06 May 2023
Posted:
08 May 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Despite the fact that COVID-19 treatment and management are now considerably regulated, SARS-CoV-2 is still one of the leading causes of death in 2022. The availability of COVID-19 vaccines, FDA-approved antivirals, and monoclonal antibodies in low-income countries still poses an issue to be addressed. Natural products, particularly traditional Chinese medicines (TCMs) and medicinal plant extracts (or their active component) have challenged the dominance of drug repurposing and synthetic compound libraries in COVID-19 therapeutics. Abundant resources and excellent antiviral performance make natural products a relatively cheap and easily available alternative for COVID-19 therapeutics. Here, we deliberately review the anti-SARS-CoV-2 mechanisms of the natural products, their potency (pharmacological profiles), and application strategies for COVID-19 intervention. In light of their advantages, this review is intended to acknowledge the potential of natural products as COVID-19 therapeutic candidates.
Keywords:
COVID-19
; SARS-CoV-2
; antiviral
; therapeutics
; natural products
; traditional Chinese medicines
; plant extracts
1. Introduction
The coronavirus disease 2019 (COVID-19) pandemic represented an unprecedented disaster to the health and welfare of humanity. Since Oct 2021, the number of cases and deaths worldwide resulting from COVID-19 has increased to an unanticipated number of ≈687 million and 6.86 million, respectively[1]. The COVID-19 pandemic has had a significant socio-economic impact globally. Low-income countries, which often have weaker health systems, higher rates of poverty, and limited access to essential resources such as medical supplies and vaccines, were most affected. Many low-income countries rely heavily on tourism and exports, which were severely disrupted by the pandemic. This led to a sharp decline in revenue for these countries, making it more difficult to invest in essential services such as healthcare and education. As a result, the availability of natural antivirals can play an important role in supporting the response to COVID-19 in low-income countries, helping to make healthcare more affordable, accessible, and sustainable.
COVID-19 is caused by the novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which belongs to the genus betacoronavirus of the family Coronaviridae[2]. SARS-CoV-2 is an enveloped, positive-sense, single-stranded RNA virus. It contains a nucleocapsid (N) encapsulated genome, which is enclosed by an envelope that is associated with three structural proteins: membrane (M) protein, S protein, and envelope (E) protein[3,4,5]. Upon entering the host cell, the viral genome is released into the cytoplasm, where it is rapidly subjected to the translation by host cell ribosomes to produce sixteen individual nsps[4,6,7], accessory proteins, and four structural proteins: S, E, M, and N proteins[4,8,9]. The replication and transcription processes are carried out in virus-induced organelles, such as double-membrane vesicles (DMVs), convoluted membranes (CMs), and vesicle packets (VPs)[10]. Newly synthesized viral RNA genomes in the specialized membranous structures are transported through double-membrane-spanning pore complexes where viral assembly and budding occur in an endoplasmic-reticulum-Golgi intermediate compartment (ERGIC) in a single-membrane vesicle (SMV) lined by viral S, E, and M glycoproteins[11]. Lastly, the fusion of the mature virion-containing vesicles with host cell membranes promotes the egress of SARS-CoV-2 virions via exocytosis[11](Figure 1).
An effective immune response against COVID-19 requires both an innate immune system and an adaptive immune system. Studies have reported the role of innate immunity in determining the severity and prognosis of COVID-19. As the first line of defense against SARS-CoV-2, innate immune cells secrete interferons and other cytokines upon virus entry, thereby preventing virus entry, replication, and assembly, eliminating infected cells, and accelerating adaptive immunity[12]. SARS-CoV-2-specific CD4+ T cells, CD8+ T cells, and antibodies produced by B cells are the key components of adaptive immunity against COVID-19. Several research groups have reviewed the roles of adaptive immunity and their interplay with innate immunity in COVID-19[13,14]. Upon 5-6 days after contact, most SARS-CoV-2 infections in patients present with mild symptoms, such as fever, dry cough, tiredness, sore throat, loss of taste and smell, and skin rashes[15]. However, in some cases, the disease may progress to life-threatening pneumonia with a high risk of developing progressive respiratory failure that may lead to death, particularly in older patients or those with underlying comorbidities[16].
As a countermeasure to the continued worsening of the COVID-19 pandemic, enormous research efforts have been thrown into vaccine development, which led to the rapid production of several notable vaccines, such as BNT162b2, AZD1222, mRNA-1273, and CoronaVac[17]. However, there are major obstacles to achieving herd immunity for COVID-19, including inequity in vaccine distributions, vaccine non-responders, the emergence of immune-escape variants of SARS-CoV-2, and vaccine hesitancy among the global population. Such skepticisms revolving around vaccines were not completely unfounded, as multiple life-threatening side effects, including thrombosis[18], myocarditis[19], pericarditis[20], and even death, have been reportedly associated with vaccine use. With the uncertain long-term efficacy of currently available vaccines, there is still a need for effective treatment and preventive options for COVID-19.
Notably, due to the alarming number of infections and deaths caused by the pandemic, the Food and Drug Administration (FDA) issued an emergency use authorization (EUA) for several monoclonal antibody therapies, such as sotrovimab, tocilizumab, casirivimab, imdevimab, and baricitinib for treating or preventing COVID-19 infection[21,22]. Additionally, remdesivir represents the first direct-acting antiviral drug fully approved by the FDA for treating COVID-19. Ongoing efforts are being made through continuous collaborations between the FDA with pharmaceutical companies, researchers, and manufacturers to accelerate the development and discovery of new therapeutic drugs to prevent and treat COVID-19. Thus far, the standard treatment options for hospitalized patients with COVID-19 include the use of antivirals (remdesivir), corticosteroids (dexamethasone), monoclonal antibodies (baricitinib and tocilizumab), and oxygen supplementation care[23]. Unfortunately, most of these interventions remain largely with nations able to pay for early access due to many patent holders resisting calls for license agreements to let generic manufacturers produce similar antivirals[24]. As a result, COVID-19 has hit poor countries the hardest while exacerbating existing inequalities. As a result, effective, conveniently accessible, and economical antiviral targeting SARS-CoV-2 is mandatory, especially for low-income countries.
Nature is a great reservoir of effective compounds that can be directly used as pharmaceuticals or further developed into new drugs. Natural compounds derived from plants have been used as medicine throughout history and are considered safe. Thus, rigorous evaluation of potential natural compounds involving controlled clinical trials, in conjunction with agricultural advancement to upscale the production of effective natural antivirals, is needed. Thereby, it might lead to the availability of anti-SARS-CoV-2 drugs at a reasonable cost while serving as a source of income in low-income countries. This review describes natural products, including naturally-derived extracts (or their active components), and traditional Chinese medicines (TCMs) as potential antivirals against SARS-CoV-2, emphasizing their efficacies, mode of action, and therapeutic potentials as novel therapies against Covid-19.
2. Pre-infection inhibitors, their proposed mechanisms, and their potency against SARS-CoV-2.
Pre-infection inhibitors primarily act against viruses before the internalization phase, such as blocking the viral attachment and entry into host cells. Moreover, antivirals that target the pre-infection stage might alter the cellular endocytic mechanisms, change the environment conditions (pH) or interact with the cellular receptor and proteolytic enzymes to prevent internalization and replication of the virus [25,26]. Pre-infection inhibitors usually use as prophylactics to prevent the development of severe viral disease, reduce the risk of disease transmission, or modulate the immune system before exposure to pathogens.
Notably, there have yet to be effective FDA-approved antivirals targeting the SARS-CoV-2 pre-infection stage. Although previously established entry inhibitor hydroxychloroquine has shown promising in vitro antiviral activity, it has not shown any meaningful in vivo benefits in both animal models and humans[27,28,29,30,31,32,33,34,35,36,37,38,39]. Furthermore, higher and longer doses were difficult to achieve due to drug-related severe adverse effects of these drugs[27,29,30,31,32]. Moreover, umifenovir, an antiviral proposed to inhibit SARS-CoV-2 viral entry, has shown some clinical efficacy but is of uncertain clinical importance due to a lack of high-quality evidence and contradicting findings from different clinical studies. Mounting evidence suggested that various natural compounds could serve as pre-infection inhibitors and potentially use as prophylaxis for COVID-19[40]. Table 1 lists potential natural compounds, their structure, potency (based on IC50), toxicity (CC50), and mechanism of action. Besides, Figure 2 shows a schematic diagram of the potential natural compounds targeting at different stages of the SARS-CoV-2 life cycle.
Epigallocatechin gallate (EGCG) found in tea extracts was reported to bind to the RBD of extracellular SARS-CoV-2 directly, thus preventing the interaction of the virus with host receptor ACE2 and subsequent viral entry[41]. In further corroboration of the findings above, EGCG in green tea displayed a dose-dependent anti-entry effect against pseudotyped lentiviral vectors carrying SARS-CoV-2 S protein with an IC50 value of 2.47 μg/mL. In live viruses, EGCG could inhibit the viral plaque formation of SARS-CoV-2, MERS-CoV, and SARS-CoV. Using the SARS-CoV-2 vitronectin ELISA kit, researchers further reported that EGCG could simultaneously bind to the RBD of SARS-CoV-2 and compete with ACE2 binding, efficiently blocking the recruitment of these viruses to host cell surfaces[42].
When highly sulfated polysaccharides were screened using SPR, RPI-27, RPI-28, fucoidans, and heparin could bind to the S-protein of SARS-CoV-2 [43]. Among all the polysaccharides tested, RPI-27, a high molecular weight branched polysaccharide, was identified as the most potent antiviral against SARS-CoV-2 with IC50 of 83 nM, more effective than remdesivir. Other sulfated polysaccharides such as heparin, TriS-heparin, and non-anticoagulant low molecular weight heparin (NACH) also showed good antiviral activity against SARS-CoV-2. In silico study demonstrated that heparin could bind to the S-protein RBD (PDB ID: 6VW1), suggesting that sulfated polysaccharides might target the entry stage of SARS-CoV-2[43]. Another study also reported that marine sulfated polysaccharides extracted from Stichopus japonicus, brown and red algae, showed dose-dependent antiviral activities against SARS-CoV-2. Among these polysaccharides, sea cucumber sulfated polysaccharide (SCSP) extracted from Stichopus japonicus was identified as the most potent polysaccharide with IC50 of 9.10 μg/mL. SCSP could bind to the S protein carried by pseudotyped virus and prevented its entry into the host cells[44].
Various bioactive compounds have been identified in Vitis vinifera leaf extract, and most of them were derivatives of quercetin, luteolin, kaempferol, apigenin, isorhamnetin, myricetin, chrysoeriol, biochanin, isookanin, and scutellarein[45]. Vitis Vinifera leaf extract was reported to have a virucidal effect against enveloped DNA virus, HSV-1, and RNA virus such as SARS-CoV-2. Cotreatment of virus and this extract to the Vero cells showed potent viral inhibition, suggesting that it might block virus attachment and thus prevent entry [45]. Temperature shift assays further confirmed this projection and gene expression study demonstrated that Vitis Vinifera leaf extract strongly reduced the S protein expression in SARS-CoV-2 infected cells[45].
HIDROX containing 40% of hydroxytyrosol (HT), is a commercial product produced by the olive oil industry and is used as a natural product to improve health. The anti-SARS-CoV-2 activity of HIDROX and HT was reported to be virucidal against the extracellular virus in a time and dose-dependent manner [46]. Besides, by comparing the efficiency of their virucidal effects, HIDROX was found to be more potent than pure HT. In addition, both HICROX and HT were suggested to alter the structure of the SARS-CoV-2 S protein, which was reflected in the molecular weight increase of S1, S2, and RBD subunits when evaluated in western blotting. This may cause the S proteins to aggregate and lead to the virucidal effects observed in cells. On the other hand, HIDROX-containing cream also had virucidal activity against SARS-CoV-2, indicating that HT-rich olive extract could be applied to the skin for protective purposes to reduce transmission of COVID-19[46].
Water extract of Prunella vilgaris (NhPV) was reported to inhibit the SARS-CoV-2 S protein-mediated virus entry [47]. It was observed that NhPV could inhibit the entry of SARS-CoV-2 pseudotyped HIV that carries full-length S protein (IC50= 30 μg/mL) or mutated S protein, D614, that enhanced virus infectivity. Time-of-drug addition assays revealed that NhPV treatment worked best when cells were pre-treated with NhPV 1 hour before viral infection. ELISA assay further confirms that NhPV could bind to ACE2 and block the interaction of viral S protein to the ACE2 receptor, thus preventing viral entry. A combination of NhPV and anti-SARS-CoV-2 neutralizing antibody (SAD-S35) enhanced the blocking effect, so similar efficiencies could be achieved by reducing the compound concentrations. Moreover, 50 μg/mL of NhPV could prevent the CPE induced by wild-type SARS-CoV-2. By titrating the production of viral progeny, researchers also reported that 100 μg/mL of NhPV could completely prevent virus infection[47].
The mixture of Agrimonia Pilosa (AP) and Galla rhois (RG) in a ratio of 6:4 (APRG64) was reported to inhibit the formation of SARS-CoV-2 plaques by blocking SARS-CoV-2 entry into host cells and their antiviral effects were comparable to remdesivir and chloroquine phosphate [48]. Ursolic acid, quercetin, and 1,2,3,4,6-penta-O-galloyl-β-d-glucose were identified as active compounds responsible for the anti-SARS-CoV-2 activity of APRG64. They significantly reduced the S protein in cell supernatants and could inhibit SARS-CoV-2 propagation. Molecular docking study showed that ursolic acid exhibited the highest binding affinity towards SARS-CoV-2 S RBD and the B.1.1.7 lineage S RBD with a binding energy of −9.5 kcal/mol and −9.0 kcal/mol, respectively[48].
Echinaforce, an alcoholic extract of Echinacea purpurea plants, has been previously evaluated for its benefit in treating the common cold and other respiratory tract infections[49,50,51]. Echinaforce was observed to have irreversible virucidal activity against HCoV-229E in viral infected pseudostratified respiratory epithelial cell culture, which better mimics the in vivo lung model. Moreover, Echinaforce also exhibited virucidal activity against MERS-CoV, SARS-CoV, and SARS-CoV-2, with complete inactivation after 50 μg/ml Echinaforce treatment[52].

Table 1.
Examples of natural compounds against SARS-CoV-2. (Compound structures are drawn using ChemDraw Professional 16.0).
Table 1.
Examples of natural compounds against SARS-CoV-2. (Compound structures are drawn using ChemDraw Professional 16.0).
