
Submitted:
06 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
Oxidative stress and inflammation are basic pathogenic factors involved in tissue injury, pain, as well as acute and chronic diseases. Since long-term uses of steroids and non-steroidal anti-inflammatory drugs (NSAIDs) cause severe adverse effects, novel effective materials with minimal side effects are required. In this study, polyphenol content and antioxidative activity of rosebud extracts from 24 newly-crossbred Korean roses were analyzed. Among them, Pretty Velvet rosebud extract (PVRE) were found to contain high polyphenols and to show in vitro antioxidative and anti-inflammatory activities. In RAW 264.7 cells stimulated with lipopolysaccharide (LPS), PVRE down-regulated mRNA expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), and thereby decreased nitric oxide (NO) and prostaglandin E2 (PGE2) production. In an air-pouch inflammation model, treatment with PVRE decreased λ-carrageenan-induced tissue exudation, infiltration of inflammatory cells including neutrophils, monocytes and lymphocytes, and tumor necrosis factor-α and interleukin-1β concentrations, as achieved with dexamethasone (a steroid). Notably, PVRE also inhibited PGE2, similar to dexamethasone and indomethacin (an NSAID). The anti-inflammatory effects of PVRE were confirmed by microscopic findings, attenuating tissue erythema, edema, and inflammatory cell infiltration. These results indicate that PVRE exhibits dual (steroid- and NSAID-like) anti-inflammatory activities by blocking both the iNOS ― NO and COX-2 ― PG pathways, and that PVRE could be a promising anti-inflammatory material for diverse issue injuries.
Keywords:
rosebud extract
; Pretty Velvet
; polyphenol
; antioxidation
; anti-inflammation
; nitric oxide
; prostaglandin E2
1. Introduction
Oxidation and oxidative phosphorylation are essential processes in our body for metabolism and energy formation, but oxidative stresses are negative reaction of the imbalance between oxidation and antioxidation caused by over-production of reactive oxygen species (ROS). Excessive ROS could attack the cellular proteins, lipids, and nucleic acids, leading to cellular dysfunction, immune reaction, and inflammation [1,2].
Inflammation is a part of the complex biological responses of vascular tissues to harmful stimuli, such as erythema and edema, which are induced by increased blood flow to the site of injury as well as enhanced vascular permeability resulting in extravasation of macromolecules [3]. Inflammation is a protective attempt by the organism to remove the injurious stimuli and to initiate the healing process [4]. However, if the host fails to eliminate the stimuli early enough or undergoes an improper recovery process, the unresolved inflammation can cause organ dysfunction and become detrimental to the host [5]. Inflammation underlies a wide variety of physiological and pathological processes [6]. Previous reports have provided evidence that inflammation plays an important role in the pathogenesis of acute and chronic diseases [7,8]. Thus, considering that inflammation is the cornerstone of many acute and chronic pathological processes, anti-inflammatory therapy is essential to increase patients’ survival rate and to improve their quality of life.
As an in vitro inflammation model, RAW 264.7 cells, a macrophage cell line, stimulated with lipopolysaccharide (LPS) are commonly used [9,10]. Since during activation of the cells, dual pathways of inflammation, that is, inducible nitric oxide synthase (iNOS) ― nitric oxide (NO) and cyclooxygenase-2 (COX-2) ― prostaglandin E2 (PGE2), are activated [11]. For the screening of anti-inflammatory compounds, the mRNA expression of iNOS and COX-2 as well as the production of NO and PGE2 can be analyzed.
Among many in vivo skin inflammation models, λ-carrageenan-induced air-pouch inflammation is recommended, in which exudate can be easily collected for the analysis of inflammatory cells and mediators in the exudate. Carrageenans are high molecular weight sulphated polygalactans obtained from marine algae [12]. Air pouch is formed by subcutaneous injection of sterile air into the backside of rats or mice, which can be later injected with λ-carrageenan to produce inflammation [13,14]. The pathogenic agent λ-carrageenan induces inflammatory responses in the lining tissue of the pouch wall [15], leading to extravasation of blood cells and fluid into the pouch. Investigators have shown the accumulation of inflammatory cells such as macrophages, white blood cells (WBCs), and lymphocytes, cytokines including tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), as well as mediators such as NO and PGE2 [8,16,17]. These endpoints can be quantified and used to determine the degree of inflammation, resolution of inflammation, or anti-inflammatory and antioxidative activities of test compounds [18].
The TNF-α ― iNOS ― NO and arachidonic acid ― COX-2 ― PGE2 inflammatory pathways are blocked by steroids and non-steroidal anti-inflammatory drugs (NSAIDs), respectively [19]. However, severe adverse effects of such drugs are easily developed [20-22]. Indeed, it is well known that high doses and/or long-term use of synthetic steroids cause tolerance, glaucoma, cataracts, diabetes, osteoporosis, and muscle weakness [22]. NSAIDS can induce gastrointestinal bleeding, congestive heart failure, chronic kidney diseases with renal papillary necrosis, and allergic reactions. Thus, consumers are interested in natural ingredients that are effective, but not come with the side effects of synthetic drugs [23,24].
Roses are dicotyledons belonging to the genus Rosa in the family Rosacea. Plants from the Rosacea family are rich in natural molecules with beneficial biological properties, and they are widely appreciated and used in the food industry, perfumery, and cosmetics [25]. Rose has been confirmed to contain a large amount of antioxidants, and is used as an edible flower in many countries, as well as being used as a cosmetic ingredient [26]. It was found that rose petals contained antioxidant ingredients including vitamins, polyphenols, anthocyanins, flavonoids, and lactones, and thus displaying antioxidative activity higher than other plants [27,28] and comparable to that of butylated hydroxytoluene (BHT), a representative synthetic antioxidant [29-31]. Related to effectiveness, anthocyanins of rose flowers have been reported to be effective in preventing cardiovascular diseases, cancers, and diabetes [32]. In many our studies, it has been confirmed that rosebud extracts have strong antimicrobial potential [33,34], and exhibit anti-allergic, anti-atopic and anti-inflammatory activities [35,36], and antioxidation and anyi-inflammation-related neuroprotective effects [37,38].
Therefore, the purpose of this study is screening of novel functional materials with high antioxdative and anti-inflammatory activities. In this context, polyphenol contents and antioxidative activities of 24 rosebud extracts from newly-crossbred roses were assessed first, and the anti-inflammatory effects of a selected candidate were confirmed in vitro and in vivo using RAW 264.7 macrophages and a λ-carrageenan-induced air-pouch inflammation model, respectively.
2. Materials and Methods
2.1. Extraction of Rosebuds
Dried rosebuds from 24 newly-crossbred Korean roses (Rosa hybrida) were from Gumi Floriculture Research Institute (Gumi, Korea) in 2021, which are 1) Lover Shy, 2) Lovely Scarlet, 3) Loving Heart, 4) Red Perfume, 5) Luminus, 6) Mirinae Gold, 7) Betty, 8) Bichina, 9) Aileen, 10) Onnuri, 11) Yunina, 12) Jaemina Red, 13) Jinseonmi, 14) Chilbaegri, 15) Tamina, 16) Tamnari, 17) Pretty Velvet, 18) Peach Grace, 19) Pink Love, 20) Pink Perfume, 21) Hanaro, 22) Hanaram, 23) Hanggina, and 24) Ice Wing.
The dried rosebuds were pulverized in a rotor mill (Laval Lab Inc., Laval, QC, Canada), immersed in 80% ethanol in an ultrasonic water bath, heated at 60~70°C for 2 hours, and then ultrasonic-extracted for 1 hour. Actually, the extraction solvent : solid ratio was set to 49 : 1 (980 mL 80% ethanol : 20 g dried rosebuds). After extraction, the mixture was cooled at room temperature, filtered, and then concentrated under a reduced pressure to 50 brix using a vacuum evaporator (Rotary Vacuum Evaporator N-N series; Eyela, Tokyo, Japan), and then used as a test sample.
2.2. Analysis of Antioxidant Ingredients
Total polyphenols as a major antioxidant component of plants were measured according to the method of Dewanto et al. (2002) [39]. That is, the contents of polyphenol were measured by the principle that Folin-Ciocalteu's phenol reagent is reduced by the polyphenolic compounds to develop a color of molybdenum. The sample concentration was adjusted to 1 mg/mL. Into 100 μL sample solution, 2 mL of 2% Na2CO3 was added, and reacted for 3 min. After adding 100 μL of 50% Folin-Ciocalteu's phenol reagent and reacting for 30 min, the absorbance was measured at 750 nm using a spectrophotometer (UV-1650; Shimadzu Corporation, Kyoto, Japan). A standard calibration curve was prepared with gallic acid (Sigma-Aldrich, St. Louis, MO, USA) as a standard material diluted 10, 20, 30, 40 and 50 times, and the polyphenol contents were expressed as mg gallic acid equivalent (GAE) in 1 g of extract.
2.3. Measurement of Antioxidative Activities
2.3.1. Analysis of ABTS-scavenging activity
The antioxidant efficacy of rosebud extracts was measured by the scavenging potential of 2,2-azino-bis (3-ethylbenzothiazoline 6-sulfonic acid) (ABTS; Sigma-Aldrich) [40]. A stock ABTS radical cation solution (7 mM) was added to a 2.45 mM potassium and sulfate solution, stirred well for 12 to 16 hours in the dark at room temperature. The solution was diluted with distilled water to obtain an absorbance of 1.4 to 1.5 at 735 nm. A diluted 1 mL ABTS radical cation solution was added to 50 μL of the test substance or distilled water (blank) to adjust to 0.5 mg/mL. After 1 hour, the absorbance was measured at 735 nm. ABTS radical scavenging activity, i.e., electron-donating ability, was expressed as mg ascorbic acid equivalent (AAE) in 1 g of extract with the difference in absorbance according to the addition and non-addition (distilled water) of the test substance.