| Natural compounds | Compound structure | genotype/strain | IC50 | CC50 | Stages of inhibition | Suggested mechanism | Reference | |
|---|---|---|---|---|---|---|---|---|
| Traditional Chinese Medicine (TCM) | ||||||||
| Shuanghuanglian preparations | NA | 2019-nCoV | 0.064-0.090 µL/mLA1 0.010 mg/mLA2 3.65-4.44 µL/mLA3 0.14 mg/mLA4 0.93-1.20 μL/mLB |
>12.5 μL/mL | Polypeptide processing and replication | Inhibited catalytic activity 3CLpro Inhibited polymerization of RdRp. |
[53,54] | |
| Baicalein | 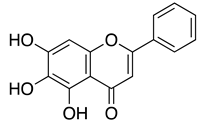 |
0.94 μMA 2.94 μMB |
>200 μM | |||||
| Baicalin | 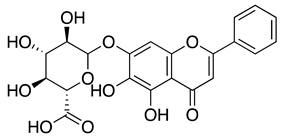 |
6.41µMA 27.87 µMB |
>200 μM | |||||
| Liu Shen | NA | MT123290.1 | 0.6024 μg/mLC | 4.930 μg/mL | Post-infection | Targeted both the virus and host factor. Down regulate NF-κB/MAPK signaling pathway and reduce the production of proinflammatory cytokines. | [55] | |
| Lianhuaqingwen (LHQW) | NA | Genebank accession no. MT123290.1 | 411.2 μg/mLC | 1157 μg/mL | Immune modulation Virucidal |
Reduced production of proinflammatory cytokines such as TNF-α, IL-6, CCL-2/MCP-1, and CXCL-10/IP-10. Altered the morphology of extracellular virus. |
[56] | |
| Jinhua Qinggan (JHQG) | NA | NA | NA | NA | Immune modulation | Reduced production of IL-6 and increased the production of IFN-γ | [57] | |
| Xuebijing (XBJ) | NA | NA | NA | NA | Immune modulation | NA | [58] | |
| NRICM101 | NA | TCDC#4 from Taiwan CDC | 0.22 mg/mLA 0.41 mg/mLG 0.28 mg/mLH 0.42-1.18 mg/mLI |
1.77 mg/mL | Pre- and post-infection Immune modulation |
Blocked the binding of S protein to ACE2 Inhibited 3CL protease Reduced production of IL-6 and increased the production of TNF-α |
[59] | |
| Mentha haplocalyx extract | NA | hCoV-19/Taiwan/4/2020 | NA | NA | Prophylactic effect | NA | [60] | |
| Natural extracts and its active ingredients | ||||||||
| Punicalagin (PUG) from Pomegranate peel extract (PPE) | 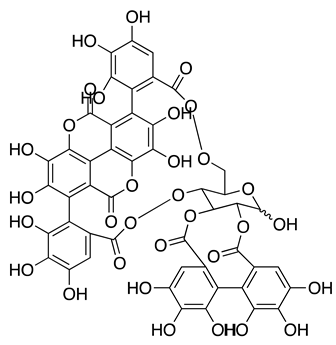 |
Isolate USA-WA1/2020 | 7.20 µMD 4.62 µMA |
100 µM | Polypeptide processing | Acted as allosteric Inhibitor and inhibited catalytic activity 3CLpro. | [61] | |
| Artemisia annua L. extracts | NA | USA/ WA12020 | 0.01-0.14 μg/mLC | >500 µg/mL | Replication | NA | [62] | |
| EGYVIR | NA | hCoV-19/Egypt/NRC-03/2020 | 0.57 μg/mLD | NA | Virucidal effect | Inactivated extracellular SARS-CoV-2. Modulated the immune system by inhibiting nuclear translocation of p50 and down-regulating Ikβα, TNF-α and IL-6, thus preventing cytokine storm. |
[63] | |
| Netrium oleander extract and oleandrin | 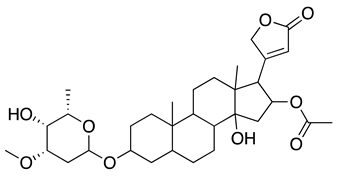 |
USA-WA1/ 2020 strain | 7.07-11.98 ng/mlD | > 1 µg/ml | Prophylactic effect | NA | [64] | |
| Vitis Vinifera leaf Extract | NA | clinical isolate from Lazzaro Spallanzani Hospital | 5-10 μg/mLD | >500 μg/mL | Attachment and entry | NA | [45] | |
| Andrographolide from Andrographis paniculata extract | 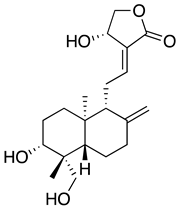 |
SARS-CoV2/ 01/ human/ Jan2020/ Thailand | 0.034 μMD 5-15.05 μMA |
13.19-81.52 μM | Polypeptide processing | Covalently linked to the active site of Mpro. | [65,66,67] | |
| Hydroxytyrosol-Rich Olive Pulp Extract (HIDROX) | NA | JPN/TY/ WK-521 strain | NA | NA | Virucidal effect | Interacted, changed the structure, and aggregated the S protein. | [46] | |
| APRG64, mixture of Agrimonia pilosa (AP) and Galla rhois (RG) | NA | NCCP43326 | NA | NA | Entry | Active ingredients such as ursolic acid, quercetin, and 1,2,3,4,6-penta-O-galloyl-β-d-glucose interacted with RBD of S protein. | [48] | |
| epigallocatechin gallate (EGCG) from green tea extract | 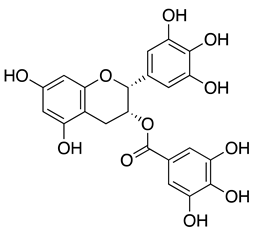 |
MUC-IMB-1 | 2.47 μg/mLE | >20 μg/mL | Entry | Bound to RBD of S protein | [42] | |
| Propolis extract | NA | NA | NA | NA | Entry Replication |
NA | [68,69] | |
| Echinaforce | NA | BetaCoV/ France/ IDF0372/ 2020 | NA | >100 µg/ml | Virucidal effect | NA | [52] | |
| Prunella vilgaris (NhPV) extract | NA | hCoV- 19/Canada/ON-VIDO-01/2020, GISAID accession# EPI_ISL_425177 | 30 μg/mLE | >200 μg/mL | Entry | Bound to ACE2 receptor and prevent viral entry | [47] | |
| Pure natural compounds isolated from natural origin | ||||||||
| Isorhamnetin | 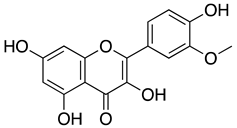 |
SARS-CoV-2 pseudovirus | NA | NA | Entry and polypeptide processing | Interacted with ACE2, S protein and inhibited catalytic activity of Mpro. | [70,71,72] | |
| Bufalin | 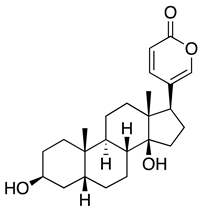 |
WIV-04 | 0.018 μMC | >2000 μM | Replication | Targeted the ion exchange function of Na+/K+-ATPase | [73] | |
| Naringenin | 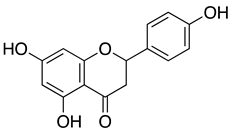 |
hCoV-19/ Egypt/ NRC-03/ 2020 (Accession Number on GSAID: EPI_ISL_430820) | 28.35 µg/mLC 92 nMA |
178.75 µg/mL | Polypeptide processing | Inhibited catalytic activity Mpro. | [74] | |
| Resveratrol | 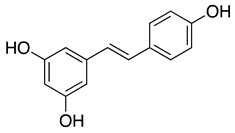 |
NL/2020 (EVAg-010V-03903) | 66 µMD | NA | Post-entry | NA | [75] | |
| Pterostilbene | 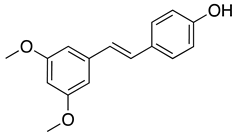 |
19 µMD | ||||||
| Sulfated polysaccharide (RPI-27) | NA | NA | 83 nMD | >500 µg/mL | Entry | Interacted with RBD on S protein. | [43] | |
| Crude polysaccharide 375 | NA | WIV04 | 0.48 µMA 27 nMB |
136 mg/Kg on mice | Polypeptide processing | Inhibited catalytic activity Mpro. | [76] | |
| Sea cucumber sulphated polysaccharide | NA | NA | 9.10 μg/mLD | NA | Entry | Interacted with S protein. | [44] | |
| Sinapic acid |  |
NA | 2.69 µg/mLC | 189.3 µg/mL | Entry Assembly and release | Interacted with E protein | [77] | |
| X-206 | NA | NA | 14 nMC | 8.2 µM | Pre- and post-infection stage | NA | [78] | |
| Niclosamide | 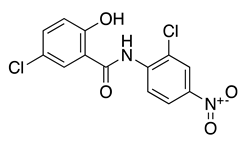 |
England/ IC19/ 2020 (IC19) | 0.34 µMD | NA | Replication | Blocked intracellular calcium release and prevented syncytia formation | [79] | |
| lycorine | 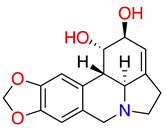 |
WIV04 | 0.31 µMB | > 40μM | Post-infection | NA | [80] | |
| Tocopherol derivative, TPGS | NA | Wa-1/USA | 212 nMA | NA | Transcription | Interacted with RdRp and inhibited viral transcription and replication | [81] | |
IC50= the inhibitory concentration of natural compounds required to cause 50% inhibition of virus. CC50= the cytotoxic concentration of natural compounds required to cause 50% cell death. 3CLpro= 3C-like protease; S= S protein; Na+/K+-ATPase= Sodium–potassium adenosine triphosphatase; RdRp= RNA-dependent RNA polymerase; Mpro= Main protease; RBS= Receptor-binding domain; E= Envelope protein; TNF-α= Tumor necrosis factor alpha; IL-6= Interleukin 6; NF-κB/MAPK = Nuclear factor kappa B/ mitogen-activated protein kinase; TPGS= D-α-tocopherol polyethylene glycol succinate. A IC50 value measured in terms of viral protein inhibition. B IC50 value measured in terms of reduction in SARS-CoV-2 RNA copy number, measured using qRT-PCR. C IC50 value measured in terms of cytopathic effects induced by SARS-CoV-2. D IC50 value measured in terms of reduction in SARS-CoV-2 plaque or foci number. E IC50 value measured in terms of inhibition of entry of psuedotyped virus. F IC50 value measured in terms of cell-cell fusion. G IC50 value measured in terms of binding of viral protein to host cell receptor. H IC50 value measured in terms of viral protein expression. I IC50 value measured in terms of cytokine reduction. 1 oral liquid form of Shuanghuanglian preparations against 3CLpro. 2 lyophilized powder of Shuanghuanglian preparations against 3CLpro. 3 oral liquid form of Shuanghuanglian preparations against PLpro. 4 lyophilized powder of Shuanghuanglian preparations against PLpro.
3. Post-infection inhibitors, their proposed mechanisms and their potency against SARS-CoV-2
Post-infection inhibitors are antivirals that work after a person has been infected with a virus. An effective post-infection drug has the potential to reduce the severity and duration of symptoms and may reduce the risk of complications from the infection. Post-infection antiviral treatment is provided by the physician specifically at the virus causing the infection after diagnosis. Significant portion of low-income countries’ populations have been infected with SARS-CoV-2, thus effective antivirals for COVID-19 targeting the post infection stage are important to prevent the further spread of COVID-19 and to mitigate the impact of the disease on the poor.
Shuanghuanglian preparations are TCM extracted from Lonicera japonica, Scutellaria baicalensis Georgi, and Forsythia suspense are widely used to treat infections of the respiratory tract[82,83]. Several independent studies revealed that Shuanghuanglian preparations and their active components, baicalin, and baicalein, exhibited potent antiviral activity against SARS-CoV-2 by inhibiting the activity of its 3CLpro and PLpro and its RdRp[53,54]. Due to the high amount of baicalin being present within Shuanghuanglian preparations and almost similar IC50 of Shuanghuanglian preparations (1.83 to 6.14 μM) with pure baicalin (6.41 μM), it was believed that baicalin is the main component that led to the proteolytic inhibition of SARS-CoV-2 PLpro. The protein-ligand interactions were further evaluated in isothermal titration calorimetry (ITC) measurements as well as electrospray ionization mass spectrometry (ESI-MS), and it was observed that baicalin and baicalein both had a high binding affinity with SARS-CoV-2 PLpro with Kd of 11.50 and 4.03 μM in ITC and 12.73 and 1.40 µM in ESI-MS, respectively. The crystal structure of SARS-CoV-2 PLpro complexed with baicalein was further investigated using X-ray crystallography. Results showed that baicalein acted as a shield, interacting with S1/S2 subsites and the oxyanion loop, thus preventing the substrate from binding with the catalytic site of SARS-CoV-2 PLpro[53]. Besides, baicalein and baicalin were reported to be potent inhibitors of SARS-CoV-2 RdRp. When the SARS-CoV-2 polymerase complex was expressed and purified, the ability of the polymerase to extend the RNA template (14-Mer RNA) in the presence of baicalein and baicalin was investigated. As expected, extended RNA products were reduced significantly in the presence of non-toxic doses of baicalein and baicalin. Molecular docking analysis also confirmed that baicalein and baicalin strongly bind to the RNA polymerase of SARS-CoV-2 with a binding energy of −8.7 and−7.8 kcal/mol, respectively. Therefore, unlike any nucleoside analogs, baicalein and baicalin do not pose a potential risk of host mutational activity, as they directly bind and inhibit SARS-CoV-2 RdRp transcriptional activity. Notably, a pharmacokinetic study in rhesus monkeys revealed that despite the low oral bioavailability of baicalein, this compound was rapidly converted to the 7-O-βglucopyranuronoside metabolite baicalin. Both compounds demonstrated relatively long half-lives as well as Cmax values (2.3 μM and 30.7 μM, respectively) that were higher than the IC50 values (1.2 μM and 6.2 μM, respectively) reported in Calu-3 human lung cells[54], suggesting that less frequent dosage may be needed to maintain therapeutic plasma concentration[84].
A natural extract of A. paniculate and its main bioactive compound, andrographolide were reported to reduce the viral progeny production in plaque assays (IC50 = 0.036 μg/mL and 0.034 μM, respectively) and in high-content imaging using an anti-SARS-CoV-2 antibody. The IC50 value of A. paniculate extract and andrographolide using foci reduction is lower than remdesivir (IC50 = 0.086 μg/mL), indicating that they are promising treatment to being use against COVID-19[67]. Furthermore, in silico molecular docking study suggested that andrographolide might target the main protease of SARS-CoV-2, Mpro, and absorption, distribution, metabolism, and excretion (ADME) profile prediction showed good pharmacodynamic property[65]. To further evaluate the interaction of andrographolide and Mpro, researchers expressed and purified 2019-nCoV Mpro and SARS-CoV Mpro and reported that andrographolide could form covalent linkages and inhibited proteolytic activities of Mpro with IC50 of 15.05 and 5.0 μM, respectively. In addition, andrographolide could dock into the catalytic pockets of Mpro with binding affinity from -7.85—9.72 kcal/mol[66].
Naringenin is a flavone found in many citrus plants and has various biological functions such as antiviral, antibacterial, and anticancer[85]. Recently, naringenin was identified as a potent antiviral agent against SARS-CoV-2 MPro (PDB entry: 6w63) from HTVS of 8793 natural compounds. Naringenin and other lead compounds selected from HTVS were reported to inhibit SARS-CoV-2 MPro in vitro using enzyme inhibition assays (>75% inhibition) at 100 µM. Based on the cell-based antiviral assays, only naringenin showed moderate anti-SARS-CoV-2 activity with IC50 of 28.347 μg/mL and SI of 6.3. Structure-activity relationship of naringenin and its closely related derivative, eriodyctiol, revealed that naringenin (7-OH position) forms H-bonds with Gln189 and Thr190 residues as well as the interaction with His41 residues might explain its potent antiviral activity in targeting SARS-CoV-2 Mpro in vitro. The molecular properties of naringenin also agreed with Lipinski’s rule. They showed a good score for topological polar surface area (TPSA) value of <140 Å, indicating that naringenin might have good oral bioavailability, intestinal permeability, and absorption[74]. Parallel with these findings, in silico study using molecular docking simulations, molecular mechanics Poisson–Boltzmann surface area (MM-PBSA), density functional theory (DFT), and binding energy analysis supported the interaction of naringenin at the active site of SARS-CoV-2 Mpro[86].
Recently, resversatrol and pterostilbene were reported to inhibit SARS-CoV-2 progeny production in Vero cells with an IC50 of 66 and 19 μM, respectively. These two compounds were also reported to have long-lasting antiviral effects against SARS-CoV-2. They maintained their antiviral activation in cell cultures up to 40 hpi, equivalent to approximately 5 rounds of viral replications. Time-of-drug-addition assays demonstrated that these compounds solely targeted the viral replication stage but not the entry stage, nor did they have virucidal activity. Besides that, the post-infection antiviral effect of RES was also observed in primary human bronchial epithelial cells cultured under air-liquid interphase (ALI) conditions[75]. Another study also showed evidence that RES could inhibit SARS-CoV-2 (IC50 =10.66 μM) and HCoV-229E (IC50 =4.6 μM) at the post-infection stage with reduced cytotoxicity (CC50=210 μM)[87]. Cotreatment with silent information regulator T1 (SIRT1) signaling antagonist, sirtinol, and RES attenuated the inhibitory effect of RES. It promoted the replication of the virus, suggesting that RES might inhibit SARS-CoV-2 infection via the SIRT1 pathway[88].
4. Multi-stage inhibitors, their proposed mechanisms, and their potency against SARS-CoV-2.
Antiviral compounds that can inhibit early and late stage of virus infection are preferable as they have the potential to target multiple stages of the virus life cycle, thus preventing viral adaptation and the emergence of drug-resistant viruses. SARS-CoV-2 is highly mutagenic due to the inherent error-prone nature of RNA replication, and many novel variants of SARS-CoV-2 have emerged. These new variants might confer resistance to antivirals that target a single stage of the virus life cycle. Therefore, the search of SARS-CoV-2 inhibitors that target both pre- and post-infection stages is crucial to reduce the likelihood of drug resistance.
Chingguan Yihau (NRICM101) is a TCM formulated in Taiwan and was used in clinical settings for COVID-19 patients [59]. Patients were administered 100 mL 3 times per day at 30 mins after meal. Clinical data showed that patients with the underlying disease who showed no improvement after 21 days of hospitalizations had benefited from NRICM101 treatment without adverse effects [59]. In vitro findings from surface plasmon resonance (SPR) analysis indicated that NRICM101 could bind to RBD protein dose-dependently. At the same time, ELISA showed that NRICM101 inhibited S protein from binding to the ACE2 receptor with an IC50 value of 0.41 mg/mL. Besides, NRICM101 could also inhibit the 3CLpro enzymatic activity with an IC50 value of 0.22 mg/mL. In addition, NRICM101 treatment also reduced the growth of SARS-CoV-2 in terms of viral protein expression (IC50= 0.28 mg/mL) and plaque formation. Besides its promising antiviral activity, NRICM101 could also reduce the production of pro-inflammatory cytokines, IL-6, and TNF-α in lipopolysaccharide (LPS)-stimulated alveolar macrophages with an IC50 value of 0.42 and 1.18 mg/mL, respectively[59]. When antiviral studies were performed on the individual herb of NRICM101, researchers reported that the Scutellaria baicalensis component was responsible for its effective anti-3CLpro activity (100%). Other components, such as Schizonepeta tenuifolia, Morus alba, Magnolia officinalis, and Mata and Mentha haplocalyx in NRICM101, effectively blocked the binding of S protein to ACE2 (>70%). In addition, Scutellaria baicalensis and Houttuynia cordata with 20-fold dilution could inhibit IL-6 and TNF-α production, and Scutellaria baicalensis was able to inhibit cytokine production at 40-fold dilution[59]. Another component of NRICM101, Mentha haplocalyx, the natural source of Chinese peppermint, has gained some attention as an antiviral agent against SARS-CoV-2. It was used as traditional medicine for minor ailments[89]. The essential oil extracted from peppermint leaves could improve muscle pain, and itching or be used as a fragrance[90]. Using a cell-based HTS assay to screen 190 traditional herbal medicines, researchers identify Mentha haplocalyx extract as an effective anti-SARS-CoV-2 compound in vitro and in vivo. Along with Mentha haplocalyx, Ganoderma lucidum extracts could also reduce the CPE of SARS-CoV-2 in Vero cells at 960-fold dilution. In the hamster model, oral administration of M. haplocalyx extracts at 200 mg/kg/day for three consecutive days after viral infection showed promising antiviral activity and significantly reduced lung viral titres[60].
Isorhamnetin is a flavonoid extracted from Sea buckthorn or Hippophae rhamnoides L. The extract from berries of this plant was reported to have anticancer, antiviral, anti-diabetic, and immune regulatory activities[91,92,93,94]. In one of the studies, the active compound of Sea buckthorn, isorhamnetin, was tested against the SARS-CoV-2 S pseudotyped virus in vitro. It was observed that isorhamnetin could block SARS-CoV-2 S pseudotyped virus entry into the cells expressing ACE by 47.7% at a non-toxic concentration of 50 μM. Quercetin, another component of Sea buckthorn, was similarly evaluated, but there was no significant reduction of viral entry. Using SPR analysis, isorhamnetin showed affinity towards immobilized ACE recombinant protein, and molecular docking analysis further proved that isorhamnetin could bind to ACE2 at K353, E37, and H34 residues[70]. In addition, high throughput virtual screening (HTVS) of naturally occurring phytochemicals also showed that along with other flavonoids, isorhamnetin could interact with the S2 domain of the SARS-CoV-2 S protein with the binding energy of -8.3 Kcal/mol[72]. On the other hand, various derivatives of isorhamnetin from Salvadora persica were reported to have a strong binding affinity towards SARS-CoV-2 Mpro with a binding orientation similar to the positive control N3, which binds to the Cys–His catalytic dyad located between domains I and II of SARS-CoV-2 Mpro. Structure-activity relationship analysis revealed the presence of disaccharide rutinose (α-L-rhamnopyranosyl-(1-6)-β-D-glucopyranose) at position carbon no. 3 of flavonoids helped to enhance the binding stability in the N3 binding site of SARS-CoV-2 Mpro[71]. Since these studies showed that isorhamnetin could interact with host protein (ACE2 receptor), viral structural protein (S protein), and a viral non-structural protein (SARS-CoV-2 Mpro), isorhamnetin should be considered for further study to develop it into a potent antiviral drug that could target multiple stages of viral life-cycle, thus preventing the replication of SARS-CoV-2.
Ionophore antibiotics are a family of natural compounds produced by microorganisms, which are well known for their antibacterial activity against gram-positive bacteria[95,96]. In addition, the antiviral activity of ionophore antibiotics against HIV, influenza virus, ZIKV, MERS-CoV, and SARS-CoV was also reported[97,98,99]. Recently, researchers screened 11 different naturally occurring polyether ionophores for their potential to prevent the CPE caused by SARS-CoV-2 in Vero E6 cells that overexpressed TMPRSS2. Ionophore antibiotics such as narasin, salinomycin, and nanchangmycin exhibited >100-fold selectivity. Surprisingly, 2 compounds: maduramycin and X-206 showed selective index of 313 and 586, respectively, which were higher than the remdesivir control (SI=>67). Compound X-206 displayed significant antiviral activity against SARS-CoV-2 by reducing CPE (IC50 = 14 nM), viral RNA copy number, viral S proteins, and viral plaque formation by SARS-CoV-2. Time-of-addition assays revealed that X-206 achieved approximately 2-fold log reduction of viral progeny at all time points from 4 hours pre-infection to 8 hours post-infection. Based on morphological profiling, which provided bioactivity fingerprints, X-206 was found to have a different mechanism of action compared to the control lysosomotropic hydroxychloroquine (HCQ). However, the molecular mechanism of this compound has not been elucidated, and further experiments should be conducted to discover the mode of action of X-206[78]. The ionophore antibiotics, salinomycin, and niclosamide, were observed to inhibit syncytia formation which indicates caused by the fusion of cells induced by viral infection. It was observed that niclosamide and salinomycin exhibited cell protection effects against SARS-CoV-2 with IC50 of 0.34 and 0.22 μM, respectively. Besides, salinomycin and niclosamide also inhibited viral replications in respiratory Calu-3 cells. Furthermore, a low concentration of 1 μM of niclosamide blocked the transmembrane member 16 (TMEM16) chloride channel. It significantly attenuated the effect of spontaneous calcium transients in the presence of the S protein, suggesting that ionophore antibiotics might block intracellular calcium release to prevent syncytia formation[79].
Oleandrin is one of the bioactive compounds found in Nerium oleander extract and was identified as a unique lipid-soluble cardiac glycoside that acted on the Na/K ATPases pump to enhance heart contraction in heart failure patients. It was also reported to be used in treating dermatological diseases and cancers[100,101,102]. Recently, oleandrin was reported to be a potent inhibitor of SARS-CoV-2. When oleandrin was present before (prophylactic) and during the whole viral life cycle, it could completely reduce the plaque formation of SARS-CoV-2 by 4 log10, with IC50 values of 11.98 and 7.07 ng/mL for 24 and 48 hpi, respectively. Additionally, when oleandrin was added 24 hours post-infection, where extensive viral replication had already occurred, oleandrin could still maintain its therapeutic effect. In the hamster model, oleander extract did not cause any toxicity effects regarding body weight, organ lesions, alkaline phosphatase (ALP), and alanine aminotransferase (ALT) levels. In vivo, the prophylactic efficacy of oleander extract against SARS-CoV-2 showed a significant reduction in viral loads in nasal turbinates after 3 days post-infection. From day 3 onwards, the viral titre was below the detection limit, and after four days of treatment, the virus was completely cleared[64]. Besides SARS-CoV-2, oleandrin was also reported to be a potent inhibitor of HCoV-OC43 when CPE (IC50 = 26 mM) and viral titre (2-3 log10 reduction) were measured as the end point[103].
5. Natural products with immunomodulatory effects.
Attenuating immunopathogenesis of COVID-19, such as cytokine storm and excessive immune cell infiltration and suppressing viral replications, are considered important hallmarks in halting the progression to severe disease and reducing mortality. Corticosteroid usage in severe COVID-19 patients was shown to be beneficial in reducing mortality and attenuating excessive inflammation[104,105,106]. Still, they carry several disadvantages, including immunosuppression[107], increased risk of secondary bacterial infections[105], increased risk of developing systemic complications[108], and prolonged viremia[105,109]. Additionally, early corticosteroid treatment was associated with an increased risk of death in COVID-19 patients, particularly those below 60 years old who were not presenting inflammation upon admission[104].
EGYVIR extract containing Curcuma longa (Turmeric) root and Piper nigrum seed extracts has demonstrated immunomodulatory properties and potent antiviral activity against SARS-CoV-2. EGYVIR reduced the viral plaque formation by up to 92% with an IC50 of 0.57 μg/mL. When treatment was performed 24 hours post-infection, EGYVIR extract remained effective in inhibiting the virus infectivity by 78%, which was higher than the control, hydroxychloroquine (66%). Additionally, the extract was found to downregulate the expression of inflammatory markers such as Ikβα, TNF-α, and IL-6 in SARS-CoV-2 infected Huh7 cells, suggesting that it might alleviate the cytokine storm induced by SARS-CoV-2 infection[63]. Similarly, TCMs such as Lianhuaqingwen (LHQW)[56], Liu Shen (LS)[55], and Chingguan Yihau[59] have been shown to suppress SARS-CoV-2 replication and excessive cytokine production in vitro. LHQW has been shown to inhibit SARS-CoV-2 replication with an IC50 of 411.2 μg/mL. Under TEM, LHQW-treated cells (48 hours) showed a reduction in the number of virions in the cytoplasm, endoplasmic reticulum, cell membrane, and intracellular vesicles. LHQW treatment was also observed to alter SARS-CoV-2 morphology. Moreover, LHQW inhibited SARS-CoV-2-induced cytokine production in vitro, as shown by significantly reduced mRNA expression levels of tumor necrosis factor-alpha (TNF-α), interleukin 6 (IL-6), C-C motif chemokine ligand 2 (CCL-2)/monocyte chemoattractant protein 1 (MCP-1), and C-X-C motif chemokine ligand 10 (CXCL-10)/interferon gamma-induced protein 10 (IP-10)[56]. In a prospective, multicenter, open-label, randomized controlled trial of 284 hospitalized COVID-19 patients, treatment with LHQW improved the recovery rate of chest computed tomography (CT) manifestations, shortened the median time to the symptom (fever, cough, and fatigue) recovery, and enhanced the rate of symptom recovery[110]. Two meta-analyses revealed that integration of LHQW with western medicines compared with western medicine alone also led to significantly improved CT imaging, reduced rate of severe disease progression, and improved recovery and disappearance rate of clinical symptoms such as fever, cough, and fatigue[111,112]. More recently, the China FDA has approved using LHQW as a therapeutic agent against COVID-19 due to ample clinical findings substantiating its immunoregulatory effect in COVID-19 patients. Another TCM, Liu Shen (LS), besides its direct antiviral effect (IC50 = 0.06024 μg/mL), could down-regulated the nuclear factor kappa-light-chain-enhancer of activated B cell/ mitogen-activated protein kinase (NF-κB/MAPK) signaling pathway, which was reflected in the decrease in expression of p65, phospho (p)-nuclear factor of kappa light polypeptide gene enhancer in B-cell inhibitor alpha (IκBα), and p-p38 MAPK. Downregulation of NF-κB/MAPK signaling pathway was postulated to decrease pro-inflammatory cytokines such as IL-6, TNF-α, IL-1β, CXCL10/IP-10, CCL2/MCP-1, and IL-8, suggesting that it may prevent cytokine storm in COVID-19[55]. Another TCM, Chingguan Yihau, formulated in Taiwan, was used in clinical settings for COVID-19 patients. Clinical data showed that patients with underlying diseases who showed no improvement after 21 days of hospitalization benefited from treatment without adverse effects. The mechanism of action of Chingguan Yihau was reported to be through its ability to reduce the production of proinflammatory cytokines, IL-6, and TNF-α, with an IC50 value of 0.42 and 1.18 mg/mL, respectively, in lipopolysaccharide (LPS)-stimulated alveolar macrophages [59]. Network pharmacology analyses revealed that LHQW[113,114], Jinhua Qinggan[115], and Xuebijing injection (XBJ)[116] could potentially treat COVID-19 by modulating various genes and signaling pathways involved in anti-inflammatory, antiviral, and immune responses. Additionally, molecular docking analyses of the bioactive constituents of these TCMs were shown to bind with various viral and host proteins, including Mpro, 3CLpro, S protein, and ACE2[113,114,115,116,117], which, together with the previous findings, suggest a multitude of mechanisms of TCMs in ameliorating COVID-19 infection and immunopathogenesis.
Some COVID-19 patients were also reported to have gastrointestinal microbiota dysregulation. Over time, probiotics have been reported to improve general health by preventing gastrointestinal dysbiosis and helping with gastrointestinal, oral, and vaginal infections[118,119]. Moreover, the immunomodulatory effects of probiotics have gained attention concerning their anti-allergy and antiviral activities[120]. Antigens from the probiotics such as lipoteichoic acids (LTA), lipopolysaccharides (LPS), and bacterial nucleic acid, could bind to specific host cell receptors such as Toll-like receptor (TLR) and give rise to immunomodulatory effects[121,122]. Moreover, probiotics may be more accessible and affordable than synthetic drugs for COVID-19 in low-income countries for several reasons. Firstly, probiotics are generally less expensive than synthetic drugs, making them more affordable for people living in low-income countries. Secondly, probiotics are widely available in many regions, including low-income countries. This means that people living in areas with limited access to synthetic drugs may be able to use probiotics as an alternative treatment option. Lastly, probiotics are generally regarded as safe. They have fewer side effects than synthetic drugs, which can be particularly important in areas where access to medical care may be limited[123]. Various probiotics, especially lactic acid bacteria (LAB), have been reported to modulate the immune system and prevent viral infection. For example, live and heat-killed Enterococcus faecalis KH2 could protect animals against influenza virus and EV-71 by modulating monocyte chemoattractant protein-1(MCP-1), a key chemokine that regulates migration and infiltration of immune cells to the site of infection. Pretreatment of E. faecalis reduced the MCP-1 production, thus preventing migration, infiltration, and differentiation of monocyte, preventing inflammatory disorder, reducing lung viral titer, and enhancing the survival rate of influenza-infected mice. Also, pretreatment of E. faecalis reduced the MCP-1 and protected mice against EV71-induced neurotoxicity and growth retardation in EV-71-infected mice[124]. LAB is also well known for altering immune cells' functional role, especially dendric cells. For example, Bifidobacteria strains in clinically active probiotic combination (VSL#3) could stimulate intestinal and blood DC to upregulate IL-10 production and downregulate the expression level of costimulatory molecules CD80 and CD40, thus preventing the generation of pro-inflammatory Th1 cells and decreasing IFNγ production by T cells[125]. Besides this, LC-plasma could also stimulate pDC to produce type 1 IFN via TLR9 and subsequently induce the upregulation of ISGs in HepG2 cells, thus inhibiting the replication of DENV[126,127]. Based on these studies, probiotics or LAB might have a promising advantage in reducing the COVID-19 severity and speed up the viral clearance. In an open-label propensity score matched retrospective cohort study, probiotic Bifidobacterium, Lactobacillus, and Enterococcus could reduce time to clinical improvement, days of hospitalization, and time to viral RNA clearance of COVID-19[128]. Lactiplantibacillus plantarum is a probiotic that was reported to modulate cytokine production by increasing TNF-α, IL-1β, IL-18, IL-8, and reducing IL-6, a critical factor in immune dysregulation in COVID-19, thus enhancing the activity of natural killer cells[129]. In a quadruple-blinded, randomized, placebo-controlled trial, Lactiplantibacillus plantarum (KABP022, KABP023, KAPB033) and Pediococcus acidilactici (KABP021) could shorten the disease recovery period and symptom duration[130]. Recently, Bacillus subtilis natto, a probiotic used to ferment soy food, was reported to stimulate human M1-phenotype macrophages by upregulating immune activation markers, CD80, and enhanced the gene expression of interferon stimulative genes (IFNβ1, IFNλ1, ISG20, and RNase L), thus was suggested to be used as antiviral against SARS-CoV-2[131]. A double-blinded, randomized, placebo-controlled trial was performed with three strains of bacillus, Bacillus coagulans LBSC (DSM 17654), Bacillus subtilis PLSSC (ATCC SD 7280), and Bacillus clausii 088 A E (MCC 0538) concerning COVID-19 symptoms. Results from this trial showed that the mental and physical fatigue score of the treatment group were significantly improved (91%)[132]. Based on the evidence mentioned above and the absence of conclusive treatments for COVID-19, probiotics supplementation is highly anticipated by the public, especially the socioeconomically challenged population, because probiotics are generally recognized as safe (GRAS), low in cost and easily found in daily diet. However, further preclinical and clinical studies should be explored to discover the role of probiotics in COVID-19. Many clinical trials investigating the benefits of probiotics for COVID-19 are ongoing.
6. Application strategies
6.1. Combinational therapy of natural compounds.
Researchers have shown interest in exploring the potential of natural products as anti-SARS-CoV-2 agents. However, there is a lack of research on their combined effectiveness with other antiviral agents. Combination therapy can help target multiple viral life cycles to promote effective antiviral effects, which far outweigh simple monotherapy. Combination therapy which involved two or more therapeutic agents, has been a cornerstone of antiviral therapy. For example, combination treatment involving at least three different antiretroviral drugs (two nucleoside reverse transcriptase inhibitors and one either an integrase strand transfer inhibitor (INSTI), a non-nucleoside reverse transcriptase inhibitor, or a protease inhibitor) is recommended for standard treatment of HIV[133]. Such a combination therapy could inhibit HIV replication, allowing the immune system to overcome the infection and prevent AIDS. Besides that, the repurposing drugs, sofosbuvir, and daclatasvir, when combined, could synergistically inhibit SARS-CoV-2 replication by targeting different processes during viral RNA synthesis, and this combination has shown promising efficacy in several clinical studies[134,135,136]. So, it may be an interesting avenue worth exploring, considering the promising preliminary evidence suggesting the potential benefit of natural products for COVID-19. It is plausible that the combinatorial use of various natural products or natural products with other anti-SARS-CoV-2 therapies (monoclonal antibodies, small chemical molecules, antiviral peptides, and repurposing drugs) could lead to better clinical outcomes in severe COVID-19 and achieving better survivability, which warrants further investigations. For example, a combination of NhPV/Suramin has been shown to synergistically inhibit SARS-CoV-2 viral entry when used with a SARS-CoV-2 neutralizing antibody (SAD-S35)[47]. Intriguingly, some of the aforementioned natural products have reported various drug-drug interactions, which, used in combination, might lead to summation (additivity), enhancement (synergism), or initiation (potentiation) of antiviral effect against COVID-19. For example, pomegranate extract has been studied for its potential use with other antiviral compounds. A study has shown that a pomegranate rind extract (PRE) and punicalagin, when co-administered with zinc (II) ion, were more effective at inhibiting the replication of the acyclovir-resistant HSV than either compound alone[137]. Besides, PUG has also been shown to have synergistic antiviral effects with oseltamivir, an antiviral drug that the human influenza A virus has conferred resistance to[138]. Besides, the impact of PUG combined with zinc sulfate monohydrate on SARS-CoV-2 3CLpro was reported, and the coefficient of drug association (CDI) was 0.272, indicating a synergistic effect of combining these two compounds[139]. Another compound, quercetin, present in APRG64, LHQW, XBJ, and vitis vinifera leaf extract, was identified as an anti-SARS-CoV-2 compound and was reported to have a synergistic antiviral effect when combined with vitamin C. The synergistic effect of quercetin and vitamin C was believed to be attributed to the potent antiviral effect of quercetin, the immunomodulatory effects of vitamin C, and the capacity of vitamin C to recycle quercetin[140]. Natural products, such as natural extracts or TCMs, are considered combination therapy. They are made up of diverse bioactive constituents and allow synergistic interactions between each ingredient and exert multiple mechanisms, such as immunomodulatory, anti-inflammatory, and antiviral responses. For example, when the active compound of Aetemisia annua L. extract, artemisinin, was tested as a pure drug, it was less effective (IC50= 70 μM) than the Aetemisia annua L. cultivar extracts (IC50=0.01 to 0.14 μg/mL), suggesting that artemisinin might not be the sole active ingredient contributing to the anti-SARS-CoV-2 activity of this extract[62]. For example, a mixture of polysaccharide 375 isolated from the seaweed Ecklonia kurome Okam, was more effective in inhibiting the enzymatic activity of SARS-CoV-2 Mpro when compared to purified homogenous polysaccharides 37501 and 37503. This finding suggested that the cocktail-like polysaccharide synergistically inhibited SARS-CoV-2, supporting that it might be a potential antiviral drug for COVID-19[76]. Since low-income countries face several challenges in combating COVID-19, a combination of antiviral therapy that is constituted by natural products can offer several advantages. First, combination therapy using natural compounds might improve treatment outcomes by reducing the risk of treatment failure. By ensuring the effectivity and sustainability of antiviral treatments, it would potentially reduce the need for future hospitalizations and other medical interventions. Second, combining multiple drugs in a single regimen might reduce the risk of drug resistance, and thus reducing the cost to develop new antiviral drugs. This can make it more feasible for low-income countries to provide treatment to a larger portion of the population. When used in conjunction with other public health measures such as vaccination and infection control, over time, combination therapy can help to reduce the burden of viral infections on low-income countries. However, further in-depth studies to evaluate the efficacy and safety of combined natural products against COVID-19 should be prioritized in the current global health pandemic.