2.3.2. Analysis of DPPH-scavenging activity
Additional antioxidant efficacy of the rosebud extracts was measured according to the method of Hwang et al. (2006) [41], based on the scavenging potential of 1,1-diphenyl-2-picrylhydrazyl (DPPH; Sigma-Aldrich). That is, into 0.8 mL of 0.2 mM DPPH solution, 0.2 mL of the extract or distilled water (blank) was added to adjust to 0.1 mg/mL concentration. After 30-min incubation at room temperature, absorbance was measured at 520 nm. DPPH radical-scavenging activity, i.e., electron-donating ability, was expressed as mg AAE in 1 g of extract with the difference in absorbance according to the addition and non-addition (distilled water) of the test substance.
2.3.3. Analysis of correlation between antioxidant contents and antioxidative activities
The correlations between antioxidant (polyphenol) contents and antioxidative activities were analyzed. Linear regression coefficients were calculated for the estimation of relationship.
2.4. Measurement of Anti-inflammatory Activities
2.4.1. Analysis of NO-inhibitory activity
RAW 264.7 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultivated with Dulbecco’s Modified Eagle’s Medium (DMEM; Biowest, Kansas City, MO, USA) supplemented with 10% fetal bovine serum (FBS; Biowest).
RAW 264.7 cells were seeded in a 96-well plate in a density of 5 × 105 cells/mL (0.1 mL/well). The cells were incubated with LPS (Sigma-Aldrich; 1 μg/mL) and 100 μg/mL of 24 each rosebud extract for 24 hours. The cell culture supernatants were harvested, and the concentrations of NO were measured by Griess reagent system (Promega, Madison, WI, USA).
2.4.2. Analysis of correlation between antioxidant contents and NO-inhibitory activities
The correlation between antioxidant (polyphenol) contents and NO-inhibitory activity of 24 rosebud extracts was analyzed. Linear regression coefficient was calculated for the estimation of relationship.
Based on the antioxidant contents, antioxidative activities and NO-inhibitory potentials of 24 rosebud extracts, Pretty Velvet rosebud extract (PVRE) was selected as a candidate for further studies on anti-inflammatory effects in vitro and in vivo, and underlying mechanisms.
2.5. Measurement of In Vitro Anti-inflammatory Activity of PVRE
2.5.1. MTT assay for cytotoxicity
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) was purchased from Sigma-Aldrich (98% purity). Briefly, MTT powder was dissolved in Dulbecco’s phosphate- buffered saline (D-PBS; Biowest) to a final concentration of 5 mg/mL (MTT stock solution).
RAW 264.7 cells were seeded in a 96-well plate at a density of 5 × 105 cells/mL (0.1 mL/well). The cells were incubated with various concentrations (10, 30 or 100 μg/mL) of PVRE for 24 hours. Into each cell culture well (0.1 mL/well), 100 μL of DMEM medium containing 10% MTT stock solution (MTT medium) was added, and incubated under 5% CO2 at 37°C for further 1 hour. The MTT medium was then discarded, and the formazan produced in the cells was extracted with 100 μL of dimethylsulfoxide (DMSO). The absorbance was read at 540 nm.
2.5.2. qPCR analysis of mRNA expressions of iNOS and COX-II
RAW 264.7 cells were seeded into a 24-well plate (5 × 105 cells/mL) and incubated under 5% CO2 at 37°C overnight. The cells were incubated with LPS (1 μg/mL) and various concentrations (10, 30 or 100 μg/mL) of PVRE for 24 hours. Total RNA was extracted using 1 mL of RNAiso PLUS (TaKaRa, Shiga, Japan). Then, 1 μg of total RNA was reverse transcribed to cDNA. Real-time qPCR samples were prepared with a PrimeScriptTM RT reagent Kit with gDNA Eraser (TaKaRa). The primer sequences for iNOS, COX-II, and GAPDH are described in Table 1. qPCR was performed with a AriaMx Real-Time PCR system (Agilent Technologies, Santa Clara, CA, USA) using a two-step protocol (40 cycles of 95°C for 30 sec followed by 60°C for 30 sec).
2.5.3. Chemical analysis of nitric oxide and prostaglandin E2
After collection of RAW 264.7 cells for the analysis of mRNA expression, the culture supernatant was harvested. The concentrations of NO and PGE2 were measured using the Griess reagent and enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA), respectively.
2.6. Measurement of In Vivo Anti-inflammatory Activity of PVRE
2.6.1. Animals
Six-week-old male Sprague-Dawley (SD) rats were purchased from DBL (Eumsung, Korea). The animals were housed at a room with a constant temperature (23 ± 2°C), relative humidity of 55 ± 10%, and 12-hour light/dark cycle, and fed with standard rodent chow and purified water ad libitum
All animal experimental procedures were approved and carried out in accordance with the Institutional Animal Care and Use Committee of Laboratory Animal Research Center at Chungbuk National University, Korea (Approval No. CBNUR-1629-21).
2.6.2. Experimental design using an air-pouch inflammation model
A λ-carrageenan-induced air-pouch model of acute inflammation was established in rats. Briefly, rats were anesthetized with isoflurane. Then, 20 mL of sterile air was injected subcutaneously into the mid-back side using a syringe. On the 5th and 6th days, 10 mL of sterile air was re-injected at the same site to maintain the size of the air pouch. Twenty four hours later, inflammation was induced by injecting 1 mL of λ-carrageenan solution (2% in D-PBS) into the pouch. Test sample (PVRE; 10, 30 or 100 mg/kg), dexamethasone (Sigma-Aldrich; 2 mg/kg) or indomethacin (Sigma-Aldrich; 2 mg/kg) were administrated intraperitoneally 30 min before λ-carrageenan injection. Six hours after λ-carrageenan injection, the exudate in the pouch was collected. An aliquot of the exudate was analyzed for inflammatory cells. Remaining exudate was centrifuged at 10,000 g at 4°C for 10 min, and the supernatant was used for the analysis of inflammatory mediators. The centrally-positioned skin tissues of the pouches were excised for Image-J analysis and histopathological examination.
2.6.3. Analysis of exudate volume and inflammation cells
After sacrifice of the rats, the air-pouch exudate was collected. Briefly, 2 mL of cold saline (0.9% NaCl) was injected into the pouch and the pouch was gently massaged. All the fluid was harvested using a syringe, the volume was recorded, and net exudate was calculated by subtracting 2 mL saline injected.
The exudate was analyzed for total white blood cells (WBCs), neutrophils, monocytes, and lymphocytes with a hematology analyzer (IDEXX Procyte Dx, Westbrook, CT, USA).
2.6.4. ELISA analysis of inflammatory cytokines
The levels of TNF-α and IL-1β in air-pouch exudate were measured using ELISA kits (R&D Systems) according to the manufacturer’s instructions.
2.6.5. Chemical analysis of NO and PGE2
The concentrations of NO and PGE2 in air-pouch exudate were measured using the Griess reagent (Sigma-Aldrich) and ELISA kit (R&D Systems), respectively, as described above.
2.6.6. Image-J analysis of dermal blood vessel area
After the rats were sacrificed, the back skin was cut off in the same area and photographed, and the area of blood vessels on the back skin were observed and calculated by Image-J software (NIH, Bethesda, MD, USA).
2.6.7. Microscopic examination of tissue inflammation
Skin tissues obtained from the pouches were fixed with 10% neutral formalin, followed by tissue processing and paraffin embedding. Paraffin blocks were sectioned (4 μm in thickness) and stained with hematoxylin-eosin. Histopathological examination was then performed under a light microscope (Nikon, Tokyo, Japan).
2.7. Statistical Analysis
The data were described as mean ± standard error. Statistical significance between the groups was analyzed by one-way analysis of variance using the SPSS statistical software (SPSS Inc., Chicago, IL, USA). P-values of less than 0.05 were considered statistically significant.
3. Results
3.1. Antioxidant Contents in 24 Rosebud Extracts
As a major antioxidant ingredient, the contents of polyphenols in 24 rosebud extracts tested were in the range of 111.7 to 385.5 mg GAE/g extract. In comparison, Lover Shy (385.5 ± 0.8 mg GAE/g extract), Pretty Velvet (372.5 ± 1.5 mg GAE/g extract), Ice Wing (338.4 ± 1.3 mg GAE/g extract), Red Perfume (335.2 ± 2.4 mg GAE/g extract), Onnuri (316.2 ± 0.8 mg GAE/g extract), Hanggina (296.6 ± 1.3 mg GAE/g extract), and Jaemina Red (295.2 ± 0.1 mg GAE/g extract) showed relatively high polyphenol contents (Figure 1A). Especially, Lover Shy and Pretty Velvet extracts were found to contain the highest antioxidant concentrations.
3.2. Antioxidant Activities of 24 Rosebud Extracts
As shown in Figure 1B, among the 24 rosebud extracts tested (100 μg/mL), the high ABTS radical-scavenging activities were confirmed in Lover Shy (425.0 ± 21.0 mg/AAE/g extract), Pretty Velvet (420.2 ± 22.3 mg/AAE/g extract), Red Perfume (360.1 ± 20.7 mg/AAE/g extract), Jaemina Red (345.4 ± 38.2 mg/AAE/g extract), Onnuri (330.3 ± 45.2 mg/AAE/g extract), Ice Wing (325.3 ± 29.9 mg/AAE/g extract), and Hanggina (310.0 ± 36.8 mg/AAE/g extract).
Similarly, high DPPH radical-scavenging activities were also observed in Lover Shy (420.3 ± 24.8 mg/AAE/g extract), Pretty Velvet (405.4 ± 35.2 mg/AAE/g extract), Onnuri (365.4 ± 43.9 mg/AAE/g extract), Red Perfume (345.1 ± 24.6 mg/AAE/g extract), Jaemina Red (330.2 ± 38.0 mg/AAE/g extract), Ice Wing (295.4 ± 30.7 mg/AAE/g extract), and Hanggina (285.1 ± 23.3 mg/AAE/g extract) (Figure 1C).
Collectively, Lover Shy and Pretty Velvet were found to have the highest antioxidant activities against ABTS and DPPH.