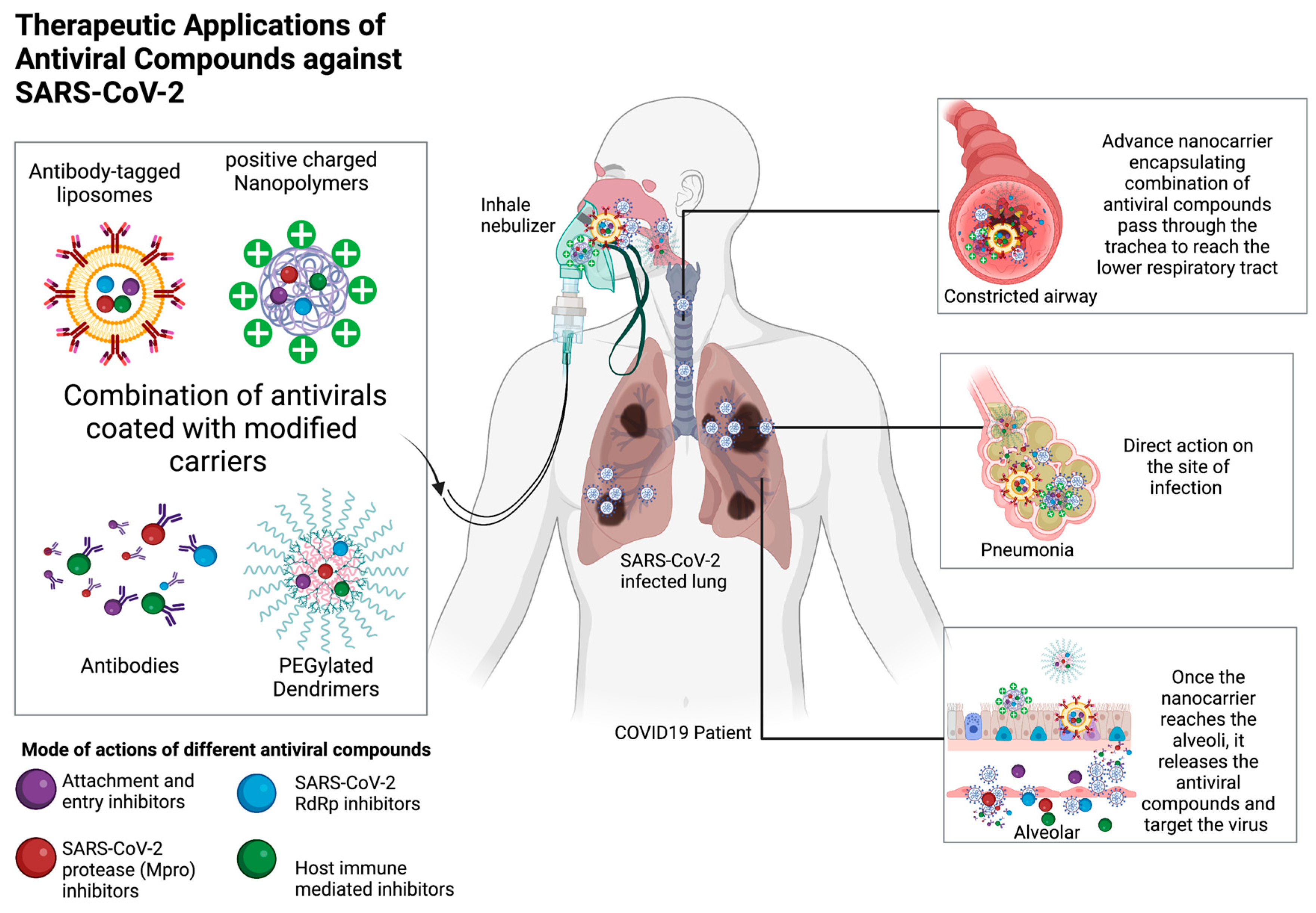
Figure 3.
Therapeutic applications of antiviral compounds against SARS-CoV-2. Different types of carriers such as liposomes, polymers, dendrimers, and antibodies can deliver antiviral compounds via the inhaling nebulizer route, concentrating the antiviral compounds in the lung where SARS-CoV-2 mostly reside. Modifications such as antibody tagging, polyethylene glycol PEGylation, and surface charge alteration can advance the pharmacodynamics of these antiviral compounds. Various antiviral compounds with different modes of action can be simultaneously encapsulated in a carrier to reduce toxicity and prevent the development of antiviral-resistant mutants. (Figure drawn by biorender).
Figure 3.
Therapeutic applications of antiviral compounds against SARS-CoV-2. Different types of carriers such as liposomes, polymers, dendrimers, and antibodies can deliver antiviral compounds via the inhaling nebulizer route, concentrating the antiviral compounds in the lung where SARS-CoV-2 mostly reside. Modifications such as antibody tagging, polyethylene glycol PEGylation, and surface charge alteration can advance the pharmacodynamics of these antiviral compounds. Various antiviral compounds with different modes of action can be simultaneously encapsulated in a carrier to reduce toxicity and prevent the development of antiviral-resistant mutants. (Figure drawn by biorender).
6.2. Potential delivery methods of natural compounds.
To reduce the viral load in COVID-19 patients, it is crucial to inhibit viral replication and virus production by host cells. Although various antiviral compounds may be active in the free form, their ability to concentrate at the sites of infection may be limited, requiring the administration of a high dose with accompanying adverse effects. Thus, an effective delivery system using nanocarriers such as dendrimers, liposomes, and polymers to transport antiviral drugs against SARS-CoV-2 may be an added advantage[141]. Antiviral natural compounds can be entrapped or coupled with these nanocarriers to enhance their solubility, stability, and bioavailability[141]. For example, polyamidoamine (PAMAM) dendrimer conjugation could enhance the inhibitory effect of sialyl lactose against a wide range of human and avian influenza virus[142]. In addition, PAMAM dendrimer is effective in pulmonary delivery of the poorly soluble anti-asthma drug, beclometasone dipropionate. Since the main SARS-CoV-2 tropism occurred in the respiratory system, aerosol-based PAMAM dendrimer might be an excellent delivery system for antiviral natural compounds against COVID-19. Liposomes consisting of mainly amphoteric phospholipids bilayer could effectively enhance the delivery of water-soluble and lipid-soluble antiviral compounds, thus enhancing the bioavailability of the compound[143]. Long-circulating nanoliposome (LCNs) containing baicalein (BAI) have been proven to yield greater LCNs-BAI oral bioavailability compared to free BAI by 4.52 times in Kunming mice[144]. Since baicalein was reported to be an effective antiviral against various viruses including SARS-CoV-2, this study provides a beneficial and promising therapeutic platform for baicalein for COVID-19 when incorporated with liposome technology. In another study, liposome formulation could enhance the antiviral activity of propolis in inhibiting the 3CL protease (IC50 = 1.183 ± 0.06) compared to free-form propolis (IC50 = 2.452 ± 0.11) and significantly reduced the viral replication of SARS-CoV-2 with an inhibitory effect comparable to remdersivir[145]. On the other hand, liposome was also suggested to be an effective delivery system for TCM to improve targetability and reduce side effects[146]. A recent study has shown that tripterin, a TCM, was successfully encapsulated into liposomes and significantly reduced the pulmonary pathological alterations and viral burden of severe COVID-19 human ACE2-transgenic (hACE2) mice model[147]. Next, polymeric nanoparticles such as chitosan are also valuable for developing nanoformulations for COVID-19. Studies have suggested that amphiphilic positively charged chitosan might electrostatically bind to the virus and prevent viral entry into the host cells[148]. Besides having potent anti-SARS-CoV-2 activity, N-palmitoyl-N-monomethyl-N,N-dimethyl-N,N,N-trimethyl-6-O-glycolchitosan (GCPQ) has been reported to reside (13.1%) in mouse nares up to 24 hours after nasal dosing, suggesting that it could use as a prophylactic nasal spray to prevent COVID-19. Since chitosan has good antiviral activity, it may be used to encapsulate antiviral drugs to achieve combinational effects, thus developing a comprehensive antiviral therapy for COVID-19. Various studies have proven that using chitosan as an effective delivery system positively affects the physiological function of natural compounds. For example, punicalagin-loaded alginate/chitosan-gallol hydrogels enhanced tissue adhesiveness and promoted wound repair and hemostasis[149]. Recently, baicalein-encapsulated chitosan has been successfully developed and has achieved high loading (74.2%) and encapsulation (96.1%) capacity. The delivery system was also reported to provide a more controlled and constant effect on tissue targets without adverse effects [150]. Although the antiviral effect of baicalein-loaded chitosan has not been tested, it has been reported that such a delivery system could effectively control immune-allergic-inflammation in asthmatic mice[150]. On the other hand, EGCG-loaded silver nanoparticles (EGCG-AgNPs), combined with zinc sulfate, could exert a triple effect on multiple steps of the infectious cycle of H5N1[151]. This strongly demonstrated that co-administering zinc sulfate with a natural compound and using silver nanoparticles as a delivery system might be a potential platform for developing SARS-CoV-2 inhibitors, which can prevent microbial resistance by restricting adaptation. Furthermore, advanced modifications of these nanocarrier-coated antiviral drugs are feasible to reduce the potential adverse effect or elevate the draggability of the antiviral compounds. For example, surface modification of cationic dendrimers to more negatively charged is directly linked to reduced cytotoxicity and may be used to transport antiviral compounds against SARS-CoV-2[152]. Moreover, coupling polyethylene glycosylated (PEG) to nanocarrier surfaces could prevent the binding of plasma proteins to the nanocarriers, avoid degradation by the immune system[153], enhance the half-life, and extend the circulation time of these antiviral compounds[154]. For instance, PEGylated PAMAM dendrimer has been reported to improve the drug loading capacity of the anti-HIV drug, Lamivudine and reduce the high surface toxicity of PAMAM while helping to control the release of antiviral drugs in a prolonged manner[155]. Also, antiviral compound-containing nanocarriers may be coated with SARS-CoV-2-antibody to direct them to the infection site, thus effectively targeting infected cells. This approach has been constructive in anticancer treatment when antibody-modified PAR28-PEG-liposome effectively delivered the drug into the targeting cells[156]. Overall, a potent antiviral delivery system may promote drug disposition at the sites of infection, enhance intracellular drug delivery, prevent drug inactivation, and prevent toxicity caused by overdosing. As a result, incorporating nanotechnology with natural compounds should be the prospective research topic for COVID-19 drug discovery.
6.3. Potential route of administration
Promising natural compounds have been proposed to inhibit SARS-CoV-2, but the draggability of these antiviral compounds remains the most challenging task. An effective antiviral should achieve a concentration level within the therapeutic range at the infection site to produce the desired clinical outcome. As a result, the administration route is one way to ensure adequate drug concentration at the targeted organ while avoiding toxicity (overdose) and ineffective treatment (under-dose). Although intravenous administration of antiviral drugs is frequently applied, it has various drawbacks. For example, it requires trained personnel, specialized equipment, and consumables, and it causes mild discomfort to the patient. Due to the intravenous use of remdesivir in COVID-19 patients, it is limited to hospital settings. Poor levels of health service coverage, low levels of government funding, and lack of political support are some of the causes of inadequate response to injecting antiviral drugs in low-income countries[157]. Recently, inhalation treatment using an inhaler portable device was suggested for COVID-19 [158]as it is convenient and easily accessible, which could provide early doses of antiviral, thus curtailing the development and transmission of COVID-19. Since the SARS-CoV-2 infection is spread through airborne virions, the ciliated nasopharyngeal epithelium and oral cavity were believed to be the primary site of infection [159]. Studies have reported that the ACE2 receptor is highly expressed in the nasal cavity compared to peripheral lung tissue[159], supporting that the nasal cavity is the primary portal for the virus to internalize into the human body[160,161]. As a result, the application of an antiviral nasal or oral spray might reduce infectious virus load and the risk of transmission. Recent research has demonstrated that nasal sprays could inhibit several respiratory viruses replication[162]. For example, AM-301 or Bentrio, a nasal spray containing bentonite (magnesium aluminum silicate), was demonstrated to reduce the viral load in human airway epithelial cells without causing toxicity to the nasal epithelium[163]. Next, povidone-iodine (PVP-I) nasal antiseptic preparation was reported to have strong virucidal activity against extracellular SARS-CoV-2[164]. The efficiency of PVP-I nasal sprays in reducing SARS-CoV-2 nasopharyngeal titer in the clinical setting was also evaluated (NCT04347954). Moreover, when the cocktail of F61 and H121, neutralizing antibodies were administered via the intranasal route, it significantly improved the survival rate and reduced the lung virus of delta SARS-CoV-2 (B.1.617.2) challenged K18-hACE2 mice[165]. Thus, nasal drops or sprays might be an excellent route for administering prophylactic natural compounds. The nasal spray of grapefruit seed extract, which contains bioactive metabolites such as limonin, naringenin, and hesperidin, has been commercialized and was suggested by physicians to be used for COVID-19 in case series[166]. In addition, Product I, an oral spray containing a mixture of natural essential oils (Anise oil, eucalyptus oil, levomenthol, myrrh extract, clove oil, peppermint oil ratanhia root extract, tormentil root extract), showed potent virucidal activity and reduced the virus load to lower limit of detection (≥4.69 log10 TCID50/mL), compared to other chemical sprays[167]. The repurposed herbal spray, Allergic Rhinitis Nose Drops (ARND), contains mainly Herba Centipedae, mint, and Scutellaria baicalensis have been reported to inhibit pseudovirus infection, modulate the immune system and prevent binding between the spike protein (Delta) and ACE2 of SARS-CoV-2[168]. These preliminary data of commercialized natural nasal or oral sprays provide the basis for further clinical investigations and future development of the antiviral hygienic product for COVID-19. Although nasal sprays are an effective tool to protect and prevent initial contact with SARS-CoV-2, they might not be effective when SARS-CoV-2 spreads further to the lower respiratory tract. In this respect, an oral inhalation nebulizer is an alternative tool to concentrate the antiviral drugs in the lung and prevent pneumonia, acute respiratory distress syndrome (ARDS), sepsis, and even superinfection[169,170]. An inhalation nebulizer is suitable for antiviral compounds with low oral bioavailability and a shorter half-life[171]. Inhalation of antiviral compounds into the lung is readily absorbed and distributed systemically compared to oral dosing, as the latter depends on gastrointestinal tract (GIT) absorption and systemic metabolism in GIT and liver[172,173,174]. For instance, when budesonide drug with high first-pass metabolism and low GIT absorption properties was administered through inhalation, it reduced the likelihood of intensive care and the recovery period in COVID-19 patients[175]. On the other hand, a natural essential oil blend made up of lemon oil, oregano oil, tea tree oil, java citronella oil, turmeric oil, peppermint oil, lavender, ginger, frankincense, eucalyptus, wheat germ, basil, cedarwood, holy basil, cinnamon oil, sage oil, and clove oil (registered batch no. 250320211) was reported to be safe and could be used to relieve respiratory symptoms of mild COVID-19 patients, when administered via nebulization[176]. Unfortunately, using nebulizers is concerning as the aerosol particles generated by nebulizers are small (1–5 μm) and can carry contaminants such as viruses and bacteria into the deep lung, thus increasing the risk of COVID-19 transmission[177]. As a result, care should be taken when using an antiviral nebulizer[178].
7. Conclusion
The SARS-CoV-2 pandemic represents an unprecedented crisis to healthcare facilities and socioeconomics worldwide. The urgent need for effective treatment for COVID-19 has led to the extensive repurposing of drugs as a convenient strategy to identify antiviral agents as a rapid treatment response to this global crisis. However, despite many drugs showing promising preclinical efficacy against SARS-CoV-2, most have failed to translate into clinical benefits in humans. Although remdesivir has been FDA-approved for treating COVID-19, it does not cure or prevent the spread of COVID-19, given its modest overall efficacy. Also, the use of many antiviral drugs, including remdesivir, is only limited to hospital use, and low-income countries commonly do not have the privilege to afford such treatment as they are facing challenges such as lack of healthcare providers, personal protection equipment, healthcare system, financial leverage and more. As a result, the search for new effective, widely accessible, affordable, and broad-spectrum antiviral drugs against SARS-CoV-2 is still warranted. Much work has been dedicated to using in silico, in vitro, ex vivo, and in vivo methods to predict, evaluate, test, and develop efficient antiviral treatments for SARS-CoV-2. In silico study uses computer simulations and modeling software to predict the compounds’ behavior in biological systems. Useful molecular docking software such as AutoDock Vina, UCSF DOCK6, Glide, and Surflex-Dock allow researchers to perform high-throughput screening to determine the binding affinity of potential compounds to SARS-CoV-2 proteins. However, in silico software rely on assumptions and simplifications that may not accurately reflect the complex biological systems. For example, the proteins may be modeled as a rigid structure rather than flexible and dynamic molecules, or the ligand affinity towards the proteins was assumed based on empirical data, known as force fields. Although in silico studies provide valuable information on the potential of the antivirals, it is important to validate the predictions made by in silico models using in vitro and in vivo studies. While in vitro studies have proven useful for selecting antivirals, it is important to acknowledge their limitation in mimicking the physiological environment. Some in vitro systems, such as organoids or microfluidic chips, can better mimic the complex cellular and physiological interactions in living organisms. Also, since the in vitro findings in this review were collected from different labs, the results should be interpreted with caution, and the potency of different natural antivirals may not be directly compared but only use as a reference because the experimental conditions, settings, and techniques used may differ. Taken together, nature is an excellent reservoir of potent antiviral compounds that can be directly used or modified into new drugs. This review introduced some promising natural products, including TCM, natural extracts (or their active compound), and probiotics all with reported activity against SARS-CoV-2, and further discussed their efficacy and mechanism. Although most of the natural products listed in this study are potential inhibitors of SARS-CoV-2, the results are primarily derived from in vitro research. Thus, future work should critically evaluate the bioavailability, suitable dosage, mode of delivery, therapeutic window, and combinatorial potential of these natural products in animal models before advocating their use as proof-of-concept antiviral in human COVID-19 clinical trials. Additionally, future efforts should focus on improving the draggability and efficacy of potential and existing antiviral drugs for COVID-19 treatment. Such efforts might include developing aerosol-based antivirals to be administered via inhalation, allowing rapid accumulation in the lungs where SARS-CoV-2 viruses mainly infect, or utilizing a nano-carrier delivery system to enhance targeted drug delivery and bioavailability. In summary, while natural products for COVID-19 have shown promise, further research is needed to fully understand its efficacy and safety, particularly in different populations and in conjunction with new treatments.