3.3. Inhibitory Activities of 24 Rosebud Extracts on NO Production
NO produced from LPS-stimulated RAW 264.7 cells was analyzed with Griess reagent. As seen in Figure 1D, LPS increased the NO concentration up to 51 μM. However, all the 24 rosebud extracts at 100 μg/mL displayed NO-inhibitory effects. It was found that Jaemina Red (82.4%), Pretty Velvet (80.4), Hanggina (78.4%), Red Perfume (68.6%), Onnuri (66.7%), Ice Wing (64.7%), and Lover Shy (58.8%) have high NO-inhibitory activity, and especially, Jaemina Red, Pretty Velvet, and Hanggina were the most effective.
3.4. Correlation between Polyphenols and Antioxidative Activities
The correlations between the antioxidant contents and antioxidant efficacies are shown in Figure 2. As seen in Figure 2A, the correlation between polyphenols and ABTS radical-scavenging activity was R² = 0.9173, indicating a high relationship that the more antioxidants, the better the ABTS-scavenging ability. In parallel with the relationship in ABTS-scavenging potential, the correlation between polyphenols and DPPH radical-scavenging activity was very high (R² = 0.8732), too (Figure 2B).
Therefore, it was confirmed that polyphenols are the main antioxidants in the 24 rosebud extracts, and that among them, Lover Shy, Red Perfume, Onnuri, Jaemina Red, Pretty Velvet, Hanggina, and Ice Wing have the highest antioxidative efficacies.
3.5. Correlation between Polyphenols and NO-Inhibitory Activities
The correlation between the antioxidant contents (polyphenols) and NO-inhibition activities was R²= 0.5773 (Figure 2C), indicating a high relationship that the more antioxidants, the better the NO-inhibitory capacity.
Collectively, Jaemina Red, Pretty Velvet, and Hanggina were found to have the highest anti-inflammatory activity.
3.6. Cytotoxicity of PVRE in RAW 264.7 Cells
In order to assess the cytotoxicity of PVRE itself, RAW 264.7 cells were treated with various concentrations (0.3 - 100 μg/mL), and 24 hours later, the cell viability was measured via MTT assay. PVRE did not exerted cytotoxicity up to 100 μg/mL, rather facilitated proliferation at 10 - 30 μg/mL (Figure 3A). Accordingly, PVRE was used at non-toxic concentrations of 10, 30, and 100 μg/mL in following studies for anti-inflammatory activities.
3.7. Effects on iNOS and COX-II Expressions in RAW 264.7 Cells
To elucidate underlying mechanisms of anti-inflammatory activity of PVRE, RAW 264.7 cells were treated with LPS (1 μg/mL) and various concentrations (10, 30 or 100 μg/mL) of PVRE, and the mRNA expressions of iNOS and COX-2 were analyzed via qPCR. LPS treatment greatly increased the iNOS mRNA expression (Figure 3B). Such LPS-induced increase in iNOS expression was significantly attenuated by PVRE treatment in a concentration-dependent manner, suggestive of a steroid-like effect. COX-2 mRNA expression in RAW 264.7 cells was also markedly increased after exposure to LPS (Figure 3C). COX-2 expression was down-regulated following treatment with PVRE in a concentration-dependent manner, too, indicating that PVRE possesses an NSAID-like activity.
3.8. Effects on NO and PGE2 Production in RAW 264.7 Cells
Inflammatory mediators NO and PGE2 produced from LPS-activated RAW 264.7 cells were analyzed with Griess reagent and ELISA kit, respectively. LPS increased the NO concentration up to 64.0 μM from 1.8 μM in untreated resting cells (Figure 3D). Notably, the LPS-induced production of NO was significantly inhibited by PVRE treatment up to 46.9% of peak value, similarly to a steroid’s effect. PGE2 production from RAW 264.7 cells was also markedly enhanced after exposure to LPS (Figure 3E). However, such LPS-mediated increase in PGE2 production was markedly lowered by treatment with PVRE up to 32.4% of peak value, indicative of an NSAID-like property.
3.9. Effect on Exudation in Air-Pouch Inflammation
Inflammation causes extravasation of the blood cells and plasma components as well as leakage of tissue fluid. Body fluid transports diverse blood and tissue ingredients such as proteins, inflammatory cells, and inflammatory mediators into the inflamed area. As shown in Figure 4A. the λ-carrageenan-induced inflammation greatly increased the exudate volume in the air-pouch. However, the increased exudation was potentially attenuated by treatment with dexamethasone (a synthetic steroid) and indomethacin (a representative NSAID). Notably, PVRE inhibited the vascular exudation, especially at 100 mg/kg, it significantly reduced the exudate volume comparable with dexamethasone and indomethacin (2 mg/kg).
3.10. Effect on Inflammatory Cells in Exudate
Total WBCs including neutrophils as well as monocytes and lymphocytes markedly increased in the exudate from the air-pouch inflammation (Figure 4B-E). The inflammatory cell infiltration was significantly inhibited by both dexamethasone and indomethacin, although dexamethasone was superior to indomethacin. Interestingly, PVRE (10-100 mg/kg) remarkably inhibited the infiltration of all types of inflammatory cells in a dose-dependent manner, and notably to the levels achieved with dexamethasone (2 mg/kg) at 30-100 mg/kg.
3.11. Effect on Inflammatory Cytokine Concentration in Exudate
The major inflammatory cytokines TNF-α and IL-1β in the inflamed exudate greatly increased (Figure 5A,B). Both the cytokines increased by λ-carrageenan challenge was significantly inhibited only by dexamethasone, but not by indomethacin. Notably, TNF-α and IL-1β were also inhibited by PVRE in a dose-dependent manner, suggestive of steroid-like activity of PVRE.
3.12. Effects on NO and PGE2 Concentrations in Exudate
In parallel with the inflammatory cytokines, the concentrations of NO and PGE2 in the exudate following λ-carrageenan exposure markedly increased (Figure 5C,D). The NO accumulation was attenuated by treatment with dexamethasone. As inferred from the inhibition of cytokines, the NO was also significantly suppressed by PVRE. Although TNF-α, IL-1β, and NO were not affected by indomethacin, PGE2 was remarkably inhibited by both dexamethasone and indomethacin. It was also lowered by PVRE in a dose-dependent manner, suggesting that PVRE have both steroid- and NSAID-like activities.
3.13. Effect on Blood Vessel Area
Inflammation occurs along with vasodilatation, leading to erythema and edema. The dermal blood vessel dilatation was observed in the skin (Figure 6A). Accordingly, the blood vessel area measured with Image-J increased more than 6 times, which was significantly attenuated by dexamethasone (Figure 6B). Interestingly, PVRE (100 mg/kg) exhibited a potential effect similar to that of dexamethasone (2 mg/kg).
3.14. Microscopic Findings
From the histopathological examination of the tissues surrounding the λ-carrageenan-exposed air-pouches, severe infiltration of many inflammatory cells into the dermal and subcutaneous tissues was observed (Figure 7A). In addition, blood vessel dilatation as well as edema in the dermal and muscular layers were also seen, as confirmed by the thickness of lining and muscle layers (Figure 7B). The inflammatory responses were inhibited by dexamethasone and indomethacin. Notably, PVRE also exerted strong anti-inflammatory activity in a dose-dependent manner, wherein PVRE’s effect at 100 mg/kg was comparable to dexamethasone and superior to indomethasone (2 mg/kg).
4. Discussion
Most of the animals consuming oxygen for metabolism suffer from tissue damage, called oxidative stress, and aging due to the generation of ROS [42]. Organisms synthesize or ingest antioxidants in the body to block cell and tissue damages caused by ROS. If oxidative stress in the body goes beyond the antioxidant defense capacity that counteracts it, theoretically, supplementation with antioxidants can limit oxidative damage. As such, the search for a compound capable of scavenging ROS or a substance that inhibits the formation of oxides is being actively conducted. Such compounds mainly containing a polyphenol structure are rich in plants, called phytochemicals which are largely divided into phenolics, carotenoids, alkaloids, organosulfur derivatives, and nitrogen-containing compounds [30].
ABTS and DPPH radical-scavenging activities are common tools for the rapid assessment of antioxidative potentials of diverse natural products [43]. In the present study, the polyphenol contents and antioxidant activities of 24 rosebud extracts showed high relationships, and among them, Lover Shy and Pretty Velvet were found to be the highest in both antioxidant ingredients and their activities.
Tissue damage caused by ROS is involved in the most inflammatory responses. Inflammation is the basic mechanism available for tissue repair after an injury and consists of a cascade of cellular and microvascular reactions. Therefore, the anti-inflammatory effects of the 24 rosebud extracts were evaluated in LPS-stimulated RAW 264.7 macrophages, from which Pretty Velvet, Jaemina Red, and Hanggina displayed the highest NO-inhibitory activities. Thus, PVRE possessing the highest antioxidative and anti-inflammatory potentials was selected as a candidate for further studies on therapeutic effects and underlying mechanisms in vitro and in vivo.
The process of inflammation is accompanied by exudation, infiltration of inflammatory cells, and production of cytokines and inflammatory mediators. The accumulation of circulating leukocytes and monocytes, followed by lymphocytes, is one of the most visible signs of inflammation [6,44]. In this process, an important role has been attributed to the release of chemo-attractants by resident cells inducing the inflammatory cell recruitment to the inflammatory focus [45]. Inflammatory cytokines are produced predominantly by activated macrophages, and mediate upregulation of inflammatory reactions [46]. Especially, certain inflammatory cytokines such as TNF-α and IL-1β are involved in the process of pathologic pain [47,48]. Although NO produced by constitutive NOS (cNOS) is an important cellular signaling molecule that participates in diverse physiological functions in mammals, excessive production of NO from cytokine- or bacterial endotoxin-activated iNOS cause acute pain, edema, and even shock [49,50]. It has been well defined that TNF-α ― iNOS ― NO pathway is controlled by steroids [9,10]. Interestingly, PVRE strongly inhibited the iNOS mRNA expression, and ensuing NO production in LPS-stimulated RAW 264.7 macrophages. From such phenomena, it is inferred that PVRE has steroid-like action mechanism.
PGs produced from arachidonic acid by enzyme COX-1 are important physiological intrinsic regulators for blood blow, gastric mucosal protection, and renal and uterine functions. Among diverse PGs, PGE2 synthesis is affected by the expression of COX-2, levels of which are highly inducible in many tissues by inflammatory factors including cytokines and growth factors. Thus, PGE2 is an important lipid mediator of inflammatory and immune responses during acute and chronic infections [51]. By comparison with the TNF-α ― iNOS ― NO pathway, arachidonic acid ― COX-2 ― PGE2 pathway is regulated by NSAIDs. In the present study, the COX-2 expression and PGE2 production were markedly inhibited by PVRE, indicative of an NSAID-like activity of PVRE. Based on the dual effects of PVRE on NO and PGE2 regulation, it was expected that PVRE could be a potential candidate for anti-inflammation. So, further studies in vivo were conducted.
In vivo air-pouch inflammation, λ-carrageenan caused vascular dilatation (erythema) and dermal edema, resulting in the great increase in exudate volume, which are indicative of vascular leakage of serum components [52]. The vasodilatation and exudation were effectively attenuated by dexamethasone (a steroid), indomethacin (an NSAID), and PVRE. In addition, the huge increases in inflammatory cells (WBCs, neutrophils, monocytes, and lymphocytes), cytokines (TNF-α and IL-1β), and NO in the exudate was also markedly inhibited by dexamethasone and PVRE, but not by indomethacin. It is of interest to note that λ-carrageenan induces inflammatory cell infiltration and activation to release cytokines, iNOS expression, and NO production, and that PVRE blocks the cell chemo-attraction, activation, and iNOS expression similarly to the steroids’ activity.
λ-Carrageenan greatly increased the PGE2 concentration in the exudate indicating the activation of COX-2 inflammatory pathway, which was inhibited by indomethacin as well as PVRE. Therefore, it was confirmed that PVRE has both steroid- and NSAID-like activities. Separately, the PGE2-reducing activity of dexamethasone may be due to the inhibitory effect of steroids on the arachidonic acid-degrading phospholipase A2 (PLA2) [53].
The anti-inflammatory effects of PVRE on vascular exudation, edema, and inflammatory cell infiltration were confirmed in microscopic examinations. The dermal tissues exposed to λ-carrageenan exhibited severe vasodilatation, increasing the blood vessel area under Image-J analysis, as well as tissue edema, increasing the dermal and muscular layer thickening. In addition, severe infiltration of inflammatory cells was observed in the dermal and subcutaneous tissues. Interestingly, such inflammation was substantially suppressed by PVRE and dexamethasone, which were superior to indomethacin. Such phenomena indicate that λ-carrageenan-induced vascular response and inflammatory cell infiltration are mainly mediated by TNF-α ― iNOS ― NO pathway.
From the present study results, several rosebud extracts were found to contain high concentrations of antioxidant ingredients, and exhibited close relationships with antioxidative and NO-inhibitory activities. At the same time, PVRE inhibited inflammatory reactions in RAW 264.7 macrophages by down-regulating iNOS and COX-2 mRNA expression. In addition, PVRE decreased the λ-carrageenan-induced air-pouch inflammation in vivo: i.e., it blocked vascular leakage, inflammatory cell infiltration, and inflammatory cytokine and mediator production in dual mechanisms on TNF-α ― iNOS ― NO and arachidonic acid ― COX-2 ― PGE2 pathways, which are controlled by steroids and NSAIDs, respectively.
Therefore, the results indicate that rosebud extracts containing large amounts of antioxidants, including PVRE, from newly-crossbred roses could be candidates for the development of antioxidative and anti-inflammatory functional products. In addition, it is expected that novel medicinal plants through cross-breeding could be produced in the near future.
Author Contributions
C.W., I.-J.K., H.-R.S. and C.H.N. performed cell and animal experiments; S.P., K.Y.K. and S.T.K. prepared test materials; T.M.K. and H.S.J analyzed ingredients; H.-G.Y., S.C.K., E.-K.C. and Y.-B.K. conceived/designed the study and wrote paper. All authors read approved the final manuscript.
Funding
This result was supported by "Regional Innovation Strategy (RIS)" through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (MOE) (2021RIS-001).
Acknowledgments
The authors express special thanks to Gumi Floriculture Research Institute for providing rosebud samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: pathogenesis and therapeutic strategies. Life Sci 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Dong, F. The relevant targets of anti-oxidative stress: a review. J Drug Target 2021, 29, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: novel pro-resolving lipid mediators in resolution. Semin Immunol 2015, 27, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Nonato, F.R.; Santana, D.G.; De Melo, F.M.; Dos Santos, G.G.L.; Brustolim, D.; Camargo, E.A.; De Sousa, D.P.; Soares, M.B.P.; Villarreal, C.F. Anti-inflammatory properties of rose oxide. Int Immunopharmacol 2012, 14, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.; Wang, B.; Lv, L.L.; Liu, B.C. Extracellular vesicle-based nanotherapeutics: emerging frontiers in anti-inflammatory therapy. Theranostics 2020, 10, 8111–8129. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Schönbein, G.W. Analysis of inflammation. Annu Rev Biomed Eng 2006, 8, 93–151. [Google Scholar] [CrossRef]
- Tsai, D.S.; Huang, M.H.; Tsai, J.C.; Chang, Y.S.; Chiu, Y.J.; Lin, Y.C.; Wu, L.Y.; Peng, W.H. Analgesic and anti-inflammatory activities of Rosa taiwanensis Nakai in mice. J Med Food 2015, 18, 592–600. [Google Scholar] [CrossRef]
- Liu, Y.; Su, W.W.; Wang, S.; Li, P.B. Naringgin inhibits chemokine production in an LPS-induced RAW 264.7 macrophage cell line. Mol Med Rep 2012, 6, 1343–1350. [Google Scholar] [CrossRef]
- Cheng, L.; Ren, Y.; Lin, D.; Peng, S.; Zhong, B.; Ma, Z. The anti-inflammatory properties of Citrus wilsonii Tananka extract in LPS-induced RAW 264.7 and primary mouse bone marrow-derived dendritic cells. Molecules 2017, 22, 1213. [Google Scholar] [CrossRef]
- Shin, S.; Joo, S.S.; Jeon, J.H.; Park, D.; Jang, M.J.; Kim, T.O.; Kim, H.K.; Hwang, B.Y.; Kim, K.Y.; Kim, Y.B. Anti-inflammatory effects of a Houttuynia cordata supercritical extract. J Vet Sci 2010, 11, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.W.; Horne, C.H.W. Toxicity of various carrageenans in the mouse. Br J Exp Pathol 1976, 57, 455–459. [Google Scholar] [PubMed]
- Edwards, J.C.W.; Sedgwick, A.D.; Willoughby, D.A. The formation of a structure with the features of synovial lining by subcutaneous injection of air: an in vivo tissue culture system. J Pathol 1981, 134, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Vandooren, J.; Berghmans, N.; Dillen, C.; Van Aelst, I.; Ronsse, I.; Israel, L.L.; Rosenberger, I.; Kreuter, J.; Lellouche, J.P.; Michaeli, S.; et al. Intradermal air pouch leukocytosis as an in vivo test for nanoparticles. Int J Nanomedicine 2013, 8, 4745–4756. [Google Scholar] [CrossRef] [PubMed]
- Claxson, A.; Grootveld, M.; Chander, C.; Earl, J.; Haycock, P.; Mantle, M.; Williams, S.R.; Silwood, C.J.L.; Blake, D.R. Examination of the metabolic status of rat air pouch inflammatory exudate by high field proton NMR spectroscopy. Biochim Biophys Acta 1999, 1454, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Nijampurkar, B.; Qureshi, F.; Jain, N.; Banerjee, T.; Kumar, A.; Parmar, H.S. Anti-inflammatory role of thyroid hormones on rat air pouch model of inflammation. Inflamm Allergy Drug Targets 2015, 14, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Eteraf-Oskouei, T.; Shafiee-Khamneh, A.; Heshmati-Afshar, F.; Delazar, A. Anti-inflammatory and anti-angiogenesis effect of bee pollen methanolic extract using air pouch model of inflammation. Res Pharm Sci 2020, 15, 66–75. [Google Scholar] [CrossRef]
- Fehrenbacher, J.C.; McCarson, K.E. Models of inflammation: carrageenan air pouch. Curr Protoc 2021, 1, e183. [Google Scholar] [CrossRef]