Author Contributions
Conceptualization, Z.L. and R.L; validation, Z.L. and R.L.; formal analysis, Z.L.; investigation, Z.L and R.L.; resources, P.H. E.T. and SAB; data curation, Z.L.; writing—original draft preparation, Z.L.; writing—review and editing, R.L.; visualization, R.L.; supervision, P.H., E.T. and SAB.; project administration, P.H.; funding acquisition, P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Higher Education for niche research under the Higher Institution Centre of Excellence (HICoE) program, Project Code: MO002-2019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data availability statement: Not applicable.
Acknowledgments
We acknowledge the funding from the Ministry of Higher Education, Malaysia for niche area research under the Higher Institution Centre of Excellence (HICoE) program (Project MO002-2019). We would like to acknowledge the staff of Tropical Infectious Disease Research and Education Center (TIDREC) and the staff of the Department of Medical Microbiology, Faculty of Medicine, Universiti Malaya for their unwavering support throughout this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Worldometer Coronavirus. Available online: https://www.worldometers.info/coronavirus/ (accessed on 17 September 2021).
- Pal, M.; Berhanu, G.; Desalegn, C.; Kandi, V. Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2): An Update. Cureus 2020, 12, e7423. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.; Benvenuto, D.; Giovanetti, M.; Angeletti, S.; Ciccozzi, M.; Pascarella, S. Sars-CoV-2 Envelope and Membrane Proteins: Structural Differences Linked to Virus Characteristics? Biomed Res Int 2020, 2020, 4389089. [Google Scholar] [CrossRef] [PubMed]
- Mariano, G.; Farthing, R.J.; Lale-Farjat, S.L.M.; Bergeron, J.R.C. Structural Characterization of SARS-CoV-2: Where We Are, and Where We Need to Be. Front Mol Biosci 2020, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, C.; Xu, X.; Xu, W.; Liu, S. Structural and Functional Properties of SARS-CoV-2 Spike Protein: Potential Antivirus Drug Development for COVID-19. Acta Pharmacol Sin 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Moustaqil, M.; Ollivier, E.; Chiu, H.-P.; Tol, S. van; Rudolffi-Soto, P.; Stevens, C.; Bhumkar, A.; Hunter, D.J.B.; Freiberg, A.N.; Jacques, D.; et al. SARS-CoV-2 Proteases PLpro and 3CLpro Cleave IRF3 and Critical Modulators of Inflammatory Pathways (NLRP12 and TAB1): Implications for Disease Presentation across Species. Emerg Microbes Infect 2021, 10, 178–195. [Google Scholar] [CrossRef]
- Yan, S.; Wu, G. Potential 3-Chymotrypsin-like Cysteine Protease Cleavage Sites in the Coronavirus Polyproteins Pp1a and Pp1ab and Their Possible Relevance to COVID-19 Vaccine and Drug Development. The FASEB Journal 2021, 35, e21573. [Google Scholar] [CrossRef]
- Michel, C.J.; Mayer, C.; Poch, O.; Thompson, J.D. Characterization of Accessory Genes in Coronavirus Genomes. Virol J 2020, 17, 1–13. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V. v.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 Protein Interaction Map Reveals Targets for Drug Repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Knoops, K.; Kikkert, M.; Worm, S.H.E. van den; Zevenhoven-Dobbe, J.C.; Meer, Y. van der; Koster, A.J.; Mommaas, A.M.; Snijder, E.J. SARS-Coronavirus Replication Is Supported by a Reticulovesicular Network of Modified Endoplasmic Reticulum. PLoS Biol 2008, 6, e226. [Google Scholar] [CrossRef]
- Mendonça, L.; Howe, A.; Gilchrist, J.B.; Sheng, Y.; Sun, D.; Knight, M.L.; Zanetti-Domingues, L.C.; Bateman, B.; Krebs, A.-S.; Chen, L.; et al. Correlative Multi-Scale Cryo-Imaging Unveils SARS-CoV-2 Assembly and Egress. Nat Commun 2021, 12, 4629. [Google Scholar] [CrossRef]
- Diamond, M.S.; Kanneganti, T.D. Innate Immunity: The First Line of Defense against SARS-CoV-2. Nature Immunology 2022 23:2 2022, 23, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Primorac, D.; Vrdoljak, K.; Brlek, P.; Pavelić, E.; Molnar, V.; Matišić, V.; Erceg Ivkošić, I.; Parčina, M. Adaptive Immune Responses and Immunity to SARS-CoV-2. Front Immunol 2022, 13, 2035. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Crotty, S. Adaptive Immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef] [PubMed]
- Sheikhi, K.; Shirzadfar, H.; Sheikhi, M. A Review on Novel Coronavirus (Covid-19): Symptoms, Transmission and Diagnosis Tests. Research in Infectious Diseases and Tropical Medicine 2020, 2, 1–8. [Google Scholar]
- An, P.; Song, P.; Lian, K.; Wang, Y. CT Manifestations of Novel Coronavirus Pneumonia: A Case Report. Balkan Med J 2020, 37, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Sharma, O.; Sultan, A.A.; Ding, H.; Triggle, C.R. A Review of the Progress and Challenges of Developing a Vaccine for COVID-19. Front Immunol 2020, 11, 585354. [Google Scholar] [CrossRef] [PubMed]
- Smadja, D.M.; Yue, Q.-Y.; Chocron, R.; Sanchez, O.; Lillo-Le Louet, A. Vaccination against COVID-19: Insight from Arterial and Venous Thrombosis Occurrence Using Data from VigiBase. European Respiratory Journal 2021, 2100956. [Google Scholar] [CrossRef]
- Albert, E.; Aurigemma, G.; Saucedo, J.; Gerson, D.S. Myocarditis Following COVID-19 Vaccination. Radiol Case Rep 2021, 16, 2142–2145. [Google Scholar] [CrossRef]
- McLean, K.; Johnson, T.J. Myopericarditis in a Previously Healthy Adolescent Male Following COVID-19 Vaccination: A Case Report. Academic Emergency Medicine 2021, 8, 918–921. [Google Scholar] [CrossRef]
- Ison, M.G.; Wolfe, C.; Boucher, H.W. Emergency Use Authorization of Remdesivir: The Need for a Transparent Distribution Process. JAMA - Journal of the American Medical Association 2020, 323, 2365–2366. [Google Scholar] [CrossRef]
- Chen, P.; Nirula, A.; Heller, B.; Gottlieb, R.L.; Boscia, J.; Morris, J.; Huhn, G.; Cardona, J.; Mocherla, B.; Stosor, V.; et al. SARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19. New England Journal of Medicine 2021, 384, 229–237. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Treatment Guidelines Hospitalized Adults: Therapeutic Management. Available online: https://www.covid19treatmentguidelines.nih.gov/management/clinical-management/hospitalized-adults--therapeutic-management/ (accessed on 20 October 2021).
- Usher, A.D. The Global COVID-19 Treatment Divide. Lancet 2022, 399, 779. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.; Helenius, A. Virus Entry: Open Sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef]
- Martín, C.S.S.; Liu, C.Y.; Kielian, M. Dealing with Low PH: Entry and Exit of Alphaviruses and Flaviviruses. Trends Microbiol 2009, 17, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Goyal, A.; IV, A.C.; Lahan, S.; Dhaliwal, H.S.; Bhyan, P.; Bhattad, P.B.; Aslam, F.; Ranka, S.; Dalia, T.; et al. Hydroxychloroquine: A Comprehensive Review and Its Controversial Role in Coronavirus Disease 2019. Ann Med 2020, 53, 117–134. [Google Scholar] [CrossRef]
- Misra, D.P.; Gasparyan, A.Y.; Zimba, O. Benefits and Adverse Effects of Hydroxychloroquine, Methotrexate and Colchicine: Searching for Repurposable Drug Candidates. Rheumatol Int 2020, 40, 1. [Google Scholar] [CrossRef] [PubMed]
- Gevers, S.; Kwa, M.S.G.; Wijnans, E.; Nieuwkoop, C. van Safety Considerations for Chloroquine and Hydroxychloroquine in the Treatment of COVID-19. Clinical Microbiology and Infection 2020, 26, 1276. [Google Scholar] [CrossRef]
- Karolyi, M.; Omid, S.; Pawelka, E.; Jilma, B.; Stimpfl, T.; Schoergenhofer, C.; Laferl, H.; Seitz, T.; Traugott, M.; Wenisch, C.; et al. High Dose Lopinavir/Ritonavir Does Not Lead to Sufficient Plasma Levels to Inhibit SARS-CoV-2 in Hospitalized Patients With COVID-19. Front Pharmacol 2021, 12, 704767. [Google Scholar] [CrossRef]
- Owa, A.B.; Owa, O.T. Lopinavir/Ritonavir Use in Covid-19 Infection: Is It Completely Non-Beneficial? Journal of Microbiology, Immunology and Infection 2020, 53, 674–675. [Google Scholar] [CrossRef]
- Vecchio, G.; Zapico, V.; Catanzariti, A.; Carboni Bisso, I.; Heras, M. Las Adverse Effects of Lopinavir/Ritonavir in Critically Ill Patients with COVID-19. Medicina (B Aires) 2020, 80, 439–441. [Google Scholar]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe Covid-19. N Engl J Med 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Gagliardini, R.; Cozzi-Lepri, A.; Mariano, A.; Taglietti, F.; Vergori, A.; Abdeddaim, A.; Di Gennaro, F.; Mazzotta, V.; Amendola, A.; D’Offizi, G.; et al. No Efficacy of the Combination of Lopinavir/Ritonavir Plus Hydroxychloroquine Versus Standard of Care in Patients Hospitalized With COVID-19: A Non-Randomized Comparison. Front Pharmacol 2021, 12, 621676. [Google Scholar] [CrossRef] [PubMed]
- Reis, G.; Silva, E.A. dos S.M.; Silva, D.C.M.; Thabane, L.; Singh, G.; Park, J.J.H.; Forrest, J.I.; Harari, O.; Santos, C.V.Q. dos; Almeida, A.P.F.G. de; et al. Effect of Early Treatment With Hydroxychloroquine or Lopinavir and Ritonavir on Risk of Hospitalization Among Patients With COVID-19: The TOGETHER Randomized Clinical Trial. JAMA Netw Open 2021, 4, e216468–e216468. [Google Scholar] [CrossRef] [PubMed]
- Axfors, C.; Schmitt, A.M.; Janiaud, P.; van’t Hooft, J.; Abd-Elsalam, S.; Abdo, E.F.; Abella, B.S.; Akram, J.; Amaravadi, R.K.; Angus, D.C.; et al. Mortality Outcomes with Hydroxychloroquine and Chloroquine in COVID-19 from an International Collaborative Meta-Analysis of Randomized Trials. Nat Commun 2021, 12, 2349. [Google Scholar] [CrossRef] [PubMed]
- Rosenke, K.; Jarvis, M.A.; Feldmann, F.; Schwarz, B.; Okumura, A.; Lovaglio, J.; Saturday, G.; Hanley, P.W.; Meade-White, K.; Williamson, B.N.; et al. Hydroxychloroquine Proves Ineffective in Hamsters and Macaques Infected with SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Funnell, S.G.P.; Dowling, W.E.; Muñoz-Fontela, C.; Gsell, P.-S.; Ingber, D.E.; Hamilton, G.A.; Delang, L.; Rocha-Pereira, J.; Kaptein, S.; Dallmeier, K.H.; et al. Emerging Preclinical Evidence Does Not Support Broad Use of Hydroxychloroquine in COVID-19 Patients. Nat Commun 2020, 11, 1–4. [Google Scholar] [CrossRef]
- Saghir, S.A.; AlGabri, N.A.; Alagawany, M.M.; Attia, Y.A.; Alyileili, S.R.; Elnesr, S.S.; Shafi, M.E.; Al-shargi, O.Y.; Al-balagi, N.; Alwajeeh, A.S.; et al. <p>Chloroquine and Hydroxychloroquine for the Prevention and Treatment of COVID-19: A Fiction, Hope or Hype? An Updated Review</P>. Ther Clin Risk Manag 2021, 17, 371–387. [Google Scholar] [CrossRef]
- Mhatre, S.; Srivastava, T.; Naik, S.; Patravale, V. Antiviral Activity of Green Tea and Black Tea Polyphenols in Prophylaxis and Treatment of COVID-19: A Review. Phytomedicine 2021, 85, 153286. [Google Scholar] [CrossRef]
- Ohgitani, E.; Shin-Ya, M.; Ichitani, M.; Kobayashi, M.; Takihara, T.; Kawamoto, M.; Kinugasa, H.; Mazda, O. Significant Inactivation of SARS-CoV-2 In Vitro by a Green Tea Catechin, a Catechin-Derivative, and Black Tea Galloylated Theaflavins. Molecules 2021, Vol. 26, Page 3572 2021, 26, 3572. [Google Scholar] [CrossRef]
- Henss, L.; Auste, A.; Schürmann, C.; Schmidt, C.; Rhein, C. von; Mühlebach, M.D.; Schnierle, B.S. The Green Tea Catechin Epigallocatechin Gallate Inhibits SARS-CoV-2 Infection. Journal of General Virology 2021, 102, 001574. [Google Scholar] [CrossRef]
- Kwon, P.S.; Oh, H.; Kwon, S.-J.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated Polysaccharides Effectively Inhibit SARS-CoV-2 in Vitro. Cell Discov 2020, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory Activities of Marine Sulfated Polysaccharides against SARS-CoV-2. Food Funct 2020, 11, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Giugliano, R.; Chianese, A.; Buonocore, C.; Vitale, G.A.; Sanna, G.; Sarno, F.; Manzin, A.; Nebbioso, A.; Termolino, P.; et al. Antiviral Activity of Vitis Vinifera Leaf Extract against SARS-CoV-2 and HSV-1. Viruses 2021, Vol. 13, Page 1263 2021, 13, 1263. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Jamsransuren, D.; Matsuda, S.; Crea, R.; Ogawa, H. The SARS-CoV-2-Inactivating Activity of Hydroxytyrosol-Rich Aqueous Olive Pulp Extract (HIDROX®) and Its Use as a Virucidal Cream for Topical Application. Viruses 2021, Vol. 13, Page 232 2021, 13, 232. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z.; Chan, M.; Ouyang, M.J.; Olukitibi, T.A.; Mahmoudi, M.; Kobasa, D.; Yao, X. Identification and Evaluation of the Inhibitory Effect of Prunella Vulgaris Extract on SARS-Coronavirus 2 Virus Entry. PLoS One 2021, 16, e0251649. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-G.; Kang, K.W.; Hong, W.; Kim, Y.H.; Oh, J.T.; Park, D.W.; Ko, M.; Bai, Y.-F.; Seo, Y.-J.; Lee, S.-M.; et al. Potent Antiviral Activity of Agrimonia Pilosa, Galla Rhois, and Their Components against SARS-CoV-2. Bioorg Med Chem 2021, 45, 116329. [Google Scholar] [CrossRef]
- Rauš, K.; Pleschka, S.; Klein, P.; Schoop, R.; Fisher, P. Effect of an Echinacea-Based Hot Drink Versus Oseltamivir in Influenza Treatment: A Randomized, Double-Blind, Double-Dummy, Multicenter, Noninferiority Clinical Trial. Current Therapeutic Research 2015, 77, 66–72. [Google Scholar] [CrossRef]
- Schapowal, A. Efficacy and Safety of Echinaforce® in Respiratory Tract Infections. Wiener Medizinische Wochenschrift 2012, 163, 102–105. [Google Scholar] [CrossRef]
- Jawad, M.; Schoop, R.; Suter, A.; Klein, P.; Eccles, R. Safety and Efficacy Profile of Echinacea Purpurea to Prevent Common Cold Episodes: A Randomized, Double-Blind, Placebo-Controlled Trial. Evidence-based Complementary and Alternative Medicine 2012, 2012, 841315. [Google Scholar] [CrossRef]
- Signer, J.; Jonsdottir, H.R.; Albrich, W.C.; Strasser, M.; Züst, R.; Ryter, S.; Ackermann-Gäumann, R.; Lenz, N.; Siegrist, D.; Suter, A.; et al. In Vitro Virucidal Activity of Echinaforce®, an Echinacea Purpurea Preparation, against Coronaviruses, Including Common Cold Coronavirus 229E and SARS-CoV-2. Virol J 2020, 17, 1–11. [Google Scholar] [CrossRef]
- Su, H. xia; Yao, S.; Zhao, W. feng; Li, M. jun; Liu, J.; Shang, W. juan; Xie, H.; Ke, C. qiang; Hu, H. chen; Gao, M. na; et al. Anti-SARS-CoV-2 Activities in Vitro of Shuanghuanglian Preparations and Bioactive Ingredients. Acta Pharmacol Sin 2020, 41, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Musall, K.; Oo, A.; Cao, D.; Liang, B.; Hassandarvish, P.; Lan, S.; Slack, R.L.; Kirby, K.A.; Bassit, L.; et al. Baicalein and Baicalin Inhibit Sars-Cov-2 Rna-Dependent-Rna Polymerase. Microorganisms 2021, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Pan, W.; Li, R.; Liu, B.; Li, C.; Xie, Y.; Wang, Z.; Zhao, J.; Jiang, H.; Huang, J.; et al. Liu Shen Capsule Shows Antiviral and Anti-Inflammatory Abilities against Novel Coronavirus SARS-CoV-2 via Suppression of NF-ΚB Signaling Pathway. Pharmacol Res 2020, 158, 104850. [Google Scholar] [CrossRef] [PubMed]
- Runfeng, L.; Yunlong, H.; Jicheng, H.; Weiqi, P.; Qinhai, M.; Yongxia, S.; Chufang, L.; Jin, Z.; Zhenhua, J.; Haiming, J.; et al. Lianhuaqingwen Exerts Anti-Viral and Anti-Inflammatory Activity against Novel Coronavirus (SARS-CoV-2). Pharmacol Res 2020, 156, 104761. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, Y.; Aida, K.; Kawauchi, K.; Morimoto, M.; Ebisui, T.; Akiyama, T.; Nakamura, T. Jinhua Qinggan Granule, a Chinese Herbal Medicine against COVID-19, Induces Rapid Changes in the Plasma Levels of IL-6 and IFN-γ. medRxiv 2020. 2020.06.08.20124453. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Zhang, S.; Wang, W.; Jiang, X. Clinical Observation of Xuebijing in the Treatment of COVID-19. Chinese journal of hospital pharmacy 40, 964–967. [CrossRef]
- Tsai, K.-C.; Huang, Y.-C.; Liaw, C.-C.; Tsai, C.-I.; Chiou, C.-T.; Lin, C.-J.; Wei, W.-C.; Lin, S.J.-S.; Tseng, Y.-H.; Yeh, K.-M.; et al. A Traditional Chinese Medicine Formula NRICM101 to Target COVID-19 through Multiple Pathways: A Bedside-to-Bench Study. Biomedicine & Pharmacotherapy 2021, 133, 111037. [Google Scholar] [CrossRef]
- Jan, J.-T.; Cheng, T.-J.R.; Juang, Y.-P.; Ma, H.-H.; Wu, Y.-T.; Yang, W.-B.; Cheng, C.-W.; Chen, X.; Chou, T.-H.; Shie, J.-J.; et al. Identification of Existing Pharmaceuticals and Herbal Medicines as Inhibitors of SARS-CoV-2 Infection. Proceedings of the National Academy of Sciences 2021, 118, e2021579118. [Google Scholar] [CrossRef]
- Du, R.; Cooper, L.; Chen, Z.; Lee, H.; Rong, L.; Cui, Q. Discovery of Chebulagic Acid and Punicalagin as Novel Allosteric Inhibitors of SARS-CoV-2 3CLpro. Antiviral Res 2021, 190, 105075. [Google Scholar] [CrossRef]
- MS, N.; Y, H.; DA, F.; SJ, P.; J, W.; MJ, T.; PJ, W. Artemisia Annua L. Extracts Inhibit the in Vitro Replication of SARS-CoV-2 and Two of Its Variants. J Ethnopharmacol 2021, 274, 114016. [Google Scholar] [CrossRef]
- Roshdy, W.H.; Rashed, H.A.; Kandeil, A.; Mostafa, A.; Moatasim, Y.; Kutkat, O.; Shama, N.M.A.; Gomaa, M.R.; El-Sayed, I.H.; Guindy, N.M. El; et al. EGYVIR: An Immunomodulatory Herbal Extract with Potent Antiviral Activity against SARS-CoV-2. PLoS One 2020, 15, e0241739. [Google Scholar] [CrossRef]
- Plante, K.S.; Dwivedi, V.; Plante, J.A.; Fernandez, D.; Mirchandani, D.; Bopp, N.; Aguilar, P. V.; Park, J.G.; Tamayo, P.P.; Delgado, J.; et al. Antiviral Activity of Oleandrin and a Defined Extract of Nerium Oleander against SARS-CoV-2. Biomedicine & Pharmacotherapy 2021, 138, 111457. [Google Scholar] [CrossRef]
- Enmozhi, S.K.; Raja, K.; Sebastine, I.; Joseph, J. Andrographolide as a Potential Inhibitor of SARS-CoV-2 Main Protease: An in Silico Approach. J Biomol Struct Dyn 2020, 39, 3092–3098. [Google Scholar] [CrossRef]
- Shi, T.-H.; Huang, Y.-L.; Chen, C.-C.; Pi, W.-C.; Hsu, Y.-L.; Lo, L.-C.; Chen, W.-Y.; Fu, S.-L.; Lin, C.-H. Andrographolide and Its Fluorescent Derivative Inhibit the Main Proteases of 2019-NCoV and SARS-CoV through Covalent Linkage. Biochem Biophys Res Commun 2020, 533, 467. [Google Scholar] [CrossRef]
- Sa-ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis Paniculata Extract and Its Major Component Andrographolide in Human Lung Epithelial Cells and Cytotoxicity Evaluation in Major Organ Cell Representatives. J Nat Prod 2021, 84, 1261–1270. [Google Scholar] [CrossRef]
- Khayrani, A.C.; Irdiani, R.; Aditama, R.; Pratami, D.K.; Lischer, K.; Ansari, M.J.; Chinnathambi, A.; Alharbi, S.A.; Almoallim, H.S.; Sahlan, M. Evaluating the Potency of Sulawesi Propolis Compounds as ACE-2 Inhibitors through Molecular Docking for COVID-19 Drug Discovery Preliminary Study. J King Saud Univ Sci 2021, 33, 101297. [Google Scholar] [CrossRef]
- Dewi, L.K.; Sahlan, M.; Pratami, D.K.; Agus, A.; Agussalim; Sabir, A. Identifying Propolis Compounds Potential to Be Covid-19 Therapies by Targeting Sars-Cov-2 Main Protease. International Journal of Applied Pharmaceutics 2021, 13, 103–110. [Google Scholar] [CrossRef]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential Antiviral Activity of Isorhamnetin against SARS-CoV-2 Spike Pseudotyped Virus in Vitro. Drug Dev Res 2021, 10, 1002. [Google Scholar] [CrossRef]
- Owis, A.; S. El-Hawary, M.; Amir, D.E.; M. Aly, O.; Ramadan Abdelmohsen, U.; S. Kamel, M. Molecular Docking Reveals the Potential of Salvadora Persica Flavonoids to Inhibit COVID-19 Virus Main Protease. RSC Adv 2020, 10, 19570–19575. [Google Scholar] [CrossRef]
- Pandey, P.; Rane, J.S.; Chatterjee, A.; Kumar, A.; Khan, R.; Prakash, A.; Ray, S. Targeting SARS-CoV-2 Spike Protein of COVID-19 with Naturally Occurring Phytochemicals: An in Silico Study for Drug Development. J Biomol Struct Dyn 2020, 36, 6306–6316. [Google Scholar] [CrossRef]
- Zhang, Z.R.; Zhang, Y.N.; Li, X.D.; Zhang, H.Q.; Xiao, S.Q.; Deng, F.; Yuan, Z.M.; Ye, H.Q.; Zhang, B. A Cell-Based Large-Scale Screening of Natural Compounds for Inhibitors of SARS-CoV-2. Signal Transduct Target Ther 2020, 5, 218. [Google Scholar] [CrossRef]
- Abdallah, H.M.; El-Halawany, A.M.; Sirwi, A.; El-Araby, A.M.; Mohamed, G.A.; Ibrahim, S.R.M.; Koshak, A.E.; Asfour, H.Z.; Awan, Z.A.; Elfaky, M.A. Repurposing of Some Natural Product Isolates as SARS-COV-2 Main Protease Inhibitors via In Vitro Cell Free and Cell-Based Antiviral Assessments and Molecular Modeling Approaches. Pharmaceuticals 2021, 14, 213. [Google Scholar] [CrossRef]
- Ellen, B.M. ter; Kumar, N.D.; Bouma, E.M.; Troost, B.; Pol, D.P.I. van de; Ende-Metselaar, H.H. van der; Apperloo, L.; Gosliga, D. van; Berge, M. van den; Nawijn, M.C.; et al. Resveratrol And Pterostilbene Potently Inhibit SARS-CoV-2 Replication In Vitro. Viruses 2021, 13, 1335. [Google Scholar] [CrossRef]
- Zhang, S.; Pei, R.; Li, M.; Sun, H.; Su, M.; Ding, Y.; Chen, X.; Du, Z.; Jin, C.; Huang, C.; et al. Structural Characterization of Cocktail-like Targeting Polysaccharides from Ecklonia Kurome Okam and Their Anti-SARS-CoV-2 Activities Invitro. bioRxiv 2021. 2021.01.14.426521. [Google Scholar] [CrossRef]
- Orfali, R.; Rateb, M.E.; Hassan, H.M.; Alonazi, M.; Gomaa, M.R.; Mahrous, N.; GabAllah, M.; Kandeil, A.; Perveen, S.; Abdelmohsen, U.R.; et al. Sinapic Acid Suppresses SARS CoV-2 Replication by Targeting Its Envelope Protein. Antibiotics 2021, 10, 420. [Google Scholar] [CrossRef]
- Svenningsen, E.B.; Thyrsted, J.; Blay-Cadanet, J.; Liu, H.; Lin, S.; Moyano-Villameriel, J.; Olagnier, D.; Idorn, M.; Paludan, S.R.; Holm, C.K.; et al. Ionophore Antibiotic X-206 Is a Potent Inhibitor of SARS-CoV-2 Infection in Vitro. Antiviral Res 2021, 185, 104988. [Google Scholar] [CrossRef]
- Braga, L.; Ali, H.; Secco, I.; Chiavacci, E.; Neves, G.; Goldhill, D.; Penn, R.; Jimenez-Guardeño, J.M.; Ortega-Prieto, A.M.; Bussani, R.; et al. Drugs That Inhibit TMEM16 Proteins Block SARS-CoV-2 Spike-Induced Syncytia. Nature 2021, 594, 88–93. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Zhang, Q.-Y.; Li, X.-D.; Xiong, J.; Xiao, S.-Q.; Wang, Z.; Zhang, Z.-R.; Deng, C.-L.; Yang, X.-L.; Wei, H.-P.; et al. Gemcitabine, Lycorine and Oxysophoridine Inhibit Novel Coronavirus (SARS-CoV-2) in Cell Culture. Emerg Microbes Infect 2020, 9, 1170–1173. [Google Scholar] [CrossRef]
- Pacl, H.T.; Tipper, J.L.; Sevalkar, R.R.; Crouse, A.; Crowder, C.; Institute, U.P.M.; Ahmad, S.; Ahmad, A.; Holder, G.D.; Kuhlman, C.J.; et al. Water-Soluble Tocopherol Derivatives Inhibit SARS-CoV-2 RNA-Dependent RNA Polymerase. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Q.; Zhou, W.; Gao, S.; Lin, H.; Ye, S.; Xu, Y.; Cai, J. Chinese Medicine Injection Shuanghuanglian for Treatment of Acute Upper Respiratory Tract Infection: A Systematic Review of Randomized Controlled Trials. Evid Based Complement Alternat Med 2013, 2013, 987326. [Google Scholar] [CrossRef]
- Gu, S.; Yin, N.; Pei, J.; Lai, L. Understanding Traditional Chinese Medicine Anti-Inflammatory Herbal Formulae by Simulating Their Regulatory Functions in the Human Arachidonic Acid Metabolic Network. Mol Biosyst 2013, 9, 1931–1938. [Google Scholar] [CrossRef]
- Tian, S.; He, G.; Song, J.; Wang, S.; Xin, W.; Zhang, D.; Du, G. Pharmacokinetic Study of Baicalein after Oral Administration in Monkeys. Fitoterapia 2012, 83, 532–540. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef]
- Varughese, J.K.; Libin, K.L.J.; Sindhu, K.S.; Rosily, A. V.; Abi, T.G. Investigation of the Inhibitory Activity of Some Dietary Bioactive Flavonoids against SARS-CoV-2 Using Molecular Dynamics Simulations and MM-PBSA Calculations. J Biomol Struct Dyn 2021, 23, 1–6. [Google Scholar] [CrossRef]
- Pasquereau, S.; Nehme, Z.; Ahmad, S.H.; Daouad, F.; Assche, J. Van; Wallet, C.; Schwartz, C.; Rohr, O.; Morot-Bizot, S.; Herbein, G. Resveratrol Inhibits HCoV-229E and SARS-CoV-2 Coronavirus Replication In Vitro. Viruses 2021, 13, 354. [Google Scholar] [CrossRef]
- Yang, M.; Wei, J.; Huang, T.; Lei, L.; Shen, C.; Lai, J.; Yang, M.; Liu, L.; Yang, Y.; Liu, G.; et al. Resveratrol Inhibits the Replication of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in Cultured Vero Cells. Phytother Res 2021, 35, 1127–1129. [Google Scholar] [CrossRef]
- She, G.-M.; Xu, C.; Liu, B.; Shi, R.-B. Polyphenolic Acids from Mint (the Aerial of Mentha Haplocalyx Briq.) with DPPH Radical Scavenging Activity. J Food Sci 2010, 75, C359–C362. [Google Scholar] [CrossRef]
- Tiryaki Sönmez, G.; Çolak, M.; Sönmez, S.; Schoenfeld, B. Effects of Oral Supplementation of Mint Extract on Muscle Pain and Blood Lactate. Biomed Hum Kinet 2010, 2, 66–69. [Google Scholar] [CrossRef]
- Pengfei, L.; Tiansheng, D.; Xianglin, H.; Jianguo, W. Antioxidant Properties of Isolated Isorhamnetin from the Sea Buckthorn Marc. Plant Foods Hum Nutr 2009, 64, 141–145. [Google Scholar] [CrossRef]
- Olas, B.; Skalski, B.; Ulanowska, K. The Anticancer Activity of Sea Buckthorn [Elaeagnus Rhamnoides (L.) A. Nelson]. Front Pharmacol 2018, 9, 232. [Google Scholar] [CrossRef]
- Dayem, A.A.; Choi, H.Y.; Kim, Y.B.; Cho, S.-G. Antiviral Effect of Methylated Flavonol Isorhamnetin against Influenza. PLoS One 2015, 10, e0121610. [Google Scholar] [CrossRef]
- Matboli, M.; Saad, M.; Hasanin, A.H.; A. Saleh, L.; Baher, W.; Bekhet, M.M.; Eissa, S. New Insight into the Role of Isorhamnetin as a Regulator of Insulin Signaling Pathway in Type 2 Diabetes Mellitus Rat Model: Molecular and Computational Approach. Biomedicine & Pharmacotherapy 2021, 135, 111176. [Google Scholar] [CrossRef]
- Liu, C.-M.; Westley, J.W.; Hermann, T.E.; Prosser, B.L.T.; Palleroni, N.; Evans, R.H.; Miller, P.A. Novel Polyether Antibiotics X-14873A, G and H Produced by a Streptomyces: Taxonomy of the Producing Culture, Fermentation, Biological and Ionophorous Properties of the Antibiotics. J Antibiot (Tokyo) 1986, 39, 1712–1718. [Google Scholar] [CrossRef]
- Liu, C.M.; Westley, J.W.; Chu, J.; Hermann, T.E.; Liu, M.; Palleroni, N. Three Novel Polyether Antibiotics X-14889a, c, and d from a Streptomycete Taxonomy of the Producing Ogranism, Fermentation Production and Biological Properties of the Antibiotics. Journal of Antibiotics 1993, 46, 275–279. [Google Scholar] [CrossRef]
- Rausch, K.; Hackett, B.; Weinbren, N.; Reeder, S.; Sadovsky, Y.; Hunter, C.; Schultz, D.C.; Coyne, C.; Cherry, S. Screening Bioactives Reveals Nanchangmycin as a Broad Spectrum Antiviral Active against Zika Virus. Cell Rep 2017, 18, 804–815. [Google Scholar] [CrossRef]
- Nakamura, M.; Kunimoto, S.; Takahashi, Y.; Naganawa, H.; Sakaue, M.; Inoue, S.; Ohno, T.; Takeuchi, T. Inhibitory Effects of Polyethers on Human Immunodeficiency Virus Replication. Antimicrob Agents Chemother 1992, 36, 492–494. [Google Scholar] [CrossRef]
- Velthuis, A.J.W. te; Worm, S.H.E. van den; Sims, A.C.; Baric, R.S.; Snijder, E.J.; Hemert, M.J. van Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture. PLoS Pathog 2010, 6, e1001176. [Google Scholar] [CrossRef]
- Kanwal, N.; Rasul, A.; Hussain, G.; Anwar, H.; Shah, M.A.; Sarfraz, I.; Riaz, A.; Batool, R.; Shahbaz, M.; Hussain, A.; et al. Oleandrin: A Bioactive Phytochemical and Potential Cancer Killer via Multiple Cellular Signaling Pathways. Food and Chemical Toxicology 2020, 143, 111570. [Google Scholar] [CrossRef]
- Afaq, F.; Saleem, M.; Aziz, M.H.; Mukhtar, H. Inhibition of 12-O-Tetradecanoylphorbol-13-Acetate-Induced Tumor Promotion Markers in CD-1 Mouse Skin by Oleandrin. Toxicol Appl Pharmacol 2004, 195, 361–369. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, Y.; Zhao, W.; Zhou, X.; Wang, C.; Deng, F. The Cardiac Glycoside Oleandrin Induces Apoptosis in Human Colon Cancer Cells via the Mitochondrial Pathway. Cancer Chemotherapy and Pharmacology 2017 80:1 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Yang, C.-W.; Lee, Y.-Z.; Hsu, H.-Y.; Jan, J.-T.; Lin, Y.-L.; Chang, S.-Y.; Peng, T.-T.; Yang, R.-B.; Liang, J.-J.; Liao, C.-C.; et al. Inhibition of SARS-CoV-2 by Highly Potent Broad-Spectrum Anti-Coronaviral Tylophorine-Based Derivatives. Front Pharmacol 2020, 11, 2056. [Google Scholar] [CrossRef]