- Dinarello, C.A. Anti-inflammatory agents: Present and future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, S.H. Risk and benefit of steroid therapy. J Korean Soc Intern Med 2009, 77, 298–303. [Google Scholar]
- Eteraf-Oskouei, T.; Mirak, S.M.; Najafi, M. Anti-inflammatory and anti-angiogenesis effects of verapamil on rat air pouch inflammation model. Adv Pharm Bull 2017, 7, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Cho, S.S.; Li, Y.C.; Bae, C.S.; Park, K.M.; Park, D.H. Anti-inflammatory effect of Curcuma longa and Allium hookeri co-treatment via NF-κB and COX-2 pathways. Sci Rep 2020, 10, 5718. [Google Scholar] [CrossRef] [PubMed]
- Boukhatem, M.N.; Kameli, A.; Ferhat, M.A.; Saidi, F.; Mekarnia, M. Rose geranium essential oil as a source of new and safe anti-inflammatory drugs. Libyan J Med 2013, 8, 22520. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Park, Y.L.; Song, A.Y.; Kim, W.G.; Je, C.Y.; Jung, D.H.; Kim, Y.J.; Oh, J.; Cho, J.Y.; Kim, D.J.; Park, J.H. Water extract of Artemisia scoparia Waldst. & Kitam suppresses LPS-induced cytokine production and NLRP3 inflammasome activation in macrophages and alleviates carrageenan-induced acute inflammation in mice. J Ethnopharmacol 2021, 268, 113606. [Google Scholar] [PubMed]
- Mileva, M.; Ilieva, Y.; Jovtchev, G.; Gateva, S.; Zaharieva, M.M.; Georgieva, A.; Dimitrova, L.; Dobreva, A.; Angelova, T.; Vilhelmova-Ilieva, N.; Valcheva, V.; Najdenski, H. Rose flowers — a delicate perfume or a natural healer? Biomolecules 2021, 11, 127. [Google Scholar] [CrossRef]
- Gorji, A. Pharmacological treatment of headache using traditional Persian medicine. Trends Pharmacol Sci 2003, 24, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J Nutr 2003, 133, 2812–2819. [Google Scholar] [CrossRef]
- Cho, E.K.; Son, J.Y.; Kang, K.O. Antioxidant activities of rose, camellia and cockscomb flower extracts. Food Serv Indust J 2015, 11, 21–33. [Google Scholar]
- Lee, H.R.; Lee, J.M.; Choi, N.S.; Lee, J.M. The anti-oxidative and anti-microbial ability of ethanol extracts from Rosa hybrida. Korean J Food Sci Technol 2003, 35, 373–378. [Google Scholar]
- Park, D.; Jeon, J.H.; Kwon, S.C.; Shin, S.; Jang, J.Y.; Jeong, J.H.; Lee, H.S.; Kim, D.I.; Kim, Y.B.; Joo, S.S. Antioxidative activities of white rose flower extract and pharmaceutical advantages of its hexane fraction via free radical scavenging effects. Biochem Cell Biol 2009, 87, 943–952. [Google Scholar] [CrossRef]
- Seong, H.; Heo, J.; Lee, K.H.; Lee, Y.B.; Kim, Y.B.; Han, N.S. Enhancing the antioxidant activities of wines by addition of white rose extract. J Microbiol Biotechnol 2017, 27, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Konczak, I.; Zhang, W. Anthocyanins-more than nature’s colours. J Biomed Biotechnol 2004, 2004, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Özkan, G.; Sağdiç, O.; Baydar, N.G.; Baydar, H. Anti-oxidant and anti-bacterial activities of Rosa damascena flower extracts. Food Sci Technol Int 2004, 10, 277–281. [Google Scholar] [CrossRef]
- Park, D.; Shin, K.; Choi, Y.; Guo, H.; Cha, Y.; Kim, S.H.; Han, N.S.; Joo, S.S.; Choi, J.K.; Lee, Y.B.; Choi, E.K.; Kim, J.B.; Kim, Y.B. Antimicrobial activities of ethanol and butanol fractions of white rose petal extract. Regul Toxicol Pharmacol 2016, 76, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Kwon, S.; Park, D.; Shin, S.; Jeong, J.; Park, S.; Hwang, S.; Kim, Y.; Joo, S. Anti-allergic effects of white rose petal extract and anti-atopic properties of its hexane fraction. Arch Pharm Res 2009, 32, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.K.; Guo, H.; Choi, J.K.; Jang, S.K.; Shin, K.; Cha, Y.S.; Choi, Y.; Seo, D.W.; Lee, Y.B.; Joo, S.S.; Kim, Y.B. Extraction conditions of white rose petals for the inhibition of enzymes related to skin aging. Lab Anim Res 2015, 31, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Park, D.; Lee, S.; Bae, D.; Yang, Y.; Kyun, J.; Kim, D.; Choi, E.; Hong, J.; Jeong, H.; Kim, H.; Jang, S.; Joo, S.; Kim, Y. Neuroprotective effects of a butanol fraction of Rosa hybrida petals in a middle cerebral artery occlusion model. Biomol Ther 2013, 21, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Yon, J.; Kim, Y.B.; Park, D. The ethanol fraction of white rose petal extract abrogates excitotoxicity-induced neuronal damage in vivo and in vitro through inhibition of oxidative stress and proinflammation. Nutrients 2018, 10, 1375. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Liu, R.H. Processed sweet corn has higher antioxidant activity. J Agric Food Chem 2002, 50, 4959–4964. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.M.; Chun, J.; Lee, H.B.; Lee, J. Influence of heat treatment on the antioxidant activities and polyphenolic compounds of Shiitake (Lentinus edodes) mushroom. Food Chem 2006, 99, 381–387. [Google Scholar] [CrossRef]
- Hwang, I.G.; Woo, K.S.; Kim, T.M.; Kim, D.J.; Yang, M.H.; Jeong, H.S. Change of physicochemical characteristics of Korean pear (Pyrus pyrifolia Nakai) juice with heat treatment conditions. Korean J Food Sci Technol 2006, 38, 342–334. [Google Scholar]
- Hwang, M.R.; Kim, H.E.; Park, D.K.; Heu, Y.C.; Lee, H.J.; Kang, N.J. Induction of oxidative stress and activation of antioxidant enzymes by infection of powdery mildew in cucurbita plants. J Agric Life Sci 2013, 47, 75–81. [Google Scholar]
- Que, F.; Mao, L.; Zhu, C.; Xie, G. Anti-oxidant properties of Chinese yellow wine, its concentrate and volatiles. LWT-Food Sci Technol 2006, 39, 111–117. [Google Scholar] [CrossRef]
- Teixeira de Moraes Costa, M.M.; Penha de Oliveira, S.H.; Gomes-Filho, J.E. Mechanism of calcium hydroxide-induced neutrophil migration into air-pouch cavity. Oral Surgery Oral Med Oral Pathol Oral Radiol Endod 2008, 105, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Perez, D.A.; Vago, J.P.; Athayde, R.M.; Reis, A.C.; Teixeira, M.M.; Sousa, L.; PPinho, V. Switching off key signaling survival molecules to switch on the resolution of inflammation. Mediators Inflamm 2014, 2014, 829851. [Google Scholar] [CrossRef]
- Zhang JM, An JX, Cytokines, inflammation, and pain. Int Anesth Clin 2009, 45, 27–37.
- Winyard, P.G.; Willoughby, D. Inflammation protocols. In Methods in Molecular Biology; Humana Press: New Jersey, 2003; p. 225. [Google Scholar]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: regulation, structure, and inhibition. Med Res Rev 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, S.Q.; Takeda, R.; Okazaki, K.; Sekita, M. Anti-inflammatory activities of amber extract in lipopolysaccharide-induced RAW 264.7 macrophages. Biomed and Pharmacother 2021, 141, 111854. [Google Scholar] [CrossRef]
- Agard, M.; Asakrah, S.; Morici, L.A. PGE2 suppression of innate immunity during mucosal bacterial infection. Front Cell Infect Microbiol 2013, 3, 45. [Google Scholar] [CrossRef]
- Shin, S.; Jeon, H.; Park, D.; Jang, J.Y.; Joo, S.; Hwang, Y.; Choe, S.Y.; Kim, Y.B. Anti-inflammatory effects of an ethanol extract of Angelica gigas in a carrageenan-air pouch inflammation model. Exp Anim 2009, 58, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Goppelt-Struebe, M.; Wolter, D.; Resch, K. Glucocorticoids inhibit prostaglandin synthesis not only at the level of phospholipase A2 but also at the level of cyclo-oxygenase/PGE isomerase. Br J Pharmacol 1989, 98, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antioxidant contents in 24 rosebud extracts and their antioxidative and anti-inflammatory activities (100 μg/mL). (A) Polyphenol contents. (B) 2,2-Azino-bis(3-ethylbenzothiazoline 6-sulfonic acid) (ABTS) radical-scavenging activity. (C) 1,1-Diphenyl-2-picrylhydrazyl (DPPH) radical- scavenging activity. (D) Inhibitory activity on nitric oxide (NO) production from RAW 264.7 macrophages stimulated with lipopolysaccharide (LPS, 1 μg/mL).
Figure 1.
Antioxidant contents in 24 rosebud extracts and their antioxidative and anti-inflammatory activities (100 μg/mL). (A) Polyphenol contents. (B) 2,2-Azino-bis(3-ethylbenzothiazoline 6-sulfonic acid) (ABTS) radical-scavenging activity. (C) 1,1-Diphenyl-2-picrylhydrazyl (DPPH) radical- scavenging activity. (D) Inhibitory activity on nitric oxide (NO) production from RAW 264.7 macrophages stimulated with lipopolysaccharide (LPS, 1 μg/mL).