- Dupuis, C.; Montmollin, E. de; Buetti, N.; Goldgran-Toledano, D.; Reignier, J.; Schwebel, C.; Domitile, J.; Neuville, M.; Ursino, M.; Siami, S.; et al. Impact of Early Corticosteroids on 60-Day Mortality in Critically Ill Patients with COVID-19: A Multicenter Cohort Study of the OUTCOMEREA Network. PLoS One 2021, 16, e0255644. [Google Scholar] [CrossRef]
- van Paassen, J.; Vos, J.S.; Hoekstra, E.M.; Neumann, K.M.I.; Boot, P.C.; Arbous, S.M. Corticosteroid Use in COVID-19 Patients: A Systematic Review and Meta-Analysis on Clinical Outcomes. Crit Care 2020, 24, 1–22. [Google Scholar] [CrossRef]
- Group, T.R.C. Dexamethasone in Hospitalized Patients with Covid-19. N Engl J Med 2020, 384, 693–704. [Google Scholar] [CrossRef]
- Tang, X.; Feng, Y.-M.; Ni, J.-X.; Zhang, J.-Y.; Liu, L.-M.; Hu, K.; Wu, X.-Z.; Zhang, J.-X.; Chen, J.-W.; Zhang, J.-C.; et al. Early Use of Corticosteroid May Prolong SARS-CoV-2 Shedding in Non-Intensive Care Unit Patients with COVID-19 Pneumonia: A Multicenter, Single-Blind, Randomized Control Trial. Respiration 2021, 100, 116–126. [Google Scholar] [CrossRef]
- Berton, A.M.; Prencipe, N.; Giordano, R.; Ghigo, E.; Grottoli, S. Systemic Steroids in Patients with COVID-19: Pros and Contras, an Endocrinological Point of View. J Endocrinol Invest 2021, 44, 873–875. [Google Scholar] [CrossRef]
- Wang, J.; Yang, W.; Chen, P.; Guo, J.; Liu, R.; Wen, P.; Li, K.; Lu, Y.; Ma, T.; Li, X.; et al. The Proportion and Effect of Corticosteroid Therapy in Patients with COVID-19 Infection: A Systematic Review and Meta-Analysis. PLoS One 2021, 16, e0249481. [Google Scholar] [CrossRef]
- Hu, K.; Guan, W. jie; Bi, Y.; Zhang, W.; Li, L.; Zhang, B.; Liu, Q.; Song, Y.; Li, X.; Duan, Z.; et al. Efficacy and Safety of Lianhuaqingwen Capsules, a Repurposed Chinese Herb, in Patients with Coronavirus Disease 2019: A Multicenter, Prospective, Randomized Controlled Trial. Phytomedicine 2021, 85, 153242. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, H.; Wu, M.; Zhou, Y.; Che, J.; Zhang, S.; Meng, D.; Liu, J.; Ma, L.; Peng, Y.; et al. Integrated Lianhua-Qingwen and Western Medicine Versus Western Medicine-Only Therapies Against COVID-19: A Systematic Review and Meta-Analysis. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Wang, S.X.; Li, M.Y.; Chen, X.L.; Ma, M.Y.; Hu, J.H. Clinical Efficacy of Lianhua Qingwen Integrated with Western Medicine on COVID-19 by Meta-Analysis. Chinese Traditional and Herbal Drugs 2020, 51, 3763–3769. [Google Scholar] [CrossRef]
- Yan, H.; Zou, C. Mechanism and Material Basis of LianhuaQingwen Capsule for Improving Clinical Cure Rate of COVID-19: A Study Based on Network Pharmacology and Molecular Docking Technology. Journal of Southern Medical University 2021, 41, 20. [Google Scholar] [CrossRef]
- Ling, X.Y.; Tao, J.L.; Sun, X.; Yuan, B. Exploring Material Basis and Mechanism of Lianhua Qingwen Prescription against Coronavirus Based on Network Pharmacology. Chinese Traditional and Herbal Drugs 2020, 51, 1723–1730. [Google Scholar] [CrossRef]
- Zhang, Y.; Yao, Y.; Yang, Y.; Wu, H. Investigation of Anti-SARS, MERS, and COVID-19 Effect of Jinhua Qinggan Granules Based on a Network Pharmacology and Molecular Docking Approach: Nat Prod Commun 2021, 16, 1–9. [CrossRef]
- Xing, Y.; Hua, Y.R.; Shang, J.; Ge, W.H.; Liao, J. Traditional Chinese Medicine Network Pharmacology Study on Exploring the Mechanism of Xuebijing Injection in the Treatment of Coronavirus Disease 2019. Chin J Nat Med 2020, 18, 941–951. [Google Scholar] [CrossRef]
- Ren, Y.; Yin, Z.; Dai, J.; Yang, Z.; Ye, B.; Ma, Y.; Zhang, T.; Shi, Y. Evidence-Based Complementary and Alternative Medicine Exploring Active Components and Mechanism of Jinhua Qinggan Granules in Treatment of COVID-19 Based on Virus-Host Interaction: Nat Prod Commun 2020, 15, 1–11. [CrossRef]
- Nader-Macías, M.E.F.; De Gregorio, P.R.; Silva, J.A. Probiotic Lactobacilli in Formulas and Hygiene Products for the Health of the Urogenital Tract. Pharmacol Res Perspect 2021, 9, e00787. [Google Scholar] [CrossRef]
- Nava, G.M.; Bielke, L.R.; Callaway, T.R.; Castañeda, M.P. Probiotic Alternatives to Reduce Gastrointestinal Infections: The Poultry Experience. Anim Health Res Rev 2005, 6, 105–118. [Google Scholar] [CrossRef]
- Michail, S. The Role of Probiotics in Allergic Diseases. Allergy, Asthma and Clinical Immunology 2009, 5, 1–7. [Google Scholar] [CrossRef]
- Makino, S.; Ikegami, S.; Kano, H.; Sashihara, T.; Sugano, H.; Horiuchi, H.; Saito, T.; Oda, M. Immunomodulatory Effects of Polysaccharides Produced by Lactobacillus Delbrueckii Ssp. Bulgaricus OLL1073R-1. J Dairy Sci 2006, 89, 2873–2881. [Google Scholar] [CrossRef]
- Kim, K.W.; Kang, S.S.; Woo, S.J.; Park, O.J.; Ahn, K.B.; Song, K.D.; Lee, H.K.; Yun, C.H.; Han, S.H. Lipoteichoic Acid of Probiotic Lactobacillus Plantarum Attenuates Poly I:C-Induced IL-8 Production in Porcine Intestinal Epithelial Cells. Front Microbiol 2017, 8, 1827. [Google Scholar] [CrossRef]
- Zielińska, D.; Sionek, B.; Kołożyn-Krajewska, D. Safety of Probiotics. Diet, Microbiome and Health 2018, 131–161. [Google Scholar] [CrossRef]
- Chen, M.F.; Weng, K.F.; Huang, S.Y.; Liu, Y.C.; Tseng, S.N.; Ojcius, D.M.; Shih, S.R. Pretreatment with a Heat-Killed Probiotic Modulates Monocyte Chemoattractant Protein-1 and Reduces the Pathogenicity of Influenza and Enterovirus 71 Infections. Mucosal Immunology 2016, 10, 215–227. [Google Scholar] [CrossRef]
- Hart, A.L.; Lammers, K.; Brigidi, P.; Vitali, B.; Rizzello, F.; Gionchetti, P.; Campieri, M.; Kamm, M.A.; Knight, S.C.; Stagg, A.J. Modulation of Human Dendritic Cell Phenotype and Function by Probiotic Bacteria. Gut 2004, 53, 1602–1609. [Google Scholar] [CrossRef]
- Tsuji, R.; Yamamoto, N.; Yamada, S.; Fujii, T.; Yamamoto, N.; Kanauchi, O. Induction of Anti-Viral Genes Mediated by Humoral Factors upon Stimulation with Lactococcus Lactis Strain Plasma Results in Repression of Dengue Virus Replication in Vitro. Antiviral Res 2018, 160, 101–108. [Google Scholar] [CrossRef]
- Suzuki, H.; Tsuji, R.; Sugamata, M.; Yamamoto, N.; Yamamoto, N.; Kanauchi, O. Administration of Plasmacytoid Dendritic Cell-Stimulative Lactic Acid Bacteria Is Effective against Dengue Virus Infection in Mice. Int J Mol Med 2019, 43, 426–434. [Google Scholar] [CrossRef]
- Zhang, L.; Han, H.; Li, X.; Chen, C.; Xie, X.; Su, G.; Ye, S.; Wang, C.; He, Q.; Wang, F.; et al. Probiotics Use Is Associated with Improved Clinical Outcomes among Hospitalized Patients with COVID-19. Therap Adv Gastroenterol 2021, 14. [Google Scholar] [CrossRef]
- Kageyama, Y.; Nishizaki, Y.; Aida, K.; Yayama, K.; Ebisui, T.; Akiyama, T.; Nakamura, T. Lactobacillus Plantarum Induces Innate Cytokine Responses That Potentially Provide a Protective Benefit against COVID-19: A Single-Arm, Double-Blind, Prospective Trial Combined with an in Vitro Cytokine Response Assay. Exp Ther Med 2022, 23. [Google Scholar] [CrossRef]
- Gutiérrez-Castrellón, P.; Gandara-Martí, T.; Abreu, A.T.; Nieto-Rufino, C.D.; López-Orduña, E.; Jiménez-Escobar, I.; Jiménez-Gutiérrez, C.; López-Velazquez, G.; Espadaler-Mazo, J. Efficacy and Safety of Novel Probiotic Formulation in Adult Covid19 Outpatients: A Randomized, Placebo-Controlled Clinical Trial. medRxiv 2021. 2021.05.20.21256954. [Google Scholar] [CrossRef]
- Chaiyasut, B.; Sundaram, S.; Kesika, P.; Fujii, K.; Kubo, Y.; Noguchi, T.; Tobita, K. Effects of Bacillus Subtilis Natto Strains on Antiviral Responses in Resiquimod-Stimulated Human M1-Phenotype Macrophages. 2023. [CrossRef]
- Rathi, A.; Jadhav, S.B.; Shah, N. A Randomized Controlled Trial of the Efficacy of Systemic Enzymes and Probiotics in the Resolution of Post-COVID Fatigue. Medicines 2021, Vol. 8, Page 47 2021, 8, 47. [Google Scholar] [CrossRef]
- What to Start: Initial Combination Regimens for the Antiretroviral-Naive Patient | NIH. Available online: https://clinicalinfo.hiv.gov/en/guidelines/hiv-clinical-guidelines-adult-and-adolescent-arv/what-start-initial-combination-regimens (accessed on 16 March 2023).
- Jockusch, S.; Tao, C.; Li, X.; Chien, M.; Kumar, S.; Morozova, I.; Kalachikov, S.; Russo, J.J.; Ju, J. Sofosbuvir Terminated RNA Is More Resistant to SARS-CoV-2 Proofreader than RNA Terminated by Remdesivir. Sci Rep 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Jácome, R.; Campillo-Balderas, J.A.; Ponce de León, S.; Becerra, A.; Lazcano, A. Sofosbuvir as a Potential Alternative to Treat the SARS-CoV-2 Epidemic. Sci Rep 2020, 10, 1–5. [Google Scholar] [CrossRef]
- Sacramento, C.Q.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Silva, A. de P.D. Da; Dias, S. da S.G.; Silva, C. dos S. da; Ferreira, A.C.; Mattos, M.; Pão, C.R.R.; Freitas, C.S. de; et al. In Vitro Antiviral Activity of the Anti-HCV Drugs Daclatasvir and Sofosbuvir against SARS-CoV-2, the Aetiological Agent of COVID-19. J Antimicrob Chemother 2021, 76, 1874–1885. [Google Scholar] [CrossRef]
- Houston, D.M.J.; Bugert, J.J.; Denyer, S.P.; Heard, C.M. Potentiated Virucidal Activity of Pomegranate Rind Extract (PRE) and Punicalagin against Herpes Simplex Virus (HSV) When Co-Administered with Zinc (II) Ions, and Antiviral Activity of PRE against HSV and Aciclovir-Resistant HSV. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Haidari, M.; Ali, M.; Ward Casscells, S.; Madjid, M. Pomegranate (Punica Granatum) Purified Polyphenol Extract Inhibits Influenza Virus and Has a Synergistic Effect with Oseltamivir. Phytomedicine 2009, 16, 1127–1136. [Google Scholar] [CrossRef]
- Saadh, M.; Almaaytah, A.; Alaraj, M.; Dababneh, M.; Sa’adeh, I.; Aldalaen, S.; Kharshid, A.; Alboghdadly, A.; Hailat, M.; Khaleel, A.; et al. Punicalagin and Zinc (II) Ions Inhibit the Activity of SARS-CoV-2 3CL-Protease in Vitro. Eur Rev Med Pharmacol Sci 2021, 25, 3908–3913. [Google Scholar] [CrossRef]
- Colunga Biancatelli, R.M.L.; Berrill, M.; Catravas, J.D.; Marik, P.E. Quercetin and Vitamin C: An Experimental, Synergistic Therapy for the Prevention and Treatment of SARS-CoV-2 Related Disease (COVID-19). Front Immunol 2020, 11, 1451. [Google Scholar] [CrossRef]
- Wakaskar, R.R. General Overview of Lipid–Polymer Hybrid Nanoparticles, Dendrimers, Micelles, Liposomes, Spongosomes and Cubosomes. 2017, 26, 311–318. [CrossRef]
- Günther, S.C.; Maier, J.D.; Vetter, J.; Podvalnyy, N.; Khanzhin, N.; Hennet, T.; Stertz, S. Antiviral Potential of 3′-Sialyllactose- and 6′-Sialyllactose-Conjugated Dendritic Polymers against Human and Avian Influenza Viruses. Scientific Reports 2020 10:1 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Lee, M.-K. Liposomes for Enhanced Bioavailability of Water-Insoluble Drugs: In Vivo Evidence and Recent Approaches. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Liang, J.; Wu, W.; Liu, Q.; Chen, S. Long-Circulating Nanoliposomes (LCNs) Sustained Delivery of Baicalein (BAI) with Desired Oral Bioavailability in Vivo. 2013, 20, 319–323. [CrossRef]
- Refaat, H.; Mady, F.M.; Sarhan, H.A.; Rateb, H.S.; Alaaeldin, E. Optimization and Evaluation of Propolis Liposomes as a Promising Therapeutic Approach for COVID-19. Int J Pharm 2021, 592, 120028. [Google Scholar] [CrossRef]
- Fu, M.; Han, X.; Chen, B.; Guo, L.; Zhong, L.; Hu, P.; Pan, Y.; Qiu, M.; Cao, P.; Chen, J. Cancer Treatment: From Traditional Chinese Herbal Medicine to the Liposome Delivery System. Acta Materia Medica 2022, 1, 486–506. [Google Scholar] [CrossRef]
- Que, H.; Hong, W.; Lan, T.; Zeng, H.; Chen, L.; Wan, D.; Bi, Z.; Ren, W.; Luo, M.; Yang, J.; et al. Tripterin Liposome Relieves Severe Acute Respiratory Syndrome as a Potent COVID-19 Treatment. Signal Transduct Target Ther 2022, 7. [Google Scholar] [CrossRef]
- Boroumand, H.; Badie, F.; Mazaheri, S.; Seyedi, Z.S.; Nahand, J.S.; Nejati, M.; Baghi, H.B.; Abbasi-Kolli, M.; Badehnoosh, B.; Ghandali, M.; et al. Chitosan-Based Nanoparticles Against Viral Infections. Front Cell Infect Microbiol 2021, 0, 175. [Google Scholar] [CrossRef]
- Ju, J.; Kim, J.; Choi, Y.; Jin, S.; Kim, S.; Son, D.; Shin, M. Punicalagin-Loaded Alginate/Chitosan-Gallol Hydrogels for Efficient Wound Repair and Hemostasis. Polymers 2022, Vol. 14, Page 3248 2022, 14, 3248. [Google Scholar] [CrossRef]
- Wang, D.; Mehrabi Nasab, E.; Athari, S.S. Study Effect of Baicalein Encapsulated/Loaded Chitosan-Nanoparticle on Allergic Asthma Pathology in Mouse Model. Saudi J Biol Sci 2021, 28, 4311–4317. [Google Scholar] [CrossRef]
- Saadh, M.J.; Aldalaen, S.M. Inhibitory Effects of Epigallocatechin Gallate (EGCG) Combined with Zinc Sulfate and Silver Nanoparticles on Avian Influenza A Virus Subtype H5N1. Eur Rev Med Pharmacol Sci 2021, 25, 2630–2636. [Google Scholar] [CrossRef]
- M, C.; JF, P.; M, F.; A, J.; JB, C.; B, K.; M, B. Surface Modification of PAMAM Dendrimer Improves Its Biocompatibility. Nanomedicine 2012, 8, 815–817. [Google Scholar] [CrossRef]
- TA, E.; VP, T. Liposomes for Targeted Delivery of Antithrombotic Drugs. Expert Opin Drug Deliv 2008, 5, 1185–1198. [Google Scholar] [CrossRef]
- A, G.; H, S.; Y, B. Pharmacokinetics of Pegylated Liposomal Doxorubicin: Review of Animal and Human Studies. Clin Pharmacokinet 2003, 42, 419–436. [Google Scholar] [CrossRef]
- Kumar, P.D.; Kumar, P.V.; Selvam, T.; Rao, K.S. Prolonged Drug Delivery System of PEGylated PAMAM Dendrimers with a Anti-HIV Drug. undefined 2015. [Google Scholar]
- Hama, S.; Sakai, M.; Itakura, S.; Majima, E.; Kogure, K. Rapid Modification of Antibodies on the Surface of Liposomes Composed of High-Affinity Protein A-Conjugated Phospholipid for Selective Drug Delivery. Biochem Biophys Rep 2021, 27, 101067. [Google Scholar] [CrossRef]
- Low-Income, Uninsured Face Hurdles to Obtain Covid Antivirals. Available online: https://www.nbcnews.com/health/health-news/antiviral-hurdles-low-income-people-furthers-divisions-covid-rcna14565 (accessed on 26 April 2023).
- Eedara, B.B.; Alabsi, W.; Encinas-Basurto, D.; Polt, R.; Ledford, J.G.; Mansour, H.M. Inhalation Delivery for the Treatment and Prevention of COVID-19 Infection. Pharmaceutics 2021, 13, 1077. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 Entry Factors Are Highly Expressed in Nasal Epithelial Cells Together with Innate Immune Genes. Nature Medicine 2020 26:5 2020, 26, 681–687. [Google Scholar] [CrossRef]
- JH, A.; J, K.; SP, H.; SY, C.; MJ, Y.; YS, J.; YT, K.; HM, K.; MDT, R.; MK, C.; et al. Nasal Ciliated Cells Are Primary Targets for SARS-CoV-2 Replication in the Early Stage of COVID-19. J Clin Invest 2021, 131. [Google Scholar] [CrossRef]
- GL, D. Infection Site Concentrations: Their Therapeutic Importance and the Macrolide and Macrolide-like Class of Antibiotics. Pharmacotherapy 2005, 25. [Google Scholar] [CrossRef]
- Fazekas, T.; Eickhoff, P.; Pruckner, N.; Vollnhofer, G.; Fischmeister, G.; Diakos, C.; Rauch, M.; Verdianz, M.; Zoubek, A.; Gadner, H.; et al. Lessons Learned from a Double-Blind Randomised Placebo-Controlled Study with a Iota-Carrageenan Nasal Spray as Medical Device in Children with Acute Symptoms of Common Cold. BMC Complement Altern Med 2012, 12, 147. [Google Scholar] [CrossRef]
- Fais, F.; Juskeviciene, R.; Francardo, V.; Mateos, S.; Constant, S.; Borelli, M.; Hohenfeld, I.P.; Meyer, T. Drug-Free Nasal Spray as a Barrier against SARS-CoV-2 Infection: Safety and Efficacy in Human Nasal Airway Epithelia. bioRxiv 2021. 2021.07.12.452021. [Google Scholar] [CrossRef]
- JS, P.; B, T.; S, F.; JB, W.; SM, B.; JA, C. Efficacy of Povidone-Iodine Nasal and Oral Antiseptic Preparations Against Severe Acute Respiratory Syndrome-Coronavirus 2 (SARS-CoV-2). Ear Nose Throat J 2021, 100, 192S–196S. [Google Scholar] [CrossRef]
- Lu, J.; Yin, Q.; Pei, R.; Zhang, Q.; Qu, Y.; Pan, Y.; Sun, L.; Gao, D.; Liang, C.; Yang, J.; et al. Nasal Delivery of Broadly Neutralizing Antibodies Protects Mice from Lethal Challenge with SARS-CoV-2 Delta and Omicron Variants. Virol Sin 2022, 37, 238–247. [Google Scholar] [CrossRef]
- Go, C.C.; Pandav, K.; Sanchez-Gonzalez, M.A.; Ferrer, G. Potential Role of Xylitol Plus Grapefruit Seed Extract Nasal Spray Solution in COVID-19: Case Series. Cureus 2020, 12. [Google Scholar] [CrossRef]
- Meister, T.L.; Todt, D.; Brüggemann, Y.; Steinmann, J.; Banava, S.; Brill, F.H.H.; Pfaender, S.; Steinmann, E. Virucidal Activity of Nasal Sprays against Severe Acute Respiratory Syndrome Coronavirus-2. J Hosp Infect 2022, 120, 9. [Google Scholar] [CrossRef]
- Yip, K.M.; Lee, K.M.; Ng, T.B.; Xu, S.; Yung, K.K.L.; Qu, S.; Cheung, A.K.L.; Sze, S.C.W. An Anti-Inflammatory and Anti-Fibrotic Proprietary Chinese Medicine Nasal Spray Designated as Allergic Rhinitis Nose Drops (ARND) with Potential to Prevent SARS-CoV-2 Coronavirus Infection by Targeting RBD (Delta)- Angiotensin Converting Enzyme 2 (ACE2) Binding. Chin Med 2022, 17. [Google Scholar] [CrossRef]
- Ng, K.K.; Ng, M.K.; Zhyvotovska, A.; Singh, S.; Shevde, K. Acute Respiratory Failure Secondary to COVID-19 Viral Pneumonia Managed With Hydroxychloroquine/Azithromycin Treatment. Cureus 2020, 12. [Google Scholar] [CrossRef]
- Buehler, P.K.; Zinkernagel, A.S.; Hofmaenner, D.A.; Wendel Garcia, P.D.; Acevedo, C.T.; Gómez-Mejia, A.; Mairpady Shambat, S.; Andreoni, F.; Maibach, M.A.; Bartussek, J.; et al. Bacterial Pulmonary Superinfections Are Associated with Longer Duration of Ventilation in Critically Ill COVID-19 Patients. Cell Rep Med 2021, 2, 100229. [Google Scholar] [CrossRef]
- Labiris, N.R.; Dolovich, M.B. Pulmonary Drug Delivery. Part II: The Role of Inhalant Delivery Devices and Drug Formulations in Therapeutic Effectiveness of Aerosolized Medications. Br J Clin Pharmacol 2003, 56, 600. [Google Scholar] [CrossRef]
- A, O.-M.; OJ, H.; D, T.; A, G.; L, A.; A, R.-H. The Use of ROC Analysis for the Qualitative Prediction of Human Oral Bioavailability from Animal Data. Pharm Res 2014, 31, 720–730. [Google Scholar] [CrossRef]
- Price, G.; Patel, D.A. Drug Bioavailability. xPharm: The Comprehensive Pharmacology Reference 2020, 1–2. [Google Scholar]
- Strong, P.; Ito, K.; Murray, J.; Rapeport, G. Current Approaches to the Discovery of Novel Inhaled Medicines. Drug Discov Today 2018, 23, 1705–1717. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Nicolau, D. V; Langford, B.; Mahdi, M.; Jeffers, H.; Mwasuku, C.; Krassowska, K.; Fox, R.; Binnian, I.; Glover, V.; et al. Inhaled Budesonide in the Treatment of Early COVID-19 (STOIC): A Phase 2, Open-Label, Randomised Controlled Trial. Lancet Respir Med 2021, 9, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Rathod, R.; Mohindra, R.; Vijayakumar, A.; Soni, R.K.; Kaur, R.; Kumar, A.; Hegde, N.; Anand, A.; Sharma, S.; Suri, V.; et al. Essential Oil Nebulization in Mild COVID-19(EONCO): Early Phase Exploratory Clinical Trial. J Ayurveda Integr Med 2022, 13, 100626. [Google Scholar] [CrossRef] [PubMed]
- Amirav, I.; Newhouse, M.T. Transmission of Coronavirus by Nebulizer: A Serious, Underappreciated Risk. CMAJ : Canadian Medical Association Journal 2020, 192, E346. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M.; Ora, J.; Bianco, A.; Rogliani, P.; Matera, M.G. Guidance on Nebulization during the Current COVID-19 Pandemic. Respir Med 2021, 176, 106236. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of SARS-CoV-2 (a) structure, (b) genome encoding for structural, non-structural, and accessory proteins, and (c) replication cycle. ACE2= angiotensin-converting enzyme 2, pp= polyprotein, RBD= receptor-binding domain, RTC= replication and transcription complex, SPs= structural proteins, ssRNA= single stranded RNA, TMPRSS2= transmembrane serine protease 2, S= spike, N= neucleocapsid, M= membrane, E= envelop, nsp= nonstructural proteins, PLpro= papain-like protease, 3CLpro= chymotrypsin-like protease, RdRp= RNA-dependent RNA polymerase, MTase= methyltransferase, ExoN= exonuclease, EndoN= endonuclease (Figure drawn by biorender).
Figure 1.
Schematic diagram of SARS-CoV-2 (a) structure, (b) genome encoding for structural, non-structural, and accessory proteins, and (c) replication cycle. ACE2= angiotensin-converting enzyme 2, pp= polyprotein, RBD= receptor-binding domain, RTC= replication and transcription complex, SPs= structural proteins, ssRNA= single stranded RNA, TMPRSS2= transmembrane serine protease 2, S= spike, N= neucleocapsid, M= membrane, E= envelop, nsp= nonstructural proteins, PLpro= papain-like protease, 3CLpro= chymotrypsin-like protease, RdRp= RNA-dependent RNA polymerase, MTase= methyltransferase, ExoN= exonuclease, EndoN= endonuclease (Figure drawn by biorender).
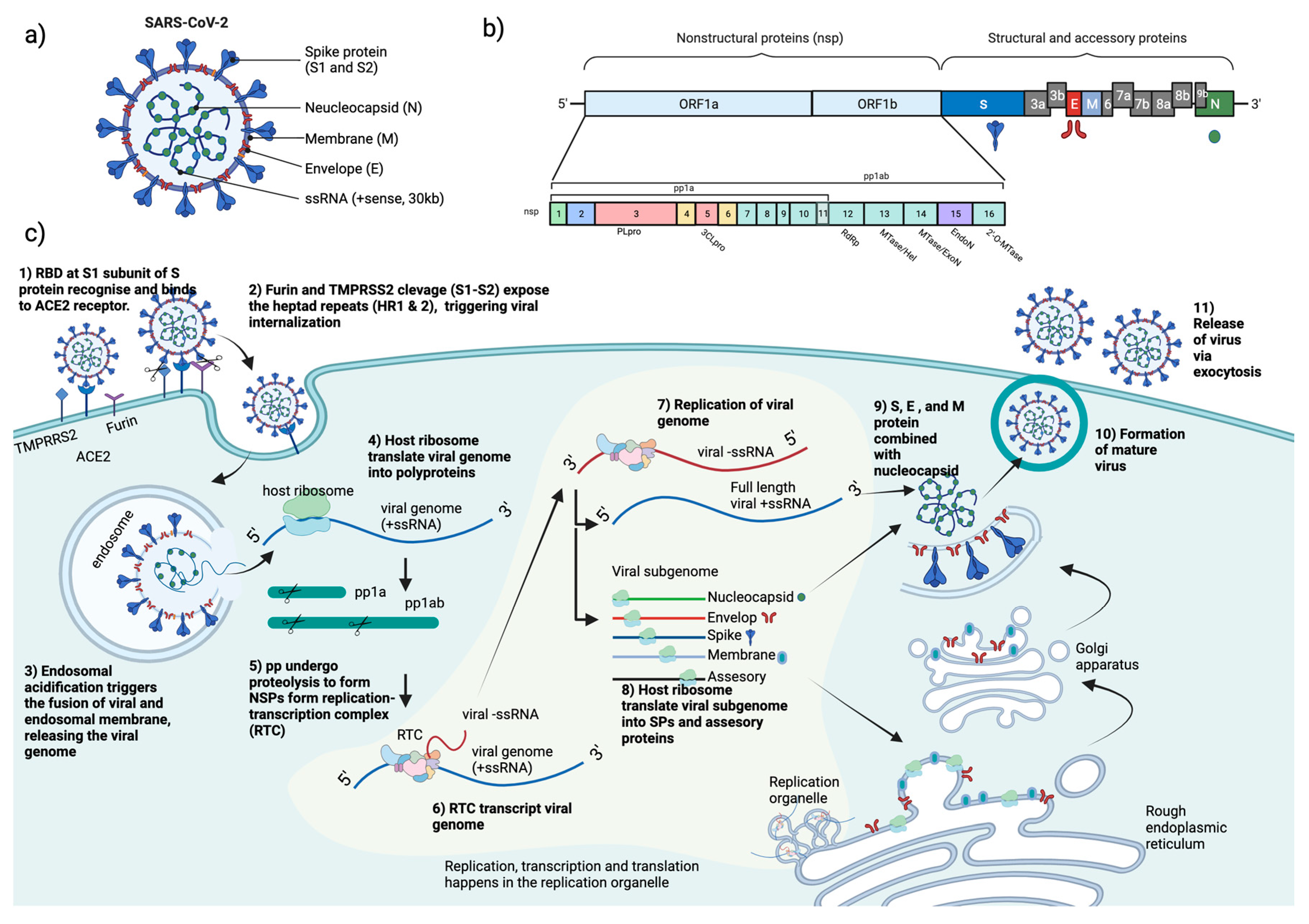
Figure 2.
The antiviral activity of natural compounds, including natural extracts and their active ingredients in the life cycle of SARS-CoV-2. Natural compounds reported against SARS-CoV-2 were categorized according to the mechanism of inhibition at different stages of the SARS-CoV-2 life cycle. Natural compounds affecting uncoating have not been reported. RTC= Replication-transcription complex, +ssRNA= positive single-stranded ribonucleic acid, -ssRNA= negative single-stranded ribonucleic acid TMPRRS2= Transmembrane serine protease 2, ACE2= angiotensin-converting enzyme 2, pp= polypeptide, NF-kB= Nuclear factor-kB, MAPK= Mitogen-activated protein kinase, PPE= pomegranate peel extracts, PUG= punicalagin EGCG= epigallocatechin gallate LHQW=Lianhuaqungwen, JHQG=Jinhua Qinggan, XBJ= Xuebijing. (Figure drawn by biorender).
Figure 2.
The antiviral activity of natural compounds, including natural extracts and their active ingredients in the life cycle of SARS-CoV-2. Natural compounds reported against SARS-CoV-2 were categorized according to the mechanism of inhibition at different stages of the SARS-CoV-2 life cycle. Natural compounds affecting uncoating have not been reported. RTC= Replication-transcription complex, +ssRNA= positive single-stranded ribonucleic acid, -ssRNA= negative single-stranded ribonucleic acid TMPRRS2= Transmembrane serine protease 2, ACE2= angiotensin-converting enzyme 2, pp= polypeptide, NF-kB= Nuclear factor-kB, MAPK= Mitogen-activated protein kinase, PPE= pomegranate peel extracts, PUG= punicalagin EGCG= epigallocatechin gallate LHQW=Lianhuaqungwen, JHQG=Jinhua Qinggan, XBJ= Xuebijing. (Figure drawn by biorender).
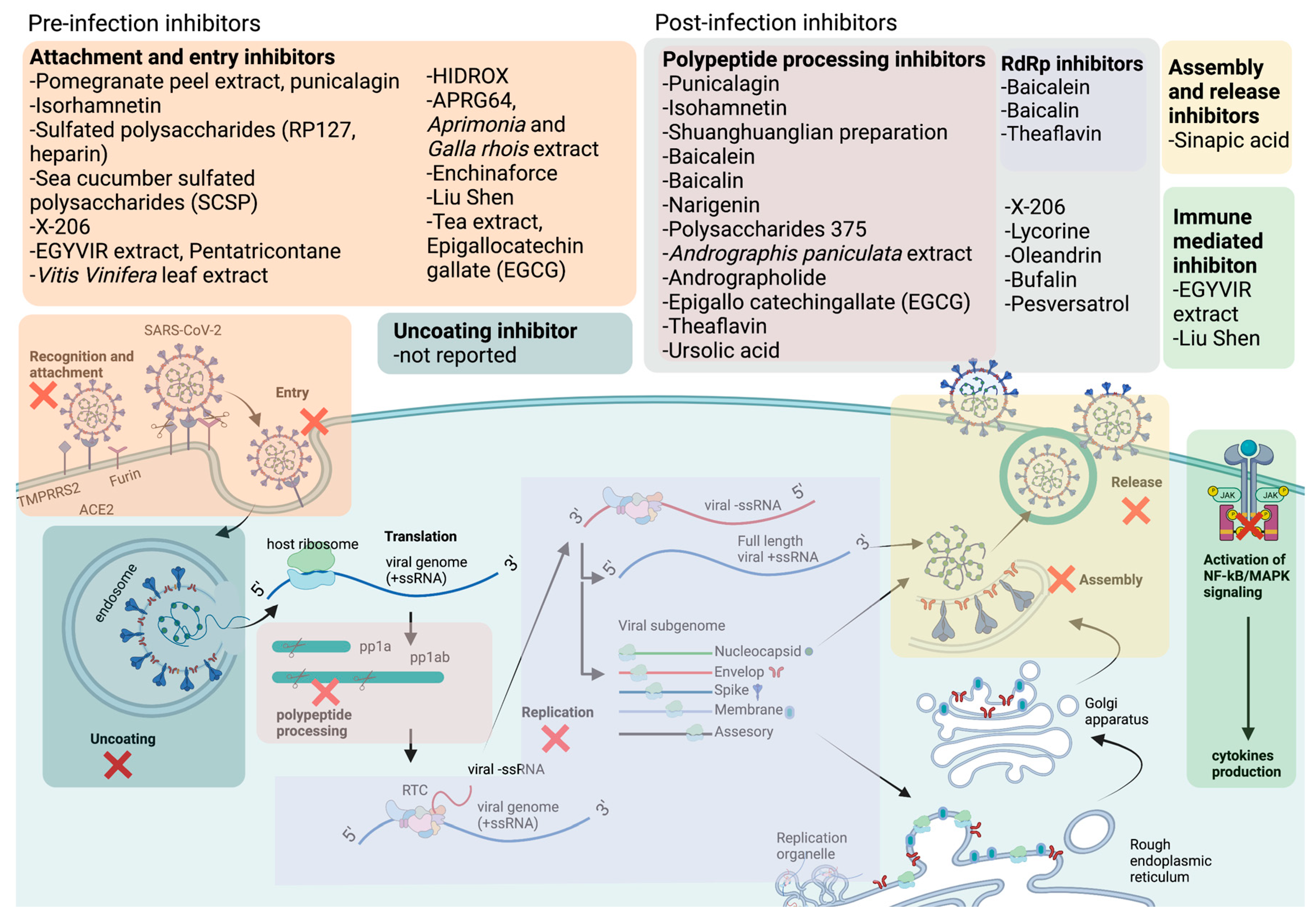
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