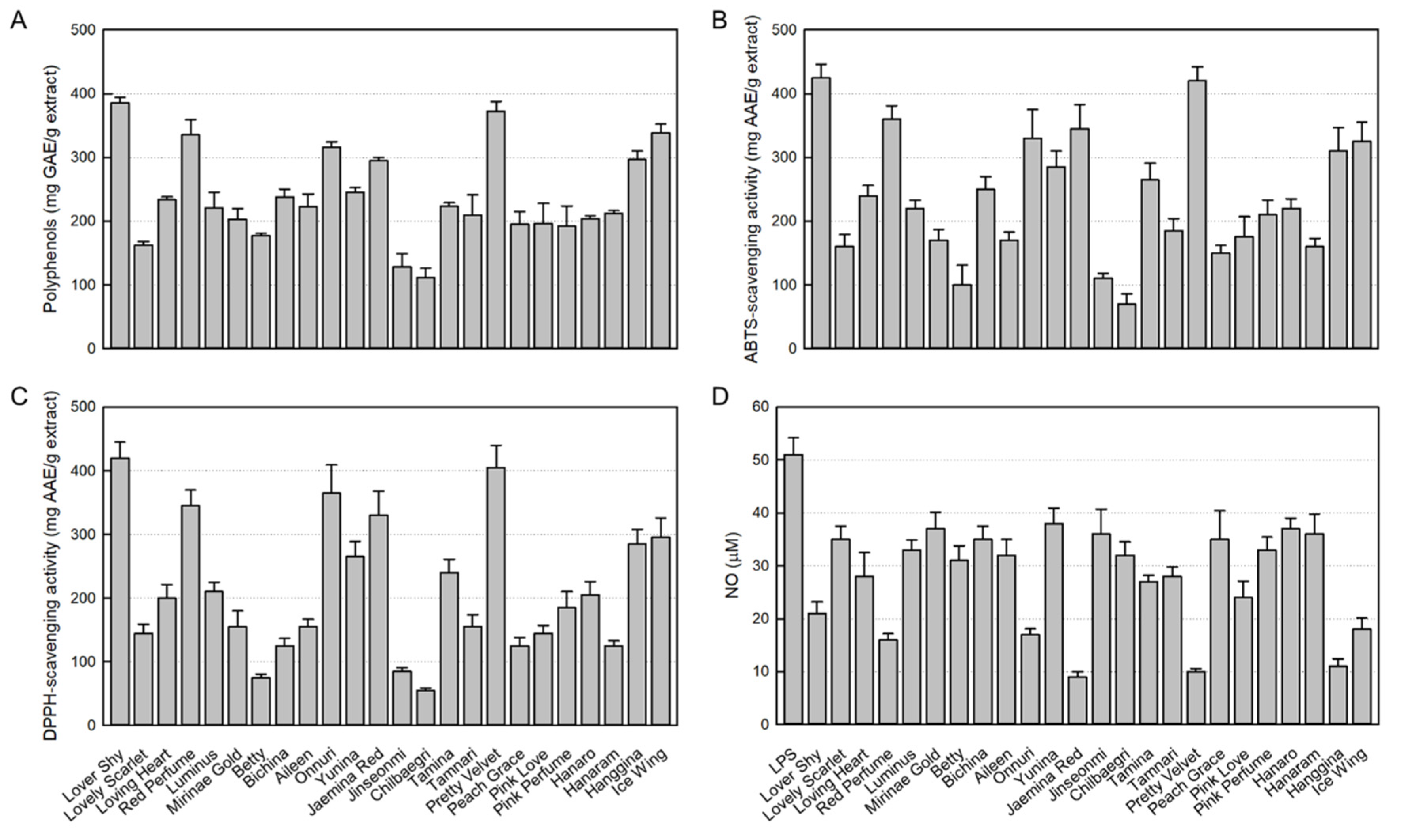
Figure 2.
Correlations between antioxidant ingredients in 24 rosebud extracts and their antioxidative or anti-inflammatory activities (100 μg/mL). (A) Correlation between polyphenols and 2,2-azino-bis(3-ethylbenzothiazoline 6-sulfonic acid) (ABTS) radical-scavenging activity. (B) Correlation between polyphenols and 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical- scavenging activity. (C) Correlation between polyphenols and nitric oxide (NO)-inhibitory activities.
Figure 2.
Correlations between antioxidant ingredients in 24 rosebud extracts and their antioxidative or anti-inflammatory activities (100 μg/mL). (A) Correlation between polyphenols and 2,2-azino-bis(3-ethylbenzothiazoline 6-sulfonic acid) (ABTS) radical-scavenging activity. (B) Correlation between polyphenols and 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical- scavenging activity. (C) Correlation between polyphenols and nitric oxide (NO)-inhibitory activities.
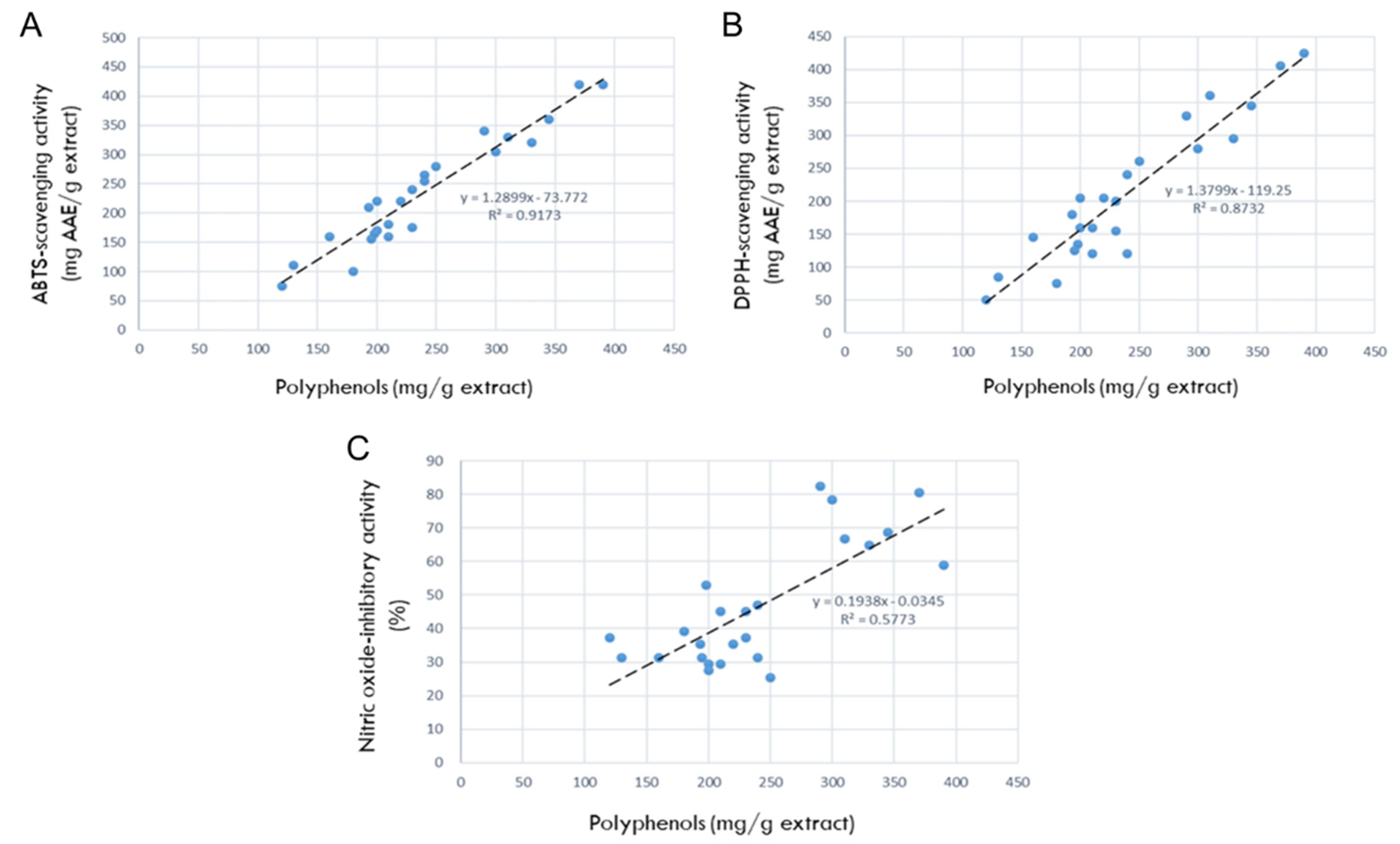
Figure 3.
Cytotoxicity and anti-inflammatory activity of Pretty Velvet rosebud extract (PVRE) in RAW 264.7 cells. (A) Cytotoxicity of PVRE. (B-E) Inhibitory activities on inducible nitric oxide synthase (iNOS) expression (B), cyclooxygenase-2 (COX-2) expression (C), nitric oxide (NO) production (D), and prostaglandin 2 (PGE2) production from RAW 264.7 macrophages stimulated with lipopolysaccharide (LPS). *Significantly different from control (P<0.05). #Significantly different from LPS alone (P<0.05).
Figure 3.
Cytotoxicity and anti-inflammatory activity of Pretty Velvet rosebud extract (PVRE) in RAW 264.7 cells. (A) Cytotoxicity of PVRE. (B-E) Inhibitory activities on inducible nitric oxide synthase (iNOS) expression (B), cyclooxygenase-2 (COX-2) expression (C), nitric oxide (NO) production (D), and prostaglandin 2 (PGE2) production from RAW 264.7 macrophages stimulated with lipopolysaccharide (LPS). *Significantly different from control (P<0.05). #Significantly different from LPS alone (P<0.05).
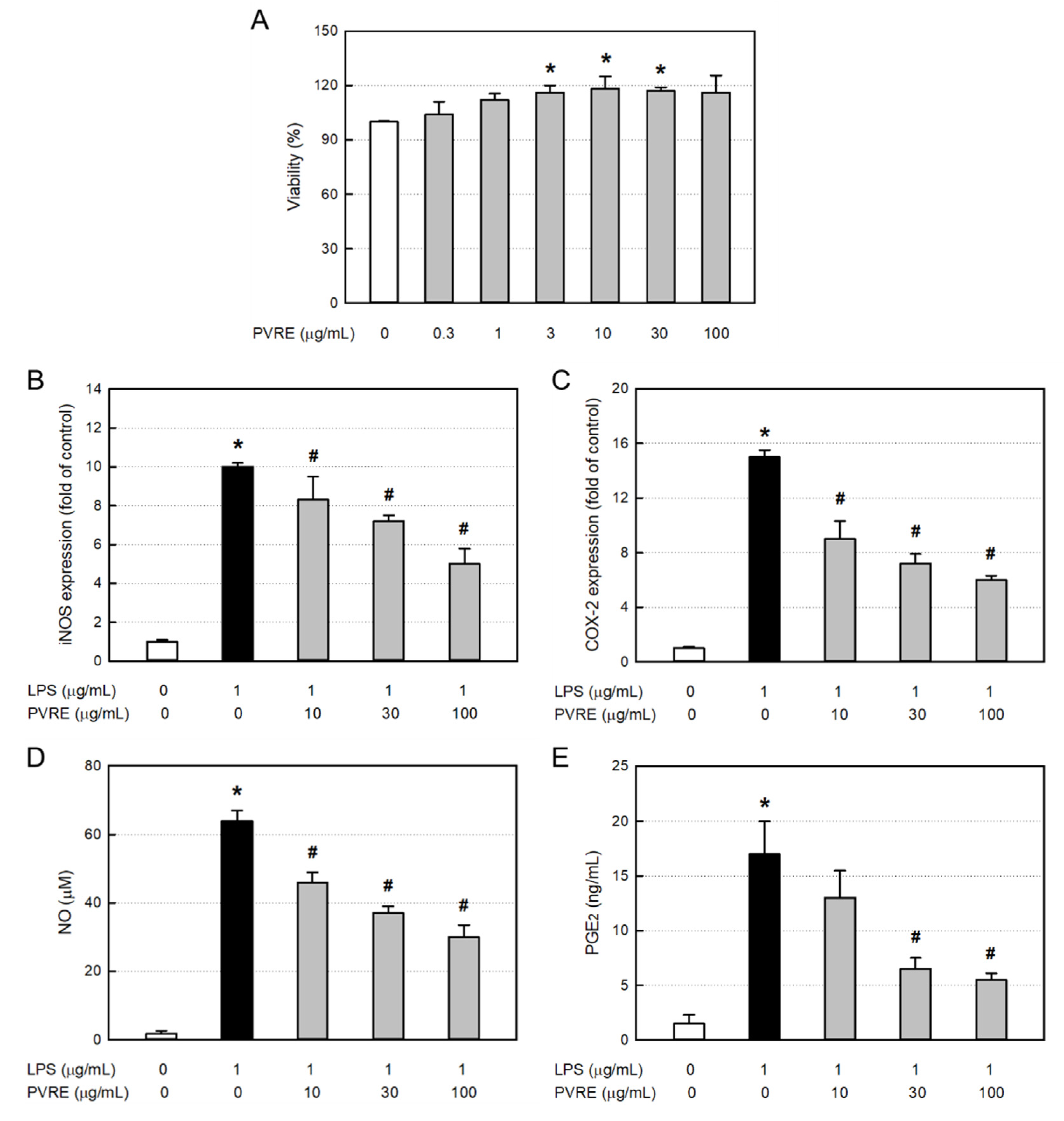
Figure 4.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Effects on λ-carrageenan-induced exudation. (B-E) Effects on the infiltration of white blood cells (WBCs, B), neutrophils (C), monocytes (D), and lymphocytes (E) in the exudate. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
Figure 4.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Effects on λ-carrageenan-induced exudation. (B-E) Effects on the infiltration of white blood cells (WBCs, B), neutrophils (C), monocytes (D), and lymphocytes (E) in the exudate. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
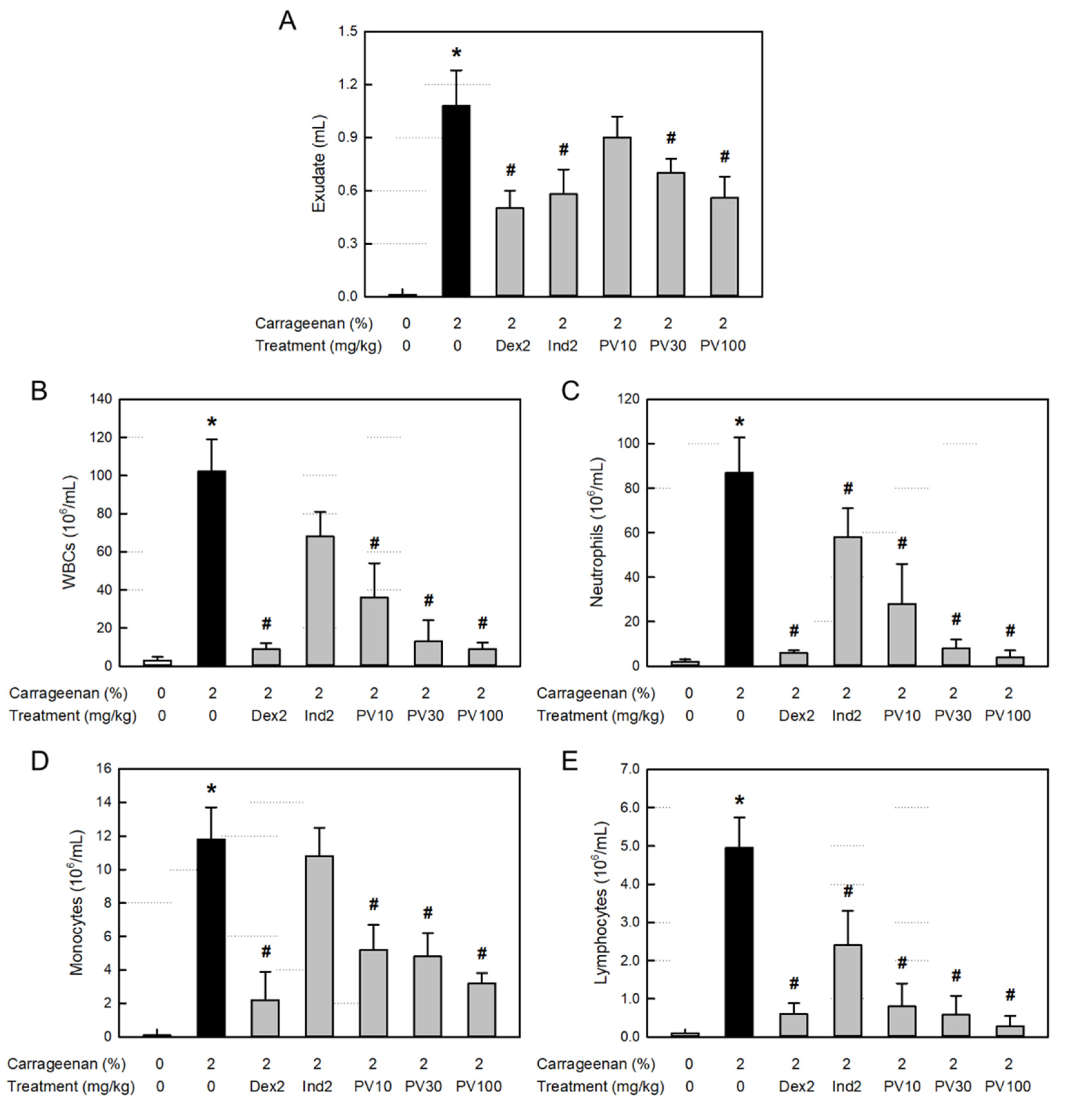
Figure 5.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A & B) Effects on the λ-carrageenan-induced accumulation of tumor necrosis factor-α (TNF-α, A) and interleukin-1β (IL-1β, B). (C & D) Effects on the accumulation of nitric oxide (NO, C) and prostaglandin E2 (PGE2, D). Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
Figure 5.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A & B) Effects on the λ-carrageenan-induced accumulation of tumor necrosis factor-α (TNF-α, A) and interleukin-1β (IL-1β, B). (C & D) Effects on the accumulation of nitric oxide (NO, C) and prostaglandin E2 (PGE2, D). Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
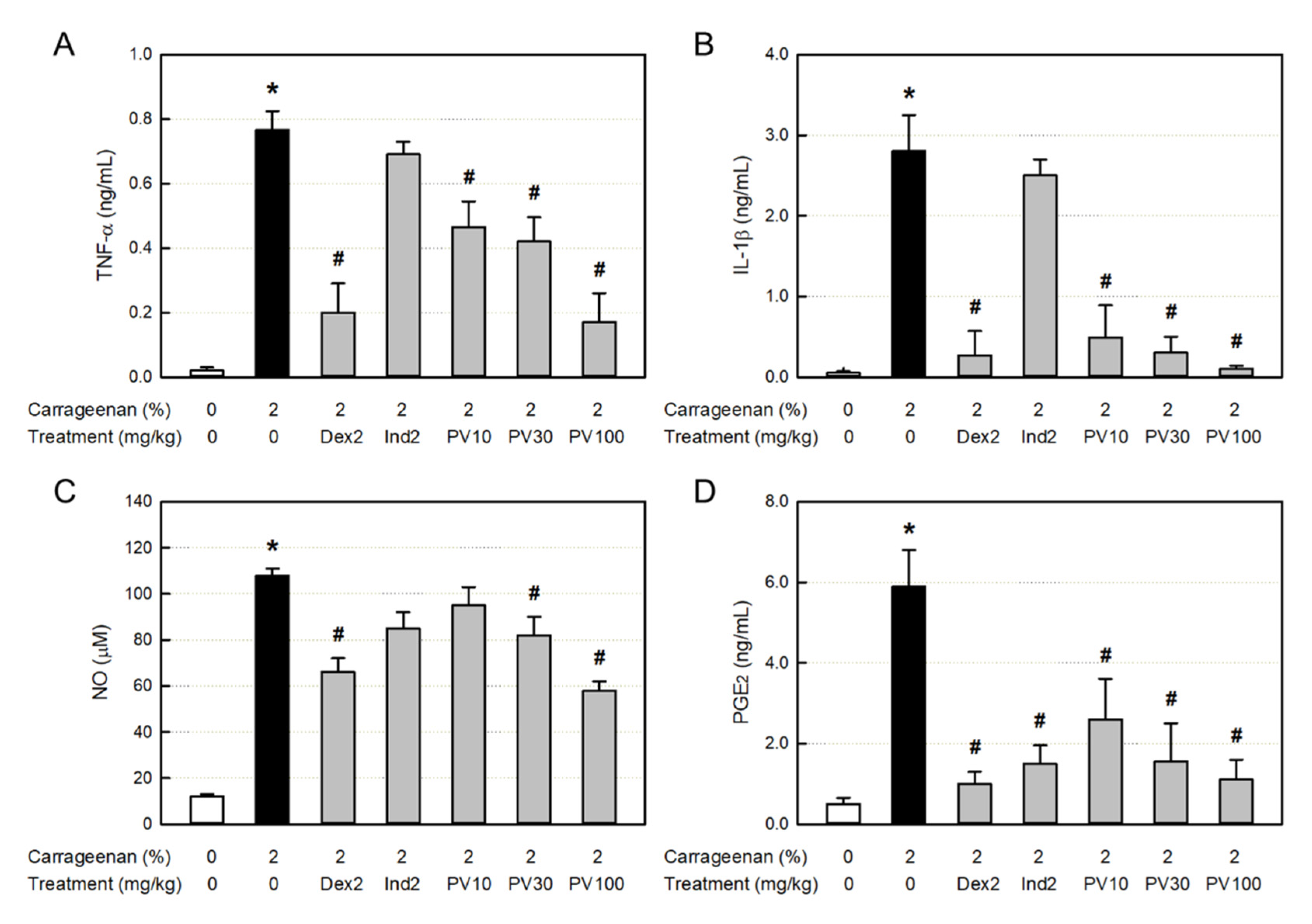
Figure 6.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Representative findings of λ-carrageenan-induced erythema (vascular swelling) in the skin (Upper: gross findings, Lower: Image-J findings). (B) Blood vessel area analyzed with Image-J. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
Figure 6.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Representative findings of λ-carrageenan-induced erythema (vascular swelling) in the skin (Upper: gross findings, Lower: Image-J findings). (B) Blood vessel area analyzed with Image-J. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05).
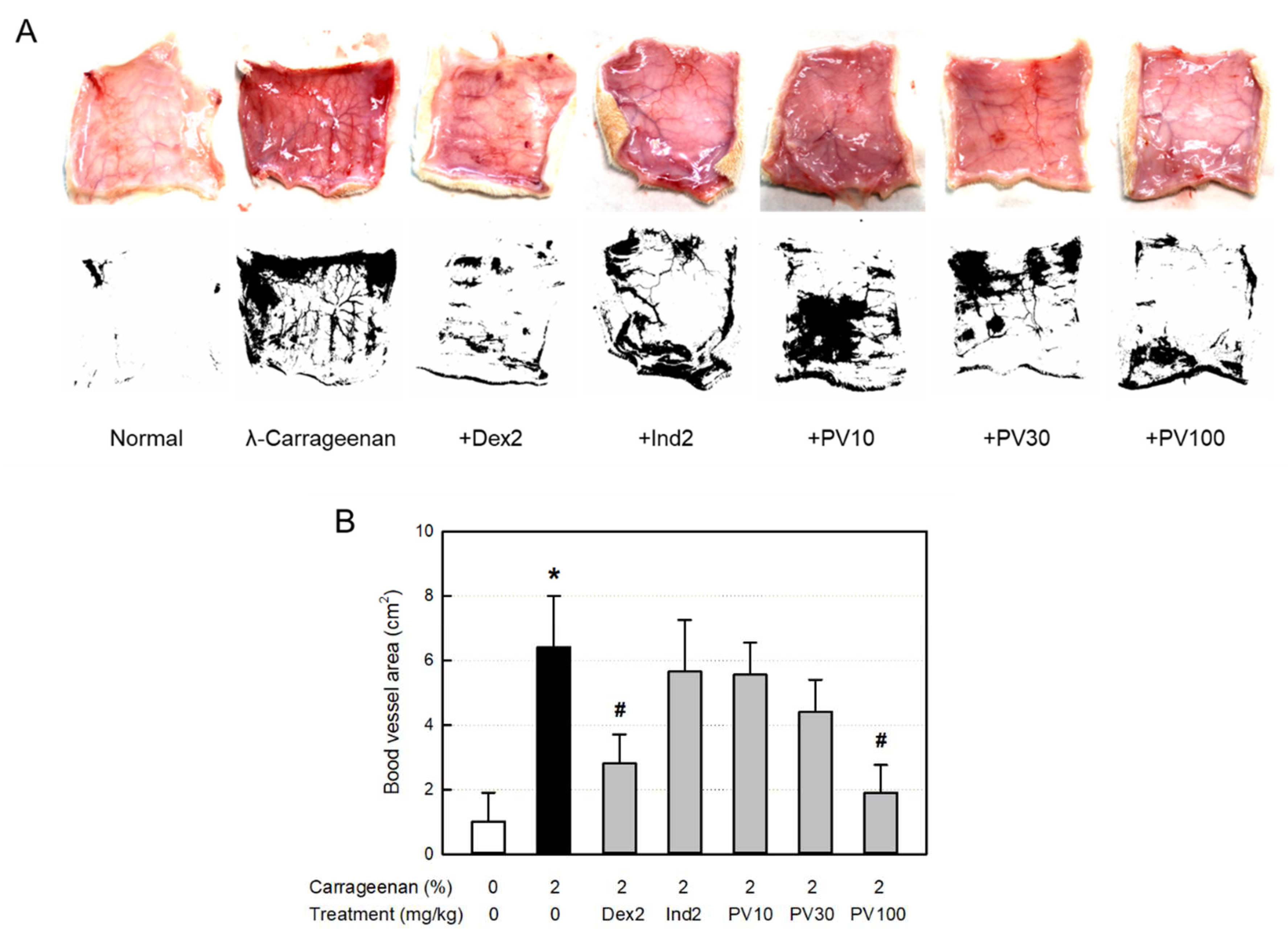
Figure 7.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Representative microscopic findings of λ-carrageenan-induced edema (tissue thickening) and inflammatory cell infiltration in the skin. (B) The thickness of air-pouch lining tissue. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05). Scale bar = 200 μm.
Figure 7.
Anti-inflammatory activities of Pretty Velvet (PV) rosebud extract (PVRE) in a dermatitis animal model. (A) Representative microscopic findings of λ-carrageenan-induced edema (tissue thickening) and inflammatory cell infiltration in the skin. (B) The thickness of air-pouch lining tissue. Dex2: 2 mg/kg dexamethasone, Ind2: 2 mg/kg indomethacin, PV10: 10 mg/kg PVRE, PV30: 30 mg/kg PVRE, PV100: 100 mg/kg PVRE. *Significantly different from control (P<0.05). #Significantly different from λ-carrageenan alone (P<0.05). Scale bar = 200 μm.
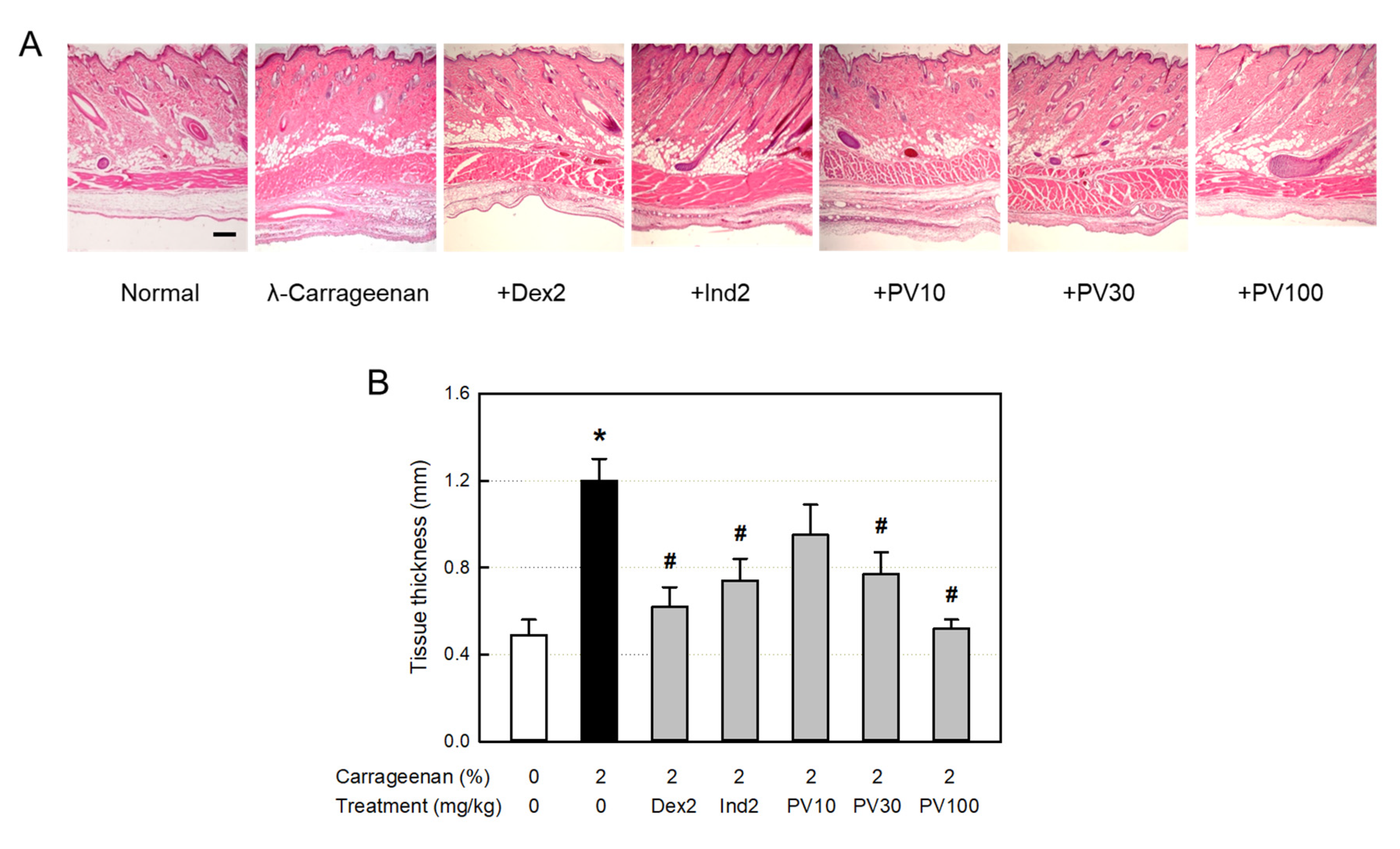

Table 1.
Primer sequences used in this study.
| Genes | Sequence 5’-3’ | Temperature |
|---|---|---|
| iNOS | Forward: CAGGATCCAGTGGTCCAACC Reverse: CGTACCGGATGAGCTGTGAA |
60°C59°C |
| COX-2 | Forward: GTACAAGCAGTGGCAAAGGC Reverse: ACGAGGTTTTTCCACCAGCA |
60°C60°C |
| GAPDH | Forward: GACCTCATGGCCTACATGGC Reverse: GCCCCTCCTGTTATTATGGGG |
60°C59°C |
iNOS: inducible nitric oxide synthase, COX-2: cyclooxygenase-2, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



